
Abstract
Water use efficiency (or transpiration efficiency) describes the intrinsic trade-off between carbon fixation and water loss that occurs in dryland plants because water evaporates from the interstitial tissues of leaves whenever stomata open for CO2 acquisition. The transpiration efficiency of crop plants is generally low as they typically lose several 100-fold more water than the equivalent units of carbon fixed by photosynthesis. With the increasing demand for sustainable water use and increasing agricultural productivity, the need to improve transpiration efficiency (TE) of crops has received much attention, although this trait may not be beneficial in all water-limited environments. This chapter shows that TE is predominantly driven by hydraulic properties and genes that modulate TE are mostly involved in gas exchange. Genetic variation exists in crop plants for most of the respective traits, but more research is needed to determine their relative influence on TE under well-watered as well as water-limited conditions. Moreover, research is needed to demonstrate that improvements in TE will improve yields in different environments.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
- Hydraulic Conductance
- Vapor Pressure Deficit
- Leaf Water Potential
- Stomatal Density
- Carbon Isotope Discrimination
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
6.1 Introduction
CO2 acquisition by dryland plants inherently results in water loss. Vast amounts of water are transpired in comparison with the small amounts of carbon that are fixed by photosynthesis. Typical crop plants transpire 200–1,000 g of water per g of assimilated carbon (Martin et al. 1976). With diminishing fresh water supplies, the threat of more frequent droughts due to climate change, and the increasing demand for more food crops, the possibility of increasing “agricultural water productivity” through agronomic and genetic means has received much attention (Araus et al. 2008; Reynolds and Tuberosa 2008; Passioura and Angus 2010). Although conservation agriculture, improved irrigation management, and other agronomic practices have made significant contributions to improving yields under water-limited conditions (Anderson et al. 2005; Turner and Asseng 2005), genotypes that are better matched to their target environments are also needed.
One theoretical avenue for improving yields with less water is through manipulation of the relationship between carbon gain (photosynthesis) and water loss (transpiration). The ratio between these two parameters, called water use efficiency (WUE and see below for full definition), essentially describes how “efficient” the plant is at optimizing carbon gain while minimizing water loss. Plants that are drought tolerant or have evolved in environments with limited available water tend to have higher WUE than plants adapted to conditions with freely available water (Smith et al. 1989). It is, therefore, not surprising that different plant species vary in their WUE (Briggs and Shantz 1913; Rawson and Begg 1977; Siddique et al. 2001). Many species also display plasticity in their WUE (Smith et al. 1989), acclimating to drier conditions, but caution needs to be taken in comparing studies from different environments as differences in transpiration associated with vapor pressure deficits need to be taken into account (Rawson and Begg 1977). Identification that there is intraspecific genetic variation for WUE has encouraged breeders to develop selection programs for improving WUE as a way to improve drought resistance (Farquhar and Richards 1984; Condon et al. 1990; Bonhomme et al. 2009; Barbour et al. 2010; Galmés et al. 2010; Devi et al. 2011). However, traits that are associated with high WUE are often associated with low yield potential, and therefore, there is debate whether selection for high WUE results in higher yield under water-limited conditions (Blum 2005, 2009). Selection for high WUE may only be appropriate for irrigated crops or crops that are grown on stored soil moisture (Blum 2005, 2009). With this caveat in mind, we will explore the traits that are associated with high WUE in this chapter and indicate their relationship to yield production.
Genetic variation in photosynthetic efficiency and transpiration has also driven research to identify mechanisms and genes controlling WUE in the hope of speeding up breeding efforts. Given the multifarious hydraulic and biochemical processes involved in controlling water flow through the plant and photosynthesis rates, many genes and their interaction are likely to be involved in determining the WUE of a plant during development and under water-limited conditions. The problem arises in identifying which of the plethora of genes will have the most impact if their activity is modified in such a complex system. The outlook however is promising, as a few signaling and transcription factors have been identified that control a cascade of subsequent events and development of relevant traits (Masle et al. 2005; Karaba et al. 2007), making these individual genes propitious candidates for genetic manipulation or targeting by breeders. Because these individual genes have the potential to most rapidly influence development of new crop varieties with modified WUE, we will focus on these genes in this chapter.
6.2 Definition of Water Use Efficiency
WUE is used in various ways depending on the level of the observations. At the level of the crop in the field, WUE is usually the ratio of aboveground biomass or economic yield to water use or evapotranspiration (ET):
-
WUEb = aboveground biomass/water use
-
WUEg = grain yield/water use
Water use or ET needs to be measured for the whole growing season. In Mediterranean climatic regions where crops grow on current rainfall and there is no runoff or deep drainage from the soil, growing-season rainfall may be a useful measure of crop water use (French and Schultz 1984a, b; Siddique et al. 1990a, 2001). However, in glasshouse experiments where pots can be weighed and roots extracted, WUE may be on a total biomass basis:
-
WUEt = total biomass/water use
One of the problems associated with the measurement of WUE in the field is that water use or ET includes water loss by soil evaporation as well as transpiration by the crop so that changes in WUE may reflect changes in soil evaporation rather than changes in plant production/transpiration. It is difficult to measure soil evaporation within the crop in the field as it will differ from bare soil evaporation even when measured nearby. In pots it is possible to cover the soil to reduce the soil evaporation, so that water use is now a measure of transpiration. Transpiration efficiency (TE) over the growing season or part of the growing season can be measured where
-
TEt = total biomass/transpiration
-
TEb = aboveground biomass/transpiration
-
TEg = grain yield or economic yield/transpiration
For comparisons between locations or seasons, differences in humidity or vapor pressure deficits will influence the value of the measured transpiration, so that WUE and TE are likewise affected. Average values of the vapor pressure deficit for the period of measurement of transpiration can be used to make the values of WUE and TE comparable.
At the level of the leaf, the instantaneous TE (sometimes incorrectly termed instantaneous WUE) can be measured by gas exchange equipment as the ratio of the rate of net leaf photosynthesis (A N) to rate of leaf transpiration (E):
where p a and p i are the partial pressures of carbon dioxide and e a and e i are the vapor pressure deficits of the air external (a) and internal (i) to the leaf. Integrated over the life of the plant, the losses of carbon by respiration (ϕ c) and the losses of water through the cuticle and partly closed stomata (ϕ w) must be also accounted for when determining TE (Hubick and Farquhar 1989; Turner 1997):
Thus TE is affected by both environmental factors, primarily vapor pressure deficit, and plant factors. It has long been known that some species such as maize and sorghum have much higher values of WUE, and particularly TE, than others (Briggs and Shantz 1913), and this was shown to be due to the higher rates of assimilation and higher ratios of internal to external partial pressures for CO2 in C4 species than C3 species (Tanner and Sinclair 1983). Initially it was considered that the TE of a species did not vary among genotypes (Fischer 1981), but subsequent analysis and measurement demonstrated that TE did vary among genotypes and that it could be selected in breeding programs (Farquhar and Richards 1984; Condon et al. 1990; Richards 2006). Analysis has shown that variation among genotypes can arise from either or both differences in photosynthetic efficiency and differences in stomatal conductance (g s) (Condon et al. 1990). Differences on the discrimination of 12C to 13C due to different diffusion rates through the stomatal pore and uptake within the stomatal cavity can be used to measure integrated TE. Carbon isotope discrimination (CID) also varies with drought, which generally increases TE. Thus, genetic differences in TE are usually identified in well-watered plants in the vegetative phase and not on the seed as differences in harvest index and CID in the conversion of carbon to starch or sugars can cloud differences in TE (Richards 2006).
6.3 Plant Mechanisms and Characteristics That Influence WUE
Figure 6.1 shows plant traits that influence water use and photosynthetic efficiency. Essentially, the plant is a hydraulic conduit between the soil and atmosphere. Canopy and root architecture, organ morphology, anatomy, and aquaporin activity influence a plant’s water uptake capacity, transport efficiency, and water loss. These in turn influence plant water status (Fig. 6.2). A well-hydrated status is needed for growth and many biochemical processes. The trade-off between water loss (transpiration) and CO2 assimilation exists because the atmosphere is almost always drier than the substomatal cavities of leaves. Consequently, when the stomata open, water evaporates from the interstitial tissues into the atmosphere. The four attributes listed above also influence net assimilation rates, but photosynthetic efficiency is also influenced by leaf anatomy and biochemical pathways (Fig. 6.2).

Attributes that influence water use and photosynthetic efficiency

Schematic diagram of the complex interaction between hydraulic and biochemical components that influence transpiration efficiency
The C4 biochemical pathway, in which the first products of photosynthesis are C4 carboxylic acids, and specific bundle sheath anatomy of leaves enable higher rates of photosynthesis than the C3 biochemical pathway. As C4 plants frequently, but not always, have lower stomatal conductances, the TE of C4 species is considerably greater than that of C3 species when directly compared in the same environment. However, C4 species are often grown in warmer and drier environments than C3 species, so that WUE may be similar because of the greater vapor pressure deficits in the warmer and drier environments or seasons. The highest values of TE are found in those plants that fix carbon via the carboxylic acid metabolism (CAM) pathway, whereby the plant opens its stomata and takes up CO2 in the dark when vapor pressure deficits and water loss are low. CAM plants store the carbon as the C4 malate in mesophyll cells and use sunlight to convert the malate to carbohydrates behind closed stomata (Ting 1985). While the CAM biochemical pathway is very efficient in terms of water productivity, dry matter production is very low and is generally found in specialized plants living in extremely dry environments. However, a crop species that has the CAM pathway is pineapple (Ananas comosus), but as some species can switch between CAM and C3 pathways depending on water supply (Dodd et al. 2002), it is likely that pineapple uses the C3 pathway when irrigated for fruit production.
6.3.1 Physical Attributes That Influence Transpiration and Photosynthesis
6.3.1.1 Canopy Size and Architecture
Canopy size and architecture will influence WUE and TE through its effects on ET, E, and A N. For example, more-closed canopies will have higher humidity and, hence, potentially lower rates of transpiration. The water-conserving benefits of a denser canopy may, however, be outweighed by reduced light penetration and lower rates of photosynthesis. Canopy architecture affects light interception and reflectance, such that erectophile canopies with leaves more vertically orientated allow greater light penetration and reflectance during the day due to the angle of the sun, compared with planophile canopies where leaves are orientated more in a horizontal plane (Pinter et al. 1985; Yunasa et al. 1993).
Canopy size and architecture in the field affects ET through soil moisture evaporation. While agronomic methods such as stubble retention and application of mulches can help reduce evaporation from the soil (Yunasa et al. 1994; Richards et al. 2002; Gregory 2004), soil moisture evaporation will also be ameliorated by a crop canopy that shades the soil surface (Siddique et al. 1990b). Soil moisture evaporation may be greater early in the season, before the canopy has fully established, depending on temperature and relative humidity. The trait for early vigor may be a way of establishing canopy cover quicker to minimize soil evaporation (Richards et al. 2002). Wheat genotypes with early vigor traits have been tested in a number of environments (Regan et al. 1992; Turner and Nicolas 1998; Botwright et al. 2002). However, although vigorous growth is supported by higher rates of photosynthesis in wheat, these genotypes appear to have much higher rates of transpiration resulting in lower instantaneous TE in comparison with commercial cultivars (Dias de Oliveira et al. 2013). These genotypes may, therefore, deplete the soil moisture more quickly, leaving insufficient water supply for the reproductive and grain filling stages, which would be a problem for crops growing on predominantly stored soil moisture.
While early vigor may not be suitable where high TE is required, this does not exclude it as a suitable trait in other water-limited environments. For example, in southwestern Australia, winter rainfall during the growing season usually exceeds crop demand for water, but terminal drought during spring and early summer is a common occurrence (Turner and Asseng 2005). Terminal drought is the greatest limitation to crop yields (Boyer 1982; Loss and Siddique 1994; Dracup et al. 1998). Early vigor genotypes flower earlier, thereby avoiding terminal drought, and their high growth rates facilitated by high photosynthesis and transpiration rates mean that there is sufficient biomass to support grain production after flowering despite a shorter vegetative stage (Dias de Oliveira et al. 2012). In this type of environment, more effective use of available water such as early vigor rather than high TE may be the key to improving and sustaining crop yields (Siddique et al. 1989; Turner 1997). Blum (2005, 2009) has argued in support of this paradigm, and Sinclair (2012) has demonstrated that although improvements in TE are possible, it is unlikely to dramatically improve yields, but more effective water use may be a better focus for breeding efforts.
6.3.1.2 Leaf Morphology and Anatomy
The attributes of leaves are probably the greatest determinants of plant TE. Natural variation in the complexity of leaf shape is controlled in most eudicots by expression of a KNOX1 gene (Bharathan et al. 2002; Kimura et al. 2008), but monocots have only simple leaves where KNOX is downregulated. Auxin is also involved in regulating whether leaf margins are serrated or lobed and many other genes are, of course, involved in regulating the length and width of leaf laminas (see review by Nicotra et al. 2011). The function of leaf shape (excepting leaf size) is still a matter of debate (Nicotra et al. 2011) and is mainly relevant in an ecological context, but it should not be dismissed completely from this discussion because the water relations of leaves are strongly influenced by leaf shape (Nicotra et al. 2011), and therefore, manipulation of leaf shape may be an avenue to improving TE in crop species with complex or lobed leaves.
The size of a leaf, its color, orientation, and its topology determine the amount of light intercepted and its radiation and heat load. Small and thick leaves are associated with drought tolerance and high TE (Abrams et al. 1990, 1994). In many crop species, leaf size decreases with water deficit, and in fact, leaf expansion is one of the most sensitive processes to water stress (Boyer 1970; Hsiao et al. 1985). Leaf thickness also tends to increase in many crop species in response to water deficit, particularly genotypes with greater drought tolerance and higher TE (Nobel 1980; Wright et al. 1994). Increasing leaf thickness is associated with water conservation. Increasing leaf thickness will increase the boundary layer thickness and, hence, decrease the rate of evaporation from the leaf (see below), but boundary layer thickness will be conversely reduced by a reduction in leaf size. Thicker cell walls, as well as increased suberization of bundle sheaths, help protect from dehydration but would probably also decrease leaf hydraulic conductance (see below). Leaf thickness also contributes to higher TE through an increase in A N (Dornhoff and Shibles 1976; Schulze et al. 2006) due to greater abundance of photosynthetic apparatus.
Topological features such as pubescence and glaucousness are associated with drought tolerance and high TE in xerophytes, and many plant species adapted to semiarid environments and environments with seasonal rainfall (Mulroy 1979; Ehleringer 1980; Ehleringer and Clark 1987; Rieger and Duemmel 1992; Abrams et al. 1994). Despite the limited research on pubescence and glaucousness in crop species providing no clear indication that they will lead to improvements in yield by increasing TE, there is increasing recent interest in selection of glaucousness and pubescence for drought tolerance by plant breeders (Reynolds et al. 2001).
Depending on their density and length, leaf trichomes help to keep the surface of the leaf more humid, thereby reducing transpiration, and trichomes may also help reduce the heat load of a leaf by reflecting light (Ehleringer 1980, 1984). Trichome density varies within and across species (Ehleringer 1984), and trichome density of some species and genotypes increases in response to water deficit (Ehleringer 1982). Varying trichome density has been observed in several crop species (Sharma and Waines 1994; Webster et al. 1994; Sagaram et al. 2007). Trichomes, like leaf thickness, increase the thickness of the undisturbed air surrounding the leaf, that is, the boundary layer (Schuepp 1993). The conductance of water vapor across the boundary layer decreases with increasing boundary layer thickness, and therefore, the rate of evaporation from the leaf surface will be lower (Nobel 2009). However, the conductance of CO2 across the boundary layer will be similarly reduced, and so the presence of leaf trichomes may negatively impact on photosynthetic rate. It should be noted, however, that the boundary layer resistance to CO2 diffusion will only impact photosynthetic rates if it is greater than stomatal and internal resistance to CO2 (Nobel 1999). Selecting for leaf pubescence in crops would, therefore, only have a beneficial effect on TE if the increased resistance to water vapor diffusion out of the leaf was greater than the increased resistance to CO2 diffusion into the leaf. Pubescence as a trait may be beneficial in the drier climates expected for the future because greater atmospheric CO2 due to climate change will increase the gradient for CO2 diffusion into the leaf. Leaf pubescence in Brachypodium distachyon has been reported to be under the control of a single dominant gene, SPUB (Garvin et al. 2008), which provides the opportunity to manipulate expression of this trait and evaluate its influence on the physiology of a species closely related to the major crops wheat and barley.
The aboveground parts of almost all land plants are covered by a waxy cuticle, which helps reduce evaporation from the nonstomatal epidermis. Transpiration across the leaf cuticle varies between species and across genotypes probably due to differences in thickness and hydrophilic properties of the epicuticular wax (Jordan et al. 1984; Larsson and Svenningsson 1986; Svenningsson and Liljenberg 1986; Clarke and Richards 1988; González and Ayerbe 2009). As cuticular transpiration is usually measured when the stomata are assumed to be closed in the dark, it needs to be recognized that some of the putative variation in cuticular transpiration may be the result of incomplete stomatal closure. High glaucousness and low cuticular transpiration are associated with drought tolerance, which can vary with environmental conditions depending on the species (Oppenheimer 1960). Glaucousness may also help in reflecting light to reduce radiation load. Evidence in support of this comes from a study on durum wheat lines isogenic for glaucousness (Richards et al. 1986). Glaucous lines had slightly cooler leaf temperatures under water deficit in the field, compared with nonglaucous wheat lines (Richards et al. 1986). In some species, transpiration through the stomata can increase with glaucousness (Kerstiens 1997, 2006), which would cool the leaf surface, but Richards et al. (1986) reported similar transpiration rates for glaucous and nonglaucous lines. Glaucousness of wheat flag leaves had no effect on TE, or photosynthetic and transpiration rates, but increased leaf duration under drought (Richards et al. 1986). Although glaucousness reduced photosynthesis and transpiration rates of wheat ears, the greater proportional reduction in transpiration resulted in increased TE of ears in comparison with nonglaucous ears (Richards et al. 1986).
Glaucousness did not correlate with yield or yield components in a doubled haploid population derived from a cross between a drought-tolerant and a relatively drought-intolerant bread wheat line (Bennett et al. 2012), despite glaucousness of parental material being associated with higher TE and yield under water deficit (Izanloo et al. 2008). The disparity between these two studies may be related to differences in behavior in glasshouse compared with field conditions, although Bennett et al. (2012) speculated that their method of visually scoring for glaucousness may not correlate with physical wax content, especially those lines that were given mid-high scores, as was observed by Clarke et al. (1993). Epicuticular wax content should also have been measured (Bennett et al. 2012). In barley breeding lines, increased drought resistance and greater yields compared with commercial cultivars were associated with greater epicuticular wax load (González and Ayerbe 2009). Water deficit increased the epicuticular wax load and reduced nonstomatal transpiration. Epicuticular wax load was negatively associated with nonstomatal transpiration and positively associated with grain yield (González and Ayerbe 2009). The negative association between epicuticular wax load and nonstomatal transpiration has also been observed for sorghum (Jordan et al. 1984), but the converse was observed in earlier studies on barley and oats (Larsson and Svenningsson 1986; Svenningsson and Liljenberg 1986).
Stomata are small pores on leaf and stem surfaces that facilitate the exchange of gases between the leaf interior and the atmosphere. They are an important determinant of TE, being physically located as the gateway controlling the trade-off between carbon capture and water loss, and, as such, have warranted the greatest amount of research. Stomatal conductance has a greater effect on transpiration than photosynthesis, so maximum TE will occur through coordination of stomatal opening and capability for CO2 fixation (Nobel 2009; Yoo et al. 2009).
Stomatal density and size of their aperture determine a leaf’s conductance to CO2 and water vapor (assuming gas exchange across the nonstomatal epidermis is negligible). Stomatal density increases stomatal conductance to water vapor (g s) of a leaf in a linear dependent manner, but conductance to water vapor through a single pore increases with radius of the aperture to the fourth power, i.e., doubling the number of stomata will double g s, but doubling stomatal aperture (keeping density the same) will increase g s 16-fold. Hetherington and Woodward (2003) showed that there is a strong negative curvilinear relationship between stomatal density and stoma size across a range of plant groups and that the relationship between g s and A N approximately fit within one of two curvilinear relationships depending on whether the plants have a C3 or C4 photosynthetic pathway. Stoma aperture size is regulated by turgor pressure of the flanking guard cells. An uptake of ions (K+ and Cl−1) (Turner 1972) generates the osmotic gradient to induce water uptake, which probably occurs through aquaporins (Heinen et al. 2009). Guard cell volume increases with turgor pressure causing the stoma to open due to radial orientation of the cellulose microfibrils of the guard cells walls, causing the cells to swell more in one direction. The reverse happens to cause a stoma to close. Guard cell length, therefore, ultimately determines aperture size of the stoma. It has been suggested that small stomata can open and close more rapidly and by being generally associated with high densities provide the capacity for rapid changes in g s with fluctuating environmental conditions (Hetherington and Woodward 2003).
Stomatal density is under strict genetic control, but the number of stomata can also be influenced by environmental factors (see review by Casson and Gray (2008) and Sect. 6.4.1). Stomatal density tends to decrease when plants are grown under elevated atmospheric CO2 and has been shown to be modulated by the HIGH CARBON DIOXIDE gene in Arabidopsis. Light intensity also induces changes in stomatal density in developing leaves, but the genes regulating the response are not yet known (Casson and Gray 2008). Drought can influence stomatal density of developing leaves, depending on the species (Quarrie and Jones 1977; Clifford et al. 1995; Xu and Zhou 2008; Hamanishi et al. 2012), but this may be confounded by inhibited leaf expansion at low soil water potentials (see above).
Stomatal opening depends on the water status of the plant and is influenced by irradiance, CO2 concentration, relative humidity, phytohormones such as ABA, and stress (see reviews by Buckley 2005; Shimazaki et al. 2007). Evapotranspirational demand (temperature, relative humidity, and wind speed) also determines g s in field conditions. Variation in g s across species in response to soil water deficit has been observed (Henson et al. 1989a). Stomatal closure occurred in lupin, but not wheat, before changes in the leaf water potential were detected. The disparity in the responses was later attributed to differences in sensitivity to ABA (Henson et al. 1989b). However, field-grown wheat has also been observed closing stomata before leaf water potential decreased under drought (Ali et al. 1999), indicating that there may be genotypic variation in ABA production and/or stomatal sensitivity to soil water deficit. Genotypic variation in the relationship between g s and leaf water potential under water-limited conditions has been observed for a number of other crop species (Ackerson et al. 1980; González et al. 1999).
Modern phenotyping programs are using canopy temperature, as assessed through thermal imaging, as a surrogate for selecting genotypes with high g s (Jones et al. 2009; Munns et al. 2010). Lower canopy temperatures have correlated well with greater yields in genotypes of wheat (Reynolds et al. 2007) and rice (Garrity and O’Toole 1995), but not soybean (Harris et al. 1984). Low canopy temperature was also correlated with yield in a comparison between four legume species (Pandey et al. 1984), whereas lower canopy temperature was associated with higher susceptibility to drought in genotypes of potato (Stark et al. 1991). Lower canopy temperatures imply higher g s facilitating transpirational cooling and favoring net photosynthesis. Higher g s also indicates enhanced capacity for water uptake or better ability at maintaining plant water status. In these situations, low TE may in fact be selected since canopy temperature depression is related to transpirational cooling. Transpiration is linearly related to g s, whereas photosynthesis saturates at high g s (Yoo et al. 2009) due to metabolic limitations of carbon assimilation and fixation (Farquhar and Sharkey 1982). The ideal then for improved TE is to select genotypes with less than maximum g s, when the slope of the relationship between A N and g s is greater than the slope of the relationship between E and g s (Yoo et al. 2009).
The internal anatomy of the leaf determines the pathway for water flow from the petiole to the substomatal cavities and the distance for diffusion of water vapor from the evaporation sites to the external atmosphere as well as the diffusion distance for CO2 to reach the photosynthetic apparatus. These properties, therefore, determine leaf hydraulic conductance (water transport efficiency) and internal conductance to CO2, respectively, although measurements of the internal conductance to CO2 also include the permeability of mesophyll plasma membranes to CO2.
The reader is referred to Sack and Holbrook (2006) for an in-depth review of leaf hydraulics, but we will highlight aspects relevant to TE in crop plants for the purpose of this chapter. Leaf hydraulic resistance accounts for up to a third of the total plant resistance to liquid water transport (Sack and Holbrook 2006), which is an overlooked factor potentially affecting TE of crops (see more detailed discussion below on roots, regarding constraints of hydraulic resistance to gas exchange). Leaf hydraulics has mainly been studied in tree leaves in an ecophysiological context, but many aspects would also be applicable to crop species. The hydraulic conductance of leaves from annual crop species is apparently two- to threefold greater than woody species, although annual crop species accounted for only 7 % of the species so far examined (Sack and Holbrook 2006). The high hydraulic conductance of crop leaves may account for the small differences in water potential observed between leaves and stems (Tsuda and Tyree 2000). Leaf hydraulic conductance is variable, changing with development and environmental conditions (Tsuda and Tyree 2000; Brodribb and Holbrook 2004; Nardini et al. 2005; Simonin et al. 2012). Irradiance can also stimulate increased hydraulic conductance (independently of stomatal opening) in some species, which has been attributed to increased aquaporin activity (Cochard et al. 2007; Baaziz et al. 2012). Although not yet shown to involve aquaporins, the hydraulic conductance of sunflower shoots was on average fourfold higher when the leaves were illuminated than under ambient light in the laboratory, but only when leaves were well hydrated (Guyot et al. 2012). The function of this irradiance-induced stimulation of leaf hydraulic conductance is not yet fully understood but when combined with stomatal regulation of g s provides flexible regulation of leaf water status (Guyot et al. 2012).
The myriad of leaf hydraulic properties and responses reflects differences in anatomy, principally vein density, and extravascular pathways for water flow (Cochard et al. 2004; Brodribb et al. 2007). When water enters the leaves, it travels through the veins, the density and pattern of which determine the distribution of water within the leaf. Water distribution (and assimilate translocation) within C4 grass leaves is superior to C3 grasses because of a more denser network of the smaller longitudinal and transverse veins (Ueno et al. 2006). Water exits the major or minor veins into the bundle sheath, but then its journey is less clear (Sack and Holbrook 2006). Water may travel apoplastically, around the cells, or transcellulary, through the cells of the mesophyll. Transcellular water transport may occur through the symplast via plasmodesmata or across aquaporin-embedded membranes. The contribution of these pathways is not yet known, but the rapid and reversible changes in leaf hydraulic conductance stimulated by temperature or irradiance indicate significant transcellular transport in those species. Moreover, suberin deposition in the walls of the bundle sheath, such as those found in the mestome sheath of wheat and oats (O’Brien and Carr 1970), may obstruct apoplastic water flow. The sites of water evaporation within the leaf are also not yet known but will likely be determined by anatomy and the pathway for water flow through the leaf.
Soybean is one of the few crop species where genotypic differences in leaf hydraulics have been examined. High diversity of leaf morphological features was observed in soybean cultivars, where g s was significantly correlated with stomatal density (Tanaka et al. 2010). The greater productivity of US cultivars compared with Asian cultivars of soybean was attributed to greater g s due to greater stomatal density. Interestingly, US cultivars had slightly smaller guard cells, smaller leaves, and higher vein density, which indicate that leaves of US cultivars also potentially had greater water transport efficiency. Conversely, a low leaf hydraulic conductance contributed to higher TE in a soybean genotype through water conservation when transpirational demands were high (Sinclair et al. 2008). Either high or low hydraulic conductance may, therefore, aid adaptation to drought, the significance of either being dependent on coordination with other leaf properties.
The internal conductance to CO2 (g i) is the remaining part of this discussion about leaf properties to be considered with regard to TE of crop plants. The greatest physical constraint to photosynthetic rates, excepting the biochemical process, was long considered to be the stomatal resistance to CO2 diffusion into the leaf. However, g i is also a major constraint (Warren 2008). The internal conductance includes the diffusion of CO2 once inside the leaf to the sites of carboxylation. The diffusion distance through the intercellular air spaces of the mesophyll is, therefore, a major determinant of g i as well as the permeability of the mesophyll membranes to CO2. The dissolution of gaseous CO2 to the liquid phase at the mesophyll cell wall and diffusion across the cytoplasm and chloroplast envelope should also not be discounted.
There is high variability among species in the relative limitation of A N due to g i, but generally A N increases with increasing g i (see review by Warren 2008). Herbaceous species tend to have higher g i, so the capacity for manipulating this property to improve TE, by reducing a constraint to A N, in crop species may be limited, but g i will be influenced by anatomical changes in response to water stress. Moreover, aquaporin-mediated CO2 diffusion and carbonic anhydrase activity can change g i rapidly (Flexas et al. 2007).
There are few genotypic comparisons of g i but we can make some inferences from anatomical studies. In flag leaves of diploid, tetraploid, and hexaploid wheat and Aegilops species, the surface area of the mesophyll cells per unit volume of mesophyll tissue was similar, but diploid genotypes had the highest photosynthetic rates at light saturation (Austin et al. 1982; Kaminski et al. 1990). This occurred despite thinner leaves and lower chlorophyll abundance. Diploid genotypes may have had a greater g i due to shorter diffusion distance between the leaf surface and carboxylation sites in thin leaves; diffusion across the cytoplasm would also have been shorter because diploid mesophyll cells were smaller. Interestingly, greater efficiency in the import of nutrients and export of assimilates was also related to improved productivity as vein density was also positively correlated with A N (Austin et al. 1982). Light-saturated A N was also negatively correlated with mesophyll cell size in ryegrass leaves, but this was dependent on growth temperatures (Wilson and Cooper 1969). Conversely, slower growth rates and lower photosynthetic rates were correlated with thinner leaves in bean genotypes (Sexton et al. 1997). However, RuBisCO may have been limiting in bean genotypes with lower photosynthetic rates, as indicated by the lower leaf nitrogen contents.
6.3.1.3 Stem Anatomy
Apart from the obvious structural role in supporting leaves above the ground, stems are the conveyors of water, nutrients, and photosynthetic assimilates between roots and leaves. Their anatomy, especially the vascular bundles, will determine transport efficiency which, similar to roots and leaves, will influence TE. The primary structure of stems is highly variable with the vascular tissue forming either a continuous cylinder sandwiched between the cortex and pith or a cylinder of discrete strands within the undifferentiated ground tissue, or in monocots such as maize, the vascular bundles may be spread throughout the ground tissue. Stems may also be involved in storage and remobilization of assimilates or water, particularly in response to abiotic stress. Remobilization of water-soluble carbohydrates is a major contributor to drought adaptation in a number of crop species (McIntyre et al. 2012), but it can confound measurements of TE if the whole plant biomass at maturity is considered, and therefore, TE is usually measured during the vegetative stages.
Within the vascular bundles, the number of xylem vessels varies. Xylem vessels are analogous to pipes and hence, their abundance and diameters largely determine the hydraulic conductance of a stem. Accordingly, the hydraulic conductance of stems is generally much greater than leaves or roots (Steudle 2000). Studies on the resistance to lodging indicate that there is high genetic variation for the number of vascular bundles in grass species, including the major cereal crops (Jellum 1962; Dunn and Briggs 1989; Khanna 1991; Kaack et al. 2003). Genetic variation in vascular bundles also exists in other crop species. In tomato, thicker stems with more conducting tissue (xylem vessels) were related to drought resistance (Kulkarni and Deshpande 2006).
Water flow through the stem, and for that matter roots and leaves, can be disrupted if tensions in the xylem become large and cavitations occur. The susceptibility of xylem to cavitation-induced hydraulic disruption depends on the species, but most research has examined this phenomenon in woody species. Xylem cavitation can occur in annual crop species at fairly high water potentials in comparison with woody species (Tyree et al. 1986). However, herbaceous species tend to generate high root pressures when they are not transpiring (Steudle and Jeschke 1983; Bramley et al. 2010), and so it is possible that embolized vessels may be refilled during the night. Although embolisms may be reversed, when cavitation occurs, water transport to the leaves will be disrupted, which will ultimately affect crop productivity. The occurrence and susceptibility of crop species to hydraulic disruption should, therefore, be investigated.
6.3.1.4 Root Architecture and Anatomy
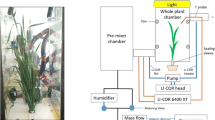
The architecture of a root system, or spatial distribution within the soil, is described by its length and branching pattern, which in turn are determined by the rate, angle, and duration of root growth. The root system architecture, therefore, determines the capacity for water and nutrient uptake from the soil (Lynch 1995, 2007). Root systems of different species display a high degree of phenotypic plasticity, responding to environmental conditions, but developmental instability has also been proposed as another adaptive trait generating nongenetic variation (Forde 2009). Intraspecific variation in root system architecture of major crop species is receiving increasing interest as a potential means to improve water and nutrient uptake efficiency (Ratnakumar and Vadez 2011; Chen et al. 2012; Wasson et al. 2012), but the fact that roots are below ground and the soil environment is generally heterogeneous presents an enormous challenge in root research. Phenotyping is, thus, generally undertaken on roots growing in artificial medium (Chen et al. 2011; Clark et al. 2011) or in glass-walled chambers (Liao et al. 2006; Manschadi et al. 2006; Bramley et al. 2011). Moreover, breeding programs need to ensure that traits identified in the laboratory are translated into the field (Wasson et al. 2012).
In order to identify which root traits are relevant for improving TE, we need to consider the target environment because this will determine temporal and spatial variation in available water, as well as physical and chemical constraints to root growth. For example, crops growing on deep stored moisture may need long roots and high branch-root density at depth (Wasson et al. 2012), whereas high shallow-root density may be more advantageous for crops growing on sandy soils relying on intermittent in-season rainfall (Manschadi et al. 2006). There is considerable genetic variation for root length density that could be exploited (Siddique et al. 1990a; Mwanamwenge et al. 1998; Chen et al. 2011; Ratnakumar and Vadez 2011).
We also need to consider the particular species when selecting root traits for high TE because root hydraulic properties determine the region of root involved in water uptake and the transport efficiency of the root system (hydraulic conductance), which vary between species. For example, cereal roots absorb water preferentially in a region just behind the root tip, whereas legumes absorb water along the entire root length (Bramley et al. 2009). The hydraulic conductivity of cereal roots will, hence, be influenced by the number of branches, whereas the hydraulic conductivity of legume roots will be influenced by total root length.
The root system is a major constraint to gas exchange and, consequently, TE because it forms the largest resistance to liquid water flow in the plant (Steudle 2000). Root hydraulic conductivity needs to be high enough to supply the leaf with sufficient water to maintain leaf hydration so stomata can remain open. High root hydraulic conductivity will also minimize the drop in water potential needed to drive water uptake from the soil and transport it to the shoot. However, root hydraulic conductivity also needs to be variable and coordinated with stomatal opening and environmental conditions to maximize hydraulic efficiency; otherwise, the plant runs the risk of losing water to the soil when it becomes extremely dry or excessive water loss to the atmosphere if the vapor pressure deficit is high. While water transport is a passive process, roots themselves are not passive structures.
Hydraulic conductivity of a root system is determined by morphology, anatomy of individual roots, and aquaporin activity. All roots are cylindrical, comprising several concentric tissue layers: epidermis, exodermis (if present), cortex, and endodermis. The endodermis surrounds the stele, which contains the pericycle (origin of lateral roots), parenchyma, and vascular tissue. The outer surface of the epidermis, and walls of the exodermis and endodermis, can contain lignin or suberin, which reduces the permeability of the apoplast to water and nutrients (Steudle et al. 1993; Zimmermann et al. 2000). Development of these wall modifications depends on the species, developmental stage, and environmental conditions (Barrowclough et al. 2000; Zimmermann et al. 2000; Bramley et al. 2009; Vandeleur et al. 2009). An exodermis does not develop in wheat, barley, or perennial ryegrass (Perumalla et al. 1990) probably because the root cortex dies back in mature roots (Hamblin and Tennant 1987), revealing the heavily suberized endodermis. Lupin and chickpea roots also do not form an exodermis, but this has not been tested in response to soil water deficit (Hartung et al. 2002; Bramley et al. 2009). Increased suberization of the exodermis and endodermis has been observed in many crop species in response to water deficit, especially woody perennials, and was correlated with decreased root hydraulic conductivity (Lo Gullo et al. 1998; Rieger and Litvin 1999; Vandeleur et al. 2009). Maize is one of the few annual crop species where an exodermis has been reported, and it becomes more suberized under drought (Stasovski and Peterson 1991). The fact that increased exodermal suberization can also be stimulated by growing maize roots in a mist culture has demonstrated the reduction in root hydraulic conductance caused by this feature (Zimmermann et al. 2000).
Air-filled spaces called aerenchymas can also develop in the cortex, particularly in adventitious (nodal) roots, in response to waterlogging-induced oxygen deficiency, some nutrient deficiencies, and occasionally water deficit (Drew et al. 1989; Stasovski and Peterson 1991; Colmer 2003; Zhu et al. 2010). Aerenchyma development may be a more common root response to soil water deficit than has been reported because very few field studies have examined root anatomy, and cell plasmolysis or death occurs with dehydration. Aerenchymas also serve as radial barriers to water and nutrient transport (see review by Bramley and Tyerman 2010), but they also reduce the plant’s respiratory costs (Zhu et al. 2010). Aerenchyma development, then, may be a beneficial trait for drought tolerance (Zhu et al. 2010) and even TE but only if water is preferentially absorbed in a region close to the root tip, where aerenchymas do not form.
Principal anatomical differences occur between monocotyledon and eudicotyledon roots, especially in vasculature development and because dicots have secondary growth (Klepper 1983). Monocot roots generally develop a ring of small xylem vessels surrounding a central pith or large xylem vessel, whereas xylem development in dicot roots initially occurs in poles and develops centripetally in a diarch or tetrarch pattern. Consequently, the axial hydraulic conductance of dicots is generally greater than monocots, and axial hydraulic conductance increases with root length in dicots because of increasing vessel development (Bramley et al. 2009).
Genetic variation in xylem vessel diameter has been exploited in wheat to reduce axial hydraulic conductance (Richards and Passioura 1989). The philosophy was based on the idea that reduced axial hydraulic conductance would reduce the rate of water uptake, conserving soil water for later in the season when it would be available for grain filling. Passioura (1972) first tested this theory by growing a wheat cultivar entirely on stored soil moisture and removing all but one seminal root to simulate reduced axial conductance, as all water uptake must flow through the one remaining central xylem vessel (discounting the ring of smaller vessels). Nodal roots were allowed to grow but were mostly restricted from accessing the soil moisture. Single-rooted plants compensated for the initially small root system with increased growth rate and branching, so that total root mass at maturity was the same as the control plants. Single-rooted plants initially maintained a similar water use as the control plants, but after a few weeks and until after anthesis, plant water use was lower than the control plants. Single-rooted plants ultimately had greater grain yield because control plants ran out of water for grain production. Passioura (1972) did not measure root hydraulic conductance or other physiological parameters, but he speculated that the estimated threefold reduction in axial conductance (three seminal roots reduced to one) would have caused stomatal closure due to much lower leaf water potentials, thereby conserving water. However, translocation of stored carbon in response to water deficit also probably contributed to grain yield (Passioura 1976). Remobilization of water-soluble carbohydrates is a well-recognized drought-adaptive trait in wheat (Reynolds et al. 2009). Moreover, partial excision of roots in young wheat plants increased hydraulic conductivity of the remaining root, probably by an ABA-induced change in aquaporin activity (Vysotskaya et al. 2004), so single-rooted wheat plants growing on stored moisture may be more efficient in water use through a number of mechanisms.
Reducing the diameter of the central xylem vessel by 10 μm through plant breeding increased wheat yield in the field by about 10 % in one of the selected genotypes, in comparison with unselected controls, but only in the low-yielding or drier environments (Richards and Passioura 1989). Theoretically this would reduce axial hydraulic conductance of the seminal root system by about half (vessel diameter decreased from 65 to 55 μm). It is not clear what effect the selection for narrower xylem vessels in seminal roots had on morphology and anatomy of nodal roots, anatomy of stems, or other physiological processes such as gas exchange and leaf water status. Moreover, the yield increase was not observed in an environment relying on in-season rainfall. Despite narrower xylem vessels being a potential drought-adaptive trait for environments relying on stored soil moisture, no current commercial wheat varieties appear to have been developed with narrower xylem vessels. Subsequent investigations of genetic differences in xylem anatomy in rice have led to the conclusion that manipulating vessel radii will not improve drought resistance, but in that study the aim was to increase axial hydraulic conductance (Yambao et al. 1992). Conservative water use has also been shown to be important in tolerance of chickpea genotypes to drought, which was correlated with lower g s but not root growth components (Zaman-Allah et al. 2011) and suggests that either greater sensitivity to ABA or lower root hydraulic conductance induced stomatal closure.
Surprisingly, studies on manipulating xylem anatomy to improve drought tolerance or TE have discounted the fact that it is the radial, not axial, hydraulic conductance that mostly constrains total root hydraulic conductance (Steudle and Peterson 1998). When water is taken up by the root, it has to cross the cylinder of living tissue described above, either by crossing membranes or the symplast (via plasmodesmata) or through cell walls (apoplast), and it is for this reason that the radial hydraulic conductance is several orders of magnitude less than the axial conductance. The radial pathway, though, is the main source attributing to variable root hydraulic conductance through anatomical changes (described above) and changes in aquaporin activity (see Sect. 6.4.4), depending on the pathway taken by water across the root radius. If water crosses membranes at some point in the radial pathway, aquaporins can facilitate rapid and reversible changes in hydraulic conductance. Of greatest significance to TE are the aquaporin-mediated changes in hydraulic conductance that can match transpirational demands (Henzler et al. 1999; Vandeleur et al. 2009), but also those that improve water transport efficiency in response to soil water deficit (Sade et al. 2010). However, more research is needed linking aquaporins to their physiological roles.
Differences in radial root anatomy and aquaporin activity have been found between genera, closely related species, and cultivars (Bramley et al. 2009; Vandeleur et al. 2009). Genotypic differences in root anatomy have also been observed in a number of crop species (Saliendra and Meinzer 1992; Zhu et al. 2010). However, the only study linking genotypic differences in aquaporin activity to root hydraulics within a species has been undertaken on accessions of Arabidopsis (Sutka et al. 2010).
6.3.2 Biochemicals and Enzymes
6.3.2.1 ABA and Sensitivity to ABA
The phytohormone abscisic acid (ABA) is an important stress hormone produced in the roots as the soil dries (Davies and Zhang 1991; Turner and Hartung 2012) and is transported in the xylem to the leaves where it closes the stomata (Henson et al. 1989b; Zhang and Davies 1989, 1990). It is, therefore, an important hormone in controlling water loss. While ABA triggers stomatal closure in the leaves, it also induces a reduction in leaf growth (Davies et al. 2005), an increase in root growth at low water potentials (Spollen et al. 2000), and an increase in the hydraulic conductivity of roots (Hose et al. 2000; Aroca et al. 2006), all of which provide an integrated response to water shortage. Although the influence of ABA on root hydraulic conductivity may be transient (Hose et al. 2000) or inhibitory (Fiscus 1981), ABA production has been shown to increase yields of wheat under moderate stress conditions (Travaglia et al. 2010) and improve TE in tree seedlings, particularly when subject to a water deficit (Duan et al. 2007). As there is genetic variation in the response of stomata to ABA concentration (Henson and Turner 1991), this provides another genetic mechanism for differences in TE to be expressed (see Sect. 6.4.4 for further details).
6.3.2.2 Isotopic Discrimination
Farquhar et al. (1982) first developed the theory that predicted that in C3 plants the discrimination of the small amounts of the naturally occurring stable isotope of carbon, 13C, in photosynthesis will be least in those plants that fix the most carbon per unit amount of water transpired, i.e., in those that have the highest TE. This was confirmed by Farquhar and Richards (1984) who showed that the ratio of 13C to 12C in the leaves of wheat genotypes was correlated with the TE measured from changes in dry weight and water use for the period before sampling for CID. This relationship between CID and TE has been shown to exist in a range of crop species (Turner 1993), but not all crop species (Turner et al. 2007b). The relationship has been used to select wheat genotypes that yield better under drought (Richards 2006).
6.3.2.3 Maintenance of Turgor and Leaf Water Potential
Crop plants have been shown to maintain their turgor potential/pressure as the plant water status decreases by the accumulation of solutes (Jones et al. 1980; Turner and Jones 1980), a process known as osmotic adjustment or osmoregulation (Turner and Jones 1980; Morgan 1984). Wheat with genes for high osmotic adjustment has been shown to have increased yields in water-limited environments (Richards 2006). However, osmotic adjustment in chickpea was shown to be not highly inherited, and the degree of osmotic adjustment was not associated with yield under dryland conditions in the field (Turner et al. 2007a). Other crop species such as lupin show limited capacity for osmotic adjustment, and they are relatively intolerant to drought (see review by Palta et al. 2012).
Changes in tissue elasticity is another feature of adaptation to drought (Bowman and Roberts 1985; Lo Gullo and Salleo 1988). The properties of cell walls predominantly determine leaf elastic modulus and, therefore, water-holding capacity. Leaf density correlates strongly with bulk leaf elastic modulus (Niinemets 2001). The capacity to reduce leaf elastic modulus in response to soil moisture deficit enhances the water-holding capacity and when combined with osmotic adjustment increases turgor maintenance under lower relative water contents (Jensen and Henson 1990). High water-holding capacity, therefore, aids in maintaining leaf water status. Conversely, with a high leaf elastic modulus (low elasticity), a leaf’s water potential decreases more for a given change in tissue water content, which may aid in water uptake from dry soils (Bowman and Roberts 1985) or may induce stomatal closure. Measurement of leaf elastic modulus is laborious, which is probably the reason why few crop studies have investigated this attribute, but genetic diversity exists (Johnson et al. 1984; Rascio et al. 1988). Moreover, leaf elastic modulus and hydraulic conductance may not be mutually exclusive properties because a high water-holding capacity would buffer against water loss when the root or shoot resistance to water flow is high. Inferences about leaf hydraulic capacitance can be made by plotting leaf water potential against transpiration rates; hysteresis between dehydration and rehydration indicates that either the hydraulic conductance is variable or stored water is initially used in transpiration (Zhang and Davies 1989). Whether these relationships are relevant to crop species, particularly annual crops remains to be seen.
6.3.2.4 Photosynthesis and Respiration
Earlier, TE was shown to be a ratio between the rates of A N and E. While this is controlled at the level of the leaf by leaf conductance to CO2 and water vapor, A N is also affected by other anatomical and biochemical factors in the carbon assimilation pathway. Measurement under similar light intensity reveals variation in the rate of leaf net photosynthesis among plant and crop species (e.g., Turner et al. 1984; Leport et al. 1998). The reasons for this variation can be several. The concentration of chlorophyll that captures the light energy may vary, but as the rate of photosynthesis is maintained over a relatively wide range of leaf chlorophyll concentrations, a low concentration may not necessarily result in lower rates of photosynthesis. The conversion of CO2 to carbohydrates is not the subject of this chapter, but with the energy from sunlight captured in the thylakoid membranes of the chloroplasts, the CO2 is converted into C3 sugars via a series of reactions in the Calvin cycle. One of the key enzymes in the Calvin cycle is ribulose-1,5-bisphosphate carboxylase/oxygenase (RuBisCO), which comprises 50 % (20–30 % of the total leaf nitrogen) of the soluble protein in the leaf of C3 plants and 30 % (5–9 % of the total leaf nitrogen) of the soluble leaf protein in C4 plants. However, it is notoriously inefficient in converting ribulose-1,5-bisphosphate to glycerate-3-phosphate and glucose, and therefore, there has been considerable effort to increase the efficiency of the enzyme and, hence, the photosynthetic efficiency of crop plants (Spreitzer and Salvucci 2002; Parry et al. 2003). Although Spreitzer and Salvucci (2002) concluded that this will not be easy due to the range of atomic structures of the enzyme, attempts to genetically modify the enzyme and increase photosynthetic efficiency have continued (see Masle et al. (2005) and discussion in Sect. 6.4.1), but with limited success to date in crop plants. In the presence of high concentrations of CO2 in the leaf, RuBisCO converts the CO2 to sugars, but if the CO2 concentration decreases, for example, when stomata close with water deficits, RuBisCO also has an affinity for oxygen and reduces ribulose-1,5-bisphosphate to glycerate-3-phosphate at a reduced rate and greater metabolic cost. This process is known as photorespiration and is estimated to reduce photosynthesis by as much as 25 % in C3 plants (Sharkey 1988).
However, photosynthetic efficiency is much higher in C4 plants because they are able to increase the CO2 concentration in the leaf, thereby reducing the oxygenation of ribulose-1,5-bisphosphate and reducing the rate of photorespiration. In order to bypass the photorespiration pathway, C4 plants have a mechanism to efficiently deliver CO2 to the enzyme RuBisCO. They have a different anatomy from C3 plants in that chloroplasts exist not only in the mesophyll cells but also in specialized bundle sheath cells that surround the leaf xylem. Instead of direct fixation to RuBisCO via the Calvin cycle, CO2 is incorporated into the 4-carbon oxaloacetate by the enzyme PEP carboxylase and then converted to malate, which can be transferred to the bundle sheath cells that have the ability to regenerate CO2 in the chloroplasts. Bundle sheath cells can then utilize this CO2 to generate carbohydrates by the C3 pathway (Slack and Hatch 1967). There have been many attempts to increase the photosynthetic efficiency of C3 plants by the incorporation of the C4 photosynthetic pathway, but to date all attempts have been unsuccessful.
Respiration, the conversion of carbohydrates back to CO2 with the release of energy, is required for growth, metabolism, and nutrient uptake. It consumes between 25 and 70 % of the carbon fixed and also varies among species and genotypes (Lambers et al. 2005). Attempts to reduce carbon losses without affecting growth have met with little success.
Finally, as mentioned above, the specialized leaf anatomy of C4 plants enables them to fix CO2 more efficiently than C3 plants. Additionally, leaf anatomy such as thicker leaves also affects the rate of photosynthesis. Schulze et al. (2006) compared 65 species of Eucalyptus at 73 sites in Australia and showed that thicker leaves were associated with higher nitrogen content (and presumably higher RuBisCO content) and higher TE as determined by CID.
6.4 Genes That Influence WUE
The interactive networks of traits and biochemical processes that influence or determine TE imply that TE is under the control of a whole suite of genes. The findings of Masle et al. (2005) suggested that TE may be under the control of a single gene. During the last decade a number of other individual genes have also been identified as having dominant roles in influencing WUE or TE (Table 6.1). These genes are predominantly involved in signaling, transcription, or water transport (Table 6.1). A summary of the genes that have been shown to influence WUE or TE follows in the next section.
6.4.1 Signaling Factors
One of the most promising discoveries for manipulation of TE arose in the last decade when a leucine-rich repeat receptor-like kinase (LRR-RLK) called ERECTA was identified as being a major contributor to the genetic locus for CID (Masle et al. 2005). Transmembrane receptor kinases, such as ERECTA, sense external signals via extracellular ligands, which activate the molecule’s intracellular kinase domain resulting in downstream signal transduction and gene regulation (Torii 2004). ERECTA, known for its involvement in regulating organ shape (Torii et al. 1996), was found to regulate the coordination between photosynthesis and transpiration in Arabidopsis thaliana (Masle et al. 2005). Complementation of erecta mutants with the wild-type allele restored TE to the same levels as wild-type plants under both well-watered and water-deficient conditions. ERECTA coordinates the early phase of stomatal development that determines stomatal density (see for review Bergmann and Sack 2007), which subsequently influences stomatal conductance and transpiration. More densely packed spongy mesophyll cells, greater electron transport capacity, and higher RuBisCO carboxylation rates in complemented ERECTA lines contributed to their greater CO2 assimilation rates (Masle et al. 2005). Overexpression of ERECTA from Populus nigra in Arabidopsis produced similar results and improved dry biomass (Xing et al. 2011). These results indicate that manipulation of ERECTA may be a way of potentially improving crop performance (Masle et al. 2005).
Despite the significance of the ERECTA discovery in regulating TE, it has not yet directly translated into crop varieties with greater yield under water-limited conditions due to improved TE, although the gene has presumably been selected in some crop species where breeding programs for drought tolerance used genotypic screening of CID to select parental material (Condon et al. 2002). Wheat genotypes with improved TE have already been produced from these programs (Condon et al. 2004). Homologs of the A. thaliana ERECTA locus have been identified as well as ERECTA genes isolated from the crop species rice, sorghum, maize, and wheat, which will aid marker-assisted selection and breeding programs (Masle et al. 2004). More recently, transgenic maize plants overexpressing ERECTA (ZmERECTA A) showed more vigorous growth in the field compared with nontransformed controls, but yield data was not provided (Guo et al. 2011). Interestingly, leaves on transgenic maize had not only decreased stomatal density but also increased pubescence (Guo et al. 2011).
ERECTA, along with the other members of its family (ERECTA-LIKE1 and ERECTA-LIKE2), and the leucine-rich repeat receptor-like protein (LRR-RLP), TOO MANY MOUTHS, are all needed to coordinate stomatal production and spacing, but the interacting relationships of these genes and their involvement in signal transduction are not yet fully understood (Bergmann and Sack 2007; Rowe and Bergmann 2010). ERECTA-family members have subtly different functions, and TOO MANY MOUTHS modulates ERECTA signaling in a tissue-specific manner (Rowe and Bergmann 2010), possibly through formation of heterodimers (Torii 2004). Rapid progress is being made in identifying the many other genes and signaling peptides that act upstream and downstream of the ERECTA-family and TOO MANY MOUTHS (Table 6.2) in the stomatal development pathway. While the specific details of stomatal development are not a primary focus of this chapter, the reader is referred to the excellent reviews of Bergmann and Sack (2007), Nadeau (2009), and Rowe and Bergmann (2010). Expression of any of these genes, in addition to ERECTA, could potentially influence TE by modulating stomatal conductance given their interdependent relationships and influence on stomatal density. Almost all of the stomatal development research has been undertaken on the model plant Arabidopsis, but to have greater relevance to agricultural crops, more research needs to be undertaken on other species to determine whether the mechanisms identified in Arabidopsis are conserved across species. B. distachyon would be an ideal model for cereals because of the close synteny between the genomes (Huo et al. 2009) and similar root and shoot development (Watt et al. 2009). Greater understanding of how the environment interacts with stomatal development is also needed as plasticity in TE may be a useful trait in different crops and different growing regions.
6.4.2 Transcription Factors Influencing WUE
Transcription factors control the expression of other genes, and therefore, those implicated in influencing stomatal density and aperture (ABA signaling) and root or shoot growth are likely to play a significant role in TE. Several transcription factors that affect TE have been identified (Table 6.1), but not all of the downstream events that they regulate. GTL1 appears to be involved in the development of stomata as it interacts with the promoter of STOMATAL DENSITY AND DISTRIBUTION1 (Yoo et al. 2010) (Table 6.2), a subtilizing protease, which is believed to be one of the ligands that interacts with TOO MANY MOUTHS (Bergmann and Sack 2007). Lower transpiration of gtl1-mutant Arabidopsis plants was associated with upregulated STOMATAL DENSITY AND DISTRIBUTION1 and fewer stomates on the abaxial leaf surface. GTL1 is one of the first genes to be shown to be involved in stomatal development in response to environmental conditions as it was shown to be downregulated with water stress (Yoo et al. 2010).
Several other transcription factors influencing TE independently from the genes involved in the stomata developmental pathway described in the section above have been identified (Table 6.1). HARDY, an AP2-ERF-like transcription factor, was first identified in phenotypic screening of Arabidopsis mutants (Karaba et al. 2007). Overexpression of HARDY in Arabidopsis induced thicker leaves due to extra palisade and mesophyll cells, and the root system was denser. Transformed plants survived longer periods when water was withheld and were more tolerant to salinity than wild-type plants. To test whether HARDY could contribute to improved TE in crop plants, Karaba et al. (2007) expressed the Arabidopsis HARDY gene in the rice cultivar Nipponbare. HARDY rice lines had greater TE, growth, and consequently biomass, but yield data was not reported. The root system contributed most to the greater biomass. In comparison with nontransformed plants, HARDY rice lines also had higher TE and instantaneous TE under well-watered and water-deficient conditions. Greater TE was facilitated by lower stomatal conductance and greater assimilation rates; however, the mechanism responsible for this is not known (Karaba et al. 2007). HRD overexpression induced clusters of genes that are normally expressed under drought stress, which probably contributed to the greater drought tolerance of the transgenic rice.
Overexpression of the Arabidopsis HARDY gene in Trifolium alexandrinum was consistent with the results in rice (Abogadallah et al. 2011). HARDY-Trifolium alexandrinum lines had greater instantaneous TE due to lower transpiration rates under water-deficient conditions. In the field, transgenic T. alexandrinum plants were more drought tolerant because of slower soil water depletion and had improved biomass in comparison with wild-type plants.
Other transcription factors that have been associated with TE include DREB1 and ABP9 (Table 6.1). Both of these genes are known for their association with drought tolerance (Lata and Prasad 2011). The Arabidopsis genes of DREB1 were expressed in groundnut driven by a stress-responsive promoter (Bhatnagar-Mathur et al. 2007; Devi et al. 2011). Five transgenic lines were compared against wild-type parental material under well-watered and water-deficient conditions in the field. TE varied across the lines and was correlated with specific leaf area and leaf chlorophyll content, but only under water deficit and was not correlated with CID under either water treatment. High TE lines maintained gas exchange longer, until the soil water content was lower than low TE genotypes (Devi et al. 2011). DREB1’s involvement in increased drought tolerance is associated with increased antioxidant activities (Li et al. 2011). DREB1 is also reported to stimulate root growth (Vadez et al. 2007). DREB homologous genes have been isolated in a variety of plant species, and some are also responsive to other abiotic stresses such as heat and cold, but overexpression does not always result in phenotypic changes possibly because constitutive promoters are not always functional in other species (Nakashima et al. 2009).
ABP9 is involved in the ABA-dependent signaling pathway. Transgenic Arabidopsis plants overexpressing ABP9 were more tolerant to drought and high temperature compared with wild-type plants and maintained higher rates of A N and g s (Zhang et al. 2008). Increased ABA content, photosynthetic pigments, and stress-responsive genes were some of the positive changes associated with increased instantaneous TE of transgenic plants.
6.4.3 Aquaporins Influencing TE
Aquaporins are membrane intrinsic proteins that predominantly form water-conducting pores, although some isoforms appear to conduct a wide range of other neutral molecules (see for review Hachez and Chaumont 2010). Although aquaporins are found in every organism, their abundance and diversity is greatest in plants (Tyerman et al. 2002). More than 30 aquaporin genes have been identified in Arabidopsis, maize, rice, and tomato (Chaumont et al. 2001; Johanson et al. 2001; Sakurai et al. 2005; Sade et al. 2009); at least 23 aquaporins have been detected in grapevine (Shelden et al. 2009) and 55 in poplar (Populus trichocarpa) (Gupta and Sankararamakrishnan 2009). Wheat apparently has an even greater number of aquaporin genes, as 24 PIPs (plasma membrane intrinsic proteins) and 11 TIPs (tonoplast intrinsic proteins) have already been identified (Forrest and Bhave 2008) and more may be discovered because allohexaploid wheat has three diploid genomes.
Aquaporin gene expression varies in different parts the plant, but they tend to be concentrated in tissue and cells needing greatest osmotic regulation and regulation of water flows (Maurel 1997), with the majority of aquaporin isoforms being expressed in roots (Bramley et al. 2007). Aquaporins that are located in plasma membranes (PIPs) may be primarily responsible for controlling transcellular water flow (Javot and Maurel 2002). TIPs and NIPs (nodulin-like intrinsic proteins) may also be involved in osmoregulation and transport of some osmolytes (Maurel 1997; Maurel et al. 2009), processes that are also important in drought resistance. Aquaporins that belong to the other subgroups, SIPs (small basic intrinsic proteins), and XIPs (X-intrinsic proteins) are rarely expressed (Chaumont et al. 2001; Danielson and Johanson 2008). In Arabidopsis, SIPs have been observed in endoplasmic reticulum membranes of specific cell types (Maeshima and Ishikawa 2008), and XIPs of the Solanaceae family transport uncharged molecules, but not water (Bienert et al. 2011).
The hydraulic conductance of membranes and tissue, if water takes the transcellular pathway, can be controlled by the abundance and gating (opening/closing) of aquaporins (Tyerman et al. 1999). The high diversity and complex expression patterns of aquaporins is one of the reasons why there is limited understanding of the role of individual isoforms. Nevertheless, pharmacological agents, that block aquaporin “pores” or change protein conformation, and reverse genetics have demonstrated the influence of aquaporins on controlling water flow across membranes and through roots or leaves of a number of species (Cochard et al. 2007; Bramley et al. 2009; Ehlert et al. 2009; Vandeleur et al. 2009; Sutka et al. 2010). Aquaporin-mediated regulation of hydraulic conductance endows the capability for rapid and reversible regulation of water flow through the plant to match transpiration and in response to the environment, unlike changes in morphology and anatomy, which are slow and growth dependent, i.e., efficiency in water transport can match the needs of the plant and environmental conditions (Tyerman et al. 1999). However, the majority of aquaporin studies have focused on details at the molecular level; a greater understanding of their physiology is needed.
A detailed discussion about the mechanisms controlling aquaporin activity is beyond the scope of this chapter, but it should be pointed out that aquaporin gating is directly or indirectly regulated by apoplastic water potential, free Ca2+ concentration, cytosolic pH, H2O2, and phosphorylation (see for review Bramley and Tyerman 2010), all of which are pertinent to cellular responses to water stress. The expression of various aquaporin isoforms has also been shown to change in response to drought and osmotic stress (Jang et al. 2004; Lian et al. 2004; Parent et al. 2009; Vandeleur et al. 2009), and ABA has been implicated in either stimulating gating or changes in aquaporin expression (Wan et al. 2004; Aroca et al. 2006; Parent et al. 2009). There have been few studies demonstrating the role of aquaporins in maintaining leaf water status. Moreover, these proteins are prime candidates for modulating TE, particularly if plant hydraulic conductance is tightly coupled with g s. Those aquaporins shown to be involved in TE (Table 6.1) or productivity will be discussed in the remainder of this chapter.
In contrast with the signaling and transcription factors described above, instantaneous TE of tomato plants was increased through overexpression of the tobacco aquaporin NtAQP1 (Sade et al. 2010). This aquaporin is one of the most intensely studied isoforms, and while highly abundant in roots (see references in Bramley et al. 2007), expression has been detected in all plant organs, including leaves where it is concentrated in the spongy mesophyll (Biela et al. 1999; Siefritz et al. 2001). Antisense tobacco plants with inhibited NtAQP1 expression demonstrated the importance of this aquaporin in maintaining plant water status, especially under water stress (Siefritz et al. 2002). Moreover, NtAQP1 is induced by stress, possibly through the production of the phytohormones ABA or gibberellic acid (Siefritz et al. 2001). In addition to transporting water (Siefritz et al. 2002; Sade et al. 2010), the NtAQP1 pore is also permeable to CO2 and influences A N through moderation of a leaf’s internal conductance to CO2 (or mesophyll conductance) (Uehlein et al. 2003, 2008). Interestingly, NtAQP1-transformed tomato plants had not only higher A N but also higher g s that was facilitated by larger stomatal apertures and not through increased stomatal density (Sade et al. 2010). The difference in instantaneous TE between transformed and wild-type tomato plants was even greater under salt stress.
Manipulation of aquaporin expression can cause changes in plant growth or hydraulic conductance (Martre et al. 2002; Siefritz et al. 2002), but overexpression of NtAQP1 in tomato did not affect root hydraulic conductivity (normalized to xylem cross-sectional area). Indeed, overexpression of this aquaporin in tobacco increased leaf growth, but did not affect root mass or plant height (Uehlein et al. 2003), whereas plant size and root mass were affected by inhibited expression (Siefritz et al. 2002). Although NtAQP1-transformed and wild-type tomato plants had similar root hydraulic conductivities under control conditions, salinity reduced root hydraulic conductivity of NtAQP1-transformed plants considerably less than wild-type plants. The maintenance of root hydraulic conductivity in NtAQP1-transformed plants likely supported their higher daily rates of transpiration under salinity (Sade et al. 2010).
Rice has several aquaporin genes that respond to abiotic stresses. A plasma membrane aquaporin, OsPIP1;3, was upregulated in an upland subspecies that is more tolerant of drought than a lowland subspecies (Lian et al. 2004). When OsPIP1;3 was overexpressed in transgenic lowland plants and exposed to osmotic stress (decreased water potential of the root solution culture), transgenic plants had higher root hydraulic conductivity, leaf water potential, and transpiration (Lian et al. 2004). In contrast, overexpression of PIP1b in Arabidopsis had an adverse effect on drought resistance, which caused premature wilting (Aharon et al. 2003). The productivity of transgenic plants was improved under well-watered conditions due to greater transpiration rates and photosynthetic efficiency, which was facilitated by increased stomatal density. These results indicate that selection of the relevant isoforms should be conducted under water-deficient conditions if the aim is to improve TE under drought.
Modulation of aquaporin activity to improve TE would need to lead to improved yields before this trait would be considered for selection in breeding programs. NtAQP1 is a promising candidate because there was no yield penalty associated with overexpression of this isoform in tomato (Sade et al. 2010). Furthermore, individual fruit of NtAQP1-transformed tomato plants weighed more than wild-type plants under salt treatment (salt did not reduce individual fruit weight of transformed plants). Under control conditions the number of fruit per plant was higher on transformed plants and overall total fruit fresh weight was slightly higher in transformed plants under salinity compared with wild type (Sade et al. 2010), but whether this was due to higher water content or greater dry weight is not clear. Sade et al. (2010) also demonstrated that the beneficial effect of NtAQP1 overexpression on plant production is not isolated to tomato because transformed NtAQP1-Arabidopsis plants produced more biomass under control and saline conditions compared with wild-type plants.
Sade et al. (2009) tested overexpression of a TIP in tomato, SlTIP2;2, on yield performance under salt and drought stress in the field. TOM-SlTIP2;2 plants had more biomass and fruit yields under drought stress compared with nontransgenic plants. Transgenic TOM-SlTIP2;2 plants had higher transpiration rates than controls when grown in pots and irrigated with saline or nonsaline solution. Higher and sustained rates of transpiration under drought led to lower relative water contents in leaves of transgenic plants. TIP aquaporins are expressed in the tonoplast, and it has been speculated that their function may be buffering of the cytoplasm against deleterious changes in volume (Maurel 1997; Tyerman et al. 1999) or they may function as osmotic and turgor pressure sensors (Hill et al. 2004). The novel results of Sade et al. (2009) demonstrate that SlTIP2;2 is involved in leaf water status, but rather than maintaining leaf water status, the leaves dehydrated to lower relative water contents suggesting that overexpression of this TIP served a protective role.
6.4.4 Other Regulatory Genes of TE
Several other direct or indirect regulators of TE have been identified (Table 6.1), the most studied of which is ESKIMO1, which encodes a protein with unknown function (Xin et al. 2007). ESKIMO1 was first identified for its negative effect on freezing tolerance, which was associated with changes in expression of transcription factors, signaling components, and stress-responsive genes (Xin et al. 2007). The overlap between the genes regulated by ESKIMO1 with those that respond to osmotic, salt, and ABA treatments suggested that ESKIMO1 may be involved in drought tolerance, but esk1-mutants wilted earlier than wild-type plants (Xin et al. 2007). In contrast, Bouchabke-Coussa et al. (2008) observed improved drought tolerance in esk1-mutants through a lower transpiration rate and improved TE. Esk1 mutants were smaller than wild-type plants and produced fewer seeds under well-watered conditions, but esk1-mutants stayed green longer and wilted later than wild-type plants when water was withheld. Interestingly, of the 135 genes that were differentially expressed between wild-type and esk1-mutants under water deficit, a TIP and a PIP aquaporin were downregulated in the mutant plants. The PIP aquaporin (PIP1;5) is predominantly expressed in roots and has consistently been shown to be downregulated in response to abiotic stress (Jang et al. 2004; Alexandersson et al. 2005). Downregulation of PIP1;5 could have reduced the water transport capability of esk1-mutant roots and may explain why rosettes of esk1-mutants had lower relative water contents (Bouchabke-Coussa et al. 2008; Lugan et al. 2009). Lower root hydraulic conductance in esk1-mutants compared with wild-type plants was confirmed in a subsequent study, but this was correlated with xylem deformation (Lefebvre et al. 2011). Genes involved in cell wall synthesis (CesA7) have also been found to influence TE through xylem deformation (Liang et al. 2010). The rosettes of Esk1-mutants also had higher ABA contents, which may be a water-conserving mechanism inducing stomatal closure because of the decreased water uptake capability (Lefebvre et al. 2011). Changes in metabolite content and osmotic potential further indicate that ESKIMO1 is involved in maintaining water balance (Lugan et al. 2009).
The relationship between ABA synthesis and g s has long been known, but recent discoveries about the genetic control of ABA biosynthesis and its role in TE provide new avenues for genetic manipulation of crop plants. Overexpression of NCED1, an enzyme (Table 6.1) that catalyzes a rate-limiting step in ABA biosynthesis, increased ABA content and reduced g s in transgenic tomato in comparison with wild-type plants (Thompson et al. 2007). Biomass of transgenic and wild-type plants was not significantly different, but one of the transgenic lines had the lowest total water use and highest TE. Drought reduced the g s of wild-type plants but not transgenic plants. The reduced g s were associated with greater ABA production in wild-type plants and not increased sensitivity to ABA. Because of the reduced rate of water loss, the decrease in leaf water potential of transgenic plants was slower than wild-type plants, and the transgenic plants retained their leaves longer. In a separate experiment, hydroponically grown transgenic plants tended to have greater root hydraulic conductivities, which were calculated from the exudation rate of detopped roots and the difference in osmotic pressure between the root exudates and the solution bathing the roots (Thompson et al. 2007). The authors speculated that ABA stimulated an increased solute flux into the xylem stream to drive the greater exudation rates, but the osmolarity of the exudates was lower than wild-type plants (Thompson et al. 2007). However, a decrease in the force driving water efflux is compensated by the increased hydraulic conductance. Exogenous application of ABA to root systems has been shown previously to stimulate increased root hydraulic conductance, in some species, probably via an aquaporin-mediated mechanism (Quintero et al. 1999; Aroca et al. 2006), and therefore, the overproduction of ABA in transgenic plants may have stimulated increased aquaporin activity. It should be noted that interpretations based on root pressure-driven exudation rates from detopped plants should be made with caution as the water flow rates and driving forces are not representative of transpiring plants.
Another gene associated with ABA signaling and shown to improve TE is the ABA-inducible HVA1 gene (Sivamani et al. 2000). Expression of the barley HVA1 in transgenic wheat improved TE in all transgenic lines except one. Transgenic plants had larger root systems under moderate water deficit compared with controls. Shoot dry weight was also greater, but seed weight was only greater in one homozygous transgenic line. In the field, some of the HVA1-transgenic wheat lines outyielded the wild-type plants but only in some seasons (Bahieldin et al. 2005).
Manipulation of stomatal aperture by perturbation of the enzyme that converts malic acid to pyruvate in guard cells can also influence TE (Laporte et al. 2002). Malate is one of the anions that balance the positive charge generated by the influx of potassium ions during turgor-driven stomatal opening. Expressing the maize NADP-malic enzyme (ME) in transgenic tobacco increased the malate content of leaves, decreased g s, and improved TE compared with wild-type plants. Under drought conditions, the lower g s of ME-transformed plants drew soil water content down more slowly so that the wild type wilted earlier than transformed plants.
6.5 Conclusion and Future Directions
This chapter has demonstrated that there are many hydraulic and biochemical processes involved in controlling water flow through the plant and in controlling photosynthesis. As a consequence, there are potentially many genes and their interaction involved in determining the TE of a plant during development and under water-limited conditions. Nevertheless, TE is predominantly driven by hydraulic properties, such as stomatal conductance and anatomy of the water transport pathway, or factors that regulate those hydraulic properties, such as stomatal density (Fig. 6.2). Thus, most of the genes associated with modulation of TE are involved in regulating gas exchange, and although g s will also affect A N, A N is further limited by the efficiency of the photosynthetic apparatus. In C3 plants, the greatest limitation is RuBisCO, which provides few avenues for improvement. These results indicate that the main way of improving TE of agricultural crops will be through a reduction in water use rather than increases in photosynthetic efficiency. Many studies have observed that higher genotypic TE is generally due to lower transpiration rates (see reviews by Blum 2005, 2009).
Moreover, despite the extensive research on TE and aspirations to improve TE of crops, evidence demonstrating the benefits of improved TE in the field and for increasing yield is substantially deficient. Most of the studies discussed in this chapter indicate that higher TE is predominantly associated with slower growth. Most of the transgenic plants with improved TE were smaller than their related parental wild types. Higher TE was also related to more gradual depletion of the soil moisture, which suggests that high TE could be a suitable trait for improving drought tolerance under water-limited conditions, but when the water supply is not limiting, it may lead to yield penalties. In addition, slower growth is not a suitable trait in environments that experience terminal drought, but a more conservative water use would be beneficial in environments where crops are grown predominantly on stored soil moisture. Future research should, therefore, be directed towards examining the influence of high TE on yield production in targeted environments, as the research to date indicates that it is not a trait that would be beneficial in all growing regions.
References
Abogadallah GM, Nada RM, Malinowski R, Quick P (2011) Overexpression of HARDY, an AP2/ERF gene from Arabidopsis, improves drought and salt tolerance by reducing transpiration and sodium uptake in transgenic Trifolium alexandrinum L. Planta 233:1265–1276
Abrams MD, Kubiske ME, Steiner KC (1990) Drought adaptations and responses in five genotypes of Fraxinus pennsylvanica Marsh: photosynthesis, water relations and leaf morphology. Tree Physiol 6:305–315
Abrams MD, Kubiske ME, Mostoller SA (1994) Relating wet and dry year ecophysiology to leaf structure in contrasting temperate tree species. Ecology 75:123–133
Ackerson RC, Krieg DR, Sung FJM (1980) Leaf conductance and osmoregulation of field-grown Sorghum genotypes. Crop Sci 20:10–14
Aharon R, Shahak Y, Winiger S, Bendov R, Kapulnik Y, Galili G (2003) Overexpression of a plasma membrane aquaporin in transgenic tobacco improves plant vigor under favorable growth conditions but not under drought or salt stress. Plant Cell 15:439–447
Alexandersson E, Fraysse L, Sjövall-Larsen S, Gustavsson S, Fellert M, Karlsson M, Johanson U, Kjellbom P (2005) Whole gene family expression and drought stress regulation of aquaporins. Plant Mol Biol 59:469–484
Ali M, Jensen CR, Mogensen VO, Bahrun A (1999) Drought adaptation of field grown wheat in relation to soil physical conditions. Plant Soil 208:149–159
Anderson WK, Hamza MA, Sharma DL, D’Antuono MF, Hoyle FC, Hill N, Shackley BJ, Amjad M, Zaicou-Kunesch C (2005) The role of management in yield improvement of the wheat crop-a review with special emphasis on Western Australia. Aust J Agric Res 56:1137–1149
Araus JL, Slafer GA, Royo C, Serret MD (2008) Breeding for yield potential and stress adaptation in cereals. Crit Rev Plant Sci 27:377–412
Aroca R, Ferrante A, Vernieri P, Chrispeels MJ (2006) Drought, abscisic acid and transpiration rate effects on the regulation of PIP aquaporin gene expression and abundance in Phaseolus vulgaris plants. Ann Bot 98:1301–1310
Austin RB, Morgan CL, Ford MA (1982) Flag leaf photosynthesis of Triticum aestivum and related diploid and tetraploid species. Ann Bot 49:177–189
Baaziz KB, Lopez D, Rabot A, Combes D, Gousset A, Bouzid S, Cochard H, Sakr S, Venisse J-S (2012) Light-mediated Kleaf induction and contribution of both the PIP1s and PIP2s aquaporins in five tree species: walnut (Juglans regia) case study. Tree Physiol 32:423–434
Bahieldin A, Hesham HT, Eissa HF, Saleh OM, Ramadan AM, Ahmed IA, Dyer WE, Madkour MA (2005) Field evaluation of transgenic wheat plants stably expressing the HVA1 gene for drought tolerance. Physiol Plant 123:421–427
Barbour M, Warren C, Farquhar G, Forrester G, Brown H (2010) Variability in mesophyll conductance between barley genotypes, and effects on transpiration efficiency and carbon isotope discrimination. Plant Cell Environ 33:1176–1185
Barrowclough DE, Peterson CA, Steudle E (2000) Radial hydraulic conductivity along developing onion roots. J Exp Bot 51:547–557
Bennett D, Izanloo A, Reynolds MP, Kuchel H, Langridge P, Schnurbusch T (2012) Genetic dissection of grain yield and physical grain quality in bread wheat (Triticum aestivum) under water-limited environments. Theor Appl Genet 125:255–271
Bergmann DC, Sack FD (2007) Stomatal development. Annu Rev Plant Biol 58:163–181
Bharathan G, Goliber TE, Moore C, Kessler S, Pham T, Sinha NR (2002) Homologies in leaf form inferred from KNOX1 gene expression during development. Science 296:1858–1860
Bhatnagar-Mathur P, Devi MH, Reddy DS, Lavanya M, Vadez V, Serraj R, Yamaguchi-Shinozaki K, Sharma KK (2007) Stress-inducible expression of AtDREB1A in transgenic peanut (Arachis hypogaea L.) increases transpiration efficiency under water-limiting conditions. Plant Cell Rep 26:2071–2082
Biela A, Grote K, Otto B, Hoth S, Hedrich R, Kaldenhoff R (1999) The Nicotiana tabacum plasma membrane aquaporin NtAQP1 is mercury-insensitive and permeable for glycerol. Plant J 18:565–570
Bienert GP, Bienert MD, Jahn TP, Boutry M, Chaumont F (2011) Solanaceae XIPs are plasma membrane aquaporins that facilitate the transport of many uncharged substrates. Plant J 66:306–317
Blum A (2005) Drought resistance, water-use efficiency, and yield potential – are they compatible, dissonant, or mutually exclusive? Aust J Agric Res 56:1159–1168
Blum A (2009) Effective use of water (EUW) and not water-use efficiency (WUE) is the target of crop yield improvement under drought stress. Field Crops Res 112:119–123
Bonhomme L, Monclus R, Vincent D, Carpin S, Lomenech AM, Plomion C, Brignolas F, Morabito D (2009) Leaf proteome analysis of eight Populus x euramericana genotypes: genetic variation in drought response and in water-use efficiency involves photosynthesis-related proteins. Proteomics 9:4121–4142
Botwright TL, Condon AG, Rebetzke GJ, Richards RA (2002) Field evaluation of early vigour for genetic improvement of grain yield in wheat. Aust J Agric Res 53:1137–1145
Bouchabke-Coussa O, Quashie ML, Seoane-Redondo J, Fortabat MN, Gery C, Yu A, Linderme D, Trouverie J, Granier F, Téoulé E, Durand-Tardif M (2008) ESKIMO1 is a key gene involved in water economy as well as cold acclimation and salt tolerance. BMC Plant Biol 8:125
Bowman WD, Roberts SW (1985) Seasonal changes in tissue elasticity in chaparral shrubs. Physiol Plant 65:233–236
Boyer JS (1970) Leaf enlargement and metabolic rates in corn, soybean and sunflower at various leaf water potentials. Plant Physiol 43:1056–1062
Boyer JS (1982) Plant productivity and environment. Science 218:543–548
Bramley H, Tyerman S (2010) Root water transport under waterlogged conditions and the roles of aquaporins. In: Mancuso S, Shabala S (eds) Waterlogging signalling and tolerance in plants. Springer, Berlin, pp 151–180
Bramley H, Turner DW, Tyerman SD, Turner NC (2007) Water flow in the roots of crop species: the influence of root structure, aquaporin activity, and waterlogging. Adv Agron 96:133–196
Bramley H, Turner NC, Turner DW, Tyerman SD (2009) Roles of morphology, anatomy, and aquaporins in determining contrasting hydraulic behavior of roots. Plant Physiol 150:348–364
Bramley H, Turner NC, Turner DW, Tyerman SD (2010) The contrasting influence of short-term hypoxia on the hydraulic properties of cells and roots of wheat and lupin. Funct Plant Biol 37:183–193
Bramley H, Tyerman SD, Turner DW, Turner NC (2011) Root growth of lupins is more sensitive to waterlogging than wheat. Funct Plant Biol 38:910–918
Briggs LJ, Shantz HL (1913) The water requirement of plants. Bureau of Plant Industry Bulletin. US Department of Agriculture, Washington, DC, pp 284–285
Brodribb T, Holbrook N (2004) Diurnal depression of leaf hydraulic conductance in a tropical tree species. Plant Cell Environ 27:820–827
Brodribb TJ, Feild TS, Jordan GJ (2007) Leaf maximum photosynthetic rate and venation are linked by hydraulics. Plant Physiol 144:1890–1898
Buckley TN (2005) The control of stomata by water balance. New Phytol 168:275–292
Casson S, Gray JE (2008) Influence of environmental factors on stomatal development. New Phytol 178:9–23
Chaumont F, Barrieu F, Wojcik E, Chrispeels MJ, Jung R (2001) Aquaporins constitute a large and highly divergent protein family in maize. Plant Physiol 125:1206–1215
Chen YL, Dunbabin VM, Postma J, Diggle AJ, Palta JA, Lynch JP, Siddique KHM, Rengel Z (2011) Phenotypic variability and modelling of root structure of wild Lupinus angustifolius genotypes. Plant Soil 348:345–364
Chen YL, Dunbabin VM, Diggle AJ, Siddique KHM, Rengel Z (2012) Assessing variability in root traits of wild Lupinus angustifolius germplasm: basis for modelling root system structure. Plant Soil 354:141–155
Clark RT, MacCurdy RB, Jung JK, Shaff JE, McCouch SR, Aneshansley DJ, Kochian LV (2011) Three-dimensional root phenotyping with a novel imaging and software platform. Plant Physiol 156:455–465
Clarke JM, Richards RA (1988) The effects of glaucousness, epicuticular wax, leaf age, plant height, and growth environment on water loss rates of excised wheat leaves. Can J Plant Sci 68:975–982
Clarke JM, McCaig TN, Depauw RM (1993) Relationship of glaucousness and epicuticular wax quantity of wheat. Can J Plant Sci 73:961–967
Clifford SC, Black CR, Roberts JA, Stronach IM, Singleton-Jones PR, Mohamed AD, Azam-Ali SN (1995) The effect of elevated atmospheric CO2 and drought on stomatal frequency in groundnut (Arachis hypogaea (L.)). J Exp Bot 46:847–852
Cochard H, Nardini A, Coll L (2004) Hydraulic architecture of leaf blades: where is the main resistance? Plant Cell Environ 27:1257–1267
Cochard H, Venisse J-S, Barigah TS, Brunel N, Herbette S, Guilliot A, Tyree MT, Sakr S (2007) Putative role of aquaporins in variable hydraulic conductance of leaves in response to light. Plant Physiol 143:122–133
Colmer TD (2003) Long-distance transport of gases in plants: a perspective on internal aeration and radial oxygen loss from roots. Plant Cell Environ 26:17–36
Condon AG, Farquhar GD, Richards RA (1990) Genotypic variation in carbon isotope discrimination and transpiration efficiency in wheat. Leaf gas exchange and whole plant studies. Funct Plant Biol 17:9–22
Condon AG, Richards RA, Rebetzke GJ, Farquhar GD (2002) Improving intrinsic water-use efficiency and crop yield. Crop Sci 42:122–131
Condon AG, Richards RA, Rebetzke GJ, Farquhar GD (2004) Breeding for high water-use efficiency. J Exp Bot 55:2447–2460
Danielson J, Johanson U (2008) Unexpected complexity of the Aquaporin gene family in the moss Physcomitrella patens. BMC Plant Biol 8:45
Davies WJ, Zhang J (1991) Root signals and the regulation of growth and development of plants in drying soil. Annu Rev Plant Biol 42:55–76
Davies WJ, Kudoyarova G, Hartung W (2005) Long-distance ABA signalling and its relation to other signalling pathways in the detection of soil drying and the remediation of the plant’s response to drought. J Plant Growth Regul 24:285–295
Devi MJ, Bhatnagar-Mathur P, Sharma KK, Serraj R, Anwar SY, Vadez V (2011) Relationships between transpiration efficiency and its surrogate traits in the rd29A:DREB1A transgenic lines of groundnut. J Agron Crop Sci 197:272–283
Dias de Oliveira E, Bramley H, Siddique KHM, Henty S, Berger J, Palta JA (2013) Can elevated CO2 combined with high temperature ameliorate the effect of terminal drought in wheat? Funct Plant Biol 40:160–171
Dodd AN, Borland AM, Haslam RP, Griffiths H, Maxwell K (2002) Crassulacean acid metabolism: plastic, fantastic. J Exp Bot 53:569–580
Dornhoff GM, Shibles R (1976) Leaf morphology and anatomy in relation to CO2-exchange rate of soybean leaves. Crop Sci 16:377–381
Dracup M, Reader M, Palta JA (1998) Timing of terminal drought is an important cause of yield variability in lupin grown on duplex soils in souther Australia. Aust J Agric Res 49:799–810
Drew MC, He C-J, Morgan PW (1989) Decreased ethylene biosynthesis, and induction of aerenchyma, by nitrogen- or phosphate-starvation in adventitious roots of Zea mays L. Plant Physiol 91:266–271
Duan B, Yang Y, Lu Y, Korpelainen H, Berninger F, Li C (2007) Interactions between water deficit, ABA, and provenances in Picea asperata. J Exp Bot 58:3025–3036
Dunn GJ, Briggs KG (1989) Variation in culm anatomy among barley cultivars differing in lodging resistance. Can J Bot 67:1838–1843
Ehleringer J (1980) Leaf morphology and reflectance in relation to water and temperature stress. In: Turner NC, Kramer PJ (eds) Adaptation of plants to water and high temperature stress. Wiley Interscience, New York, pp 295–308
Ehleringer J (1982) The influence of water stress and temperature on leaf pubescence development in Encelia farinosa. Am J Bot 69:670–675
Ehleringer J (1984) Ecology and ecophysiology of leaf pubescence in North American desert plants. In: Rodrigues E, Healy PL, Mehta I (eds) Biology and chemistry of plant trichomes. Plenum, New York, pp 113–132
Ehleringer JR, Clark C (1987) Evolution and adaptation in Encelia (Asteraceae). In: Gottlieb LD, Jain SK (eds) Plant evolutionary biology. Chapman and Hall, London, pp 221–248
Ehlert C, Maurel C, Tardieu F, Simonneau T (2009) Aquaporin-mediated reduction in maize root hydraulic conductivity impacts cell turgor and leaf elongation even without changing transpiration. Plant Physiol 150:1093–1104
Farquhar GD, Richards RA (1984) Isotopic composition of plant carbon correlates with water-use efficiency of wheat genotypes. Aust J Plant Physiol 11:539–552
Farquhar GD, Sharkey TD (1982) Stomatal conductance and photosynthesis. Annu Rev Plant Physiol 33:317–345
Farquhar GD, O’Leary MH, Berry JA (1982) On the relationship between carbon isotope discrimination and the intercellular carbon dioxide concentration in leaves. Aust J Plant Physiol 9:121–137
Fischer RA (1981) Optimizing the use of water and nitrogen through breeding of crops. Plant Soil 58:249–278
Fiscus EL (1981) Effects of abscisic acid on the hydraulic conductance and the total ion transport through Phaseolus root systems. Plant Physiol 68:169–174
Flexas J, Ribas-Carbo M, Diaz-Espejo A, Galmes J, Medrano H (2007) Mesophyll conductance to CO2: current knowledge and future prospects. Plant Cell Environ 31:602–621
Forde BG (2009) Is it good noise? The role of developmental instability in the shaping of a root system. J Exp Bot 60:3989–4002
Forrest K, Bhave M (2008) The PIP and TIP aquaporins in wheat form a large and diverse family with unique gene structures and functionally important features. Funct Integr Genomics 8:115–133
French RJ, Schultz JE (1984a) Water use efficiency of wheat in a Mediterranean-type environment. 1. The relation between yield, water use and climate. Aust J Agric Res 35:743–764
French RJ, Schultz JE (1984b) Water use efficiency of wheat in a Mediterranean-type environment. 2. Some limitations to efficiency. Aust J Agric Res 35:765–775
Galmés J, Conesa MA, Ochogavía JM, Perdomo JA, Francis DM, Ribas-Carbó M, Savé R, Flexas J, Medrano H, Cifre J (2010) Physiological and morphological adaptations in relation to water use efficiency in Mediterranean accessions of Solanum lycopersicum. Plant Cell Environ 34:245–260
Garrity DP, O’Toole JC (1995) Selection for reproductive stage drought avoidance in rice, using infrared thermometry. Agron J 87:773–779
Garvin DF, Gu YQ, Hasterok R, Hazen SP, Jenkins G, Mockler TC, Mur LAJ, Vogel JP (2008) Development of genetic and genomic research resources for Brachypodium distachyon, a new model system for grass crop research. Crop Sci 48:S69–S84
González A, Ayerbe L (2009) Effect of terminal water stress on leaf epicuticular wax load, residual transpiration and grain yield in barley. Euphytica 172:341–349
González A, Martín I, Ayerbe L (1999) Barley yield in water-stress conditions: the influence of precocity, osmotic adjustment and stomatal conductance. Field Crops Res 62:23–34
Gray JE (2007) Plant development: three steps for stomata. Curr Biol 17:R213–R215
Gregory P (2004) Agronomic approaches to increasing water use efficiency. In: Bacon M (ed) Water use efficiency in plant biology. Blackwell, Oxford, pp 142–170
Guo M, Rupe M, Simmons C, Sivasankar S (2011) The maize ERECTA genes for improving plant growth, transpiration efficiency and drought tolerance in crop plants. United States Patent Application, US 2011/0035844 A1
Gupta A, Sankararamakrishnan R (2009) Genome-wide analysis of major intrinsic proteins in the tree plant Populus trichocarpa: characterization of XIP subfamily of aquaporins from evolutionary perspective. BMC Plant Biol 9:134
Guyot G, Scoffoni C, Sack L (2012) Combine impacts of irradiance and dehydration on leaf hydraulic conductance: insights into vulnerability and stomatal control. Plant Cell Environ 35:857–871
Hachez C, Chaumont F (2010) Aquaporins: a family of highly regulated multifunctional channels. In: Jahn TP, Bienert GP (eds) MIPs and their role in the exchange of metalloids. Springer, New York, pp 1–17
Hamanishi ET, Thomas BR, Campbell MM (2012) Drought induces alterations in the stomatal development program in Populus. J Exp Bot 63:4959–4971
Hamblin A, Tennant D (1987) Root length density and water uptake in cereals and grain legumes: how well are they correlated? Aust J Agric Res 38:513–527
Harris DS, Schapaugh WT, Kanemasu ET (1984) Genetic diversity in soybeans for leaf canopy temperature and the association of leaf canopy temperature and yield. Crop Sci 24:839–842
Hartung W, Leport L, Ratcliffe RG, Sauter A, Duda R, Turner NC (2002) Abscisic acid concentration, root pH and anatomy do not explain growth differences of chickpea (Cicer arietinum L.) and lupin (Lupinus angustifolius L.) on acid and alkaline soils. Plant Soil 240:191–199
Heinen RB, Ye Q, Chaumont F (2009) Role of aquaporins in leaf physiology. J Exp Bot 60:2971–2985
Henson IE, Turner NC (1991) Stomatal responses to abscisic-acid in 3 lupin species. New Phytol 117:529–534
Henson IE, Jensen CR, Turner NC (1989a) Leaf gas exchange and water relations of lupins and wheat. I. Shoot responses to soil water deficits. Aust J Plant Physiol 16:401–413
Henson IE, Jensen CR, Turner NC (1989b) Leaf gas exchange and water relations of lupins and wheat. III. Abscisic acid and drought-induced stomatal closure. Aust J Plant Physiol 16:429–442
Henzler T, Waterhouse RN, Smyth AJ, Carvajal M, Cooke DT, Schäffner AR, Steudle E, Clarkson DT (1999) Diurnal variations in hydraulic conductivity and root pressure can be correlated with the expression of putative aquaporins in the roots of Lotus japonicus. Planta 210:50–60
Hetherington AM, Woodward FI (2003) The role of stomata in sensing and driving environmental change. Nature 424:901–908
Hill AE, Shachar-Hill B, Shachar-Hill Y (2004) What are aquaporins for? J Membr Biol 197:1–32
Hose E, Steudle E, Hartung W (2000) Abscisic acid and hydraulic conductivity of maize roots: a study using cell- and root-pressure probes. Planta 211:874–882
Hsiao TC, Silk WW, Jing J (1985) Leaf growth and water deficits: biophysical effects. In: Baker NR, Davies WJ, Ong CK (eds) Control of leaf growth. Cambridge University Press, Cambridge, pp 239–266
Hubick KT, Farquhar GD (1989) Genetic variation in carbon isotope discrimination and the ratio of carbon gained to water lost in barley. Plant Cell Environ 12:795–804
Huo N, Vogel JP, Lazo GR, You FM, Ma Y, McMahon S, Dvorak J, Anderson OD, Luo M-C, Gu YQ (2009) Structural characterization of Brachypodium genome and its syntenic relationship with rice and wheat. Plant Mol Biol 70:47–61
Izanloo A, Condon AG, Langridge P, Tester M, Schnurbusch T (2008) Different mechanisms of adaptation to cyclic water stress in two South Australian bread wheat cultivars. J Exp Bot 59:3327–3346
Jang JY, Kim DG, Kim YO, Kim JS, Kang H (2004) An expression analysis of a gene family encoding plasma membrane aquaporins in response to abiotic stresses in Arabidopsis thaliana. Plant Mol Biol 54:713–725
Javot H, Maurel C (2002) The role of aquaporins in root water uptake. Ann Bot 90:301–313
Jellum MD (1962) Relationships between lodging resistance and certain culm characters in oats. Crop Sci 2:263–267
Jensen CR, Henson IE (1990) Leaf water relations characteristics of Lupinus angustifolius and L. cosentini. Oecologia 82:114–121
Johanson U, Karlsson M, Johansson I, Gustavsson S, Sjovall S, Fraysse L, Weig AR, Kjellbom P (2001) The complete set of genes encoding major intrinsic proteins in Arabidopsis provides a framework for a new nomenclature for major intrinsic proteins in plants. Plant Physiol 126:1358–1369
Johnson RC, Nguyen HT, Croy LI (1984) Osmotic adjustment and solute accumulation in two wheat genotypes differing in drought resistance. Crop Sci 24:957–962
Jones MM, Osmond CB, Turner NC (1980) Accumulation of solutes in leaves of sorghum and sunflower in response to water deficits. Aust J Plant Physiol 7:193–205
Jones HG, Serraj R, Loveys BR, Xiong L, Wheaton A, Price AH (2009) Thermal infrared imaging of crop canopies for the remote diagnosis and quantification of plant responses to water stress in the field. Funct Plant Biol 36:978–989
Jordan WR, Shouse PJ, Blum A, Miller FR, Monk RL (1984) Environmental physiology of sorghum. II. Epicuticular wax load and cuticular transpiration. Crop Sci 24:1168–1173
Kaack K, Schwarz K-U, Brander PE (2003) Variation in morphology, anatomy and chemistry of stems of Miscanthus genotypes differing in mechanical properties. Ind Crops Prod 17:131–142
Kaminski A, Austin RB, Ford MA, Morgan CL (1990) Flag leaf anatomy of Triticum and Aegilops species in relation to photosynthetic rate. Ann Bot 66:359–365
Karaba A, Dixit S, Greco R, Aharoni A, Trijatmiko KR, Marsch-Martinez N, Krishnan A, Nataraja KN, Udayakumar M, Pereira A (2007) Improvement of water use efficiency in rice by expression of HARDY, an Arabidopsis drought and salt tolerance gene. Proc Natl Acad Sci USA 104:15270–15275
Kerstiens G (1997) In vivo manipulation of cuticular water permeance and its effect on stomatal response to air humidity. New Phytol 137:473–480
Kerstiens G (2006) Water transport in plant cuticles: an update. J Exp Bot 57:2493–2499
Khanna VK (1991) Relationship of lodging resistance yield to anatomical characters of stem in wheat [Triticum spp.], triticale and rye. Wheat Inform Serv 73:19–24
Kimura S, Koenig D, Kang J, Yoong FY, Sinha N (2008) Natural variation in leaf morphology results from mutation of a novel KNOX gene. Curr Biol 6:672–677
Klepper B (1983) Managing root systems for efficient water use: axial resistances to flow in root systems – anatomical considerations. In: Taylor HM, Jordan WR, Sinclair TR (eds) Limitations to efficient water use in crop production. American Society of Agronomy, Madison, WI, pp 115–125
Kulkarni M, Deshpande U (2006) Comparative studies in stem anatomy and morphology in relation to drought resistance in tomato (Lycopersicon esculentum). Am J Plant Pathol 1:82–88
Lambers H, Robinson SA, Ribos-Carbo M (2005) Regulation of respiration in vivo. In: Lambers H, Ribas-Carbo M (eds) Plant respiration. Springer, Dordhecht, pp 1–15
Laporte MM, Shen B, Tarczynski MC (2002) Engineering for drought avoidance: expression of maize NADP-malic enzyme in tobacco results in altered stomatal function. J Exp Bot 53:699–705
Larsson S, Svenningsson M (1986) Cuticular transpiration and epicuticular lipids of primary leaves of barley (Hordeum vulgare). Physiol Plant 68:13–19
Lata C, Prasad M (2011) Role of DREBs in regulation of abiotic stress responses in plants. J Exp Bot 62:4731–4748
Lefebvre V, Fortabat MN, Ducamp A, North HM, Maia-Grondard A, Trouverie J, Boursiac Y, Mouille G, Durand-Tardif M (2011) ESKIMO1 disruption in Arabidopsis alters vascular tissue and impairs water transport. PLoS One 6:e16645
Leport L, Turner NC, French RJ, Tennant D, Thomson BD, Siddique KHM (1998) Water relations, gas exchange and growth of cool-season grain legumes in a Mediterranean-type environment. Eur J Agron 9:295–303
Li X, Cheng X, Liu J, Zeng H, Han L, Tang W (2011) Heterologous expression of the Arabidopsis DREB1A/CBF3 gene enhances drought and freezing tolerance of transgenic Lolium perenne plants. Plant Biotechnol Rep 5:61–69
Lian H-L, Yu X, Ye Q, Ding X-S, Kitagawa Y, Kwak S-S, Su W-A, Tang Z-C (2004) The role of aquaporin RWC3 in drought avoidance in rice. Plant Cell Physiol 45:481–489
Liang YK, Xie X, Lindsay SE, Wang YB, Masle J, Williamson L, Leyser O, Hetherington AM (2010) Cell wall composition contributes to the control of transpiration efficiency in Arabidopsis thaliana. Plant J 64:679–686
Liao M, Palta JA, Fillery IRP (2006) Root characteristics of vigorous wheat improve early nitrogen uptake. Aust J Agric Res 57:1097–1107
Lo Gullo MA, Salleo S (1988) Different strategies of drought resistance in three Mediterranean sclerophyllous trees growing in the same environmental conditions. New Phytol 108:267–276
Lo Gullo MA, Nardini A, Salleo S, Tyree MT (1998) Changes in root hydraulic conductance (KR) of Olea oleaster seedlings following drought stress and irrigation. New Phytol 140:25–31
Loss SP, Siddique KHM (1994) Morphological and physiological traits associated with wheat yield increases in Mediterranean environments. Adv Agron 52:229–276
Lugan R, Niogret M-F, Kervazo L, Larher FR, Kopka J, Bouchereau A (2009) Metabolome and water status phenotyping of Arabidopsis under abiotic stress cues reveals new insight into ESK1 function. Plant Cell Environ 32:95–108
Lynch JP (1995) Root architecture and plant productivity. Plant Physiol 109:7–13
Lynch JP (2007) Roots of the second green revolution. Aust J Bot 55:494–512
Maeshima M, Ishikawa F (2008) ER membrane aquaporins in plants. Pflugers Arch Eur J Physiol 456:709–716
Manschadi AM, Christopher J, deVoil P, Hammer GL (2006) The role of root architectural traits in adaptation of wheat to water-limited environments. Funct Plant Biol 33:823–837
Martin JH, Leonard WH, Stamp DL (1976) Principles of field crop production. Macmillan, New York
Martre P, Morillon R, Barrieu F, North GB, Nobel PS, Chrispeels MJ (2002) Plasma membrane aquaporins play a significant role during recovery from water deficit. Plant Physiol 130:2101–2110
Masle J, Farquhar GD, Gilmore SR (2004) Method of producing plants having enhanced transpiration efficiency and plants produced there from. International application, Patent Cooperation Treaty, WO 2004/005555 A1
Masle J, Gilmore SR, Farquhar GD (2005) The ERECTA gene regulates plant transpiration efficiency in Arabidopsis. Nature 436:866–870
Maurel C (1997) Aquaporins and water permeability of plant membranes. Annu Rev Plant Physiol Plant Mol Biol 48:399–429
Maurel C, Santoni V, Luu D-T, Wudick MM, Verdoucq L (2009) The cellular dynamics of plant aquaporin expression and functions. Curr Opin Plant Biol 12:690–698
McIntyre CL, Seung D, Casu RE, Rebetzke GJ, Shorter R, Xue GP (2012) Genotypic variation in the accumulation of water soluble carbohydrates in wheat. Funct Plant Biol 39:560–568
Morgan JM (1984) Osmoregulation and water stress in higher plants. Annu Rev Plant Physiol 35:299–319
Mulroy TW (1979) Spectral properties of heavily glaucous and non-glaucous leaves of a succulent rosette-plant. Oecologia 38:349–357
Munns R, James RA, Sirault XRR, Furbank RT, Jones HG (2010) New phenotyping methods for screening wheat and barley for beneficial responses to water deficit. J Exp Bot 61:3499–3507
Mwanamwenge J, Loss SP, Siddique KHM, Cocks PS (1998) Growth, seed yield and water use of faba bean (Vicia faba L) in a short-season Mediterranean-type environment. Aust J Exp Agric 38:171–180
Nadeau JA (2009) Stomatal development: new signals and fate determinants. Curr Opin Plant Biol 12:29–35
Nakashima K, Ito Y, Yamaguchi-Shinozaki K (2009) Transcriptional regulatory networks in response to abiotic stresses in Arabidopsis and grasses. Plant Physiol 149:88–95
Nardini A, Salleo S, Andri S (2005) Circadian regulation of leaf hydraulic conductance in sunflower (Helianthus annuus L. cv Margot). Plant Cell Environ 28:750–759
Nicotra AB, Leigh A, Boyce CK, Jones CS, Niklas KJ, Royer DL, Tsukaya H (2011) The evolution and functional significance of leaf shape in the angiosperms. Funct Plant Biol 38:535–552
Niinemets Ü (2001) Global-scale climatic controls of leaf dry mass per area, density, and thickness in trees and shrubs. Ecology 82:453–469
Nobel PS (1980) Leaf anatomy and water use efficiency. In: Turner NC, Kramer PJ (eds) Adaptation of plants to water and high temperature stress. Wiley Interscience, New York, pp 43–55
Nobel PS (1999) Physicochemical and environmental plant physiology. Academic, San Diego, CA
Nobel PS (2009) Physicochemical and environmental plant physiology. Academic, Oxford
O’Brien TP, Carr DJ (1970) A suberized layer in the cell walls of the bundle sheath of grasses. Aust J Biol Sci 23:275–288
Oppenheimer HR (1960) Adaptation to drought, xerophytism. Arid zone research. XV. Plant water relationships in arid and semi-arid conditions. UNESCO, Paris, pp 105–138
Palta JAP, Berger J, Bramley H (2012) Physiology of the yield under drought: lessons from studies with lupin. In: Aroca R (ed) Plant responses to drought stress: from morphological to molecular features. Springer, Heidelberg, pp 417–440
Pandey RK, Herrera WAT, Pendleton JW (1984) Drought response of grain legumes under irrigation gradient. II. Plant water status and canopy temperature. Agron J 76:553–557
Parent B, Hachez C, Redondo E, Simonneau T, Chaumont F, Tardieu F (2009) Drought and abscisic acid effects on aquaporin content translate into changes in hydraulic conductivity and leaf growth rate: a trans-scale approach. Plant Physiol 149:2000–2012
Parry MAJ, Andralojc PJ, Mitchell RA, Madgwick PJ, Keys AJ (2003) Manipulation of RuBisCO: the amount, activity, function and regulation. J Exp Bot 54:1321–1333
Passioura JB (1972) The effect of root geometry on the yield of wheat growing on stored water. Aust J Agric Res 23:745–752
Passioura JB (1976) Physiology of grain yield in wheat growing on stored water. Aust J Plant Physiol 3:559–565
Passioura JB, Angus JF (2010) Improving productivity of crops in water-limited environments. Adv Agron 106:37–75
Perumalla CJ, Peterson CA, Enstone DE (1990) A survey of angiosperm species to detect hypodermal Casparian bands. I. Roots with a uniseriate hypodermis and epidermis. Bot J Linn Soc 103:93–112
Pinter PJ, Jackson RD, Ezra CE (1985) Sun-angle and canopy-architecture effects on the spectral reflectance of six wheat cultivars. Int J Remote Sens 6:1813–1825
Quarrie SA, Jones HG (1977) Effects of abscisic acid and water stress on development and morphology of wheat. J Exp Bot 28:192–203
Quintero JM, Fournier JM, Benlloch M (1999) Water transport in sunflower root systems: effects of ABA, Ca2+ status and HgCl2. J Exp Bot 50:1607–1612
Rascio A, Cedola MC, Sorrentino G, Pastore D, Wittmer G (1988) Pressure-volume curves and drought resistance in two wheat genotypes. Physiol Plant 73:122–127
Ratnakumar P, Vadez V (2011) Groundnut (Arachis hypogaea) genotypes tolerant to intermittent drought maintain a high harvest index and have small leaf canopy under stress. Funct Plant Biol 38:1016–1023
Rawson HM, Begg JE (1977) The effect of atmospheric humidity on photosynthesis, transpiration and water use efficiency of leaves of several plant species. Planta 134:5–10
Regan KL, Siddique KHM, Turner NC, Whan BR (1992) Potential for increasing early vigour and total biomass in spring wheat. II. Characteristics associated with early vigour. Aust J Agric Res 43:541–553
Reynolds MP, Tuberosa R (2008) Translational research impacting on crop productivity in drought-prone environments. Curr Opin Plant Biol 11:171–179
Reynolds MP, Ortiz-Monasterio JI, McNab A (eds) (2001) Application of physiology of wheat breeding. CIMMYT, DF Mexico
Reynolds MP, Pierre CS, Saad ASI, Vargas M, Condon AG (2007) Evaluating potential genetic gains in wheat associated with stress-adaptive trait expression in elite genetic resources under drought and heat stress. Crop Sci 47:S172–S189
Reynolds MP, Manes Y, Izanloo A, Langridge P (2009) Phenotyping approaches for physiological breeding and gene discovery in wheat. Ann Appl Biol 155:309–320
Richards RA (2006) Increasing crop productivity when water is scarce—from breeding to field management. Agric Water Manage 80:176–196
Richards RA, Passioura JB (1989) A breeding program to reduce the diameter of the major xylem vessel in the seminal roots of wheat and its effect on grain yield in rain-fed environments. Aust J Agric Res 40:943–950
Richards RA, Rawson HM, Johnson DA (1986) Glaucousness in wheat: its development and effect on water-use efficiency, gas exchange and photosynthetic tissue temperatures. Aust J Plant Physiol 13:465–473
Richards RA, Rebetzke GJ, Condon AG, van Herwaarden AF (2002) Breeding opportunities for increasing the efficiency of water use and crop yield in temperate cereals. Crop Sci 42:111–121
Rieger M, Duemmel MJ (1992) Comparison of drought resistance among Prunus species from divergent habitats. Tree Physiol 11:369–380
Rieger M, Litvin P (1999) Root system hydraulic conductivity in species with contrasting root anatomy. J Exp Bot 50:201–209
Rowe MH, Bergmann DC (2010) Complex signals for simple cells: the expanding ranks of signals and receptors guiding stomatal development. Curr Opin Plant Biol 13:548–555
Sack L, Holbrook NM (2006) Leaf hydraulics. Annu Rev Plant Biol 57:361–381
Sade N, Vinocur BJ, Diber A, Shatil A, Ronen G, Nissan H, Wallach R, Karchi H, Moshelion M (2009) Improving plant stress tolerance and yield production: is the tonoplast aquaporin SlTIP2;2 a key to isohydric to anisohydric conversion? New Phytol 181:651–661
Sade N, Gebretsadik M, Seligmann R, Schwartz A, Wallach R, Moshelion M (2010) The role of tobacco aquaporin1 in improving water use efficiency, hydraulic conductivity, and yield production under salt stress. Plant Physiol 152:245–254
Sagaram M, Lombardini L, Grauke LJ (2007) Variation in leaf anatomy of pecan cultivars from three ecogeographic locations. J Am Soc Hortic Sci 132:592–596
Sakurai J, Ishikawa F, Yamaguchi T, Uemura M, Maeshima M (2005) Identification of 33 rice aquaporin genes and analysis of their expression and function. Plant Cell Physiol 46:1568–1577
Saliendra NZ, Meinzer FC (1992) Genotypic, developmental and drought-induced differences in root hydraulic conductance of contrasting sugarcane cultivars. J Exp Bot 43:1209–1217
Schuepp PH (1993) Leaf boundary layers. New Phytol 125:477–507
Schulze E-D, Turner NC, Nicoll D, Schumacher J (2006) Leaf and wood carbon isotope ratios, specific leaf areas and wood growth of Eucalyptus species across a rainfall gradient in Australia. Tree Physiol 26:479–492
Sexton PJ, Peterson CM, Boote KJ, White JW (1997) Early-season growth in relation to region of domestication, seed size, and leaf traits in common bean. Field Crops Res 52:69–78
Sharkey TD (1988) Estimating the rate of photorespiration in leaves. Physiol Plant 63:147–152
Sharma HC, Waines JG (1994) Inheritance of leaf pubescence in diploid wheat. J Hered 85:286–288
Shelden MC, Howitt SM, Kaiser BN, Tyerman SD (2009) Identification and functional characterisation of aquaporins in the grapevine, Vitis vinifera. Funct Plant Biol 36:1065–1078
Shimazaki K-i, Doi M, Assmann SM, Kinoshita T (2007) Light regulation of stomatal movement. Annu Rev Plant Biol 58:219–247
Siddique KHM, Belford RK, Perry MW, Tennant D (1989) Growth, development and light interception of old and modern wheat cultivars in a Mediterranean-type environment. Aust J Agric Res 40:473–487
Siddique KHM, Belford RK, Tennant D (1990a) Root:shoot ratios of old and modern, tall and semi-dwarf wheats in a Mediterranean environment. Plant Soil 121:89–98
Siddique KHM, Tennant D, Perry MW, Belford RK (1990b) Water use and water use efficiency of old and modern wheat cultivars in a Mediterranean-type environment. Aust J Agric Res 41:431–447
Siddique KHM, Regan KL, Tennant D, Thomson BD (2001) Water use and water use efficiency of cool season grain legumes in low rainfall Mediterranean-type environments. Eur J Agron 15:267–280
Siefritz F, Biela A, Eckert M, Otto B, Uehlein N, Kaldenhoff R (2001) The tobacco plasma membrane aquaporin NtAQP1. J Exp Bot 52:1953–1957
Siefritz F, Tyree MT, Lovisolo C, Schubert A, Kaldenhoff R (2002) PIP1 plasma membrane aquaporins in tobacco: from cellular effects to function in plants. Plant Cell 14:869–876
Simonin KA, Limm EB, Dawson TE (2012) Hydraulic conductance of leaves correlates with leaf lifespan: implications for lifetime carbon gain. New Phytol 193:939–947
Sinclair TR, Zwieniecki MA, Holbrook NM (2008) Low leaf hydraulic conductance associated with drought tolerance in soybean. Physiol Plant 132:446–451
Sinclair TR (2012) Is transpiration efficiency a viable plant trait in breeding for crop improvement? Funct Plant Biol 39:359–365
Sivamani E, Bahieldin A, Wraith JM, Al-Niemi T, Dyer WE, Ho TD, Qu R (2000) Improved biomass productivity and water use efficiency under water deficit conditions in transgenic wheat constitutively expressing the barley HVA1 gene. Plant Sci 155:1–9
Slack CR, Hatch MD (1967) Comparative studies on the activity of carboxylases and other enzymes in relation to the new pathway of photosynthetic carbon dioxide fixation in tropical grasses. Biochem J 103:660–665
Smith JAC, Popp M, Lüttge U, Cram WJ, Diaz M, Griffiths H, Lee HSJ, Medina E, Schäfer C, Stimmel KH, Thonke B (1989) Ecophysiology of xerophytic and halophytic vegetation of a coastal alluvial plain in northern Venezuela. VI. Water relations and gas exchange of mangroves. New Phytol 111:293–307
Spollen WG, LeNoble ME, Sammuels TD, Bernstein N, Sharp RE (2000) Abscisic acid accumulation maintains maize primary root elongation at low water potentials by restricting ethylene production. Plant Physiol 122:967–976
Spreitzer RJ, Salvucci ME (2002) RuBisCO: structure, regulatory interactions, and possibilities for a better enzyme. Annu Rev Plant Biol 53:449–475
Stark JC, Pavek JJ, McCann IR (1991) Using canopy temperature measurements to evaluate drought tolerance of potato genotypes. J Am Soc Hortic Sci 116:412–415
Stasovski E, Peterson CA (1991) The effects of drought and subsequent rehydration on the structure and vitality of Zea mays seedling roots. Can J Bot 69:1170–1178
Steudle E (2000) Water uptake by roots: effects of water deficit. J Exp Bot 51:1531–1542
Steudle E, Jeschke WD (1983) Water transport in barley roots. Planta 158:237–248
Steudle E, Peterson CA (1998) How does water get through roots? J Exp Bot 49:775–788
Steudle E, Murrmann M, Peterson CA (1993) Transport of water and solutes across maize roots modified by puncturing the endodermis. Plant Physiol 103:335–349
Sutka M, Li G, Boudet J, Boursiac Y, Doumas P, Maurel C (2010) Natural variation of root hydraulics in Arabidopsis grown in normal and salt-stressed conditions. Plant Physiol 155:1264–1276
Svenningsson M, Liljenberg C (1986) Changes in cuticular transpiration rate and cuticular lipids of oat (Avena sativa) seedlings induced by water stress. Physiol Plant 66:9–14
Tanaka Y, Fujii K, Shiraiwa T (2010) Variability of leaf morphology and stomatal conductance in soybean [Glycine max (L.) Merr.] cultivars. Crop Sci 50:2525–2532
Tanner CB, Sinclair TR (1983) Efficient water use in crop production: research and re-search? In: Taylor HM, Jordan WR, Sinclair TR (eds) Limitations to efficient water use in crop production. American Society of Agronomy, Madison, WI, pp 1–27
Thompson AJ, Andrews J, Mulholland BJ, McKee JMT, Hilton HW, Horridge JS, Farquhar GD, Smeeton RC, Smillie IRA, Black CR, Taylor IB (2007) Overproduction of abscisic acid in tomato increases transpiration efficiency and root hydraulic conductivity and influences leaf expansion. Plant Physiol 143:1905–1917
Ting IP (1985) Crassulacean acid metabolism. Annu Rev Plant Physiol 36:595–622
Torii KU (2004) Leucine-rich repeat receptor kinases in plants: structure, function, and signal transduction pathways. Int Rev Cytol 234:1–46
Torii KU (2012) Mix-and-match: ligand-receptor pairs in stomatal development and beyond. Trends Plant Sci 17(12):711–719
Torii KU, Mitsukawa N, Oosumi T, Matsuura Y, Yokoyama R, Whittier RF, Komeda Y (1996) The Arabidopsis ERECTA gene encodes a putative receptor protein kinase with extracellular leucine-rich repeats. Plant Cell 8:735–746
Travaglia C, Reinoso H, Cohen A, Luna C, Tommasino E, Castillo C, Bottina R (2010) Exogenous ABA increases yield in field-grown wheat with moderate water restriction. J Plant Growth Regul 29:366–374
Tsuda M, Tyree MT (2000) Plant hydraulic conductance measured by the high pressure flow meter in crop plants. J Exp Bot 51:823–828
Turner NC (1972) K+ uptake of guard cells stimulated by fusicoccin. Nature 235:341–342
Turner NC (1993) Water use efficiency of crop plants: potential for improvement. In: Buxton DR, Shibles R, Forsberg RA, Blad BL, Asay KH, Paulsen GM, Wilson RF (eds) International crop science 1. Crop Science Society of America, Madison, WI, pp 75–82
Turner NC (1997) Further progress in crop water relations. Adv Agron 58:293–337
Turner NC, Asseng S (2005) Productivity, sustainability, and rainfall-use efficiency in Australian rainfed Mediterranean agricultural systems. Aust J Agric Res 56:1123–1136
Turner NC, Hartung W (2012) Dehydration of isolated roots of seven Lupinus species induces synthesis of different amounts of free, but not conjugated, abscisic acid. Plant Growth Regul 66:265–269
Turner NC, Jones MM (1980) Turgor maintenance by osmotic adjustment: a review and evaluation. In: Turner NC, Kramer PJ (eds) Adaptation of plants to water and high temperature stress. Wiley Interscience, New York, pp 87–103
Turner NC, Nicolas ME (1998) Early vigour: a yield-positive characteristic for wheat in drought-prone Mediterranean-type environments. In: Behl RK, Singh DP, Lodhi GP (eds) Crop improvement for stress tolerance. CCS Haryana Agricultural University/Max Mueller Bhawan, Hisar/New Delhi
Turner NC, Schulze E-D, Gollan T (1984) The responses of stomata and leaf gas exchange to vapour pressure deficits and soil water content. I. Species comparisons at high soil water contents. Oecologia 63:338–342
Turner NC, Abbo S, Berger JD, Chaturvedi SK, French RJ, Ludwig C, Mannur DM, Singh SJ, Yadava HS (2007a) Osmotic adjustment in chickpea (Cicer arietinum L.) results in no yield benefit under terminal drought. J Exp Bot 58:187–194
Turner NC, Palta JA, Shrestha R, Ludwig C, Siddique KHM, Turner DW (2007b) Carbon isotope discrimination is not correlated with transpiration efficiency in three cool-season grain legumes (pulses). J Integr Plant Biol 49:1478–1483
Tyerman SD, Bohnert HJ, Maurel C, Steudle E, Smith JAC (1999) Plant aquaporins: their molecular biology, biophysics and significance for plant water relations. J Exp Bot 50:1055–1071
Tyerman SD, Niemietz CM, Bramley H (2002) Plant aquaporins: multifunctional water and solute channels with expanding roles. Plant Cell Environ 25:173–194
Tyree MT, Fiscus EL, Wullschleger SD, Dixon MA (1986) Detection of xylem cavitation in corn under field conditions. Plant Physiol 82:597–599
Uehlein N, Lovisolo C, Siefritz F, Kaldenhoff R (2003) The tobacco aquaporin NtAQP1 is a membrane CO2 pore with physiological functions. Nature 425:734–737
Uehlein N, Otto B, Hanson DT, Fischer M, McDowell N, Kaldenhoff R (2008) Function of Nicotiana tabacum aquaporins as chloroplast gas pores challenges the concept of membrane CO2 permeability. Plant Cell 20:648–657
Ueno O, Kawano Y, Wakayama M, Takeda T (2006) Leaf vascular systems in C3 and C4 grasses: a two-dimensional analysis. Ann Bot 97:611–621
Vadez V, Krishnamurthy L, Kashiwagi J, Kholova J, Devi JM, Sharma KK, Bhatnagar-Mathur P, Hoisington DA, Hash CT, Bidinger FR, Keatinge JDH (2007) Exploiting the functionality of root systems for dry, saline, and nutrient deficient environments in a changing climate. SAT eJournal. ICRISAT, Pattancheru, pp 1–61
Vandeleur RK, Mayo G, Shelden MC, Gilliham M, Kaiser BN, Tyerman SD (2009) The role of plasma membrane intrinsic protein aquaporins in water transport through roots: diurnal and drought stress responses reveal different strategies between isohydric and anisohydric cultivars of grapevine. Plant Physiol 149:445–460
Vysotskaya LB, Kudoyarova GR, Jones HG (2004) Unusual stomatal behaviour on partial root excision in wheat seedlings. Plant Cell Environ 27:69–77
Wan X, Steudle E, Hartung W (2004) Gating of water channels (aquaporins) in cortical cells of young corn roots by mechanical stimuli (pressure pulses): effects of ABA and of HgCl2. J Exp Bot 55:411–422
Warren CR (2008) Stand aside stomata, another actor deserves centre stage: the forgotten role of the internal conductance to CO2 transfer. J Exp Bot 59:1475–1487
Wasson AP, Richards RA, Chatrath R, Misra SC, Prasad SVS, Rebetzke GJ, Kirkegaard JA, Christopher J, Watt M (2012) Traits and selection strategies to improve root systems and water uptake in water-limited wheat crops. J Exp Bot 63:3485–3498
Watt M, Schneebeli K, Dong P, Wilson IW (2009) The shoot and root growth of Brachypodium and its potential as a model for wheat and other cereal crops. Funct Plant Biol 36:960–969
Webster JA, Inayatullah C, Hamissou M, Mirkes KA (1994) Leaf pubescence effects in wheat on yellow sugarcane aphids and greenbugs (Homoptera: Aphididae). J Econ Entomol 87:231–240
Wilson D, Cooper JP (1969) Effect of temperature during growth on leaf anatomy and subsequent light-saturated photosynthesis among contrasting Lolium genotypes. New Phytol 68:1115–1123
Wright GC, Nageswara R, Farquhar GD (1994) Water-use efficiency and carbon isotope discrimination in peanut under water deficit conditions. Crop Sci 34:92–97
Xin Z, Mandaokar A, Chen J, Last RL, Browse J (2007) Arabidopsis ESK1 encodes a novel regulator of freezing tolerance. Plant J 49:786–799
Xing HT, Guo P, Xia XL, Yin WL (2011) PdERECTA, a leucine-rich repeat receptor-like kinase of poplar, confers enhanced water use efficiency in Arabidopsis. Planta 234:229–241
Xu Z, Zhou G (2008) Responses of leaf stomatal density to water status and its relationship with photosynthesis in a grass. J Exp Bot 59:3317–3325
Yambao EB, Ingram KT, Real JG (1992) Root xylem influence on the water relations and drought resistance of rice. J Exp Bot 43:925–932
Yoo CY, Pence HE, Hasegawa PM, Mickelbart MV (2009) Regulation of transpiration to improve crop water use. Crit Rev Plant Sci 28:410–431
Yoo CY, Pence HE, Jin JB, Miura K, Gosney MJ, Hasegawa PM, Mickelbart MV (2010) The Arabidopsis GTL1 transcription factor regulates water use efficiency and drought tolerance by modulating stomatal density via transrepression of SDD1. Plant Cell 22:4128–4141
Yunasa IAM, Siddique KHM, Belford RK, Karimi MM (1993) Effect of canopy structure on efficiency of radiation interception and use in spring wheat cultivars during the pre-anthesis period in a Mediterranean-type environment. Field Crops Res 35:113–122
Yunasa IAM, Sedgley RH, Siddique KHM (1994) Influence of mulching on the pattern of growth and water use by spring wheat and moisture storage on a fine textured soil. Plant Soil 160:119–130
Zaman-Allah M, Jenkinson DM, Vadez V (2011) A conservative pattern of water use, rather than deep or profuse rooting, is critical for the terminal drought tolerance of chickpea. J Exp Bot 62:4239–4252
Zhang J, Davies WJ (1989) Sequential response of whole plant water relations to prolonged soil drying and the involvement of xylem sap ABA in the regulation of stomatal behaviour of sunflower plants. New Phytol 113:167–174
Zhang J, Davies WJ (1990) Changes in the concentration of ABA in xylem sap as a function of changing soil water status can account for changes in leaf conductance and growth. Plant Cell Environ 13:277–285
Zhang X, Wollenweber B, Jiang D, Liu F, Zhao J (2008) Water deficits and heat shock effects on photosynthesis of a transgenic Arabidopsis thaliana constitutively expressing ABP9, a bZIP transcription factor. J Exp Bot 59:839–848
Zhu J, Brown KM, Lynch JP (2010) Root cortical aerenchyma improves the drought tolerance of maize (Zea mays L.). Plant Cell Environ 33:740–749
Zimmermann HM, Hartmann K, Schreiber L, Steudle E (2000) Chemical composition of apoplastic transport barriers in relation to radial hydraulic conductivity of corn roots (Zea mays L.). Planta 210:302–311
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2013 Springer-Verlag Berlin Heidelberg
About this chapter
Cite this chapter
Bramley, H., Turner, N.C., Siddique, K.H.M. (2013). Water Use Efficiency. In: Kole, C. (eds) Genomics and Breeding for Climate-Resilient Crops. Springer, Berlin, Heidelberg. https://doi.org/10.1007/978-3-642-37048-9_6
Download citation
DOI: https://doi.org/10.1007/978-3-642-37048-9_6
Published:
Publisher Name: Springer, Berlin, Heidelberg
Print ISBN: 978-3-642-37047-2
Online ISBN: 978-3-642-37048-9
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)