
Abstract
Trehalose-containing glycolipids are mainly produced by Gram-positive, high GC content bacteria of Actinomycetales. Their structures are quite diverse in hydrophobic moiety, varying from short simple to long complex fatty acids. Correspondingly, functions and physiochemical properties vary upon structures. From the view of practical use as a biosurfactant, the trehalose lipids from Rhodococcus and the genera other than Mycobacterium are of high potential in application. While, like other kinds of biosurfactants, their relative low productivity limits practical use. And yet, the biosynthesis mechanism of trehalose lipids has been less exploited and needs further investigations.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

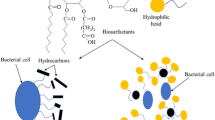

Keywords
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
1 Introduction
Trehalolipids, or trehalose lipids, are glycolipids containing trehalose hydrophilic moiety. They are among the best known biosurfactants but distinguished from rhamnolipids and sophorolipids in both composition and activity. They are active to lower water surface and interface tension. In addition, they function uniquely in many other ways.
The occurrence of trehalose in lipids was first noticed in 1933 in fat extracts from tubercle bacilli (Anderson and Newman 1933). At that time, the authors did not pay attention to the property as a biosurfactant but found intriguing that trehalose was in place of glycerol as the alcoholic component of a fat. However, the pure compound of trehalose lipid was first successfully purified in 1956 from the lipids of Mycobacterium tuberculosis and identified as 6, 6′-dimycoloyl-α′, α′-d-trehalose (cord factor). Later, several groups of trehalose-containing lipids were isolated, mainly from Mycobacteria, Nocardia, and Corynebacteria, as reviewed by Asselineau and Asselineau (Asselineau and Asselineau 1978).
Interest in trehalose lipids as a general surfactant can be traced back to the discovery that the emulsion layer of Arthrobacter paraffineus culture broths contained trehalose dimycolates when the cells were grown on hydrocarbon substrates (Suzuki et al. 1969). Various trehalose lipids varying in fatty acid part have been found. The glycolipids from mycobacteria usually have branched long-chain hydroxyaliphatic acids as their hydrophobic moiety; while some bacteria choose short-chain fatty acids, such as some cases of Rhodococcus. Trehalose lipids are quite diverse in chemical structure and attractive in applications as other biosurfactants.
2 Trehalolipid-Producing Bacteria
Production of trehalolipids mainly occurs in the following bacteria of Actinomycetales, including the genera of Rhodococcus, Mycobacterium, Micrococcus, Nocardia, Gordonia, Corynebacterium, Brevibacteria, Arthrobacter etc. (Table1). However, yeast and other fungus have seldom been reported as a producer of trehalose lipids.
Among the reported trehalolipid-producing bacteria, most are alkane-degrading bacteria. Surfactive compounds are generated only when bacteria feed on hydrophobic substrates such as n-hexadecane (listed in Table1), such as Rhodococcus erythropolis, Rhodococcus opacus, Rhodococcus wratislaviensis, Gordonia amarae, Arthrobacter parafineus, Micrococcus luteus, Mycobacterium paraffinicum Corynebacteria, etc. They have different isolation origins, some of which are from marine environments (Passeri et al. 1991; Schulz et al. 1991; Yakimov et al. 1999; Satpute et al. 2010); some are from alkaline soils or polar soils. These bacteria seem to be tolerant with high salt concentrations (up to 6% salinity). Among them, isolates of Rhodococcus are the most potential in biosurfactant production (Lang and Philp 1998).
Rhodococcus bacteria usually produce trehalose lipids when they grow with alkanes as carbon sources (Espuny et al. 1995; Rapp et al. 1979; Singer et al. 1990). Among the bacteria of this genus, isolates of R. erythropolis are most frequently reported as a producer of trehalose lipids (Lang and Philp 1998; Peng et al. 2007). On the other hand, glycolipids other than trehalose lipids have been also found recently in bacteria of Rhodococcus, e.g., a glucolipid was found in R. erythropolis isolate 3C-9 in addition to a trehalolipid (Peng et al. 2007); for another example, rhamnose glycolipid was found as the main biosurfactant in the case of Rhodococcus fascians isolate A-3 grown with glucose or kerosene as a substrate (Gesheva et al. 2010).
R. erythropolis 51T7 is a relatively intensively investigated bacterium. It was once named Corynebacterium sp. 51T7 at the beginning (Martin et al. 1991), then renamed as Rhodococcus sp. 51T7 (Espuny et al. 1996), and now named as R. erythropolis 51T7 (Marques et al. 2009). This bacterium has been confirmed to produce tetraesters of trehalose with short fatty acid chains (C8–C11).
Mycobacterium is another group of trehalolipid-producing bacteria. But the trehalolipids produced by Mycobacterium are seldom reported as a biosurfactant, interests to them are mainly attributed to their role in pathogenicity and molecular immunology (Imasato et al. 1990; Ryll et al. 2001; Hunter et al. 2006; Ortiz et al. 2008, 2009; Zaragoza et al. 2009). Many species of this genus have been reported to produce trehalose-containing lipids, such as Mycobacterium tuberculosis, Mycobacterium bovis, Mycobacterium paraffinicum, Mycobacterium phlei, Mycobacterium flavescens and Mycobacterium avium (Mompon et al. 1978; Batrakov et al. 1981; Fujita et al. 2005). In addition to their effects in health, bacteria of this genus also frequently reported as degraders of alkanes and polycyclic aromatic hydrocarbons (Hallas and Vestal 1978; Berekaa and Steinbuchel 2000; Kotani et al. 2006; Churchill et al. 2008; Vila and Grifoll 2009). The trehalolipids of mycobacteria might also involve in dissolving hydrophobic substrates. One example supporting this assumption is that a paraffin-oxidizing bacterium of M. paraffinicum can produce at least five trehalose lipids including the cord factor or two analogs (Batrakov et al. 1981).
3 Chemical Structures
Like other glycolipids, trehalose lipids are composed of a carbohydrate group in combination with fatty acids groups. Differently, their hydrophobic moieties are more diverse, including aliphatic acids and hydroxylated branched-chain fatty acids (mycolic acids) of varied chain lengths. The numbers of hydrophobic chain in each molecule of trehalose lipids are usually 1, 2, and 4, forming mono-, di-, and tetraesters, correspondingly. Triesters can also be the main glycolipid component in some cases, such as in Mycobacteriurn fortuitum (Gautier et al. 1992). Among the trehalose lipids, the trehalose esters produced by R. erytropolis have been studied most extensively, in addition to the cord factors of Mycobacteria.
Trehalose is a nonreducing sugar formed from two glucose units joined by a 1–1 alpha bond, giving it the name of α-d-glucopyranosyl-(1→1)-α-d-glucopyranoside (Fig.1). The bonding makes trehalose very resistant to acid hydrolysis, and therefore is stable in solution at high temperatures, even under acidic conditions. Trehalose is the carbohydrate group of cell wall glycolipids in Mycobacteria and Corynebacteria.

Trehalose structure
Mycolic acids were first identified in an unsaponifiable lipid extract isolated from M. tuberculosis. They are complex hydroxylated branched-chain fatty acids with 60–90 carbon atoms, while those from other species (Corynobacterium, Nocardia) are shorter and named corynomycolic (22–36 carbons) or nocardomycolic (44–60 carbons) acids. They may also contain diverse functional groups such as methoxy, keto, or epoxy ester group and cyclopropane ring (Fig.2). The structure, physiological function, and biosynthesis of mycolic acids have been reviewed by Barry et al. (1998).

Mycolic acids of the unsaturation and cyclopropane chains (cited from web page www.cyberlipid.org/fa/acid0006.htm)
3.1 Cord Factor: A Trehalose Diester from Mycobacteria
Trehalose lipids in mycobacteria have been extensively exploited in chemical composition (Mompon et al. 1978; Batrakov et al. 1981; Fujita et al. 2005). Of all trehalose lipids, cord factor is the best known. It has two mycolic acids of variable length, esterified to the 6-hydroxyl group of each glucose to give trehalose 6,6′-dimycolate (TMD) (Ryll et al. 2001) (Fig.3). TDM is the most prominent and best-studied mycolic acid-containing glycolipid of mycobacteria. The structure varies greatly among mycobacterial species, and the mycolyl moiety is related with toxicity and antigenicity, thereby constituting potential virulence or immunostimulating mechanisms (Ryll et al. 2001; Hunter et al. 2006; Guidry et al. 2007; Ishikawa et al. 2009).

Trehalose 6,6′-dimycolate(TDM ) from Isolated from M. tuberculosis
With MALDI-TOF mass spectrometry, cord factors were characterized by Fujita et al.(2005) from nine species of human-virulent and nonvirulent mycobacteria as follows, M. tuberculosis H37Rv (ATCC 27294), M. tuberculosis Aoyama B (ATCC 31726), M. bovis BCG Tokyo 172 (ATCC 35737), M. bovis BCG Connaught (ATCC 35745), Mycobacterium intracellulare serotype 4 (ATCC 35767), M. intracellulare serotype 16 (ATCC 13950), Mycobacterium kansasii (ATCC 12478), Mycobacterium phlei (ATCC 11758), and Mycobacterium flavescens (ATCC 14474). They found that α-mycolic acid was a ubiquitous component among the mycobacterial species, with carbon numbers ranging from C74 to C88 and with two cyclopropane rings or equivalent double bonds. Ketomycolates were also widely distributed, with carbon numbers ranging from C76 to C91, but the ranges of the carbon number of ketomycolates differed greatly among the species (Fujita et al. 2005).
3.2 Trehalose Lipids from Rhodococcus
The surface-active lipids in rhodococci have been reviewed by Lang S (Lang 1999). The biosurfactants produced by members of the genus Rhodococcus is dominated by trehalose-containing glycolipids. They are quite diverse in the fatty acid moiety, such as dicorynomycolates, monocorynomycolates, and 2, 2′, 3, 4-tetraester (Kim et al. 1990; Espuny et al. 1996; Rapp et al. 1979; Kretschmer et al. 1982; Kretschmer and Wagner 1983). Additionally, succinoyl-trehalose and octaacyl-trehalose have also been detected in R. erythropolis (Uchida et al. 1989; Singer and Finnerty 1990; Tokumoto et al. 2009).
3.2.1 Trehalose Diesters
3.2.1.1 Trehalose Dimycolates from R. erythropolis DSM 43215
The trehalose lipids produced by R. erythropolis DSM 43215 were characterized as α-d-Glucopyranosyl-α-d-glucopyranoside-6,6′-di-(2-alkyl-3-hydroxy)-carboxylic ester(Rapp et al. 1979; Kretschmer et al. 1982) (Fig.4). The bacterium was grown on 2% (w/v) n-alkanes (chain length C12–C18). The glycolipid was extracted from the biomass with n-hexane and purified by repeated chromatography on silica gel. NMR revealed that its lipid moiety was consisted of saturated long-chain α-branched β-hydroxy fatty acids (mycolic acids) ranging from C32H64O3 to C38H76O3, of which C34H38O3 and C35H70O3 predominated. Prior to this study and afterwards, similar trehalose dimycolates were found in Rhodococcus, Nocardia, and Rhodochrous grown on glycerol.

α, α-trehalose-6,6′-dicorynomycolates from R. erythropolis DSM 43215 (Rapp et al. 1979). The mycolic acids range from C32H64O3 to C38H76O3.
3.2.1.2 Novel Trehalose Dimycolate from Rhodococcus opacus 1CP
Glycolipids were purified from culture of R. opacus 1CP growing with n-decane as sole carbon source. They are characterized to be trehalose dimycolate with 1H-NMR spectroscopy. The fatty acids are nocardiomycolic acids of total chain lengths between 48 and 54 carbons (Fig.5). In addition, in each mycolic acid, two double bonds exist (Niescher et al. 2006). These features (longer chain and unsaturation) distinguish the trehalose dimycolates from those of R. erythropolis DSM43215 (Rapp et al. 1979; Kretschmer et al. 1982) and Rhodococcus ruber IEGM231 (Philp et al. 2002).

Structure of the trehalose dinocardiomycolate of R. opacus 1CP (Niescher et al. 2006). The mycolic acid components of carbon number ranging from C-48 to C-54 (n+x+y+z from 37 to 43)
3.2.1.3 Trehalose Lipids of Rhodococcus ruber IEGM231
Three kinds of glycolipids were purified from R. ruber culture, namely C1, C2, and C3. Each has analogs with varied chain lengths (Philp et al. 2002). Their structures were determined by NMR and ESI-MS (Fig.6). In common, they share trehalose as the hydrophilic moiety. Among them, C1 was a typical trehalose dicorynomycolate with chain α-branched-β-hydroxy fatty acids (the total carbon number ranging from 34 to 46, with C40 as a main component), which is longer than that in R. erythropolis DSM 43215 (Rapp et al. 1979). In the case of C2 component, the major component was equivalent to a molecule carrying two C16-fatty acid units and other analogs of the C11–C14 acids. The major C3 component is C14-glycolipids containing fatty acids of both the saturated and unsaturated.

Trehalose lipids of Rhodococcus ruber IEGM231 (Philp et al. 2002)
3.2.1.4 Succinoyl Trehalose Lipids from R. erythropolis SD-74
The bacterium strain SD-74 can abundantly produce extracellular trehalose lipids from n-alkanes. The characteristic of this trehalose lipid is succinic acid containing. Two main components named STL-1 and STL-2 have been characterized from culture of strain SD-74 (Uchida et al. 1989; Tokumoto et al. 2009). They are not only powerful surfactants, but they also show versatile biochemical actions, such as induction of human cell differentiation (Isoda et al. 1995; Sudo et al. 2000).
The STL structure is primarily determined by NMR and GC-MS. STL1 and STL2 were 2,3,4,2″-di-O-succinoyl-di-O-alkanoyl-α,α-trehalose and 2,3,4-mono-O-succinoyl-di-O-alkanoyl-α,α-trehalose, respectively(Uchida et al. 1989). This is the first report of succinoyl trehalose lipids (STLs) (Uchida et al. 1989).
Recently, the structure of STL1 produced by Rhodococcus sp. SD-74 was subjected to further characterization (Tokumoto et al. 2009). The exact structure of STL-1 was depictured on the basis of NMR, MALDI-TOF/MS, and GC-MS analyses. The STL-1 in this report was produced from n-hexadecane and was the main component. It was identified to be 3,4-di-O-alkanoyl-2-O-succinoyl-α-d-glucopyranosyl-2′-O-succinoyl-α-d-glucopyranoside (Fig.7). The major fatty acid of STL-1 was C16, indicating that n-hexadecane as the carbon source is possibly directly incorporated into the carbohydrate moiety via terminal oxidation.

Succinoyl trehalose lipids (STL-1) from R. erythropolis SD-74
3.2.1.5 Trehalose Diesters and Triesters of Mycobacteriurn fortuitum
M. fortuitum produced a family of trehalose-containing glycolipids named mycoside F (Gautier et al. 1992). Three main glycoconjugates were detected and their structures established as 2, 3-diacyl, 2, 3, 4- and 2, 3, 6-triacyl trehalose. The nature of the fatty acyl substituents identified primarily as 2-methyl-octadecen-2-oyl.
In another case, a trehalose-containing glycolipid was detected in several strains of M. fortuitum and characterized as 2,3-di-O-acyltrehalose (DAT) (Ariza et al. 1994). In the report, lipid constituents were identified as a mixture of straight-chain (14–18 carbon atoms) and methyl-branched-chain (17–21 carbon atoms) fatty acyl groups. DAT was further fractionated by reverse phase TLC into four fractions that were designated DAT- I to IV. DAT-I contained 70–75% straight-chain acyl substituents (hexadecanoyl and octadecanoyl predominating) and 25–30% 2-methyl branched substituents (mainly 2-methyl octadecadienoyl). DAT-II was composed of a mixture in which the acyl groups were almost exclusively 2-methyl branched, with 2-methyl octadecadienoyl and 2-methyl octadecen-2-oyl predominating. DAT-III, which was the major isolated fraction, consisted of compounds in which the ratio linear to branched acyl groups varied between 0.8 and 0.9, 2-methyl octadecen-2-oyl, hexadecanoyl and octadecanoyl being the most abundant. Finally, DAT-IV comprised a mixture of DAT molecules containing mostly 2-methyl octadecadienoyl, 2-methyl octadecen-2-oyl, 2-methyl eicosadienoyl, and 2-methyl eicosen-2-oyl groups. Actually, early in 1964, trehalose lipids of M. fortuitum had been described having hydrophobia moiety like above described, i.e., C16:0 and sometimes with C19:0 and C20:0 in fatty acids moiety (Vilkas and Rojas 1964).
3.2.2 Trehalose Tetraesters Produced by Rhodococcus and Related Bacteria
In this section, the structure of the trehalose lipids produced by Arthrobacter, Micrococcus and Rhodococcus is presented.
3.2.2.1 Arthrobacter sp. EK 1 (Passeri et al. 1991)
Arthrobacter sp. EK 1 is a marine n-alkane-utilizing bacterium. After purification by column and thick layer chromatography, the main fraction, an anionic 2,3,4,2′-trehalose tetraester, was obtained. The chain lengths of fatty acids ranged from 8 up to 14; furthermore, succinate was detected. The exact position of succinate is confirmed at C 2 atom of trehalose. This is the first report of succinate on trehalose of a glycolipid (Passeri et al. 1991).
3.2.2.2 Rhodococcus wratislaviensis BN38 (Tuleva et al. 2008)
Glycolipids were produced by R. wratislaviensis BN38 when grown on 2% n-hexadecane. The glycolipids were isolated by chromatography on silica gel columns, and the main product was characterized as 2,3,4,2′-trehalose tetraester with molecular mass of 876, esterified with two decanoic, one octanoic, and one succinic acid (Tuleva et al. 2008). Trace amounts of another 2,3,4,2′-trehalose tetraester of molecular mass 849 were also detected.
3.2.2.3 Micrococcus luteus BN56 (Tuleva et al. 2009)
The soil strain M. luteus BN56 can produce a mixture of two trehalose tetraester biosurfactants when grown aerobically on n-hexadecane. The biosurfactants were extracted and purified from whole cultures and characterized by NMR and mass spectrometry (Fig.8). The two major components consisted trehalose tetraesters with molecular mass of 876 and 848. In both cases, α,α-trehalose was linked at C-2 or C-4 with a succinic and at C-2′ with a decanoic acid. The trehalose lipid with molecular mass of 876 was esterified at C-2, C-3, or C-4 with an octanoic and a decanoic acid. The other one with MW 848, of lower molecular mass, was esterified at the same positions with two octanoic acids (Tuleva et al. 2009).

Chemical structure of trehalose tetraesters produced by M. luteus BN56 (n=6–11; R2=succinic acid) (Tuleva et al. 2009)
3.2.2.4 Rhodococcus erythropolis 51T7 (Marques et al. 2009)
The chemical structure of trehalose lipids produced by strain 51T7 was further studied by LC-MS, with tetradecane as the carbon source (Marques et al. 2009). This analysis revealed that this surfactant is a mixture of at least six components, with the pseudomolecular ions being between m/z 905 and 821 (using negative ion mode). The most abundant component is the pseudomolecular ion 876, occupying 43.9% of the total lipids after 72h in culture. This molecular weight may correspond to either trehalose–succinic acid–C9–C9–C10 or trehalose–succinic acid–C11–C10–C7. The remaining components had a lower and more constant concentration during growth. Worth to note, the trehalose tetraesters of R. erythropolis 51T7 differed with the results of a previous report in the main component (Espuny et al. 1996). The authors postulated that this may have resulted from variations in the carbon source used.
3.2.3 Octaacyltrehalose from Rhodococcus sp. H13-A
Growing with hexadecane, Rhodococcus sp. H13-A produced a mixture of trehalose lipids with one major and ten minor components(Singer et al. 1990). The components of hydrophobic tail are relatively complex, constituting of normal C10–C22 saturated and unsaturated fatty acids, C35–C40 mycolic acids, hexanedioic and dodecanedioic acids, and 10-methyl hexadecanoic and 10-methyl octadecanoic acids. The major glycolipid was identified as 2,3,4,6,2′,3′,4′,6′-octaacyltrehalose, plus minor components of di-, tetra- and hexa-acyltrehalose derivatives.
4 Physicochemical Property
Physicochemical properties of trehalose lipids have been mainly examined in the genus Rhodococcus while the glycolipids from mycobacteria are seldom investigated in this aspect due to their long hydrophobic moiety tightly attached on cell wall and limitations as a practical biosurfactant.
Recently, some trehalose lipids come to purification (Tuleva et al. 2008, 2009; Marques et al. 2009; Tokumoto et al. 2009; Peng et al. 2007; Niescher et al. 2006; Kretschmer et al. 1982; Kretschmer and Wagner 1983; Kim et al. 1990). Most of them showed strong surface activity by lowering water surface tension below 30mNm−1 (ranging from 19 to 43mNm−1), lowering the interfacial tension against hexadecane (decane or kerosene) to 5mNm−1, even <1mNm−1 (Table1), while the CMC values can reach 0.7mgl−1 in case of dicorynomycolates (Kretschmer and Wagner 1983), and the CMC values of most trehalose lipids varied from 0.7 to 37mgl−1. Examples are as follows.
The trehalose corynomycolates generated by R. erythropolis DSM43215 showed extremely low CMC in high-salinity solutions, and the interfacial properties were stabile in solutions with a wide range of pH and ionic strength(Kretschmer et al. 1982). The dicorynomycolates reduced interfacial tension from 44 to 18mNm−1 and are less sensitive to salt concentrations than synthesized surfactants, therefore of potential application in enhanced oil recovery.
Succinoyl trehalose lipids (STL) from R. erythropolis SD-74 are quite efficient to lower water surface tension. The estimated CMC and γCMC values for STL-1 were 5.6×10−6 M (equivalent to 5mgl−1) and 19.0mNm−1, and those for sodium salt (Na-STL-1) were 7.7×10−6 M and 23.7mNm−1, respectively. Thus, STL-1 holds an excellent surface activity at remarkably low concentrations (Tokumoto et al. 2009).
The glycolipid synthesized by Rhodococcus species H13-A (Singer et al. 1990) exhibited a CMC of 1.5mgl−1 and minimum interfacial tension value of 0.02mNm−1 against decane, even much lower in interfacial tension to 6×10−5mNm−1 in the presence of pentanol. In the case of the glycolipids purified from M. luteus BN56, they own CMC value of 25mgl−1 and can reduce the surface tension of water to 24.1mNm−1 and lower the interfacial tension against hexadecane to 1.7mNm−1. In addition, they have emulsion-stabilizing activity.
5 Biological Activity
Biosurfactants play an essential role in bacterial swarming motility and participate in cell signaling and differentiation as well as in biofilm formation. In addition, they can increase the bioavailability of hydrophobic substrates, trap heavy metals, and function as an antibiotic (Singh and Cameotra 2004; Rodrigues et al. 2006). Trehalose lipids accept the general merits being a biosurfactant. Moreover, some particular functions also exist, and worthy to be noted.
5.1 Trehalose Lipids in Mycobacteria
Trehalose lipids in mycobacteria draw attentions due to their role in pathogenicity and molecular immunology (Imasato et al. 1990; Ryll et al. 2001; Hunter et al. 2006; Ortiz et al. 2008, 2009; Zaragoza et al. 2009). TDM (cord factor) is the most abundant, granulomagenic, and toxic lipid in the cell surface of virulent M. tuberculosis(Hunter et al. 2006). TDM has been identified as the factor responsible for cord formation. It has been also detected in nontuberculous mycobacteria, including M. avium complex species (Fujita et al. 2005). In the mycobacterial cell envelope, together with arabinogalactan mycolate and trehalose monomycolate, TDM forms an integral part of the cell wall skeleton, resulting in highly hydrophobic cell surface properties and acid-fastness (Barry et al. 1998). In addition, it is believed that TDM is responsible for the low permeability of the membranes conferring appreciable drug resistance to the organisms.
On the other hand, TDM and its stereoisometric derivatives (TDCMs) were evidenced to inhibit tumor metastasis, and TDCMs are more potential to suppress tumor growth and inhibit tumor metastasis than TDM (Watanabe et al. 1999).
5.2 Trehalose Lipids from Rhodococcus
Beside their known industrial applications, trehalose lipids from Rhodococcus recently attracted attention due to their functions in cell membrane interaction and the prospects as a therapeutic agent (Isoda et al. 1995; Zaragoza et al. 2009, 2010; Aranda et al. 2007; Harland et al. 2009; Ortiz et al. 2008, 2009; Imasato et al. 1990).
-
The succinoyl trehalose lipid (STL-1) of R. erythropolis SD-74 shows effects on cell differentiation. Isoda et al. (1995) reported that STL-1 markedly inhibited the growth of a human monocytoid leukemic cell line and induced its morphological alteration along a monocyte-macrophage lineage. STL-1 markedly increased differentiation-associated characteristics in macrophage, such as phagocytic activities in U937. Furthermore, the U937 cells activated with STL-1 exhibited cytotoxic activity against human carcinoma cell line, while it has low cytotoxicity against normal human cells (Isoda et al. 1995).
-
Hemolytic activity of a succinoyl-containing trehalose lipid: the trehalose lipid produced by Rhodococcus sp. was observed to cause the swelling of human erythrocytes followed by hemolysis at concentrations well below its CMC value. It works by a colloid-osmotic mechanism, most likely by formation of enhanced permeability domains, or “pores” enriched in the biosurfactant, within the erythrocyte membrane (Zaragoza et al. 2010).
-
Interactions with phosphatidylethanolamine membrane: Ortiz et al. (2008) purified a trehalose lipid from Rhodococcus sp. and examined its effect on the thermotropic and structural properties of phosphatidylethanolamine membranes of different chain length and saturation. They find that the trehalose lipid presents good miscibility both in the gel and the liquid crystalline phases of the membrane and affects the gel to liquid crystalline phase transition. The trehalose lipid was suggested to incorporate into the membrane bilayers and produce structural perturbations, which might affect the function of the membrane. Similar conclusions were drawn based on the results with phosphatidylserine membranes (Ortiz et al. 2009):
-
Mechanism of membrane permeabilization by the STL (Zaragoza et al. 2009): The partition constant of the trehalose lipid to palmitoyloleoylphosphatidylcholine membranes indicates that trehalose lipid behaves as a weak detergent, which prefers membrane incorporation over micellization. Addition of the trehalose lipid to large unilamellar vesicles results in a size-selective leakage of entrapped solutes to the external medium. Further studies support that the trehalose lipid incorporates into phosphatidylcholine membranes and segregates within lateral domains, which may constitute membrane defects or “pores.”
-
Synthetic trehalose glycolipids confer desiccation resistance to supported lipid monolayers (Harland et al. 2009). Harland et al. (2009) presented the first synthetic trehalose glycolipids capable of providing desiccation protection to membranes of which they are constituents. They deduced that interactions between the trehalose headgroup and surrounding molecules are the determining factor in dehydration protection.
6 Biosynthesis
The biosynthesis pathways of biosurfactant have been extensively studied in rhamnolipids and lipopeptides, and related genes have been determined in some cases (Nakano et al. 1991, 1992; Ochsner et al. 1994; Campos-Garcia et al. 1998). However, the corresponding researches on trehalose lipids are hard to be retrieved from literatures.
With an aim to clarify the synthesis pathway of trehalose dicorynomycolates, Kretschmer and Wagner (1983) characterized biosynthetic intermediates of the trehalose lipids in R. erythropolis DSM 43215. Free corynomycolic acid intermediates were observed. The corynomycolic acid moiety and the trehalose moiety therefore are synthesized independently and are subsequently esterified. Trehalose is first esterified to form the monomycolate and then the dimycolate. Based on the intermediates, a biosynthetic pathway was suggested (Fig.9) (Kretschmer and Wagner 1983).

Trehalose dicorynomycolate synthesis pathway from n-tetradecane by Rhodococcus erythropolis DSM 43215 (Kretschmer and Wagner 1983)
7 Production
The production of most trehalose lipids is growth-associated, whereas production is also observed under growth-limiting conditions or by resting cells. Compared to rhamnolipids, the yield of trehalose lipids reported in the literature is lower, usually below 3gl−1, considering the production of purified products.
7.1 Substrates Used for Trehaloselipids Production
-
n-alkanes: In the case of oil degrading bacteria of Rhodococcus, n-alkanes are usually the optimal substrates for glycolipids production. Although bacteria of Rhodococcus can utilize alkanes of a wide length range, C14 and C16 alkanes seem to be the best for high yielding (Niescher et al. 2006), especially for the trehalose lipids with medium length chains(Tokumoto et al. 2009).
-
Nonalkanes: Trehalose dicorynomycolates have also been produced in the absence of n-alkanes or other lipophilic carbon sources by Brevibacterium vitarumen 12143 (Laneelle and Asselineau 1976), Corynebacterium diphtheriae (Adam et al. 1967), and different pathogenic Mycobacteriaceae (Asselineau and Asselineau 1978).
7.2 Cell Wall Association
The usual pattern of production of nonionic trehalose glycolipids is generally cell wall-associated (Suzuki et al. 1969; Kretschmer et al. 1982). Usually, the lipids of long-chain fatty acids tend to be cell wall-associated. Especially at early stage of cultivation, the products mainly attach to the cell wall.
Growing on n-alkanes as a sole source of carbon and energy, R. fascians produced both an extracellular and cell-bound surface-active mixture of trehalose lipids that reduced the surface tension of water to 32mNm (Yakimov et al. 1999). When R. erythropolis strain 51T7 was cultivated with both alkanes and waste lubricant oil as a sole carbon substrate, it produced extracellular glycolipids with surface activity. Rhodococcus sp. strain 51T7 produced trehalose lipids between 0.48 and 1.12gl−1(Espuny et al. 1996). However, in a previous report of this strain (named Coryneform bacterium 51T7 at first), trehalose lipids are associated mainly with the cell wall on incubation up to 80h. After 80h, the trehalose lipids are mainly extracellular (Martin et al. 1991).
7.3 Growth-Associated Production
Growth-associated production of trehalose lipids is popular (Table1), such as the production of a trehalose dicorynomycolate in R. erythropolis DSM43215 (Rapp et al. 1979). The production of a trehalose dicorynomycolate in R. opacus 1CP is also growth-associated (Niescher et al. 2006) (Fig.10).

Correlation of cell growth (protein), alkane consumption, and glycolipid formation of R. opacus 1CP (Niescher et al. 2006). Batch cultures (20ml), each containing mineral medium and 1% (w/v) n-tetradecane, were inoculated with identical amounts of glucose-grown R. opacus 1CP and incubated at 30°C with constant shaking (130rpm)
However, the formation of anionic trehalose ester by R. erythropolis DSM43215 seems to be uncoupled from growth and occurs in the stationary phase (Ristau and Wagner 1983; Kim et al. 1990). The growth-uncoupled production also was observed in a paraffin-oxidizing bacterium R. erythropolis cultivated in shake flasks on a mixture of C14 and C15 n-alkanes or kerosene. The growth-limiting conditions, such as nitrogen deficiency, caused the formation of α, α-trehalose-2,3,4,2′-tetraesters.
Contrarily, also being a trehalose 2,3,4,2′-tetraester, one of the Rhodococcus sp 51T7 is produced in growth-associated mode. When grown on hydrocarbon, cells were highly segmented and accumulated lipid granules in the cytoplasm. Under optimal concentrations of sodium nitrate, potassium phosphate and iron (2.5gl−1, 1.5gl−1 and 0.01gl−1, respectively), production was increased from 0.5 to 3gl−1 (Espuny et al. 1996).
References
Adam A, Senn M, Vilkas E, Lederer E (1967) Spectrométrie de masse de glycolipides. Eur J Biochem 2:460–468
Anderson RJ, Newman MS (1933) The chemistry of the lipids of tubercle bacilli: xxxiii. isolation of trehalose from the acetone-soluble fat of the human tubercle Bacillus. J Biol Chem 101:499–504
Aranda FJ, Teruel JA, Espuny MJ, Marques A, Manresa A, Palacios-Lidon E, Ortiz A (2007) Domain formation by a Rhodococcus sp. biosurfactant trehalose lipid incorporated into phosphatidylcholine membranes. Biochim Biophys Acta 1768:2596–2604
Ariza MA, Martin-Luengo F, Valero-Guillen PL (1994) A family of diacyltrehaloses isolated from Mycobacterium fortuitum. Microbiology 140(Pt 8):1989–1994
Asselineau C, Asselineau J (1978) Trehalose-containing glycolipids. Prog Chem Fats Other Lipids 16:59–99
Barry CE 3rd, Lee RE, Mdluli K, Sampson AE, Schroeder BG, Slayden RA, Yuan Y (1998) Mycolic acids: structure, biosynthesis and physiological functions. Prog Lipid Res 37:143–179
Batrakov SG, Bv R, Koronelli TV, Bergelson LD (1981) Two novel types of trehalose lipids. Chem Phys Lipids 29:241–266
Berekaa MM, Steinbuchel A (2000) Microbial degradation of the multiply branched alkane 2, 6, 10, 15, 19, 23-hexamethyltetracosane (Squalane) by Mycobacterium fortuitum and Mycobacterium ratisbonense. Appl Environ Microbiol 66:4462–4467
Besra GS, Bolton RC, McNeil MR, Ridell M, Simpson KE, Glushka J, van Halbeek H, Brennan PJ, Minnikin DE (1992) Structural elucidation of a novel family of acyltrehaloses from Mycobacterium tuberculosis. Biochemistry 31:9832–9837
Campos-Garcia J, Caro AD, Najera R, Miller-Maier RM, Al-Tahhan RA, Soberon-Chavez G (1998) The Pseudomonas aeruginosa rhlG gene encodes an NADPH-dependent beta-ketoacyl reductase which is specifically involved in rhamnolipid synthesis. J Bacteriol 180:4442–4451
Churchill PF, Morgan AC, Kitchens E (2008) Characterization of a pyrene-degrading Mycobacterium sp. strain CH-2. J Environ Sci Health B 43:698–706
Datta AK, Takayama K (1993) Isolation and purification of trehalose 6-mono- and 6, 6′-di-corynomycolates from Corynebacterium matruchotii. Structural characterization by 1H NMR. Carbohydr Res 245:151–158
Desai AJ, Patel KM, Desai JD (1988) Emulsifier production by Pseudomonas fluorescens during the growth on hydrocarbons. Current Science Association, Bangalore
Esch SW, Morton MD, Williams TD, Buller CS (1999) A novel trisaccharide glycolipid biosurfactant containing trehalose bears ester-linked hexanoate, succinate, and acyloxyacyl moieties: NMR and MS characterization of the underivatized structure. Carbohydr Res 319:112–123
Espuny MJ, Egido S, Mercadé ME, Manresa A (1995) Characterization of trehalose tetraester produced by a waste lubricant oil degrader Rhodococcus sp. Toxicol Environ Chem 48:83–88
Espuny M, Egido S, Rodón I, Manresa A, Mercadé M (1996) Nutritional requirements of a biosurfactant producing strain Rhodococcus sp 51T7. Biotechnol Lett 18:521–526
Fujita Y, Naka T, McNeil MR, Yano I (2005) Intact molecular characterization of cord factor (trehalose 6, 6′-dimycolate) from nine species of mycobacteria by MALDI-TOF mass spectrometry. Microbiology 151:3403–3416
Gautier N, Lopez Marin LM, Laneelle MA, Daffe M (1992) Structure of mycoside F, a family of trehalose-containing glycolipids of Mycobacterium fortuitum. FEMS Microbiol Lett 77:81–87
Gesheva V, Stackebrandt E, Vasileva-Tonkova E (2010) Biosurfactant production by halotolerant Rhodococcus fascians from Casey Station, Wilkes Land, Antarctica. Cur Microbiol 61(2):112–117
Guidry TV, Hunter RL Jr, Actor JK (2007) Mycobacterial glycolipid trehalose 6, 6′-dimycolate-induced hypersensitive granulomas: contribution of CD4+ lymphocytes. Microbiology 153:3360–3369
Hallas LE, Vestal JR (1978) The growth of Mycobacterium convolutum on solid n-alkane substrates: effect on cellular lipid composition. Can J Microbiol 24:1197–1203
Harland CW, Botyanszki Z, Rabuka D, Bertozzi CR, Parthasarathy R (2009) Synthetic trehalose glycolipids confer desiccation resistance to supported lipid monolayers. Langmuir 25:5193–5198
Hunter RL, Olsen M, Jagannath C, Actor JK (2006) Trehalose 6, 6′-dimycolate and lipid in the pathogenesis of caseating granulomas of tuberculosis in mice. Am J Pathol 168:1249–1261
Imasato H, Procopio J, Tabak M, Ioneda T (1990) Effect of low mole fraction of trehalose dicorynomycolate from Corynebacterium diphtheriae on water permeability and electrical capacitance of lipid bilayer membranes. Chem Phys Lipids 52:259–262
Ishikawa E, Ishikawa T, Morita YS, Toyonaga K, Yamada H, Takeuchi O, Kinoshita T, Akira S, Yoshikai Y, Yamasaki S (2009) Direct recognition of the mycobacterial glycolipid, trehalose dimycolate, by C-type lectin Mincle. J Exp Med 206:2879–2888
Isoda H, Shinmoto H, Matsumura M, Nakahara T (1995) Succinoyl trehalose lipid induced differentiation of human monocytoid leukemic cell line U937 into monocyte-macrophages. Cytotechnology 19:79–88
Kim JS, Powalla M, Lang S, Wagner F, Lunsdorf H, Wray V (1990) Microbial glycolipid production under nitrogen limitation and resting cell conditions. J Biotechnol 13:257–266
Kotani T, Kawashima Y, Yurimoto H, Kato N, Sakai Y (2006) Gene structure and regulation of alkane monooxygenases in propane-utilizing Mycobacterium sp. TY-6 and Pseudonocardia sp. TY-7. J Biosci Bioeng 102:184–192
Kretschmer A, Wagner F (1983) Characterization of biosynthetic intermediates of trehalose dicorynomycolates from Rhodococcus erythropolis grown on n-alkanes. Biochim Biophys Acta/Lipids Lipid Metabol 753:306–313
Kretschmer A, Bock H, Wagner F (1982) Chemical and physical characterization of interfacial-active lipids from Rhodococcus erythropolis grown on n-alkanes. Appl Environ Microbiol 44:864–870
Laneelle MA, Asselineau J (1976) Glycolipids of Brevibacterium vitarumen. Biochim Biophys Acta 486:205–208
Lang S (1999) Production of microbial glycolipids. In: Bucke C (ed) Methods in Biotechnology, Vol. 10, Carbohydrate biotechnology protocols. Humana Press Inc., Totowa, NJ, pp 103–118
Lang S, Philp JC (1998) Surface-active lipids in rhodococci. Antonie Leeuwenhoek 74:59–70
Marques AM, Pinazo A, Farfan M, Aranda FJ, Teruel JA, Ortiz A, Manresa A, Espuny MJ (2009) The physicochemical properties and chemical composition of trehalose lipids produced by Rhodococcus erythropolis 51T7. Chem Phys Lipids 158:110–117
Martin M, Bosch P, Parra JL, Espuny MJ, Virgili A (1991) Structure and bioconversion of trehalose lipids. Carbohydr Res 220:93–100
Mompon B, Federici C, Toubiana R, Lederer E (1978) Isolation and structural determination of a “cord-factor” (trehalose 6, 6′ dimycolate) from Mycobacterium smegmatis. Chem Phys Lipids 21:97–101
Nakano MM, Magnuson R, Myers A, Curry J, Grossman AD, Zuber P (1991) srfA is an operon required for surfactin production, competence development, and efficient sporulation in Bacillus subtilis. J Bacteriol 173:1770–1778
Nakano MM, Corbell N, Besson J, Zuber P (1992) Isolation and characterization of sfp: a gene that functions in the production of the lipopeptide biosurfactant, surfactin, in Bacillus subtilis. Mol Gen Genet 232:313–321
Niescher S, Wray V, Lang S, Kaschabek SR, Schlomann M (2006) Identification and structural characterisation of novel trehalose dinocardiomycolates from n-alkane-grown Rhodococcus opacus 1CP. Appl Microbiol Biotechnol 70:605–611
Ochsner UA, Koch AK, Fiechter A, Reiser J (1994) Isolation and characterization of a regulatory gene affecting rhamnolipid biosurfactant synthesis in Pseudomonas aeruginosa. J Bacteriol 176:2044–2054
Ortiz A, Teruel JA, Espuny MJ, Marques A, Manresa A, Aranda FJ (2008) Interactions of a Rhodococcus sp. biosurfactant trehalose lipid with phosphatidylethanolamine membranes. Biochim Biophys Acta 1778:2806–2813
Ortiz A, Teruel JA, Espuny MJ, Marques A, Manresa A, Aranda FJ (2009) Interactions of a bacterial biosurfactant trehalose lipid with phosphatidylserine membranes. Chem Phys Lipids 158:46–53
Passeri A, Lang S, Wagner F, Wray V (1991) Marine biosurfactants, II. Production and characterization of an anionic trehalose tetraester from the marine bacterium Arthrobacter sp. EK 1. Z Naturforsch C 46:204–209
Peng F, Liu Z, Wang L, Shao Z (2007) An oil-degrading bacterium: Rhodococcus erythropolis strain 3C-9 and its biosurfactants. J Appl Microbiol 102:1603–1611
Philp JC, Kuyukina MS, Ivshina IB, Dunbar SA, Christofi N, Lang S, Wray V (2002) Alkanotrophic Rhodococcus ruber as a biosurfactant producer. Appl Microbiol Biotechnol 59:318–324
Rapp P, Bock H, Wray V, Wagner F (1979) Formation, Isolation and characterization of trehalose dimycolates from Rhodococcus erythropolis Grown on n-Alkanes. J Gen Microbiol 115:491–503
Ristau E, Wagner F (1983) Formation of novel anionic trehalosetetraesters from Rhodococcus erythropolis under growth limiting conditions. Biotechnol Lett 5:95–100
Rodrigues L, Banat IM, Teixeira J, Oliveira R (2006) Biosurfactants: potential applications in medicine. J Antimicrob Chemother 57:609–618
Ryll R, Kumazawa Y, Yano I (2001) Immunological properties of trehalose dimycolate (cord factor) and other mycolic acid-containing glycolipids – a review. Microbiol Immunol 45:801–811
Satpute SK, Banat IM, Dhakephalkar PK, Banpurkar AG, Chopade BA (2010) Biosurfactants, bioemulsifiers and exopolysaccharides from marine microorganisms. Biotechnol Adv 28(4):436–450
Schulz D, Passeri A, Schmidt M, Lang S, Wagner F, Wray V, Gunkel W (1991) Marine biosurfactants. I. Screening for biosurfactants among crude oil degrading marine microorganisms from the North Sea. Z Naturforsch C 46:197–203
Singer V, Finnerty WR (1990) Physiology of biosurfactant synthesis by Rhodococcus species H13-A. Can J Microbiol 36:741–745
Singer V, Finnerty WR, Tunelid A (1990) Physical and chemical properties of a biosurfactant synthesized by Rhodococcus species H13-A. Can J Microbiol 36:746–750
Singh P, Cameotra SS (2004) Potential applications of microbial surfactants in biomedical sciences. Trends Biotechnol 22:142–146
Sudo T, Zhao X, Wakamatsu Y, Shibahara M, Nomura N, Nakahara T, Suzuki A, Kobayashi Y, Jin C, Murata T, Yokoyama KK (2000) Induction of the differentiation of human HL-60 promyelocytic leukemia cell line by succinoyl trehalose lipids. Cytotechnology 33:259–264
Suzuki T, Tanaka K, Matsubara J, Kimoshita S (1969) Trehalose lipid and α-branched-β-hydroxy fatty acid formed by bacteria grown on n-alkanes. Agric Biol Chem 33:1619–1627
Tokumoto Y, Nomura N, Uchiyama H, Imura T, Morita T, Fukuoka T, Kitamoto D (2009) Structural characterization and surface-active properties of a succinoyl trehalose lipid produced by Rhodococcus sp. SD-74. J Oleo Sci 58:97–102
Tuleva B, Christova N, Cohen R, Stoev G, Stoineva I (2008) Production and structural elucidation of trehalose tetraesters (biosurfactants) from a novel alkanothrophic Rhodococcus wratislaviensis strain. J Appl Microbiol 104:1703–1710
Tuleva B, Christova N, Cohen R, Antonova D, Todorov T, Stoineva I (2009) Isolation and characterization of trehalose tetraester biosurfactants from a soil strain Micrococcus luteus BN56. Process Biochem 44:135–141
Uchida Y, Tsuchiya R, Chino M, Hirano J, Tabuchi T (1989) Extracellular accumulation of mono- and di-succinoyl trehalose lipids by a strain of Rhodococcus erythropolis grown on n-alkanes. Agric Biol Chem 53:757–763
Vila J, Grifoll M (2009) Actions of Mycobacterium sp. strain AP1 on the saturated- and aromatic-hydrocarbon fractions of fuel oil in a marine medium. Appl Environ Microbiol 75:6232–6239
Vilkas E, Rojas A (1964) On the Lipids of Mycobacterium Fortuitum. Bull Soc Chim Biol (Paris) 46:689–701
Watanabe R, Yoo YC, Hata K, Mitobe M, Koike Y, Nishizawa M, Garcia DM, Nobuchi Y, Imagawa H, Yamada H, Azuma I (1999) Inhibitory effect of trehalose dimycolate (TDM) and its stereoisometric derivatives, trehalose dicorynomycolates (TDCMs), with low toxicity on lung metastasis of tumour cells in mice. Vaccine 17:1484–1492
Yakimov MM, Giuliano L, Bruni V, Scarfi S, Golyshin PN (1999) Characterization of Antarctic hydrocarbon-degrading bacteria capable of producing bioemulsifiers. New Microbiol 22:249–256
Zaragoza A, Aranda FJ, Espuny MJ, Teruel JA, Marques A, Manresa A, Ortiz A (2009) Mechanism of membrane permeabilization by a bacterial trehalose lipid biosurfactant produced by Rhodococcus sp. Langmuir 25:7892–7898
Zaragoza A, Aranda FJ, Espuny MJ, Teruel JA, Marques A, Manresa A, Ortiz A (2010) Hemolytic activity of a bacterial trehalose lipid biosurfactant produced by Rhodococcus sp.: evidence for a colloid-osmotic mechanism. Langmuir 26(11):8567–8572
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2011 Springer-Verlag Berlin Heidelberg
About this chapter
Cite this chapter
Shao, Z. (2011). Trehalolipids. In: Soberón-Chávez, G. (eds) Biosurfactants. Microbiology Monographs, vol 20. Springer, Berlin, Heidelberg. https://doi.org/10.1007/978-3-642-14490-5_5
Download citation
DOI: https://doi.org/10.1007/978-3-642-14490-5_5
Published:
Publisher Name: Springer, Berlin, Heidelberg
Print ISBN: 978-3-642-14489-9
Online ISBN: 978-3-642-14490-5
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)