
Abstract
Although epigenetic aberrations frequently occur in aging and cancer and form a core component of these conditions, perhaps the most useful aspect of epigenetic processes is that they are readily reversible. Unlike genetic effects that also play a role in cancer and aging, epigenetic aberrations can be relatively easily corrected. One of the most widespread approaches to the epigenetic alterations in cancer and aging is dietary control. This can be achieved not only through the quality of the diet, but also through the quantity of calories that are consumed. Many phytochemicals such as sulforaphane from cruciferous vegetables and green tea have anticancer epigenetic effects and are also efficacious for preventing or treating the epigenetic aberrations of other age-associated diseases besides cancer. Likewise, the quantity of calories that are consumed has proven to be advantageous in preventing cancer and extending the lifespan through control of epigenetic mediators. The purpose of this chapter is to review some of the most recent advances in the epigenetics of cancer and aging and to provide insights into advances being made with respect to dietary intervention into these biological processes that have vast health implications and high translational potential.
Access provided by Autonomous University of Puebla. Download conference paper PDF
Similar content being viewed by others


Keywords
1 Introduction
Although there are many different variations of the definition of epigenetics, perhaps the most widespread is that epigenetic processes involve changes that are heritable but are not encoded with the DNA sequence itself. There are numerous types of epigenetic mechanisms, and the three most important in mammals include changes in DNA methylation, histone modifications, and non-coding RNAs.
DNA methylation is the most studied of the epigenetic processes and is based on the addition of a methyl moiety (CH3) donated enzymatically from S-adenosylmethionine (SAM) to the 5-position of cytosine, primarily occurring in CpG dinucleotides. This is carried out by three major methyltransferases (DNMT1, DNMT3A, and DNMT3B) in mammalian systems. DNMT1 is responsible for maintaining the methylation pattern that is largely preserved with each mitotic division, and DNMT3A and DNMT3B are more involved with de novo methylation, the creation of new methylated cytosines (5-methylcytosines) at cytosine that were not previously methylated. In general, the more methylated a gene regulatory region becomes, the less transcription will occur from the promoter although there are notable exceptions to this dogma as occurs with the gene that encodes human telomerase reverse transcriptase (hTERT), the key regulatory gene of telomerase [11, 44].
Epigenetic changes are also mediated by histone modifications. Although histone acetylation and methylation are the most studied of these modifications, others also occur such as histone phosphorylation, ubiquitination, biotinylation, sumoylation, and ADP-ribosylation. The number of enzymes that carry out histone modifications is large relative to those that mediate DNA methylation and the two that often attract interest, especially with regard to cancer and aging, are the histone acetyltransferases (HATs) and the histone deactylases (HDACs) [26]. In general, the more acetylated the histone amino tails become, the more likely it is that the gene promoter region that contains those histones will have increased transcriptional activity [7].
Non-coding RNA, the third major type of epigenetic control in mammalian systems, is also important in gene expression. For example, microRNA (miRNA) consists of single-stranded non-coding RNAs that are usually about 21–23 nucleotides in length. These sequences suppress gene expression by altering the stability of gene transcripts and also by targeting the transcripts for degradation although miRNA may also lead to an increase in gene transcription [36]. Many miRNAs have now been identified and may regulate a large percentage of genes in mammals [14].
2 Cancer Epigenetics and Dietary Intervention
Environmental factors such as the diet are well known to influence gene expression and to contribute to cancer through epigenetic mechanisms [20]. To illustrate the importance of this, it has been reported that over half of the gene defects that occur in cancer are through epigenetic alterations when compared to genetic mutations [23]. General DNA hypomethylation not uncommonly occurs in cancer cells, while gene-specific hypermethylation often leads to inactivation of key genes such as the tumor suppressors [3]. It is thought that these changes in DNA methylation play an early role in cancer genesis and lead to aberrations in cellular proliferation as well as immortalization of previously normal cells. Changes in histone modifications are also prevalent in cancer cells. For example, increased activity of the HDACs is a very common feature of cancer cells and can lead to tumorigenesis through effects on epigenetic gene expression [13]. Alterations in miRNA are also common in cancer, and this occurs in many cases through defects in the expression of genes that play a role in cancer initiation or progression [14]. Perhaps most importantly, a growing interest has been the collective interactions of epigenetic processes in the control of gene regulation aberrations in cancer. For instance, it is not uncommon for epigenetic mechanisms to act in concert which lead to alterations in gene expression in cancer cells [15].
2.1 The Epigenetics Diet in Cancer Prevention and Treatment
Many natural dietary agents which consist of bioactive compounds have been shown to be effective in cancer prevention and treatment, and these nutraceuticals often mediate favorable epigenetic changes [38, 50]. In fact, we have suggested an “epigenetics diet” that impacts the epigenome and that may be used in conjunction with other cancer prevention and chemotherapeutic strategies [18].
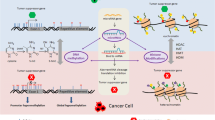
One of the foremost compounds that have been shown in a vast number of studies to have anticancer properties is (-)-epigallocatechin-3-gallate (EGCG) of green tea. Many studies have shown a positive connection between green tea (EGCG) consumption and the inhibition of numerous cancers [6, 25, 48]. Although EGCG has varied effects on cancer cells such as antioxidant properties, it has also been shown to have important epigenetic effects in that it can inhibit DNMT activity by directly interacting with the DNMTs [16]. This can effectively lead to the reversal of tumor suppressor epigenetic silencing of cancer cells and induce apoptosis of these cells. Further, we have found that the inhibition of DNMTs by EGCG can lead to the suppression of telomerase in cancer cells by down-regulating hTERT [4, 41]. hTERT activity in cancer cells is associated with increased DNA methylation of its gene regulatory region due to repressors binding to its promoter [4]. Since telomerase is central to tumor progression, this EGCG-mediated down-regulation of telomerase activity may have major implications in cancer prevention and treatment (Fig. 1).

Effects of components of the epigenetics diet on epigenetics, telomerase (hTERT), and aging and cancer. Plant products such as sulforaphane from cruciferous vegetables and (-)-epigallocatechin-3-gallate (EGCG) (structures shown) modify epigenetic processes that can have a direct impact on aging and cancer. They also lead to the down-regulation of hTERT which is central to both aging and cancer. The mechanisms for epigenetic modifications of the phytochemicals can vary depending on the particular compound. Glucose restriction also can impact epigenetic processes and affect aging and cancer
Another key dietary component that has epigenetic effects is sulforaphane (SFN) of cruciferous vegetables such as broccoli, cauliflower, Brussels sprouts, cabbage, and kale. However, although we have shown that SFN can inhibit DNMT1 and DNMT3A [39], its most powerful effects are through inhibition of HDAC activity [12, 42, 43, 47]. As aforementioned, HDACs are often increased in cancer cells and inhibition of HDAC activity by SFN may have considerable potential in preventing HDAC increases in cancer cells. Since hTERT is also controlled though histone acetylation/deacetylation [11], we tested whether SFN may have an impact on hTERT gene regulation in breast cancer cells. We found that SFN treatment of MCF-7 and MDA-MB-231 cells leads to a dose- and time-dependent down-regulation of hTERT (Fig. 1). The SFN-induced hyperacetylation facilitated the binding of many hTERT repressor proteins such as MAD1 and CTCF to the hTERT gene regulatory region [39]. We also found that depletion of CTCF using siRNA attenuated the SFN-induced hTERT down-regulation in breast cancer cells [39].
Although dietary compounds such as EGCG in green tea and SFN in cruciferous vegetables have many other effects, it is clear that these compounds can affect the epigenetic control of key genes and greatly influence the initiation and progression of cancer [38]. In addition to EGCG and SFN, a number of other dietary compounds are also quite effective in cancer prevention. For instance, curcumin (tumeric), resveratrol (grapes and red wine), and genistein (soybeans) as well as many other phytochemicals have created considerable excitement for their epigenetic potential [38]. We feel that a diet consisting of epigenetic-modifying foods and beverages could have a major impact on the incidence of cancer worldwide and have therefore encouraged the epigenetic diet as a means to not only prevent cancers, but also perhaps treat many early stage cancers [18]. It is also highly feasible that a combination of these phytochemicals in the diet may show synergistic effects to further reduce the incidence of cancer [40].
3 Aging Epigenetics: The Impact of Dietary Factors
The single most important risk factor for developing cancer is age, and therefore, many investigations have explored the role of epigenetics in both cancer and aging with the intention that there may be links between these two important biological processes. In fact, as with cancer, aging is associated with general genomic hypomethylation and regional or gene-specific hypermethylation [22, 37] which may be due to changes in the expression of the DNMTs [5, 34]. These modulations in DNA methylation during aging likely contribute to a number of changes in the regulation of epigenetically controlled genes such as hTERT to contribute to the aging process [33]. Moreover, histone modifications appear to play a major role in aging. In fact, sirtuin 1 (SIRT1) is a class III NAD+-dependent HDACs that has shown remarkable effects on aging through increasing the lifespan of a diverse range of animal models [8, 24]. The SIRT1 enzyme appears to be an important nutrient sensor linked to metabolic rate. The redox potential, simplified as the NAD/NADH ratio, may be important in regulating SIRT1 activity which is a key indicator for oxygen consumption and the respiratory chain. Although many other epigenetic changes occur during the aging process, the regulation of DNA methylation and histone modifications as well as epigenetic control of hTERT and the role of SIRT1 in modulating aging biological processes have captured considerable interest.
3.1 Nutrient Quantities, Epigenetics and Aging
As aforementioned, not only is the quality of the diet an important factor, but the quantity of nutrients consumed is also a major player in cancer and aging. Caloric restriction (CR) is the most effective intervention into the aging process and maximum lifespan, and it is mediated in part by epigenetic mechanisms [49, 53]. The restriction of total calories by 25–60 % relative to normally fed controls while providing essential nutrients can lead to a 50 % increase in lifespan [9, 10, 21, 31, 45, 52]. DNA methylation may be altered in response to CR through its effects on specific gene loci leading to increased longevity [19]. Moreover, SIRT1, an important HDAC in the aging process, is strongly linked to CR. For example, many studies have shown that its activity is affected by CR both in vitro and in vivo [8, 17, 27, 32]. The longevity–extension effects of sirtuin were originally discovered in yeast [17], and activation of SIRT1 is often observed in various tissues of animals subjected to CR, while inactivation of SIRT1 may lead to ablation of the lifespan-extending effects of CR. It is therefore apparent that epigenetic processes are not only central to the aging process, but also that they are involved with key mediators of aging such as DNA methylation and SIRT1.
3.2 The Epigenetics Diet and Aging
Resveratrol, a dietary polyphenol phytochemical, is an important mediator of CR and acts as a SIRT1 mimic that leads to increased longevity both in vitro and in vivo [1, 2, 51, 54]. Besides resveratrol, many other polyphenols such as EGCG have been shown to have beneficial effects on the aging process [46]. Other epigenetic diet components such as cruciferous vegetables and soybeans may also have advantageous effects on longevity through their cancer preventive properties [28, 31, 38]. For instance, consumption of components of the epigenetics diet over a period of time may lead to a decrease in age-associated diseases such as cancer and cardiovascular disease [35].
3.3 Glucose Restriction and Extension of the Hayflick Limit
The Hayflick limit refers to the cellular senescence process that is considered to be fundamental to biological aging. Since studies of CR have been confined to analysis of the organism longevity (including single-cell yeast), we sought to test whether CR could affect the Hayflick limit in mammalian systems. Moreover, the effects of CR on human aging have not yet been resolved largely because of the impracticality of CR studies using relatively long-lived human populations. Since cellular senescence is considered a fundamental basis of aging, we restricted glucose in cultures of human fibroblast cells and monitored the Hayflick limit of the cells [29]. The human cells that received only about 15 mg/L of glucose had a significantly reduced lifespan when compared to control human cells that received the normal level of 4.5 g/L of glucose in culture. We further noted that epigenetic alterations occurred in these cells in response to glucose restriction that led to a relatively modest increase (compared to cancer cells) in hTERT and a decrease in p16, a gene which slows cellular replication. We found that the epigenetic changes in these cells in response to CR were at least in part due to changes in DNA methylation and histone modifications in these cells (Fig. 2). Further, precancerous cells were found to have the opposite effects on hTERT and p16 in response to glucose restriction and to lead to epigenetic-induced apoptosis of the cells [29] (Fig. 2).

Effects of glucose restriction on longevity and cancer inhibition through epigenetic regulation. Glucose restriction can impact epigenetic regulation in both normal and cancer cells. In normal cells, it leads to p16 repression and hTERT activation to extend the Hayflick limit. In precancerous cells, the opposite effects on p16 and hTERT lead to apoptosis, cellular senescence, and cancer inhibition of the glucose-restricted cells
In additional investigations, our studies have indicated that SIRT1 becomes elevated in the glucose-restricted human cells that lead to lifespan extension through epigenetic effects of SIRT1 on p16 as well as genetic effects of SIRT1 on p16 through the Akt/p70S6K1 pathway [30]. Therefore, CR is effective in extending the lifespan of not only animals, but also their individual cells. Since the Hayflick limit is a basic aspect of aging and there were no metabolic systemic factors involved in these studies, this suggests that CR works primarily at the cellular senescence level and that an important mediator of cellular senescence and aging is changes in epigenetic mechanisms in response to CR.
4 Conclusions
Epigenetic mechanisms are central to many aspects of both aging and cancer, and dietary factors are important means of alleviating many of the adverse effects of these biological processes. Both the quality and the quantity of the diet are crucial in healthy aging and also significantly impact cancer. The quality of the diet is illustrated though the epigenetics diet consisting of consumption of phytochemicals that modulate epigenetic processes such as DNA methylation, histone modifications, and non-coding RNA. Substantial data that have been accumulated worldwide clearly show that the epigenetics diet has considerable potential in not only preventing cancer, but also reducing the incidence of age-related diseases. The quantity of the diet also has epigenetic effects in that reduction in calories impacts many epigenetic mechanisms such as the activity of SIRT1 which is an epigenetic mediator of a number of cellular processes. We have shown that CR at the cellular level can extend the lifespan of human cells and is likely a fundamental basis of the life-extending process of the epigenetic effects of CR. Many questions remain regarding the role of epigenetics in cancer and aging, but it is now clear that epigenetic mechanisms are basic aberrations in both cancer and aging and that the epigenetics diet is moving to the forefront of cancer and aging research as a safe and efficacious means to reduce the morbidity and mortality of these biological processes that claim so many human lives.
Abbreviations
- CR:
-
Caloric restriction
- DNMT:
-
DNA methyltransferases
- EGCG:
-
(-)-epigallocatechin-3-gallate
- HAT:
-
Histone methyltransferases
- HDAC:
-
Histone deacetylase
- hTERT:
-
Human telomerase reverse transcriptase
- miRNA:
-
microRNA
- SAM:
-
S-adenosylmethionine
- SFN:
-
Sulforaphane
- siRNA:
-
Short-interfering RNA
- SIRT1:
-
Sirtuin 1
References
Barger JL, Kayo T, Vann JM et al (2008) A low dose of dietary resveratrol partially mimics caloric restriction and retards aging parameters in mice. PLoS ONE 3(6):e2264
Bass TM, Weinkove D, Houthoofd K et al (2007) Effects of resveratrol on lifespan in Drosophila melanogaster and Caenorhabditis elegans. Mech Ageing Dev 128:546–552
Baylin SB, Herman JG, Graff JR et al (1998) Alterations in DNA methylation: a fundamental aspect of neoplasia. Adv Cancer Res 72:141–196
Berletch JB, Liu C, Love WK et al (2008) Epigenetic and genetic mechanisms contribute to telomerase inhibition by EGCG. J Cell Biochem 103:509–519
Casillas MA Jr, Lopatina N, Andrews LG et al (2003) Transcriptional control of the DNA methyltransferases is altered in aging and neoplastically-transformed human fibroblasts. Mol Cell Biochem 252:33–43
Chen PN, Chu SC, Kuo WH et al (2011) Epigallocatechin-3 gallate inhibits invasion, epithelial-mesenchymal transition, and tumor growth in oral cancer cells. J Agric Food Chem 59:3836–3844
Clayton AL, Hazzalin CA, Mahadevan LC (2006) Enhanced histone acetylation and transcription: a dynamic perspective. Mol Cell 23:289–296
Cohen HY, Miller C, Bitterman KJ et al (2004) Calorie restriction promotes mammalian cell survival by inducing the SIRT1 deacetylase. Science 305:390–392
Colman RJ, Anderson RM, Johnson SC et al (2009) Caloric restriction delays disease onset and mortality in rhesus monkeys. Science 325:201–204
Cruzen C, Colman RJ (2009) Effects of caloric restriction on cardiovascular aging in non-human primates and humans. Clin Geriatr Med 25:733–743
Daniel M, Peek GW, Tollefsbol TO (2012) Regulation of the human catalytic subunit of telomerase (hTERT). Gene 498:135–146
Dashwood RH, Ho E (2008) Dietary agents as histone deacetylase inhibitors: sulforaphane and structurally related isothiocyanates. Nutr Rev 66(Suppl 1):S36–S38
Dokmanovic M, Marks PA (2005) Prospects: histone deacetylase inhibitors. J Cell Biochem 96:293–304
Esquela-Kerscher A, Slack FJ (2006) Oncomirs—microRNAs with a role in cancer. Nat Rev Cancer 6:259–269
Esteller M (2008) Epigenetics in cancer. N Engl J Med 358:1148–1159
Fang MZ, Wang Y, Ai N et al (2003) Tea polyphenol (-)-epigallocatechin-3-gallate inhibits DNA methyltransferase and reactivates methylation-silenced genes in cancer cell lines. Cancer Res 63:7563–7570
Guarente L, Picard F (2005) Calorie restriction–the SIR2 connection. Cell 120:473–482
Hardy TM, Tollefsbol TO (2011) Epigenetic diet: impact on the epigenome and cancer. Epigenomics 3:503–518
Hass BS, Hart RW, Lu MH et al (1993) Effects of caloric restriction in animals on cellular function, oncogene expression, and DNA methylation in vitro. Mutat Res 295:281–289
Herceg Z (2007) Epigenetics and cancer: towards an evaluation of the impact of environmental and dietary factors. Mutagenesis 22:91–103
Holloszy JO, Fontana L (2007) Caloric restriction in humans. Exp Gerontol 42:709–712
Issa JP (1999) Aging, DNA methylation and cancer. Crit Rev Oncol Hematol 32:31–43
Issa JP (2008) Cancer prevention: epigenetics steps up to the plate. Cancer Prev Res (Phila) 1:219–222
Kanfi Y, Peshti V, Gozlan YM et al (2008) Regulation of SIRT1 protein levels by nutrient availability. FEBS Lett 582:2417–2423
Kim JW, Amin AR, Shin DM (2010) Chemoprevention of head and neck cancer with green tea polyphenols. Cancer Prev Res (Phila) 3:900–909
Kouzarides T (2007) Chromatin modifications and their function. Cell 128:693–705
Leibiger IB, Berggren PO (2006) Sirt1: a metabolic master switch that modulates lifespan. Nat Med 12:34–36
Li Y, Liu L, Andrews LG et al (2009) Genistein depletes telomerase activity through cross-talk between genetic and epigenetic mechanisms. Int J Cancer 125:286–296
Li Y, Liu L, Tollefsbol TO (2010) Glucose restriction can extend normal cell lifespan and impair precancerous cell growth through epigenetic control of hTERT and p16 expression. FASEB J 24:1442–1453
Li Y, Tollefsbol TO (2011) p16(INK4a) suppression by glucose restriction contributes to human cellular lifespan extension through SIRT1-mediated epigenetic and genetic mechanisms. PLoS ONE 6:e17421
Li Y, Daniel M, Tollefsbol TO (2011) Epigenetic regulation of caloric restriction in aging. BMC Med 9:98
Lin SJ, Defossez PA, Guarente L (2000) Requirement of NAD and SIR2 for life-span extension by calorie restriction in Saccharomyces cerevisiae. Science 289:2126–2128
Liu L, Wylie RC, Andrews LG et al (2003) Aging, cancer and nutrition: the DNA methylation connection. Mech Ageing Dev 124:989–998
Lopatina N, Haskell JF, Andrews LG et al (2002) Differential maintenance and de novo methylating activity by three DNA methyltransferases in aging and immortalized fibroblasts. J Cell Biochem 84:324–334
Mathers JC (2006) Nutritional modulation of ageing: genomic and epigenetic approaches. Mech Ageing Dev 127:584–589
Mathers JC, Strathdee G, Relton CL (2010) Induction of epigenetic alterations by dietary and other environmental factors. Adv Genet 71:3–39
Mays-Hoopes LL (1989) DNA methylation in aging and cancer. J Gerontol 44:35–36
Meeran SM, Ahmed A, Tollefsbol TO (2010a) Epigenetic targets of bioactive dietary components for cancer prevention and therapy. Clin Epigenetics 1:101–116
Meeran SM, Patel SN, Tollefsbol TO (2010b) Sulforaphane causes epigenetic repression of hTERT expression in human breast cancer cell lines. PLoS ONE 5(7):e11457
Meeran SM, Patel SN, Li Y et al (2012) Bioactive dietary supplements reactivate ER expression in ER-negative breast cancer cells by active chromatin modifications. PLoS ONE 7(5):e37748
Mittal A, Pate MS, Wylie RC et al (2004) EGCG down-regulates telomerase in human breast carcinoma MCF-7 cells, leading to suppression of cell viability and induction of apoptosis. Int J Oncol 24:703–710
Myzak MC, Tong P, Dashwood WM et al (2007) Sulforaphane retards the growth of human PC-3 xenografts and inhibits HDAC activity in human subjects. Exp Biol Med (Maywood) 232:227–234
Nian H, Delage B, Ho E et al (2009) Modulation of histone deacetylase activity by dietary isothiocyanates and allyl sulfides: studies with sulforaphane and garlic organosulfur compounds. Environ Mol Mutagen 50:213–221
Poole JC, Andrews LG, Tollefsbol TO (2001) Activity, function, and gene regulation of the catalytic subunit of telomerase (hTERT). Gene 269:1–12
Pugh TD, Oberley TD, Weindruch R (1999) Dietary intervention at middle age: caloric restriction but not dehydroepiandrosterone sulfate increases lifespan and lifetime cancer incidence in mice. Cancer Res 59:1642–1648
Queen BL, Tollefsbol TO (2010) Polyphenols and aging. Curr Aging Sci 3:34–42
Schwab M, Reynders V, Loitsch S et al (2008) The dietary histone deacetylase inhibitor sulforaphane induces human beta-defensin-2 in intestinal epithelial cells. Immunology 125(2):241–251
Shanmugam MK, Kannaiyan R, Sethi G (2011) Targeting cell signaling and apoptotic pathways by dietary agents: role in the prevention and treatment of cancer. Nutr Cancer 63:161–173
Sinclair DA (2005) Toward a unified theory of caloric restriction and longevity regulation. Mech Ageing Dev 126:987–1002
Su LJ, Mahabir S, Ellison GL et al (2011) Epigenetic contributions to the relationship between cancer and dietary intake of nutrients, bioactive food components, and environmental toxicants. Front Genet 2:91
Subramanian L, Youssef S, Bhattacharya S et al (2010) Resveratrol: challenges in translation to the clinic–a critical discussion. Clin Cancer Res 16:5942–5948
Sun D, Krishnan A, Su J, Lawrence R et al (2004) Regulation of immune function by calorie restriction and cyclophosphamide treatment in lupus-prone NZB/NZW F1 mice. Cell Immunol 228:54–65
Weindruch R, Walford RL, Fligiel S et al (1986) The retardation of aging in mice by dietary restriction: longevity, cancer, immunity and lifetime energy intake. J Nutr 116:641–654
Wood JG, Rogina B, Lavu S et al (2004) Sirtuin activators mimic caloric restriction and delay ageing in metazoans. Nature 430:686–689
Acknowledgments
This work was supported in part by grants from the National Cancer Institute (RO1 CA129415), the American Institute for Cancer Research, and the Norma Livingston Foundation.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2014 Springer-Verlag Berlin Heidelberg
About this paper
Cite this paper
Tollefsbol, T.O. (2014). Dietary Epigenetics in Cancer and Aging. In: Zappia, V., Panico, S., Russo, G., Budillon, A., Della Ragione, F. (eds) Advances in Nutrition and Cancer. Cancer Treatment and Research, vol 159. Springer, Berlin, Heidelberg. https://doi.org/10.1007/978-3-642-38007-5_15
Download citation
DOI: https://doi.org/10.1007/978-3-642-38007-5_15
Published:
Publisher Name: Springer, Berlin, Heidelberg
Print ISBN: 978-3-642-38006-8
Online ISBN: 978-3-642-38007-5
eBook Packages: MedicineMedicine (R0)