
Abstract
Contaminants may enter vegetables and fruits by several pathways: by uptake with soil pore water, by diffusion from soil or air, by deposition of soil or airborne particles, or by direct application. The contaminant-specific and plant-specific properties that determine the importance of these pathways are described in this chapter. A variety of models have been developed, specific for crop types and with steady-state or dynamic solutions. Model simulations can identify sensitive properties and relevant processes. Persistent, polar (log K OW < 3) and non-volatile (K AW < 10–6) contaminants have the highest potential for accumulation from soil, and concentrations in leaves may be several hundred times higher than in soil. However, for most contaminants the accumulation in vegetables or fruits is much lower. Lipophilic (log K OW > 3) contaminants are mainly transported to leaves by attached soil particles, or from air. Volatile contaminants have a low potential for accumulation because they quickly escape to air. Experimental data are listed that support these model predictions, but underline also the high variability of accumulation under field conditions. Plant uptake predictions are uncertain, due to the immense variation in environmental and plant physiological conditions. Uptake of organic contaminants into vegetables and fruits may lead to human health risks, but it may also be used to delineate subsurface plumes and monitor Natural Attenuation. Most models mentioned in this chapter are freely available from the authors.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
1 Introduction
A major concern of European citizens is residues of pesticides in fruits and vegetables (EFSA 2006). But not just pesticides pose a risk. In fact, for a whole range of organic contaminants, uptake via diet is often the primary contribution to human exposure (SCF 2002; Travis and Hattemer-Frey 1991). Contaminants may be taken up into plants and subsequently accumulate in the human food chain (Czub and McLachlan 2004) and affect the health of humans. Consequently, uptake of contaminants into plants is an essential part of most exposure models, for example CSOIL (Brand et al. 2007), CLEA (DEFRA 2002) and EUSES (EC 2003).
About 20,000 plant species are used by the human race, and about 600 species are cultivated (Franke 1987). Additionally, about 250,000 wild plants grow on earth (Sitte et al. 1991). Thus, there is a large variability in plant properties. Also the growth conditions vary, depending on soil type, soil properties, climatic conditions and agricultural practice. Similarly, the number of organic contaminants is very high. More than 5 million compounds have been synthesized. Around 30,000 compounds are marketed in Europe, and contaminants released to the environment may also be metabolized. Thus, from a researcher’s point of view, the number of possible combinations of plant species, contaminants and environmental conditions is close to infinite. Nevertheless, general patterns are known and process-oriented models have been established.
This chapter will give an overview of uptake processes of organic contaminants from soil into plants, on prediction methods and on experimental results. Model simulations will be carried out to identify the chemical properties that control the accumulation in food crops. These predictions will be compared with experimental results, in order to determine the potential of soil contaminants for accumulation in food crops. For the calculation of exposure through vegetable consumption, see Chapter 11 by Elert et al., of this book.
2 Uptake and Transport Processes
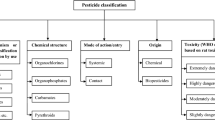
Contaminants in the environment can enter plants by various ways (Fig. 9.1). The main passive transport and uptake processes from soil are:
-
uptake with transpiration water;
-
diffusion from soil into roots;
-
attachment of soil particles, eventually followed by diffusion into plant tissue.

Transport and uptake processes in the soil-air-plant system
However, contaminants can also be present in air. The main uptake processes from air are:
-
diffusive (gaseous) exchange with air;
-
wet and dry particle deposition from air on plant surfaces followed by diffusion into plant tissue.
Inside the plant, the phloem and xylem flux may distribute the contaminants. The xylem sap flows from the roots into the stem to the leaves and, to some extent, to fruits. The phloem sap flows from the leaves to all growing parts of the plant and to fruits and storage organs, such as tubers. The relative importance of these processes varies with plant type, environmental conditions and properties of contaminants. Active uptake processes, which involve energy or enzymes of the plant, may also play a role, but have not yet been shown to be of large relevance for environmental organic contaminants.
3 Empirical Methods for Estimating Uptake of Contaminants into Plants
Laboratory and field experiments have been conducted to determine the uptake of contaminants from soil into plants. Single bioconcentration factors for contaminants as well as regressions were established for predicting the uptake of organic contaminants from soil into plants.
3.1 Bioconcentration Factors
The ratio of contaminant concentration in an organism to contaminant concentration in the surrounding medium is called the bioconcentration factor (BCF). Measurements of concentrations in plant tissues and concentrations in soil will yield a BCF plant to soil, which is defined as
where C Plant is the concentration in plant tissues and C Soil is the concentration in soil (ideally at steady state, but practically at harvest). This BCF will only apply to the specific contaminant and soil type used for the determination.
Care must be taken in cases where a measurable background concentration in plants is present. Because then, for low soil concentration (C Soil → 0), the concentration ratio BCF can be very high (C Plant/C Soil → ∞). For higher soil concentrations, however, the BCF decreases and approaches a constant value. This pattern was occasionally interpreted as a variable BCF with soil concentration, i.e. a decreasing BCF with increasing soil concentration. A real-world example is the ratio between the measured concentration of p,p'-DDT in radishes and in soil. The concentration ratio is high at low soil concentrations, and decreases for higher soil concentrations. A plausible explanation for this pattern is that plants have a limited sorption capacity for organic contaminants, which becomes saturated at higher soil concentrations. However, a more likely interpretation is that the uptake into plants is from two different and independent sources, namely from soil and from air. When soil concentrations are very low there still is a background contamination of the plant tissue originating from air (Mikes et al. 2009).
Instead of simply calculating the concentration ratio of plant to soil, the relationship between concentrations in plant and soil can be quantified by a linear regression between both if measurements at different concentration levels are available. The slope of the regression between soil concentration as predictor variable and plant concentration as estimated variable can be interpreted as the BCF plant to soil, while the y-axis-intercept can be interpreted as the constant background concentration due to uptake from air.
where C Background is the constant concentration due to uptake from air.
This method has several advantages:
-
all measured values contribute to the calculated BCF;
-
variations in the measured concentrations are adequately considered;
-
the y-axis gives the concentration in plants due to the (constant) concentration in air;
-
the square of the correlation coefficient (r 2) describes how much of the variance in the measured concentration in plants is explained by the variance of the concentrations in soil.
An example is shown in Fig. 9.2. It shows the BCFs for p,p′-DDT in radishes which decrease with increasing soil concentration (Mikes et al. 2009), but with C plant plotted as a function of C Soil. The slope of the regression curve, i.e. the value 0.17, is the BCF derived from all measured values minus the background concentration in air. The BCF is statistically highly significant. The explained variance r 2 is 0.98, which means that the increase of concentrations in plants can be explained almost completely by the increase of concentration in soil.

Calculation of BCF from the slope of the regression between concentrations in soil and plant root; calculation of background concentration due to uptake from air from the y-axis intercept of the regression. Example from (Mikes et al. 2009), p,p′-DDT in roots and soil
3.2 Regression Equations
BCFs for neutral organic contaminants are related to contaminant properties. This is applied in regression equations where physico-chemical properties like lipophilicity of the contaminants are correlated to the BCFs. These regressions based on measured data allow us to estimate concentrations of contaminants in plant tissue from the octanol-water partitioning coefficient, K OW , in case of uptake from soil (e.g. (Travis and Arms 1988)) or from the octanol-air partitioning coefficient, K OA , in case of uptake from air (Kömp and McLachlan 1997).
A frequently applied regression for the uptake of organic contaminants from soil into above-ground plants stems from Travis and Arms (1988):
where BCF v is the bioconcentration factor of vegetation to soil (kg kgdw –1) and K OW (L L–1) expresses the lipophilicity of the contaminant. The curve was fitted with 29 data points in the range 1.15 ≤ log K OW ≤ 9.35. A limitation of the regression is that the uptake from air remains unknown. This may lead to false predictions.
3.3 Root Concentration Factor
Phase equilibrium is the endpoint of diffusion and is achieved when the activity of the contaminant in the root tissue is equal to the activity of the contaminant in the external solution (Lewis 1907). The concentration ratio between root and aqueous solution in phase equilibrium is called the root concentration factor RCF (L kg–1) (Shone and Wood 1974).
The most widely applied regression for the RCF is from Briggs et al. (1982). It has been fitted to data derived from experiments with intact and macerated barley roots. For neutral organic chemicals (phenylureas and o-methylcarbamoyloximes), the relation between log K OW and sorption to roots was:
The dependency on the log K OW was explained by lipophilic sorption of the contaminants to plant lipids. The value of 0.82 was interpreted as water content of the roots. A similar result was obtained for cut pieces of bean roots and stems for N-methyl-arylcarbamates (Trapp and Pussemier 1991):
This equation gives lower root concentration factors for lipophilic contaminants (Fig. 9.3). Both RCF-regressions describe partitioning to water. But roots typically grow in soil. The sorption of organic contaminants to soil is related to the organic carbon content, OC. Many regressions for the organic carbon normalised sorption to soil, K OC (L kgdw –1), were established, for example for neutral organic contaminants (EC 2003)

Bulk soil consists of solids, water and air. For the concentration ratio between bulk soil and soil pore water, K SW (L kg–1) follows
where OC is the fraction of organic carbon in soil (kg kg–1), θ is the water content of soil (L kg–1), ρ W is the density of water (1 kg L–1), ρ S,dry is the soil dry density (kgdw L–1) and ρ S,wet is the soil wet density (kgww L−1). Division of RCF with K SW gives the equilibrium concentration of the ratio of roots to soil.
Both RCF-regressions (Eqs. 9.5 and 9.6), the experimental data of Briggs et al. (1982) and the concentration ratio between bulk soil and soil pore water, K SW , for a typical soil (OC = 0.025 kg kg−1 and W = 0.2 kg kg−1) are plotted in Fig. 9.3. For low K OW values RCF is higher than K SW , due to the higher water content of roots. For higher K OW values Briggs’ RCF regression and the K SW equation (Eq. 9.8) yield similar results. This suggests that the sorption capacity of roots equals that of soil, because the content of organic carbon in soil (in this case 2.5%) is similar to the lipid content of roots (about 2−3%, including waxes and lignin), and the slope of the log K OW in the regressions is similar (0.81 for K OC in Eq. 9.7 and 0.77 for RCF in Eq. 9.5).
3.4 Partition Coefficients for Stem and Leaves
Briggs et al. (1983) measured the sorption to macerated barley stems and predicted K stem/xylem sap (L kg−1), which is the concentration of contaminants in stem tissue divided by the concentration in xylem sap, related to the log K OW of the contaminants:
Trapp et al. (1994) interpreted the regressions derived for sorption to roots and stems as equilibrium partition coefficients between plant tissue and water, K PW (L kg−1), and introduced the general equation:
where W (L kg−1) and L (kg kg−1) are water and lipid content of the plant, b is a correction factor for differences between solubility in octanol and sorption to plant lipids (in the regressions of Briggs et al. (1982, 1983) b was 0.77 for roots and 0.95 for leaves), and a is a factor correcting density differences between water and n-octanol (1/ρ Octanol = 1.22 L kg−1, where ρ Octanol is the density of octanol). When parameterized accordingly, this equilibrium approach gives the same results for roots as the Briggs RCF-regression.
Stems and leaves are in contact with air. The sorption equilibrium of contaminants between leaves and air can be described as follows:
where K LA is the partition coefficient between leaves and air (L kg−1), K LW (L kg−1) is the partition coefficient between leaves and water (Eq. 9.10) and K AW (L L−1) is the partition coefficient between air and water (also known as the dimensionless Henry’s Law constant). Instead of estimating K LA from K OW and K AW , K LA was often directly fitted to K OA , i.e. the partition coefficient between octanol and air (e.g., Kömp and McLachlan 1997).
3.5 Translocation from Roots into Stem and Leaves
Translocation of contaminants from roots into stems in the xylem is often described by the transpiration stream concentration factor TSCF (Russell and Shorrocks 1959):
From experimental data, the concentration in xylem sap is found from the mass of contaminant in shoots divided by the volume of transpired water. Briggs et al. (1982) fitted a Gaussian optimum curve to their data:
Later experiments by other research groups yielded similar results (Burken and Schnoor 1998; Hsu et al. 1990; Sicbaldi et al. 1997). However, recent research has shown that these regressions might not be valid for the polar contaminants (log K OW < 1) and a sigmoidal relationship between TSCF and log K OW has been established (Dettenmaier et al. 2009):
4 Mechanistic Models for Estimating Uptake of Contaminants into Plants
The first author met D. Mackay at his talk at the University of Bayreuth in 1986, where he encouraged his audience to develop mechanistic models for the plant uptake of organic contaminants. So we did (Trapp et al. 1990). D. Mackay and his team also developed some of the early models for this purpose, formulated with the fugacity approach (Paterson et al. 1994). These models were later simplified by Hung and Mackay (1997) to three mass balance equations and solved numerically. At about the same time, the numerical model PlantX was developed (Trapp et al. 1994). Later, crop-specific models were derived, i.e. specific models for roots (Trapp 2002), potatoes (Trapp et al. 2007a), leaves (Trapp and Matthies 1995) and fruits (Trapp 2007). These models are all based on the same physico-chemical principles and describe the same basic processes, such as advective uptake into plants, diffusive uptake, chemical equilibrium, transport in xylem and phloem, dilution by growth, and particle deposition from soil and air. The actually occurring processes and their parameterization depend on the type of crop (Fig. 9.4). Plant models were also developed by other groups, for example a partition-limited model by Chiou et al. (2001) and the models for pesticide uptake by Fujisawa et al. (2002a, b). For electrolytes, such as acids and bases, very different approaches were developed. Examples of such models are the phloem transport model by Kleier (1988), the model relating to pesticide spray application by Satchivi (2000a, b) and the cell model (Trapp 2000, 2004).

Overview of crop-specific plant uptake processes (Legind and Trapp 2009)
4.1 Processes to Include in a Plant Uptake Model
A plant uptake model should consider both uptake from soil and air into plants. A relatively simple model includes the compartments soil, roots and leaves (or fruits or grains) and the processes:
-
continuous and pulse input to all compartments (soil, roots and leaves);
-
degradation, leaching, run-off and plant uptake, resulting in loss from soil;
-
uptake into roots with the transpiration water;
-
growth dilution, degradation and metabolism in roots;
-
translocation from roots to leaves (or fruits) with the transpiration stream;
-
loss from leaves to air;
-
deposition from air to leaves;
-
transport to leaves with attached soil;
-
growth dilution, degradation and metabolism in leaves.
4.2 Mass Balance for a Dynamic Plant Uptake Model
Though not in the scope of this chapter, a mass balance for soil can be described as follows.
Change of contaminant mass in soil is deposition from air minus leaching, run-off, volatilization, degradation and uptake into roots. Division by soil mass, M S , results in the concentration in soil:
where I S (mg d−1) is input to soil (including deposition from air), and k S (d−1) is the sum of all first-order loss rates from soil. For parameters for the equation, see other sections of this book.
The mass balance for thick roots, such as carrots, can be described as follows.
Change of contaminant mass in roots is influx with water minus outflux with xylem sap. Diffusive uptake is not considered, since it only makes a small change in the concentration in roots. The root is described with the following equation:
where m R is the mass of contaminant in roots (mg), Q is the transpiration stream (L d−1), C W is the concentration in soil pore water (mg L−1) and C Xy is the concentration in the xylem at the outflow of the root (mg L−1). If the xylem sap is in equilibrium with the root, the concentration is C Xy = C R /K RW . K RW (L kg−1) is the partition coefficient between root and water (Eq. 9.10). The concentration in soil pore water, C W , is C S × K WS . Substituting these expressions in Eq. 9.16 gives the following equation:
If plant growth is exponential, and the ratio Q/M R (transpiration stream Q to root mass M R (kg)) is constant, the growth by exponential dilution can be considered by a first-order growth rate k R (d−1). If first order degradation or metabolism occurs, the rate k R is the sum of the loss processes and the growth dilution. Division by mass of the root results in the concentration in roots:
The mass balance for leaves can be described as follows.
Change of contaminant mass in leaves is influx with transpiration water plus gaseous and particulate deposition from air minus diffusion to air. This results in the following equation:
where A L is leaf area (m2), K LA is the partition coefficient between leaves and air (L kg−1), C A is the total concentration in air (mg m−3) and f P (−) is the fraction of the total concentration in air that is adsorbed on particles. Uptake from air can either be by diffusive exchange in the gas phase with conductance g L (m d−1), or by deposition of particles on the surface of the leaves (A L /2) with velocity v dep (m d−1). The concentration in leaves is as follows:
where k L (d−1) again is the first-order rate that includes growth dilution and biotic and abiotic (photolysis) degradation processes. The first term of the equation quantifies translocation from roots to leaves and replaces the TSCF in earlier model versions (Trapp and Matthies 1995). The advantages of this new formulation are as follows:
-
There is a relation between concentrations in roots and in leaves. This allows, for example, calculation of the fate of metabolites formed in roots.
-
The TSCF is related to plant physiological parameters, such as transpiration Q, growth rate k and partitioning between root tissue and xylem, K RW (Eq. 9.10).
The calculated concentration ratio between the xylem and the external solution is close to the calculated concentration resulting from the empirical TSCF-regression by Dettenmaier et al. (2009) for all contaminants, and to the TSCF-regressions following a Gaussian curve for contaminants with log K OW > 2 (Trapp 2007). Trapp (2007) speculates, based on this equation, that plants growing in soil outdoors would have a different TSCF-curve than plants grown in hydroponic solutions. This is due to the formation of root hairs in soil, which leads to better diffusive uptake of polar contaminants and subsequently higher TSCF-values (the TSCF remains high (i.e. near 1) for contaminants with a log K OW < 1). Dettenmaier et al. (2009) suggests that differences in experimental methods and plant growth conditions cause the disparity.
An additional process not considered in Eq. 9.20 is the contamination of leaves with attached soil, R (kg soil kg plant−1 (wet weight)). A convenient way of calculation is to add the concentration due to attachment of particles from soil with subsequent deposition on leaves to the calculated C L , as follows:
Default values for R range from 0.001 kg kg−1 to 0.01 kg kg−1 (see Section 9.6.3).
Mass balances for fruit and grain can be set up analogous to Eq. (9.20). However, the processes and parameters differ. For grain, we assume no particle deposition and a lower soil attachment value.
4.3 Steady-State Solution for the Root and Leaf Model
The steady-state (t → ∞) concentration in roots, C R , with constant concentration in soil, C Soil, is as follows:
For leaves, the steady state concentration is:
where I is the sum of all input terms (mg kg−1 d−1):
and a is the sum of all loss processes (d−1):
4.4 General Solutions for a Cascade Model
The system of three linear differential equations (Eqs. 9.15, 9.18 and 9.20) can be solved analytically or numerically for continuous or pulse input. Continuous input occurs from atmospheric deposition to soil and leaves, whereas pulse inputs vary, e.g. inputs from accidents, pesticide spray application and application of manure or compost.
The differential equations for the contaminant concentration in soil, root and leaves can be treated as a diagonal matrix, so that:
where indices 1–3 refer to soil, roots and leaves, respectively; C (mg kg−1) is concentration; k 1 , k 2 and k 3 are the sum of all first-order loss processes in compartment 1, 2 and 3, respectively, and k 12 and k 23 are the transfer rates from compartment 1 to 2 and 2 to 3, respectively. I i (mg d−1) describes the constant input to the compartments, e.g., from air, and M i (kg) is the mass of compartment i, i = 1, 2, 3. The matrix elements k and I can be derived from the differential equations above (Eqs. 9.15, 9.18 and 9.20).
Linear differential equations approach steady state for t → ∞, i.e. the change of concentration with time is zero, dC/dt = 0. The steady-state solutions for matrix equations 1 (soil), 2 (roots) and 3 (leaves) with continuous input are as follows:
The steady-state solution follows the general scheme:
where n is the compartment number.
The analytical solutions for the differential equations 1 (soil), 2 (roots) and 3 (leaves) for a pulse input is the same as for initial concentrations C(0) ≠ 0:
The general solution scheme for pulse input to soil only, i.e. C 1 (0) ≠ 0 and C n (0) = 0 with n > 2 is as follows:
and for pulse input into all compartments, i.e. C n (0) ≠ 0 with \(n \ge 1\):
This solution scheme can offer solutions for several soil-plant-air cascade system variations. For example, in analogy with this example, compartment 1 could be soil, compartment 2 roots, and compartment 3 leaves. Or, alternatively, compartment 1 could be parent contaminant in soil, compartment 2 metabolite in soil, compartment 3 metabolite in root, compartment 4 metabolite in leaves, and compartment 5 metabolite of metabolite in leaves.
Also for repeated applications there is a solution. This is a situation in which one, two or more subsequent pulse inputs occur. In that case, the resulting concentration can be calculated by adding the concentrations resulting from steady state and one, two or more pulse inputs. To this purpose, the simulation is split up into several periods. The concentration vector C (t) at the end of a specific period serves as initial concentration vector C (0) for the next period. This refers to concentrations in any compartment (i.e. soil, roots and leaves).
4.5 Input Data for the Root and Leaf Model
Input data are the same for the steady state and the dynamic model version and are mostly taken from the carrot model (Trapp 2002) and the leafy vegetables model (Trapp and Matthies 1995) (Table 9.1).
5 Influence of Contaminant-Specific Parameters
The uptake of contaminants into plants and their accumulation depends on contaminant-specific parameters. The importance of some of these parameters is investigated in this section by using the models described before. Also, experimental results are reported, not only with the intention to give a review, but also in order to confirm (or to falsify) the processes and the principal outcomes of the model simulations. Many experimental studies on plant uptake of organic contaminants are available, but most of them are from laboratory or greenhouse experiments, and the results may not always be applicable to field conditions. Results from field studies, however, are less often published. This may be due to high expenses, analytical difficulties or other research priorities. But a reason may also be that the results of uptake studies from outdoors often show a very large variation and are difficult to interpret.
5.1 KOW on Accumulation in Roots and Potatoes
Figure 9.5 shows the calculated concentration in roots (steady-state solution Eq. 9.22) and potatoes (Trapp et al. 2007a) for a constant soil concentration of 1 mg kg−1 (wet weight). The most relevant contaminant-specific parameter in the root and potato model is the log K OW , which is varied from 0 (polar contaminants) to 8 (super-lipophilic contaminants). The concentrations are compared to the concentrations resulting from the equilibrium partition coefficient approach (RCF, Eqs. 9.5 and 9.10) and to the Travis and Arms-regression (“T&A”, Eq. 9.3). For very polar contaminants, the concentration in roots and potatoes is predicted to be higher than the concentration in soil. This is because very polar contaminants are mainly found in aqueous phases, and roots and potatoes contain usually more water (up to 95%) than soil (about 30%). For more lipophilic contaminants, the RCF (here related to soil, i.e. RCF/K SW ) approaches a value of 1, due to the similar sorption capacity of roots and soil.

Concentration in root (fresh weight) with varying log K OW , predicted with the regression of Travis and Arms (T&A), the root concentration factor (RCF), the root model and the potato model. Concentration in soil is equal to 1 mg kg−1 (wet weight)
For polar contaminants, the regression of T&A and the two dynamic models give results close to the equilibrium (RCF). With increasing lipophilicity, the predicted concentration decreases and the deviation from equilibrium increases. The root and the potato model include growth dilution, and the effect of this growth dilution increases with increasing log K OW . The BCF predicted by the root model at log K OW 7 is more than a factor 1000 below chemical equilibrium. Chemical equilibrium may be found in the peel or close to the peel. But the inner substance of roots and potatoes will have much lower concentrations (Trapp 2002, 2007).
Interestingly, the T&A regression was originally established for above-ground crops. But Fig. 9.5 shows that the predicted bioconcentration in plants is very close to that predicted by the root model (advective uptake) and the potato model (diffusive uptake).
In current chemical Risk Assessment (EC 2003) and in some Risk Assessment tools for contaminated soils, predictions of concentrations in root vegetables are based on the equilibrium approach. This will lead to an overestimation of the concentration in roots and, hence, of human exposure (Legind and Trapp 2009). Recent studies showed that uptake into root vegetables (radishes) may also be from air (Mikes et al. 2009). This has not yet been considered in any available root uptake model.
5.2 KOW and KAW on Accumulation of Contaminants in Leaves
Simulations were done with the leaf model described in Section 9.4.3 (Eq. 9.23). A value of 0.1% attached soil was chosen as default. This gives a minimum BCF of 0.001 kg kg−1 (wet weight) for all contaminants.
Figure 9.6a shows the calculated concentration in leaves for a concentration in soil of 1 mg kg−1, and in air of 0 mg m−3. Log K OW (x-axis) is varied for volatile (K AW : 0.2 L L−1), semi-volatile (K AW : 0.001 L L−1) and non-volatile contaminants (K AW : is 10−6 L L−1). For all volatile contaminants, the concentration in leaves is low (0.001 mg kg−1) over the whole range of log K OW . This means that their accumulation due to translocation to leaves is low; the plotted concentration is due to attachment of soil particles only (default 0.1%).

Simulated concentration in leaves for compounds with varying log K OW and varying partition coefficient air-water (K AW ), for a concentration in soil equal to 1 mg kg−1 (wet weight). Concentration in air is (a) 0 and (b) in phase equilibrium with soil
The semi-volatile contaminants show some accumulation, up to a BCF of 0.01 kg kg−1 for the most polar contaminants. With increasing lipophilicity of the contaminants the accumulation decreases. The largest accumulation is seen for non-volatile contaminants, in particular for the polar ones (low K OW ). These contaminants are readily soluble in soil pore water, taken up by plants with the soil pore water and translocated to the leaves. From the leaves, the water evaporates, but the contaminants remain. For non-volatile chemicals (K AW → 0), calculated concentrations of polar contaminants in leaves are more than 100 times higher than in soil.
Sulfolane (tetrahydrothiophene 1,1-dioxide) is an organo-sulfur contaminant used as solvent that accumulates in leaves. Sulfolane is neutral, polar (log K OW of –0.77) and non-volatile (K AW of 2.14×10−4). The uptake of sulfolane into wetland vegetation was measured in field and greenhouse studies (Doucette et al. 2005). The measured RCF was between 0.3 and 1.4 L kg−1 (related to the initial concentration in solution). High translocation to leaves was found. The BCF values for shoots were up to 160. Another experiment with sulfolane was done for apples (Chard et al. 2006). The concentration ratio of fruit to soil was 2.8. For leaves, a BCF of 652 was found. As far as the authors are aware, this is the highest BCF plant to soil that was ever measured, and confirms model predictions in which polar, non-volatile contaminants are best translocated to and accumulated in leaves. Also measured BCFs for hexahydro-1,3,5-trinitro-1,3,5-triazine (RDX) (McKone and Maddalena 2007) confirm model predictions for polar non-volatile contaminants.
Contrarily, trichloroethene (TCE) is a volatile chlorinated solvent (K AW : 0.5 L L−1) that does not accumulate in leaves. A study on trichloroethylene uptake by apple and peach trees and transfer to fruit was performed by Chard et al. (2006). No TCE could be detected in fruits, but 14C from unidentified metabolites was found. In leaves, the metabolites dichloroacetic acid (DCAA) and trichloroacetic acid (TCAA) could be detected. The article cites a field study where TCE could be detected in several fruits, but only in traces. Overall, the findings confirm the model prediction (Fig. 9.6a) that volatile contaminants do not show high accumulation in above-ground plant parts like leaves.
5.3 Uptake from Air Versus Uptake from Soil
A frequent experimental result is that contaminants are found in moderate or even high concentrations in plants even though concentrations in soil are low (Delschen et al. 1996, 1999; Mikes et al. 2009). This is typically the case when uptake is mainly from air (compare Section 9.3.1). The simulations displayed in Fig. 9.6b were done for identical conditions as for Fig. 9.6a, except that the concentration in air was set to phase equilibrium to soil (i.e., C Air = K AW × C Soil /K SW ), with concentration in soil equal to 1 mg kg−1). The development of the concentration in plants is completely different from Fig. 9.6a (note that the figure was rotated and the z-axis crosses now at C Leaf equal to 1 mg kg−1). The concentration in leaves is higher than in Fig. 9.6a where there was no contaminant present in the air, in particular for volatile contaminants (K AW : 0.2 L L−1). Also, the concentration is less variable, with most values between 1 and 10 mg kg−1. This is because for most contaminants the system is close to equilibrium in regard to soil with air and air with leaves. An exception are the non-volatile contaminants, their predicted concentration does not change substantially. For the polar and non-volatile contaminants, the calculated concentration in leaves is particularly high.
From Fig. 9.6a and b it can be seen that the partition coefficient between air and water (also known as the dimensionless Henry’s Law constant) is a very important parameter for calculation of the accumulation in leaves, because K OA (the ratio of K OW and K AW ) determines partitioning into leaves. Leaves have a very high exchange with air (that is their role in plant physiology), and any volatile contaminant (with high K AW ) will escape from leaves into air and will not accumulate.
The pattern of uptake of contaminants from soil into fruits is very similar (not shown), although the level of concentrations is typically about a factor of 10 lower (Trapp 2007). This means that also, in fruits, polar and non-volatile contaminants have the highest potential for accumulation from soil.
Uptake into fruits of lipophilic contaminants is preferably from air. An example is the transfer of PCDD/F from contaminated sites into field crops, which has been intensively studied. Müller et al. (1994) found an increase of PCDD/F concentrations in the peel of carrots, when grown at a PCDD/F-contaminated site. No increase of the concentration in lettuce and peas was found. In a similar study, it could be shown that the transfer of PCDD/F into apples and pears is exclusively from air (Müller et al. 1993). The results from these studies confirm the model predictions in which uptake of lipophilic contaminants into above-ground plant parts is primarily from air. However, there are exceptions. Hülster et al. (1994) could proof a transfer of PCCD/F into zucchini and pumpkins in field experiments, which was much higher than for other fruits.
Another example is benzo(a)pyrene (BaP). The range of measured concentrations of BaP in food crops is quite large, from 0.01 to 48 μg kg−1. Generally, leafy vegetables and lettuce had the highest concentrations, followed by grains, potato and root vegetables (Kazerouni et al. 2001; Samsøe-Petersen et al. 2002; SCF 2002). This indicates air as the primary source for contamination of vegetables with PAHs.
Uptake of organochlorine pesticides and polychlorinated biphenyls (PCBs) from soil and air into radishes were measured at a contaminated field site in the Czech Republic (Mikes et al. 2009). Root concentration factors (RCF) and bioconcentration factors for leaves (BCF L ) were determined by linear regression. The transfer from soil into leaves (average BCF L 0.08 kg kgdw −1) was rather constant for all contaminants and could best be explained by soil particle attachment. Generally, uptake from air was higher than uptake from soil. Uptake from air into radish roots and bulbs was observed. The example in Section 9.3.1 was taken from this study.
5.4 Dissipation from Soil
The simulations above were done for the steady-state situation, with constant concentration in soil. However, contaminants can rapidly dissipate from soil by several removal processes, such as degradation, plant uptake, leaching, volatilization and sequestration.
An example is shown in Fig. 9.7. Ortho-xylene is a moderately lipophilic contaminant which is rapidly degraded in soil. Søvik et al. (2002) give a first-order degradation rate constant of 0.11 d−1, i.e. a half-life of about 1 week. After a few weeks, the concentration in soil has approached zero. The simulated concentration in roots closely follows this pattern: About four days after the pulse input, the calculated concentration in root is already close to equilibrium to soil, and parallel with the concentration in soil, concentration in roots falls to very low values within a few weeks, in any case before the harvest. A steady state simulation with constant soil concentration would overpredict concentrations in the harvest product by several orders of magnitude.

Simulated concentration of o-xylene in soil and roots as a function of time, with a degradation rate of 0.11 d−1
The degradation or total dissipation rate is a key variable. To some extent, elimination in soil is related to physico-chemical properties. Bacterial degradation depends on the bioavailable fraction of the contaminants (Alexander 2000; Reichenberg and Mayer 2006), which is typically higher at low sorption and low sequestration. Plant uptake and leaching are important for soluble contaminants with low K OW . Volatilization to air depends on K AW (or K OA ). Therefore, polar and volatile contaminants (low K OW , high K AW ) show shorter residence time in soil. The actual dissipation rate, however, cannot be predicted from physico-chemical properties alone but is an input data usually derived from experiments.
5.5 Impact of pKa and pH on Uptake of Ionisable Contaminants
Ionisable contaminants, i.e. acids, bases, zwitterions or amphoters, may be present in soils as neutral or ionic molecules. The neutral and the ionic molecules have completely different physico-chemical properties. The ion is usually much more polar and water soluble and has a very low vapor pressure compared to the neutral species. Thus, the ion is a polar and non-volatile contaminant, and as such has a high potential for accumulation in plants. On the other hand, cations have a strong tendency to adsorb to soil organic matter and/or clay (Franco and Trapp 2008). Besides, charged contaminants cross biomembranes slowly (Trapp 2004), which limits their uptake.
A process that may lead to high accumulation of ionisable contaminants is the ion trap. This principle is described in Fig. 9.8, for an acid. If the pH outside in the soil pore water is low, a weak acid is at least partly neutral. The uptake into the cell is rapid (provided that the contaminant is not too hydrophilic). The pH in cell sap (cytosol) is about 7 to 7.5, in xylem about 5.5, and in phloem about 8. In particular in cytosol and phloem, weak acids dissociate and form the anion. The membrane permeability of the anion is very low, and reverse diffusion is slow. As a consequence, the anion is trapped inside the cytosol or the phloem. The same process occurs for a base, when the cell sap is acid, compared to the outside soil pore water, for example in the vacuoles of plant cells and in the xylem (pH is 5.5).

Principle of the ion trap. AH is the neutral contaminant (here an acid), and A− is the ion (here an anion)
Summarizing, the ion trap requires a gradient in pH between outside and inside of the plant, so that the outside pH is lower (for bases: higher) than the inside pH. Second, a pKa at or somewhat below (for bases: above) the outside pH is necessary, so that a relevant fraction of contaminants is neutral outside, but most of it is ionic inside. This means that weak acids (pKa 2–6) may accumulate in the alkaline phloem (and are in consequence transported to fruits), while weak bases (pKa 6–10) tend to accumulate in the acidic xylem (and are translocated to leaves). Examples of such contaminants are herbicides like 2,4-D (Shone and Wood 1974) and sulfonylureas (Fahl et al. 1995), and drugs like the base trimethropin (Boxall et al. 2006) and the acid ibuprofen.
Briggs et al. (1987) determined the RCF and TSCF of weak organic acids with log K OW between 0.06 and 4.51 and pKa-values near 3. At pH 7 in external solution, RCF-values were low, between 0.5 and 4.5 L kg−1. At pH 4, the RCF-values were higher and ranged from 2.6 to 72 L kg−1. Similar, the TSCF-values of weak acids at pH 7 were low, 0.04 to 0.05 L L−1, while at pH 4 TSCF-values up to 4.2 L L−1 were found. Briggs et al. used the ion trap process to interpret their results. Chamberlain et al. (1998) studied the uptake into roots and translocation to shoots of two weak bases, i.e. dodemorph (pKa 7.8) and tridemorph (pKa 7.4). At pH 5, RCF-values of dodemorph were < 10 L kg−1, and of tridemorph about 20 L kg−1. With increasing pH, RCF increased to 49 L kg−1 for dodemorph and 183 L kg−1 for tridemorph at pH 8. In parallel, the TSCF increased from < 1 L L−1 for both bases at pH 5 to 24 L L−1 for dodemorph and slightly below 10 L L−1 for tridemorph at pH 8. To our knowledge, the TSCF of dodemorph in this experiment was the highest TSCF ever determined. A similar study was done by Inoue et al. (1998), who also found a large increase of uptake and translocation of weak bases in plants with increasing pH. The results from these studies underline the importance of pH in the soil pore water for uptake and translocation of weak electrolytes. However, the results have not yet been confirmed by field studies.
6 Influence of Plant-Specific Parameters
As pointed out in the introduction, there are a very high number of plant-contaminant combinations, and the simulations done for a generic plant are not necessarily valid for the whole plant empire and all contaminants. In this section, some properties of plants are discussed, which vary and may lead to different accumulation of contaminants.
6.1 Crop Types and Uptake Pathways
The crop type has a very large impact on uptake processes, e.g. roots and potatoes are in close contact to soil, while apples are not. As a consequence, the accumulation of contaminants from soil is much lower in apples and other tree fruits, whereas the accumulation by uptake directly from air is higher for fruits. It is obvious that crop-specific models will give more realistic predictions of concentrations.
Even between different species of the same crop type differences in uptake can exist. For zucchini and pumpkin (Cucurbita pepo), both members of the plant family Cucurbitaceae and the genus Cucurbita, root uptake and subsequent translocation to shoots and fruits was the main uptake route of PCDD/F, probably due to root exudates which mobilize lipophilic contaminants (Hülster and Marschner 1995) and an increased mobility in the transpiration stream (Gent et al. 2007). However, fruits and leaves from other plant species, even for the closely related cucumber plant (Cucumis sativus), were mainly contaminated by airborne PCDD/F (Hülster and Marschner 1994; Hülster et al. 1994). The exact reason for the ability of cucurbita plants to extract lipophilic contaminants from soil and transport them with the xylem sap is yet unknown, but it was observed repeatedly, and for a number of lipophilic organic contaminants, such as p,p'-DDE (White 2002), DDT (Lunney et al. 2004), PCB (Whitfield Åslund et al. 2007) and PBDE (Mueller et al. 2006). For phenanthrene, unusual high adsorption to a range of plant species was observed (Zhu et al. 2007).
6.2 Physiological Parameters
The importance of physiological plant-specific parameters, such as transpiration rate, leaf area, conductance, water and lipid contents as well as growth rate, depends largely on the properties of the contaminant. For water soluble contaminants, which are rapidly translocated from soil to leaves, the transpiration rate is among the most important parameters, since the accumulation in leaves is almost directly dependent on the transpiration.
This is illustrated with the example of carbofuran in Fig. 9.9. The log K OW of this contaminant is 1.6–2.07 (Trapp and Pussemier 1991). Two calculations were done, one with the standard transpiration of 1 L d−1, the other with an increased transpiration of 5 L d−1. The simulated concentration in roots remains nearly constant with increasing transpiration, only the time period until steady state is reached is shorter. However, the breakthrough of contaminant into leaves is faster and the calculated concentration in leaves is a factor of 5 higher for the increased transpiration rate due to their direct relationship. The translocation of carbofuran in bean plants was determined experimentally, and the concentration pattern as well as the close relation between transpiration and concentration in leaves was confirmed (Trapp and Pussemier 1991).

Simulation of the concentration of carbofuran in plants as a function of time, with low transpiration (Q is 1 L d−1) and high transpiration (Q is 5 L d−1). Dynamic model version, data Table 9.1
In turn, the amount of transpired water depends on temperature, humidity, leaf area and stomata permeability. However, the transpiration depends also on the availability of water. Students in a field course at the Technical University of Denmark noticed that concentrations of chlorinated solvents (PCE and TCE) measured in wood cores taken from trees growing at the Glostrup site (Denmark) were much lower in the very dry June 2008 than in the very wet June 2007 (Mette Broholm, personal communication). A plausible explanation is a reduced transpiration due to water stress in 2008. Transpiration in field settings can be estimated using the Penman-Monteith model by Penman (1948) and Monteith (1964) (cited in (Monteith 1995)).
Leaves grow and contaminants entering the leaf with the transpiration stream are not uniformly distributed in the leaf, but tend to accumulate in the leaf tip (Doucette et al. 2005).
6.3 Plant Morphology and Collection Efficiency for Particles
The resuspension of contaminated soil particles from soil to plant surfaces is an important transport mechanism for lipophilic contaminants. This transfer pathway was well studied using insoluble radionuclides, and large variations were found in soil attachment among plant species. Soil particles may reach leaf surfaces mainly by three mechanisms, namely rain splash, wind erosion and erosion due to agricultural practice (ploughing, harvesting et cetera). Table 9.2 shows measured values for the transfer of radionuclides (Li et al. 1994). The transfer range varies from 1.1 to 260 mg soil gplant −1 (dry weight), depending on the species. But also, the intraspecies variation was considerable, as can be seen from the standard deviations given in Table 9.2. The highest value was found for lettuce, with 260 mg soil gplant −1 (dry weight), probably due to the small size of the plant (< 40 cm), the leaf morphology where leaves are collecting particles to the interior of the plant, and the surface structure. It was also demonstrated that particle-bound organic contaminants migrate from the leaf surface to the interior of the leaf (i.e. are overgrown by surface waxes), from where they can not be washed off (Kaupp 1996). A BCF for radish leaves to soil of 0.08 kg kgdw −1 was recently measured at a Czech site for persistent lipophilic contaminants and was contributed to soil particle attachment (Mikes et al. 2009).
A default value of 10 mg soil gplant −1 (fresh weight) for transfer of attached soil was chosen for lettuce in the New Model Framework for dietary exposure of children and adults (Legind and Trapp 2009) (for lettuce, ρ L,wet = 1 kgfw L−1 and ρ L,dry = 0.2 kgdw L−1, so this corresponds to 50 mg soil gplant −1 (dry weight)). For grains, 1 mg soil gplant −1 (fresh weight) transfer with attached soil particles was assumed, due to pollution at harvest. In models predicting the exposure to radio-nuclides, the default value for the transfer of attached soil is 5 mg soil gplant −1 (fresh weight) (25 mg soil gplant −1 (dry weight)) for pasture grass, and 1 mg soil gplant −1 (fresh weight) for other plants (Paretzke and Garland 1990).
In systematic experiments, Delschen et al. (1996, 1999) investigated the uptake and the uptake pathways of PAHs (polycyclic aromatic hydrocarbons) into vegetables and crops. They found that uptake of PAHs can both be from contaminated sites and from the atmosphere. In some experiments, the soil was covered with fine sand or a gas-permeable foliage. This avoided soil particle attachment, but allowed for volatilization with subsequent adsorption to leaves. In fact, this coverage reduced the uptake of 15 PAHs from soil almost completely, indicating soil particle attachment to leaves as a major transfer pathway of PAHs (in particular benzo(a)pyrene and dibenz(a,h)-anthracene) from soil to vegetables. Thus, crops with harvested parts close to the soil surface have the highest affinity for accumulation, because soil particle attachment by rain splashing rarely affects plants with a height above 40 cm (Dreicer et al. 1984; Li et al. 1994). No systemic uptake of PAHs via plant roots was found. Concentrations of PAHs in peels of potato and carrots from organic farming were generally higher than in the core of potatoes and carrots (Zohair et al. 2006). The BCF for individual PAHs ranged from 0.0002 to 0.3 kg kg−1 and decreased with log K OW (Trapp et al. 2007a; Zohair et al. 2006). The results from the experiments are in good accordance with the model predictions.
Note that the ingestion of soil attached to the daily vegetable diet may be higher than the current estimate for direct soil ingestion used in Exposure Assessments for humans. The direct ingestion of soil is 50 mg/day for adults and 100 mg/d for children (ECETOC 2001). The average lettuce consumption in Denmark is 6 g/d for children and 9 g/d for an adult woman. A transfer of 1% (10 mg soil gplant −1 (fresh weight)) due to attached soil corresponds to 60 mg/d and 90 mg/d soil ingestion with lettuce only. Other vegetables and fruits are consumed at 389 g/d (children) and 475 g/d (mother). If on average 0.1% soil were attached, this corresponds to 389 mg/d and 475 mg/d of soil, which is much more than the value of direct ingestion of soil and deserves consideration in Human Health Exposure Assessment.
6.4 Variation of Partition Coefficients
The RCF regressions for barley (Briggs et al. 1982) and bean plants (Trapp and Pussemier 1991) differ (Fig. 9.3). Also, the partition coefficients between leaves and air, K LA , vary with plant species. The K LA depends both on plant-specific parameters (such as plant lipid content and empirical exponent b (Eq. 9.10)) and contaminant-specific parameters (K OW and K AW or K OA ). Kömp and McLachlan (1997) found differences in the uptake of PCB between five different plant species of up to a factor of 20. There was a linear relationship between log K PA and log K OA values within each plant species, but the slopes of the regressions ranged from 0.57 to 1.15. The standard value for leaves (slope = b = 0.95, Eq. 9.9) was determined by Briggs et al. (1983) and lies in this range.
6.5 Permeability
The velocity of diffusive uptake from air and loss to air is controlled by the permeability of leaves (synonyms exchange velocity, conductance or transfer velocity). Three major resistances control the exchange: air boundary layer resistance, stomata resistance and cuticle resistance (Riederer 1995). Their importance depends on the chemical properties but varies for each leaf. Volatile and gaseous contaminants will prefer stomata for the entry to or escape from plants, while lipophilic contaminants will preferably diffuse through cuticles. The stomata resistance for water and contaminants can be calculated from the transpiration rate, leaf area, temperature and humidity (Trapp 1995, 2007). The cuticle resistance depends on thickness and diffusivity inside the cuticle (Schönherr and Riederer 1989). The latter is highly variable with species and temperature (Buchholz et al. 1998): at high temperatures, the surface waxes liquidize and get more permeable.
A method for estimation of conductance g (m s−1, related to gas phase) is given by Trapp (1995, 2007). Values for g estimated with this method range from 9×10−3 m s−1 for non-volatile, lipophilic contaminants to 0.2×10–3 m s−1 for volatile or polar contaminants. A default value of 10−3 m s−1 was suggested, to avoid lenghty calculations (Trapp and Matthies 1995).
6.6 Particle Deposition
A similar variability can be observed for the particle deposition rate. Particle deposition depends on the fraction of contaminant adsorbed to particles, the deposition velocity of particles, as well as characteristics of the leaves. The fraction adsorbed to particles, f P , is often calculated from the Junge equation, using vapor pressure as input data (Junge 1977; cited in EC 2003). Newer methods are based on the K OA (Cousins and Mackay 2001).
The particle deposition rate for wet and dry deposition, v dep , changes with the diameter of the particles. Default values are given in the German TA-Luft and are listed in Table 9.3. Generally, diameters and thus deposition velocities are higher close to the source of emission. At larger distance the coarse particles have settled, and finer particles remain. The deposition rate of fine particles is similar to the conductance of gases, and the default value of v dep (10−3 m s−1) is the same as for conductance g. Thus, uncertainties in f P and v dep are not crucial for the calculated result.
Bakker et al. (1999) studied the deposition of polycyclic aromatic hydrocarbons (PAHs) on leaves from three closely related Plantago species and found variations up to a factor of five, which could be explained solely by plant morphology and surface structure.
6.7 Metabolism in Plants
Rapid metabolism in plants will significantly decrease the contaminant concentration in plants. Transformation of contaminants may occur in the rhizosphere, inside the plant and on the leaf surface. Often, it is difficult to differentiate between metabolism by fungi or bacteria living on and in plants and metabolism by plant cells. Stimulation of bacterial degradation in the root zone has been observed frequently and is a key process in phytoremediation projects (Trapp and Karlson 2001). Therefore, it may happen that non-persistent contaminants do not reach the roots but are degraded by bacteria living on or in the vicinity of roots.
The role of the plant enzyme system is detoxification, and there is an upper limit, beyond which plants suffer from toxic effects and probably die (Trapp et al. 2007b). This contamination of plants may limit activities in gardening, agriculture and forestry on contaminated sites.
Metabolism by plants has been described by the green liver concept, because plant metabolism rather resembles the processes in the animal liver than the bacterial metabolism (Sandermann 1994). The first step (phase I reaction) is typically an oxygenation with cytochrome P-450, followed by conjugation reactions (phase II) with glutathione-S-transferases (GST) (Barret 1995; Pflugmacher and Schröder 1995). Unlike animals, plants are not able to excrete conjugates via the urine. Instead, phase III of plant xenobiotic metabolism involves storage of soluble conjugates in the vacuole and of insoluble conjugates in the cell wall (Komossa et al. 1995). This may lead to so-called bound residues. These bound unextractable residues resist solubilization in common laboratory solvents and are therefore not accessible to standard residue analysis. It was found that bound residues can be present in larger amounts than the parent contaminant and could therefore represent a source of significant consumer exposure (Sandermann 2004).
Little is known about metabolism rates of contaminants by vegetation. Cyanide (HCN) was used as model contaminant to study the variation of rates among plant species. Even though inorganic, cyanide behaves like an organic contaminant in terms of lipid solubility, volatility and metabolism. The removal of free cyanide followed Michael-Menten kinetics (Larsen et al. 2004). Adding Michaelis-Menten kinetics to the mass balance equation for roots (Eq. 9.18) leads to the following non-linear equation:
where v max (mg kg−1 d−1) is the maximal metabolism velocity of the contaminant and K M (mg L−1) is the half-saturation constant and (Larsen et al. 2005).
At a low external concentration in soil pore water, all contaminants that are taken up are metabolized (Fig. 9.10). At higher concentrations, however, the enzyme system is overloaded. Then, uptake is linearly related to the external concentration. This was shown experimentally for free cyanide (HCN) by Larsen et al. (2005). A non-linear BCF relation indicating enzymatic activity of plants was found repeatedly, e.g. for phenol (Ucisik and Trapp 2006) and salt, NaCl (Trapp et al. 2008). In these cases, the BCF was low at a low external concentration in soil pore water, but increased at higher external concentrations.

Calculation of concentration in roots of cyanide as a function of the concentration in external aqueous solution (CW), using Michaelis-Menten kinetics for metabolism; v max is 6.9 mg kg−1 h−1; K M is 0.44 mg L−1 (Larsen et al. 2005)
The Michaelis-Menten parameters K M and v max varied with plant species, but less than expected. Values of v max of 12 species from nine plant families were found in a relatively narrow range between 6.7 and 21.9 mg CN kgplant −1 h−1 and were normally distributed with a mean of 13 mg CN kgplant −1 h−1 (Yu et al. 2004). The authors concluded that the variation of metabolism rates between plant species was small, and that it is likely that all investigated plants used the same enzyme for detoxification. Another sign for common pathways and rates of metabolism in plant species is the difficulty to find selectively acting herbicides: it is rare that a toxin hits weeds but not crops (Börner 1995).
7 Environmental Variables
Which contaminants are taken up into plants and the variation in the affinity for uptake, and hence accumulation, depends also on environmental parameters. The importance of some of these parameters is investigated in this section by using the models described before and some experimental results.
7.1 Climate
Principally, the models can be adapted to all climatic conditions at which plants grow. In particular temperature influences uptake and fate of xenobiotics in various ways. At higher temperatures plant physiological processes such as transpiration, growth and metabolism are stimulated. According to Claudius-Clapeyron’s law, the vapor pressure increases exponentially with temperature, so the vapor pressure of water in air is 872 Pa at 5°C but 5635 Pa at 35°C, i.e. a factor of 6.5 higher. The potential transpiration is directly proportional to the vapor pressure of water. Similarly, metabolic reactions are stimulated at higher temperatures. For example, the removal rate of cyanide was 3−5 times higher at 30°C than at 11°C (Yu et al. 2005a, 2007). Also, contaminant properties change with temperature. Like the vapor pressure, the partition coefficient air to water, K AW also increases exponentially with temperature. For chlorinated solvents, such as TCE, the K AW at 5°C is about a factor of 3 below that at 35°C (Kühne et al. 2005). This means less accumulation in leaves and more volatilization to air at higher temperatures.
7.2 Bioavailability
The concentration in the soil pore water is for neutral organic contaminants calculated from the K OC . The bioavailability of contaminants may be reduced due to aging (Alexander 2000). At contaminated sites the soil pore water concentrations can be much lower and sorption coefficients can be much higher than equilibrium partitioning models predict (Ter Laak et al. 2006). Cations are attracted by the electrical potential of living cells, but also adsorb to soil organic carbon and to negatively charged clay particles (Franco and Trapp 2008). This reduces their bioavailability and, hence, uptake. Also, a depletion of contaminants in soil due to uptake into plants should be taken into account. In Chapter 16 an extended description of bioavailability is given.
7.3 Soil pH
Soil pH directly affects the speciation of acids and bases, as described by the Henderson-Hasselbalch law (log [A−]/[HA] = pH − pKa). Uptake of anions is generally lower than of neutral molecules, due to electrical repulsion and slow transfer across membranes. Both weak acids and bases can undergo the ion trap process (Section 9.5.5). This will lead to an accumulation of weak acids from acidic soils, and of bases from alkaline soils. This conclusion is based on a model prediction and has not yet been confirmed by experiments. The pH also has an indirect effect on uptake: many plants do not grow well outside their optimum pH range. Extreme pH (high or low), will lead to reduced growth, and this may be accompanied by reduced uptake of contaminants.
7.4 Uncertainties in Predictions
Sections 9.5, 9.6 and 9.7 list parameters and variables that influence the uptake of contaminants into plants. Most likely, this list is far from complete. This may explain why under some conditions, in some experiments, a high uptake of a contaminant into a plant may be found, while this may not be the case in the next investigation, under other conditions. It also explains why data from experiments with plants often suffer from large scatter. Besides, care must be taken when results obtained from uptake studies are translated to other crop types, other climates, and other agricultural practices.
Models may help to design and interpret uptake experiments, in indicating relevant processes and pathways, and hence in translating results to other conditions. But due to the large number of parameters and their high variability in space and time, these models can not be expected to give exact results. Some studies tested the validity of model approaches (Fryer and Collins 2003; Legind and Trapp 2009; McKone and Maddalena 2007; Rikken et al. 2001; Trapp and Schwartz 2000). Often, the results were ambiguous, because the influence of concentrations in air could not be quantified (compare Section 9.3.1), but also due to large uncertainties in measured as well as predicted concentrations in plants (McKone and Maddalena 2007). The primary role of models is to indicate relevant processes and the potential for accumulation of contaminants in plants. Good decision making needs to consider both model predictions and experimental data.
In a critical review of uncertainties related to soil-to-plant bioconcentration factors by McKone and Maddalena (2007), BCF-values for the explosive RDX (hexahydro-1,3,5-trinitro-1,3,5-triazine) are listed. RDX is a quite persistent, relatively polar (log K OW : 0.87) and non-volatile contaminant (K AW : 6.3 × 10−8 atm m3 mol−1). BCF-values range from 0.06 (mg kgww plant −1: mg kgdw soil −1) in bean pods and 0.07 (mg kg ww plant −1: mg L water−1) in bean fruits at the lower end to 19.2 (mg kgww plant −1: mg Lwater −1) in bean leaves and 28.6 (mg kgdw plant −1: mg kgdw soil −1) in carrot shoots at the upper end. Overall, fruits tend to have lower BCF values than roots, and leaves have the highest accumulation, but there is a very large variation in the data. The BCF values for fruits range from 0.07 to 5.50, for roots from 0.08 to 4.50 and for leaves from 0.30 to 28.6, i.e. the maximum BCF is 79 fold, 56 fold or 93 fold larger than the minimum BCF. Even though the units are not uniform (i.e. a mix of wet weight and dry weight, soil and water), this variation is quite impressive for one single contaminant and similar crop type.
8 Uptake Potential of Specific Substance Classes
This section gives a short classification of several frequently found soil and groundwater contaminants with respect to their uptake into plants. It is based on the model simulations, measured data and the process review above, but also on the experience of the authors.
8.1 Chlorinated Solvents (PCE, TCE and Others)
Chlorinated solvents such as perchloroethene (PCE) and trichloroethene (TCE) are probably the most frequently found groundwater contaminants world-wide. They are water-soluble, relatively persistent and very volatile. Uptake of chlorinated solvents into tree trunks has been reported frequently (Chard et al. 2006; Larsen et al. 2008; Sorek et al. 2008; Vroblesky et al. 1999). However, these contaminants are highly volatile and escape rapidly from branches, small stems and leaves into the air (Baduru et al. 2008). Thus, in fruits, only metabolites of chlorinated solvents could be detected (Chard et al. 2006). Accumulation in root crops and potatoes is likely, though not described yet. During storage and cooking, chlorinated solvents could be eliminated from food stuff, due to their high volatility. Moreover, these contaminants are not expected to be present in high concentrations in top soil, i.e. the root zone, due to volatilization to the air and leaching to the groundwater.
8.2 Gasoline Contaminants
Gasoline is a mixture of light petroleum products (for example alkanes such as hexane, and aromates such as benzene and toluene) plus additives like ethanol and MTBE. They frequently occur in the groundwater due to leaching storage tanks. Gasoline contaminants are volatile and usually rapidly degraded. Thus, concentrations in aerated soil are generally low (Fig. 9.7). Accumulation in crops from soil has not been described, except in laboratory experiments. An exception is MTBE, which is less volatile and more persistent. MTBE was not metabolized in any of 24 plant species tested (Trapp et al. 2003). Uptake of benzene from air is rapid but levels are low, due to a low K OA , and insignificant for human exposure (Collins et al. 2000).
8.3 Heavy Petroleum Products
The heavy fraction of petroleum products consists of long-chain alkanes and some polycyclic aromatic hydrocarbons. These contaminants are lipophilic, volatile and degradable under aerobic conditions. Significant uptake from soil into plants has not been observed and is not expected from the properties of petroleum components.
8.4 Polycyclic Aromatic Hydrocarbons
Polycyclic aromatic hydrocarbons (PAHs) is a group of contaminants with two (naphthalene) to seven or more condensed aromatic rings with wide-spread occurrence in the environment from incomplete combustion. Most of these contaminants are very lipophilic (log K OW 5−7) and have a very low water-solubility. Uptake into plants with transpiration water is therefore unlikely. Contamination of plants is mainly via attached soil particles or from air (Delschen et al. 1999). Uptake from soil into the peel of carrots or potatoes may occur. Solubilization by cucurbita species (pumpkin and zucchini) is likely.
8.5 Persistent Organic Pollutants POPs
Persistent Organic Pollutants (POPs) are chlorinated contaminants like the pesticides DDT, dieldrin and lindane, but also polychlorinated biphenyls (PCB) and polychlorinated dibenzo-p-dioxins and -furans (PCDD/F). POPs are persistent, semi-volatile and lipophilic. Similar to PAHs, the uptake with transpiration water into plants is very unlikely, while attached soil particles can lead to contamination of plants. Solubilization with subsequent translocation by cucurbita species (pumpkin and zucchini) has been observed repeatedly.
8.6 Explosives
Explosives such as trinitrotoluene (TNT) and hexahydro-1,3,5-trinitro-1,3,5-triazine (RDX) are relatively polar contaminants with low vapor pressure. High uptake into plants has been observed (McKone and Maddalena 2007, Thompson et al. 1998). Inside plants, a reduction of the nitro groups to amino groups is likely and would lead to an increase of the toxic potential. Furthermore, bound residues may be formed.
8.7 Phenols
Phenol is the common name for a class of contaminants with a phenolic ring. A variety of chemical groups may be attached to the ring. This leads to contaminants with a large variety of properties (nitrophenols, anilines, chlorinated phenols). Phenols are less volatile than the aromatics benzene, toluene, ethylbenzene and xylene (BTEX). The contaminant phenol as well as monochlorophenol were degraded rapidly in the root zone (Ucisik and Trapp 2006, 2008), while 2,4-dichlorophenol was taken up into roots and translocated to stem, and to some extent also to leaves (Ucisik et al. 2007).
8.8 Cyanides
Cyanides are contaminants with a CN-group. They are often found at (abandoned) gasworks sites and gold mines. Free cyanide (HCN, CN−) is rapidly taken up into roots and metabolized. An accumulation in healthy plants was not observed, because free CN is highly toxic (Larsen et al. 2004, 2005; Yu et al. 2004, 2005b). Iron-complexed cyanide (ferri- and ferrocyanide) is taken up by plants and translocated upwards, while slow metabolism was observed (Ebbs et al. 2003; Larsen and Trapp 2006; Samiotakis and Ebbs 2003). Field measurements at a former gas works site showed no accumulation of total cyanide (sum of free and complexed CN) in leaves (Trapp and Christiansen 2003, and own results, unpublished). Surface contamination of crops by solid iron cyanide (such as Prussian blue) is possible and has been observed (“blue strawberries”).
9 Monitoring of Contaminants in Soils and Shallow Aquifers with Vegetation
The uptake of soil and groundwater contaminants into plants has also positive aspects: contaminants such as chlorinated solvents in soils and shallow aquifers can be monitored using plant samples. In principle, every tree could be considered as a well, a pump and a passive sampler, all in one. Plant samples can be taken from stems of trees (Fig. 9.11) and analyzed to identify and map subsurface contamination. The method is rapid and cheap. Screening an area suspect of contamination in the upper aquifer with the tree core method prior to boring observation wells can support the determination of optimal location of these wells, instead of arbritrarily placing wells. This reduces the number of wells necessary for the investigation and thus reduces overall costs. Besides, tree cores can be sampled at sites which are difficult to access with heavy equipment, for example gardens in residential areas or dense forests. Even better, the effect on trees of the contaminants in a plume is reflected by the size of tree rings, and dendrochronology can be used to determine when a plume passed below the tree (Balouet et al. 2007).

Demonstration of the tree core method by one of the authors
The potential of this monitoring procedure is large. Tree core sampling as monitoring method was successfully applied for chloroethenes (Gopalakrishnan et al. 2007; Vroblesky et al. 1999). Larsen et al. (2008) found a good agreement between the plume delineation by traditional well drilling and by tree core sampling, but only monotone (and not linear) relations. The authors could also use the method to monitor Natural Attenuation of a PCE spill. Sorek et al. (2008) even detected several before unknown spills in Tel Aviv by a random sampling of trees in the city. But there are also limitations. Tree core sampling should rather be used for assessing the presence of contaminants than for determining exact subsurface concentrations. Because from tree core measurements alone, precise evaluation of subsurface contamination will not be possible, due to varying concentrations in wood (Larsen et al. 2008; Sorek et al. 2008).
The method could also be successful for some heavy metals such as cadmium and copper, and it may also work for MTBE, perchlorate and dichlorophenol. For a number of contaminants the method was not successful, to mention are BTEX, PAHs (except naphthalene), free and iron complexed cyanide, long-chain alkanes (generally petroleum products), phenol, monochlorophenol, iron and lead.
A free guide to vegetation sampling for screening of subsurface pollution is available from the web (Trapp et al. 2009).
10 Conclusions
What can be concluded from this review? An important conclusion is that the polar contaminants are preferably translocated from soil and accumulate in leaves and fruits. Lipophilic contaminants will be retained in soil, roots and the lower stem and will not reach the leaves or fruits in significant amounts. Thus, polar and non-volatile contaminants, i.e. contaminants with low K OW and K AW , have the highest accumulation potential from soil into plants. This is in conflict with the usual concept of bioaccumulation, where high bioaccumulation is assumed to occur for highly lipophilic contaminants (EC 2003; Mackay and Fraser 2000). Furthermore, under background conditions, all contaminants except the polar and non-volatile contaminants will preferably be taken up from air and the concentration in soil does not have much impact on the concentration in leaves, unless it is far above chemical equilibrium.
The number of available experimental data on plant uptake of contaminants increases. Often, model concepts and measurements are in good agreement, but we may also encounter surprises. Plants are living organisms, and the uptake processes are complex and variable. Uncertainties in the model predictions are high, and large variations may be expected in predicted, but also in experimental data, due to the variations of contaminant-specific, plant physiological and environmental parameters as described in this chapter.
References
Alexander M (2000) Aging, bioavailability, and overestimation of risk from environmental pollutants. Environ Sci Technol 34:4259–4265
Baduru KK, Trapp S, Burken JG (2008) Direct measurement of VOC diffusivities in tree tissues: impacts on tree-based phytoremediation. Environ Sci Technol 42:1268–1275
Bakker M, Vorenhout M, Sijm DTHM, Kollöffel C (1999) Dry deposition of atmospheric polycyclic aromatic hydrocarbons in three plantago species. Environ Toxicol Chem 18, 2289–2294
Balouet J-C, Oudijk G, Smith KT, Petrisor I, Grudd H, Stocklassa B (2007) Applied dendroecology and environmental forensics. Characterizing and age dating environmental releases: fundamentals and case studies. Environ Forensics 8:1–17
Barret M (1995) Metabolism of herbicides by cytochrome P 450 in corn. Drug Metabol Drug Interactions 12:299–315
Boxall ABA, Johnson P, Smith EJ, Sinclair CJ, Stutt E, Levy LS (2006) Uptake of veterinary medicines from soils into plants. J Agric Food Chem 54(6):2288–2297
Börner H (1995) Unkrautbekämpfung. Gustav Fischer, Jena, D
Brand E, Otte PF, Lijzen JPA (2007) CSOIL 2000: an exposure model for human risk assessment of soil contamination. RIVM Report 711701054/2007, National Institute of Public Health and the Environment. http://rivm.openrepository.com/rivm/bitstream/10029/13385/1/711701054.pdf. Accessed 13 August 2010
Briggs GG, Bromilow RH, Evans AA (1982) Relationship between lipophilicity and root uptake and translocation of non-ionised chemicals by barley. Pestic Sci 13:495–504
Briggs GG, Bromilow RH, Evans AA, Williams M (1983) Relationships between lipophilicity and the distribution of non-ionised chemicals in barley shoots following uptake by the roots. Pestic Sci 14:492–500
Briggs GG, Rigitano RLO, Bromilow RH (1987) Physicochemical factors affecting uptake by roots and translocation to shoots of weak acids in barley. Pestic Sci 19:101–112
Buchholz A, Baur P, Schönherr J (1998) Differences among plane species in cuticular permeabilities and solute mobilities are not caused by differential size selectivities. Planta 206:322–328
Burken JG, Schnoor JL (1998) Predictive relationships for uptake of organic contaminants by hybrid poplar trees. Environ Sci Technol 32:3379–3385
Chamberlain K, Patel S, Bromilow RH (1998) Uptake by roots and translocation to shoots of two morpholine fungicides in barley. Pestic Sci 54:1–7
Chard BK, Doucette WJ, Chard JK, Bugbee B, Gorder K (2006) Trichloroethylene uptake by apple and peach trees: greenhouse study. Environ Sci Technol 40:4788–4793
Chiou CT, Sheng GY, Manes M (2001) A partition-limited model for the plant uptake of organic contaminants from soil and water. Environ Sci Technol 35(7):1437–1444
Collins CD, Bell JNB, Crews C (2000) Benzene accumulation in horticultural crops. Chemosphere 40:109–114
Cousins I, Mackay D (2001) Gas-particle partitioning of organic compounds and its interpretation using relative solubilities. Environ Sci Technol 35:643–647
Czub G, McLachlan MS (2004) A food chain model to predict the levels of lipophilic organic contaminants in humans. Environ Toxicol Chem 23:2356–2366
DEFRA (Department for Environment Food and Rural Affairs) (2002) The Contaminated land exposure assessment model (CLEA): technical basis and algorithms. R & D Publications CLR 10, Environment Agency. http://publications.environment-agency.gov.uk/pdf/SCLR10-e-p.pdf?lang=_e. Accessed 13 August 2010
Delschen T, Hembrock-Heger A, Necker U (1996) Systematische Untersuchungen zum Verhalten von PAK und PCB im System Boden/Pflanze auf der Lysimeter-Anlage Waldfeucht (1989–1994). In: Landesumweltamt NRW (eds) Materialien zur Ermittlung und Sanierung von Altlasten, LUA Essen, Germany, pp 1–214
Delschen T, Hembrock-Heger A, Leisner-Saaber J, Sopczak D (1999) Bedeutung verschiedener Belastungsursachen für den PAK-Gehalt von Kulturpflanzen. Umweltwissenschaften und Schadstoff-Forschung, UWSF – Z Umweltchem Ökotox 11:79–87
Dettenmaier EM, Doucette WJ, Bugbee B (2009) Chemical hydrophobicity and uptake by plant roots. Environ Sci Technol 43(2):324–329
Doucette WJ, Chard TJK, Moore BJ, Staudt WJ, Headley JV (2005) Uptake of sulfolane and diisopropanolamine (DIPA) by cattails (Typha latifolia). Microchem J 81:41–49
Dreicer M, Hakonson TE, White GC, Whicker FW (1984) Rainsplash as a mechanism for soil contamination of plant surfaces. Health Phys 46:177–187
Ebbs S, Bushey J, Poston S, Kosma D, Samiotakis M, Dzombak D (2003) Transport and metabolism of free cyanide and iron cyanide complexes by willow. Plant Cell Environ 26:1467–1478
EC (European Commission) (2003) Technical guidance document on risk assessment in support of commission directive 93/67/EEC on risk assessment for new notified substances, Commission Regulation (EC) No 1488/94 on Risk Assessment for existing substances, and Directive 98/8/EC of the European Parliament and of the Council concerning the placing of biocidal products on the market, European Communities, Italy. http://ecb.jrc.it/documents/. Accessed 13 August 2010
ECETOC (European Centre for Ecotoxicology and Toxicology of Chemicals) (2001) Exposure factors sourcebook for european populations (with focus on UK data). Technical Report No. 79, Brussels, Belgium
EFSA (European Food Safety Authority) (2006) Pesticides are EU citizens’ top food-related health concern. Available online at http://www.euractiv.com. Accessed 17 March 2006
Fahl GM, Kreft L, Altenburger R, Faust M, Boedeker W, Grimme LH (1995) pH-Dependent sorption, bioconcentration and algal toxicity of sulfonylurea herbicides. Aquat Toxicol 31(2):175–187
Franco A, Trapp S (2008) Estimation of the soil-water partition coefficient normalized to organic carbon for ionizable organic chemicals. Environ Toxicol Chem 27(10):1995–2004
Franke W (1987) Nutzpflanzenkunde. Thieme Verlag, Stuttgart, Germany
Fryer ME, Collins CD (2003) Model intercomparison for the uptake of organic chemicals by plants. Environ Sci Technol 37:1617–1624
Fujisawa T, Ichise K, Fukushima M, Katagi T, Takimoto Y (2002a) Mathematical model for the uptake of non-ionized pesticides by edible root of root crops. J Pesticide Sci 27:242–248
Fujisawa T, Ichise K, Fukushima M, Katagi T, Takimoto Y (2002b) Improved uptake models of non-ionized pesticides to foliage and seed of crops. J Agric Food Chem 50: 532–537
Gent MPN, White JC, Parrish ZD, Isleyen M, Eitzer BD, Mattina MI (2007) Uptake and translocation of p,p'-dichlorodiphenyldichloroethylene supplied in hydroponics solution to Cucurbita. Environ Toxicol Chem 26(12):2467–2475
Gopalakrishnan G, Negri MC, Minsker BS, Werth CJ (2007) Monitoring subsurface contamination using tree branches. Ground Wat Monit Rem 27:65–74
Hsu FC, Marxmiller RL, Yang AYS (1990) Study on root uptake and xylem translocation of cinmethylin and related compounds in detopped soybean roots using a pressure chamber technique. Plant Physiol 93:1573–1578
Hülster A, Marschner H (1994) PCDD/PCDF-Transfer in Zucchini und Tomaten. Veröff PAÖ 8:579–589
Hülster A, Marschner H (1995) Identifizierung PCDD/PCDF-mobilisierender Verbindungen in Wurzelexudaten von Zucchini. Veröff PAÖ 12:359–369
Hülster A, Müller JF, Marschner H (1994) Soil-plant transfer of polychlorinated dibenzo-p-dioxins and dibenzofurans to vegetables of the cucumber family (Cucurbitaceae). Environ Sci Technol 28:1110–1115
Hung H, Mackay D (1997) A novel and simple model of the uptake of organic chemicals by vegetation from air and soil. Chemosphere 35:959–977
Inoue J, Chamberlain K, Bromilow RH (1998) Physicochemical factors affecting the uptake by roots and translocation to shoots of amine bases in barley. Pestic Sci 54:8–21
Kaupp H (1996) Atmosphärische Eintragswege und Verhalten von polychlorierten Dibenzo-p-dioxinen und -furanen sowie polyzyklischen Aromaten in einem Maisbestand. PhD thesis, faculty of biology, chemistry and earth science, University of Osnabrück
Kazerouni N, Sinha R, Hsu CH, Greenberg A, Rothman N (2001) Analysis of 200 food items for benzo[a]pyrene and estimation of its intake in an epidemiologic study. Food Chem Toxicol 39:423–436
Kleier DA (1988) Phloem mobility of xenobiotics. Plant Physiol 86:803–810
Komossa D, Langebartels C, Sandermann H Jr (1995) Metabolic processes for organic chemicals in plants. In: Trapp S, Mc Farlane C (eds) Plant contamination – modeling and simulation of organic chemical processes. Lewis, Boca Raton, Florida, pp 69–103
Kömp P, McLachlan M (1997) Interspecies variability of the plant/air partitioning of polychlorinated biphenyls. Environ Sci Technol 31:2944–2948
Kühne R, Ebert RU, Schüürmann G (2005) Prediction of the temperature dependency of Henry’s Law constant from chemical structure. Environ Sci Technol 39:6705–6711
Larsen M, Trapp S (2006) Uptake of iron cyanide complexes into willow trees. Environ Sci Technol 40:1956–1961
Larsen M, Trapp S, Pirandello A (2004) Removal of cyanide by woody plants. Chemosphere 54:325–333
Larsen M, Ucisik A, Trapp S (2005) Uptake, metabolism, accumulation and toxicity of cyanide in willow trees. Environ Sci Technol 39:2135–2142
Larsen M, Burken J, Machackova J, Karlson UG, Trapp S (2008) Using tree core samples to monitor natural attenuation and plume distribution after a PCE spill. Environ Sci Technol 42:1711–1717
Legind CN, Trapp S (2009) Modeling the exposure of children and adults via diet to chemicals in the environment with crop-specific models. Environ Pollut 157:778–785
Lewis GN (1907) A new system of thermodynamic chemistry. Proc Am Acad Arts Sci 43: 259–297
Li JG, Gerzabek MH, Mück K (1994) An experimental study on mass loading of soil particles on plant surfaces. Bodenkultur 45:15–24
Lunney AI, Zeeb BA, Reimer K (2004) Uptake of weathered DDT in vascular plants: potential for phytoremediation. Environ Sci Technol 38:6147–6154
Mackay D, Fraser A (2000) Bioaccumulation of persistent organic chemicals: mechanisms and models. Environ Pollut 110:375–391
McKone TE, Maddalena RL (2007) Plant uptake of organic pollutants from soil: bioconcentration estimates based on models and experiments. Environ Toxicol Chem 26:2494–2504
Mikes O, Cupr P, Trapp S, Klanova J (2009) Uptake of polychlorinated biphenyls and organochlorine pesticides from soil and air into radishes (Raphanus sativus). Environ Pollut 157(2):488–496
Monteith JL (1995) Accommodation between transpiring vegetation and the convective boundary-layer. J Hydrol 166(3–4):251–263
Müller JF, Hülster A, Päpke O, Ball M, Marschner H (1993) Transfer pathways of PCDD/F into fruits. Chemosphere 27:195–201
Müller JF, Hülster A, Päpke O, Ball M, Marschner H (1994) Transfer of PCDD/F from contaminated sites into carrots, lettuce and peas. Chemosphere 29:2175–2181
Mueller KE, Mueller-Spitz SR, Henry HF, Vonderheide AP, Soman RS, Kinkle BK, Shann JR (2006) Fate of pentabrominated diphenyl ethers in soil: abiotic sorption, plant uptake, and the impact of interspecific plant interactions. Environ Sci Technol 40:6662–6667
Paretzke HG, Garland JA (1990) Assessment of the radiological significance of surface contamination in entrained radioactivity. Final Report, EC-contract No 90-ET-015
Paterson S, Mackay D, McFarlane C (1994) A model of organic-chemical uptake by plants from soil and the atmosphere. Environ Sci Technol 28:2259–2266
Penman HL (1948) Natural evaporation from open water, bare soil and grass. Proc R Soc Lon A 193(1032):120–145
Pflugmacher S, Schröder P (1995) Glutathione S-transferases in trees: inducibility by various organic xenobiotics. Z Pflanzenernähr Bodenk 158:71–73
Reichenberg F, Mayer P (2006) Two complementary sides of bioavailability: accessibility and chemical activity of organic chemicals in sediments and soils. Environ Toxicol Chem 25:1239–1245
Riederer M (1995) Partitioning and transport of organic chemicals between the atmospheric environment and leaves. In: Trapp S, Mc Farlane JC (eds) Plant contamination. Modeling and simulation of organic chemical processes. Lewis, Boca Raton, FL, pp 153–190
Rikken MGJ, Lijzen JPA, Cornelese AA (2001) Evaluation of model concepts on human exposure. Proposals for updating the most relevant exposure routes of CSOIL. RIVM report 711 701 022, Bilthoven, NL
Russell RS, Shorrocks VM (1959) The relationship between transpiration and the absorption of inorganic ions by intact plants. J Exp Bot 10(29):301–316
Samiotakis M, Ebbs SD (2003) Possible evidence for transport of an iron cyanide complex by plants. Environ Pollut 127:169–173
Samsøe-Petersen L, Larsen EH, Larsen PB, Bruun P (2002) Uptake of trace elements and PAHs by fruit and vegetables from contaminated soils. Environ Sci Technol 36:3057–3063
Sandermann H (1994) Higher plant metabolism of xenobiotics: the ‘green liver’ concept. Pharmacogenetics 4:225–241
Sandermann H (2004) Bound and unextractable pesticidal plant residues: chemical characterization and consumer exposure. Pest Manag Sci 60:613–623
Satchivi NM, Stoller EW, Wax LM, Briskin DP (2000a) A nonlinear dynamic simulation model for xenobiotic transport and whole plant allocation following foliar application I. Conceptual foundation for model development. Pestic Biochem Phys 68:67–84
Satchivi NM, Stoller EW, Wax LM, Briskin DP (2000b) A nonlinear dynamic simulation model for xenobiotic transport and whole plant allocation following foliar application II. Model validation. Pestic Biochem Phys 68:85–95
SCF (Scientific Committee on Food) (2002) Annex: background document to the opinion of the scientific committee on food on the risks to human health of polycyclic aromatic hydrocarbons in food. Polycyclic aromatic hydrocarbons – occurrence in foods, dietary exposure and health effects. http://ec.europa.eu/food/fs/sc/scf/out154_en.pdf. Accessed 13 August 2010
Schönherr J, Riederer M (1989) Foliar penetration and accumulation of organic-chemicals in plant cuticles. Rev Environ Contam Toxicol 108:1–70
Shone MGT, Wood AV (1974) Comparison of uptake and translocation of some organic herbicides and a systemic fungicide by barley 1. Absorption in relation to physicochemical properties. J Exp Bot 25(85):390–400
Sicbaldi F, Sacchi GA, Trevisan M, Del Re AAM (1997) Root uptake and xylem translocation of pesticides from different chemical classes. Pestic Sci 50:111–119
Sitte P, Ziegler H, Ehrendorfer F, Bresinsky A (1991) Lehrbuch der Botanik für Hochschulen, 33rd edn. Gustav Fischer, Stuttgart
Sorek A, Atzmon N, Dahan O, Gerstl, Z, Kushisin, L, Laor Y, Mingelgrin U, Nasser A, Ronen D, Tschachansky L, Weisbrod N, Graber ER (2008) “Phytoscreening”: the use of trees for discovering subsurface contamination by VOCs. Environ Sci Technol 42:536–542
Søvik AK, Alfnes E, Breedveld GD, French HK, Pedersen TS, Aagaard P (2002) Transport and degradation of toluene and o-xylene in an unsaturated soil with dipping sedimentary layers. J Environ Qual 31:1809–1823
TA-Luft (Technische Anleitung zur Reinhaltung der Luft) (1986) In: Handbuch des Umweltschutzes, Vol. II-2, appendix 3.1. ecomed, Landsberg aL, D
Ter Laak TL, Barendregt A, Hermens JLM (2006) Freely dissolved pore water concentrations and sorption coefficients of PAHs in spiked, aged, and field-contaminated soils. Environ Sci Technol 40:2184–2190
Thompson PL, Ramer LA, Guffey AP, Schnoor JL (1998) Decreased transpiration in poplar trees exposed to 2,4,6-trinitrotoluene. Environ Toxicol Chem 17:902–906
Trapp S (1995) Model for uptake of xenobiotics into plants. In: Trapp S, Mc Farlane JC (eds) Plant contamination. Modeling and simulation of organic chemical processes. Lewis, Boca Raton, FL, pp 107–152
Trapp S (2000) Modeling uptake into roots and subsequent translocation of neutral and ionisable organic compounds. Pest Manage Sci 56:767–778
Trapp S (2002) Dynamic root uptake model for neutral lipophilic organics. Environ Toxicol Chem 21:203–206
Trapp S (2004) Plant uptake and transport models for neutral and ionic chemicals. Environ Sci Pollut Res 11:33–39
Trapp S (2007) Fruit Tree model for uptake of organic compounds from soil and air. SAR QSAR Environ Res 18:367–387
Trapp S, Christiansen H (2003) Phytoremediation of cyanide-polluted soils. In: McCutcheon SC, Schnoor JL (eds) Phytoremediation: transformation and control of contaminants. Wiley, Hoboken, NJ, pp 829–862
Trapp S, Karlson U (2001) Aspects of phytoremediation of organic compounds. J Soils Sed 1:37–43
Trapp S, Matthies M (1995) Generic one-compartment model for uptake of organic chemicals by foliar vegetation. Environ Sci Technol 29:2333–2338; erratum 30, 360
Trapp S, Pussemier L (1991) Model calculations and measurements of uptake and translocation of carbamates by bean plants. Chemosphere 22:327–339
Trapp S, Schwartz S (2000) Proposals to overcome limitations in the EU chemical risk assessment scheme. Chemosphere 41:965–971
Trapp S, Matthies M, Scheunert I, Topp EM (1990) Modeling the bioconcentration of organic chemicals in plants. Environ Sci Technol 24:1246–1252
Trapp S, Matthies M, Mc Farlane C (1994) Model for uptake of xenobiotics into plants: validation with bromacil experiments. Environ Toxicol Chem 13:413–422
Trapp S, Yu X, Mosbæk H (2003) Persistence of methyl tertiary butyl ether (MTBE) against metabolism by Danish vegetation. Environ Sci Pollut Res 10:357–360
Trapp S, Cammarano A, Capri E, Reichenberg F, Mayer P (2007a) Diffusion of PAH in potato and carrot slices and application for a potato model. Environ Sci Technol 41:3103–3108
Trapp S, Ucisik AS, DelChicca Romano P, Larsen M (2007b) The role of plants and bacteria in phytoremediation – kinetic aspects. In: Heipieper HJ (ed) Bioremediation of soils contaminated with aromatic compounds. NATO science series, IV. Earth and environmental sciences – vol 76. Springer, Dordrecht, NL, pp 41–49
Trapp S, Feificova D, Rasmussen NF, Bauer-Gottwein P (2008) Plant uptake of NaCl in relation to enzyme kinetics and toxic effects. Environ Exp Bot 64: 1–7
Trapp S, Larsen M, Legind CN, Burken J, Machackova J, Karlson UG (2009) A guide to vegetation sampling for screening of subsurface pollution. Freely available at http://homepage.env.dtu.dk/stt/. Accessed 13 August 2010
Travis CC, Arms AD (1988) Bioconcentration of organics in beef, milk, and vegetation. Environ Sci Technol 22:271–274
Travis CC, Hattemer-Frey HA (1991) Human exposure to dioxin. Sci Total Environ 104:97–127
Ucisik AS, Trapp S (2006) Uptake, removal, accumulation, and phytotoxicity of phenol in willow trees (Salix viminalis). Environ Toxicol Chem 25:2455–2460
Ucisik AS, Trapp S (2008) Uptake, accumulation, phytotoxicity and removal of 4-chlorophenol in willow trees. Arch Environ Contam Toxicol 54:619–627
Ucisik AS, Trapp S, Kusk KO (2007) Uptake, accumulation, phytotoxicity and removal of 2,4-dichlorophenol in willow trees. Environ Toxicol Chem 26:1165–1171
Vroblesky DA, Nietch CT, Morris JT (1999) Chlorinated ethenes from groundwater in tree trunks. Environ Sci Technol 33:510–515
Whitfield Åslund ML, Zeeb BA, Rutter A, Reimer KJ (2007) In situ phytoextraction of polychlorinated biphenyl – (PCB) contaminated soil. Sci Total Environ 374:1–12
White JC (2002) Differential bioavailability of field-weathered p,p'-DDE to plants of the Cucurbita and Cucumis genera. Chemosphere 49:143–152
Yu X, Trapp S, Puhua Z, Chang W, Xishi Z (2004) Metabolism of cyanide by Chinese vegetation. Chemosphere 56:121–126
Yu X, Trapp S, Zhou P, Hu H (2005a) The effect of temperature on the rate of cyanide metabolism of two woody plants. Chemosphere 59:1099–1104
Yu XZ, Trapp S, Zhou P (2005b) Phytotoxicity of cyanide to weeping willow trees. Environ Sci Pollut Res 12:109–113
Yu XZ, Trapp S, Zhou PH, Chen L (2007) Effect of temperature on the uptake and metabolism of cyanide by weeping willows. Int J Phytorem 9:243–255
Zhu Y, Zhang S, Zhu Y-G, Christie P, Shan X (2007) Improved approaches for modeling the sorption of phenanthrene by a range of plant species. Environ Sci Technol 41:7818–7823
Zohair A, Salim A-B, Soyibo AA, Beck AJ (2006) Residues of polycyclic aromatic hydrocarbons (PAHs), polychlorinated biphenyls (PCBs) and organochlorine pesticides in organically-farmed vegetables. Chemosphere 63:541–553
Acknowledgement
We thank our editor, Frank Swartjes, for his initiative, patience and help. Our work is supported by the European Union 6th Framework Program of Research, Thematic Priority 6 (Global change and ecosystems), contract number GOCE-CT-2007–036976, project 2-FUN, and contract number GOCE 037017, project OSIRIS. This work received also financial support from the Danish Council for Strategic Research, project REMTEC, contract 2104-07-0009. Support for this work was furthermore provided through a PhD grant of the University of Copenhagen for Charlotte N. Legind.
Model Availability Most models mentioned in this chapter are freely available via the authors. Please send an email if you are interested.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2011 Springer Science+Business Media B.V.
About this chapter
Cite this chapter
Trapp, S., Legind, C.N. (2011). Uptake of Organic Contaminants from Soil into Vegetables and Fruits. In: Swartjes, F. (eds) Dealing with Contaminated Sites. Springer, Dordrecht. https://doi.org/10.1007/978-90-481-9757-6_9
Download citation
DOI: https://doi.org/10.1007/978-90-481-9757-6_9
Published:
Publisher Name: Springer, Dordrecht
Print ISBN: 978-90-481-9756-9
Online ISBN: 978-90-481-9757-6
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)