
Abstract
Monocytes are immune cells that can differentiate into a number of cell types including macrophages, dendritic cells, and osteoclasts upon exposure to various cytokines. The phenotypes of these differentiated cells are highly heterogeneous and their differentiation can be affected by the cyclic nucleotides, 3′-5′-cyclic adenosine monophosphate (cAMP) and 3′-5′-cyclic guanosine monophosphate (cGMP). The intracellular levels of cAMP and cGMP are controlled through regulation of production by adenylyl and guanylyl cyclases and through degradation by cyclic nucleotide phosphodiesterases (PDEs). PDE inhibition and subsequent changes in cyclic nucleotide levels can alter the final phenotype of a differentiating monocyte with regards to surface marker expression, gene expression, or changes in secreted chemokine and cytokine levels. The differentiation process itself can also be either inhibited or augmented by changes in cyclic nucleotide levels, depending on the system being studied and the timing of cyclic nucleotide elevation. This chapter explores the effects of PDE inhibition and increases in cGMP and cAMP on monocytic differentiation into osteoclasts, dendritic cells, and macrophages.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others
Keywords
1 Introduction
Monocytes are circulating peripheral immune cells that can differentiate into a number of cell types including macrophages, dendritic cells, and osteoclasts upon exposure to various cytokines (see Fig. 1). The phenotypes of these differentiated cells are highly heterogeneous and their differentiation can be affected by the cyclic nucleotides, 3′-5′-cyclic adenosine monophosphate (cAMP) and 3′-5′-cyclic guanosine monophosphate (cGMP). The intracellular levels of cAMP and cGMP are controlled through regulation of production by adenylyl and guanylyl cyclases and through degradation by cyclic nucleotide phosphodiesterases (PDEs).

Hematopoietic differentiation. Differentiation of multiple cell types from a common progenitor cell can be accomplished through their exposure to the appropriate cytokines at various stages of differentiation. Abbreviations: HSC Hematopoietic stem cell, CFU colony-forming unit, RANKL Receptor Activator for Nuclear Factor κB Ligand
Eleven different families of PDEs have been identified, and these enzymes differ in their cellular and tissue expression, substrate specificity, kinetics, modes of regulation, and sensitivity to inhibitors (Bender and Beavo 2006a). These PDEs have emerged as well-validated drug targets especially with the success of the PDE5 inhibitors Cialis, Viagra, and Levitra for the treatment of erectile dysfunction and more recently for pulmonary hypertension. In addition, several other classes of PDE inhibitors have been investigated as anti-inflammatory agents, anti-hypertensives, anti-depressants and as a therapy for muscular dystrophies. Of the most relevance to monocyte function and development are a number of clinical studies that have been conducted with PDE4 inhibitors as anti-inflammatory agents. While promising as in vitro agents and in preclinical studies, so far the PDE4 inhibitors have met with limited success as anti-inflammatory therapeutics in the clinic. Until recently, little has been known about how cyclic nucleotides and PDE inhibition can affect monocytic differentiation. Since drugs that selectively inhibit PDEs will increase cyclic nucleotide levels in targeted cells at all stages of differentiation, they are likely to affect the final phenotype of the differentiating cell as well as the differentiation process itself. PDE profiles can also change over the lifetime of the cell or during the progression of a disease state, thereby affecting its response to endogenous cyclic nucleotide-increasing agonists or to PDE inhibitor therapy (Bender et al. 2004; Essayan 2001; Gantner et al. 1997b). Indeed, in the presence of such agonists as histamine, adenosine, and prostaglandins, it is possible for macrophages to acquire a role as so-called regulatory macrophages (Mosser and Edwards 2008), while DCs are altered in their ability to activate the adaptive immune response (Morelli and Thomson 2003).
PDE profiling is a first step in analyzing the contribution of PDEs to cellular signaling pathways. In monocytes and monocyte-derived cells, PDE4 is generally considered a major PDE controlling many important inflammatory functions (Conti et al. 2003; Houslay et al. 2005; Spina 2008). However, it is important to note that PDE activity measurements can vary depending on substrate concentrations and methods used to analyze activity. PDEs also function in discrete compartments within the cell to limit the spread of the second messengers cAMP and cGMP (Bornfeldt 2006; Fischmeister et al. 2006; Houslay 2010). Therefore, a PDE whose total activity level is low could still contribute significantly to cellular function by controlling an important microdomain within the cell.
This chapter explores the role of altered cyclic nucleotide levels and inhibition of PDEs in monocytic differentiation and maturation into macrophages, dendritic cells, and osteoclasts, as well as the effect on the phenotype of the final differentiated cell.
2 Macrophage Differentiation
2.1 HL-60
Over 20 years ago, the HL-60 cell line was isolated from the peripheral blood of a patient with acute promyelocytic leukemia (Harris and Ralph 1985). HL-60 cells can be induced to differentiate to macrophage-like cells by a number of agents, including those that elicit a sustained elevation of intracellular cAMP or cGMP (Bang et al. 1994; Boss 1989; Nonaka et al. 1992).
Classic differentiation protocols for HL-60 cells to a myeloid cell phenotype involve treatment with either DMSO, phorbol-12-myristate-13-acetate (PMA), or all-trans-β-retinoic acid (RA), and this effect can be greatly potentiated by cAMP-increasing agents (Chaplinski and Niedel 1982). One group observed an effect of RA priming with pretreatment for cAMP-dependent induction of differentiation, which was not dependent on new protein synthesis (Olsson et al. 1982). Therefore, they postulated that RA priming was likely a more direct activation of existing signaling components in the cell by cAMP, presumably through PKA or Epac, or a decrease in production of an inhibitory polypeptide.
Along these lines, it was found that PMA-stimulated, cAMP-induced growth regulation is dependent on the regulatory PKA subunits PKA RIα and PKA RIIβ. Knockdown of the RIIβ subunit with an antisense oligodeoxynucleotide resulted in a decrease in cAMP-induced growth inhibition and differentiation, without affecting phorbol ester-induced differentiation (Tortora et al. 1990). Conversely, when the RIα subunit is knocked down, cell differentiation and growth arrest are increased, even in the absence of a cAMP-inducing stimulus (Tortora et al. 1991). Differentiation can also occur through stimulation of Gαs-coupled receptors and increases in cAMP alone, without concomitant phorbol ester administration. Given these data, it seems that phorbol ester and cAMP-induced pathways are complementary pathways for inducing differentiation that can act synergistically given the appropriate conditions.
The differentiation of monocytes can also be affected through stimulation of Gαs-coupled receptors, thereby raising intracellular cAMP. ATP can exert its effects on differentiation through stimulation of the Gαs-coupled P2Y11 receptor in monocytes (Boeynaems et al. 2000). The downstream effects of cAMP appeared to be mediated through PKAI, as the authors observed a decrease in differentiation using a PKA inhibitor, Rp-8-Br-cAMPS, which is more selective for PKAI than for PKAII (Jiang et al. 1997).
Cyclic nucleotide analogs are often used to dissect signaling pathways. These analogs have differing potencies and specificities, and some of the more common include hydrolysis-resistant analogs (8-Br-cAMP, 8-Br-cGMP), PKA-specific analogs (N6-Benzoyl cAMP) and Epac-specific analogs (8-CPT-2′-O-Me-cAMPS) (Poppe et al. 2008). These analogs can be used to stimulate downstream effectors directly. Some advantages of using analogs are that they are cell permeable, therefore easily administered to cells in culture, often nonhydrolyzable by PDEs, and the concentration can be carefully controlled. A major disadvantage is that most can stimulate all cyclic nucleotide downstream effectors and are not limited to endogenous signaling microdomains within the cell. Cho and colleagues differentiated HL-60 cells for 48 h in the presence of the cAMP analog 8-Br-cAMP and identified mature cells by their ability to produce superoxide in response to PMA (Cho et al. 2003). They concluded that cAMP induces ERK activation, and this activation is essential for differentiation. ERK activation appears to be PKA mediated as it was also not dependent on B-Raf, a downstream kinase of Rap1 and Epac. In fact, B-Raf activation was negatively regulated through PKA, and the differentiation could be inhibited by using an MEK inhibitor. While these results were inconclusive with respect to the entire cAMP-signaling differentiation pathway, it did establish ERK as an essential player in cAMP-mediated HL-60 cell differentiation.
Beavo and colleagues performed a set of studies where chronic elevation of cyclic nucleotides was mimicked by treating HL-60 cells with 8-Br-cAMP or 8-Br-cGMP during the 3-day differentiation process in the presence of PMA (Bender and Beavo 2006b). Morphological differences were readily observed after differentiation in the presence of the cAMP analog, but 8-Br-cGMP had a minimal effect on morphology. Cell spreading in culture was inhibited in the presence of elevated cyclic nucleotides and the cells form large clumps of round cells. These differences were not attributed to any changes in macrophage function, but one could imagine that differences in “cell spreading” could be the result of differences in migration or secretion of chemotactic molecules or cytokines. Surface molecule expression was also slightly changed with treatment but differentiation was not halted. 8-Br-cAMP significantly decreased the upregulation of CD11b, a monocyte/macrophage marker, while it increased expression of the urokinase plasminogen activator (CD87); however, 8-Br-cGMP had no effect. These observed effects were mimicked by the PKA-specific activator N6-Benzoyl-cAMP, and therefore likely mediated through PKA.
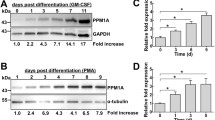
HL-60 cells have also been used as a model system emulating human granulocyte macrophage colony-stimulating factor (GM-CSF) differentiated monocytes to determine the role of PDE1B in these cells. One variant of this isoform, the cGMP-preferring enzyme PDE1B2 (Fidock et al. 2002), is specifically upregulated upon differentiation in both GM-CSF differentiated human monocytes and PMA differentiated HL-60 cells (Bender et al. 2005). PDE1B2 activity is regulated by intracellular Ca2+ and may provide a link between Ca2+ and cAMP signaling pathways. To determine the role of PDE1B2, stable knockdown of this isoform by short hairpin RNAs was performed in differentiated HL-60 cells (Bender and Beavo 2006b). This specific isoform proved to be the major regulator of atrial natriuretic factor-stimulated cGMP levels in HL-60 cells, and the authors subsequently searched for a functional role for this newly uncovered variant. It was found that suppression of PDE1B2 expression alters some aspects of the macrophage-like phenotype, because cell spreading, phagocytic ability, and CD11b expression were augmented. Unexpectedly, the cAMP analog 8-Br-cAMP, but not 8-Br-cGMP, reversed the changes caused by PDE1B2 knockdown, indicating a role for cAMP in control of these PDE1B2-dependent functions. Moreover, despite the preference of PDE1B for cGMP as substrate, knockdown of PDE1B2 caused a decrease in basal cAMP levels, but showed no change in cGMP, and a number of PKA phosphorylation consensus sites were altered. The cause of the decrease in basal cAMP levels remains unclear. The expected increase in cGMP with PDE1B2 knockdown may be contained within localized signaling microdomains, and therefore undetectable by whole cell cGMP measurements. More likely, there is some level of crosstalk between the cGMP and cAMP signaling pathways, possibly through inhibition of an adenylyl cyclase, or activation of another cAMP-preferring PDE.
2.2 U937
The U937 cell line is a human line established from a diffuse histiocytic lymphoma and displays many monocytic characteristics (Harris and Ralph 1985). The cGMP-PDE profile of this cell line is very similar to that of a macrophage differentiated from a human CD14+ monocyte in the presence of monocyte colony-stimulating factor (M-CSF) in that it exhibits high PDE2 activity and some PDE1 activity (Bender et al. 2004). Changes in PDE4 activity have been observed in conjunction with changes in cAMP levels. Characterization of PDE4 activity in monocytic U937 cells showed that PDE4 activity can be augmented with a transient increase in cAMP of 2–4 h in length. Also, the V max of PDE4 was increased two-to threefold in the presence of salbutamol, a β2 adrenergic agonist and the PDE4 inhibitor, rolipram, but only if cAMP levels were elevated for more than 2 h. It was determined that PDE4 activity was increased through a PKA- and protein synthesis-dependent mechanism (Torphy et al. 1992). These data imply that cAMP signaling feeds back to activate PDE4-mediated degradation of cAMP, thereby limiting the duration and magnitude of signaling through that microdomain.
Houslay and colleagues examined cAMP-PDE activity levels and found that PDE4 comprised the majority of PDE activity in U937 monocytic cells. However, upon differentiation with PMA, PDE3 activity predominated and PDE4 activity was decreased. Using antisera specific for PDE4 isoforms and RT-PCR, the investigators determined the splice variants of PDE4 that were changed during U937 differentiation to a macrophage-like cell. The N-terminal end of PDE4 isoforms contain unique isoform-specific regions that can vary in length with long, short, and supershort isoforms commonly observed. These isoforms contain variable truncations of the upstream conserved regions, or UCR domains. The long forms generally contain both UCR1 and UCR2 domains, while the short forms express only UCR2 and supershort forms merely a portion of the UCR2 domain (Houslay et al. 2005). A key distinguishing factor for these various isoforms is their susceptibility to regulation by phosphorylation in their N-terminal UCR domains. Upon differentiation to a macrophage, protein levels of the long isoform PDE4A10 and the short isoform PDE4B2 were markedly increased, whereas long-form PDE4A4 remains unchanged, and short forms PDE4D3 and PDE4D5 were downregulated (Shepherd et al. 2004). Additionally, it was found that phospho-ERK was increased with acute PMA challenge, and this challenge gave a time-dependent inhibition of PDE4 activity in U937 monocytes and an increase in PDE4 activity in U937 macrophages, both of which are dependent on ERK activation. The differential effects of ERK activation can be directly attributed to the remodeling of the pattern of PDE4 isoform expression. The predominant PDE4 activity in monocytes is contributed by long PDE4D isoforms, whereas in macrophages PDE4B short form activity is the major source of PDE4 activity. This remodeling has functional consequences since PDE4D long isoforms will become inhibited, PDE4B short forms will be activated upon phosphorylation by ERK, and PDE4A isoforms are unaffected as they do not contain ERK phosphorylation sites (Baillie et al. 2000). This presents an attractive therapeutic possibility if PDE4 isoform-selective inhibitors can be developed, allowing the differential regulation of monocytes and macrophages.
U-937 differentiation can be induced by increasing cAMP levels, either using PGE2, the adenylyl cyclase agonist forskolin, or a nonhydrolyzable cAMP analog, db-cAMP (Brodsky et al. 1998; Shayo et al. 1997). Surprisingly, activation of the Gαs-coupled histamine receptor is not able to induce U937 differentiation, despite its ability to activate adenylyl cyclase and cause a transient increase in intracellular cAMP (Shayo et al. 1997). Upon further investigation, it was discovered that the histamine receptor rapidly becomes desensitized through a GRK2-dependent mechanism and that a sustained elevation of cAMP is required for induction of differentiation (Fernandez et al. 2002; Legnazzi et al. 2000; Shayo et al. 2001). The stable overexpression of the histamine H2 receptor (H2R) alone also induced differentiation (Monczor et al. 2006). The increased expression of H2R leads to an increase in basal cAMP levels and a leftward shift of the dose–response curve. There was a maximal response to agonist similar to those cells expressing normal amounts of H2R, indicating that overexpression of H2Rs results in increased numbers of functional spare receptors in this system. In addition to the generation of spare receptors, there was a compensatory increase in GRK2 expression and PDE activity stimulation. Despite the onset of these regulatory mechanisms, an H2 agonist and the PDE4 inhibitor, rolipram, were still able to induce differentiation in these cells, contrary to the previous observations in naïve cells (Monczor et al. 2006).
The time course of cAMP elevation was determined to be critical for the induction of U937 differentiation. Using the cAMP-elevating agonists amthamine (a histamine H2 receptor agonist), PGE2 (an EP receptor agonist), and forskolin (an adenylyl cyclase agonist), Shayo and colleagues were able to demonstrate a time dependence to cAMP-induced differentiation in U937 cells (Shayo et al. 2004). Amthamine caused a transient increase in cAMP returning to baseline in about 3 h, whereas forskolin and PGE2 created a more prolonged elevation of cAMP, which was still sustained at the final measurement of 4 h. This prolonged cAMP elevation was necessary for differentiation of U937 cells, as indicated through the observed change in differentiation markers such as an increase in C5aR expression, arrested proliferation, and a decrease in c-myc protein levels. The mechanism of this discrepancy seems to be H2R dependent occurring through GRK2-mediated desensitization of the receptor. PGE2 and forskolin bypass H2R and therefore are able to sustain cAMP elevation. Homologous desensitization of the EP receptor by PGE2 was evident immediately upon stimulation and persisted for 40 min. However, a sustained cAMP elevation via either PGE2 or forskolin can also induce heterologous desensitization of the H2 receptor in a PKA/PKC-dependent manner. The data also showed a transient upregulation of c-Fos with amthamine and sustained c-Fos expression with PGE2 and forskolin. This correlated well with the fact that sustained c-fos expression is known to be essential for U937 differentiation. Overall, these studies indicate that cAMP levels must be sustained long term to induce U937 differentiation.
2.3 Human AB Serum and M-CSF
2.3.1 AB Serum
Human peripheral blood monocytes can be differentiated to macrophages in culture in human AB serum, and enriched through their adherence to tissue culture plates. Initially, the monocytes contain mostly PDE4 activity, with some PDE3 activity. As the cells differentiate in culture, the predominate PDEs emerge as PDE1 and PDE3, with rapidly declining amounts of PDE4 activity. PDE2 and PDE5 levels remain consistently low throughout the differentiation process (Gantner et al. 1997a). In another study with human monocytes, the majority of the PDE4 activity was found in the soluble fraction, and the PDE3 activity was associated with the particulate fraction similar to its location in many other tissues (Verghese et al. 1995). AB serum-differentiated monocytes exhibit a PDE profile strongly resembling that observed for human alveolar macrophages. A comparison of TNF-α production elicited by lipopolysaccharide (LPS) between monocytes and monocyte-derived macrophages showed inhibition of TNF-α production with increases in cAMP brought on by PDE4 inhibition in both cell types (Gantner et al. 1997a). PDE4 controlled the largest portion of TNF-α production in monocytes, as about 80% could be blocked with a PDE4 inhibitor, with PDE3 inhibition decreasing TNF-α production approximately 10%. In macrophages, PDE3 and PDE4 controlled more equal amounts, but PDE inhibition was only effective when administered in the presence of an additional cAMP stimulus, perhaps indicating a lower cAMP tone in macrophages than monocytes.
Human monocytes cultured in human AB serum also have a very similar PDE profile to that present in the cell line Mono-Mac-6. Using this cell line as a surrogate, a closer look at the changes in the PDE4 isoform levels during challenge with cAMP-increasing agents showed that db-cAMP, PGE1, or LPS transiently increased PDE4A, B, and D mRNA and protein levels (Verghese et al. 1995). However, the time course and magnitude of induction of each isoform was not identical. PDE4B levels rose early, mRNA increased from 2 to 4 h, protein increased from 2 to 8 h; with PDE4A and PDE4D rising later in the time course, mRNA increased from 2 to 8 h and protein increased after 5–24 h of treatment. This increase was specific for PDE4 isoforms as there was no concomitant increase in PDE3 activity with differentiation. An additional level of regulation was observed for the PDE4 isoforms as differences in the induction of mRNA when compared to protein levels were apparent over the time courses tested, suggesting that the PDE4 isoforms and splice variants were transcriptionally regulated.
2.3.2 M-CSF + AB Serum
While PDE activities, and therefore the duration of cAMP and cGMP signals, can change during differentiation, the levels of their downstream signaling effector proteins can also be altered. The immediate downstream mediators for cAMP are PKA, Epac, and nucleotide-gated ion channels. The levels of these mediators can also change over the course of monocyte to macrophage differentiation and maturation. Taskén’s group identified a number of signaling molecules downstream of cAMP in monocytes and macrophages. Monocytes contained the regulatory PKA subunits RI-α, RII-α and the catalytic C subunit, with high levels of RI-α and lower levels of RII-α (Bryn et al. 2006). Monocytes differentiated in the presence of human AB serum and M-CSF produce macrophages that exhibit a threefold increase in Epac1 mRNA levels. Presumably, this would translate into an increased role for Epac and its downstream signaling through Rap1 and the MAP kinase pathways in macrophage function. Strangely, these investigators found that Epac activation had a minimal effect on mature macrophage functions for most conditions tested, such as inhibition of TNF-α or IL-12 secretion. The majority of macrophage functions remained PKA-dependent and therefore unaffected by treatment with the Epac-specific activator, 8-CPT-2′-O-Me-cAMP. In one functional assay for phagocytic capability, however, FcR-mediated phagocytosis was reduced equally by both PKA and Epac-specific activators in mature macrophages, while in undifferentiated monocytes, there was no effect of the Epac-specific cAMP analog alone. Although this study was not exhaustive, it definitively showed a minimal involvement of Epac in the regulation of monocyte/macrophage inflammatory function and a major role for PKA.
2.3.3 M-CSF
When monocytes are instead differentiated using M-CSF and fetal calf serum, a distinct set of characteristics are acquired (Geissmann et al. 2010). These cells have moderate levels of a number of PDEs involved in degradation of both cGMP and cAMP and changes in guanylyl cyclase activity. M-CSF-derived macrophages retain modest PDE4, PDE3, and PDE1 levels. For the cGMP PDEs, PDE2 activity increases slightly with M-CSF differentiation, and PDE5 activity decreases to undetectable levels (Bender and Beavo 2004; Bender et al. 2004). In addition to the regulation of the cyclic nucleotide-degrading PDEs, these investigators also observed changes in the levels of the cGMP-producing guanylyl cyclases, likely affecting the availability of cGMP in the cells. The levels of GC-A were undetectable in monocytes, with slightly increased levels in M-CSF macrophages, while soluble GC mRNA levels were dramatically downregulated upon differentiation to a macrophage.
The physiological implications of these changes are not immediately clear. Expression of particular PDEs or cyclases is often linked with specific signal transduction pathways through localization with a specific receptor, anchoring protein or kinase. For example, if PDE5 is tightly linked with other members of the cGMP pathway, the loss of PDE5 with M-CSF differentiation may be coupled to a loss in sGC and a decrease in responsiveness to NO (Bender et al. 2004). Similar correlations could also occur with cAMP signaling pathways, leading to changes in cAMP-PDEs, PKA, Epac, adenylyl cyclases, or any number of downstream signaling components.
2.4 GM-CSF
Monocyte to macrophage differentiation can also occur when monocytes are exposed to the cytokine GM-CSF (Geissmann et al. 2010). These macrophages have a phenotype similar to that observed for alveolar macrophages. The cGMP-PDE profile in CD14+ monocytes isolated from human blood consists of mostly PDE5 activity, with some PDE1 and PDE2 activity and low PDE3 and modest PDE4. Upon differentiation for 7 days in GM-CSF, PDE1 emerges as the predominant cGMP-PDE present in these macrophages, consisting mostly of the PDE1B isoform and the cAMP-PDE activity was attributed equally to PDE3 and PDE4 (Bender and Beavo 2004; Bender et al. 2004; Hertz et al. 2009a). Also characterized in these studies was the cGMP-PDE composition of the monocytic cell line THP-1 which was found to be similar to GM-CSF-derived macrophages upon differentiation with PMA. THP-1 is a human acute monocytic leukemia cell line with high levels of PDE1B, but very low levels of PDE1A and PDE1C and very little PDE2 activity. These cells can be differentiated to a macrophage-like cell upon stimulation with PMA. While these THP-1-derived macrophages contained high levels of PDE1B, the levels of all PDEs were consistently high in the undifferentiated monocyte and throughout differentiation, in contrast to CD14+ monocyte-derived macrophages (Bender et al. 2004).
Upon further investigation, it was discovered that a unique splice variant, PDE1B2 (Fidock et al. 2002), is the form upregulated upon monocyte to macrophage differentiation in the presence of GM-CSF (Bender et al. 2005). Interestingly, this isoform of PDE1B has a threefold lower EC50 for calmodulin than its counterpart PDE1B1. This calmodulin-sensitive PDE has a separate transcriptional start site that can be activated by monocytic differentiation to a macrophage, but is suppressed upon the addition of IL-4 and subsequent differentiation of the monocyte to a dendritic cell. The authors postulate that selective upregulation of a calcium/calmodulin-sensitive cGMP-preferring PDE could provide a mechanism by which calcium transients can control the duration and amplitude of the cGMP signal.
The cAMP-PDE activities in GM-CSF-differentiated monocytes were determined using a PDE activity assay, with PDE4 showing the largest amount of activity in monocytes, with decreasing amounts of activity during differentiation. However, in fully differentiated macrophages PDE3 and PDE4 comprised equivalent amounts of cAMP PDE activity (Gantner et al. 1997a, b, 1999; Hertz et al. 2009a). It should also be noted that PDE1B is a dual specificity PDE, able to hydrolyze both cAMP and cGMP. Although it prefers to hydrolyze cGMP over cAMP, a moderate amount of cAMP-PDE activity is contributed by PDE1B in these cells.
The activity and type of cyclase present is another factor that can affect cyclic nucleotide levels in any cell. CD14+ monocytes were found to have high levels of soluble guanylyl cyclase, which was decreased slightly upon differentiation to a macrophage with GM-CSF (Bender et al. 2004). The levels of GC-A were undetectable in monocytes, with greatly increased levels in macrophages. These changes could affect the cells’ ability to respond to a number of cGMP-increasing factors including nitric oxide and atrial natriuretic peptide (ANP).
More recently, the effects of chronic elevated cyclic nucleotide levels on the function and final phenotype of monocytes differentiating to alveolar-type macrophages were investigated (Hertz et al. 2009b). Human CD14+ monocytes were differentiated in the presence of GM-CSF for 6 days in combination with cAMP-increasing agents, such as the adenylyl cyclase agonist forskolin, the EP receptor agonist PGE2, and selective PDE inhibitors to assess the effects of elevated cAMP levels on the final phenotype of the macrophage. Initially, monocytes were treated with a high dose of forskolin in culture for 6 days. A microarray analysis was performed on RNA extracted from the resulting macrophages and, unexpectedly, a number of proinflammatory genes were upregulated, especially chemokines binding to the CXCR2 and CCR2 classes of receptors. Expression of the cytokines IL-10 and IL-6, as well as a number of genes involved in wound-healing, were also increased upon differentiation in the presence of cAMP (see Fig. 2a). A phenotypic analysis of these forskolin-treated cells showed that differentiation was not halted as was seen previously with GM-CSF + IL-4-induced dendritic cell differentiation (Giordano et al. 2003), but the expression of a number of surface markers was altered. Notably, the downregulation of CD14, the coreceptor for LPS, and CD163, a scavenger receptor, was prevented with forskolin treatment.

Effects of cAMP on monocytic differentiation to macrophages and dendritic cells. Monocytes isolated from human blood can be differentiated in the presence of cytokines GM-CSF or GM-CSF plus IL-4 to give rise to macrophages and dendritic cells, respectively. When differentiated in the presence of elevated cyclic nucleotides, the final phenotypes of these cells are altered. Differences in cytokine and chemokine production are observed as well as changes in surface marker expression levels
To determine which PDEs were controlling the pools of cAMP regulating surface marker and chemokine expression, selective inhibitors of PDE2, 3, and 4 were used in combination with a low dose of forskolin. The low dose of forskolin showed a minimal change in mRNA and protein levels of these genes (Hertz et al. 2009a). However, when that same dose of forskolin was administered with a PDE inhibitor it caused a large response. This is exactly what would be predicted for a synergistic effect of PDE inhibition coupled with adenylyl cyclase activation. Using this technique, it was determined that PDE4 primarily controls the signaling microdomain regulating expression of surface markers and chemokines.
The intracellular signaling pathways involved in regulation of these chemokines seem to be largely downstream of Epac, not PKA as seen for cAMP's effect on most of the proinflammatory cytokines (Hertz et al. 2009a). Cyclic nucleotide analogs which specifically activate Epac induce chemokine expression at both the mRNA and the protein levels, while a PKA-specific activator had a minimal effect on chemokine levels. Downstream signaling pathways leading to changes in macrophage inflammatory gene expression are generally regulated through the canonical NF-κB pathway. Chemokine production was attenuated with a general inhibitor of the NF-κB signaling pathway, the IKK1/2 inhibitor BMS-345541, but surprisingly remained elevated after treatment with a peptide inhibitor of the canonical p50/p65 NF-κB heterodimer. While inflammatory cytokine production is normally regulated through the p50/65 NF-κB pathway in macrophages, it seems that production of these chemokines via elevated cAMP may occur through the noncanonical p50/p105 NF-κB pathway. Also affecting this pathway was a negative regulator of NF-κB, the transcription factor ATF3, which exhibits both reduced mRNA levels and attenuated binding to the CXCL7 promoter when forskolin is present during differentiation (Hertz et al. 2009a). The relief of inhibition of ATF3 on promoter regions containing NF-κB would promote expression of many inflammatory genes (Gilchrist et al. 2006).
The signaling pathways downstream of cAMP can also be changed during differentiation. Human monocytes stimulated with LPS produce large amounts of TNF-α, IL-6, and IL-8. The release of these chemokines can be reduced with a p38 kinase inhibitor. However, in macrophages differentiated in GM-CSF for 12 days, p38 inhibition has a lesser effect on production of these cytokines while TNF-α production is effectively blocked with an ERK inhibitor (Tudhope et al. 2008). This suggests that kinase usage can also change as a monocyte differentiates to a macrophage, with monocytes favoring p38 kinase and macrophages preferring ERK to control cytokine production.
3 Dendritic Cell Differentiation
Monocytes differentiated in the presence of GM-CSF + IL-4 gain a dendritic cell phenotype (Geissmann et al. 2010). The PDE profile of these dendritic cells (DCs) showed lower PDE4 levels than monocytes, increased PDE3, and a moderate upregulation of PDE1B (Gantner et al. 1999; Giordano et al. 2003). Regarding PDE4 specifically, PDE4B was found to be expressed at high levels in monocytes, with PDE4A being the predominant PDE4 isoform in DCs (Heystek et al. 2003).
Elevation of cyclic nucleotides through administration of PGE2 has been shown to enhance the differentiation and maturation of DCs under serum-free cell culture conditions. In lieu of fetal calf serum, a monocyte conditioned medium was used to culture the monocytes in the presence of GM-CSF and IL-4 and the addition of PGE2 greatly enhanced their differentiation (Jonuleit et al. 1997). It was hypothesized that preexposure of monocytes and DCs to cAMP-elevating agents, such as PGE2, in tissues could have functional consequences for the final phenotype of the cell and the subsequent priming of naïve T cells in lymph nodes.
In other studies to test the effects of elevated cAMP on DC function, monocytes were isolated from human peripheral blood and differentiated for 6 days in the presence of fetal calf serum, GM-CSF and IL-4 in the presence or absence of PGE2 (Kalinski et al. 1997) or with the addition of the adenylyl cyclase activator forskolin, cyclic nucleotide analogs (8-Br-cGMP and 8-Br-cAMP) or the nonselective PDE inhibitor, isobutylmethylxanthine (IBMX) (Giordano et al. 2003). The phenotype of this cAMP-generated DC was characterized by analyzing its surface marker and cytokine expression profile to determine whether the cell was directed to a DC, or another cell type. (see Fig. 2b) Normally during monocytic differentiation to a DC, the surface markers CD14 and CD1a are reciprocally regulated with the cells showing a downregulation of the monocytic marker CD14 and an upregulation of DC marker CD1a. The reciprocal regulation of these surface markers was prevented with increased cyclic nucleotides (Giordano et al. 2003). However, cyclic nucleotides do not seem to affect the regulation of other makers such as DC-SIGN, a DC-restricted C-type lectin involved in the early interaction between DCs and naive T cells and also DC trafficking and internalization of Ags, or the downregulation of the high affinity IgG-binding receptor, CD64/FcγRI. Terminal differentiation of myeloid cells is generally characterized by a switch from CD64/FcγRI to the low affinity CD32/FcγRII. Interestingly, CD32/FcγRII expression was not blocked by elevated cyclic nucleotides, but was slightly enhanced and the final phenotype was more reminiscent of the levels seen on macrophages (Giordano et al. 2003). The regulation of the costimulatory molecules CD80 and CD86 was unaffected by PGE2 (Kalinski et al. 1997) but CD86 and MHC class II were upregulated with cAMP analog treatment over the 6-day differentiation process and the reason for this is unclear (Giordano et al. 2003). The authors concluded that DC differentiation was halted at an intermediate cell stage between monocytes, macrophages, and DCs.
This intermediate population of cells was not homogeneous; however, since differentiation in the presence of elevated cyclic nucleotides also upregulated a subset of cells expressing CD16 (Giordano et al. 2003), a surface marker upregulated in some pathological conditions such as HIV and autoimmune diseases (Grage-Griebenow et al. 2001). CD16 is expressed on less than 10% of circulating monocytes and is generally expressed on the cells that are CD14lo. After cyclic nucleotide treatment in vitro, the percentage of CD16+ cells increased dramatically, up to as much as 50% of the total cell population (Giordano et al. 2003). The authors speculate that cyclic nucleotide signaling may be a part of the intracellular mechanisms used to modulate expansion of this subpopulation in pathological conditions.
DCs generated in the presence of physiological concentrations of PGE2, and therefore elevated cAMP, demonstrated Th-cell stimulation abilities comparable to the control DCs differentiated in the absence of PGE2. However, the ability of PGE2-DCs to produce IL-12 was markedly reduced when stimulated with IL-12-inducing agents such as soluble CD40L, and even in the presence of the normally potent IL-12 inducing agent, IFN-γ (Kalinski et al. 1997). When the concentration of PGE2 was increased to 100 nM, the cells’ IL-12 producing ability was completely abolished. Conversely, production of the anti-inflammatory cytokine IL-10 was markedly increased with PGE2 pretreatment, but decreased with IFN-γ cotreatment. This profile was maintained over 48 h after the removal of PGE2, and the ability of these DCs to produce other Th cell-polarizing cytokines was also altered. PGE2-DCs retained higher IL-4 and IL-5 producing capacity and produced 70% less IFN-γ than control DCs. The ability of PGE2-DCs to promote Th2-type differentiation of naïve T cells was largely due to the absence of IL-12, which would normally drive the cell to a Th1 phenotype, not due to the presence of an additional secreted factor. Similar results were observed by another group when a PDE4 inhibitor was administered during differentiation of monocytes to dendritic cells, prior to final maturation (Heystek et al. 2003).
In a pathological situation, cyclic nucleotide levels will be elevated not only during differentiation of monocytes, but also during maturation into the final macrophage or dendritic cell. The effects of PGE2 on DC maturation were found to act as a cofactor and synergize with TNF-α and IL-1β to induce final maturation of immature DCs (Kalinski et al. 1998). The adenylyl cyclase agonist, forskolin, mimicked the effects of PGE2, indicating that final maturation can be influenced directly by cAMP levels. After final DC maturation, PGE2 and IL-10 were no longer able to affect the phenotype of the DC, indicating that shaping of the mature DC phenotype occurs while the immature DC is a tissue resident and capable of taking up antigen. Therefore, the levels of IL-12 production and mature DC phenotype are predetermined at the stage of immature DC in peripheral tissues. The stability of the phenotype for up to 2 days after exposure to PGE2 implies that the DC phenotype will be retained upon its arrival in the draining lymph node, allowing it to contribute to the development of Th2-biased responses without picking up irrelevant signals as it migrates to the lymph node.
This effect on maturation is not isolated only to cAMP, similar effects have been observed with elevated intracellular levels of cGMP. In one set of studies, mature DCs were derived from CD14+ human monocytes and matured/activated with LPS (Morita et al. 2003). When these cells were pretreated with the guanylyl cyclase agonist ANP, the LPS-induced expression of IL-12 and TNF-α was decreased and IL-10 expression was augmented similar to what was observed for cAMP (Morita et al. 2003). This phenotype is also able to polarize CD4+ T cells toward promoting Th2 responses through interactions in the absence of IL-12 production.
DCs can mature upon exposure to LPS and acquire a distinct set of surface markers that are regulated by cyclic nucleotides. Cyclic nucleotide treatment inhibited the increase in DC-LAMP and CD83 normally brought on with LPS stimulation. DC-LAMP is a lysosome-associated membrane glycoprotein that belongs to DC-specific apparatus for antigen processing. Different antigen-presenting cells have distinct endocytic routes used for processing antigen, with specific markers characterizing each step. CD68 or macrosialin is another lysosomal marker expressed on both macrophages and DCs and is inversely regulated from DC-LAMP; as DC-LAMP appears in these compartments, CD68 levels are reduced. Curiously, while DC-LAMP expression was reduced in the presence of cyclic nucleotides, CD68 remained unaffected. Therefore, it seems that an increase in cyclic nucleotide levels during differentiation could impair the ability of DCs to process antigen, but does not inhibit antigen-processing machinery shared with other cells of the myeloid lineage.
DCs differentiated in the absence of PGE2, but matured in its presence, also acquire an increased ability to migrate toward the lymph nodes via CCR7 and exhibit improved ability to activate CD4 T cells suggesting the involvement of PGE2 in facilitating CCR7 signaling (Luft et al. 2002; Scandella et al. 2002). However, when DCs are differentiated in the presence of cyclic nucleotide analogs, the induction of CCR7 by LPS is reduced (Giordano et al. 2003). CCR7 confers the ability onto dendritic cells to migrate toward CCL19, which is a vital step in leading DCs to lymph nodes and downstream T-cell activation. Despite a potential reduction in CCR7 levels, these DCs do still acquire strong antigen-presenting capabilities, as they are able to activate CD4 T cells in allogenic mixed leukocyte reactions, a measure of their function as DCs. This is likely due to the upregulation of the costimulatory molecule CD86 and the antigen-presenting molecules MHCII DQ and DR (Giordano et al. 2003). This effect of cyclic nucleotides seems to be dependent on the timing of exposure to cAMP-inducing agents during differentiation or maturation, as somewhat disparate results are obtained depending on the methodology used for differentiation, maturation, and cyclic nucleotide elevation.
Interestingly, dendritic cell maturation in serum-free conditions, but in the presence of a cytokine cocktail, leads to a DC capable of a nonbiased Th2 response (Morelli and Thomson 2003; Scandella et al. 2002). While this is in contrast with previous in vitro data using fetal calf serum-containing media, it is likely that in the context of an inflammatory response in vivo the effect of PGE2 is to permit the generation of DCs that are capable of secreting the full range of cytokines, not just Th2 cytokines.
Stimulation of adenosine receptors and subsequent elevation of cAMP during monocyte to DC differentiation results in a distinct cell population characterized by changes in expression of cell surface markers common to both DCs and monocyte/macrophages, but remaining morphologically similar to DCs (Haskó et al. 2009). Using knockout animals and pharmacologic inhibition of adenosine receptors, it was discovered that the Gαs-coupled A2B adenosine receptor was the mediator of the phenotypic observations. Adenosine-differentiated DCs express high levels of angiogenic (VEGF), proinflammatory (IL-8, COX-2, IL-6), immune suppressive (IL-10, TGF-β), and tolerogenic factors (indoleamine 2,3-dioxygenase) (Novitskiy et al. 2008). Consequently, these cells are impaired in their ability to induce T-cell proliferation, by rendering the T cells anergic with indoleamine 2,3-dioxygenase, and also exhibited a reduced capacity for IFN-γ production. As expected, these phenotypic changes result in drastic alterations in the capacity of these cells to respond properly to abnormalities or inflammatory stimuli. For example, if these adenosine-differentiated DCs were injected directly into a subcutaneous tumor in mice, they actually promoted tumor growth through increased angiogenesis in the tumor, rather than resolving the abnormality (Novitskiy et al. 2008).
The role of the A2B receptor in cAMP-mediated regulation of pathological conditions and cell differentiation has been historically difficult to study due to its low affinity for adenosine and other selective agonists. A2B receptors are stimulated by micromolar concentrations of adenosine, rather than the normal physiological submicromolar levels commonly observed and associated with the A1, A2A, and A3 receptors. However, this low affinity may carry physiological importance as high concentrations of adenosine are often found at the site of traumatic injury, ischemia, or inflammation (Haskó et al. 2009). Overall, DC differentiation and maturation is retarded in the presence of elevated cyclic nucleotides, and this results in altered phenotypic characteristics affecting many key DC functions.
4 Osteoclast Differentiation
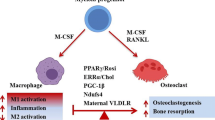
Osteoclasts are a unique cell type involved in calcium homeostasis in the body. They perform this function through the resorption of bone in response to either direct or indirect hormonal regulation by parathyroid hormone, calcitonin, and 1,25-dihydroxyvitamin D3 (Bar-Shavit 2007; Nakashima and Takayanagi 2009). Osteoclasts work in harmony with their bone-building counterparts, osteoblasts, to maintain the level of free calcium required for many bodily functions. Osteoblasts regulate formation and differentiation of osteoclasts and so are major players in homeostatic control of bone remodeling. cAMP and PDEs have profound effects on osteoblastic function, namely the regulation of receptor activator of nuclear factor-kappa B ligand (RANKL) production, but this will not be discussed in depth in this chapter, as osteoblasts are derived from the mesenchymal lineage, not the hematopoietic/monocytic lineage.
Osteoclasts are derived from the hematopoietic lineage with many steps along their differentiation pathway, but require only two major cytokines to complete their maturation. First, the hematopoietic stem cell can be started along the osteoclastic pathway by responding to cytokines such as M-CSF to become an osteoclast precursor cell or CFU-M (see Fig. 1). These cells are monocyte-like and can be differentiated to macrophages or dendritic cells at this stage, if they do not receive additional signals guiding them toward an osteoclast. This major driving signal for osteoclast formation is the RANKL cytokine produced by osteoblasts in response to hormonal stimulation. RANKL causes the CFU-M to commit to becoming an osteoclast and soon becomes a mononucleated osteoclast. Osteoclasts must fuse together and become multinucleated (three or more nuclei) and fully mature before they are able to perform their bone-resorbing functions (Bar-Shavit 2007; Nakashima and Takayanagi 2009).
Elevated cAMP, increased through treatment with PGE2, has been reported to both increase and decrease osteoclast formation. These discrepancies seem to be attributable to a number of factors: the dose of PGE2 being administered, the timing of PGE2 administration, the type of cells being studied, and the methods used to prepare osteoclasts and/or osteoblasts. It has been demonstrated that addition of a cAMP-elevating agonist such as PGE2 or forskolin or an inhibition of PDE activity can increase osteoclast formation from either human or mouse bone marrow cultures. Several groups have reported an increase in osteoclast formation from bone marrow with PGE2 (Fujita et al. 2003; Noh et al. 2009; Take et al. 2005; Wani et al. 1999), 8-isoprostaglandin E2 (an oxidized form of PGE2) (Tintut et al. 2002), forskolin (Ransjö et al. 1999), a cAMP analog (Wani et al. 1999), or PDE inhibition (Noh et al. 2009; Park and Yim 2007; Takami et al. 2005; Yamagami et al. 2003). This increase in osteoclast number and function can derive from two separate mechanisms: the stimulation of osteoblasts to produce RANKL, and direct effects on osteoclast precursors.
The concentration of PGE2 is vitally important in the regulation of osteoclast differentiation from bone marrow. Moderate doses of PGE2, ranging from 1 to 100 nM, seem to stimulate osteoclastogenesis in adherent cell-depleted hematopoietic monocultures in the presence of M-CSF and RANKL for both mouse and human bone marrow cells (Fujita et al. 2003; Ono et al. 2005). One group reported a synergistic increase in osteoclast formation when stimulated with RANKL and 1 μM PGE2 in bone marrow cultures (Wani et al. 1999). However, several groups report that when the concentration of PGE2 is 1 μM or higher, osteoclastogenesis is inhibited and subsequent bone resorption is decreased (Fujita et al. 2003; Ono et al. 2005; Park and Yim 2007). This does not hold true for all cell types as in RAW 264.7 cells, a mouse monocyte-like cell line, PGE2 concentrations at 1 μM stimulate osteoclast formation through PKA-dependent phosphorylation of TAK1 (Kobayashi et al. 2005a).
A second model of osteoclast generation uses CD14+ monocytes from human blood differentiated in the presence of M-CSF and RANKL to form osteoclasts. This monocyte to osteoclast transformation is inhibited by PGE2 at all concentrations of PGE2 tested, 0.1–100 nM (Itonaga et al. 1999; Take et al. 2005). However, it was found that the timing of administration of PGE2 also played a critical role in the ability of osteoclast precursors to differentiate into osteoclasts. If PGE2 was added at days 0–2 of culture, differentiation was prevented, if added later in the maturation process at days 6–8 there was no change in the number of mature osteoclasts after 8 days (Take et al. 2005). Similarly, the differentiation of bone marrow cells was stimulated when PGE2 was present during days 0–2 of culture, but no change was observed when added at days 6–8.
In order for the cells to be responsive to PGE2 levels, they must carry the appropriate receptors. The prostaglandin receptors are EP1, 2, 3, and 4 coupled to Gαq, Gαs, Gαi, and Gαs, respectively. The high level expression of the prostaglandin receptors EP2 and EP4 in precursor cells allows the cells to be responsive to stimuli like PGE2 and produce cAMP through adenylyl cyclase stimulation. Upon differentiation, the prostaglandin receptors are downregulated, preventing the stimulation or inhibition of the mature osteoclast by PGE2 (Kobayashi et al. 2005b; Take et al. 2005). If EP4 is reintroduced using an adenoviral transfection system, osteoclastic function was drastically inhibited (Kobayashi et al. 2005b). PGE2 also can directly inhibit bone resorbing function in mature OCs thru EP4 (Mano et al. 2000) and an EP4 antagonist inhibited osteoclast formation (Ono et al. 1998). EP1 and EP3 do not seem to play a major role in osteoclast formation because no increase in osteoclast formation was observed with the administration of butaprost, an EP1/3 agonist, while EP2-selective agonists slightly increase osteoclast formation (Ono et al. 1998). The Gαs-coupled prostaglandin receptors EP2 and EP4 seem to have nonredundant roles in osteoclast formation as EP2 knockout mice exhibit impaired osteoclast development, likely due to both a defect in the osteoblast and having direct effects on PGE2-stimulated osteoclast function (Li et al. 2000).
PDEs also play a role in controlling cAMP levels during differentiation of bone marrow precursors to osteoclasts. Inhibition of PDE4 with rolipram indirectly stimulates osteoclast formation through the ERK, p38, MAP kinase pathways by leading to a change in COX2 expression in osteoblasts. Elevated COX2 expression leads to an increase in PGE2, which in turn increased RANKL production by the osteoblasts (Cho et al. 2004; Park et al. 2007; Takami et al. 2005). PDE4 inhibition can also have effects directly on osteoclast formation from their precursors. Yamagami and colleagues showed that when the PDE4 inhibitor XT-611 was added to either a culture of bone marrow-derived osteoclastic precursors, or a coculture with osteoblasts, osteoclast formation was blocked when given concomitantly with PGE2. This effect seemed to be specific for PDE4 inhibition, as inhibition with either a PDE3-selective inhibitor milrinone, or the cGMP-PDE inhibitor zaprinast, had no effect on osteoclast differentiation in the presence of PGE2 (Yamagami et al. 2003). In a similar study, it was found that both PDE3 and PDE4 inhibitors had an effect on osteoclastic differentiation; however, these effects were largely on the osteoblasts, not on the osteoclasts themselves (Noh et al. 2009). PDE4 inhibitors can also potentiate the effects of endogenous agonists such as calcitonin. Calcitonin acts directly on osteoclasts to decrease osteoclastogenesis and this process was found to be PKA dependent and rolipram potentiated these effects (Miyamoto et al. 2006).
PDE4 inhibitors as a therapeutic intervention for osteoporosis show some promise based on animal models of osteopenia. Additionally, single-nucleotide polymorphisms in the PDE4D gene have been shown to correlate with variations in bone mineral density, possibly implicating PDE4D as a contributing genetic factor in human osteoporosis (Reneland et al. 2005). Both rolipram and pentoxifylline were reported to increase bone density in normal mice after 5 weeks of treatment (Kinoshita et al. 2000). In rats, a PDE4 inhibitor decreased bone loss in a model of osteoporosis in rats (Miyamoto et al. 1997) and increased bone density in ovarectomized, neurectomized rats (Waki et al. 1999).
One report regarding cGMP and osteoclastogenesis suggests that cGMP can block the differentiation of osteoclasts, but only when present during the final 3 days of a 6-day differentiation process (Holliday et al. 1997). As there has been some debate as to the pathways used for intracellular signaling by nitric oxide, both nitric oxide releasers such as sodium nitroprusside, used at low concentrations, and the cGMP analogues 8-Br-cGMP and dibutryl-cGMP, were used to demonstrate reduced osteoclast formation in the presence of elevated cGMP. Elevating cGMP using an inhibitor of PDE5, zaprinast, also inhibited osteoclast formation when administered through the final 3 days of differentiation, while decreasing cGMP using nitric oxide synthase inhibitors increased osteoclast formation. It was concluded that elevated cGMP inhibits osteoclast formation, but only when present during the final stages of differentiation.
5 Murine Bone Marrow Differentiation
Bone marrow progenitors can be differentiated to macrophages in the presence of a number of factors, such as M-CSF, GM-CSF, and PMA. It has long been observed that agents that can elevate cAMP can inhibit macrophage formation from progenitors. M-CSF can also induce proliferation of differentiated bone marrow macrophages, and PGE2 negatively regulates this process (Kurland et al. 1977, 1978a, b). Several agents that can increase cAMP, such as PGE2, 8-Br-cAMP, and IBMX, can suppress the mitogenic actions of M-CSF, GM-CSF, and PMA through the inhibition of DNA synthesis. This effect of cAMP does not appear to affect the cells’ early response to colony-stimulating factors, but exerts its effect during the late G1 phase in the cell cycle (Vairo et al. 1990). Inhibition by PGE2 seems to be part of a negative feedback loop controlling proliferation of monocytes and macrophages. PGE2 may feedback to control macrophage formation by inhibiting proliferation and differentiation of immature monocytoid cells.
M-CSF-induced proliferation of mouse bone marrow macrophages is suppressed by adenosine-mediated signaling through the Gαs-coupled A2B receptors. The downstream cAMP signaling mechanism involved a PKA-dependent induction of a cyclin-dependent kinase inhibitor known as p27kip-1 that leads to growth arrest at the G1 phase of the cell cycle (Xaus et al. 1999). This molecular mechanism could potentially apply to the negative regulation of monocytic proliferation caused with elevated cAMP, regardless of the source, but this has not been fully tested in all cell systems. Using A2B knockout mice and pharmacological inhibition of adenosine receptors several groups demonstrated that adenosine or ATP signaling through this receptor inhibits TLR-dependent IL-12p70 and TNF-α production in bone marrow derived DCs while enhancing IL-10 production (Addi Abduelhakem et al. 2008; Wilson et al. 2009). These cells were also less effective at stimulating T-cell proliferation, likely due to reduced MHC II and CD86 levels.
Studies were also performed in a mouse myeloid cell line to demonstrate the effects of cAMP on differentiation. Increasing cAMP levels using either 8-Br-cAMP or PGE1 can induce macrophage differentiation in M1 myeloid cells, which were engineered to express the M-CSF receptor (Wilson et al. 2005). Increased cAMP can potentiate M-CSF-induced differentiation in cells containing the receptor through ERK activation, as differentiation was halted with the addition of a MEK inhibitor. These cells also retain the ability to inhibit M-CSF stimulated macrophage proliferation through phosphorylation of the M-CSF receptor. It has also been shown that cAMP can modulate M-CSF-induced MAPK activation of ERK, JNK, and p38 in a time-dependent manner, thereby inhibiting macrophage development (Zhu et al. 2008).
6 Conclusions
PDEs are a class of enzymes with a number of isoforms distributed throughout all tissues and cell types in the body. PDE5 inhibition has emerged as a proven method to treat erectile dysfunction and pulmonary hypertension. The ability to selectively inhibit a PDE in order to modulate cellular function is an exciting prospect for many future therapies. However, the administration of these inhibitors may carry unintended consequences when they affect the differentiation or activation of cells also containing the targeted PDE. For example, chronic administration of a PDE inhibitor would presumably increase the cyclic nucleotide levels in all cell types containing the targeted PDE. Observations of these increased levels are difficult to confirm in vivo and in vitro work utilizing monocyte or bone marrow-derived cells has been widely used as a surrogate for in vivo differentiation conditions.
Elevated cyclic nucleotide levels can have profound effects on the final phenotype of the cell when present during differentiation or maturation. Dendritic cell differentiation can be arrested in the presence of increased cAMP or cGMP at an intermediate stage between monocyte and dendritic cell. The capabilities of these dendritic cells are also altered, skewing its T-cell activation capacity toward a Th2 type response, largely due to the lack of IL-12 produced. Macrophage differentiation does not seem to be halted by elevated cyclic nucleotides, but a number of characteristics are altered. cAMP normally has anti-inflammatory actions on macrophages, but recent work has uncovered the role of cAMP in increasing chemokine and cytokine expression.
The alteration of hematopoietic differentiation by cyclic nucleotides and PDE inhibitors is also important regarding clinical efforts to generate therapeutic cell types from monocytes ex vivo. Generation of specific cell types from pluripotent and multipotent cells ex vivo for therapeutic usage holds great promise. For example, numerous efforts have been undertaken to develop DC or macrophage populations ex vivo that target specific antigens for fighting cancer (Paczesny et al. 2003), to treat chest wound infections after open heart surgery (Orenstein et al. 2005), to treat other refractory wounds (Zuloff-Shani et al. 2004), as a treatment for spinal cord injury (Knoller et al. 2005), and as gene therapy delivery vehicles (Burke et al. 2002; Pastorino et al. 2001).
Although the phenotypes of the macrophages used for some therapeutic applications have been partly characterized, it is unclear which features are critical for their therapeutic effect. Currently, macrophages and other hematopoietic-derived cells are being generated ex vivo using serum and other mixed indeterminate stimuli. Using specific combinations of cytokines and humoral factors may allow generation of a more defined cell type with specific features that make it more therapeutically effective.
References
Addi Abduelhakem B, Lefort A, Hua X, Libert F, Communi D, Ledent C, Macours P, Tilley SL, Boeynaems J-M, Robaye B (2008) Modulation of murine dendritic cell function by adenine nucleotides and adenosine: involvement of the A2B receptor. Eur J Immunol 38:1610–1620
Baillie GS, MacKenzie SJ, McPhee I, Houslay MD (2000) Sub-family selective actions in the ability of Erk2 MAP kinase to phosphorylate and regulate the activity of PDE4 cyclic AMP-specific phosphodiesterases. Br J Pharmacol 131:811–819
Bang B, Ericsen C, Aarbakke J (1994) Effects of cAMP and cGMP elevating agents on HL-60 cell differentiation. Pharmacol Toxicol 75:108–112
Bar-Shavit Z (2007) The osteoclast: a multinucleated, hematopoietic-origin, bone-resorbing osteoimmune cell. J Cell Biochem 102:1130–1139
Bender AT, Beavo JA (2004) Specific localized expression of cGMP PDEs in Purkinje neurons and macrophages. Neurochem Int 45:853–857
Bender AT, Beavo JA (2006a) Cyclic nucleotide phosphodiesterases: molecular regulation to clinical use. Pharmacol Rev 58:488–520
Bender AT, Beavo JA (2006b) PDE1B2 regulates cGMP and a subset of the phenotypic characteristics acquired upon macrophage differentiation from a monocyte. Proc Natl Acad Sci USA 103:460–465
Bender AT, Ostenson CL, Giordano D, Beavo JA (2004) Differentiation of human monocytes in vitro with granulocyte-macrophage colony-stimulating factor and macrophage colony-stimulating factor produces distinct changes in cGMP phosphodiesterase expression. Cell Signal 16:365–374
Bender AT, Ostenson CL, Wang EH, Beavo JA (2005) Selective up-regulation of PDE1B2 upon monocyte-to-macrophage differentiation. Proc Natl Acad Sci USA 102:497–502
Boeynaems J-M, Communi D, Savi P, Herbert J-M (2000) P2Y receptors: in the middle of the road. Trends Pharmacol Sci 21:1–3
Bornfeldt KE (2006) A single second messenger: several possible cellular responses depending on distinct subcellular pools. Circ Res 99:790–792
Boss G (1989) cGMP-induced differentiation of the promyelocytic cell line HL-60. Proc Natl Acad Sci USA 86:7174–7178
Brodsky A, Davio C, Shayo C, Lemos Legnazzi B, Barbosa M, Lardo M, Morelli A, Baldi A, Sanchez Avalos JC, Rivera E (1998) Forskolin induces U937 cell line differentiation as a result of a sustained cAMP elevation. Eur J Pharmacol 350:121–127
Bryn T, Mahic M, Enserink JM, Schwede F, Aandahl EM, Tasken K (2006) The cyclic AMP-Epac1-Rap1 pathway is dissociated from regulation of effector functions in monocytes but acquires immunoregulatory function in mature macrophages. J Immunol 176:7361–7370
Burke B, Sumner S, Maitland N, Lewis CE (2002) Macrophages in gene therapy: cellular delivery vehicles and in vivo targets. J Leukoc Biol 72:417–428
Chaplinski TJ, Niedel JE (1982) Cyclic nucleotide-induced maturation of human promyelocytic leukemia cells. J Clin Invest 70:953–964
Cho Y-J, Kim J-Y, Jeong S-W, Lee SB, Kim ON (2003) Cyclic AMP induces activation of extracellular signal-regulated kinases in HL-60 cells: role in cAMP-induced differentiation. Leuk Res 27:51–56
Cho E, Yu J, Kim M, Yim M (2004) Rolipram, a phosphodiesterase 4 inhibitor, stimulates osteoclast formation by inducing TRANCE expression in mouse calvarial cells. Arch Pharm Res 27:1258–1262
Conti M, Richter W, Mehats C, Livera G, Park J-Y, Jin C (2003) Cyclic AMP-specific PDE4 phosphodiesterases as critical components of cyclic AMP signaling. J Biol Chem 278:5493–5496
Essayan DM (2001) Cyclic nucleotide phosphodiesterases. J Allergy Clin Immunol 108:671–680
Fernandez N, Monczor F, Lemos B, Notcovich C, Baldi A, Davio C, Shayo C (2002) Reduction of G protein-coupled receptor kinase 2 expression in U-937 cells attenuates H2 histamine receptor desensitization and induces cell maturation. Mol Pharmacol 62:1506–1514
Fidock M, Miller M, Lanfear J (2002) Isolation and differential tissue distribution of two human cDNAs encoding PDE1 splice variants. Cell Signal 14:53–60
Fischmeister R, Castro LRV, Abi-Gerges A, Rochais F, Jurevicius J, Leroy J, Vandecasteele G (2006) Compartmentation of cyclic nucleotide signaling in the heart: the role of cyclic nucleotide phosphodiesterases. Circ Res 99:816–828
Fujita D, Yamashita N, Iita S, Amano H, Yamada S, Sakamoto K (2003) Prostaglandin E2 induced the differentiation of osteoclasts in mouse osteoblast-depleted bone marrow cells. Prostaglandins Leukot Essent Fatty Acids 68:351–358
Gantner F, Kupferschmidt R, Schudt C, Wendel A, Hatzelmann A (1997a) In vitro differentiation of human monocytes to macrophages: change of PDE profile and its relationship to suppression of tumour necrosis factor-[alpha] release by PDE inhibitors. Br J Pharmacol 121:221–231
Gantner F, Tenor H, Gekeler V, Schudt C, Wendel A, Hatzelmann A (1997b) Phosphodiesterase profiles of highly purified human peripheral blood leukocyte populations from normal and atopic individuals: a comparative study. J Allergy Clin Immunol 100:527–535
Gantner F, Schudt C, Wendel A, Hatzelmann A (1999) Characterization of the phosphodiesterase (PDE) pattern of in vitro-generated human dendritic cells (DC) and the influence of PDE inhibitors on DC function. Pulm Pharmacol Ther 12:377–386
Geissmann F, Manz MG, Jung S, Sieweke MH, Merad M, Ley K (2010) Development of monocytes, macrophages, and dendritic cells. Science 327:656–661
Gilchrist M, Thorsson V, Li B, Rust AG, Korb M, Kennedy K, Hai T, Bolouri H, Aderem A (2006) Systems biology approaches identify ATF3 as a negative regulator of Toll-like receptor 4. Nature 441:173–178
Giordano D, Magaletti DM, Clark EA, Beavo JA (2003) Cyclic nucleotides promote monocyte differentiation toward a DC-SIGN+ (CD209) intermediate cell and impair differentiation into dendritic cells. J Immunol 171:6421–6430
Grage-Griebenow E, Flad HD, Ernst M (2001) Heterogeneity of human peripheral blood monocyte subsets. J Leukoc Biol 69:11–20
Harris P, Ralph P (1985) Human leukemic models of myelomonocytic development: a review of the HL-60 and U937 cell lines. J Leukoc Biol 37:407–422
Haskó G, Csóka B, Németh ZH, Vizi ES, Pacher P (2009) A2B adenosine receptors in immunity and inflammation. Trends Immunol 30:263–270
Hertz AL, Bender AT, Smith KC, Gilchrist M, Amieux PS, Aderem A, Beavo JA (2009a) Elevated cyclic AMP and PDE4 inhibition induce chemokine expression in human monocyte-derived macrophages. Proc Natl Acad Sci USA. doi:10.1073/pnas.0911684106
Hertz AL, Bender AT, Smith KC, Gilchrist M, Amieux PS, Aderem A, Beavo JA (2009b) Elevated cyclic AMP and PDE4 inhibition induce chemokine expression in human monocyte-derived macrophages. Proc Natl Acad Sci 106:21978–21983
Heystek HC, Thierry A-C, Soulard P, Moulon C (2003) Phosphodiesterase 4 inhibitors reduce human dendritic cell inflammatory cytokine production and Th1-polarizing capacity. Int Immunol 15:827–835
Holliday LS, Dean AD, Lin RH, Greenwald JE, Gluck SL (1997) Low NO concentrations inhibit osteoclast formation in mouse marrow cultures by cGMP-dependent mechanism. Am J Physiol Renal Physiol 272:F283–F291
Houslay MD (2010) Underpinning compartmentalised cAMP signalling through targeted cAMP breakdown. Trends Biochem Sci 35:91–100
Houslay MD, Schafer P, Zhang KYJ (2005) Keynote review: phosphodiesterase-4 as a therapeutic target. Drug Discov Today 10:1503–1519
Itonaga I, Sabokbar A, Neale SD, Athanasou NA (1999) 1, 25-Dihydroxyvitamin D3 and prostaglandin E2 act directly on circulating human osteoclast precursors. Biochem Biophys Res Comm 264:590–595
Jiang L, Foster FM, Ward P, Tasevski V, Luttrell BM, Conigrave AD (1997) Extracellular ATP triggers cyclic AMP-dependent differentiation of HL-60 cells. Biochem Biophys Res Comm 232:626–630
Jonuleit H, Kühn U, Müller G, Steinbrink K, Paragnik L, Schmitt E, Knop J, Enk A (1997) Pro-inflammatory cytokines and prostaglandins induce maturation of potent immunostimulatory dendritic cells under fetal calf serum-free conditions. Eur J Immunol 27:3135–3142
Kalinski P, Hilkens CM, Snijders A, Snijdewint FG, Kapsenberg ML (1997) IL-12-deficient dendritic cells, generated in the presence of prostaglandin E2, promote type 2 cytokine production in maturing human naive T helper cells. J Immunol 159:28–35
Kalinski P, Schuitemaker JHN, Hilkens CMU, Kapsenberg ML (1998) Prostaglandin E2 induces the final maturation of IL-12-deficient CD1a+CD83+ dendritic cells: the levels of IL-12 are determined during the final dendritic cell maturation and are resistant to further modulation. J Immunol 161:2804–2809
Kinoshita T, Kobayashi S, Ebara S, Yoshimura Y, Horiuchi H, Tsutsumimoto T, Wakabayashi S, Takaoka K (2000) Phosphodiesterase inhibitors, pentoxifylline and rolipram, increase bone mass mainly by promoting bone formation in normal mice. Bone 27:811–817
Knoller N, Auerbach G, Fulga V, Zelig G, Attias J, Bakimer R, Marder JB, Yoles E, Belkin M, Schwartz M, Hadani M (2005) Clinical experience using incubated autologous macrophages as a treatment for complete spinal cord injury: phase I study results. J Neurosurg Spine 3:173–181
Kobayashi Y, Mizoguchi T, Take I, Kurihara S, Udagawa N, Takahashi N (2005a) Prostaglandin E2 enhances osteoclastic differentiation of precursor cells through protein kinase A-dependent phosphorylation of TAK1. J Biol Chem 280:11395–11403
Kobayashi Y, Take I, Yamashita T, Mizoguchi T, Ninomiya T, Hattori T, Kurihara S, Ozawa H, Udagawa N, Takahashi N (2005b) Prostaglandin E2 receptors EP2 and EP4 are down-regulated during differentiation of Mouse osteoclasts from their precursors. J Biol Chem 280:24035–24042
Kurland JI, Hadden JW, Moore MAS (1977) Role of cyclic nucleotides in the proliferation of committed granulocyte-macrophage progenitor cells. Cancer Res 37:4534–4538
Kurland J, Bockman R, Broxmeyer H, Moore M (1978a) Limitation of excessive myelopoiesis by the intrinsic modulation of macrophage-derived prostaglandin E. Science 199:552
Kurland J, Broxmeyer H, Pelus M, Bockman R, Moore M (1978b) Role for monocyte-macrophage-derived colony stimulating factor and prostaglandin E in the positive and negative feedback control of myeloid stem cell proliferation. Blood 52:388
Legnazzi BL, Shayo C, Monczor F, Martin ME, Fernandez N, Brodsky A, Baldi A, Davio C (2000) Rapid desensitization and slow recovery of the cyclic AMP response mediated by histamine H2 receptors in the U937 cell line. Biochem Pharmacol 60:159–166
Li X, Okada Y, Pilbeam CC, Lorenzo JA, Kennedy CRJ, Breyer RM, Raisz LG (2000) Knockout of the murine prostaglandin EP2 receptor impairs osteoclastogenesis in vitro. Endocrinology 141:2054–2061
Luft T, Jefford M, Luetjens P, Toy T, Hochrein H, Masterman K-A, Maliszewski C, Shortman K, Cebon J, Maraskovsky E (2002) Functionally distinct dendritic cell (DC) populations induced by physiologic stimuli: prostaglandin E2 regulates the migratory capacity of specific DC subsets. Blood 100:1362–1372
Mano M, Arakawa T, Mano H, Nakagawa M, Kaneda T, Kaneko H, Yamada T, Miyata K, Kiyomura H, Kumegawa M, Hakeda Y (2000) Prostaglandin E2 directly inhibits bone-resorbing activity of isolated mature osteoclasts mainly through the EP4 receptor. Calcif Tissue Int 67:85–92
Miyamoto K-i, Waki Y, Horita T, Kasugai S, Ohya K (1997) Reduction of bone loss by denbufylline, an inhibitor of phosphodiesterase 4. Biochem Pharmacol 54:613–617
Miyamoto K-i, Nishioka T, Waki Y, Nomura M, Katsuta H, Yokogawa K, Amano H (2006) Phosphodiesterase 4 inhibitor rolipram potentiates the inhibitory effect of calcitonin on osteoclastogenesis. J Bone Miner Metab 24:260–265
Monczor F, Fernandez N, Riveiro E, Mladovan A, Baldi A, Shayo C, Davio C (2006) Histamine H2 receptor overexpression induces U937 cell differentiation despite triggered mechanisms to attenuate cAMP signalling. Biochem Pharmacol 71:1219–1228
Morelli AE, Thomson AW (2003) Dendritic cells under the spell of prostaglandins. Trends Immunol 24:108–111
Morita R, Ukyo N, Furuya M, Uchiyama T, Hori T (2003) Atrial natriuretic peptide polarizes human dendritic cells toward a Th2-promoting phenotype through its receptor guanylyl cyclase-coupled receptor A. J Immunol 170:5869–5875
Mosser DM, Edwards JP (2008) Exploring the full spectrum of macrophage activation. Nat Rev Immunol 8:958–969
Nakashima T, Takayanagi H (2009) Osteoclasts and the immune system. J Bone Miner Metab 27:519–529
Noh ALSM, Yang M, Lee J-M, Park H, Lee D-S, Yim M (2009) Phosphodiesterase 3 and 4 negatively regulate receptor activator of nuclear factor-kappa B ligand-mediated osteoclast formation by prostaglandin E2. Biol Pharm Bull 32:1844–1848
Nonaka T, Mio M, Doi M, Tasaka K (1992) Histamine-induced differentiation of HL-60 cells: the role of cAMP and protein kinase A. Biochem Pharmacol 44:1115–1121
Novitskiy SV, Ryzhov S, Zaynagetdinov R, Goldstein AE, Huang Y, Tikhomirov OY, Blackburn MR, Biaggioni I, Carbone DP, Feoktistov I, Dikov MM (2008) Adenosine receptors in regulation of dendritic cell differentiation and function. Blood 112:1822–1831
Olsson IL, Breitman TR, Gallo RC (1982) Priming of human myeloid leukemic cell lines HL-60 and U-937 with retinoic acid for differentiation effects of cyclic adenosine 3′:5′-monophosphate-inducing agents and a T-lymphocyte-derived differentiation factor. Cancer Res 42:3928–3933
Ono K, Akatsu T, Murakami T, Nishikawa M, Yamamoto M, Kugai N, Motoyoshi K, Nagata N (1998) Important role of EP4, a subtype of prostaglandin (PG) E receptor, in osteoclast-like cell formation from mouse bone marrow cells induced by PGE2. J Endocrinol 158:R1–R5
Ono K, Kaneko H, Choudhary S, Pilbeam CC, Lorenzo JA, Akatsu T, Kugai N, Raisz LG (2005) Biphasic effect of prostaglandin E2 on osteoclast formation in spleen cell cultures: role of the EP2 receptor. J Bone Miner Res 20:23–29
Orenstein A, Kachel E, Zuloff-Shani A, Paz Y, Sarig O, Haik J, Smolinsky AK, Mohr R, Shinar E, Danon D (2005) Treatment of deep sternal wound infections post-open heart surgery by application of activated macrophage suspension. Wound Repair Regen 13:237–242
Paczesny S, Ueno H, Fay J, Banchereau J, Palucka AK (2003) Dendritic cells as vectors for immunotherapy of cancer. Semin Cancer Biol 13:439–447
Park H, Yim M (2007) Rolipram, a phosphodiesterase 4 inhibitor, suppresses PGE2-induced osteoclast formation by lowering osteoclast progenitor cell viability. Arch Pharm Res 30:486–492
Park H, Young Lee S, Lee D-S, Yim M (2007) Phosphodiesterase 4 inhibitor regulates the TRANCE/OPG ratio via COX-2 expression in a manner similar to PTH in osteoblasts. Biochem Biophys Res Comm 354:178–183
Pastorino S, Massazza S, Cilli M, Varesio L, Bosco MC (2001) Generation of high-titer retroviral vector-producing macrophages as vehicles for in vivo gene transfer. Gene Ther 8:431–441
Poppe H, Rybalkin SD, Rehmann H, Hinds TR, Tang X-B, Christensen AE, Schwede F, Genieser H-G, Bos JL, Doskeland SO, Beavo JA, Butt E (2008) Cyclic nucleotide analogs as probes of signaling pathways. Nat Meth 5:277–278
Ransjö M, Lie A, Mackie EJ (1999) Cholera toxin and forskolin stimulate formation of osteoclast-like cells in mouse marrow cultures and cultured mouse calvarial bones. Eur J Oral Sci 107:45–54
Reneland R, Mah S, Kammerer S, Hoyal C, Marnellos G, Wilson S, Sambrook P, Spector T, Nelson M, Braun A (2005) Association between a variation in the phosphodiesterase 4D gene and bone mineral density. BMC Med Genet 6:9
Scandella E, Men Y, Gillessen S, Forster R, Groettrup M (2002) Prostaglandin E2 is a key factor for CCR7 surface expression and migration of monocyte-derived dendritic cells. Blood 100:1354–1361
Shayo C, Davio C, Brodsky A, Mladovan AG, Legnazzi BL, Rivera E, Baldi A (1997) Histamine modulates the expression of c-fos through cyclic AMP production via the H2 receptor in the human promonocytic cell line U937. Mol Pharmacol 51:983–990
Shayo C, Fernandez N, Legnazzi BL, Monczor F, Mladovan A, Baldi A, Davio C (2001) Histamine H2 receptor desensitization: involvement of a select array of G protein-coupled receptor kinases. Mol Pharmacol 60:1049–1056
Shayo C, Legnazzi BL, Monczor F, Fernández N, Riveiro ME, Baldi A, Davio C (2004) The time-course of cyclic AMP signaling is critical for leukemia U-937 cell differentiation. Biochem Biophys Res Comm 314:798–804
Shepherd MC, Baillie GS, Stirling DI, Houslay MD (2004) Remodelling of the PDE4 cAMP phosphodiesterase isoform profile upon monocyte-macrophage differentiation of human U937 cells. Br J Pharmacol 142:339–351
Spina D (2008) PDE4 inhibitors: current status. Br J Pharmacol 155:308–315
Takami M, Cho ES, Lee SY, Kamijo R, Yim M (2005) Phosphodiesterase inhibitors stimulate osteoclast formation via TRANCE/RANKL expression in osteoblasts: possible involvement of ERK and p38 MAPK pathways. FEBS Lett 579:832–838
Take I, Kobayashi Y, Yamamoto Y, Tsuboi H, Ochi T, Uematsu S, Okafuji N, Kurihara S, Udagawa N, Takahashi N (2005) Prostaglandin E2 strongly inhibits human osteoclast formation. Endocrinology 146:5204–5214
Tintut Y, Parhami F, Tsingotjidou A, Tetradis S, Territo M, Demer LL (2002) 8-Isoprostaglandin E2 enhances receptor-activated NFkB ligand (RANKL)-dependent osteoclastic potential of marrow hematopoietic precursors via the cAMP pathway. J Biol Chem 277:14221–14226
Torphy TJ, Zhou HL, Cieslinski LB (1992) Stimulation of beta adrenoceptors in a human monocyte cell line (U937) up-regulates cyclic AMP-specific phosphodiesterase activity. J Pharmacol Exp Ther 263:1195–1205
Tortora G, Clair T, Cho-Chung YS (1990) An antisense oligodeoxynucleotide targeted against the type II beta regulatory subunit mRNA of protein kinase inhibits cAMP-induced differentiation in HL-60 leukemia cells without affecting phorbol ester effects. Proc Natl Acad Sci USA 87:705–708
Tortora G, Yokozaki H, Pepe S, Clair T, Cho-Chung YS (1991) Differentiation of HL-60 leukemia by type I regulatory subunit antisense oligodeoxynucleotide of cAMP-dependent protein kinase. Proc Natl Acad Sci USA 88:2011–2015
Tudhope SJ, Finney-Hayward TK, Nicholson AG, Mayer RJ, Barnette MS, Barnes PJ, Donnelly LE (2008) Different mitogen-activated protein kinase-dependent cytokine responses in cells of the monocyte lineage. J Pharmacol Exp Ther 324:306–312
Vairo G, Argyriou S, Bordun AM, Whitty G, Hamilton JA (1990) Inhibition of the signaling pathways for macrophage proliferation by cyclic AMP. Lack of effect on early responses to colony stimulating factor-1. J Biol Chem 265:2692–2701
Verghese MW, McConnell RT, Lenhard JM, Hamacher L, Jin SL (1995) Regulation of distinct cyclic AMP-specific phosphodiesterase (phosphodiesterase type 4) isozymes in human monocytic cells. Mol Pharmacol 47:1164–1171
Waki Y, Horita T, Miyamoto K-i, Ohya K, Kasugai S (1999) Effects of XT-44, a phosphodiesterase 4 Inhibitor, in osteoblastgenesis and osteoclastgenesis in culture and its therapeutic effects. Jpn J Pharmacol 79:477–483
Wani MR, Fuller K, Kim NS, Choi Y, Chambers T (1999) Prostaglandin E2 cooperates with TRANCE in osteoclast induction from hemopoietic precursors: synergistic activation of differentiation, cell spreading, and fusion. Endocrinology 140:1927–1935
Wilson NJ, Cross M, Nguyen T, Hamilton JA (2005) cAMP inhibits CSF-1-stimulated tyrosine phosphorylation but augments CSF-1R-mediated macrophage differentiation and ERK activation. FEBS J 272:4141–4152
Wilson JM, Ross WG, Agbai ON, Frazier R, Figler RA, Rieger J, Linden J, Ernst PB (2009) The A2B adenosine receptor impairs the maturation and immunogenicity of dendritic cells. J Immunol 182:4616–4623
Xaus J, Valledor AF, Cardo M, Marques L, Beleta J, Palacios JM, Celada A (1999) Adenosine inhibits macrophage colony-stimulating factor-dependent proliferation of macrophages through the induction of p27kip-1 expression. J Immunol 163:4140–4149
Yamagami H, Nishioka T, Ochiai E, Fukushima K, Nomura M, Kasugai S, Moritani S, Yokogawa K, Miyamoto K-i (2003) Inhibition of osteoclastogenesis by a phosphodiesterase 4 inhibitor XT-611 through synergistic action with endogenous prostaglandin E2. Biochem Pharmacol 66:801–807
Zhu N, Cui J, Qiao C, Li Y, Ma Y, Zhang J, Shen B (2008) cAMP modulates macrophage development by suppressing M-CSF-induced MAPKs activation. Cell Mol Immunol 5:153–157
Zuloff-Shani A, Kachel E, Frenkel O, Orenstein A, Shinar E, Danon D (2004) Macrophage suspensions prepared from a blood unit for treatment of refractory human ulcers. Transfus Apher Sci 30:163–167
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2011 Springer-Verlag Berlin Heidelberg
About this chapter
Cite this chapter
Hertz, A.L., Beavo, J.A. (2011). Cyclic Nucleotides and Phosphodiesterases in Monocytic Differentiation. In: Francis, S., Conti, M., Houslay, M. (eds) Phosphodiesterases as Drug Targets. Handbook of Experimental Pharmacology, vol 204. Springer, Berlin, Heidelberg. https://doi.org/10.1007/978-3-642-17969-3_16
Download citation
DOI: https://doi.org/10.1007/978-3-642-17969-3_16
Published:
Publisher Name: Springer, Berlin, Heidelberg
Print ISBN: 978-3-642-17968-6
Online ISBN: 978-3-642-17969-3
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)