
Abstract
Sepioteuthis lessoniana is a demersal neritic species that inhabits coral and rock reefs, seaweed, sea grass beds, and estuaries. Due to its wide distribution range in the Indo-Pacific region, S. lessoniana is an economically important resource of many countries. S. lessoniana has been successfully cultured through multiple generations since the 1960s in both open and closed seawater systems in Thailand, Japan, and the USA. The objectives of aquaculture studies are the production of human food in tropical countries and experimental animals in temperate countries. S. lessoniana hatchlings are larger than other loliginid squids, which enables good adaptation to culture conditions and a very high growth rate through the entire life cycle. In tropical waters, individuals can grow to 500 g in less than 150 days. This rapid growth results from a high feeding rate and requires a massive supply of live feed organisms during the early phase of life. The grow-out phase begins after S. lessoniana can accept dead feed. Further studies of artificial feed or mass production of live feed is required in order to make aquaculture of S. lessoniana economically viable on a large scale. The method and studies of S. lessoniana culture in tropical and temperate waters are reviewed.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
1 Importance of this Species in the Market
The fins of Sepioteuthis lessoniana are very large and broad compared to those of other loliginid squids. This has led to its common names, the bigfin reef squid, the bigfin squid, and the oval squid. The fin length (FL) of this squid is 90–100 % of the mantle length (ML), and the fin width is more than 75 % of the ML (Jereb et al. 2010). The bigfin squid is a demersal neritic species and is found between the surface and depths of about 100 m. Its habitats include coral and rock reefs, seaweed and sea grass beds, and estuaries. Its geographical distribution covers the Indo-Pacific region, 20°E–120°W, 40°N–40°S (Dunning 1998; Jereb et al. 2010; Okutani 2005). The maximum body weight (BW) of S. lessoniana is about 2 kg, while the maximum ML is 422 mm for males and 382 mm for females (Jereb et al. 2010). In the shallow waters of the Gulf of Thailand, the maximum ML of these squid is around 305 mm, and squid that are 70–260 mm in length are commonly caught (Chotiyaputta 1995). The first maturation size of female squid is 130–140 mm and their fecundity is approximately 1,000 (400–3,500) eggs (Chotiyaputta 1984, 1988, 1995; Nabhitabhata 1996; Roongratri 1997; Segawa 1987; Segawa et al. 1993b).
S. lessoniana is of commercial and artisanal interest throughout the Indo-West Pacific region. It constitutes about 7 % of Indian east coast cephalopod landings and 5 % of Thai landings (Jereb et al. 2010; Supongpan et al. 1988; Supongpan 1995). It is captured using a variety of fishing gear and comprises about 10 % of the catch by offshore trawlers and purse seine. In the Southeast Asian region where S. lessoniana is the most abundant, i.e. Thailand and Indonesia, squid traps are popular inshore gear for capturing this species. Squid trap fisheries in Thailand annually yield about 7,000 t, and 90–95 % of the catch is comprised of S. lessoniana, the main target species (> 6,000 t). The remaining 5–10 % is comprised of by-catch pharaoh cuttlefish, Sepia pharaonis (Supongpan et al. 1988; Chotiyaputta and Yamrungreung 1998).
The trap used to capture these squid is artisanal, can be made from local materials, and reflects a good understanding of squid behaviour by local fishermen. In Thailand, two types of traps are used: the semicylindrical type (Fig. 17.2; for details, see also Chap. 7, ‘Aquaculture to Restocking’) and the rectangular box type. Both types are 500–800 × 800–1,200 × 500–650 mm, and most of them are 600 × 1,000 × 500 mm (Boongerd and Rachaniyom 1990; Chotiyaputta and Yamrungreung 1998). Trap frames are made of locally available wood and are covered by a polyethylene fishing net that has a 25-mm mesh size. In fishing operations, the trap is suspended in the water column 2–3 m above the bottom using rock sinkers with Styrofoam and bamboo floats. The fishing depth is approximately 6–40 m, depending on the local fishermen’s experience regarding the optimal trap depth. Coconut fronds are used to cover the trap, making it look like a shelter. S. lessoniana egg clusters are hung inside the trap to persuade squid to enter. When egg clusters are scarce, stripped plastic shopping bags are sometimes used as a substitute. Female S. lessoniana, escorted by mated males, enter the trap in search of appropriate substrates for spawning. After entering the trap, a female attaches her eggs to the net covering. Chotiyaputta (1984, 1988) and Roongratri (1997) reported that the sex ratio of squid captured in traps in Thai waters is close to 1:1.
The squid trap is an appropriate fishing gear in view of natural resource conservation, as only full-grown stocks are exploited. Chotiyaputta (1988) reported that 80 % of the squid captured in traps are larger than 140 mm and are therefore fully mature. The fishing period consists of 20 days per month for about 9 months each year. The remaining periods are reserved for natural stock recruitment.
Aquaculture of S. lessoniana has been successfully performed in various places and times. Descriptions of aquaculture efforts include those by: Choe and Ohshima (1961, 1963), Ohshima and Choe (1961), and Choe (1966a, b) of efforts in Korea–Japan; SEAFDEC (1975) of efforts in the Philippines; Nabhitabhata (1978, 1996), Nabhitabhata and Kbinrum (1981), and Nabhitabhata et al. (1984, 1992b, 1996) of efforts in Thailand; Saso (1979), Tsuchiya (1982), Segawa (1987, 1990, 1993), and Ikeda et al. (2003, 2009a) of efforts in Japan; Sivalingam et al. (1993) of efforts in India; Lee et al. (1994, 1998a, b) of efforts in the USA; and Ahmad and Usman (1997) of efforts in Indonesia. All of these studies agree that the very high growth rate and large final size of S. lessoniana make them a promising candidate for commercial aquaculture .
In Japan, S. lessoniana rearing experiments date from the early 1960s when Ohshima and Choe (1961) reared hatchlings for up to 45 days in an open seawater system . Similar rearing experiments were also carried out by regional fisheries experimental stations. In the 1980s, Tsuchiya (1982) succeeded in rearing S. lessoniana in an open seawater system from hatchlings to maturity in 306 days. Later, Segawa (1987, 1993) reported the embryonic development and feeding characteristics of juvenile squid using observations made during rearing experiments. Since then, S. lessoniana has been recognized as an advantageous squid for aquaculture because this species can survive for long periods in captivity, as can cuttlefishes . Lee et al. (1994) succeeded in culturing S. lessoniana for three generations in a closed water system and suggested this squid might be used as a model for neuroscientific and behavioural studies . Advanced trials were carried out by Walsh et al. (2002) who cultured S. lessoniana for seven generations. Ikeda et al. (2003) also established culture traits of S. lessoniana in a closed seawater system , with the goal of using this species to study brain science.
2 State of the Art
The life cycle of S. lessoniana can be completed under aquaculture conditions . Broodstock are either collected from the wild, as they will spawn in captivity, or cultured from egg capsules collected as a by-product of fishing. This is the initial phase of culture. Hatchlings are fed for up to 30 days on live feed (i.e. mysid shrimps or shrimp postlarvae), collected from the wild or produced by hatcheries. Similar facilities and management can be used in both closed and open systems for the nursing of eggs and the nursing of young squid after hatching. Training the young to feed on dead feed is the initial and critical period of the ongrowing phase . A very high growth rate is a hallmark of the aquaculture of this species. Feeding efficiency is high from hatching to maturity. Cultured squid can reach 200 mm length and 500 g weight in 120 days. Culture of this species in floating net cages and other in situ facilities can reduce the cost of production. Future studies should focus on live and artificial feed as well as taking advantage of size dimorphism, including monosex culture and induction of sex reversion from females into larger males and differences in the growth rate of different morphs.
3 Culture Methodology
Culture methodology is expressed in the Sects. 17.3.1 ‘Open Seawater System in Tropical Countries’ and 17.3.2 ‘Open and Closed Seawater System in Temperate Countries’ because of two reasons. First, the methodology developed in different countries serves different main purposes: production of human food in tropical countries and production of experimental animals in temperate countries. Second, some aspects of biological information obtained from studies in different climate zones are contradictory, which are considered to be the results of different natural environment as well as different conditions and management in different culture systems.
3.1 Open Seawater System in Tropical Countries
3.1.1 Background
Countries in tropical zones are located near the equator, where temperatures are higher (20–30 °C), and fluctuate less, than those in temperate zones. The high temperature enables an impressively high S. lessoniana growth rate in culture. On the other hand, the high growth rate is accompanied by shorter life span and smaller final size. Studies of cephalopod culture in tropical countries are performed with the goal of providing food for human consumption. The information in this section comes mainly from Thailand and other Southeast Asian countries such as Indonesia, the Philippines, and India. Because these countries are developing countries, open seawater systems are suitable and typically used in studies of S. lessoniana culture. Open seawater systems require low investment, have low operational costs, and consume less energy than closed seawater systems . In order to reduce costs, researches in these countries are studying S. lessoniana aquaculture using in situ facilities, i.e. net cages or pens.
3.1.2 Broodstock Maintenance
S. lessoniana broodstock can be collected using hand jigs, squid traps , purse seine, or lift nets operating near the shore. Squid-trapping fishermen in Thailand maintain live squid in a ‘live chamber’ in their boats. The live chamber is a partition of the boat that water can flow through and is similar to that used for Sepia pharaonis (see Chap. 12). Jigs and lift nets are also proper gear for collection of broodstock . At landing, squid can be transferred to 500-L plastic tanks for transport to hatcheries. However, bigfin squid have a high metabolic rate and must be placed in oxygen-rich seawater immediately after landing. A change of water or a flow-through system is necessary after inking by the squid that is caused by handling stress. The high dissolved oxygen content of the water should also be maintained during transportation. In hatcheries, broodstock are maintained in 2 m3 concrete tanks at a male to female ratio of 1:1 (Nabhitabhata et al. 2005). Pieces of fishing net with sinkers are placed in the tanks as artificial substrates for the laying of eggs.
S. lessoniana reproduce in tanks, but the acclimatization period depends on the condition of the animals. A healthy pair will mate within a few hours after release into a tank. Consequent spawning takes place soon after (Nabhitabhata et al. 2005). The female swims around touching substrates with her arms in order to examine the surface and shape of materials before attaching her egg capsules. Each female spawns one to three egg masses. Each egg mass contains 100–400 egg capsules that are attached to each other at the capsule stalks. Spawned egg capsules are sometimes removed from their substrates by other males, although the male mate will attempt to defend them. The spawning period may last for as long as 10 days (Nabhitabhata 1983, 1996). After the last spawning of the female mate, the healthy male turns to another female. If another female is already accompanied by her male mate, agonistic contests between two males will occur. The larger male (regardless of the prior mating status of the male) generally prevails, provided he is healthy (normal performance of routine and ritual behaviour is observed).
In vitro fertilization between mature oocytes from the female oviducts and sperm from the spermatophores in the male Needham’s sac has been studied. About 50 % of the oocytes were successfully fertilized (Nabhitabhata et al. 2001a). The oviducal and nidamental gland jellies are not applied to the eggs. The eggs develop for about 4–10 h to the blastula stage before ceasing.
3.1.3 Nursing of Egg Capsules
3.1.3.1 Egg-Capsule Characteristics
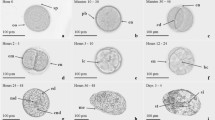
Egg capsules are white, opaque, slender, and cylindrical in the first stage of development . Between 2 and 11 eggs in separate chambers aggregate into a one-string capsule. About five to six eggs per capsule are regularly observed in tropical waters with an average temperature of 28 °C. Larger females tend to lay longer capsules with larger numbers of eggs than smaller females (Nabhitabhata 1983). Egg capsules are attached in clusters of 100–400 capsules. Egg capsules gradually absorb water and increase in size along with the size of the embryos. The shape of each egg capsule changes to a tamarind- or finger-like shape, and the capsule wall becomes more transparent as development proceeds. Eggs can better tolerate transportation and handling during early embryonic developmental stages than during later stages.
Egg capsules collected from the wild vary in size (Fig. 17.1) and embryonic stages. Therefore, the egg capsules must be graded. Newly spawned capsules are at least 50 % smaller than those almost ready to hatch. Egg capsules are graded into four stages according to their size and shape. Newly spawned egg capsules are in the first stage and have a cylindrical shape. Egg capsules in the second stage are similar in shape to those in the first stage, but have comparatively larger egg chambers. In the third stage, the capsules have a finger-like shape, and each chamber of the egg capsule is globular in shape and has become more transparent. Each chamber of the egg capsule becomes cube shaped in the fourth stage.

Sepioteuthis lessoniana egg capsules in sequential stages of development (left to right). (Photograph of J. Nabhitabhata)
3.1.3.2 Water Quality and System Requirements
Egg capsules that contain many eggs require efficient aeration. There must be enough space between egg capsules to allow a sufficient water current to carry enough oxygen to the capsules. Also, with regard to sanitation, elimination of unfertilized eggs and empty egg capsules is conveniently carried out in plastic baskets that float in concrete tanks until the eggs hatch (Nabhitabhata 1978, 1996; Nabhitabhata and Kbinrum 1981; Chindamaikul et al. 1994b). Before spreading in plastic baskets, the capsules must be separated from their clusters.
Nursing of egg capsules can continue under controlled conditions in the tanks previously used for acclimatization in hatcheries. In an open system, changes in physical and chemical parameters should be minimized. Egg capsules are visually checked every day, and those that die, containing abnormally developed embryos, or are empty are discarded in order to prevent infection of others or decomposition and reduction of water quality. Mechanical stimuli and brief changes in temperature or salinity can cause premature hatching. A water flow-through method is used to exchange tank water and to minimize temperature fluctuation in open systems, and flow is maintained at a rate of 1 L min−1. Fresh seawater is pumped through a carbon filter and stored in a concrete tank before it is gravity-fed to the hatchery. Using this system, the water in nursing tanks can be maintained at an average temperature of 28 °C, salinity of 30–33 psu, and pH of 6.0–8.0 (Nabhitabhata et al. 2005). The salinity required to achieve a hatching rate of at least 50 % is 22–37 psu (Nabhitabhata et al. 1991b, 2001c).
Other important physical conditions include turbidity, or suspended solids, and light. Turbidity should be kept as low as possible (Nabhitabhata 1993) through filtration and/or prior sedimentation, particularly in open systems. High levels of suspended solids or high turbidity are critical detrimental parameters. The use of lighting and unnatural daily light–dark periods should be avoided. The growth of algae and fungi on the surface of the egg capsules due to excess light reduces the hatching rate by blocking oxygen supply to the embryos. The attachment of algae can also initiate fungal growth on the capsule membrane and lead to infection of the embryo. The most convenient way to reduce incident light is by curtaining the hatchery with a camouflage net that reduces light efficiency by 80 % (Nabhitabhata et al. 2005).
3.1.3.3 Hatching Event
The hatching rate depends on the biotic and abiotic history during incubation, and embryonic development. The average embryonic period is 20 days (a range of 17–23 days) at about 28 °C (Nabhitabhata 1978, 1996; Nabhitabhata and Kbinrum 1981) and is longer at lower temperatures. The hatching period, from the hatching of the first egg to the hatching of the last eggs in the same cluster of the same spawning, is 3–7 days. More than 50 % of the hatching occurs at night after 2400 h on the second and third day from the first hatching. The hatching rate is generally more than 90 % but may vary depending on the embryonic stage at the time of collection from the wild. Hatching rates of eggs spawned in captivity are low due to a larger percentage of unfertilized eggs and abnormally developed embryos than are found in eggs collected from the wild. The hatchlings pass through the basket mesh into the nursing tank. When the optimum density of hatchlings in the tank is achieved, the egg baskets are transferred to another tank containing (previously prepared) water of similar quality.
Egg baskets are transferred between tanks to control the hatchling density in tanks and to grade the size of hatchlings. Egg-capsule-nursing tanks are then used to nurse the young. Hatchlings may be handled if extreme caution is used to avoid skin damage that could subsequently cause mortality. Transfer of hatchlings is better avoided, and it is much more appropriate to transfer egg capsules sometime before hatching.
3.1.4 Nursing of Young
3.1.4.1 Characters and Habits of Young
S. lessoniana hatchlings are planktonic (Fig. 17.2) and, compared to other loliginids, are much larger with mantles that are 3.5–6.4 mm, or approximately 5.4 mm, in length (Lee et al. 1994; Nabhitabhata 1978, 1996), and with BWs of approximately 0.04 g. Hatchlings in good health swim with their heads down and hover at a 45–60° angle at approximately 75 % of the total water depth above the bottom of the tank. They begin to feed immediately after hatching. Hatchlings are photopositive and prefer low-intensity light (Nabhitabhata 1978; Nabhitabhata and Kbinrum 1981). Hatchlings are dark brown in the light and are pale or transparent in dim light or dark areas. The arms are still comparatively short, but they are strong enough to catch prey that are equal to, or sometimes larger than, their own ML. When feeding on small crustaceans, hatchlings seize prey using their arms and the prey are bitten at the posterior part of the carapace. The tail parts of the prey are eaten first, and then the flesh is gradually consumed. The hard structures of the prey are left uneaten.

Planktonic hatchling of Sepioteuthis lessoniana. (Photograph of J. Nabhitabhata)
3.1.4.2 Water Quality and System Requirements
The planktonic habit of hatchlings requires a directed current that allows them to hover in the water column. Aeration has to be managed to generate a directional flow in the tanks. Adjusting the current velocity to an optimum velocity can reduce stress and the energy consumption required for countercurrent swimming. The optimum current velocity can be observed from the angle of hovering. The hovering angle should be 45–60° to the tank bottom at all times. A simple air-lift system is used to generate the appropriate horizontal flow in the cylindrical tank. In a circular tank that is 2–3 m in diameter with a water depth of 0.6 m, such a system can be constructed from a PVC pipe that is 50 mm in diameter and 800 mm in length and that has been cut longitudinally in half (for details of a similar system on a smaller scale, see Chap. 15 ‘Euprymna hyllebergi’; Fig. 15.3). Each half is drilled at one end where an air pipe and air stone are inserted into the interior face. Both prepared half-pipes are placed facing the same direction at a 45° angle with the air stone at the bottom of the tank. The number of these flow-generating pipe sets used in one tank depends on the size and shape of the tank. At least two sets are required in each circular tank and at least four sets are needed in each rectangular tank, one set at each corner. The current velocity can be adjusted by adjusting the aeration rate.
The maintenance of water quality and tank management in the nursing phase may need to be adjusted from those of the egg-nursing phase. Hatchlings can tolerate a range of salinity from 20 to 40 psu for at least 24 h. The salinity required for more than 50 % survival is 23–36 psu and the pH required for more than 50 % survival is 6.3–8.4 (Nabhitabhata et al. 1991a, 1992a, 2001c; Nabhitabhata and Kbinrum 1984). S. lessoniana hatchlings can tolerate ammonia concentrations (NH4-N) up to 0.06 mg·L−1, nitrite (NO2-N) up to 0.08 mg·L−1, and phosphate (PO4-P) up to 6.0 mg·L−1 (Chindamaikul et al. 2001; Chainak et al. 2003). The routine management of nursing tanks follows directly from the egg-nursing phase.
In larger tanks, the water level in the nursery tanks is adjusted to accommodate the number and size of live feed organisms. An appropriate level of the feed and cephalopod biomass in the tank reduces excess food-hunting activity. The initial water depth is 500 mm at hatching and the density of hatchlings is 5–10 individuals·L−1. As the squid grow, the water depth is increased by 50 mm every day or every second day. Squid are graded by size every 10 days, and squid of similar size are grouped together for subsequent nursing. The density of squid is decreased by about 20–25 % in each group after grading.
3.1.4.3 Feeding
S. lessoniana hatchlings begin to feed immediately after hatching and must be provided with live feed of appropriate size, which is 50–200 % of their ML. Wild-caught live mysids (Mesopodopsis orientalis) that are about 6–7 mm in total length are key to the successful nursing of hatchlings (Nabhitabhata 1978). The live feed is stocked and fed to the squid ad libitum. Pygmy goby fry (Stigmatogobius romeri) are also a good feed organism (Chankaew et al. 2003a). Other wild zooplankton and fish larvae are suitable food, but as the supply of these organisms is not consistent, they are not reliable enough for the mass production of squid.
Feed organisms that are commercially produced can ensure more reliable and consistent success of squid hatchery operations. Postlarvae of the penaeid shrimp (Penaeus merguiensis) that are 5–10 mm in total length and sea bass fry (Lates calcarifer) that are 5 mm in length can be used as the primary foodstock. About 7 (1–22) postlarvae or fish are consumed daily by each squid hatchling (Chankaew et al. 2003b; Nabhitabhata et al. 1996). Other crustaceans that have been tried did not result in good squid production . The crab mysis Portunus pelagicus can initiate seizures in squid hatchlings if fed in the first 2–3 days after hatching. Young squid fed freshwater prawn mysis (Macrobrachium rosenbergii) each consumed about 140 individuals per day, but only 8 % of the hatchlings survived longer than 10 days (Nabhitabhata et al. 1992b). One reason for this low rate of hatchling survival was that the prawn mysis could not survive in 30 psu seawater long enough to provide the required quantity of feed for the squid hatchlings.
The survival rate of young squid fed shrimp postlarvae (P. merguiensis) is approximately 80 % (22–100 %) from hatching to 10 days of age (Chindamaikul et al. 1994a; Nabhitabhata 1978; Nabhitabhata and Kbinrum 1981; Nabhitabhata et al. 1996). The survival rate during this period decreases if the supply of feed organisms is insufficient (Nabhitabhata 1978, 1996; Nabhitabhata and Kbinrum 1981).
The daily feeding rate of squid hatchlings is about 28 % of their BW and the average conversion rate is more than 50 % (28–95 %; Nabhitabhata et al. 1992b, 1996). The unit cost of production in Thailand has been estimated to be about US$ 10 for 100 hatchlings with a feed cost of about US$ 9 in 1996 (Nabhitabhata et al. 1996), when US$ 1 is approximately equal to Thai Baht 25 (US$ 1 equals Thai Baht 30 in 2013). At 30 days after hatching, survival is about 60 % (10–74 %; Nabhitabhata 1978; Nabhitabhata and Kbinrum 1981; Nabhitabhata et al. 1996; Table 17.1).
Young squid can alternatively be nursed in a floating net cage. Ahmad and Usman (1997) nursed their S. lessoniana from hatchlings in a net cage that was 1.5 × 1.5 × 1.5 m and provided Mesopodopsis mysids as feed organisms. The growth was rapid with an increase of 2.9–4 g after 30 days, as compared to about 2 g in concrete tanks. The cost of production using this method may be lower than that in tanks because the cost of the aeration and water quality maintenance required for both open (water flow-through) and closed systems is eliminated.
3.1.4.4 Growth
Beginning 10 days after hatching, the habit of the young S. lessoniana obviously changes from planktonic to pelagic or nektonic. The young lower their level of hovered swimming from near the surface level to a depth of about 50 % of the total water depth. They begin to gather in schools of similar body size and respond to stimuli in a synchronized manner. The ritual colour pattern on the dorsum changes from dark brown to green and golden brown with white spots. The FL increases from approximately 33 % of the ML at hatching to 74 % of the ML at 30 days of age (Nabhitabhata 1978, 1983, 1996; Nabhitabhata and Kbinrum 1981).
The rate of growth is highest over the first 10-day period. In open systems, S. lessoniana grow to 11 mm and 0.55 g by the tenth day after hatching, with a daily growth rate of 6.7 % for ML and 17.3 % for weight (Fig. 17.3). During the first month (0–30 days), the daily growth rate for ML is 2.8–6.7 % and the daily growth rate for weight is 6.4–17.3 %. The relationship between ML (mm) and weight (W; g; Fig. 17.4) in the first (nursing) phase from hatching to 30 days of age at 28 °C in open seawater systems (Nabhitabhata 2002) is given by:

Growth in terms of the relationships between mantle length (mm, above), weight (g, below), age (d, solid line), and daily growth rate (%, broken lines) in length and weight in an open seawater system. Arrows indicate mating (m) and spawning (s). (From Nabhitabhata 2002)

Growth in terms of the relationship between mantle length (mm) and weight (g) in an open seawater system. Arrows indicate mating (m) and spawning (s). (From Nabhitabhata 2002)
Growth in terms of the ML–age (T; d) relationship (Fig. 17.5) in the first (nursing) phase at 28 °C is given by:

Growth in terms of the relationship between mantle length (mm) and age (d) in an open seawater system. Arrows indicate mating (m) and spawning (s). (From Nabhitabhata 2002)
3.1.5 Ongrowing
3.1.5.1 Water Quality and System Requirements
The ongrowing tanks and water management systems for open systems are similar to those for nursing of the young. Open systems operate at 24–30 °C, a higher temperature range than closed systems (20–24 °C). This results in a shorter life span and smaller final size.
3.1.5.2 Feeding
Training S. lessoniana to feed on dead feed is a critical process which affects the subsequent success of the ongrowing phase. At 20 days after hatching, young S. lessoniana are trained to feed on dead meat. Squid are fed on sliced fish meat to satiation twice daily at 0800 and 1600 h. There has been no sign of specific preferences for the species of fish consumed. The sizes of the fish chips correspond to the size of the squid, and are one to two times the ML. Squid seize their food in the water column and eat it while hovering. When squid feed on live prey, the prey is seized with the tentacles and retained using the arms. When the squid are fed dead feed, they change their feeding behaviour ; they use only their arms to seize the food and do not perform a positioning step. This change indicates the success of the training period and that the squid are now accustomed to the dead feed and culture conditions (human appearance with feed and the environment of the culture tank). One reason for the change in behaviour is the lower motion of the dead feed compared to active live feed. This kind of behavioural change is similar to that observed for cultured cuttlefish , Sepia pharaonis and Sepiella inermis (see Chaps. 12 and 13). Aggressive behaviour and cannibalism become prominent at this age whenever food is in short supply and/or there are squid with a wide range of sizes in the same tank. Larger starved squid that have not learnt to consume dead feed may attack and feed on smaller squid in the same tank. Human trainers must ensure sufficient feeding and that every squid is fed.
3.1.5.3 Growth
Growth is very rapid although the rate of increase in size and weight decreases as maturity approaches. In open seawater systems , the mantle growth rate decreases to less than 3 % daily after 40 days of age and the rate of weight gain decreases to less than 5 % after 50 days of age. For S. lessoniana reared at 28 °C, the overall average daily growth rate from hatching to 130 days of age is 2.8 % for length and 6 % for weight (Nabhitabhata 1996). As in the first 30 days after hatching, the relationship between ML and W (Fig. 17.4) is exponential in the second phase (30–60 days after hatching) and the third phase (60–176 days after hatching; Nabhitabhata 2002). The ‘b’ values from aquaculture batches, 2.738 and 2.502, are higher than those from wild stocks, 2.218–2.477 (Rattana-anant 1978, 1979, 1980; Roogratri 1997; Sivashanthini et al. 2009), revealing higher growth rates under culture conditions than those that occur in the wild. Growth models for cultured batches at 28 °C in open seawater systems (Nabhitabhata 2002) are:
Growth, in terms of the ML–age relationship (Fig. 17.5), in the second phase and the third phase can be modelled as:
Models that fit plots of the weight–age relationship (Fig. 17.6) from hatching to 60 days of age (early phase) and from 60 to 176 days of age (ongrowing phase) are:

Growth in terms of the relationship between weight (g) and age (d) in an open seawater system. Arrows indicate mating (m) and spawning (s). (From Nabhitabhata 2002)
In open seawater systems at 28 °C, mating occurs at about 90 days of age and spawning is 1–3 weeks later, at about 110 days of age. The average life span of cultured batches is about 130–140 days, or within 1 month after spawning. The final size of cultured squid is about 250 mm in length and 600 g in weight (Nabhitabhata 1978, 1983, 1996; Nabhitabhata and Kbinrum 1981; SEAFDEC 1975; Tables 17.1 and 17.2).
3.1.5.4 Alternative Culture Facilities
Ongrowing in net cages that float in natural waters is another option for S. lessoniana culture. Nabhitabhata et al. (1984) suggested that the most appropriate structure was a box-type cage, hanging from a floating buoy or a bamboo raft with sinkers at the corners. The cages are made from a monofilament nylon net with a 13-mm mesh size, corresponding to the initial size of squid. Underwater hard frames for cages are avoided in order to prevent collisions or rubbing of squid that cause skin damage and consequent infectious disease. In order to fit into the existing net-cage facilities, the cage dimensions are 2 × 2.5 × 2 m and are submerged to a depth of 1.5 m. Cages therefore contain 7.5 m3 seawater and have a 5-m2 surface area. The optimum squid density is reported to be about 5–10 individuals·m−2 (3–7 ind·m−3) through a 60-day period for squid with an initial ML of about 50 mm length and weight of 16 g (40 days; Nabhitabhata 1978, 1996; Nabhitabhata and Kbinrum 1981; Nabhitabhata et al. 1996). Because the nets are made from monofilament, cages should be left at the ongrowing sites until fouling organisms are observed growing on the net. The fouled net has enhanced visibility to the squid and reduces their collisions with the net.
3.2 Open and Closed Seawater System in Temperate Countries
Methods of aquaculture for S. lessoniana in tropical countries and temperate countries can share many things. In this section, the methodology specifically applied to closed water systems and open water systems in temperate countries will be described for comparison.
3.2.1 Broodstock Maintenance
In Japan, jigging and set net are the main fishery tools for collecting subadult and adult S. lessoniana. Munekiyo and Kawagishi (1993) and Ueta (2000a) found that S. lessoniana caught by set net follow a lunar cycle; the catch amount increases at the full moon and decreases at the new moon. The sex ratio of S. lessoniana caught by set net is estimated to be 1:1 (Ueta and Jo 1989). Precise statistical data for the total landing of S. lessoniana from all Japanese waters are not available. Ueta (1992) estimated the annual S. lessoniana catch per year in the coastal waters of Tokushima Prefecture, where S. lessoniana is abundant, to be 37–57 t by set net and 8–24 t by jigging (resource from 1985 to 1988). Based on 13 years of survey data (resource from 1984 to 1996), Ueta (2000a) estimated the annual catch per year for this squid in Tokushima Prefecture to be 79.1 ± 44.1 t.
Because squid caught by set nets can be kept alive and in good condition in net cages, set nets are suitable for supplying live specimens, at a relatively high price, to traditional Japanese restaurants and sushi bars. Squid caught by set net are also suitable for broodstock maintenance and aquaculture. Ikeda et al. (2004) transported young and subadult S. lessoniana from set nets and reared them as the origin for culture trials. The approximately 740-km transfer took 22–23 h, and both water quality and temperature had decreased by arrival (pH < 7.0, NH4-N 5.0 mg·L−1, temperature > 4 °C). Squid survival is high when the BW to seawater volume ratio is below 30 at shipping. Survival is also relatively high in large containers, such as Styrofoam boxes with large (> 1,620 cm2) bottom areas. An acclimation period of suitable duration is necessary when transported squid are moved to new aquaria. Squid with a short acclimation (< 1 h) period die soon after being transferred to the aquaria, but those with a longer acclimation (> 2 h) can survive (e.g. shipping container water differs from aquarium water by 5 °C and 1.2 pH units).
Set nets also act as a spawning bed for S. lessoniana. Therefore, it is easy to get freshly laid egg capsules for use in aquaculture. Three types of artificial S. lessoniana spawning beds were examined in the spawning grounds in the coastal waters of Tokushima in Japan (Ueta et al. 1995; Ueta and Kitakado 1996). These were fibreglass-reinforced plastic (FRP) type, mid-water cage type, and steel type. The FRP type consisted of 9- or 12-mm-diameter FRP that were spaced 5, 10, 15, or 20 cm apart in 1 × 1 × 0.5 m concrete blocks. The mid-water cage type consisted of 0.5-, 1-, or 2-m-high cylindrical cages with a 9-mm rope that was stretched between the walls, which were linked to three other cages, a float, and a concrete block. The steel type consisted of 49 vertical and 0–6 horizontal 19-mm steel poles fixed to a 1 × 1 × 0.5 m concrete block (Ueta et al. 1995; Ueta and Kitakado 1996). Egg capsules were laid by S. lessoniana on these artificial spawning beds with the number of egg capsules laid on FRP type > mid-water cage type > steel type. However, the differences between the numbers of eggs laid on each spawning bed type were small. When using the FRP-type spawning bed, female squid spawn many more egg capsules on poles set close together than on poles that are widely spaced. Furthermore, female squid prefer spawning on these artificial spawning beds to natural spawning beds composed of Sargassum and Zostera.
Late-stage egg capsules can be collected in order to decrease the necessary incubation time and to avoid cessation of embryonic development at earlier stages in captivity. However, special care must be taken when late-stage egg capsules are collected because accidental pre-hatching often occurs due to physical shock during transportation.
The availability of artificially fertilized eggs would be convenient for both culture programs and experimental studies. Techniques for artificial insemination have not yet been established for loliginid squid. These squid have relatively large ova that are wrapped in a thick gelatinous layer (i.e. egg capsule) during embryonic development. Artificial insemination of the ommastrephid squid Todarodes pacificus is successfully performed by the addition of oviducal gland jelly at, and after, insemination of ova from the oviduct with spermatozoa from the seminal receptacle (Ikeda et al. 1993). The gelatinous substance of oviducal gland origin is necessary for formation of the perivitelline space that allows embryonic development to proceed normally. Oviducal gland jelly would act similarly in S. lessoniana.
3.2.2 Nursing of Egg Capsules
3.2.2.1 Egg-Capsule Characteristics
Structural abnormality of spawned egg capsules occurs in later generations of cultured S. lessoniana (Ikeda et al. 2009a). The outer gelatinous coat appears to be normal but the arrangement of eggs in the capsules is abnormal. The ova connect at one end of the egg capsule (Fig. 17.7). Most of the abnormally arranged eggs do not develop, which results in a low rate of hatching. Walsh et al. (2002) also reported a decrease in egg viability in their cultured squid. Abnormalities, such as the abnormal arrangement of eggs in egg capsules, may be a consequence of inbreeding.

Egg capsules spawned by cultured Sepioteuthis lessoniana, with arrows indicating ova. Note that one end of the egg capsule is widened due to the abnormal distribution of ova. (From Ikeda et al. 2009a)
The number of ova contained in a single S. lessoniana egg capsule varies in temperate waters . For example, egg capsules are reported to contain the following number of ova: 2–8 (average 3.0 and 5.8) at Iriomote Island of the Ryukyu Archipelago (Tsuchiya 1981), 2 and 4–8 (mode 6) at Ishigaki Island of the Ryukyu Archipelago (Segawa et al. 1993b), 0–9 (mean 3.5–6.4) at Kominato of Chiba Prefecture (Segawa 1987), and 0–9 (mean 5.5) at Kii Channel in Tokushima Prefecture (Ueta et al. 1992). Ueta et al. (1992) provided the following equation for a natural population of S. lessoniana that describes the relationship between the average number of ova in a single egg capsule (X) and the number of ova per egg mass (Y):
3.2.2.2 Water Quality and System Requirements
If large-capacity tanks that have enough space for floating several baskets cannot be provided, egg capsules can be separated and suspended from Styrofoam pillars that are 2 cm in diameter. Each egg capsule then freely floats on the surface layer of a smaller circular tank (30 cm in diameter, 27 cm in depth, 20 L; Ikeda et al. 2005) or a large circular tank (70 cm in diameter, 34 cm in depth, 120 L; Ikeda and Sugimoto 2013; Fig. 17.8). With this method, every surface of the egg capsules is completely exposed to water. This allows for sufficient oxygen exchange with the eggs, and gentle flow continuously cleans the capsule surface.

Lee et al. (1994); Forsythe et al. (2001), and Walsh et al. (2002; all at the Natural Resource Center for Cephalopods, Marine Biomedical Institute, University of Texas Medical Branch, USA) used 1.8-m-diameter cylindrical tanks for egg nursing in a closed water system. The tanks were supplied by fresh natural seawater drawn through an under-gravel filter and pumped back to the tanks after passing through a particle filter, an activated carbon filter, and two 30-W ultraviolet (UV) sterilizers. The concentration of NH4-N, NO2-N, and NO3-N was maintained under 0.10, 0.10, and 50 mg·L−1, respectively (Walsh et al. 2002). Ikeda et al. (2003, 2009a) circulated the water from 20- and 50-L tanks through coral gravel filters and UV sterilizers. Another option involves pumping water into eight cubic tanks used for growing macro-algae before gravity-feeding the water back to the main tank.
3.2.2.3 Hatching Event
Variation in the size of hatchlings occurs due to different hatching times. Those that hatch early tend to be smaller than those that hatch later (Ikeda et al. 1999). Smaller, early hatchlings are less able to move in order to feed or escape than their later-hatching counterparts. Special care must be taken for these early-hatched small juveniles.
3.2.3 Nursing of Young
3.2.3.1 Characters and Habit of Young
Compared to other loliginids, S. lessoniana hatchlings are much larger, with a 3.5–6.4-mm ML (Lee et al. 1994). Hatching usually occurs at midnight and seldom occurs during daylight hours. After hatching, hatchlings (of ML 5–7 mm) swim gently to maintain their position in the middle to upper water layer, keeping their mantle upward and head obliquely downward (Segawa 1987). Hatchlings also assume an upward or downward V-shaped position, while body colour is kept either all dark or transparent. In the upward V-shaped position, the arms are divided into two rows, like the letter ‘V’, while in the downward V-shaped position, all arms droop downward (Ikeda, unpublished observation). Hatchlings show positive phototaxis to a beam of light at night (Segawa 1987). Because a similar behaviour is also observed for hatchlings of Heterololigo bleekeri, another loliginid squid (Ikeda et al. 2005), this may be a shared characteristic of loliginids at hatching. This characteristic likely contributes to keeping hatchlings at the surface layer where they are most likely to encounter prey organisms and/or avoid predators.
The young begin to form schools in which individuals are positioned parallel to their neighbours, and schools are completely formed within 2 months of hatching (Sugimoto and Ikeda 2012).
3.2.3.2 Water Quality and System Requirements
In closed water systems, nursing of the young proceeds in the 20- or 50-L cylindrical egg-nursing tanks at a temperature of about 23–24 °C (Ikeda et al. 2003, 2009a; Lee et al. 1994, 1998a, b). Such small tanks are advantageous in that prey organisms aggregate in them and they are convenient for aquaculturists to manage. In addition, less seawater is required for closed systems than for open systems and they are therefore economical. A 120-L circular tank can also be used to rear juveniles and the young in a closed system; this relatively large tank enables the observation of squid behaviours that require space (e.g. schooling and social interaction).
3.2.3.3 Feeding
Various species of mysids can be successfully provided as prey, e.g. the brackish water crustaceans Neomysis japonica (Choe and Ohshima 1961; Ohshima and Choe 1961; Choe (1966a, b), Siriella longipes (Segawa 1990), and Mysidopsis almyra (Lee et al. 1994). Other potential live feed organisms include small-sized or freshwater fish fry, e.g. guppy, Poecilia reticulata, or medaka, Oryzias latipes (Lee et al. 1994; Ikeda et al. 2003, 2009a).
Juvenile squid older than 1 month of age can adapt to feeding on dead organisms that are thrown by humans into the water column. At first, squid neglect the dead diet when caretakers throw it into the tank, but after repeated exposure, they begin to pay attention to it and catch it immediately. Juvenile squid will also seize dead diet that is provided by caretakers with forceps. This method ensures that all squid are fed, even when individuals do not hunt food that is thrown in the water column. Providing the diet ad libitum can avoid starvation and cannibalism and is important for culturing squid successfully.
3.2.3.4 Growth
A large decrease in survivorship occurs in cultures within 30 days after hatching (Lee et al. 1994; Ikeda et al. 2003, 2009a). The most likely reason for the high death rate of S. lessoniana, as for other squids, during the early phase of life is starvation as a result of insufficient feeding. Hatchlings of S. lessoniana consume both inner yolk and live dietary specimens for up to 1 week (Shimazoe 2010). Hatchlings may learn by trial and error how to catch food during this phase, as do other loliginid squid (Chen et al. 1996).
3.2.4 Ongrowing
3.2.4.1 Feeding
Because a live diet of marine organisms (e.g. zooplankton and fish larvae) is only obtainable by laboratories situated near oceans, dead diets provide a suitable and economical alternative for inland squid cultures. Young S. lessoniana are fed on a mixture of live and dead frozen prey items, including Japanese sardine Sardinops sp. and Japanese anchovy Engraulis sp. (Ikeda et al. 2003, 2009a). Subadult and adult squid can be fed on a variety of prey items, either alive or dead. Lee et al. (1994) estimated the feeding rate of captive S. lessoniana (60–300 days of age) in a closed water system to be 20–35 % BW·day−1.
3.2.4.2 Growth and Survival
Fins of S. lessoniana hatchlings are subterminal, small, and round. However, they elongate with growth. Segawa (1987) used the following equation to describe the relationship between FL and ML for squid with ML 2.6–206 mm:
Kanamaru and Itoh (1996) reported that the ratio of FL to the length of the mantle margin reaches an adult-like value when S. lessoniana grow to 50–60 mm in ML.
Segawa (1987) used the following equation to describe the relationship between BW and ML for S. lessoniana with ML 5.7–128.2 mm cultured in open water systems (> 20 °C):
Lee et al. (1994) estimated the instantaneous growth rate of cultured S. lessoniana (temperature of 23.2 ± 0.6 °C) to be 3.9–5.3 BW·day−1 for the entire life span and 8.2–12.0 BW·day−1 for the first 100 days after hatching in closed water systems. Forsythe et al. (2001) examined the effect of temperature on the growth of cultured S. lessoniana in a closed water system. They estimated that squid grown at approximately 27 °C attained a size of 10 g in as little as 45 days at a sustained growth rate of 12.2 % BW·day−1, while squid cultured at 20 °C required almost 100 days to attain the same size at a rate of 5.7 % BW·day−1. Kanamaru and Itoh (1996) did not find any difference in the growth of females and males with ML 45–160 mm. On the other hand, Ueta (2000a) used the following equation to describe the relationship between BW and ML for a natural population of S. lessoniana with ML 7–470 mm in Japanese waters:
Survival of squid older than 30 days does not often change drastically, but intermittent death from unknown causes sometimes occurs in closed water systems. Between 70 and 140 days after hatching, only 1–2 of 25 squid die daily, and those that die are always the smallest in the rearing population (Fig. 17.9; Sugimoto and Ikeda 2013). In addition to the size variation that occurs at hatching, much greater size variation occurs in culture tanks 2 months after hatching (Sugimoto and Ikeda 2013; Fig. 17.10). At 60 days post hatching, the largest squid may be more than three times the size of the smallest squid. Whether this difference is genetic or environmental is unknown, and size differences may become much larger in later phases of life under both cultured and wild conditions (Fig. 17.10; Ikeda et al. 2009a). Because smaller squid often become the target of cannibalism (Segawa 1993), selective ongrowing of squid with similar-sized squid in the same tank may contribute to successful culture.

Survivorship of Sepioteuthis lessoniana in culture conditions. Triangles indicate survival rate; diamonds indicate dead individuals; circles and bars indicate the average and standard deviation of dorsal mantle length (ML). (Sugimoto and Ikeda 2013)

Size variation of Sepioteuthis lessoniana during the post-hatching phase in culture conditions. Circles and bars indicate the average and standard deviation of dorsal mantle length (ML), respectively. (Sugimoto and Ikeda 2013)
Kanamaru and Itoh (1996) found that it is possible to completely determine the sex of S. lessoniana based on characteristic colour patterns on the dorsal mantle (for squid with ML > 165 mm) and the hectocotylized fourth left arm of males (for squid with ML > 85 mm). These criteria can be used to regulate the number of female and male squid in a single tank. Because male squid often become aggressive and engage in agonistic contests after they reach maturity, maintaining a suitable balance of males and females (e.g. male < female) leads to successful reproduction in the tank.
S. lessoniana are estimated to live for about 1 year in Japanese waters (Ueta and Jo 1989; Ueta 2000b). In general, cephalopods reach maturity earlier in captive conditions than in the wild (Mangold 1987). This is also true for S. lessoniana. Tsuchiya (1982), who first cultured S. lessoniana in an open water system (temperature of 20.81–29.89 °C), observed spawning at 136 days post hatching. Ikeda et al. (2003, 2009a) also observed precocious maturation of cultured squid (temperature of 21.1–23.8 °C; Fig. 17.11). They observed that males reached maturity at 140 days of age, 3–8 months earlier than wild-caught squid of the same generation, and initiated copulation repeatedly. This is also similar for females. Cultured squid begin to mature at least 2 months earlier than wild-caught squid. The constant, relatively high temperature in captivity may accelerate maturation. Female squid spawn 160–196 days post hatching. Sometimes, a single female will spawn more than twice within 40–48 days. Intermittent spawning by a single female was also observed by Wada and Kobayashi (1995) and Ueta (2000b) in an open water system. Both males and females die restively after repeated copulation and spawning. The life span of cultured S. lessoniana in closed water systems is reported to be 4–6 months (Lee et al. 1994), 220–300 days (Forsythe et al. 2001), 169–262 days (Walsh et al. 2002), and 161–315 days (Ikeda et al. 2009a). Because cephalopod life spans may be controlled by programmed death (Wodinsky 1977), and cephalopods are terminal spawners, the early maturation of cultured squid must also induce early death (i.e. short life span).

Temperature regime for Sepioteuthis lessoniana measured in captivity (solid line) and estimated in nature (dotted line), the progress of dorsal mantle length (ML), female gonad somatic index (GSI), and male testis index (TI) of cultured and wild-caught squid by date (X, cultured squid of unknown sex; +, cultured squid; W, wild-caught squid). Data for wild-caught squid are expressed as means with maximum and minimum values. Lines in the ML graph show hypothesized growth lines. (From Ikeda et al. 2009a)
Mating pairs touch their newly laid egg capsules and stay near them to guard them from other pairs (Fig. 17.12). On average, mating pairs touch their eggs for approximately 6 s and guard them for 45 s. Similar behaviour is observed in both cultured (Lee et al. 1994) and wild squid (Segawa 1987).

Egg-touching behaviour by cultured Sepioteuthis lessoniana. a Male partner touches egg capsules. b Male partner stays near egg capsules to guard against other squid. (Ikeda, original observation)
Cultured female squid have larger genital organs than wild-caught squid (Ikeda et al. 2009a). This is because captive females store a larger number of ripe ova in their oviducts, presumably due to inhibition of spawning. Spawning may be inhibited by particular environmental factors, such as strong lighting (Fig. 17.11). Wild females store 55–303 ripe ova, whereas cultured females store 4–2,096 ripe ova (Ikeda et al. 2009a). Mature females may be motivated to spawn, even without mating and without stored spermatangia and spermatozoa (Ikeda et al. 2009b).
3.2.4.3 Water Quality and System Requirements
Closed systems used for the ongrowing phase have filtration systems similar to those used for nursing the young, but use tanks of larger size. Lee et al. (1994, 1998a, b) and Walsh et al. (2002) used a 15,000-L raceway (2.4 × 6.1 × 0.9 m) and a 50,000-L circular tank (6.5 m circular line, 1.75 m depth) filled with artificial seawater. Juvenile squid of 30–40 mm ML were transferred from their nursing tanks to these ongrowing tanks . The ongrowing density of this system was 5.6 individuals·m−3. An average water temperature of 23 °C is a major factor that can extend the life span and the final size of cultured squid. After about 300 days, squid are 214–260 mm in length and 682–1,073 g in weight. Six generations have been cultured using this system. Ikeda et al. (2003, 2009a) used a large circular tank with a 10,000-L capacity (4 m diameter, 1 m water depth) connected to an algae tank (Fig. 17.8) and smaller 20-, 50-, and 1,700-L-capacity tanks. The squid grew to 268 mm in length 243 days after hatching (20–23 °C). Three consecutive generations were cultured using this system. Lee et al. (1998b) suggested that, in a closed system, removal of ink (ejected by frightened squid) was an important consideration peculiar to filtration systems.
Closed water systems maintained at 21–26 °C have been used to successfully culture three to seven consecutive generations of S. lessoniana (Lee et al. 1994; Walsh et al. 2002; Ikeda et al. 2009a; Fig. 17.11). Ueta (2000b) reported that the mortality rate in a long-term rearing experiment of S. lessoniana using an open water system increased when the temperature fell below 20 °C. In the wild, lower temperatures (15 °C) affect the movement and distribution of S. lessoniana (Ueta and Jo 1990). Therefore, temperatures should be maintained between 20 and 30 °C for S. lessoniana culture.
Both natural light and artificial light are used for the culture of S. lessoniana in open and closed water systems (Tsuchiya 1982; Lee et al. 1994; Walsh et al. 2002; Ikeda et al. 2003, 2009a). In closed water systems, fluorescent and metal halide bulbs are used for illumination. Young S. lessoniana generally avoid both the brightest part (85 Lx) and darkest part (5 Lx) of the tank (Lee et al. 1994), whereas adult squid avoid the brightest areas of tanks (Ikeda, unpublished observation).
The required physical and chemical parameters of the seawater used for squid culture are, on average, pH 7.9–8.2, NH4-N 0.009–0.044 mg·L−1, NO2-N 0.023–0.069 mg·L−1, and NO3-N 9.6–36.0 mg·L−1 (Walsh et al. 2002) or pH 7.8 ± 0.2, ammonia 4.7 ± 11.7 mg·L−1, nitrite 0.3 mg·L−1, nitrate 114 ± 73 mg·L−1, and salinity 35.8 ± 1.2 psu (Ikeda et al. 2009a). As with other loliginids, ammonia concentrations that are too high cause sudden death of S. lessoniana. In addition, drastic changes in water quality and temperature can also cause immediate squid death.
4 Trends in Research and Industrial Level
One of the main focuses of future research should include the development of feed for large-scale culture, with live and artificial feed that are similar to that used for the cuttlefish, Sepia pharaonis and Sepiella inermis (see also Chaps. 12 and 13). Problems such as the cost and supply of live feed and the availability of an acceptable form of artificial feed are still encountered (Nabhitabhata et al. 2001b; Sangpradab et al. 1984).
Nursing and ongrowing in net cages and pens in situ should be studied as a means of reducing the cost of S. lessoniana culture. At selected sites, such facilities could substitute for land-based hatcheries. In tropical countries, such facilities must be protected from strong sunlight, as excess lighting causes stress to cultured squid, fouling of egg capsules, and fouling of cage cover nets.
Monosex culture may be a method for postponing reproduction and prolonging squid growth. To accomplish this, squid must be separated by sex before proceeding to, or as early as possible in, the ongrowing phase. The sex of S. lessoniana can be determined from their colour patterns 60 days after hatching (Nabhitabhata 1978, 1983).
Manipulation of the sex ratio or artificial sex induction for S. lessoniana can increase production. The degree of sexual size dimorphism is high. S. lessoniana males are much larger than females. Batch cultures with many large males will produce a higher yield than cultures with few large males. Manipulation of the development of the male external characters through the application of sex hormones, e.g. testosterone, is feasible. The methodology for this type of manipulation can be adapted from that which is already used in the finfish culture industry for tilapia (Oreochromis niloticus) and groupers (Epinephelus spp.). For example, developing S. lessoniana egg capsules can be immersed in water that contains a particular hormone and/or the young can be fed hormone-added feed. These methods need further study.
S. lessoniana is a complex species (Dunning 1998) comprised of at least three morphs with different final sizes. Segawa et al. (1993a, b) reported morphological, behavioural, and ecological differences among three morphs of S. lessoniana with the local names shiro-ika (white squid), aaka-ika (red squid), and kuwa-ika (small squid) from Ishigaki Island of the Ryukyu Archipelago, Japan. The three morphs have different final sizes, chromatophore arrangements, egg-capsule characteristics, spawning sites, and seasons (Izuka et al. 1996b; Segawa et al. 1993a, b). Molecular evidence indicates genetic differences between the three morphs (Izuka et al. 1994, 1996a; Yokokawa and Ueta 2000; Triantafillos and Adams 2005; Aoki et al. 2008). Further investigation and taxonomic description are required to determine whether these differences are at the specific or subspecific level. Differences in the growth rate of different morphs should be a very interesting research topic for the maximization of aquaculture production.
Although S. lessoniana is widely distributed in the Indo-Pacific region, collection sites for materials (i.e. eggs) used for previous culture or long-term rearing are predominantly located in Asian waters. For example, all of the S. lessoniana eggs used by Lee et al. (1994); Forsythe et al. (2001), and Walsh et al. (2002) were collected from Asian waters, i.e. Thailand, Mainland Japan, and Okinawajima Island of Japan. These Asian countries have major fisheries for this species (Ueta et al. 1992, 2000a; Chotiyaputta et al. 2002; Lu 2002). These countries are ideal locations for studying the culture of S. lessoniana. Aquaculture facilities for cephalopods and other marine organisms are already established in Asian countries. Squid cultured in such facilities can be used as scientific experimental models, species for restocking , and as human food. The network of S. lessoniana culture to be established in Asian countries is expected to form the future cephalopod culture centre of the world.
5 Conclusions
Sepioteuthis lessoniana is highly adaptable to captive conditions and can be cultured for multiple generations in either open or closed seawater systems . This success is mainly due to the relatively large hatchling size of S. lessoniana and behavioural characteristics that are advantageous for limited tank space. This is an advantage not only for industrial culturing per se but also for other purposes, such as providing this squid for restocking and as an experimental animal for ethology, physiology, and neuroscience for which live material is usually necessary. On the other hand, similar to other cephalopods, the life history of S. lessoniana has not been entirely documented in either tropical or temperate waters . This includes the reproductive process, survival after hatching, and population genetics. Modern techniques, such as bio-logging and DNA markers, are expected to provide new information about these unresolved questions. In addition, because the fisheries’ demand for S. lessoniana is high, policies for the protection of this squid must be established.
The process of S. lessoniana culture includes broodstock collection , nursing of egg capsules, nursing of the young in the hatchery, and the grow-out phase. Water quality control procedures are different in closed and open seawater systems and are fully controlled in the former and partially controlled in the latter. The young are fed from hatching with live prey organisms, particularly mysids. The size of prey has to be similar to that of the squid. Sufficient supply of live feed ensures good survival. The young can be trained to accept dead feed, starting in the grow-out phase. The growth rate, final size, and longevity of these squid can vary, particularly with temperature and seawater system. Research and development of artificial feed is urgently required in order to resolve the live feed bottleneck and to reduce the cost of production.
References
Ahmad T, Usman (1997) Bigfin squid culture: the Indonesian experience. Phuket Mar Biol Cent Spec Pub 17:285–287
Aoki M, Imai H, Naruse T, Ikeda Y (2008) Low genetic diversity of oval squid Sepioteuthis lessoniana (Cephalopoda: Loliginidae), in Japanese waters inferred from a mitochondrial DNA non-coding region. Pac Sci 62:403–411
Boongerd S, Rachaniyom S (1990) Squid trap fishing. Tech Pap 1/1990, Fish Tech Subdiv, Mar Fish Div, Dept Fish
Chainak P, Chindamaikul T, Nabhitabhata J, Kwandee V (2003) Effect of phosphate concentration on survival of hatchlings of bigfin squid Sepioteuthis lessoniana Lesson, 1830. In: Nabhitabhata J (ed) Report on cephalopod culture research and development programme, 2002–2004. Rayong Coast Fish Res Dev Cent Contrib 27
Chankaew K, Nabhitabhata J, Sanein N, Pattanakul S (2003a) Life history and management of pygmy goby as live feed for nursing of young bigfin squid. In: Nabhitabhata J (ed) Report on cephalopod culture research and development programme, 2002–2004. Rayong Coast Fish Res Dev Cent Contrib 27
Chankaew K, Nabhitabhata J, Sanein N, Pattanakul S (2003b) Management of sea bass fries as live feed of hatchlings of bigfin squid and pharaoh cuttlefish. In: Nabhitabhata J (ed) Report on cephalopod culture research and development programme, 2002–2004. Rayong Coast Fish Res Dev Cent Contrib 27
Chen DS, Dykhuizen GV, Hodge J, Gilly WF (1996) Ontogeny of copepod predation in juvenile squid (Loligo opalescens). Biol Bull 190:69–81
Chindamaikul T, Nabhitabhata J, Krainara T, Somsueb S (1994a) Survival of bigfin squid, Sepioteuthis lessoniana Lesson, in different stocking density. Tech Pap 30/1994, Phuket Coast Aquacult Dev Cent, Coast Aquacult Div, Dept Fish
Chindamaikul T, Nabhitabhata J, Somsueb S, Krainara T (1994b) Comparison on different methods for hatching of bigfin squid, Sepioteuthis lessoniana. Tech Pap 7/1994, Phuket Coast Aquacult Dev Cent, Coast Aquacult Div, Dept Fish
Chindamaikul T, Kaoian K, Chainak P, Nabhitabhata J, Samrarnrat N (2001) Effect of nitrite-nitrogen and ammonia-nitrogen on survival of hatchlings of bigfin squid, Sepioteuthis lessoniana Lesson, 1830. In: Nabhitabhata J (ed) Report on cephalopod culture research and development programme, 2002–2004. Rayong Coast Fish Res Dev Cent Contrib 27
Choe S (1966a) On the egg, rearing, habits of the fry, and growth of some cephalopoda. Bull Mar Sci 16:330–348
Choe S (1966b) On the growth, feeding rates and the efficiency of food conversion for cuttlefishes and squids. Korea J Zool 9:12–20
Choe S, Ohshima Y (1961) On the embryonal development and the growth of the squid, Sepioteuthis lessoniana Lesson. Venus 21:462–476
Choe S, Ohshima Y (1963) Rearing of cuttlefishes and squids. Nature 197:307
Chotiyaputta C (1984) Spawning season of Sepioteuthis lessoniana. Ann Rep 1984, Invertebr Sect, Mar Fish Div, Dept Fish
Chotiyaputta C (1988) Reproductive biology of bigfin reef squid from squid trap. Tech Pap 5, Mar Life Hist Sect, Mar fish Div, Dept Fish
Chotiyaputta C (1995) Biology of cephalopods. In: Nabhitabhata J (ed) Biology and culture of cephalopods. Rayong Coast Aquacult St, Rayong
Chotiyaputta C, Yamrungreung A (1998) Trap fisheries for squid and their impact on spawning. S Afr J Mar Sci 20:285–291
Chotiyaputta C, Nootmorn P, Jirapunpipat K (2002) Review of cephalopod fishery production and long term changes in fish communities in the Gulf of Thailand. Bull Mar Sci 71:223–238
Dunning MC (1998) Loliginidae inshore squids, pencil squids. In: Carpenter KE, Niem VH (eds) The living marine resources of the western central Pacific vol 2 cephalopods, crustaceans, holothurians and sharks. Food and Agriculture Organization of the United Nations, Rome
Forsythe JW, Walsh LS, Turk PE, Lee PG (2001) Impact of temperature on juvenile growth and age at first egg-laying of the Pacific reef squid Sepioteuthis lessoniana reared in captivity. Mar Biol 138:103–112
Ikeda Y, Sakurai Y, Shimazaki K (1993) Fertilizing capacity of squid (Todarodes pacificus) spermatozoa collected from various sperm storage sites, with special reference to the role of gelatinous substance from oviducal gland in fertilization and embryonic development. Invertebr Reprod Dev 23:39–44
Ikeda Y, Wada Y, Arai N, Sakamoto W (1999) Note on size variation of body and statolith in the oval squid Sepioteuthis lessoniana hatchlings. J Mar Biol Assoc UK 79:757–759
Ikeda Y, Sakurazawa I, Sakurai Y, Matsumoto G (2003) Initials trials of squid rearing, maintenance and culture at the Brain Science Institute of RIKEN. Suisanzoshoku 51:391–400
Ikeda Y, Ueta Y, Sakurazawa I, Matsumoto G (2004) Transport of the oval squid Sepioteuthis lessoniana Férussac, 1831 in Lesson 1830–1831 (Cephalopoda: Loliginidae) for up to 24 h and subsequent transfer to an aquarium. Fish Sci 70:21–27
Ikeda Y, Sakurazawa I, Ito K, Sakurai Y, Matsumoto G (2005) Rearing of squid hatchlings, Heterololigo bleekeri (Keferstein 1866) up to 2 months in a closed seawater system. Aquac Res 36:409–412
Ikeda Y, Ueta Y, Anderson FE, Matsumoto G (2009a) Reproduction and life span of the oval squid Sepioteuthis lessoniana (Cephalopoda: Loliginidae): comparison between laboratory-cultured and wild-caught squid. Mar Biodiversity Rec 2:e50. doi:10.1017/S175526720900061X
Ikeda Y, Oshima Y, Sugimoto C, Imai H (2009b) Multiple spawning by uncopulated oval squid (Sepioteuthis lessoniana) in captivity. Aquac Sci 57:39–43
Izuka T, Segawa S, Okutani T, Numachi K (1994) Evidence on the existence of three species in the oval squid, Sepioteuthis lessoniana complex in Ishigaki Island, Okinawa, South-western Japan, by isozyme analyses. Venus 53:217–228
Izuka T, Segawa S, Okutani T (1996a) Biochemical study of the population heterogeneity and distribution of the oval Squid Sepioteuthis lessoniana complex in southwestern Japan. Amer Malac Bull 12:129–135
Izuka T, Segawa S, Okutani T (1996b) Identification of three species in oval squid, Sepioteuthis lessoniana complex by chromatophore arrangements on the funnel. Venus 55:139–142
Jereb P, Vecchione M, Roper CFE (2010) Family Loliginidae. In: Jereb P, Roper CFE (eds) Cephalopods of the world an annotated and illustrated catalogue of cephalopod species known to date vol. 2 myopsid and oegopsid squids FAO Species Catalogue for Fishery Purposes No. 4, Vol. 2. FAO, Rome, pp 38–117
Kanamaru H, Itoh Y (1996) Development of fin base and appearance of secondary sexual characters in juvenile and young of the oval squid Sepioteuthis lessonina. Saibai Giken 25:15–20
Lee PG, Turk PE, Yang WT, Hanlon RT (1994) Biological characteristics and biomedical applications of the squid Sepioteuthis lessoniana cultured through multiple generations. Biol Bull 186:328–341
Lee PG, Turk PE, Forsythe JW, DiMarco FP (1998a) Cephalopod culture: physiological, behavioral and environmental requirements. Suisanzoshoku 46:417–422
Lee PG, Walsh L, Turk PE, DiMarco FP (1998b) Large-scale culture of the loliginid squid, Sepioteuthis lessoniana. In: Tiensongrasmee B, Tedengren M, Jarayabhand P, Tandavanitj N, Popongviwat A, Nujsawat A, Soisodsri S (eds) Mollusk research in Asia. Thailand Research Fund, Bangkok
Lu CC (2002) Cephalopod production from the waters around Taiwan. Bull Mar Sci 71:465–471
Mangold K (1987) Reproduction. In: Boyle PR (ed) Cephalopod life cycles vol. 2. Academic Press, London
Munekiyo M, Kawagishi M (1993) Diurnal behaviors of the oval squid, Sepioteuthis lessoniana, and fishing strategy of a small-sized set net (preliminary report). In: Okutani T, O’Dor RK, Kubodera T (eds) Recent advances in cephalopod fisheries biology. Tokai University Press, Tokyo
Nabhitabhata J (1978) Rearing experiment on economic cephalopod -I: long-finned squid Sepioteuthis lessoniana Lesson. Tech Pap 1978, Rayong Brackish Fish Sect, Rayong Fish Stn, BrackishFish Div, Dept Fish
Nabhitabhata J (1983) External sex characteristics and sexual activities of long-finned squid, Sepioteuthis lessoniana Lesson. Tech Pap 1/1983, Rayong Brackish Fish Stn, Brackish Fish Div, Dept Fish
Nabhitabhata J (1993) Concept, materials and method of cephalopod culture. Contrib 6, Rayong Coast Aquacult Stn, Coast Aquacult Div, Dept Fish
Nabhitabhata J (1996) Life cycle of cultured bigfin squid, Sepioteuthis lessoniana Lesson. Phuket Mar Biol Cent Spec Pub 16:83–95
Nabhitabhata J (2002) Life history and behaviour of reared neritic cephalopods from Thai Waters. PhD Thesis, Tokyo University of Fisheries
Nabhitabhata J, Kbinrum S (1981) The culture of long-finned squid, Sepioteuthis lessoniana Lesson. Rayong Brackish Fish Stn, Brackish Fish Div, Dept Fish, Ann Rep 1981:74–115
Nabhitabhata J, Kbinrum S (1984) Biological studies on economic cephalopods-III: note on survival of young cephalopods during continuous change of salinity. Tech Pap 6/1984, Rayong Brackish Fish Stn, Brackish Fish Div, Dept Fish
Nabhitabhata J, Kbinrum S, Kumpang P (1984) On floating net cage culture of bigfin squid, Sepioteuthis lessoniana Lesson, at different stocking densities. Tech Pap 7/1985, Rayong Brackish Fish Stn, Brackish Fish Div, Dept Fish
Nabhitabhata J, Kbinrum S, Soodmee Y (1991a) Effect of pH on survival rate of cephalopods hatchlings. Tech Pap 17/1991, Rayong Coast Aquacult Stn, Coast Aquacult Div, Dept Fish
Nabhitabhata J, Kbinrum S, Soodmee Y (1991b) Effect of salinity on hatching rate of Sepioteuthis lessoniana Lesson. Tech Pap 16/1991, Rayong Coast Aquacult Stn, Coast Aquacult Div, Dept Fish
Nabhitabhata J, Soodmee Y, Petrasatian K (1992a) Effect of salinity on survival rate of bigfin squid hatchlings, Sepioteuthis lessoniana Lesson. Tech Pap 3/1992, Rayong Coast Aquacult Stn, Coast Aquacult Div, Dept Fish
Nabhitabhata J, Jaroenvittayakul Y, Kakhai N, Soodmee Y, Nilaphat P (1992b) On live feed supply management for seed production of bigfin squid, Sepioteuthis lessoniana Lesson, and cost of production. Tech Pap 13/1992, Rayong Coast Aquacult Stn, Coast Aquacult Div, Dept Fish
Nabhitabhata J, Kakhai N, Suriyawarakul J, Nilaphat P (1996) Feeding and cost of seed production of cephalopods, Sepioteuthis lessoniana Lesson; Sepia pharaonis Ehrenberg; Sepiella inermis Fer. & d’Orb. Tech Pap 1/1996, Rayong Coast Aquacult Stn, Coast Aquacult Div, Dept Fish
Nabhitabhata J, Nilaphat P, Promboon P (2001a) Artificial fertilization and embryonic development of bigfin squid, Sepioteuthis lessoniana Lesson, 1830. In: Nabhitabhata J (ed) Report of cephalopod culture research and development programme, 2002–2004. Rayong Coast Fish Res Dev Cent Contrib 27
Nabhitabhata J, Nilaphat P, Promboon P (2001b) Behavioural approaches on the acceptable form of artificial feed of bigfin squid, Sepioteuthis lessoniana Lesson, 1830. In: Nabhitabhata J (ed) Report of cephalopod culture research and development programme, 2002–2004. Rayong Coast Fish Res Dev Cent Contrib 27
Nabhitabhata J, Asawangkune P, Amornjaruchit S, Promboon P (2001c) Tolerance of eggs and hatchlings of neritic cephalopods to salinity changes. Phuket Mar Biol Cent Spec Pub 25:91–99
Nabhitabhata J, Nilaphat P, Promboon P, Jaroongpattananon C, Nilaphat G, Reunreng A (2005) Performance of simple large-scale cephalopod culture system in Thailand. Phuket Mar Biol Cent Res Bull 66:337–350
Ohshima Y, Choe S (1961) On the rearing of young cuttlefish and squid. Bull Japan Soc Sci Fish 27:978–986
Okutani T (2005) Cuttlefish and squid of the world. Seizando-Shoten Publishing Co., Ltd., Tokyo
Rattana-anant T (1978) Biological studies on Sepioteuthis lessoniana Lesson (Cephalopoda; Loliginidae) in the Gulf of Thailand. Invertebr Sect, Mar Fish Div, Dept Fish
Rattana-anant T (1979) Biological studies on Sepioteuthis lessoniana Lesson (Cephalopoda; Loliginidae) in the Gulf of Thailand. Invertebr Sect, Mar Fish Div, Dept Fish
Rattana-anant T (1980) Biological studies on Sepioteuthis lessoniana Lesson (Cephalopoda; Loliginidae) in the Gulf of Thailand. Invertebr Sect, Mar Fish Div, Dept Fish
Roongratri M (1997) Biology of bigfin reef squid (Sepioteuthis lessoniana) in the eastern coast of the Gulf of Thailand. Tech Pap 65, East Mar Fish Dev Cent, Mar Fish Div, Dept Fish
Sangpradab S, Veschasatra V, Kbinrum S (1984) Experiments on feeding of long-finned squid (Sepioteuthis lessoniana Lesson) with three different kinds of feed. Tech Pap 1/1984, Rayong Brackish Fish Stn, Brackish Fish Div, Dept Fish
Saso N (1979) On the culture of aori-ika, Sepioteuthis lessoniana. In: Japan Association of Fisheries Cooperatives (ed) Proc 25 Japan Nat Fish Activ Convn
SEAFDEC (1975) Observations on the culture and life history of the broad finned squid Sepioteuthis lessoniana Fer. and Orb. SEAFDEC-MSU-Inst Fish Res Develop Ann Rep 1975:115–119
Segawa S (1987) Life history of the oval squid, Sepiotuethis lessoniana in Kominato and adjacent waters Central Honshu, Japan. J Tokyo Univ Fish 74:67–105
Segawa S (1990) Food consumption, food conversion and growth rates of the oval squid Sepioteuthis lessoniana by laboratory experiments. Nippon Suisan Gakk 56:217–222
Segawa S (1993) Field and laboratory feeding studies of the Japanese oval squid, Sepioteuthis lessoniana. In: Okutani T, O’Dor RK, Kubodera T (eds) Recent advances in cephalopod fisheries biology. Tokai University Press, Tokyo
Segawa S, Hirayama S, Okutani. T (1993a) Is Sepioteuthis lessoniana in Okinawa a single species? In: Okutani T, O’Dor RK, Kubodera T (eds) Recent advances in cephalopod fisheries biology. Tokai University Press, Tokyo
Segawa S, Izuka T, Tamashiro T, Okutani T (1993b) A Note on mating and egg deposition by Sepioteuthis lessoniana in Ishigaki Island, Okinawa, southwestern Japan. Venus 52:101–108
Shimazoe K (2010) Post hatching cephalopod nutrition and growth in tropical waters. Master degree thesis, University of the Ryukyus
Sivalingam D, Ramadoss K, Gandhi AD, Enose M (1993) Hatchery rearing of the squid, Sepioteuthis lessoniana and the cuttlefish, Sepia pharaonis. Mar Fish Info Serv 122:12–14
Sivashanthini K, Charles GA, Thulasitha WS (2009) Length-weight relationship and growth pattern of Sepioteuthis lessoniana Lesson 1830 (Cephalopoda: Teuthida) from the Jaffna Lagoon, Sri Lanka. J Biol Sci 9:357–361
Sugimoto C, Ikeda Y (2012) Ontogeny of schooling behavior in the oval squid Sepioteuthis lessoniana. Fish Sci 78:287–294
Sugimoto C, Ikeda Y (2013) Comparison of the ontogeny of hunting behavior in pharaoh cuttlefish (Sepia pharaonis) and oval squid (Sepioteuthis lessoniana). Biol Bull 225(1):50–59
Supongpan M (1995) Cephalopod resources in the Gulf of Thailand. In: Nabhitabhata J (ed) Biology and culture of cephalopods. Rayong Coast Aquacult Stn, Rayong
Supongpan M, Kongmuag K, Jittrapong S (1988) Bigfin reef squid fisheries in the Gulf of Thailand. Tech Pap 1/1988, Mar Fish Div, Dept Fish
Triantafillos L, Adams. M (2005) Genetic evidence that the northern calamary, Sepioteuthis lessoniana, is a species complex in Australian Waters. ICES J Mar Sci 62:1665–1670
Tsuchiya M (1981) On the spawning of the squid, Sepioteuthis lessoniana Lesson at Amitori Bay, Iriomote Island, Okinawa. Inst Ocean Res Develop. Tokai Univ Note 3:53–75
Tsuchiya M (1982) On the culture of the squid Sepioteuthis lessoniana Lesson (Loliginidae), from hatching to the whole life in Iriomote Island, Okinawa. Inst Ocean Res Develop. Tokai Univ Note 4:49–70
Ueta Y (1992) Distribution on the oval squid, Sepioteuthis lessoniana and its fisheries around Tokushima Prefecture. Fish Biol Oceanogr SW Water Jpn 8:61–69
Ueta Y (2000a) Fishery biological studies of the oval squid, Sepioteuthis lessoniana around Tokushima Prefecture. Bull Tokushima Pref Fish Exp Stn 1:1–79
Ueta Y (2000b) Feeding and growth of the oval squid, Sepioteuthis lessoniana in captivity. Tokushima Suishi Jiho 125–132
Ueta Y, Jo Y (1989) Notes on ecology of the oval squid Sepioteuthis lessoniana in outer waters adjacent to the Kii Channel. Nippon Suisan Gakk 55:1699–1702
Ueta Y, Jo Y (1990) Migration of the oval squid, Sepioteuthis lessoniana around Tokushima Prefecture. Suisanzoshoku 38:221–226
Ueta Y, Kitagado I (1996) Experimental studies on the form of spawning bed by oval squid, Sepioteuthis lessoniana. Suisanzoshoku 44:67–72
Ueta Y, Segawa S, Tenjin M, Jo Y, Kitakado I, Fukunaga M, Kangawa Y (1992) Occurrence and character of egg masses and estimation of egg-laying and hatching period on oval squid, Sepioteuthis lessoniana in outer waters adjacent to the Kii Channel. Suisanzoshoku 40:469–474
Ueta Y, Kitakado I, Segawa S, Tenjin M, Jo Y, Fukunaga M, Kanagawa Y (1995) Selection of spawning ground and spawning bed in oval squid Sepioteuthis lessoniana. Fish Eng 31:189–194
Wada Y, Kobayashi T (1995) On an iteroparity of the oval squid Sepioteuthis lessoniana. Nippon Suisan Gakk 61:151–158
Walsh LS, Turk PE, Forsythe JW, Lee PG (2002) Mariculture of the loliginid squid Sepioteuthis lessoniana through seven successive generations. Aquaculture 212:245–262
Wodinsky J (1977) Hormonal inhibition of feeding and death in Octopus: control by optic gland secretion. Science 198:948–951
Yokokawa K, Ueta Y (2000) Genetic analysis of oval squid (Sepioteuthis lessoniana) around Japan. Venus 59:45–55
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2014 Springer Science+Business Media Dordrecht
About this chapter
Cite this chapter
Nabhitabhata, J., Ikeda, Y. (2014). Sepioteuthis lessoniana . In: Iglesias, J., Fuentes, L., Villanueva, R. (eds) Cephalopod Culture. Springer, Dordrecht. https://doi.org/10.1007/978-94-017-8648-5_17
Download citation
DOI: https://doi.org/10.1007/978-94-017-8648-5_17
Published:
Publisher Name: Springer, Dordrecht
Print ISBN: 978-94-017-8647-8
Online ISBN: 978-94-017-8648-5
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)