
Abstract
Injudicious application of chemical fertilizers in India has a considerable negative impact on economy and environmental sustainability. There is a growing need to turn back to nature or sustainable agents that promote evergreen agriculture. Potassium (K) is an important and well-known constraint to crop production. Very low rates of potash fertilizer application in agricultural production lead to rapid depletion of K in the soil. Depletion of plant-available K in soils results in a variety of negative impacts of the crops yield and soil health. Microorganisms play important role in determining plant productivity. For successful functioning of introduced microbial bioinoculants, exhaustive efforts have been made to explore soil microbial diversity of indigenous community, their distribution, and behavior in soil habitats. Soil microorganisms are directly responsible for recycling of nutrients. K is the third major essential macronutrient for plant growth. The concentrations of soluble potassium in the soil are usually very low, and more than 90% of potassium in the soil exists in the form of insoluble rocks. Use of plant growth-promoting microorganisms (PGPMs) helps in increasing yields in addition to conventional plant protection. The most important PGPMs are Azotobacter, Azospirillum, Acidithiobacillus ferrooxidans, Bacillus circulans, B. edaphicus, B. globisporus, B. mucilaginosus, B. subtilis, Burkholderia cepacia, Enterobacter hormaechei, Paenibacillus kribensis, P. mucilaginosus, and Pseudomonas putida potassium solubilizes; these are eco-friendly and environmentally safe. Therefore, the efficient K-solubilizing microbes (KSM) should be applied for solubilization of a fixed form of K to an available form of K in the soils. This available K can be easily taken up by the plant for growth and development. In this chapter has been discussed isolation, characterization, diversity, and distribution of KSM from diverse stresses such as low and high temperatures, acidity, alkalinity, salinity, drought, and plant-associated applications. These studies elaborate on indigenous K-solubilizing microbes to develop efficient microbial bioinoculant for solubilization of K in different conditions of soil which enhances the plant growth and yield of crops.
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others

Keywords
7.1 Introduction
Countries such as Brazil, China, and India are important food producers and consumers of high amounts of potassium-based fertilizers. In Brazil, around 90% of the potassium required for agriculture is imported (Barbosa Filho et al. 2006). Plants can uptake potassium (K) through the soil minerals, organic materials, and synthetic fertilizers. Consumption of K was exceeded 260 lakh tons for 2 consecutive years (2011 and 2012) in India, and all the K fertilizers were imported across the globe to meet the demand for agricultural productivity (Nagendran et al. 2013), indicating the injudicious application of K fertilizers. K deficiency in the rhizosphere of economically important crops has become an important limiting factor responsible for sustainable development of evergreen agriculture in India (Naidu et al. 2011).
Potassium (K) is one of the major plant macronutrients influencing plant growth, development, and grain quality; its plays a key role in the synthesis of cells, enzymes, proteins, starch, cellulose, and vitamins. Moreover, K not only participates in nutrient transportation and uptake but also confers resistance to abiotic and biotic stresses, leading to enhanced production of quality crops and providing resistance to plant diseases (Epstein 1972; Epstein and Bloom 2005; Maqsood et al. 2013; Pettigrew 2008). The K is absorbed by plants in large amount than any other mineral element except nitrogen (N) and, in some cases, calcium (Ca). Chemical or synthetic K fertilizers are the largest available sources of K rhizosphere; therefore, larger amounts of K fertilizers can be used to promote the availability of K for plant uptake (Li et al. 2007). The concentration of K in straw and grain serves as an indicator whether the K status of crop is deficient or sufficient (Rao et al. 2010). However, K uptake by aboveground parts of plants is assimilated mainly into the straw but not into the grain (Basak and Biswas 2009).
Release of non-exchangeable K to the third exchangeable form occurs when level of exchangeable and solution K is decreased by crop removal, runoff, erosion, and/or leaching (Sparks 1987). With the introduction of high-yielding crop varieties/hybrids and the progressive intensification of agriculture, the soils are getting depleted in potassium reserve at a faster rate. Moreover, due to imbalanced fertilizer application, potassium deficiency is becoming one of the major constraints in crop production. This emphasized the search to find an alternative indigenous source of K for plant uptake and to maintain K status in soils for sustaining crop production (Sindhu et al. 2014; Supanjani et al. 2006).
Plant growth-promoting microbes are heterogeneous groups of microbes associated with plants in diverse ways. The plant-associated microbes colonize the rhizosphere (rhizospheric microbes), the phyllosphere (epiphytes), and inside of the plant tissue (endophytes). The word “endophyte” means “inside the plant” (derived from the Greek words “endon” meaning “within” and “phyton” meaning “plant”). Although there are diverse meanings for the term, endophytes are most commonly defined as those organisms whose “infections are inconspicuous, the infected host tissues are at least transiently symptomless, and the microbial colonization can be demonstrated to be internal”. While microbes are intimately involved in biogeochemical cycling of metals, anthropogenic release of metals has increased bacterial exposure to a high level of metals in some environments (Nies 1999; Suman et al. 2016a, b). A metal may be regarded as toxic if it impairs growth or metabolism of an organism above a certain threshold concentration: both essential and inessential metals may be toxic when supplied at high enough concentrations (Bowen 1966; Gadd 1992).
Many studies on microbial interactions with toxic metals have been made in the context of functions in metalloenzymes, resistance, and transport, but several aspects of metal “metabolism” remain unclear, particularly the mechanisms employed to obtain metals and associated nutrients from insoluble resources (Wakatsuki 1995). Frequently, microorganisms need to solubilize insoluble metal compounds occurring in the natural environment prior to uptake of essential metals and utilization of associated nutrients, e.g., P and S. Different bacterial species, such as species in the genera Pseudomonas, Agrobacterium, Bacillus, Rhizobium, and Flavobacterium, have been tested for their ability to solubilize inorganic phosphate compounds, such as tricalcium phosphate, hydroxyapatite, and rock phosphate (Goldstein 1986). Silicate bacteria were found to resolve potassium, silicon, and aluminum from insoluble minerals (Aleksandrov et al. 1967). K-solubilizing bacteria exert beneficial effects upon plant growth. Their uses as biofertilizers or control agents for agriculture improvement and environmental protection have been a focus of recent research (Deng et al. 2003; Glick 1995). Imbalanced or overdose use of chemical fertilizers have the negative environmental impacts and also increasing costs of crop production; therefore, there is an urgent need to imply eco-friendly and cost-effective agro-technologies to increase crop production. Therefore, the utilization of KSM is considered to be a sound strategy in improving the productivity of agricultural lands.
7.2 Isolation and Identification of Potassium-Solubilizing Microorganism
Potassium-solubilizing bacteria have been isolated and purified on Aleksandrov agar plates (Hu et al. 2006). The composition of the medium (g/liter) is 5.0 g glucose, 0.5 g magnesium sulfate, 0.005 g ferric chloride, 0.1 g calcium carbonate, 2 g calcium phosphate, and 2 g potassium-bearing minerals. Aleksandrov agar medium with different pH (3–11), NaCl concentration (5–20%), temperatures (5–50 °C), and PEG 8000 (−0.5 to −1.5 MPa) were used to isolate diverse groups of K-solubilizing microbes, viz., acidophilic, alkaliphilic, halophilic, psychrophilic, thermophilic, or drought tolerant. Plates were incubated at different temperatures, and time as described earlier by Yadav et al. (2015a). Cultures were purified and maintained at 4 °C as slant and glycerol stock (20%) at −80 °C for further use. Potassium aluminosilicates and mica were used as insoluble potassium-bearing minerals. Microbes showed halo zone on plates were selected and measured the diameter of halo zone. Quantitative potassium solubilization was carried out in Aleksandrov broth. Microbial cultures showed the K solubilization qualitatively were inoculated separately in conical flasks (150 mL) containing 40 mL broth. Available potassium in culture supernatant was determined by using flame photometer.
For identification and phylogenetic profiling of K-solubilizing microbes, the genomic DNA should be extracted for the identification of microbes by the method described by Verma et al. (2016a, b)). The amount of DNA was extracted and assessed by electrophoresis on a 0.8% agarose gel. The 16S rRNA/ITS gene should be amplified as described earlier (Verma et al. (2016a, b)) using the universal primers pA and pH for bacteria and ITS 1 and ITS 2 for fungus (Edwards et al. 1989). The PCR-amplified 16S rRNA/ITS gene was purified. The nucleotide sequences of purified 16S/ITS rDNA have been sequenced with fluorescent terminators (BigDye, Applied Biosystems) and run in 3130xl Applied Biosystems ABI prism automated DNA sequencer. The DNA sequence should be double-checked by sequencing both strands using primers forward and reverse reaction, respectively. The partial 16S rRNA/ITS gene sequences of the isolated strains have been compared with those available in the databases. Identification at the species level has determined using a 16S rRNA/ITS gene sequence similarity of ≥97% with that of a prototype strain sequence in the GenBank. Sequence alignment and comparison have been performed, using the program ClustalW. One sequence from each group was selected as a representative operational taxonomic unit (OTU). The phylogenetic tree was constructed on the aligned datasets using the neighbor-joining method implemented in the program MEGA 4.0.2 (Tamura et al. 2007).
7.3 Diversity of Potassium-Solubilizing Microorganism
Microbial world unique in each ecosystem niche forms the basis of the diversity associated. Agriculture is highly on soils and climatic conditions. The ever-increasing need for food to support the growing population in the country demands a systematic appraisal of its soil and climate resources in order to prepare effective lands. The diversity of K-solubilizing microorganisms inhabiting different environments has been extensively investigated in the past few years. The different groups of microbes have been reported such as bacteria and fungi, which included bacterial phylum Actinobacteria, Bacteroidetes, Firmicutes, Proteobacteria, α-Proteobacteria, β-Proteobacteria, and γ-Proteobacteria (Fig. 7.1), and only two fungal phyla were reported to solubilize potassium, namely, Ascomycota and Glomeromycota (Table 7.2). The last few decades, potassium-solubilizing bacterial genera have been recovered, that is, Acidithiobacillus, Agrobacterium, Aminobacter, Arthrobacter, Azotobacter, Bacillus, Clostridium, Delftia, Enterobacter, Klebsiella, Methylobacterium, Microbacterium, Myroides, Paenibacillus, Pseudomonas, Rhizobium, Salmonella, and Sphingomonas. Very least research has been done in K-solubilizing fungus, with only five genera reported being Aspergillus, Cladosporium, Fusarium, Glomus, and Penicillium (Fig. 7.2).

Diversity of potassium-solubilizing bacteria

Diversity of plant growth-promoting K-solubilizing fungi
7.3.1 Bacteria
Soil bacteria that colonize plant roots and promote growth when added to seeds, roots, or tubers have been termed plant growth-promoting rhizobacteria (PGPR). Different plant growth-promoting rhizosphere bacteria, including associative bacteria such as Azospirillum, Bacillus, Pseudomonas, and Enterobacter groups, have been used for their beneficial effects on plant growth. The mechanisms of plant growth stimulation by associative bacteria are mobilization of nutrients, stimulation of root growth by production of phytohormones, and antagonism against soil-borne plant pathogens. Several studies clearly showed the effect of plant growth-promoting bacteria on plant growth of different crops at different climates and soils. The survival of inoculated PGPR in the plant rhizosphere is in most cases a precondition for a potential plant stimulation effect during the vegetation time or at least during early plant development.
A wide range of rhizospheric bacteria reported as K solubilizers included B. mucilaginosus (Zarjani et al. 2013), B. edaphicus (Sheng 2005), B. circulans (Lin et al. 2002), Burkholderia, Acidithiobacillus ferrooxidans, B. mucilaginosus (Zhang and Kong 2014), Bacillus edaphicus (Sheng and He 2006), Arthrobacter spp. (Zarjani et al. 2013), Enterobacter hormaechei (Prajapati et al. 2013), Paenibacillus mucilaginosus (Liu et al. 2012; Hu et al. 2006), P. frequentans, Cladosporium (Argelis et al. 1993), Aminobacter, Sphingomonas, Burkholderia (Uroz et al. 2007), and Paenibacillus glucanolyticus (Sangeeth et al. 2012). These microbial strains have the ability to solubilize K from K-bearing minerals, but only few bacteria, such as B. edaphicus and B. mucilaginosus, have high capacity for mobilizing and solubilizing of K from minerals (Zhao et al. 2008).
Verma et al. (2016a, b) reported that most of the bacilli solubilized potassium such as Bacillus aerophilus, Bacillus atrophaeus, Bacillus cereus, Bacillus circulans, Bacillus horikoshii, Bacillus licheniformis, Bacillus megaterium, Bacillus mojavensis, Bacillus pumilus, Bacillus sphaericus, Exiguobacterium antarcticum, Paenibacillus amylolyticus, Paenibacillus dendritiformis, Paenibacillus polymyxa, Planococcus citreus, and Planococcus salinarum. The K-solubilizing bacteria may have use in the amelioration of K-deficient soil in agriculture. Diversity analysis of potassium solubilizing bacteria has been reported in different phylum such as 32% γ-Proteobacteria, 21% α-Proteobacteria, 16% Firmicutes, 11% β-Proteobacteria, 10% Proteobacteria, and 5% both Actinobacteria and Bacteroidetes. Maximum potassium-solubilizing bacterial genera have been report from γ-Proteobacteria.
7.3.2 Fungi
The alteration of rock minerals in natural environments is a well-known process mainly caused by the action of water and organic acids produced by plant roots and by microorganisms that accelerate this alteration. Molds are capable of solubilizing elements immobilized in silicates during the decomposition of organic matter, resulting in the production of organic acid. Potassium solubilization has been obtained using molds such as Aspergillus, Penicillium, and Fusarium. The filamentous fungus Aspergillus niger is an exceptionally efficient producer of organic acids, which is one of the reasons for its relevance to industrial processes and its commercial importance. The production of organic acids by A. niger is dependent on the pH of the medium, since the greatest quantities of oxalic acid are produced at a pH between 5 and 8, while it is completely absent below pH 3.0.
Arbuscular mycorrhiza can increase the solubility of the mineral form of potassium by releasing protons, H+, or CO2 and organic acid anions such as citrate, oxalate, and malate. This also increased the nitrogen, potassium, calcium, and iron in the plant leaves and fruits (Veresoglou et al. 2011; Yousefi et al. 2011). The inoculants of the two arbuscular mycorrhizal fungi (AMF) species G. intraradices and G. mosseae was applied in soil on a weight basis, and the increasing potassium uptake by maize crop was recorded (Wu et al. 2005). Ectomycorrhizal fungi particularly isolated UFSC-Pt22 and UFSC-Pt186 and contributed to the increase of the efficiency of alkaline breccias as a source of P and K to the plant growth of Eucalyptus dunnii seedlings, respectively (Alves et al. 2010).
Prajapati et al. (2012) reported that potassium-solubilizing fungi (KSF) strains such as Aspergillus terreus and Aspergillus niger were isolated from various K-rich soil samples and observed that A. terreus and A. niger could solubilize insoluble potassium and showed the highest available potassium in liquid medium by using two various insoluble sources of potassium, i.e., feldspar and potassium aluminum silicate. Aspergillus spp., Aspergillus terreus (Prajapati et al. 2013), Aspergillus niger (Prajapati et al. 2012), and Penicillium spp. (Sangeeth et al. 2012) enhanced K solubilization by mobilizing inorganic and organic K and release of structural K from rocks and minerals (Fig. 7.2). Diverity analysis of potassium solubilizing fungi has been reported in two phylum Ascomycota and Glomeromycota with five genera: Aspergillus 50%, Glomus 20%, Cladosporium 10%, Fusarium 10%, and Penicillium 10%. Aspergillus has been the more frequent potassium-solubilizing fungal genera.
7.4 Distribution of Potassium-Solubilizing Microorganisms
Microbial communities are found in most diverse conditions, including extremes of temperature, salinity, water deficiency, and pH. In order to survive under such extreme conditions, these organisms, referred to as extremophiles, have developed adaptive features that permit them to grow optimally under one or more environmental extremes, while polyextremophiles grow optimally under multiple conditions (Rothschild and Mancinelli 2001). Global work on PGPR for different crops is brief carried out on a hypothesis that PGPR can overcome the burden caused by chemical fertilizer on environment. There are diverse conditions for crops growing in different abiotic stresses of pH, salinity, temperature, and drought (Glick et al. 1999; Verma et al. 2015a).
In an efforts to understand the diversity and distribution of culturable K-solubilizing microbes associated with different crops growing in the diverse environments which included saline soil, acidic soil, water deficiency/drought stress, high temperature, and low temperature, many researchers isolated, enumerated, and characterized potassium-solubilizing microbes for tolerances to abiotic stresses. Tolerance to stress provided by microbial inoculants become more significant with the perspectives of crop production that has losses due to the severity of abiotic stresses (Grover et al. 2011).
7.4.1 Acidophiles
Acidophilus study indicated that lower pH, increase in number of cells, and the consequent increase in viscosity due to EPS are allied factors affecting K solubilization from feldspar. Fourier-transform infrared spectroscopic spectra also showed the functional groups related to them which in turn indicated the presence of EPS, organic acids, and proteins (Cao et al. 2011; Yadav et al. 2011). Numerous studies have shown that Bacillus sp. can promote the release of K from silicate minerals (Badar et al. 2006; Barker et al. 1998). Acidophilic microorganisms has been numerously studied from different crops, rhizospheric soil, cold deserts, etc. in which some microbes were reported as acidophiles such as Bacillus aerophilus, Bacillus amyloliquefaciens, Bacillus atrophaeus, Bacillus cereus, Bacillus circulans, Bacillus licheniformis, Bacillus pumilus, Lysinibacillus fusiformis, Paenibacillus polymyxa, and Planomicrobium sp. could grow at 3 pH (Yadav et al. 2015b). Verma et al. (2013) reported K-solubilizing microbes Bacillus cereus, Bacillus pumilus, Bacillus thuringiensis, Lysinibacillus fusiformis, Planococcus salinarum, Pseudomonas rhodesiae, and Variovorax soli.
7.4.2 Alkaliphiles
Alkaliphilic organisms have a pH optimum for growth above pH 9 and no growth at pH 7. Spore-forming alkaliphilic organism growing at pH 8–10 but not at pH 7 has been described which was so peculiar in its properties that the authors established a new genus for it: Amphibacillus xylanzls (Niimura et al. 1990). Its lack of cytochromes, quinones, or catalase and its ability to form spores under aerobic as well as anaerobic conditions clearly distinguished this organism from the genera Bacillus, Clostridium, and Sporolactobacillus. A wide range of rhizospheric alkaliphilic microorganisms are reported as potassium solubilizers including Achromobacter, Aerobacter, Agrobacterium, Arthrobacter, Bacillus, Burkholderia, Duganella, Exiguobacterium, Klebsiella, Lysinibacillus, Micrococcus, Paenibacillus, Planococcus, Pseudomonas, Psychrobacter, Rhizobium, Stenotrophomonas, and Variovorax (Meena and Kanwar 2015; Verma et al. 2016a, b). These bacteria have been isolated from a variety of rhizospheric and non-rhizospheric soils including sugarcane (Rosa-Magri et al. 2012), tea (Bagyalakshmi et al. 2012), tobacco (Zhang and Kong 2014), and wheat (Verma et al. 2013, 2014, 2015a, 2016a, b).
7.4.3 Halophiles
Salinity affects nearly a third of the agricultural land area worldwide. Due to the upward movement of salts in soil solution in arid and semiarid climates, it is a particular problem in irrigation agriculture under those conditions (Shabala and Cuin 2007). Salinity exerts a twofold stress on the crop (Munns and Tester 2008): it causes an osmotic stress due to decreased soil water potential and an accumulation of salts in the plant cell walls. Microbial research in saline environments has also attracted the interest of researchers due to various biotechnological applications (Sahay et al. 2012). Bacillus alcalophilus, Bacillus aquimaris, Bacillus siamensis, Halobacillus, Paenibacillus dendritiformis, and Lysinibacillus xylanilyticus were reported as halophiles (Yadav et al. 2015c).
A study of potassium transport in the haloarchaeon, Haloferax volcanii, has shown, however, that the intracellular concentrations of potassium observed in this organism cannot be accounted for by passive processes alone and ATP hydrolysis is required to actively transport potassium into the cell to reach the 3.6 M intracellular concentrations that are maintained by Hfx. volcanii (Meury and Kohiyama 1989; Oren 1999). Presently, some K-solubilizing halophilic archaea have been reported: Haloarcula marismortui, Haloarcula vallismortis, and Haloferax volcanii (Ouellette et al. 2015). These microbes have been used for composting of different waste products and materials.
7.4.4 Psychrophiles
Cold-adapted microbes have attracted the attention of the scientific community due to their ability to promote plant growth and produce cold-active enzymes, with potential biotechnological applications in a broad range of industrial, agricultural, and medical processes. Psychrotrophic microbes could be valuable in agriculture as bioinoculants and biocontrol agents for low-temperature habitats. Many cold-tolerant PGPBs have been reported from low-temperature environments including Arthrobacter, Bacillus, Exiguobacterium, Pseudomonas, and Providencia (Mishra et al. 2011; Selvakumar et al. 2011; Bisht et al. 2013; Yadav et al. 2014). Psychrophiles as biofertilizers, biocontrol agents, and bioremediators would be of great use in agriculture under cold climatic conditions.
Cold-adapted microorganisms have been reported from Antarctic subglacial, permanently ice-covered lakes, cloud droplets, ice cap cores from considerable depth, snow, and ice glaciers (Yadav et al. 2015a, b). Many K-solubilizing microbes have been sorted out from different crops growing in cold environments such as Achromobacter piechaudii, Bacillus amyloliquefaciens, Bacillus horikoshii, Bacillus megaterium, Bacillus sp., Exiguobacterium antarcticum, Klebsiella sp., Stenotrophomonas maltophilia, and Stenotrophomonas sp. (Verma et al. 2015c). Among the isolated microbes, four efficient lignocellulolytic psychrotrophic microbes Eupenicillium crustaceum, Paecilomyces sp., Bacillus atrophaeus, and Bacillus sp. and commercial fungal consortia Aspergillus awamori, Aspergillus nidulans, Trichoderma viride, and Phanerochaete chrysosporium were used in the present study.
7.4.5 Thermophiles
Global warming and its associated effects are expected to impose abiotic stresses, such as extremes of temperatures, drought, and flooding, which are bound to have adverse effects on food production. Climate change affects agriculture and the food production system in many ways (Godfray et al. 2011). Crop production is affected by climatic variables such as rising temperatures, changing precipitation regimes, and increased atmospheric CO2 levels. It is also affected by biological variables such as the lengths of the crop growth periods and the crop cycle. Over the past decades, climate change has directly affected the plant growth with different abiotic stresses and change ecosystems.
Thermotolerant microbes are used as plant growth promoters to protect the diverse stresses have resulted in more production and yield in many crops. Verma et al. (2016a, b)) have reported thermotolerant K-solubilizing microbes represented by Bacillus altitudinis, Bacillus siamensis, Bacillus subtilis, Delftia acidovorans, Delftia sp., Methylobacterium sp., Methylobacterium mesophilicum, Pseudomonas aeruginosa, and Salmonella bongori from wheat crop growing in peninsular zone of India. Bacillus altitudinis were also reported as thermotolerant bacteria from thermal springs (Verma et al. 2015b). These bacteria produced different hydrolytic types of enzymes at high temperature.
7.4.6 Xerophiles
Being the quantitatively most important osmoticum in plants, K is a main determinant of cell turgor (White 2013). Since an adequate turgor pressure is required for cell expansion, this parameter is particularly important in growing plants (Mengel and Busch 1982). However, for a crop growing in an increasingly dry soil, the maintenance of turgidity and water uptake from the soil requires a further reduction of the plant’s osmotic potential by an increase in cellular osmolyte concentration. This “osmotic adjustment” may be accomplished by the synthesis of compatible solutes, such as sugar alcohols or amino acids (Hu and Schmidhalter 2005). However, as this process is dependent on the provision of photoassimilates, it is very costly to the plant. In contrast, the uptake and storage of increased amounts of K is an energetically “cheaper” alternative. Accordingly, hyperosmotic treatments, imitating a low soil water potential, cause a sustained K uptake into roots, e.g., of barley (Chen et al. 2005). In the field, an ample K supply will thus support osmotic adjustment and sustain cell expansion at low soil water potentials (Grzebisz et al. 2013).
Microbes have solubilized potassium in water stress condition; these are some species of microbes reported: Paenibacillus polymyxa, Sporosarcina sp., Planococcus salinarum, Bacillus pumilus, Acidithiobacillus ferrooxidans, Bacillus mucilaginosus, Bacillus edaphicus, and Bacillus megaterium (Sheng et al. 2002). Verma et al. (2014) have identified drought-tolerant K-solubilizing microbes such as Bacillus megaterium, Duganella violaceusniger, Paenibacillus dendritiformis, Paenibacillus amylolyticus, Pseudomonas thivervalensis, Psychrobacter fozii, Pseudomonas monteilii, Pseudomonas lini, Stenotrophomonas maltophilia, and Stenotrophomonas sp. from wheat crops growing on central zone of India. These types of microbes protect plants from water deficiency (Fig. 7.3).

Distribution of potassium-solubilizing microbes in diverse environments (Acidophiles:(Barker et al. 1998; Badar et al. 2006; Verma et al. 2013; Yadav et al. 2015b) Alkaliphiles: (Rosa-Magri et al. 2012; Bagyalakshmi et al. 2012; Zhang and Kong 2014; Meena and Kanwar 2015; Verma et al. 2013, 2014, 2015a, 2016a, b), Halophiles: (Meury and Kohiyama 1989; Oren 1999; Yadav et al. 2015c; Ouellette et al. 2015), Psychrophiles: (Mishra et al. 2011; Selvakumar et al. 2011; Bisht et al. 2013; Yadav et al. 2014; Verma et al. 2015c), Thermophiles: Verma et al. 2015b, 2016a, b), Xerophiles: (Sheng et al. 2002; Verma et al. 2014))
7.5 Potassium Availability in the Soil and Its Relevance for Crop Production
Since the 1960s, the world population has doubled from three to seven billion, and this trend will persist in the coming decades. Because of this rapid expansion, a massive increase in crop production is required to meet the food and energy demands of future generations, while also preserving the ecological and energy-related resources of our planet. Additionally, recent climate models predict that incidences and duration of drought and heat stress periods are increasing in many regions, negatively affecting our major crops and thus our food security. Therefore, major challenges for agriculture are to enhance crop yields in more resource-efficient systems and to stabilize plant development and yield formation under biotic and abiotic stress conditions.
7.5.1 Potassium in Soils
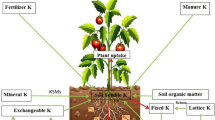
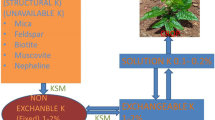
Many soils which were initially rich in K have become deficit due to luxurious utilization by crops and inadequate application of K fertilization, soil fixation, runoff, leaching, and soil erosion by different sources (Sheng and Huang 2002; Archana et al. 2012). As mineral soils contain 0.04–3% K, the total K content of the upper 0.2 m of most agricultural soils generally ranges between 10 and 20 g kg−1 (Jackson, 1964; Sparks 1987). However, in most of the soil, K (90–98%) is incorporated in the crystal lattice structure of minerals and thus not directly available for plant uptake. The availability of K differs greatly with soil type and is affected by physicochemical properties of the soil. To simplify the complex K dynamics in soil, K in soil is often classified into four groups depending on its availability to plants: water-soluble, exchangeable, non-exchangeable, and structural forms (Fig. 7.1).
Water-soluble K is directly available for plants and microbes and potentially subjected to leaching. Exchangeable K is electrostatically bound as an outer-sphere complex to the surfaces of clay minerals and humic substances (Barre et al. 2008). Both fractions are often considered to be easily available for crops. However, the size of both pools is very small. They make up only about 0.1–0.2% and 1–2% of the total K in soil, respectively (Sparks 1987). Non-exchangeable and structural forms are considered to be slowly- or non-available K sources for plants. However, these pools may also contribute significantly to the plant supply in the long term (Pal et al. 2001) (Fig. 7.4).

Effects of microbes and root exudates release different forms of K in the soil
Most of the K in soil is in the structural form, mainly comprised of K-bearing primary minerals such as muscovite, biotite, and feldspars. K-feldspars may directly release K to the soil solution, whereas interlayer K of micas is held tightly by electrostatic forces. Weathering of K-feldspars and micas inherited from soil parent materials produces secondary soil minerals which represent the potential sources of plant-available K in soils (Singh and Goulding 1997). K in trioctahedral micas (such as biotite and phlogopite) is reported to be more readily released by weathering, and to stabilize plant development and yield formation under biotic and abiotic stress conditions (Reynolds et al. 2011). In this context, among the many plant nutrients, potassium (K) plays a particularly crucial role in a number of physiological processes vital to growth, yield, quality, and stress resistance of all crops.
7.5.2 Mechanisms of Potassium Solubilization
Mechanism of potassium solubilization means by which the insoluble potassium and structurally unavailable form of potassium compounds are mobilized and solubilized due to the production of various types of organic acids which are accompanied by acidolysis and complexolysis exchange reactions and these are key processes attributed to the conversion in a soluble form. The efficiency of the K solubilization by various microorganisms was found to vary according to the nature of potassium-bearing minerals and aerobic conditions (Uroz et al. 2009). The release of various types of organic acids were reflected by microorganisms to solubilized the insoluble K to an available form of K. Solubilization of feldspar and illite via rhizospheric microorganisms is due to the production of organic acids like citric acid, tartaric acids, 2-ketogluconic acid, oxalic acid, gluconic acid, malic acid, propionic acid, and fumaric acid, which is easily taken up by the plant. Glycolic and succinic acid seems to be the most frequent agent of K solubilization of mineral (Prajapati and Modi 2012; Zarjani et al. 2013).
Potassium solubilizing microbe solubilized K is done by lowering the pH or by enhancing chelation of the cations bound to K and acidolysis of the surrounding area of microorganism. Such acidolysis by organic acids produced by the rhizospheric microorganisms can either directly dissolve the mineral K as a result of slow releases of exchangeable K, readily available exchangeable K, or can chelate both Si and Al ions associated with K minerals (Romheld and Kirkby 2010). Thus, the synthesis and discharge of organic acids by the microorganisms into the surrounding environment acidify the microbe’s cells and their surrounding environment that ultimately leads to the release of K ions from the mineral by protonation and acidification (Goldstein 1994).
7.5.3 Role of Potassium in Plant Growth Promotion
Plant species are known to differ in their K requirement and in their ability to take up K. The differences in absorption of K among different plant species are attributed to variations in root structure, such as root density, rooting depth, and root hair length for more details on the mechanisms of K uptake in plant roots (Nieves-Cordones et al. 2014) and for aspects on the distribution of K throughout the plant (Ahmad and Maathuis 2014; Wigoda et al. 2014). Positive correlations between K uptake efficiency and root hair length or density in K-depleted soils have been reported for maize, oilseed rape, tomato (Jungk 2001), pea, red clover, barley, rye, and perennial ryegrass (Hogh-Jensen and Pedersen 2003). Mengel and Steffens (1985) hypothesized that rye grass competes for K more effectively than red clover due to its longer root hairs and denser root system. Both morphological parameters may also deplete the K in larger volumes of soil solution, and this depletion of K can initiate the release of non-exchangeable K.
In intact plants, K uptake by leaves is stimulated by light (Blum et al. 1992). However, literature reports comparing the effect of K on photosynthesis in different plant species tend to be inconsistent. Tsonev et al. (2011) showed positive effects of K nutrition on the rate of photosynthesis only in crops subjected to some drought treatment. Similarly, Sen Gupta et al. (1989) also reported that plants supplied with elevated K levels showed similar levels of photosynthetic rates. However, when similar plants were exposed to drought, rates of photosynthesis were positively correlated with application rates of K. There are no clear explanations for how K starvation or suboptimal K nutrition downregulate photosynthesis, e.g., under drought conditions. Therefore, further research is needed to explain these findings, especially for crops. In this context, techniques such as chlorophyll fluorescence imaging may be used for noncontact detection of key physiological parameters regulating photosynthesis (e.g., quantum yield, electron transport rate) and stress defense mechanisms (heat dissipation, chlorophyll fluorescence) from the microscopic to the remote sensing scale (Chaerle et al. 2007). Capturing critical threshold values for potassium deficiencies (and quantifying optimum or slight super-optimum potassium nutrition) under high light, drought stress, or heat stress may be possible at early stages of the vegetation period. In the agronomic literature, high K concentrations in crops have often been termed “luxury consumption” which may be considered as an “insurance strategy” to enable the plant to better survive a sudden environmental stress (Kafkafi 1990).
7.5.4 Effects of Potassium-Solubilizing Microorganisms on Different Crops
The application of organo-minerals with a combination of silicate bacteria for enhancing plant growth and yield of maize and wheat was first reported by Aleksandrov et al. (1967). More importantly, research investigation conducted under field level test crops such as wheat, forage crop, maize, and Sudan grass crops has revealed that KSMs could drastically reduce the usage of chemical or organic fertilizers (Xie 1998). KSMs have been isolated from rhizospheric soil of various plants and from K-bearing mineral (Parmar and Sindhu 2013; Zhang et al. 2013); feldspar (Sheng et al. 2008); potato-soybean-cropping sequence (Biswas 2011); Iranian soils (Zarjani et al. 2013); ceramic industry soil (Prajapati and Modi 2012); mica core of Andhra Pradesh (Gundala et al. 2013); common bean (Kumar et al. 2012); biofertilizers (Zakaria 2009); sorghum, maize, bajra, and chili (Archana et al. 2013); cotton, tomato, soybean, groundnut, and banana (Archana et al. 2012); soil of Tianmu Mountain, Zhejiang Province (China) (Hu et al. 2006); rice (Muralikannan 1996); tea (Bagyalakshmi et al. 2012); Valencia orange (Shaaban et al. 2012); black pepper (Sangeeth et al. 2012); potato (Abdel-Salam and Shams 2012); thyme (Yadegari et al. 2012); eggplant (Han and Lee 2006); peanut and sesame (Youssef et al. 2010); and tobacco (Subhashini and Kumar 2014). Better crop performance was reported to be achieved from several horticultural plants, vegetables, and cereals, which were successfully inoculated with KSMs (Singh et al. 2010; Basak and Biswas 2012; Prajapati et al. 2013). Inoculation with KSMs has been reported to exert beneficial effects on growth of cotton and rape (Sheng 2005), pepper and cucumber (Han et al. 2006), khella (Hassan et al. 2010), sorghum (Badr 2006), wheat (Sheng and He 2006), tomato (Lin et al. 2002), chili (Ramarethinam and Chandra 2005), Sudan grass (Basak and Biswas 2010), and tobacco (Zhang and Kong 2014) Table 7.1.
Prajapati et al. (2012) isolated four different potassium-solubilizing fungi from soils nearby ceramic industries and found that Aspergillus niger and A. terreus possess a greater potassium-solubilizing activity. Aspergillus, Penicillium, and Fusarium were reported for their remarkable activity to solubilize different kinds of insoluble mineral salts in rocks including phosphates, zinc, and potassium salts (Gour 1990; Simine et al. 1998). Lopes-Assad et al. (2010a, b) reported that Aspergillus niger has a better ability to solubilize silicates of potassium and aluminum. Rock powder has been solubilized by Aspergillus niger as a source of potassium for agroecological systems Table 7.2.
According to Archana et al. (2012), the efficient K-solubilizing bacteria Bacillus spp. showed increase in growth and yield of maize. It indicates that the KSMs significantly increased yield, plant growth, and nutrient uptake component over absolute fertilizer control. Supanjani et al. (2006) reported that integration of P and K rocks with inoculation of K- and P-solubilizing bacteria increased K availability from 13 to 15% and P availability from 12% to 21%, respectively. Soil application of KSMs on plant has ~16% photosynthesis and 35% higher leaf area to control. The overall result of this experiment is the treatment of P and K rocks with P- and K-solubilizing bacterial strains that were sustainable and alternative of chemical fertilizer for crop production. Bagyalakshmi et al. (2012) reported that K-solubilizing strains were isolated from rhizosphere of tea and used as biofertilizers of K in tea that have a solubilizing capacity of muriate of potash (MOP) was increased as compared to mineral K sources. Supplementation of glucose and ammonium nitrate was found to be highly effective in solubilization of MOP as compared to the other sources which should be considered prior to the application of these strains in tea soils as bioinoculants.
K-solubilizing microbes as biofertilizers for agriculture improvement can reduce the use of agrochemicals and support eco-friendly crop production (Archana et al. 2012, 2013; Kloepper et al. 1989; Requena et al. 1997; Sheng et al. 2003; Sindhu et al. 2010; Prajapati et al. 2012, 2013). Therefore, it is imperative to isolate more species of mineral-solubilizing bacteria to enrich the pool of microbial species and genes as microbial fertilizers, which will be of great benefit to the ecological development of agriculture (Liu et al. 2012). Plant growth promoting bioinoculants were assumed to have greater importance in sustainable crop protection which could increase the shelf life providing tolerance to increase adverse conditions (Suman et al. 2016a, b). K-solubilizing microorganisms develop efficient indigenous microbial consortia which are required for enhancing plant growth and yield of various crops as well as improving the soil fertility. This type of microbial consortium is cost-effective and environmentally friendly for enhancing the sustainable agriculture.
7.6 Conclusions
In this book chapter, we summarize current knowledge regarding the importance of K in plant growth and quality in changing climate and discuss also the factors controlling K availability in soil. Potassium solubilizing microorganisms play an important role in plant nutrition that enhances the K acquisition of plants through soil which increase plant growth promotion activities; these KSM contributions play an important role to bio-fertilization of agricultural crops. Accordingly, further investigation is required to improve the performance and use of potassium-solubilizing microorganism as efficient microbial bioinoculants. The greater attention is needed for studies and application of new efficient combinations of potassium-solubilizing microorganisms and other plant growth-promoting microorganisms for improved results. The mechanisms explaining the synergistic interaction among KSM required further research to elucidate the molecular basis of these interactions. On the other hand, the application of biotechnological tools for genetic manipulation of potassium solubilizing microorganism increases their potassium-solubilizing efficiency/ability/capabilities and/or the insertion of this trait into other strains of plant growth-promoting effects.
7.7 Future Prospects
The K fertilizer supply is often inadequate due to economic reasons, unavailability of fertilizers, or limited knowledge. Fertilizer application techniques may be still better adjusted to the prevailing crop and growth conditions, e.g. as foliar sprays. An increased utilization of the large plant-non-available pool of soil K could decrease the fertilization requirements and improve crop performance, in particular in low-input systems. Ways to tap this resource would be the introduction of competitive K-mobilizing bacterial strains and the design of more K-efficient crop genotypes by conventional breeding or targeted biotechnological strategies. Promising targets for an improvement of K uptake are root morphology and anatomy, transporter kinetic sand regulation, as well as the release of root exudates. There is considerable variation among species and cultivars in those traits. Optimized K fertilizer application in K limited soils is crucial in order to enhance plant response especially to drought stress via enhancing adaptive/resistance mechanisms of crop plants. Especially, because the demand of K is expected to increase significantly, in particular in developing regions of the world.
References
Abdel-Salam MA, Shams AS (2012) Feldspar-K fertilization of potato (Solanum tuberosum L.) augmented by biofertilizer. J Agric Environ Sci 12(6):694–699
Ahmad I, Maathuis FJM (2014) Cellular and tissue distribution of potassium; physiological relevance, mechanisms and regulation. J Plant Physiol 171:708–714
Aleksandrov VG, Blagodyr RN, Iiiev IP (1967) Liberation of phosphoric acid from apatite by silicate bacteria. Mikrobiyol Zh (Kiev) 29:111–114
Alves L, Oliveira VL, Filho GNS (2010) Utilization of rocks and ectomycorrhizal fungi to promote growth of eucalypt. Braz J Microbio 41:676–684
Archana DS, Nandish MS, Savalagi VP, Alagawadi AR (2012) Screening of potassium solubilizing bacteria (KSB) for plant growth promotional activity. Bioinfolet 9(4):627–630
Archana DS, Nandish MS, Savalagi VP, Alagawadi AR (2013) Characterization of potassium solubilizing bacteria (KSB) from rhizosphere soil. Bioinfolet 10:248–257
Argelis DT, Gonzala DA, Vizcaino C, Gartia MT (1993) Biochemical mechanism of stone alteration carried out by filamentous fungi living in monuments. Biogeo Chem 19:129–147
Badar MA, Shafei AM, Sharaf El-Deen SH (2006) The dissolution of K and phosphorus bearing minerals by silicate dissolving bacteria and their effect on sorghum growth. Res J Agric Biol Sci 2:5–11
Bagyalakshmi B, Ponmurugan P, Marimuthu S (2012) Influence of potassium solubilizing bacteria on crop productivity and quality of tea (Camellia sinensis). Afr J Agri R 7:4250–4259
Barbosa Filho MP, Fageria NK, Santos DF, Couto PA (2006) Aplicação de rochas silicáticas como fontes alternativas de potássio para a cultura do arroz de terras altas. Revista Espaço e Geografia 9(1)
Barker WW, Welch SA, Chu S, Banfield JF (1998) Experimental observations of the effects of bacteria on aluminosilicate weathering. Am Mineralogist 83(11):1551–1563
Barre P, Montagnier C, Chenu C, Abbadie L, Velde B (2008) Clay minerals as a soil potassium reservoir: observation and quantification through X-ray diffraction. Plant Soil 302:213–220
Basak B, Biswas D (2009) Influence of potassium solubilizing microorganism (Bacillus mucilaginosus) and waste mica on potassium uptake dynamics by sudan grass (Sorghum vulgare Pers.) grown under two Alfisols. Plant and Soil 317:235–255
Basak BB, Biswas DR (2010) Co-inoculation of potassium solubilizing and nitrogen fixing bacteria on solubilization of waste mica and their effect on growth promotion and nutrient acquisition by a forage crop. Biol Fertil Soils 46:641–648
Basak B, Biswas D (2012) Modification of waste mica for alternative source of potassium: evaluation of potassium release in soil from waste mica treated with potassium solubilizing bacteria (KSB). ISBN 978-3659298424
Bisht SC, Mishra PK, Joshi GK (2013) Genetic and functional diversity among root-associated psychrotrophic Pseudomonads isolated from the Himalayan plants. Arch Microbiol 195:605–615
Biswas DR (2011) Nutrient recycling potential of rock phosphate and waste mica enriched compost on crop productivity and changes in soil fertility under potato-soybean cropping sequence in an Inceptisol of Indo-Gangetic Plains of India. Nutr Cycl Agroecosyst 89(1):15–30
Blum DE, Elzenga JTM, Linnemeyer PA, Van Volkenburgh E (1992) Stimulation of growth and ion uptake in bean leaves by red and blue light. Plant Physiol 100:1968–1975
Bowen H (1966) Of the trace elements. Nuclear activation techniques in the life sciences, p 393
Cao Y, Wei X, Cai P, Huang Q, Rong X, Liang W (2011) Preferential adsorption of extracellular polymeric substances from bacteria on clay minerals and iron oxide. Coll Surf B Biointerf 83(1):122–127
Chaerle L, Leinonen I, Jones HG, Van der Straeten D (2007) Monitoring and screening plant populations with combined thermal and chlorophyll fluorescence imaging. J Exp Bot 58:773–784
Chen Z, Newman I, Zhou M, Mendham N, Zhang G, Shabala S (2005) Screening plants for salt tolerance by measuring K+ flux: a case study for barley. Plant Cell Environ 28:1230–1246
Deng S, Bai R, Hu X, Luo Q (2003) Characteristics of a bioflocculant produced by Bacillus mucilaginosus and its use in starch wastewater treatment. App Microbiol Biotechnol 60:588–593
Epstein E (1972) Mineral nutrition of plants: principles and perspectives, p 412
Epstein E, Bloom A (2005) Inorganic components of plants. Mineral nutrition of plants: principles and perspectives, 2nd edn. Sinauer Associates, Inc, Massachusetts, pp 44–45
Gadd GM (1992) Metals and microorganisms: a problem of definition. FEMS Microbiol Lett 100:197–203
Glick BR (1995) The enhancement of plant growth by free-living bacteria. Can J Microbiol 41:109–117
Glick B, Patten C, Holguin G, Penrose D (1999) Overview of plant growth-promoting bacteria. In: Biochemical and genetic mechanisms used by plant growth promoting bacteria. Imperial College Press, London, pp 1–13
Godfray H, Pretty J, Thomas S, Warham E, Beddington J (2011) Linking policy on climate and food. Science 331:1013–1014
Goldstein AH (1986) Bacterial solubilization of mineral phosphates: historical perspective and future prospects. Am J Altern Agric 1:51–57
Goldstein, A. H. (1994). Involvement of the quinoprotein glucose dehydrogenase in the solubilization of exogenous phosphates by gram-negative bacteria. Phosphate in microorganisms: cellular and molecular biology. ASM Press, Washington, DC, pp 197–203
Gour AC (1990) Physiological functions of phosphate solubilizing micro-organisms. In: Gour AC (ed) Phosphate solubilizing micro-organisms as biofertilizers. Omega Scientific Publishers, New Delhi, pp 16–72
Grover M, Ali SZ, Sandhya V, Rasul A (2011) Role of microorganisms in adaptation of agriculture crops to abiotic stresses. World J Microbiol Biotechnol 27:1231–1240
Grzebisz W, Gransee A, Szczepaniak W, Diatta J (2013) The effects of potassium fertilization on water-use efficiency in crop plants. J Plant Nutr Soil Sci 176:355–374
Gundala PB, Chinthala P, Sreenivasulu B (2013) A new facultative alkaliphilic, potassium solubilizing, Bacillus Sp. SVUNM9 isolated from mica cores of Nellore District, Andhra Pradesh, India. Research and reviews. J Microbiol Biotechnol 2(1):1–7
Han HS, Supanjani, Lee KD (2006) Effect of co-inoculation with phosphate and potassium solubilizing bacteria on mineral uptake and growth of pepper and cucumber. Plant Soil Environ 52:130–136
Han HS, Lee KD (2006) Effect of co-inoculation with phosphate and potassium solubilizing bacteria on mineral uptake and growth of pepper and cucumber. Plant Soil Environ 52:130–136
Hassan EA, Hassan EA, Hamad EH (2010) Microbial solubilization of phosphate–potassium rocks and their effect on khella (Ammi visnaga) growth. Ann Agric Sci 55(1):37–53
Hogh-Jensen H, Pedersen MB (2003) Morphological plasticity by crop plants and their potassium use efficiency. J Plant Nutr 26:969–984
Hu Y, Schmidhalter U (2005) Drought and salinity: a comparison of their effects on mineral nutrition of plants. J Plant Nutr Soil Sci 168:541–549
Hu X, Chen J, Guo J (2006) Two phosphate- and potassium-solubilizing bacteria isolated from Tianmu Mountain, Zhejiang, China. World J Microbiol Biotechnol 22:983–990. https://doi.org/10.1007/s11274-006-9144-2
Jackson ML (1964) Chemical composition of soils. In: Bear FE (ed) Chemistry of the soil. Van Nostrand Reinhold, New York, pp 71–141
Jungk A (2001) Root hairs and the acquisition of plant nutrients from soil. J Plant Nutr Soil Sci 164:121–129
Kafkafi U (1990) The functions of plant K in overcoming environmental stress situations. In: Development of K-fertilizer recommendations: proceedings 22nd Colloquium of the International Potash Institute, pp 81–93
Kloepper JW, Lifshitz R, Zablotowicz RM (1989) Free-living bacterial inocula for enhancing crop productivity. Trends Biotechnol 7:39–43
Kumar P, Dubey RC, Maheshwari DK (2012) Bacillus strains isolated from rhizosphere showed plant growth promoting and antagonistic activity against phytopathogens. Microbiol Res 67:493–499
Li Q, Li Z, Huang Y (2007) Status of potassium studies and approaches to improving potassium content in tobacco leaves in China. Agric Sci Technol 35:452–455
Lin Q, Rao Z, Sun Y, Yao J, Xing L (2002) Identification and practical application of silicate-dissolving bacteria. Agric Sci China 1:81–85
Liu D, Lian B, Dong H (2012) Isolation of Paenibacillus sp. and assessment of its potential for enhancing mineral weathering. Geomicrobiol J 29(5):413–421
Lopes-Assad M L, Avansini SH, Erler G, Rosa MM, de Carvalho JRP, Ceccato-Antonini SR (2010a) Rock powder solubilization by Aspergillus niger as a source of potassium for agroecological systems. 19th World Congress of Soil Science
Lopes-Assad A M L, Avansini A S H, Erler A G, Rosa A M M, Porto J R (2010b). Rock powder solubilization by Aspergillus niger as a source of potassium for agroecological systems. In 19th World Congress of Soil Science
Maqsood M, Shehzad MA, Wahid A, Butt AA (2013) Improving drought tolerance in maize (Zea mays) with potassium application in furrow irrigation systems. Int J Agr Biol 15:1193–1198
Meena KR, Kanwar SS (2015) Lipopeptides as the antifungal and antibacterial agents: Applications in food safety and therapeutics. Biomed Res Int 2015
Mengel K, Busch R (1982) The importance of the potassium buffer power on the critical potassium level in soils. Soil Sci 133:27–32
Mengel K, Steffens D (1985) Potassium uptake of rye grass (Lolium perenne) and red clover (Trifolium pratense) as related to root parameters. Biol Fertil Soils 1:53–58
Meury J, Kohiyama M (1989) ATP is required for K+ active transport in the archaebacterium Haloferax volcanii. Arch Microbiol 151:530–536
Mishra PK, Bisht SC, Ruwari P, Selvakumar G, Joshi GK, Bisht JK, Bhatt JC, Gupta HS (2011) Alleviation of cold stress in inoculated wheat (Triticum aestivum L.) seedlings with psychrotolerant Pseudomonads from NW Himalayas. Arch Microbiol 193:497–513
Munns R, Tester M (2008) Mechanisms of salinity tolerance. Annu Rev Plant Biol 59:651–681
Muralikannan M (1996) Biodissolution of silicate, phosphate and potassium by silicate solubilizing bacteria in rice ecosystem. M. Sc.(Agric) thesis, Tamil Nadu Agricultural University, Coimbatore
Nagendran K, Karthikeyan G, Peeran MF, Raveendran M, Prabakar K, Raguchander T (2013) Management of bacterial leaf blight disease in rice with endophytic bacteria. World App Sci J 28:2229–2241
Naidu L, Sidhu G, Sarkar D, Ramamurthy V (2011) Emerging deficiency of potassium in soils and crops of India. Karnat J Agricult Sci 24(1)
Nath R, Sharma GD, Barooah M (2015) Plant growth promoting endophytic fungi isolated from tea (Camellia sinensis) shrubs of Assam, India. Appl Ecol Environ Res 13:877–891
Nies DH (1999) Microbial heavy-metal resistance. App Microbiol Biotechnol 51:730–750
Niimura Y, Koh E, Yanagida F, K4 S, Komagata K, Kozaki M (1990) Amphibaciffus xyfanus gen. nov., sp. nov., a facultatively anaerobic sporeforming xylan-digesting bacterium which lacks cytochrome, quinone, and catalase. Int J Syst Bacteriol 40:297–301
Oren A (1999) Bioenergetic aspects of halophilism. Mircobiol Mol Biol Rev 63:334–348
Ouellette M, Jackson L, Chimileski S, Papke RT (2015) Genome-wide DNA methylation analysis of Haloferax volcanii H26 and identification of DNA methyltransferase related PD-(D/E) XK nuclease family protein HVO_A0006. Front Microbiol 6
Pal DK, Srivastava P, Durge SL, Bhattacharyya T (2001) Role of weathering of fine-grained micas in potassium management of Indian soils. Appl Clay Sci 20:39–52
Parmar P, Sindhu SS (2013) Potassium solubilization by rhizosphere bacteria: influence of nutritional and environmental conditions. J Microbiol Res 3(1):25–31
Pettigrew WT (2008) Potassium influences on yield and quality production for maize, wheat, soybean and cotton. Physiologia Plantarum 133:670–681
Prajapati KB, Modi HA (2012) Isolation and characterization of potassium solubilizing bacteria from ceramic industry soil. CIBTech. J Microbiol 1:8–14
Prajapati K, Sharma MC, Modi HA (2012) Isolation of two potassium solubilizing fungi from ceramic industry soils. Life Sci Leaflets 5:71–75
Prajapati K, Sharma MC, Modi HA (2013) Growth promoting effect of potassium solubilizing microorganisms on Abelmoschus esculentus. Int J Agric Sci 3(1):181–188
Ramarethinam S, Chandra K (2005) Studies on the effect of potash solubilizing/mobilizing bacteria Frateuria aurantia on brinjal growth and yield. Pestology 11:35–39
Rao C, Rao AS, Rao K, Venkateswarlu B, Singh A (2010) Categorization of districts based on nonexchangeable potassium: implications in efficient K fertility management in Indian agriculture. Ind J Fert 6:40–54
Reitmeir RF (1951) Soil potassium. In: Norman AG (ed) Advances in agronomy II. Academic Press, New York, pp 113–164
Requena BN, Jimenez I, Toro M, Barea JM (1997) Interactions between plant growth promoting rhizobacteria (PGPR), arbuscular mycorrhizal fungi and Rhizobium spp. in the rhizosphere of Anthyllis cytisoides, a model legume for revegetation in Mediterranean semi-arid ecosystem. New Phytologist 136:667–677
Reynolds M, Bonnett D, Chapman SC, Furbank RT, Manes Y, Mather DE (2011) Raising yield potential of wheat. I. Overview of a consortium approach and breeding strategies. J Exp Bot 62:439–452
Romheld V, Kirkby EA (2010) Research on potassium in agriculture: needs and prospects. Plant Soil 335:155–180
Rosa-Magri MM, Avansini SH, Lopes-Assad ML, Tauk-Tornisielo SM, Ceccato-Antonini SR (2012) Release of potassium from rock powder by the yeast. Torulaspora globosa. Braz Arch Biol Technol 55:577–582
Rothschild LJ, Mancinelli RL (2001) Life in extreme environments. Nature 409:1092–1101
Sahay H, Mahfooz S, Singh AK, Singh S, Kaushik R, Saxena AK, Arora DK (2012) Exploration and characterization of agriculturally and industrially important haloalkaliphilic bacteria from environmental samples of hyper saline Sambhar lake, India. World J Microbiol Biotechnol 28:3207–3217
Sangeeth KP, Bhai RS, Srinivasan V (2012) Paenibacillus glucanolyticus, a promising potassium solubilizing bacterium isolated from black pepper (Piper nigrum L.) rhizosphere. J Spic Aromat Crops 21(2):118–124
Sebastiao MJ, Cabral JMS, Aires-Barros MR (1993) Fusarium solani pisi recombinant cutinase partitioning in PEG/potassium phosphate aqueous two-phase systems. Biotechnol Techn 7(9):631–634
Selvakumar G, Joshi P, Suyal P, Mishra PK, Joshi GK, Bisht JK, Bhatt JC, Gupta HS (2011) Pseudomonas lurida M2RH3 (MTCC 9245), a psychrotolerant bacterium from the Uttarakhand Himalayas, solubilizes phosphate and promotes wheat seedling growth. World J Microbiol Biotechnol 27:1129–1135
Sen Gupta A, Berkowitz GA, Pier PA (1989) Maintenance of photosynthesis at low leaf water potential in wheat role of potassium status and irrigation history. Plant Physiol 89:1358–1365
Shaaban EA, El-Shamma IMS, El Shazly S, El-Gazzar A, Abdel-Hak RE (2012) Efficiency of rock-feldspar combined with silicate dissolving bacteria on yield and fruit quality of Valencia orange fruits in reclaimed soils. J Appl Sci Res 8:4504–4510
Shabala S, Cuin TA (2007) Potassium transport and plant salt tolerance. Physiol Plant 133:651–669
Sheng X (2005) Growth promotion and increased potassium uptake of cotton and rape by a potassium releasing strain of Bacillus edaphicus. Soil Biol Biochem 37:1918–1922
Sheng XF, He LY (2006) Solubilization of potassium-bearing minerals by a wild type strain of Bacillus edaphicus and its mutants and increased potassium uptake by wheat. Can J Microbiol 52:66–72
Sheng XF, Huang WY (2002) Mechanism of potassium release from feldspar affected by the strain NBT of silicate bacterium. Acta Pedol Sin 39(6):863–871
Sheng XF, He LY, Huang WY (2002) The conditions of releasing potassium by a silicate dissolving bacterial strain NBT. Agric Sci China 1:662–666
Sheng XF, Zhao F, He H, Qiu G, Chen L (2008) Isolation, characterization of silicate mineral solubilizing Bacillus globisporus Q12 from the surface of weathered feldspar. Can J Microbiol 54:1064–1068
Simine CDD, Sayer JA, Gadd GM (1998) Solubilization of zinc phosphate by a strain of Pseudomonas fluorescens isolated form a forest soil. Biol Fert soils 28:87–94
Sindhu SS, Dua S, Verma MK, Khandelwal A (2010) Growth promotion of legumes by inoculation of rhizosphere bacteria. In: Khan MS, Zaidi A, Musarrat J (eds) Microbes for legume improvement. Springer-Wien, New York, pp 95–235
Sindhu SS, Parmar P, Phour M (2014) Nutrient cycling: potassium solubilization by microorganisms and improvement of crop growth. In: Geomicrobiol and biogeochemistry. Springer, pp 175–198
Singh B, Goulding KWT (1997) Changes with time in the potassium content and phyllosilicates in the soil of the Broad balk continuous wheat experiment at Rothamsted. Eur J Soil Sci 48:651–659
Singh G, Biswas DR, Marwaha TS (2010) Mobilization of potassium from waste mica by plant growth promoting rhizobacteria and its assimilation by maize (Zea mays) and wheat (Triticum aestivum L.): a hydroponics study under phytotron growth chamber. J Plant Nutr 33(8):1236–1251
Sparks DL (1987) Potassium dynamics in soils. Adv Soil Sci 6:1–63
Sparks DL, Huang PM (1985) Physical chemistry of soil potassium. In: Munson RD (ed) Potassium in agriculture. Madison, WI: American Society of Agronomy, pp 201–276
Subhashini DV, Kumar AV (2014) Phosphate solubilizing Streptomyces spp. obtained from the rhizosphere of Ceriops decandra of Corangi mangroves. Indian J Agric Sci 84(5):560–564
Suman A, Verma P, Yadav AN, Srinivasamurthy R, Singh A, Prasanna R (2016a) Develop lications in food safety and therapeutics. ment of hydrogel based bio-inoculant formulations and their impact on plant biometric parameters of wheat (Triticum aestivum L.) Int J Curr Microbiol App Sci 5(3):890–901
Suman A, Yadav AN, Verma P (2016b) Endophytic microbes in crops: diversity and beneficial impact for sustainable agriculture. In: Microbial Inoculants in Sustainable Agricultural Productivity. Springer, New Delhi, pp 117–143
Supanjani HH, Jung JS, Lee KD (2006) Rock phosphate potassium and rock solubilizing bacteria as alternative sustainable fertilizers. Agron Sustain Dev 26:233–240
Tamura K, Dudley J, Nei M, Kumar S (2007) MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol Biol Evol 24:1596–1599
Tsonev T, Velikova V, Yildiz-Aktas L, Gurel A, Edreva A (2011) Effect of water deficit and potassium fertilization on photosynthetic activity in cotton plants. Plant Biosyst 145:841–847
Uroz S, Calvaruso C, Turpault MP, Pierrat JC, Mustin C, Frey-Klett P (2007) Effect of the mycorrhizosphere on the genotypic and metabolic diversity of the bacterial communities involved in mineral weathering in a forest soil. Appl Environ Microbiol 73:3019–3027
Uroz S, Calvaruso C, Turpault MP, Frey-Klett P (2009) Mineral weathering by bacteria: ecology, actors and mechanisms. Trends Microbiol 17:378–387
Veresoglou SD, Mamolos AP, Thornton B, Voulgari OK, Sen R, Veresoglou S (2011) Medium-term fertilization of grassland plant communities masks plant species-linked effects on soil microbial community structure. Plant Soil 344:187–196
Verma P, Yadav AN, Kazy SK, Saxena AK, Suman A (2013) Elucidating the diversity and plant growth promoting attributes of wheat (Triticum aestivum) associated acidotolerant bacteria from southern hills zone of India. Natl J Life Sci 10(2):219–226
Verma P, Yadav AN, Kazy SK, Saxena AK, Suman A (2014) Evaluating the diversity and phylogeny of plant growth promoting bacteria associated with wheat (Triticum aestivum) growing in central zone of India. Int J Curr Microbiol Appl Sci 3(5):432–447
Verma P, Yadav AN, Khannam KS, Panjiar N, Kumar S, Saxena AK, Suman A (2015a) Assessment of genetic diversity and plant growth promoting attributes of psychrotolerant bacteria allied with wheat (Triticum aestivum) from the northern hills zone of India. Ann Microbiol 65(4):1885–1899
Verma P, Yadav AK, Khanna KS, Kumar S, Saxena AK, Suman A (2015b) Alleviation of cold stress in wheat seedlings by Bacillus amyloliquefaciens IARI-HHS2-30 an endophytic psychrotolerant K-solubilizing bacterium from NW Indian himalayas. Nat J Life Sci 12:105–110
Verma P, Yadav AN, Shukla L, Saxena AK, Suman A (2015c) Hydrolytic enzymes production by thermotolerant Bacillus altitudinis IARI-MB-9 and Gulbenkiania mobilis IARI-MB-18 isolated from Manikaran hot springs. International. J Adv Res 3(9):1241–1250
Verma P, Yadav AN, Khannam KS, Kumar S, Saxena AK, Suman A (2016a) Molecular diversity and multifarious plant growth promoting attributes of Bacilli associated with wheat (Triticum aestivum L.) rhizosphere from six diverse agro-ecological zones of India. J Basic Microbiol 56:44–58
Verma P, Yadav AN, Khannam KS, Mishra S, Kumar S, Saxena AK, Suman A (2016b) Appraisal of diversity and functional attributes of thermotolerant wheat associated bacteria from the peninsular zone of India. Saudi J Biol Sci. https://doi.org/10.1016/j.sjbs.2016.01.042
Wakatsuki T (1995) Metal oxidoreduction by microbial cells. J Ind Microbiol 14:169–177
White PJ (2013) Improving potassium acquisition and utilisation by crop plants. J Plant Nutr Soil Sci 176:305–316
Wigoda N, Moshelion M, Moran N (2014) Is the leaf bundle sheath a “smart flux valve” for K+ nutrition? J Plant Physiol 171:715–722
Wu SC, Cao ZH, Li ZG, Cheung KC, Wong MH (2005) Effects of biofertilizer containing N fixer, P and K solubilizers and AM fungi on maize growth: a greenhouse trial. Geoderma 125:155–166
Xie JC (1998) Present situation and prospects for the world’s fertilizer use. Plant Nutri Fertil Sci 4:321–330
Yadav V, Prappulla SG, Jha A, Poonia A (2011) A novel exopolysaccharide from probiotic Lactobacillus fermentum CFR 2195: production, purification and characterization. Biotechnol Bioinform Bioeng 1:415–421
Yadav AN, Sachan SG, Verma P, Saxena AK (2014) Prospecting cold deserts of north western Himalayas for microbial diversity and plant growth promoting attributes. J Biosci Bioeng 119(6):683–693. https://doi.org/10.1016/j.jbiosc. 2014.11.006
Yadav AN, Sachan SG, Verma P, Tyagi SP, Kaushik R, Saxena AK (2015a) Culturable diversity and functional annotation of psychrotrophic bacteria from cold desert of Leh Ladakh (India). World J Microbiol Biotechnol 31(1):95–108
Yadav AN, Verma P, Kumar M, Pal KK, Dey R, Gupta A, Padaria JC, Gujar GT, Kumar S, Suman A, Prasanna R (2015b) Diversity and phylogenetic profiling of niche-specific Bacilli from extreme environments of India. Ann Microbiol 65(2):611–629
Yadav AN, Sachan SG, Verma P, Kaushik R, Saxena AK (2015c) Cold active hydrolytic enzymes production by psychrotrophic Bacilli isolated from three sub-glacial lakes of NW Indian Himalayas. J Basic Microbiol 56(3):294–307
Yadav AN, Sachan SG, Verma P, Saxena AK (2016) Bioprospecting of plant growth promoting psychrotrophic Bacilli from cold desert of north western Indian Himalayas. Indian J Exp Biol 54:142–150
Yadegari M, Farahani GHN, Mosadeghzad Z (2012) Biofertilizers effects on quantitative and qualitative yield of Thyme (Thymus vulgaris). Afr J Agric Res 7(34):4716–4723
Yousefi AA, Khavazi K, Moezi AA, Rejali F, Nadian NH (2011) Phosphate solubilizing bacteria and arbuscular mycorrhizal fungi impacts on inorganic phosphorus fractions and wheat growth. World Appl Sci J 15(9):1310–1318
Youssef GH, Seddik WMA, Osman MA (2010) Efficiency of natural minerals in presence of different nitrogen forms and potassium dissolving bacteria on peanut and sesame yields. J Am Sci 6(11):647–660
Zakaria AAB (2009) Growth optimization of potassium solubilizing bacteria isolated from biofertilizer. Malaysia Pahang: Bachelor of Chemical Engeneening (Biotech.), Faculty of Chemical and Natural Resources Engineering. Univ, p 40
Zarjani JK, Aliasgharzad N, Oustan S, Emadi M, Ahmadi A (2013) Isolation and characterization of potassium-solubilizing bacteria in some Iranian soils. Arch Agron Soil Sci 59:1713–1723
Zhang C, Kong F (2014) Isolation and identification of potassium-solubilizing bacteria from tobacco rhizospheric soil and their effect on tobacco plants. App Soil Ecol 82:18–25
Zhang A, Zhao G, Gao T, Wang W, Li J, Zhang S, Zhu BC (2013) Solubilization of insoluble potassium and phosphate by Paenibacillus kribensis CX-7: a soil microorganism with biological control potential. Afr J Microbiol Res 7(1):41–47
Zhao F, Sheng X, Huang Z, He L (2008) Isolation of mineral potassium-solubilizing bacterial strains from agricultural soils in Shandong Province. Biodiv Sci 16:593–600
Acknowledgment
The authors are thankful to the Division of Microbiology, ICAR-Indian Agricultural Research Institute, New Delhi, and the Department of Biotechnology (DBT), Ministry of Science and Technology, for providing the facilities and financial support, to undertake the investigations.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2017 Springer Nature Singapore Pte Ltd.
About this chapter

Cite this chapter
Verma, P., Yadav, A.N., Khannam, K.S., Saxena, A.K., Suman, A. (2017). Potassium-Solubilizing Microbes: Diversity, Distribution, and Role in Plant Growth Promotion. In: Panpatte, D., Jhala, Y., Vyas, R., Shelat, H. (eds) Microorganisms for Green Revolution. Microorganisms for Sustainability, vol 6. Springer, Singapore. https://doi.org/10.1007/978-981-10-6241-4_7
Download citation
DOI: https://doi.org/10.1007/978-981-10-6241-4_7
Published:
Publisher Name: Springer, Singapore
Print ISBN: 978-981-10-6240-7
Online ISBN: 978-981-10-6241-4
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)