
Abstract
Bacilli are aerobic or facultatively anaerobic, Gram-positive or variable, endospore-forming bacteria that exhibit resistance to environmental stress and produce peptide antibiotics, peptide signal molecules, and extracellular enzymes. Bacillus and Paenibacillus genera include the best knowing nitrogen-fixing species. Another characteristic of bacilli is their great potential in producing substances that promote direct plant growth by the production of phytohormones (mainly indolic compounds), low-molecular-weight iron-binding molecules with a high affinity for Fe3+ (siderophores), and phosphate solubilization. It is well known that many species of Bacillus and Paenibacillus can contribute to plant growth and health in many ways, and most commercially developed PGPR are species of Bacillus which form endospores that confer population stability during formulation and storage of products. The identification and the isolation of PGP bacilli from soils, which combine the ability to fix nitrogen with the production of substances capable of promoting plant growth, could also significantly increase the productivity of many grain crops around the world.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
- Indole Acetic Acid
- Rhizosphere Soil
- Promote Plant Growth
- Indole Acetic Acid Production
- Paenibacillus Species
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
1.1 Introduction
Aerobic endospore-forming bacteria (AEFB) are essentially ubiquitous in agricultural systems. Common physiological traits important to their survival include the production of a multilayered cell wall structure, formation of stress-resistant endospores, and secretion of peptide antibiotics, peptide signal molecules, and extracellular enzymes. However, significant variation exists in other key traits, including nutrient utilization, motility, and physiochemical growth optima. Their microscopic size and omnipresence in soils facilitates their colonization of plants and animals, but the degree of niche localization of most species has not been thoroughly studied. Multiple Bacillus and Paenibacillus spp. can be readily cultured from both bulk and rhizosphere soils. Indeed, the ecological significance of the genotypic and phenotypic diversity of named species of Bacillus and related genera remains largely a mystery (Gardener 2004).
Analysis of the DNA extracted directly from soil samples, especially those that use the sequencing of the 16S ribosomal RNA genes (16S rRNA), has confirmed the occurrence of easily cultivable bacterium species, as well as a wide variety of noncultivable strains of species that belong to the genera Bacillus and Paenibacillus (Borneman et al. 1996; Felske et al. 1999; Garbeva et al. 2003; Smalla et al. 2001). Nevertheless, evidence of the relative number of cultivable and noncultivable representatives of these genera in different soils is surrounded by controversy. Some studies have suggested that most 16S rRNA sequences of bacilli isolated directly from soil samples are very similar to the sequences of cultivable and named species (Garbeva et al. 2003; Smalla et al. 2001). Other authors report that the predominant sequences found in different soils are not the same as those presented by bacilli isolated and easily cultivable (Borneman et al. 1996; Felske et al. 1999).
In turn, niche specificity and important ecological activities in Bacillus and Paenibacillus spp. appear to span phylogenetic boundaries. Most species can survive as saprophytes in soils, which are considered the primary reservoirs of these bacteria; however, most viable cells probably occur as inactive spores at any given time (Nicholson 2002). Furthermore, multiple species can be recovered as epiphytes and endophytes of plants and animals, as well as foodstuffs and composts derived from them (Priest 1993; Stahly et al. 1992; Slepecky and Hemphill 1992). The rich variety of organic substrates and microniches present in those environments supports a complex milieu of microbial species, so it is perhaps not surprising that multiple species of Bacillus and Paenibacillus inhabit them (Gardener 2004).
Numerous Bacillus and Paenibacillus species may contribute to the health status of plants in several ways. A considerable number of isolates belonging to these genera have been used as agents of biological control against phytopathogens (Lacey et al. 2001; Siddiqui and Mahmood 1999). Yet, the successful application of these agents requires comprehensive knowledge of their ecology. Additionally, the safety and efficacy of inoculants are largely determined by the ecological success of strains in the environments they are to be introduced. Better knowledge of the diversity, distribution, and action of bacteria of the Bacillus and Paenibacillus genera is extremely important to the identification of new strains, to the formulation of inoculants, and to the determination of the kinds of plantation they may be employed in Gardener (2004).
Members of the genus Paenibacillus are facultatively anaerobic organisms that produce spores in definitely swollen sporangia and have G + C contents ranging from 45 to 54 mol%. Some of these organisms excrete diverse assortments of extracellular polysaccharide-hydrolyzing enzymes to hydrolyze complex carbohydrates, including alginate, chondroitin, chitin, curdlan, and other polysaccharides (Shida et al. 1997). Also, a number of species under these genera are known to produce polysaccharides (Yoon et al. 2002), antifungal, and antimicrobial agents, such as polymyxin, octopityn, and baciphelacin (Chung et al. 2000).
Paenibacillus species have been isolated from a wide range of sources, including soil, water, plant rhizosphere, tree roots, plant material, foods, forage, feces, and insect larvae (Daane et al. 2002). Bacterial isolates from this genus have also been detected in Lake Vostok, Antarctica (Christner et al. 2001), in the ice caps of mountains, China (Christner et al. 2000), in paintings that have undergone biological deterioration (Heyrman and Swings 2001), in seawaters (Siefert et al. 2000), in mercury-reducing biofilms (Wagner-Döbler et al. 2000), and in estuarial sediments contaminated with crude oil (Daane et al. 2002). The genus was also observed to be abundant in companies that produce paper used in the food industry (Raaska et al. 2002).
Several species described as nitrogen-fixing bacilli belong to the Paenibacillus genus, as for example P. polymyxa (Grau and Wilson 1962), P. macerans, P. durus (P. azotofixans), P. peoriae (Montefusco et al. 1993), P. borealis (Elo et al. 2001), P. graminis and P. odorifer (Berge et al. 2002), P. brasilensis (von der Weid et al. 2002), P. massiliensis (Roux and Raoult 2004), P. wynnii (Rodríguez-Diaz et al. 2005), P. sabinae (Ma et al. 2007a), P. zanthoxyli (Ma et al. 2007b), P. donghaensis (Choi et al. 2008), P. forsythiae (Ma and Chen 2008), P. sonchi (Hong et al. 2009), and P. riograndensis (Beneduzi et al. 2010).
Concerning the bacteria belonging to the Bacillus genus, in 1998, Xie et al. detected the activity of the nitrogenase enzyme in B. megaterium, B. cereus, B. pumilus, B. circulans, B. licheniformis, B. subtilis, B. brevis, and B. firmus. These same authors reported, in 2003, the isolation of 14 Bacillus strains that were able to reduce acetylene in paddy fields of eight locations on the banks of river Yangtze, China. Li et al. (1992) had also identified a Bacillus species that fixed nitrogen in association with ectomycorrhizas, while Ahmad et al. (2008) have detected nitrogen-fixing Bacillus isolates in different rhizosphere soils in Aligarh, India. Bacillus fusiformis (strains PM-5 and PM-24) has also been identified as a nitrogen-fixing bacterium using the acetylene reduction test. This species was shown to exhibit an intense nitrogenase activity in different plant cultures in the province of Chungbuk, South Korea. Diazotrophic Bacilllus were even found in the rhizosphere of pine and oak trees (Rózycki et al. 1999). Ding et al. (2005) detected biological nitrogen activity in B. marisflavi and Paenibacillus massiliensis and also identified fragments of the nifH gene in B. megaterium and B. cereus. B. alkalidiazotrophicus (Sorokin et al. 2008a), a low salt-tolerant alkaliphile isolated from Mongolian soda soil, was also described as a nitrogen-fixing bacterium. Therefore, it was demonstrated that both genera, Bacillus and Paenibacillus, presented species that are able to fix nitrogen. It is interesting to note that Natronobacillus, a new bacilli genus, was created specifically for N. azotifigens, its only species, which is an anaerobic diazotrophic haloalkaliphile bacterium that was isolated from soda-rich habitats (Sorokin et al. 2008b).
In recent years, the interest in soil microorganisms has increased as they play an important role in the maintenance of soil fertility. A major challenge for the development of sustainable agriculture lies in the use of nitrogen-fixing bacteria, which are able to assimilate gaseous N2 from the atmosphere (Seldin et al. 1998).
1.2 Plant Growth Promoting Rhizobacteria
Rhizosphere can be defined as any volume of soil specifically influenced by plant roots and/or in association with roots and hairs, and plant-produced material. This space includes the soil surrounding plant roots, often extending a few millimeters from the root surface and can include as well the plant root epidermal layer. Plant exudates in the rhizosphere, such as amino acids and sugars, provide a rich source of energy and nutrients for bacteria, resulting in higher bacterial populations in this area (Gray and Smith 2005). In fact, the concentration of bacteria (per gram of soil) found around plant roots (i.e., in the rhizosphere) is generally much bigger than the bacterial density, or concentration, detected in the rest of the soil (Lynch 1990). Despite the large number of bacteria in the rhizosphere, only 7–15% of the total root surface is generally occupied by microbial cells (Gray and Smith 2005).
The interaction between bacteria and plant roots may be beneficial, harmful, or neutral for the plant, and sometimes the effect of a particular bacterium may vary as a consequence of soil conditions (Lynch 1990). Thus, for example, a particular organism that facilitates plant growth by fixing nitrogen, which is usually present in the soil in limited amounts, is unlikely to provide benefit to a plant in a place where exogenous fixed nitrogen is added to the soil (Glick 1995).
Bacteria that provide some benefit to plants are of two general types: those that establish a symbiotic relationship with the plant and those that are free-living in the soil, but are often found near on or even within plant roots (Kloepper et al. 1988). Symbiotic bacteria, especially rhizobia, have been deeply studied, and although the world market for these microorganisms is nowadays relatively small, they have been developed as a biological means of increasing crop yields in certain circumstances. Beneficial free-living soil bacteria are usually referred to as plant growth promoting rhizobacteria (PGPRs, Kloepper et al. 1989), or, according to Piao et al. (1992), as yield increasing bacteria (YIBs). However, for Gray and Smith (2005), PGPRs also include rhizobia, due to the well-characterized legume–rhizobium symbiosis. Independently of the mechanisms of vegetal growth promotion, PGPRs have to colonize the rhizosphere around the roots, the rhizoplane (root surface), or the root itself (within radicular tissues). Among rhizobacteria, there is a gradient of root proximity and intimacy as follows: (1) bacteria living in the soil near to the roots, which utilize metabolites released by the roots as C and N sources, (2) bacteria colonizing the rhizoplane (radicular surface), (3) bacteria residing inside the root tissues, inhabiting spaces between cortical cells, and lastly (4) bacteria living inside cells in specialized root structures, or nodules, which generally fall into two groups: the legume-associated rhizobia and the woody plant-associated Frankia species. The cyanobacterial nitrogen-fixing symbionts of the cycads could also be included as PGPR (Gray and Smith 2005). A number of different bacteria may be considered to be PGPR, including species of the genera Azotobacter, Azospirillum, Pseudomonas, Acetobacter, Burkholderia, and bacilli (Bashan and Levanony 1990; Brown 1974; Elmerich 1984; Kloepper et al. 1988, 1989; Okon and Labandera-González 1994; Tang et al. 1994).
To aid in this conceptualization, two simple terms have been adopted: intracellular PGPRs (iPGPRs), which are bacteria that live inside plant cells, being localized in the nodules, and extracellular PGPRs (ePGPRs), which are those bacteria that live outside plant cells, being able to enhance plant growth through the production of signal compounds that directly stimulate plant growth, to the improvement of plant disease resistance, or to the mobilization of soil nutrients to the plant. ePGPR can be subdivided into three types, based on the degree of association with plant roots: those living near but not in contact with the roots; those colonizing the root surface; and those living in the spaces between cells of the root cortex (Gray and Smith 2005).
PGPRs can affect plant growth in two different ways: indirectly or directly. The direct promotion of plant growth by PGPRs entails either providing the plant with a compound that is synthesized by the bacterium or facilitating the uptake of certain nutrients from the environment (Glick 1995). PGPRs can fix atmospheric nitrogen and supply it to plants; they synthesize siderophores that can solubilize and sequester iron from the soil and provide it to plant cells; they synthesize several different phytohormones that can act to enhance various stages of plant growth; they may have mechanisms for the solubilization of minerals such as phosphorus that then become more readily available for plant growth; and they may synthesize some less well-characterized low-molecular mass compounds or enzymes that can modulate plant growth and development (Brown 1974; Kloepper et al. 1988, 1989). A particular PGPR may affect plant growth and development by using any one, or more, of these mechanisms. The indirect promotion of plant growth occurs when PGPRs lessen or prevent the deleterious effects of one or more phytopathogenic organisms.
Researches on ePGPRs were initially focused on Bacillus and Anthrobacter spp. (Brown 1974). Applications of these associations have been investigated in maize, wheat, oat, barley, peas, canola, soy, potatoes, tomatoes, lentils, radicchio, and cucumber (Gray and Smith 2005). Among the most widely studied ePGPR bacilli are B. cereus (Handelsman et al. 1990; Ryder et al. 1999), B. circulans (Berge et al. 1990), B. firmus, B. lichenformis (Chen et al. 1996), B. subtilis (Turner and Blackman 1991; Zhang and Smith 1996), and B. thuringiensis (Bai et al. 2002a, b).
1.3 Direct Benefits of ePGPRs for Plants
The extensive biochemical and molecular biological studies of symbiotic diazotrophs, such as rhizobia, have served as a conceptual starting point for the understanding of the mechanisms of growth promotion by PGPRs. Since one of the major benefits that rhizobia provide to the plants is fixed nitrogen, it was initially thought that diazotrophic PGPRs might also function in this way. However, not all PGPRs are diazotrophic, and many of those that are diazotrophic fix only limited amounts of nitrogen. Frequently, this amount is only enough for their own needs and is not sufficient to the host plant’s nitrogen requirements (Hong et al. 1991). Some diazotrophic PGPRs still provide their plant hosts with a portion of the fixed nitrogen that they require; however, even for these PGPRs, nitrogen fixation is only a minor component of the benefit they offer to the plant (Chanway and Holl 1991).
ePGPRs influence vegetal growth in different ways, depending on the species and strain. To promote plant growth free-living rhizobacteria resort to more than one mechanism (Glick et al. 1999). Apart from nitrogen fixation, several ePGPRs are also capable of providing sufficient iron (Fe) levels to the plant in soils that exhibit low concentrations of this element (Wang et al. 1993) or even of other important minerals, such as phosphates (Singh and Kapoor 1998). Some ePGPRs can produce and secrete low-molecular-weight (400–1,000 Da) iron-binding molecules (siderophores) with a high affinity for Fe3+ (Gray and Smith 2005). Although iron is one of the most abundant minerals on Earth, in the soil it is relatively unavailable for direct assimilation by several organisms. This is because in aerobic soils iron is found predominantly in the form of Fe3+, mainly as a constituent of oxyhydroxide polymers with extremely low solubility, about 10−18 M at neutral pH. Minimal concentrations of iron required for normal growth of plants range from 10−9 to 10−4 M, depending on other nutritional factors (Neilands et al. 1987). Similarly, minimal iron concentrations for the optimal growth of many microbes are approximately 10−5 to 10−7 M (Lankford 1973). To overcome this problem, soil microorganisms secrete low-molecular-weight iron-binding molecules (siderophores) that bind Fe3+, transport it back to the microbial cell, and then make it available for the microbial growth (Leong 1986).
The iron–siderophore binding also prevents the proliferation of pathogens due to the sequestration of iron from the environment. Some ePGPRs that present this property include Pseudomonas putida and P. aeruginosa. P. putida inhibits the growth of Fusarium oxysporum, a pathogen that affects tomatoes (Vandendergh and Gonzalez 1984), while P. aeruginosa inhibits the growth of Pythium, another pathogen that attacks tomatoes (Buysens et al. 1994).
Oppositely to microbial pathogens, plants do not suffer negative effects of iron depletion by ePGPRs. Some plants may capture the bacterial iron–siderophore complex, transporting it into its cells, where iron is released from the siderophores and is made available to the plant (Crowley et al. 1988). The production of siderophores has also been reported for Azospirillum lipoferum (Saxena et al. 1986; Shah et al. 1992), A. brasilense (Bachhawat and Ghosh 1987), and Azotobacter vinelandii (Demange et al. 1988; Knosp et al. 1984).
Phosphorus (P) exists in nature in a variety of organic (derived from microorganisms and plants) and inorganic (originating from applied P fertilizers) forms that are insoluble or very poorly soluble (Paul and Clark 1989). In fact, phosphorus is one of the least soluble elements in the natural environment, with less than 5% of the total soil phosphate content being available to the plants (Dobbelaere et al. 2003). Therefore, the addition of phosphate fertilizers has become a common practice in modern agriculture. However, a large portion of the soluble inorganic phosphate applied to soil as fertilizer is rapidly immobilized by the iron and aluminum in acid soils and by calcium in calcareous soils soon after application, thus becoming unavailable to plants (Holford 1997).
Soil microorganisms are able to solubilize insoluble mineral phosphate by producing various organic acids. This results in acidification of the surrounding soil, releasing soluble orthophosphate ions that can be readily taken up by plants. Furthermore, they are able to solubilize organic P compounds through the action of phosphatase enzymes (Garcia et al. 1992). In fact, the major source of phosphatase activity in soil is considered to be of microbial origin. In particular, phosphatase activity is substantially increased in the rhizosphere (Rodríguez and Fraga 1999). Phosphate solubilization has often been cited as a likely mechanism of promotion of vegetable growth by ePGPRs.
A large number of P-solubilizing bacteria (PSB) have been isolated from the rhizosphere of several crops. It was estimated that P-solubilizing microorganisms may constitute 20–40% of the cultivable population of soil microorganisms and that a significant proportion of them can be isolated from rhizosphere soil (Chabot et al. 1993). There have been a number of reports on plant growth promotion by bacteria that have the ability to solubilize inorganic and/or organic P from soil after their inoculation in soil or plant seeds. The production (by these strains) of other metabolites beneficial to the plant, such as phytohormones, antibiotics, or siderophores, among others, has created confusion about the specific role of phosphate solubilization in plant growth and yield stimulation (Kloepper et al. 1989).
Experiments performed with P-solubilizing diazotrophs are scarce, and the results obtained are quite diverse, varying according to plant or bacterial species. B. megaterium and P. polymyxa are able to enhance growth and yield but not the P uptake of canola, indicating that P-solubilization is not the main mechanism responsible for positive growth response (de Freitas et al. 1997). Nevertheless, studies with B. firmus (Datta et al. 1982) and P. polymyxa (Gaur and Ostwal 1972) have confirmed the increase in phosphorus uptake in rice and wheat yields, respectively, after inoculation with these bacteria.
Several reports have examined the ability of different bacterial species to solubilize insoluble inorganic phosphate compounds, such as tricalcium phosphate, dicalcium phosphate, hydroxyapatite, and rock phosphate. Among the bacterial genera with this capacity are Pseudomonas, Bacillus, Rhizobium, Burkholderia, Achromobacter, Agrobacterium, Micrococcus, Aerobacter, Flavobacterium, and Erwinia. There are considerable populations of phosphate-solubilizing bacteria in soil and in plant rhizospheres. These include both aerobic and anaerobic strains, with a prevalence of aerobic strains in submerged soils. A considerably higher concentration of phosphate-solubilizing bacteria is commonly found in the rhizosphere in comparison with nonrhizosphere soil (Rodríguez and Fraga 1999).
Phytohormones are natural plant growth regulators that influence physiological processes, when at low concentrations. Plant growth regulators are classified as auxins (cell differentiation, root and fruit growth, and abscission control), cytokinins (growth regulation, cell differentiation, and plant senescence), gibberellins (cell division and elongation, interruption of dormancy, and increase in fruit growth), abscisic acid (transpiration regulation, interruption of dormancy, and initial seed development), and ethylene (ripening of fruits, promotion of leaf, fruit and leaf abscission, and influence in female sexual expression) (Arshad and Frankenberger 1998; Raven et al. 1996). The production of these growth regulators has been reported in several bacteria including Gluconacetobacter, Azospirillum, Herbaspirillum, Methylobacterium, Erwinia, Pantoea, and Pseudomonas (Bastián et al. 1998; Cassán et al. 2001; Fuentes-Ramirez et al. 1993; Koenig et al. 2002; Lucangeli and Bottini 1997; Patten and Glick 1996; Verma et al. 2001) and has been consistently observed in bacteria that live in association with plants.
Phytohormone production is a bacterium–plant interaction mechanism influenced by numerous factors, as well as the plant and microorganism genotypes. Jain and Patriquin (1985) showed that pure Azospirillum endophytic strains, or culture filtrates, induced the ramification of wheat roots. The authors also observed that this phenomenon was influenced by the bacterial genome, at strain level, and by the plant’s genome, at cultivar level. The same work reported the isolation and characterization of indole acetic acid (IAA), an auxin produced by the bacterium, which was shown to play a role in root growth.
Although some Paenibacillus species are highly efficient in fixing nitrogen, the reason for the stimulation of plant growth is not directly related to this ability, but to its capacity to produce and secrete phytohormones, such as cytokinins (Timmusk et al. 1999) and auxins, especially IAA (Lebuhn et al. 1997).
Auxins (from the Greek auxein, to increase) are plant hormones originating from the amino acid tryptophan. The natural auxin is called IAA. Tryptophan is the precursor of IAA. Although there are four different pathways for IAA biosynthesis, all of them originated from tryptophan (Raven et al. 1996). The ability to synthesize phytohormones is widely observed in bacteria associated with plants – 80% of the bacteria isolated from rhizospheres are capable to produce IAA (Zakharova et al. 1999). The first studies were conducted in the 1970s. Tien et al. (1979) demonstrated that Azospirillum brasiliense, when exposed to tryptophan, produces IAA and lactic acid, and that IAA production increases by the time. Bacillus spp. isolated from the Phaseolus vulgaris rhizosphere produces significant amounts of IAA (Srinivasan et al. 1996). The isolates obtained from the rhizosphere of the genera Enterobacter, Xanthomonas, Pseudomonas, Alcaligenes, Azotobacter, Acetobacter, and Agrobacterium produce higher amounts of IAA than the soil isolates not associated with the roots (Asghar et al. 2002). In phytopathogenic bacteria, such as Agrobacterium tumefaciens and Pseudomonas syringae pathovars, IAA is produced from tryptophan via the intermediary indole acetamide and has been related to plant tumors. The beneficial bacteria synthesize IAA mainly through a tryptophan-dependent alternative pathway, from indole-pyruvic acid.
Promotion of root growth is one of the major markers by which the beneficial effect of PGPB is measured. Rapid establishment of roots, whether by elongation of primary roots or by proliferation of lateral and adventitious roots, is advantageous for young seedlings as it increases their ability to anchor themselves to the soil and to obtain water and nutrients from their environment, thus enhancing their chances for survival. Most root-promoting bacteria synthesize IAA, and their effect on plants mimics that of exogenous IAA (Patten and Glick 2002).
Production of IAA, a plant hormone that does not apparently function as a hormone in bacterial cells, may have evolved in bacteria due to its importance in the bacterium–plant relationship. IAA secreted by a bacterium may promote root growth directly by stimulating plant cell elongation or cell division or indirectly by influencing bacterial ACC deaminase activity (1-aminocyclopropane-1-carboxylate, a precursor of acetylene). However, the role of bacterial IAA in plant growth promotion remains undetermined (Patten and Glick 2002).
The response of plants to external (microbially released) IAA can vary from beneficial to deleterious effects, depending on the concentration of IAA in the plant roots. Low IAA concentrations may stimulate growth, while high IAA concentrations may inhibit root development. IAA levels released by rhizosphere bacteria depend on bacterial growth, metabolic activity, and the expression of genes encoding enzymes for IAA biosynthesis (Lambrecht et al. 2000). To assess these effects, different methodologies have been employed, such as the inoculation of roots with mutant bacteria for the production of IAA or the application of bacterial inocula at different concentrations. Patten and Glick (2002) utilized a P. putida strain that secreted high IAA contents and reported an increase of 35–50% in canola’s primary root growth. These authors provided direct evidence that bacterial IAA plays an important role in root elongation, when the producing bacterium is associated with the plant. Apart from this, these researchers provided stronger support to the hypothesis that beneficial bacteria produce IAA from pyruvic acid (IPyA), while pathogenic bacteria produce IAA from indole acetamide (IAM) (Manulis et al. 1998; Patten and Glick 1996; Prinsen et al. 1993). Extensive research has been carried out with mutant bacteria lacking the ability to synthesize IAA. These studies reported the existence of more than one tryptophan-dependent biosynthesis pathway for IAA production in the same microorganism. These biosynthesis pathways are classified in terms of their intermediary compounds, such as IAM or IPyA (Dobbelaere et al. 1999; Patten and Glick 1996).
The detection of auxins may be carried out according to a very precise, quantitative method, such as high performance liquid chromatography (HPLC) or even by a less accurate method, a specific and sensitive colorimetric reaction that uses the Salkowski reagent (Crozier et al. 1988). Different methodologies utilize the Salkowski reagent; some use microplates while others resort to nitrocellulose (Bric et al. 1991; Sarwar and Kremer 1992).
In a study with P. polymyxa strains isolated from wheat roots, Lebuhn et al. (1997) reported the production of IAA and indolic metabolites related to this compound in media supplemented or not with tryptophan. It was also observed that tryptophan greatly increased auxin production by these strains. These findings point out the importance of tryptophan as an IAA precursor. The isolates obtained from the rhizosphere produced more IAA as compared to those obtained directly from soil not associated with rhizosphere, suggesting that specific bacterial genotypes are selected by the host plants.
1.4 Promotion of Plant Growth by Bacilli
There is a long list of studies that demonstrate the beneficial effects of PGPRs, reporting that they promote significant improvement in plant growth and in the yield of several types of crops.
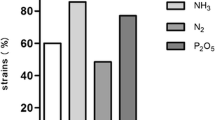
Beneduzi et al. (2008a) investigated several strains of bacilli, mainly species of the genera Bacillus and Paenibacillus, displaying important (PGP) characteristics isolated from seven distinct rice production zones of the Rio Grande do Sul State, south Brazil. Of those 296 isolates, 94 and 148 produced between 0.1 and 30 mg of (IAA) ml−1 in vitro after 72 and 144 h of incubation, respectively. Twenty-two isolates were able to solubilize phosphate and 32 isolates produced siderophores. Paenibacillus and Bacillus genera were the most prominent groups in the rhizosphere and soil populations analyzed. Paenibacillus borealis was the most frequent species in both locations. The isolate SVPR30, identified by 16S rRNA gene sequence analysis as a strain of Bacillus sp., was chosen for in vivo greenhouse experiments and proved to be very efficient in promoting a significant increase in the roots and shoot parts of rice plants (Fig. 1.1).

The effect of inoculation of a native PGPR isolate on rice growth promotion, 30 days after sprouting. (1) Plant was inoculated with SVPR30 strain; (2) positive control (plant was irrigated with mineral fertilizer solution); (3) negative control (plant was irrigated with distilled water); and (4) plant was inoculated with Paenibacillus polymyxa ATCC 10343 strain
In wheat crops, the genus Paenibacillus was the most prominent group in both the rhizosphere (77.8%) and soil (79%). P. borealis was the most frequently identified species, followed by P. graminis. The remainder of the isolated bacteria belonged to the genus Bacillus sp. Indolic compound production [indole-3-acetic acid (IAA), indole-pyruvic acid (IPyA), and IAM] was detected in 33.6 and 26% of the isolates from the rhizosphere and soil, respectively. Among the 311 isolates, 9 were able to solubilize phosphate and 48 were able to produce siderophores. The isolates SBR5, CSR16, and EsR7, identified by the 16S rRNA gene sequence as strains of Paenibacillus sp., were chosen for in vivo experiments in a greenhouse and proved to be very efficient in promoting a significant increase in the shoot and dry matter of wheat plants (Beneduzi et al. 2008b; Fig. 1.1).
Strains of Bacillus and Paenibacillus, which are Gram-positive ePGPRs, may be inoculated in separate or in consortium with Rhizobium or Bradyrhizobium strains, which are Gram-negative iPGPRs, to promote plant growth (Gray and Smith 2005). The inoculation of P. polymyxa strain H5, a phosphate-solubilizing bacterium, with Rhizobium led to an increase in chickpea yields due to the increase of phosphorus and nitrogen uptake (Alagawadi and Gaur 1988). Colonization and nodulation of soybean with B. japonicum strains (Li and Alexander 1988) may increase in media containing Bacillus spp., resulting in higher plant dry weight and seed yields. Bacillus spp. isolated from the rhizosphere of Phaseolus vulgaris produced significant amounts of IAA (Srinivasan et al. 1996). IAA promoted root growth and/or nodulation when R. etli was added to P. vulgaris cultures, causing an increase in nodule weight and number, in nitrogenase activity, and in leghemoglobin content. Applied alone, Bacillus spp. also plays a promoting role in the growth of several plants, which may vary depending on the type of soil (Ramos et al. 2003). Bacillus strains have also been proved to increase the growth of tomatoes, when used in the plant’s culture medium (Yan et al. 2003). B. subtilis promoted the growth of peanut seedlings, leading to an increase between 3.5 and 37% in crop yield and to improved germination, seedling emergence, plant nutrition, and root growth (Turner and Blackman 1991). B. lichenformis CECT 5106 and B. pumilus CECT 5105 inoculation enhanced growth rates of Pinus seedlings.
B. cereus UW85 increased soybean nodulation, both in laboratory tests and in the field, as well as in growth chambers (Halverson and Handelsman 1991). Interestingly, field experiments revealed differences in nodulation between 25 and 35 days after seeding; however, as the experiment evolved, differences in nodulation ceased to be detected as of 45 days of seedling. These results evince a promoter effect regarding initial growth, which nevertheless is not sustained in the long run. Yet, initial growth effects may be beneficial in the growth of young plants, assisting them to better cope with environmental stress they may have to face in their future development (Gray and Smith 2005).
Apart from fixing nitrogen, P. polymyxa solubilizes phosphorus in soils, produces antibiotics, chitinase, and other hydrolytic enzymes, increases soil porosity, and produces compounds that promote the growth of similar plants in media containing active IAA (Timmusk et al. 1999). It has also been demonstrated that this species increases nutrient uptake by the plant and acts as a biocontrol agent for pathogenic microorganisms (Chanway 1995; Mavingui and Heulin 1994), which is the reason for it been used as a biological control agent against Fusarium and Pythium (Guemouri-Athmani et al. 2000).
Paenibacillus durus is found in soils and roots of sugarcane, wheat, and other gramineous species. The interesting feature of this species lies in its capacity to biologically fix nitrogen, even in the presence of high nitrate levels, and to produce antimicrobial compounds (Neves and Rumjanek 1998). The capacity to fix nitrogen in vitro exhibited by P. durus is higher as compared to other Paenibacillus species, and because the bacterium is not affected by the presence of nitrate in the medium, fixing may occur in the presence of fertilizers (Rosado et al. 1996).
Li et al. (2008) conducted a study with soy nodules and isolated 98 nonsymbiotic endophytic bacterial strains that were characterized as being Pantoea, Serratia, Acinetobacter, Bacillus, and Burkholderia. The inoculation of endophytic bacteria did not lead to any significant effect in soy growth and nodulation, though several strains produced IAA, solubilized phosphate, and fixed nitrogen, a great promise in future studies on the plant growth promotion.
In the search for efficient PGPRs, Ahmad et al. (2008) have isolated and identified 72 bacteria as belonging to Azotobacter, Pseudomonas, Mesorhizobium, and Bacillus genera from the rhizospheres of wheat, sugarcane, onion, as well as chickpea nodules in Aligarh, India. These isolates were tested for antifungal activity. The Bacillus and Pseudomonas strains isolated were the most efficient and are candidates for further studies on plant growth promotion.
Park et al. (2005) isolated free-living fixing bacteria from the rhizosphere of seven different plants: sesame, maize, wheat, soy, lentil, pepper, and rice in Chungbuk, South Korea. The strains exhibiting nitrogenase activity were identified as Stenotrophomonas maltophila, B. fusiformis, and P. fluorescens. B. fusiformis exhibited the highest nitrogenase activity and IAA production rates, demonstrating its potential as rhizobacterium promoter of plant growth.
Other PGPRs isolated from the rhizospheres of melon, alfafa, tomato, cotton, and wheat were analyzed for the production of auxins, nitrogenase and antifungal activity, and also to the improvement in nutrient uptake in cotton and pea in a semiarid region in Uzbekistan (Egamberdiyeva and Höflich 2004). The strains were proved to produce auxins, to fix nitrogen, and to exert antagonistic action against Verticillium loteritum. The inoculation of the tested plants with Pseudomonas alcaligenes PsA15, P. denitrificans PsD6, P. polymyxa BcP26, and Mycobacterium phlei MbP18 strains led to a significant increase in the growth of root and aerial parts of plants, as well as a higher content of N, P, and K in these plants.
Several strains of B. subtilis and B. cereus have been isolated and selected in China, due to their ability to promote plant growth and control fungal diseases that affect wheat roots (Ryder et al. 1999). Among these strains is B. cereus A47, which has been used in several cultures in that country, stimulating wheat production by around 11%. In turn, B. subtilis B908 was used to control rice sheath blight, caused by Rhizoctonia solani. Both strains were also tested in two different soils in Australia, in experiments conducted in a greenhouse. These bacteria considerably reduced the severity of diseases caused by R. solani and Gaeumannomyces graminis var. tritici, stimulating also the growth of wheat seedlings (Ryder et al. 1999).
Mena-Violante and Olalde-Portugal (2007) investigated the effect of the inoculation of tomato roots with B. subtilis BS13 on the fruit’s quality and showed that production, weight, and length of tomatoes were higher in the inoculated group as compared to the negative control. The texture of fruits was also improved, demonstrating that BS13 strain plays a positive role in improving tomato quality, especially concerning size and texture.
Based on the presence of multiple PGP traits, Islam et al. (2009) isolated ten strains from paddy fields, screened for multiple PGP traits, and evaluated for their inoculation effects on canola and rice plants. Paenibacillus sp. RFNB4 significantly increased plant height and dry biomass production by 42.3 and 29.5%, respectively. Canola plants and rhizosphere soils inoculated with Bacillus sp. RFNB6 exhibited significantly higher nitrogenase activity.
B. megaterium B388, isolated from rhizosphere soil of pine belonging to a temperate Himalayan location, has been characterized by Trivedi and Pandey (2008). The carrier-based formulations of the bacterium resulted in increased plant growth in bioassays. The rhizosphere colonization and the viability of the cells entrapped in alginate beads were greater in comparison to coal- or broth-based formulations.
A greenhouse experiment was performed by Figueiredo et al. (2008) to evaluate the effects of PGPR on nodulation, biological nitrogen fixation (BNF), and growth of the common bean (Phaseolus vulgaris L. cv. tenderlake). Beans coinoculated with Rhizobium tropici (CIAT899) and P. polymyxa (DSM 36) had higher leghemoglobin concentrations, nitrogenase activity, and N2 fixation efficiency and thereby formed associations of greater symbiotic efficiency. Inoculation with Rhizobium and P. polymyxa strain Loutit (L) stimulated nodulation as well as nitrogen fixation. PGPR also stimulated specific-nodulation (number of nodules per gram of root dry weight) increases that translated into higher levels of accumulated nitrogen. The activities of phytohormones depended on their content and interactions with R. tropici and Paenibacillus and/or Bacillus (PGPR) strains, which affect the cytokinin content in the common bean.
Orhan et al. (2006) studied the growth promoting effects of two Bacillus strains (OSU-142, a nitrogen-fixing bacterium, and M3, which also fixes nitrogen and solubilizes phosphorus) used in consortium or individually in organic raspberry. The plants were evaluated for production, growth, nutrient content in leaves, and variation in nutrient composition. The results showed that Bacillus M3 stimulated plant growth and significantly increased yields. The inoculation of raspberry roots and rhizosphere with M3 and OSU-142 (in consortium or individually) caused an expressive improvement in yield (33.9 and 74.9%, respectively) as compared to the control. The levels of N, P, and Ca in leaves were also significantly increased. Similarly, inoculations affected the total levels of N, P, K, Ca, Mg, Fe, Mn, and Zn in soil. These results demonstrated that Bacillus M3, whether alone or in consortium with OSU-142, exhibited a high potential to increase yield, growth, and nutrition parameters in organic raspberry.
Karlidag et al. (2007) also tested the effects of Bacillus M3 and OSU-142 strains and that of Microbacterium FS01 isolated or in consortium on the growth of apple trees. The inoculation of these strains alone or in consortium increased plant growth (aerial part: 16.4–29.6%; diameter of aerial part: 15.9–18.4%) as well as yield (26–28%) and fruit size (13.9–25.5%). Of the nutrients investigated (N, P, K, Mg, Ca, Fe, Mn, and Zn), all were significantly affected (except Mg). The results of that study indicated that Bacillus M3, OSU-142, and Microbacterium FS0 strains are potential candidates to increase yield, growth, and nutrition of apple trees.
Bacillus OSU-142 was also investigated individually or in consortium with Pseudomonas BA-8 strain in yield, growth, and nutrient composition of cherry trees (Esitken et al. 2006). Both bacteria used individually as well as in consortium stimulated plant growth and led to a significant increase in yield. The use of BA-8, OSU-142, or a combination thereof on foliage and flowers led to a marked elevation in yield numbers (16.3, 10.9, and 21.76%), in fruit weight (4.14, 5.37, and 1.24%), and in the plants’ aerial parts (11.3, 11.8, and 29.6%), respectively, when compared to controls. Additionally, expressive increases were observed in N, P, and K contents in cherry leaves, while Fe and Zn levels increased with applications of BA-8 and OSU-142 strains in consortium. Mn contents were higher when these two bacterial strains were used separately. These results suggest that the inoculation of Pseudomonas BA-8 and Bacillus OSU-142 alone or in consortium may increase yield, growth, and nutrition levels of cherry trees.
1.5 Conclusions
The growing interest in PGPRs emerges from the wish to shed more light on a complex and fascinating biological system and from the potential these organisms have to increase productivity of crop cultures. The strains that exhibit a specific desired characteristic are selected, tested, and then used as an element in agricultural practices, though so far these strategies have not been extensively adopted – in spite of the several well-succeeded examples mentioned in this chapter. In a scenario where a greater number and more efficient bacterial strains are used, agricultural applications of PGPRs shall increase. Efforts made in the last decade have provided better insight into PGPRs and the mechanisms these bacteria utilize to promote plant growth, increasing the commercial interest in these strains (Glick 1995). While PGRPs have been characterized within several different bacterial taxa, many PGPRs developed for commercial applications are Bacillus species. These products utilize bacilli in their endospore form, which lends population stability during formulation and shelf life. Among bacilli, the strains of B. subtilis are the most widely used PGPRs, due to their capacity to produce antibiotics and consequent reduction in disease occurrence, when they are used as components of seed treatment strategies (Kokalis-Burelle et al. 2006).
References
Ahmad F, Ahmad I, Khan MS (2008) Screening of free-living rhizospheric bacteria for their multiple plant growth promoting activities. Microbiol Res 163:173–181
Alagawadi AR, Gaur AC (1988) Associative effect of Rhizobium and phosphate-solubilizing bacteria on the yield and nutrient uptake of chickpea. Plant Soil 105:241–246
Arshad M, Frankenberger WT Jr (1998) Plant growth-regulating substances in the rhizosphere: microbial production and functions. Adv Agron 62:45–151
Asghar HN, Zahir ZA, Arshad M, Khaliq A (2002) Relationship between in vitro production of auxins by rhizobacteria and their growth-promoting activities in Brassica jucea. Biol Fert Soils 35:231–237
Bachhawat AK, Ghosh S (1987) Iron transport in Azospirillum brasilense: role of the siderophores spirilobactin. J Gen Microbiol 133:1759–1765
Bai Y, Souleimanov A, Smith DL (2002a) An inducible activator produced by Serratia proteamaculans strain and its soybean growth promoting activity under greenhouse conditions. J Exp Bot 53:1495–1502
Bai Y, D’Aoust F, Smith DL, Driscoll BT (2002b) Isolation of plant growth-promoting Bacillus strains from soybean root nodules. Can J Microbiol 48:230–238
Bashan Y, Levanony H (1990) Current status of Azospirillum inoculation technology: Azospirillum as a challenge for agriculture. Can J Microbiol 36:591–608
Bastián F, Cohen A, Piccoli P, Luna V, Baraldi R, Bottini R (1998) Production of indole-3-acetic acid and gibberellins A1 and A3 by Acetobacter diazotrophicus and Herbaspirillum seropedicae in chemically defined culture media. Plant Growth Regul 24:7–11
Beneduzi A, Peres D, Vargas LK, Bodanese-Zanettini MH, Passaglia LMP (2008a) Evaluation of genetic diversity and plant growth promoting activities of nitrogen-fixing bacilli isolated from rice fields in South Brazil. Appl Soil Ecol 39:311–320
Beneduzi A, Peres D, Costa PB, Bodanese-Zanettini MH, Passaglia LMP (2008b) Genetic and phenotypic diversity of plant-growth-promoting bacilli isolated from wheat fields in southern Brazil. Res Microbiol 159:244–250
Beneduzi A, Costa PB, Parma M, Melo IS, Bodanese-Zanettini MH, Passaglia LMP (2010) Paenibacillus riograndensis sp. nov., a nitrogen-fixing species isolated from the rhizosphere of Triticum aestivum. Int J Syst Evol Microbiol 60:128–133
Berge O, Fages J, Mulard D, Balandreau J (1990) Effects of inoculation with Bacillus circulans and Azospirillum lipoferum on crop-yield in field grown maize. Symbiosis 9:259–266
Berge O, Guinebretiére M, Achouak W, Normand P, Heulin T (2002) Paenibacillus graminis sp. nov. and Paenibacillus odorifer sp. nov., isolated from plant roots, soil and food. Int J Syst Evol Microbiol 52:607–616
Borneman J, Skroch PW, O’Sullivan KM, Palus JA, Rumjanek NG, Jansen JL, Nienhuis J, Triplett EW (1996) Molecular microbial diversity of an agricultural soil in Wisconsin. Appl Environ Microbiol 62:1935–1943
Bric JM, Bostok RM, Silverstone S (1991) Rapid in situ assay for indoleacetic acid production by bacteria immobilized on a nitrocellulose membrane. Appl Environ Microbiol 57:535–538
Brown ME (1974) Seed and root bacterization. Annu Rev Phytopathol 12:181–197
Buysens S, Poppe J, Hofte M (1994) In: Ryder MH, Stephens PM, Bowen DG (eds) Improving plant productivity with rhizosphere bacteria. Commonwealth Science and Industrial Research Organization, Adelaide, pp 139–141
Cassán FD, Lucangeli CD, Bottini R, Piccoli PN (2001) Azospirillum spp. metabolize (17-172-H2) gibberellin A20 to (17-172-H2) gibberellin A1 in vivo in dy rice mutant seedlings. Plant Cell Physiol 42:763–767
Chabot R, Antoun H, Cescas MP (1993) Stimulation de la croissance du mais et de la laitue romaine par desmicroorganismes dissolvant le phosphore inorganique. Can J Microbiol 39:941–947
Chanway CP (1995) Differential response of western hemlock from low and high elevations to inoculation with plant growth-promoting Bacillus polymyxa. Soil Biol Biochem 27:767–775
Chanway CP, Holl FB (1991) Biomass increase and associative nitrogen fixation of mycorrhizal Pinus contorta Dougl. seedlings inoculated with a plant growth promoting Bacillus strain. Can J Bot 69:507–511
Chen YX, Mei RH, Lu S, Liu L, Kloepper JW (1996) In: Utkehede RS, Gupta VK (eds) Management of soil-borne disease. M/S Kalyani Publishers, New Delhi, pp 165–184
Choi JH, Im WT, Yoo JS, Lee SM, Moon DS, Kim HJ, Rhee SK, Roh DH (2008) P. donghaensis sp. nov., a xylan-degrading and nitrogen-fixing bacterium isolated from east sea sediment. J Microbiol Biotechnol 18:189–193
Christner BC, Mosley-Thompson E, Thompson LG, Zagorodnov V, Sandman K, Reeve JN (2000) Recovery and identification of viable bacteria immured in glacial ice. Icarus 144:479–485
Christner BC, Mosley-Thompson E, Thompson LG, Reeve JN (2001) Isolation of bacteria and 16S rDNAs from Lake Vostok accretion ice. Environ Microbiol 3:570–577
Chung YR, Kim CH, Hwang I, Chun J (2000) Paenibacillus koreensis sp. nov., a new species that produces an iturin-like antifungal compound. Int J Syst Evol Microbiol 50:1495–1500
Crowley DE, Reid CPP, Szaniszlo PJ (1988) Utilization of microbial siderophores in iron acquisition by oat. Plant Physiol 87:680–685
Crozier A, Arruda P, Jasmin JM, Monteiro AM, Sandberg G (1988) Analysis of indole-3-acetic acid and related indoles in culture medium from Azospirillum lipoferum and Azospirillum brasiliense. Appl Environ Microbiol 54:2833–2837
Daane LL, Harjono I, Barns SM, Launen LA, Palleroni NJ, Häggblom MM (2002) PAH-degradation by Paenibacillus spp. and description of Paenibacillus naphthalenovorans sp. nov., a naphthalene-degrading bacterium from the rhizosphere of salt marsh plants. Int J Syst Evol Microbiol 52:131–139
Datta M, Banish S, Dupta RK (1982) Studies on the efficacy of a phytohormone producing phosphate solubilizing Bacillus firmus in augmenting paddy yield in acid soils of Nagaland. Plant Soil 69:365–373
de Freitas JR, Banerjee MR, Germida JJ (1997) Phosphate-solubilizing rhizobacteria enhance the growth and yield but not phosphorus uptake of canola (Brassica napus L.). Biol Fert Soils 24:358–364
Demange P, Bateman A, Dell A, Abdallah M (1988) Structure of azotobactin D, a siderophore of Azotobacter vinelandii strain D (CCM 289). Biochemistry 27:2745–2752
Ding Y, Wang J, Liu Y, Chen S (2005) Isolation and identification of nitrogen fixing bacilli from plant rhizospheres in Beijing region. J Appl Microbiol 99:1271–1281
Dobbelaere S, Croonenborghs A, Thys A, Broek AV, Vanderleyden J (1999) Phytostimulatory effect of Azospirillum brasilense wild type and mutant strains altered in IAA production on wheat. Plant Soil 212:155–164
Dobbelaere S, Vanderleyden J, Okon Y (2003) Plant growth-promoting effects of diazotrophs in the rhizosphere. Crit Rev Plant Sci 22:107–149
Egamberdiyeva D, Höflich G (2004) Effect of plant growth-promoting bacteria on growth and nutrient uptake of cotton and pea in a semi-arid region of Uzbekistan. J Arid Environ 56:293–301
Elmerich C (1984) Molecular biology and ecology of diazotrophs associated with non-leguminous plants. Biotechnology 2:967–978
Elo S, Suominen I, Kämpfer P, Juhanoja J, Salkinoja-Salonen M, Haahtela K (2001) Paenibacillus borealis sp. nov., a nitrogen-fixing species isolated from spruce forest humus in Finland. Int J Syst Evol Microbiol 51:535–545
Esitken A, Pirlak L, Turan M, Sahin F (2006) Effects of floral and foliar application of plant growth promoting rhizobacteria (PGPR) on yield, growth and nutrition of sweet cherry. Sci Hortic 110:324–327
Felske A, Wolterink A, Van Lis R, de Vos WM, Akkermans ADL (1999) Searching for predominant soil bacteria: 16S rDNA cloning versus strain cultivation. FEMS Microbiol Ecol 30:137–145
Figueiredo MVB, Martinez CR, Burity HA, Chanway CP (2008) Plant growth-promoting rhizobacteria for improving nodulation and nitrogen fixation in the common bean (Phaseolus vulgaris L.). World J Microbiol Biotechnol 24:1187–1193
Fuentes-Ramirez LE, Jimenez-Salgado T, Abarca-Ocampo IR, Caballero-Mellado J (1993) Acetobacter diazotrophicus, an indoleacetic acid producing bacterium isolated from sugarcane cultivars of Mexico. Plant Soil 154:145–150
Garbeva P, van Veen JA, van Elsas JD (2003) Predominant Bacillus spp. in agricultural soil under different management regimes detected via PCR-DGGE. Microb Ecol 45:302–316
Garcia C, Fernadez T, Costa F, Cerranti B, Masciandaro G (1992) Kinetics of phosphatase activity in organic wastes. Soil Biol Biochem 25:361–365
Gardener BBM (2004) Ecology of Bacillus and Paenibacillus spp. in agricultural systems. Symposium: the nature and application of biocontrol microbes: Bacillus spp. Phytopathology 94:1252–1258
Gaur AC, Ostwal KP (1972) Influence of phosphate dissolving bacilli on yield and phosphate uptake of wheat crop. Indian J Exp Biol 10:393–394
Glick BR (1995) The enhancement of plant growth by free-living bacteria. Can J Microbiol 41:109–117
Glick BR, Patten CL, Holguin G, Penrose DM (1999) Biochemical and genetics mechanisms used by plant growth promoting bacteria. Imperial College Press, London
Grau FH, Wilson PW (1962) Physiology of nitrogen-fixation by Bacillus polymyxa. J Bacteriol 83:490–496
Gray EJ, Smith DL (2005) Intracellular and extracellular PGPR: commonalities and distinctions in the plant–bacterium signaling processes. Soil Biol Biochem 37:395–412
Guemouri-Athmani S, Berge O, Bourrain M, Mavingui P, Thiéry JM, Bhatnagar T, Heulin T (2000) Diversity of Paenibacillus polymyxa populations in the rhizosphere of wheat (Triticum durum) in Algerian soils. Eur J Soil Biol 36:149–159
Halverson LJ, Handelsman J (1991) Enhancement of soybean nodulation by Bacillus cereus UW85 in the field and in a growth chamber. Appl Environ Microbiol 57:2767–2770
Handelsman J, Raffel S, Mester EH, Wunderlich L, Crau CR (1990) Biological control of damping-off of alfalfa seedlings with Bacillus cereus UW85. Appl Environ Microbiol 56:713–718
Heyrman J, Swings J (2001) 16S rDNA sequence analysis of bacterial isolates from biodeteriorated mural paintings in the Servilia tomb (Necropolis of Carmona, Seville, Spain). Syst Appl Microbiol 24:417–422
Holford ICR (1997) Soil phosphorus, its measurements and its uptake by plants. Aust J Soil Res 35:227–239
Hong Y, Pasternak JJ, Glick BR (1991) Biological consequences of plasmid transformation of the plant growth promoting rhizobacterium Pseudomonas putida GR 12-2. Can J Microbiol 37:796–799
Hong YY, Ma YC, Zhou YG, Gao F, Liu HC, Chen SF (2009) Paenibacillus sonchi sp. nov., a nitrogen-fixing species isolated from the rhizosphere of Sonchus oleraceus. Int J Syst Evol Microbiol 59:2656–2661
Islam MR, Madhaiyan M, Boruah HPD, Yim W, Lee G, Saravanan VS, Fu Q, Hu H, Sa T (2009) Characterization of plant growth-promoting traits of free-living diazotrophic bacteria and their inoculation effects on growth and nitrogen uptake of crop plants. J Microbiol Biotechnol 9:1213–1222
Jain DK, Patriquin DG (1985) Characterization of a substance produced by Azospirillum wich causes branching of wheat root hairs. Can J Microbiol 31:206–210
Karlidag H, Esitken A, Turan M, Sahin F (2007) Effects of root inoculation of plant growth promoting rhizobacteria (PGPR) on yield, growth and nutrient elements contents of leaves of apple. Sci Hortic 114:16–20
Kloepper JW, Lifshitz R, Schroth MN (1988) Pseudomonas inoculants to benefit plant production. ISI Atlas Sci Anim Plant Sci 1:60–64
Kloepper JW, Lifshitz R, Zablotowicz RM (1989) Free-living bacterial inocula for enhancing crop productivity. Trends Biotechnol 7:39–43
Knosp O, von Tigerstrom M, Page JP (1984) Siderophore mediated uptake of iron in Azotobacter vinelandii. J Bacteriol 159:341–347
Koenig RL, Morris RO, Pollaco JC (2002) tRNA is the source of low-level trans-zeatin production in Methylobacterium spp. Appl Environ Microbiol 184:1832–1842
Kokalis-Burelle N, Kloepper JW, Reddy MS (2006) Plant growth-promoting rhizobacteria as transplant amendments and their effects on indigenous rhizosphere microorganisms. Appl Soil Ecol 31:91–100
Lacey LA, Frutos R, Kaya HK, Vail P (2001) Insect pathogens as biological control agents: do they have a future? Biol Control 21:230–248
Lambrecht M, Okon Y, Broek AV, Vanderleyden J (2000) Indole-3-acetic acid: a reciprocal signaling molecule in bacteria–plant interactions. Trends Microbiol 8:298–300
Lankford CE (1973) Bacterial assimilation of iron. Crit Rev Microbiol 2:273–331
Lebuhn M, Heulin T, Hartmann A (1997) Production of auxin and other indolic and phenolic compounds by Paenibacillus polymyxa strains isolated from different proximity to plant roots. FEMS Microbiol Ecol 22:325–334
Leong J (1986) Siderophores: their biochemistry and possible role in the biocontrol of plant pathogens. Annu Rev Phytopathol 24:187–209
Li D, Alexander M (1988) Co-inoculation with antibiotic-producing bacteria to increase colonization and nodulation by rhizobia. Plant Soil 108:211–219
Li CY, Massicote HB, Moore LVH (1992) Nitrogen-fixing Bacillus sp. associated with Douglas-fir tuberculate ectomycorrhizae. Plant Soil 140:35–40
Li JH, Em TW, Chen WF, Chen WX (2008) Genetic diversity and potential for promotion of plant growth detected in nodule endophytic bacteria of soybean grown in Heilongjiang province of China. Soil Biol Biochem 40:238–246
Lucangeli C, Bottini R (1997) Effects of Azospirillum spp. on endogenous gibberellin content and growth of maize (Zea mays L.) treated with uniconazole. Symbiosis 23:63–72
Lynch JM (1990) The rhizosphere. Wiley, Chichester
Ma Y, Chen S (2008) Paenibacillus forsythiae sp. nov., a nitrogen-fixing species isolated from rhizosphere soil of Forsythia mira. Int J Syst Evol Microbiol 58:319–323
Ma Y, Xia Z, Liu X, Chen S (2007a) Paenibacillus sabinae sp. nov., a nitrogen-fixing species isolated from the rhizosphere soils of shrubs. Int J Syst Evol Microbiol 57:6–11
Ma Y, Zhang J, Chen S (2007b) Paenibacillus zanthoxyli sp. nov., a novel nitrogen-fixing species isolated from the rhizosphere of Zanthoxylum simulans. Int J Syst Evol Microbiol 57:873–877
Manulis S, Haviv-Chesner A, Brandl MT, Lindow SE, Barash I (1998) Differential involvement of indole-3-acetic acid biosynthetic pathways in pathogenicity and epiphytic fitness of Erwinia herbicola pv. gypsophilae. Mol Plant Microbe Interact 11:634–642
Mavingui P, Heulin T (1994) In vitro chitinase antifungal activity of a soil, rhizosphere and rhizoplane populations of Bacillus polymyxa. Soil Biol Biochem 26:801–803
Mena-Violante HG, Olalde-Portugal V (2007) Alteration of tomato fruit quality by root inoculation with plant growth-promoting rhizobacteria (PGPR): Bacillus subtilis BEB-13bs. Sci Hortic 113:103–106
Montefusco A, Nakamura LK, Labeda DP (1993) Bacillus peoriae sp. nov. Int J Syst Bacteriol 43:388–390
Neilands JB, Konopka K, Schwyn B, Coy M, Francis RT, Paw BH, Bagg A (1987) In: Winkelmann G, van dre Helm D, Neilands JB (eds) Iron transport in microbes, plants and animals. Verlagsgesellschaft mbH, Weinheim, pp 3–33
Neves MCP, Rumjanek NG (1998) In: Melo IS, Azevedo JL (eds) Ecologia microbiana. EMBRAPA – CNPMA, Jaguariúna, pp 15–60
Nicholson WL (2002) Roles of Bacillus endospores in the environment. Cell Mol Life Sci 59:410–416
Okon Y, Labandera-González CA (1994) In: Ryder MH, Stephens PM, Bowen GD (eds) Improving plant productivity with rhizosphere bacteria. Commonwealth Scientific and Industrial Research Organization, Adelaide, pp 274–278
Orhan E, Esitken A, Ercisli S, Turan M, Sahin F (2006) Effects of plant growth promoting rhizobacteria (PGPR) on yield, growth and nutrients contents in organically growing raspberry. Sci Hortic 111:38–43
Park M, Kim C, Yang J, Lee H, Shin W, Kim S, Sa T (2005) Isolation and characterization of diazotrophic growth promoting bacteria from rhizosphere of agricultural crops of Korea. Microbiol Res 160:127–133
Patten CL, Glick BR (1996) Bacterial biosynthesis of indole-3-acetic acid. Can J Microbiol 42:207–220
Patten CL, Glick BR (2002) Role of Pseudomonas putida indoleacetic acid in development of the host plant root system. Appl Environ Microbiol 68:3795–3801
Paul EA, Clark FE (1989) Soil microbiology and biochemistry. Academic, San Diego, CA
Piao CG, Tang WH, Chen YX (1992) Study on the biological activity of yield-increasing bacteria. Chin J Microecol 4:55–62
Priest F (1993) Systematics and ecology of Bacillus. In: Losick R, Hoch JA, Sonenshein AL (eds) Bacillus subtilis and other gram-positive bacteria, biochemistry, physiology, and molecular genetics. American Society for Microbiology Press, Washington, DC, pp 3–16
Prinsen E, Costacurta A, Michiels K, Vanderleyden J, Van Onckelen H (1993) Azospirillum brasilense indole-3-acetic acid biosynthesis: evidence for a non-tryptophan dependent pathway. Mol Plant Microbe Interact 6:609–615
Raaska L, Sillanpää J, Sjöberg A, Suihko M (2002) Potencial microbiological hazards in the production of refined paper products for food applications. J Ind Microbiol Biotechnol 28:225–231
Ramos B, Lucas Garcia JA, Probanza A, Barrientos ML, Gutierrez Manero FJ (2003) Alterations in the rhizobacterial community associated with European alder growth when inoculated with PGPR strain Bacillus licheniformis. Environ Exp Bot 49:61–68
Raven PH, Evert RF, Eichhorn SE (1996) In: Raven PH, Evert RF, Eichhorn SE (eds) Regulacāo do crescimento e do desenvolvimento: os hormônios vegetais. Biologia vegetal. Guanabara Koogan SA, Rio de Janeiro, Brazil, pp 508–533
Rodríguez H, Fraga R (1999) Phosphate solubilizing bactéria and their role in plant growth promotion. Biotechnol Adv 17:319–339
Rodríguez-Diaz M, Lebbe L, Rodelas B, Heyrman J, de Vos P, Logan NA (2005) Paenibacillus wynnii sp. nov., a novel species harbouring the nifH gene, isolated from Alexander Island, Antarctica. Int J Syst Evol Microbiol 55:2093–2099
Rosado AS, Seldin L, Wolters AC, van Elsas JD (1996) Quantitative rDNA 16S-targeted polymerase chain reaction and oligonucleotide hybridization for the detection of Paenibacillus azotofixans in soil and the wheat rhizosphere. FEMS Microbiol Ecol 19:153–164
Roux V, Raoult D (2004) Paenibacillus massiliensis sp. nov., Paenibacillus sanguinis sp. nov. and Paenibacillus timonensis sp. nov, isolated from blood cultures. Int J Syst Evol Microbiol 54:1049–1054
Rózycki H, Dahm H, Strzelczyk E, Li CY (1999) Diazotrophic bacteria in root-free soil and in the root zone of pine (Pinus sylvestris L.) and oak (Quercus robur L.). Appl Environ Microbiol 12:239–250
Ryder MH, Yan Z, Terrace TE, Rovira AD, Tang W, Correll RL (1999) Use of strains of Bacillus isolated in China to suppress take-all and rhizoctonia root rot, and promote seedling growth of glasshouse grown wheat in Australian soils. Soil Biol Biochem 31:19–29
Sarwar M, Kremer RJ (1992) Determination of bacterially derived auxins using a microplate method. Lett Appl Microbiol 20:282–285
Saxena B, Modi M, Modi VV (1986) Isolation and characterization of siderophores from Azospirillum lipoferum D-2. J Gen Microbiol 132:2219–2224
Seldin L, Rosado A, Cruz DW, Nobrega A, van Elsas JD, Paiva E (1998) Comparison of Paenibacillus azotofixans strains isolated from rhizoplane, rhizosphere and non-root-associated soil from maize planted in two different Brazilian soils. Appl Environ Microbiol 64:3860–3868
Shah S, Karkhanis V, Desai A (1992) Isolation and characterization of siderophore, with antimicrobial activity, from Azospirillum lipoferum. Curr Microbiol 25:347–351
Shida O, Takagi H, Kodowaki K, Nakamura LK, Komagata K (1997) Transfer of Bacillus alginolyticus, Bacillus chondroitinus, Bacillus curdlanolyticus, Bacillus glucanolyticus, B. kobensis and Bacillus thiaminolyticus to the genus Paenibacillus and emended description of the genus Paenibacillus. Int J Syst Bacteriol 47:289–298
Siddiqui ZA, Mahmood I (1999) Role of bacteria in the management of plant parasitic nematodes: a review. Bioresour Tech 69:167–179
Siefert JL, Larios-Sanz M, Nakamura LK, Slepecky RA, Paul JH, Moore ER, Fox GE, Jurtshuk P (2000) Phylogeny of marine Bacillus isolates from the Gulf of Mexico. Curr Microbiol 41:84–88
Singh S, Kapoor KK (1998) Effects of inoculation of phosphate-solubilizing microorganisms and an arbuscular mycorrhizal fungus on mungbean grown under natural soil conditions. Mycorrhiza 7:249–253
Slepecky RA, Hemphill HE (1992) In: Balows A, Trüper HG, Dworkin M, Harder W, Schleifer KH (eds) The prokaryotes, a handbook on the biology of bacteria: ecophysiology, isolation, identification, applications, vol 2. Springer, New York, NY, pp 1663–1697
Smalla K, Wieland G, Buchner A, Zock A, Parzy J, Kaiser S, Roskot N, Heuer H, Berg G (2001) Bulk and rhizosphere soil bacterial communities studied by denaturing gradient gel electrophoresis: plant dependent enrichment and seasonal shifts revealed. Appl Environ Microbiol 67:4742–4751
Sorokin ID, Kravchenko IK, Tourova TP, Kolganova TV, Boulygina ES, Sorokin DY (2008a) Bacillus alkalidiazotrophicus sp. nov., a diazotrophic, low salt-tolerant alkaliphile isolated from Mongolian soda soil. Int J Syst Evol Microbiol 58:2459–2464
Sorokin ID, Zadorina EV, Kravchenko IK, Boulygina ES, Tourova TP, Sorokin DY (2008b) Natronobacillus azotifigens gen. nov., sp. nov., an anaerobic diazotrophic haloalkaliphile from soda-rich habitats. Extremophiles 12:819–827
Srinivasan M, Peterson DJ, Holl FB (1996) Influence of IAA producing Bacillus isolates on the nodulation of Phaseolus vulgaris by Rhizobium etli. Can J Microbiol 42:1006–1014
Stahly DP, Andrews RE, Yousten AA (1992) In: Balows A, Trüper HG, Dworkin M, Harder W, Schleifer KH (eds) The prokaryotes, a handbook on the biology of bacteria: ecophysiology, isolation, identification, applications. Springer, New York, NY, pp 1697–1745
Tang W, Pasternak JJ, Glick BR (1994) Stimulation of canola root growth by Pseudomonas putida GR 12-2 and its genetically engineered derivatives. Life Sci Adv 13:89–95
Tien TM, Gaskins MH, Hubbell DH (1979) Plant growth substances produced by Azospirillum brasilense and their effect on the growth of pearl millet (Pennisetum americanum L.). Appl Environ Microbiol 37:1016–1024
Timmusk S, Nicander B, Granhall U, Tillberg E (1999) Cytokinin production by Paenibacillus polymyxa. Soil Biol Biochem 31:1847–1852
Trivedi P, Pandey A (2008) Plant growth promotion abilities and formulation of Bacillus megaterium strain B 388 (MTCC6521) isolated from a temperate Himalayan location. Indian J Microbiol 48:342–347
Turner JT, Blackman PA (1991) Factors related to peanut yield increases following Bacillus subtilis seed treatment. Plant Dis 75:347–353
Vandendergh PA, Gonzalez CF (1984) Methods for protecting the growth of plants employing mutant siderophore producing strains of Pseudomonas putida. US Patent No. US4 479936
Verma SC, Ladha JK, Tripathi AK (2001) Evaluation of plant growth promoting and colonization ability of endophytic diazotrophs from deep water rice. J Biotechnol 91:127–141
von der Weid I, Duarte GF, van Elsas JD, Seldin L (2002) Paenibacillus brasilensis sp. nov., a new nitrogen-fixing species isolated from the maize rhizosphere in Brazil. Int J Syst Evol Microbiol 52:2147–2153
Wagner-Döbler I, Lünsdorf H, Lübbehüsen T, von Canstein HF, Li Y (2000) Structure and species composition of mercury-reducing biofilms. Appl Environ Microbiol 66:4559–4563
Wang Y, Brown HN, Crowley DE, Szaniszlo PJ (1993) Evidence for direct utilization of a siderophore, ferrioxamine B, in axenically grown cucumber. Plant Cell Environ 16:579–585
Xie GII, Su BL, Cui ZJ (1998) Isolation and identification of N2-fixing strains of Bacillus in rice rhizosphere of the Yangtze River valley. Acta Microbiol Sin 38:480–483
Yan Z, Reddy MS, Kloepper JW (2003) Survival and colonization of rhizobacteria in a tomato transplant system. Can J Microbiol 49:383–389
Yoon J, Seo W, Shin YK, Kho YH, Kang KH, Park Y (2002) Paenibacillus chinjuensis sp. nov., a novel exopolysaccharide-producing bacterium. Int J Syst Evol Microbiol 52:415–421
Zakharova EA, Shcherbakov AA, Brudnik VV, Skripko NG, Bulkhin NS, Ignatov VV (1999) Biosynthesis of indole-3-acetic acid in Azospirillum brasilense insights from quantum chemistry. Eur J Biochem 259:572–576
Zhang F, Smith DL (1996) Genistein accumulation in soybean (Glycine max L. Merr.) root systems under suboptimal root zone temperatures. J Exp Bot 47:785–792
Acknowledgments
The authors are grateful to Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq/Brazil).
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2011 Springer-Verlag Berlin Heidelberg
About this chapter
Cite this chapter
Beneduzi, A., Passaglia, L.M.P. (2011). Genetic and Phenotypic Diversity of Plant Growth Promoting Bacilli. In: Maheshwari, D. (eds) Bacteria in Agrobiology: Plant Growth Responses. Springer, Berlin, Heidelberg. https://doi.org/10.1007/978-3-642-20332-9_1
Download citation
DOI: https://doi.org/10.1007/978-3-642-20332-9_1
Published:
Publisher Name: Springer, Berlin, Heidelberg
Print ISBN: 978-3-642-20331-2
Online ISBN: 978-3-642-20332-9
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)