
Abstract
The Rupestrian Grassland is a vegetational complex with grassy to shrubby formations that occur throughout the high mountains of Brazil, usually formed by structurally resistant rocks, little affected by late tectonics, and strongly eroded and weathered under long term geological stability. RGC is closely associated with high altitude landsurfaces, in which several factors have a determinant role: (1) extreme oligotrophy, and acid, nutrient-depleted parent materials; (2) resistance to weathering and erosion (chemical and physical); (3) constant wind exposure; (4) intense fire regime. Variations of RG phytophysiognomies are basically due to soil depth (edaphic factor), drainage and landscape stability and evolution. Landforms (geomorphological attributes) affect the RGC at continental scales (high landsurfaces), regional scales (regional landforms, such as escarpments, valleys, slopes) and local scales (soil depth, stoniness, rockiness, drainage). The most common occurrence of RGC in Brazil is on Quartzite and metarenites, followed by canga and other Fe-rich substrates, igneous rocks and metamorphics, hence displaying a high diversity of substrates, with a major trait of extreme soil oligotrophy and acidity, and crucial variations in soil depth. The occurrence of well-documented areas of RG on Granitic and gneissic terrains imply that even richer rocks, submitted to long term weathering and erosion, can lead to similar soils on Highlands, where rock outcrops are also common (e.g. Caparaó, Itatiaia, Brigadeiro, Serra dos Órgãos). RGC can occur immersed in different domains (Atlantic Forest, Caatinga, Cerrado), regardless of present day climates, since it represents an edaphic climax of long-term development. The widespread distribution of comparable RGC, from Amazonia (Carajás, Roraima, Pacaás Novos, Cachimbo) to the Central Plateau (Santa Barbara, Ricardo Franco, Pirineus) and Northeastern and Southeastern Brazil (Caparaó, Espinhaço, Sincorá, Jacobina, Itatiaia, Serra dos Órgãos, Brigadeiro, Carangola) raises the unresolved question of phylogenetic ancestry, age and similarities (floristic, structural) between those isolated islands of Rupestrian vegetation. These aspects are central to the evolution of Brazilian Biomes, representing key issues to resolve the late Quaternary Refuge Theory controversy, and test the validity of island biogeographical isolation theories.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
1 The Geological Control
The rupestrian grassland constitute a singular kingdom of plants that form an ancient vegetation complex (rupestrian grassland complex—RGC). In Brazil, rupestrian grassland refers to rupicolous or rupestrian (rocky habitat) vegetation that grows on a range of geologies, including quartzite–sandstone (Harley and Simmons 1986), granite-gneiss (Queiroz et al. 1996) and itabirite—an iron-rich metal sedimentary rock (Jacobi and Carmo 2011; Jacobi et al. 2007; Vincent and Meguro 2008; Schaefer et al. 2009). Semir (1991) considered rupestrian grassland to include any open vegetation type associated with rocky outcrops. Veloso et al. (1991) classified it as “vegetation refuges or relic vegetation types” floristically different from the dominant surrounding flora. Bucher et al. (2007) proposed that such areas have acted as refuges during dry climate phases associated with major glaciations, as well as centers of recent speciation.
Rupestrian grassland is widely distributed in Brazil (Fig. 2.1), and forms a complex of sclerophyllous vegetation ranging from open rupestrian grassland strictu sensu, to denser physiognomies, varying from shrubs to low forests and thickets. In terms of vegetation life forms, RGC is rich in geophytes (bulbous plants), ericoids (heath-like shrubs), proteoids (protea shrubby with large leaves) and restiods (reed-like plants), acaulescent rossettes, all forming a grassy to shrubby physiognomy. The extreme oligothrophy, fire regime and wind exposure induce a great number of r-strategists, adapted to multiple environmental pressures (Grime 2001; Alves et al. 2014). The RGC are mainly associated with pre-Cambrian quartzites and metarenites, but important disjunct areas are also found on many contrasting lithologies (granites, gneisses, schists, syenites, metapelites, itabirites), all of which representing the most resistant rocks in terms of chemical and physical conditions. A general map of such distribution in relation to parent material (geology) is outlined in Fig. 2.2. There, the majority of RGC is developed on quartzite, and is clearly associated with the old Mobile Belts of Proterozoic age, as well as minor areas of younger plutonic rocks. Though tectonically stable today, these areas are the “roots” of ancient mountain ranges former of greater development, during epochs of tectonic collision between South American and African Plates. Hence, the mountains associated with RGC are very old, residual cores of strongly weathered, folded, faulted and eroded landmasses, later subjected to regional uplift during renewed tectonic events of lower magnitude. Therefore, the diversified landforms of Proterozoic rock terrains are the result of former tectonic activity and mountain-building processes, creating strong regional heterogeneity at landscape scale that remains to this time.

The distribution of rupestrian grassland complex in relation to the geological substrate and Brazilian structural provinces. The main rocks are grouped for facilitating the comprehension of the lithological control. Areas of mixed and variable rock types form lithological associations. The numbers represent: 1 Tepuis, 2 Tunuí, 3 Serra dos Pacaás Novos, 4 Serra Ricardo Franco, 5 Santa Bárbara, 6 Serra do Cachimbo, 7 Serra da Seringa, 8 Serra dos Carajás, 9 Morro do Chapéu, 10 Chapada Diamantina (Serra do Sincorá), 11 Northern Espinhaço, 12 Southern Espinhaço, 13 Highlands of Central Plateau, 14 Pontões de Santo Antônio do Jacinto, 15 Serra do Cipó, 16 Iron Quadrangle, 17 Caparaó, 18 Araxá, 19 Canastra, 20 Itatiaia, 21 Serra dos Órgãos, 22 Macaé, 23 Ibitipoca. In the Appendix I, all pictures are related to the different areas of RGC throughout the Brazilian territory, as indicated by the number

The overlay between the distribution of rupestrian grassland complex (RGC) in function of the geological era on which it developed. The main areas mapped are associated with Pre-cambrian rocks, ranging from neoarchean (Carajás, for example) to meso- and neoproterozoic. All these areas have a common attribute of very poor, acid quartzite/camglomerate/metapelite substrates, upon which the soils formed are, by extension, acid, nutrient depleted, shallow and extremely weathered/eroded. It is remarkable that the central parts of the amazon and São Francisco cratons are practically devoid of significant RGC
The predominance of quartzite in RGC is based on the fact that these rocks combine all prerequisites for RGC development: (1) extreme oligotrophy; (2) chemical resistance to weathering; (3) Physical resistance; (4) High mountain occurrence due to differential erosion; and (5) widespread distribution across the Brazilian Mobile Belts (Schaefer 2013), or the Ancient Brasiliano Orogenic Systems (Alkmim 2015), of Neoproterozoic age. In addition, these highlands are very susceptible to frequent, severe fire regimes, so that most plants are fire-adapted and tolerant.
The main attribute for controlling the RGC occurrence is rock resistance and chemical impoverishment. Whenever a given rock is weathered and form deeper soil, other types of vegetation will develop, so that the RGC is, in fact, an edaphic climax related to resistant lithologies under a certain set of environmental conditions. They always occur in rocky terrains, which form resistant masses in the mid of deeply weathered saprolites. In the case of granites, magmatites or syenites, these massive rocks are more resistant to weathering and erosion than the surrounding gneiss, so that they stand out as protruding masses known as sugar loaves (type of inselbergs), with very steep topography (Fig. 2.3) (Martinelli 1989). In this case, the massivity of rock combined with the style and degree os fracturing, are additional factors controlling RGC formation.

The rupestrian grassland complex (RGC) with its physiognomic variations on granitic rocks, as illustrated by the high surfaces at o caparaó massif, the highest mountains of southeastern Brazil. Traditionally viewed as “Campos de Altitude” these RGC are, in fact, true rupestrian in many parts, where rocky surfaces predominate. There, subtle variations in soils depth, drainage and organic matter accumulation lead to varied niches, where a set of well-adapted, R-strategists, coexist. In some areas with rocky fields, low lying rocky terrains allow the adaptations of shrubby with xeromorphic adaptations, whereas rocky pools are associated with water accumulations and cyanobacteria mats and mosses growth, followed by abundant cyperaceae, xyridaceae and eriocaulaceae. rupiculous formation are found on rock outcrops with large blocks forming inselbergs, where lichens, mosses, bromeliads and shrubs coexist, with dissolution pathways (canelures) formed by the constant input of water saturated by organic acids. Biodiversity and endemism are characteristic of these isolated mountains, surrounded by a matrix of rainforest, where most rupestrian species are completely absent. (Drawing by C. Schaefer)
The rugged mountainous relief of RGC provides a complex and varied combination of substrates, slope, aspect, altitude and edaphic condition, jointly promoting species diversification through niche specialization. The island-like distribution of RGC in Brazil points to a gene flow cut-off, from surrounding forested areas, keeping its singular floristic identity, by long-term biogeographic isolation (Fig. 2.1).
2 Landforms and Geomorphological Control
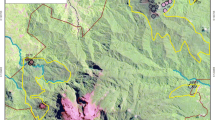
Looking at a general digital elevation model (topographic relief) of Brazil, it is clearly seen a close relation between the highlands above 1000 meters and the distribution of RGC (Fig. 2.4). In these highest landscape segments, rock outcrops are common, related to stripping of the weathering mantle and exhumation of resistant rock cores, exposed by differential erosion (Schaefer 2013). This process is variable in scale and depends on climate (and paleoclimate) conditions responsible for the balance between weathering, soil formation and erosion. At a local scale, three examples are offered here, to illustrate the variability of landforms in the RGC at local scale. In the first example, the Ibitipoca Mountains of Minas Gerais, RGC is found on quartzite, and minor variation on shallow soils on schists (Fig. 2.5), which favor forest vegetation in deeper soils (Dias et al. 2002). In another example, at the Serra do Cipó, western border of the Espinhaço Range, quartzite on the high plateau is closely associated with RGC with may different physiognomies (Fig. 2.6). Soils are invariably nutrient–poor, even though shallow and poorly developed. This apparent paradox is due to the extreme chemical poverty of the substrates, which have little nutrient to offer following exposure, weathering and soil formation, since these substrates are virtually depleted in most macronutrients (Ca, P, Mg, K and S) and micronutrients (Cu, Mn, Zn, B and Mo) (Schaefer 2013), as well as pre-weathered by long term evolution under tropical conditions (Silveira et al. 2016). A closer look on the soil aspect of RGC will be found in the next section.

The overlay of rupestrian grassland complex (RGC) distribution in Brazil in relation to topography. The close coincidence of highlands above 1000 m and RGC is striking. Most highlands are formed by residual, resistant tocks, rather than tectonically displaced mountains

A cross-section of the geology, landforms and soils on the rupestrian grassland complex of the Ibitipoca state park. In the second picture, a view from the Ibitipoca peak to the Pião peak (Background), showing the rupestrian complex as influenced by soil properties (depth, drainage, sandy texture)

Block-diagram of the western slopes of Serra do Cipó (Quartzite of Espinhaço Range), showing the rupestrian grassland on quartzites of the upper structural plateau, forming a topographical/vegetacional gradient with the São Francisco depression, to the left of the picture. The Espinhaço range form a mountains chain of residual, structural nature, by the extreme resistance of its quartzitic substrate. In the right picture, a sequence of soils at local scale is shown, as related to depth and drainage, affecting the distribution of open and shrubby physiognomies
Fire is an integral part of RGC ecosystems. It plays a key role in RGC development and survival of plants since it controls the cycles of destruction, regeneration, maturation and reproduction, hence having a major influence in the selective pressure, that allowed the enormous diversity to emerge in rupestrian grassland.
3 Soils, Landforms and Rupestrian Grassland Vegetation on Different Lithologies in Brazil
Together with geology and landforms, the soil factor plays a key role in controlling the physiognomies of rupestrian grasslands, as further described and discussed in Chap. 3. In this section, we describe the combined landscape attributes (soils, landforms, vegetation, geology) and their relationships, in four case studies representing the main types of rupestrian grasslands, and associated plant communities: (1) the canga vegetation on ironstone from Carajás, Amazonia; (2) The Ferruginous Rupestrian Complex on itabirites from the Iron Quadrangle (Moeda plateau) (3) the rupestrian metapellite to metarenite from the semiarid Serra Central State Park; and (4) the rupestrian grasslands on quartzite from the Espinhaço Mountains in Sempre-Vivas National Park.
3.1 Ferruginous Rupestrian Grasslands
The ferruginous rupestrian grasslands are herbaceous and shrubby physiognomy, associated with outcrops of Fe-rich rocks (itabirites/jaspelites), usually formed from the decomposition of ferruginous substrates (Schaefer et al. 2015). The ferruginous rupestrian grasslands are included in the open vegetation types associated with rocky outcrops (Rizzini 1979; Jacobi and Carmo 2011), called Rocky Complex by Semir (1991). Veloso et al. (1991) classifies such vegetation as refuges or relics, floristically different from the surrounding dominant flora. The vegetation associated with ferruginous rock outcrops was also called Rocky Fields over canga (Morellato and Rosa 1991; Silva 1992). In Brazil, they occur mainly as open vegetation islands in the region of the Iron Quadrangle (Minas Gerais) and in the Serra dos Carajás (Pará State) (Figs. 2.6, 2.7). In both cases, the occurrence of peculiar soils reveal an extraordinary adaptation of plants to an extremely restrictive edaphic scenario, both physically and chemically. From a climate and landscape viewpoint, the two regions are very different. Hence, the Amazonian Serra dos Carajás represent a very useful soil system for comparison to the canga soils of Iron Quadrangle, all extremely weathered rocky substrates.

Block-diagram of the Carajás plateau (Serra Sul) associated with ferrugineous rupestrian grasslands on the top surfaces, where ironstone (canga) is developed on Petric plinthosols. In the right picture, a sequence of soils at local scale is shown, as related to depth and drainage, affecting the distribution of open and shrubby physiognomies in these high landsurfaces according to topography
The Iron Quadrangle is a mountainous region located in the southern portion of the Espinhaço Range. The Iron Quadrangle possesses a structural landform, heavily controlled by the resistance of quartzite and itabirite rocks that form the “frame” of the quadrangular highland region (Fig. 2.8). Because of its mineral wealth, extraction of mineral assets has taken place since the colonial period, when towns and cities emerged, as Mariana, Ouro Preto, Congonhas, Sabara, Nova Lima, among others. Currently, some studies have hypothesized an increase in mining activity and urbanization, exerting different pressures on remaining canga ecosystems (Jacobi et al. 2007; Carmo and Jacobi 2012; Diniz et al. 2014; Nunes et al. 2015). However, no quantification of these pressures (Jacobi et al. 2011), nor a realistic picture of the extent of current remaining areas exist.

Distribution of rupestrian formations in the iron quadrangle (Quadrilatero Ferrífero) as predicted from automatic classification using high resolution RapidEye and IKONOS imaging
In this mountainous region, the substrates rich in iron show the development of Rocky Fields, also called Ferruginous Rocky Fields, or “canga” vegetation (Vincent 2004; Viana and Lombardi 2007). Such ecosystems are distributed in the higher plateaus and in the mountain crests, mainly between 900–1800 m, on rocky substrates or ferruginous laterite. They are colonized by plants well adapted to oligotrophic environments, able to tolerate a many limiting environmental factors such as shallow depth soils, drought, low fertility, high concentration of oxidized iron, low water retention, marked daily temperature ranges, incidence frequent fire, high sun exposure and constant winds (Santos and Varajão 2004; Vincent 2004; Jacobi et al. 2007). Hence, the vegetation reveals numerous anatomical, morphological, physiological and reproductive adaptations that enable their survival in this harsh environment (Silveira et al. 2016; Chap. 11).
Based on the geological mapping available (Schobbenhaus and Brito-Neves 2003), an approximate total of 700,000 ha of Iron Quadrangle, approximately 90,800 ha (13 % of the area) are made up of iron formation. These formations have generally shallow soils, such as Litholic Neosols, Cambisols and Petric Plinthosols, with a complex of rupestrian grasslands vegetation, with savanna to montane forest formations, the latter in the pockets of Oxisols.
Although there is a relative knowledge about the flora found in the iron formations of the Iron Quadrangle, revealing its rich diversity, presence of endemic and endangered species (Porto and Silva 1989; Mendonça and Lins 2000; Vincent 2004; Jacobi et al. 2007; Mourão and Stehmann 2007; Viana and Lombardi 2007), there is still a lack of information about associated soils, due to its unsuitability to cultivation and extreme poverty. However, soil studies in the Iron Quadrangle can be very useful to developing tailored techniques for regeneration of native vegetation in mining, degraded areas by mining activities.
In the following sections, we present some of the main soils found in the canga environments of the Carajás and Iron Quadrangle, emphasizing its morphological chemical, physical and mineralogical attributes, and ecological relationships with the ferruginous rupestrian grasslands.
4 Canga: A Definition
The crusts of lateritic canga, or ferruginous “cuirasse” (ferricrete), comprise the surface formations resulting from processes associated with changes in landscape and relief, directly linked to polycyclic weathering and pedogenesis (Schaefer et al. 2015). Superficial formations on ironstone, in most cases, can be classified as soils originated through sub-aerial changes in banded–iron formations, and exerting control in the distribution of vegetation.
The dynamics of formation of the ferricrete or ferruginous “cuirasse” at landscape level is an ongoing process, and strongly conditioned by Quaternary climatic variations, as illustrated in a hypothetical cycle in Fig. 2.9, starting with a dry phase and culminating in a wet phase. During the wet stages the canga degrades to produce a concretionary oxisol, with greater or lesser depth depending on the intensity of pedobioturbation, promoted by soil micro and mesofauna, especially termites and ants. This phenomenon promotes the selection and uptake of finer particles (clays and silts), which accumulates on the surface by biological turnover (Schaefer 2001). In the wet-dry climatic transitions, or vice versa, seasonality increases, and fire starts to rule the vegetation dynamics selecting resistant and/or tolerant species. Erosion losses increase, and the friable soil produced in the wet phase is markedly lost. In drier climate phases, there are outcrops of cemented canga (petroplinthite), where shallow soils prevail, with a litoplintic contact. This cycle can occur several times over the geological time, with variable duration of each phase without reaching a steady-state situation (equilibrium).

Hypothetical cycle of canga formation and association with ferruginous rupestrian grassland evolution during dry and wet phases in the late quaternary (based on Schaefer et al. 2015). In the boxes, the most characteristic features of each phase are presented
The canga formation is revealed by vertical and lateral movements of soluble and suspended products of weathering, responsible for the formation of soil horizons (Tardy and Nahon 1985). The alteration mantle on ferriferous itabiritic consists of a thick saprolite (30–300 m), wich forms the commercial interest ore. It is composed of iron oxy-hydroxides, kaolinite, gibbsite and eventually quartz, all alteration products of itabiritic rocks. In the upper part of the canga (petroplinthite), the degradation of the laterite is marked by the presence of pisolitic canga, with underground cavities, filled by small spherical aggregates (pisolites), composed by Fe and Al oxides, and several generations of coatings forming a typical coalesced hardened structure, with cracks, biological channels and tubules, with small laterite fragments and grains cemented by hematite/goethite (Schaefer et al. 2004).
These features are formed in acidic conditions and seasonal reduction, probably due to the accumulation of organic matter and water at the surface, which can cause dissolution of kaolinite, gibbsite and some of goethite, with remobilization of Si, Fe, and Al (Schaefer et al. 2004, 2008). While most of the Si is leached out of the profile, Al and Fe are repreciptated, after downward migration of colloidal solutions, followed by changing chemical conditions with depth and degradation of Al–Fe unstable organic complexes. Therefore, there is a cyclical succession of formation and destruction of the canga, with limited lowering of the landscape (Schaefer et al. 2008).
Subsurface degradation of the laterite crust occurs with the progressive lowering of the water table level, caused by regional uplift, which intensifies the vertical and lateral migration of aqueous solutions, reactivating the change process. For their insolubility, Al and Fe remobilized from the surface canga degradation is deposited in the form of gibbsite and iron oxyhydroxide along fractures and cracks on the top of the saprolite, forming a dense system of veins of gibbsite or goethite/hematite on the regolith.
5 Case Study 1—Island of Ferruginous Rupestrian Physiognomy in the Midst of the Amazonia Forest: A Sequence of Carajás Plateau, Brazilian Eastern Amazonia
Banded iron formations and associated ironstone soils and landscapes are hotspots of plant diversity around the world (Gibson et al. 2012; Porembski et al. 1994, 1997; Valentin et al. 1999; Vincent et al. 2002; Yates et al. 2007), supporting plant communities distinctive both structurally and floristically from surrounding vegetation (Gibson et al. 2010, 2012; Jacobi et al. 2008; Yates et al. 2011). These ironstone substrates present a series of restrictions to plant establishment, including shallow and patchy soils (resulting in low water-retention capacities and low nutrient availability), high insolation levels, elevated temperatures, and the presence of potentially toxic metal concentrations (Silva and Rosa 1984; Meirelles et al. 1997). The topographic and edaphic characteristics of rupestrian grassland rock outcrops likely influence the floristic composition of their vegetation cover, as described for many rocky outcrop environments elsewhere in the world (Gibson 2004a, b; Markey and Dillon 2008; Meissner and Caruso 2008 ; Porembski et al. 1994; Schaefer et al. 2009).
In this section, we present three distinct vegetation types that make up the RGC on the ferruginous canga, ranging from open rupestrian grasslands and shrubby rupestrian grasslands on rocky sites of the upper plateau (746–762 m asl), to low forest at lower elevations. Open rupestrian grasslands is an open formation of small shrubs, with Vellozia glochidea (Velloziaceae) dominant over a ground layer of grasses; shrubby rupestrian grassland is supported on more fragmented canga soil that allow greater root penetration, with a mix of tall shrubs and herbaceous plants; “capão” forest is a semi-deciduous forest formation isolated as small islands within the matrix of herbaceous and shrubby rupestrian grassland (see Chap. 7).
5.1 Study Site
The study was located on a plateau of ironstone canga (lateritic capping) known as Serra Sul (738–762 m a.s.l.) in the Carajás National Forest (FLONA) conservation unit, southeastern Pará State, Brazil (Fig. 2.10a). Ferruginous rupestrian grassland islands cover approximately 9,031 ha (2.28 %) of the total FLONA area of 395,827 ha.

National forest of Carajás, Pará state, Brazil. a Serra Sul plateau (06 24’ 36,4” S and 50 21’ 29,4” W), with the sequence of physiognomies found on ironstone. b Open rupestrian grassland. c Shrubby rupestrian grassland. d “capão forest
The climate in the region is tropical, hot and humid (type “Aw” in the Köppen system; Ab’Saber 1986), with a dry season between May and October (average precipitation < 60 mm in the driest months), and a rainy season between November and April. Rainfall increases with altitude, lowland areas receiving annual averages of 1500 mm and more elevated sites receiving up to 1900 mm/year (IBAMA 2003). Average monthly temperatures vary between 19 and 31°C.
The RGC on ferruginous canga in Carajás ranges from open rupestrian grassland and shrubby rupestrian grassland on rocky sites to “capão” forest and fringing montane forest where the ironstone is fragmented and weathered to greater depths (see Fig. 2.7).
5.2 Open Rupestrian Grassland
The open rupestrian grassland (ORG) represents areas where rocky Fe-rich canga outcrops, with soil pockets rarely exceeding 10 cm, usually less than 5 cm of organic-rich soil. This organic surface layer is basically derived from termite nest surface erosion, spreading a friable soil that fills fractures and micro depressions on canga, where roots of Vellozia glochidea and tussocks of Sobralia liliastrum are dominating. At wetter areas usually shaded, these species display a taller development, where water deficit is reduced. The frequent fire finds difficult propagation due to the existence of natural barriers of canga (lithoplintic) outcrops where no vegetation grows (Figs. 2.7, 2.10b).
The grassy, herbaceous physiognomy is dominating, but shrubby are common, side by side with herbs Ipomoea carajaensis, Norantea goyasensis, Ipomoea marabaensis, Sobralia liliastrum. According to Schaefer et al. (2008), soils in the ORG are Petric Plinthosols, Lithoplintic, dystric, a very shallow soil.
5.3 Shrubby Rupestrian Grassland
The shrubby Rupestrian grassland formation (SRG) is the most common physiognomy on the Carajás Plateau, with a marked homogeneity of the shrubby stratum, with variations in dominance and abundance. Soils are slightly deeper than in the ORG previously described, with depth reaching 30 cm. The canga (ironstone) is much more fragmented, giving way to a loose soil with easy root penetration. The alternation of soils with varying depths indicate a severe morphogenesis in the canga plateau and great erosion losses. Termite activity is also a keystone, with similar abundance compared with ORG (Fig. 2.10c). Trees are mainly Callistene minor, Byrsonima coriacea, Bauhinia pulchella, Eugenia punicifolia, Alibertia cf. longiflora e Mimosa acutistipula var. ferrea.
The Petric Plinthosols are usually concretionary or lithic, but always deeper (10–35 cm) than ORG (Schaefer et al. 2008).
5.4 “Capão” Forest
With soil development and accumulation, trees are commonly found, forming small islands of forest with ovoidal or circular shapes, usually associated with internal depressions or cave systems, where fine particles and sediments sink and accumulate (Fig. 2.10d). One such forest has been studied. According to Schaefer et al. (2008), a collapse of loose saprolite underneath the cemented canga (litoplintic) layer led to colluvial accumulation of deep loose soil material, well-aggregated and with good water retention depths are typically within 30–120 cm, with Latosols, Cambisols and Petric Plinthosols (deep, concrectionary) dispersed, fragmented cores of canga forming concrections. Termite mounds are abundant, formed by hematite-rich soil, rather than organic-rich sapric, black material, as in the case of ORG. The main forest species are Pouteria ramiflora, Sacoglottis mattogrossensis and Myrcia splendens.
6 Case Study 2—Canga Soils of the Iron Quadrangle, Serra Da Moeda, Minas Gerais, Brazil
The soil sequence in the Iron Quadrangle is rather similar to that of Carajás, and similar soil depths control the same physiognomy, in Carajás and Iron Quadrangle. The soils developed on Fe-rich substrates from Moeda are perferrics (>36 % Fe2O3, by sulfuric acid digestion, EMBRAPA 1997), and have clay textures, but are strongly affected by the presence of nodules and pisolithic concretions responsible for high levels of silt present (as microaggregates difficult to disperse) and sand (micronodules of hematite), in addition to gravels (bigger concretions, nodules, etc.). The water-dispersible clay content in water is low, especially in the Bw and Bi horizons.
The soils on canga (Table 2.1) show up a very poor nutrient status, with extremely low exchange capacity, reflecting its oxidic nature. The pH values are slightly acid, as a result of high weathering degree, which practically left an exhausted cation exchange capacity, which depends almost entirely of organic matter. This organic matter, in turn, is very poor and acidic, and mostly made up of black-carbon, resulting from incomplete burning (Benites et al. 2005, 2007). Fire appears to be a fundamental factor in the genesis of canga landscapes.
Ca, Mg and K levels are extremely low, consistent with results from elsewhere. The total organic carbon shows little significant values for horizons identified as humic, where organic matter coats aggregates and micronodules. The total N levels are too low and reflect the general poverty of the soil environment.
7 General Aspects of Canga Soils—A Discussion
Diverse terminologies have been used previously to describe the (mostly shrubby) vegetation growing on ferruginous laterites (canga) in eastern Amazonia. They have been classified as canga vegetation (Morellato and Rosa 1991; Secco and Mesquita 1983; Silva 1992), banded ironstone formations, ferruginous rupestrian grassland (Jacobi et al. 2007, 2008; Rizzini 1979; Vincent and Meguro 2008), and even metallophile savanna (Porto and Silva 1989). The plant species found growing there often show dwarfing (Porto and Silva 1989) and some may retain high concentrations of metals in their tissues (Teixeira and Lemos-Filho 1998; Vincent 2004). These high soil-metal concentrations may lead to the selection of tolerant individuals in these environments (Porto and Silva 1989), resulting in vegetation that is structurally and floristically distinct from the surrounding vegetation matrix. Regardless of differences between soil variables for the four areas sampled here, all were acidic and had low phosphorus levels; a resulted so found in other studies in ironstone (Markey and Dillon 2008, 2009, 2010; Meissner et al. 2009; Meissner and Wright 2010; Messias et al. 2013; Teixeira and Lemos-Filho 2002; Vincent and Meguro 2008). According to Jacobi and Carmo (2011), the comparison of open areas with forest islands, subjected to the same climatic conditions, shows the important influence of edaphic and microclimatic conditions in determining life-form spectra and species composition in these plant communities. The isolation of the edaphic environments of inselbergs produce endemic species, or species with distributions restricted largely to these conditions (Porembski et al. 1994), resulting in vegetation that is structurally and floristically distinct from the surrounding forest matrix, as reported by Viana and Lombardi (2007).
Soil bioavailable Al were markedly higher in open rupestrian grassland than in shrubby rupestrian grassland. High concentrations of Al can be toxic to plants (Table 2.2), but high levels of soil carbon can reduce the potentially toxic effects of high concentrations of Al (Barros 1979), and this may provide amelioration of potential toxicity effects in open rupestrian grassland. Similarly, although Al concentrations are high for surface soil in “Capão” forest, most nutrient cycling here is associated with the litter layer rather than with mineral soil. Melich-extractable Fe concentrations were high in all sites, with highest levels found in the rupestrian grassland sites (open and shrubby). The values for extractable Fe found here are higher than those reported by Silva (1992) for hematite-rich canga in Serra Norte, Carajás, and by Vincent and Meguro (2008) in the Iron Quadrangle of Minas Gerais, but there is no evidence to suggest that these high levels for extractable Fe (as Fe2+) are driving vegetation differences among sites, since most Fe is present in soil as Fe3+, being insoluble and non-available. This indicates that seasonal reducing conditions occur in ferruginous rupestrian grasslands, increasing the amount of Fe2+ released from Fe-oxide dissolution. Further work on the effective role of high metal concentrations is recommended.
Botrel et al. (2002) reported that areas with dry soils and low chemical fertility tend to have fewer species, but in our study observed species richness was generally high throughout. Nevertheless, based on species richness estimators, total species richness was much higher in the extensive fringing montane forest, and also higher in “capão” forest, than in rupestrian grassland on canga (Nunes et al. 2015). In addition, the four vegetation types held few species in common, suggesting strong edaphic filtering from the available plant species pool acting most strongly in the rupestrian vegetation types. The composition of the isolated “capão” forest and rupestrian grassland sites may represent floras more specialized to the environmental filters associated with these ironstone habitat islands (Viana and Lombardi 2007).
According to Lista de Espécies da Flora do Brasil (2015), most of the species found in montane forest are widely distributed in Amazonian rainforest, including Allophyllus floribundus, Cupania hispida, Dipterix odorata, Eugenia brachypoda, Eugenia patrisii, Guatteria foliosa, Inga stipulacea, Inga falcistipula, Nealchornea yapurensis, Vismia latifolia and Myrcia paivae. In contrast, while some species in “capão” forest are also found in other Brazilian biomes (e.g. Sacoglottis mattogrossensis, Myrcia splendens, Sclerolobium paniculatum, Matayba arborescens and Tapirira guianensis), many are unique to this vegetation type in the Amazon area, including Licania impressa, Alchornea fluviatilis, Connarus perrottetii, Cupania macrophylla, Emmotum fagifolium, Mouriri angulicosta, Ocotea tomentela, and Virola callophylla. “Capão” forest also holds some species in common with the rupestrian grassland (Callisthene minor, Eugenia punicifolia, Pouteria ramiflora and Myrcia cuprea), as found also by Viana and Lombardi (2007).
The rupestrian complex (open and shrubby rupestrian grassland) of Carajás comprises a mix of widely distributed and locally endemic plant species. Several species are shared with the Brazilian Cerrado biome (e.g. Chamaecrista desvauxi, Anemopaegma scabriusculum, Lippia alba, Norantea goyazensis, Guapira ferruginea, Pouteria ramiflora, Callisthene minor) and Caatinga (e.g. Pilosocereus sp., Mimosa acutistipula, Periandra mediterranea, Eugenia punicifolia, Polygala sp.), while a few (including Erythroxylum nelson-rosae, Ipomoea marabaensis, Ipomoea carajasensis, Ipomoea cavalcantei plus two varieties of Mimosa and one of Erythroxylum) are restricted solely to ferruginous habitats in Carajás. However, ongoing studies (Schaefer et al. 2009; Nunes et al. 2015) indicate that many species considered ironstone lovers (e.g. Vellozia glochidea) are widely distributed across open vegetation supported by different lithologies in this region, not just ironstone outcrops. Overall, the species composition of vegetation types of the banded-iron canga islands is unique and shows very high beta diversity, reflecting a high diversity of habitats defined by soil physical and chemical properties. The high conservation value of these habitat islands, and their potential loss through anthropogenic impacts, requires urgent conservation management action (Nunes et al. 2015).
Nunes et al. (2015) showed that chemical and physical soil attributes (Tables 2.2, 2.3) have a role beyond the definition of different phytophysiognomies, hence affecting species composition. For the latter, the environmental partitioning model explained 43 % of the observed variance. Soil variables, including pH and soil depth provided the greatest contribution explaining species composition, while combined soil chemical and physical properties also contributed in explaining the species distribution patterns observed. Differences in soil chemical attributes allowed clear separation of the rupestrian grassland sites, whereas soil depth appears to be a key factor for establishment and development of the forest sites.
Schaefer et al. (2009) noted that soil depth seemed to play a fundamental role in the definition of the phytophysiognomies seen in the Carajás Plateaux, which is supported by the relationships observed for soil depth in differentiating forest (deeper soils) and non-forest areas (shallower soils) on the one hand, and of SRG (deeper) and ORG (shallower) on the other. There is a clear vegetation gradient from ORG to “capão” forest accompanying the gradient of soil depth: ORG < SRG < “capão” forest. Yates et al. (2011) similarly identified species habitat preferences associated with soil depth for banded-iron plant communities in Western Australia.
Despite their shallowness, non-forest (open and shrubby rupestrian grassland) soils have higher surface (0–10 cm) organic matter levels than forest soils, probably due to the presence of charred organic matter produced by frequent fire. Thus, fire maybe an important environmental filter contributing to the floristic and structural differences in the vegetation. Furthermore, higher termite activity in the rupestrian grassland sites may also contribute to higher levels of surface soil organic carbon through redistribution of organic-rich materials (Schaefer et al. 2009). In their studies of the influence of termite activity on soil characteristics in Minas Gerais, Sarcinelli et al. (2009) concluded that high concentrations of organic matter, P and nutrients in termite mounds (in relation to the surrounding soils) were associated with the active incorporation of organic material during their construction. The higher organic matter in open rupestrian grassland may be also attributed to lower water availability and slower organic matter decomposition, and to differences in the soil fauna available for organic matter processing.
Thus, we can infer that edaphic (pedological) factors are crucial in explaining the types and distributions of rupestrian grasslands vegetation associated with ferruginous ironstone uplands (canga) in Carajás, eastern Amazon. Three different ferruginous vegetation types were identified, each with distinct soil physical and chemical, and floristic and structural characteristics. Vegetation differences were associated with variations in soil chemistry, depth and acidity, with forest soils showing greater depths, and higher concentrations of P and Al, but much lower levels of organic matter and exchangeable cations at the surface, reflecting major differences in nutrient cycles between forest and non-forest sites. The higher nutrient and organic carbon contents in rupestrian grasslands may be indirectly associated with frequent fire, filtering those species able to persist through recurrent fire events. Soil organic matter content is also likely a key variable for the development of the formations studied, especially in the ORG, where most of the inorganic nutrients were correlated with organic matter (carbon) amounts in the topsoil. High soluble Fe concentrations do not appear to restrict the establishment of species in the different formations, as this element was found in similar amounts throughout the study area. As such, the use of the term metal-loving savanna (or metallophile savanna) does not appear appropriate for the ironstone canga vegetation or rupestrian grassland at Carajás, and we suggest the use of the term ferruginous rupestrian grasslands complex for the open and shrubby rupestrian grasslands formations.
8 Case Study 3—Rupestrian Grassland on Metarenites and Metapellites from Serra Geral State Park, North Minas Gerais
Within the Brazilian semi-arid, there are many outliers of Proterozoic rocks (metarenites, quartzites, metacherts, metapelites) that form imposing Mountains in the ecotonal zone between the Atlantic Forest, Caatinga and Cerrado domains. On the rocky tops and escarpments of Serra Geral State Park, rupestrian formations are observed on very shallow soils (Fig. 2.11) (Rodrigues et al. 2014). In the following section, we discuss typical geoenvironments of rupestrian grasslands of this State Park, emphasizing and illustrating the most important landscape relationships. The landscapes associated with the soils described are illustrated in Fig. 2.11a–d. The Serra Central is bordered by the Limestone of Bambuí Group, to the west face (Fig. 2.11a), or basement rocks at the east facing slope (Fig. 2.11b), whereas dry forest occur on the depression lowlands, both east and west, downslope the escarpment.

A three-dimensional picture of the landforms at the Serra Geral state park, Minas Gerais states, illustrating the typical geoenvironments of rupestrian grasslands at the park, emphasizing and illustrating the most important landscape relationships. To the western slopes, the serra central is bordered by the limestone depression of bambuí group (São Francisco Depression) (a); to the east face, the plateau escarpment connects with the Espinosa depression, under semi-arid climates (b). In both depressions, dry forest occur on the lowlands, both east and west, downslope the escarpment. There is an abrupt change in vegetation as one approach the escarpment
8.1 High Top with Open Rupestrian Grassland on Metapellites
At the highest landscape position, shallow to moderately developed soils resting on rocky surfaces are gravelly and concretionary, being classified as Cambisols on metapelites/metacherts. Soil profile PSG1 (Table 2.4; Fig. 2.11d) is a Dystric Haplic Cambisol, clayey and gravelly, with very low fertility, very high toxic Al, low cation exchange capacity and sum of bases at the surface, as well at subsurface—2Bi horizon.
These unusual clayey soils are not typical of the RGC, but occur rather ocasionally, possessing a savanna-like physionomy. In these formations, high abundance of Vochysia rufa is diagnostic, besides Tachigali aurea, Vellozia sp., Byrsonima pachyphylla, Vochysia elliptica, Chamaecrista orbiculata, Davilla rugosa and Miconia albicans. The character of these formations is the marked abundance of Al- Hyperaccumulating plants, indicating extreme acidity and Al release from clays (Campos et al. 2014).
8.2 Slope and Crests with Open Rupestrian Grassland with Vellozia on Rocky Outcrops
In the steepy slopes at the fringes of Serra Central Plateau, or within the interior valleys where deep dissection took place, the metarenites and quartzite substrates outcrop as resistant rocks. There, typical rupestrian formations develop, following the structural alignments with the main geotectonic trend (NE/SW). Due to isolation within the semiarid domain, these Rupestrian Grasslands are characterized by high endemism, and presence of rare species. On rocky surfaces, lichen and mosses are very common, along crevices and shadowed faces. Soil occur as discontinuous pockets, rich in organic matter, but with very low fertility.
A representative soil is a Litholic Neosol, dystric, fragmentary (PSG2, Table 2.4). The sandy texture with a sequence of A-AC-Cr horizons, with unaltered rock and a lithic contact within 50 cm depth. Acid pH, very low contents of available P and CEC are typical.
The rupestrian vegetation is characterized by dominance of tall Vellozia sp, with common low trees and shrubs dominated by Vochysia rufa, Lychnophora e Chamaecrista, Vochysia elliptica, Tachigali aurea and Byrsonima pachyphylla.
8.3 Rupestrian Savanna (Cerrado Rupestre) on Escarpments
This environment represent an open to shrubby rupestrian grassland, in which many species of savanna (Cerrado) are found, on deeper soils. In the steep slopes and escarpments along the Plateau borders, shallow soils under strong erosion are covered by unusual savanna formation, with a rupestrian character. Soils are usually Regolitic Neosols (e.g. PSG3), dystric, typic, with a sequence of A-Cr horizon, and saprolite down to 50 cm, rather than fresh rock (Table 2.4). Texture is sandy loam with a predominance of fine sand (0,2–0,05 mm). Roots and biological channels are common, and many termite mounds highlight the importance of pedobioturbation in nutrient cycling in these soils. Values of cation exchange capacity, available P, organic matter and base saturation are very low, with a very acid pH (4.83 in the saprolite Cr), indicating very low nutrient reserves in the parent rock.
The vegetation of rupestrian savanna (cerrado rupestre) is characterized by trees and shrubs, mainly Vochysia rufa, Byrsonima pachyphylla, Myrsine guianensis (Aubl.) Kuntze, Eremanthus erythropappus (DC.) MacLeish, Tibouchina sp. and Tachigali aurea.
9 Case Study 4—A Soil Sequence of Quartzite and Micaceous Quartzite from the Espinhaço Range in the Sempre-Vivas State Park, Minas Gerais States
9.1 Study Area
The Espinhaço Meridional Range represent an ancient orogenic belt bordered by the São Francisco Craton, to the west, extending for 300 km in a N-S direction, from the Iron Quadrangle to the northern Minas Gerais. This is essentially built by the Espinhaço Supergroup (mainly quartzites rocks) with minor phyllites, conglomerates and acid volcanics (Almeida-Abreu 1995) (Fig. 2.12).

a Three dimensional diagram of the west-facing slope fault escarpment of the Sempre-Vivas national park (Espinhaço Range) under rupestrian grassland complex; the São Francisco depression in the foreground, down the escarpment front (under dry forest). In the detailed scheme. b The most typical rupestrian grassland complex (RGC) on quartzites from the Sempre-Vivas park is illustrated. The RGC show a variety of landforms at microscales, from Karst-like rocky pinnacles of exposed cavernous (ruiniform) quartzite, to low lying depression where sand accumulated by aeolian or colluvial deposition form sandy plains, similar to Heathlands. Termite activity is key for RG development, since it concentrates most nutrients available for plants at niche, local scale. The presence of shallow spodosols is on account of intense organic matter illuviation down the soil profile, forming well-developed spodic Bh horions of dark colours. The dissolution forms indicates that, albeit resistant to weathering, quartzites are being slowly attacked by acid solutions, leading to the present-day ruiniform grounds where erosion exhumed the former subsurface rock substrate. (Drawing by C. Schaefer)
As part of Espinhaço Range, the National Park of Sempre-Vivas covers an 124,555 ha (IBAMA 2006) with an altitudinal range of 650–1,350 m (Almeida et al. 2007). The climate is wet subtropical, mesic (Cwb, according to Köppen’s type) with mean annual temperature of 20° C, and rainfall of 1,500 mm (Almeida et al. 2007).
Approximately 80 % of the Park is on quartzites of Galho do Miguel Formation (Espinhaço Supergroup), whereas the remaining are minor metapelitic to conglomerate rocks, from the Macaúbas and Bambuí Groups.
The Galho do Miguel Formation is characterized by orthoquartzites, pure and fine-grained in more than 10 % of the unit. The remaining 10 % is basically micaceous quartzites, with occasional shales. The original stratification is well preserved, and an aeolian origin is postulated (Dossin et al. 1987).
The soil sequence studied ranged from open rupestrian grassland with Vellozia, to rupestrian savanna , savanna and dwarf forest; the latter two formations are influenced by micaceaus quartzite (Fig. 2.12).
9.2 Open Rupestrian Grassland with Vellozia on Litholic Neosols
The open rupestrian physiognomy is invariably associated with very shallow soils (Litholic Neosol, BSV1) on quartzite outcrop, in which soils occur as discontinuous pockets amid the rocky surface. Abundant Velloziaceae dominates the formation, with a dense herbaceous cover of Poaceae, Cyperaceae and Xyridadeae (Fig. 2.13a). Dwarf trees and shrubby are common on fractured rocks, with their roots penetrating in fissures and weakness lives where organic matter accumulates. The rugged, mountainous landform account for steep slopes where erosion is severe with intense soil loses! The structural control on rupestrian vegetation is evident with massive rocky covers on the top land surface. The main genera are Lagenocarpus sp, Trachypogon sp, Vellozia sp, Lychnophora sp, Symplocos sp, Byrsonima sp and Chamaecrista sp. Fire is intense and frequent, but its effect is reduced by the presence of little biomass and rocky surfaces.

Sempre-Vivas national park, Minas Gerais state, Brazil. a Open rupestrian grassland with vellozia on litholic neosols. b Open and shrubby rupestrian savanna on cambisols. c Savanna (cerrado sensu stricto) on latosols and cambisols. d Dwarf forest in foreground and rupestrian grassland on the rock outcrop in background
9.3 Open and Shrubby Rupestrian Savanna on Cambisols
The rupestrian savanna is a typical open formation on metapelitic to metarenites rocks, with greater silt contents, and deeper soil, and part of the RGC. Rocky outcrops are not common, only where resistant rocky cores stand out (Fig. 2.13b). The physiognomy varies from open grassy savanna to typical savanna (cerrado sensu stricto). The herbaceous stratum is graminoid/cyperoid, and fire propagation is facilitated by a continuous grassy cover, compared with open rupestrian grassland. Soil have a cambic horizon, but are nutrient-poor and acid, as well as shallow and silt-rich. Colours are redder, resulting form higher Fe-content. Typical genera are Qualea sp, Vellozia sp, Baccharis sp, Trachypogon sp and Vochysia sp.
9.4 Savanna (Cerrado Sensu Stricto) on Latosols and Cambisols
Typical savannas occurs on deeper soils on slopes connecting the high plateau, under the influence of metapelitic rocks, savanna s.s. develops on deep Latosols or Cambisols, all very dystric and acid, with very low cation exchange capacity and virtually no nutrient reserve (acric, gibbsitic) (Fig. 2.13c). Plants are well adapted to extreme nutrient depletion, but require clayey and deeper soils. Tree typical species are Caryocar brasiliense Cambess., Xylopia aromatica (Lam.) Mart., Qualea grandiflora Mart and many siliceous Poaceae, forming a grassy cover. The main difference is the absence of rock outcrops and rupestrian character.
9.5 Dwarf Forest on Quartzarenic Neosols or Medium-Texture Latosols
The Dwarf Forests are cloud forest (rich in epiphytes) at altitudes higher than 1,000 m, usually on deep soils (PS) of medium to sandy textures (high fine sand content) (Fig. 2.13d). These dwarf forests are transitional with candeias shrubby, but with soils with better water retention. Soils are rich in organic matter at the surfaces although very poor in nutrients. Typical trees are Protium heptaphyllum (Aubl.) Marchand, Guatteria sellowiana Schltdl. and Miconia cabucu Hoehne.
10 Final Remarks
-
1.
The Rupestrian Grassland complex is widely distributed across the Brazilian Territory, and closely follows the occurrence of highland resistant rocks, in the decreasing order of importance: Quartzites ≫ Fe-rich canga and banded iron formations > Igneous rocks (Granites, Syenites) or high-grade metamorphics (Gneiss, schists) > metapelitic rocks > other minor types (conglomerate, limestone). Quartzites comprise the vast majority of RG occurrences.
-
2.
RGC is closely associated with high altitudes, in which a number of factors have a prominent role: (i) Extreme oligotrophy; (ii) chemical resistance to weathering; physical resistance; (iii) wind-prone environment at high landsurfaces; (iv) frequent fire, and harsh climatic regime.
-
3.
Variations of RG physiognomies are basically related to soil depth and drainage (edaphic factor) and landscape stability and evolution.
-
4.
The proposed map of RGC distribution included areas not well investigated by ecologists, such as Tepuis (Roraima and Amazonas), Serras de Santa Barbara and Ricardo Franco (Mato Grosso), Cachimbo (Pará). On the other hand, also included well-known areas, formerly classified as Campos de Altitude , such as Serra dos Órgãos (Rio de Janeiro), Itatiaia (Rio/Minas) and Caparaó (Minas/Esp. Santo). In all areas, true Rupestrian vegetation formation exists, so that there is no reason to constrain the limits of RGC to the quartzitic domains, as traditionally envisaged.
-
5.
There is an urgent need for comparative studies (floristic, phylogenetics, pedological, structural and phytosociological) for investigating the similarities between these postulated occurrences of RGC in Brazil, so widely separated by surrounding dominant vegetation.
-
6.
Case studies selected and presented here highlight the importance of parent materials and soil attributes (including pedobiological aspects) for determining the phytophysiognomies of RGC. In canga (Fe-rich substrates), for example, the degree of degradation of ironstone controls soils formation and, consequently, vegetation development. In granite, rockiness and soil attributes are the main drivers.
-
7.
In synthesis, we present two basic diagrams of RGC in Brazil (Figs. 2.3, 2.12) to illustrate typical situations on (1) Granite, (2) Quartzite. In the first case, the granitic parent material is a massive rock body not so limiting in terms of chemistry, but with very poor soils resulting from extreme weathering and leaching of nutrients at these old, high mountain landscapes. In the second case, the chemical and physical resistance of quartz-rich substrates account for very shallow soils, in general. Variations of soil drainage, depth, organic-matter accumulation, aspect, slope, stoniness and rockiness and termite activity are evident, and help to explain the different niches, in which resources (limiting) are differentially offered. Hence, all high landsurfaces of these resistant mountain tops are comparable, in terms of nutrient depletion, acidity, fire regime and climate, leading to a convergent evolution of RGCs.
-
8.
Traditional Brazilian vegetation classification have emphasized the differences between Campos de Altitude (High Altitude Fields) (Martinelli 1989) on granite/gneiss and Campos Rupestres (Rupestrian Grasslands) on Canga or quartzite. However, we see no reason to treat these isolated hot-spots of true Rupestrian domains on Granite as separate phytogeographical entities. By the same rationale, Tepuis and Carajás should be treated as part of this single and complex Campo Rupestre domain. They do share many species in common, have a clear rupestrian physiognomy in most places, and are subjected to similar fire and wind regimes by high ground exposure. In addition, traits related to resource conservation are very similar, since soils are generally poor, acid, shallow and water deficient.
-
9.
The importance of RGC as biodiversity pumps (sensu Silveira et al. 2016) or hot spots are really large, and unique. They occur on different rocks, are widely distributed across Brazil, within contrasting biomes, and are relatively well-preserved by mountain top isolation, under limited agricultural pressures. Other anthropic pressures, however, are mounting: mining, pasture expansion, tourism, water needs, so that urgent measures must be taken for its long term conservation. Sound scientific research is the necessary framework for any sustainable knowledge on RG, possibly one of the oldest kingdoms of the Brazilian Neotropics.
References
Ab’Saber AN (1986) Geomorfologia da Região. In: Almeida JR (ed). Carajás: desafio político, ecologia e desenvolvimento. Brasiliense, Brasília, pp. 88–124
Alkmim FF (2015) Geological Background: A Tectonic Panorama of Brazil. In: Vieira BC, Salgado AAR, Santos LJC (eds) Landscapes and landforms of Brazil. Springer, Netherlands, pp 9–17
Almeida-Abreu PA (1995) O Supergrupo Espinhaço da Serra do Espinhaço Meridional (Minas Gerais): o Rifte, a Bacia e o Orógeno. Geonomos 3:1–18
Almeida TE, Souza DT, Salino A, Teles AM (2007) Levantamento florístico e caracterização das formações vegetacionais, Parque Nacional das Sempre-Vivas. Unpub. Technical report
Alves RJV, Silva NG, Oliveira JA, Medeiros D (2014) Circumscribing campo rupestre—megadiverse Brazilian rocky montane savanas. Braz J Biol 74:355–362
Barros NF (1979) Growth and foliar nutrient concentrations of Eucalyptus grandis in relation to spodasol properties in South Florida. Ph.D. Dissertation, University of Florida
Benites VM, Sá E, Schaefer CER, Novotny EH, Reis E, Ker JK (2005) Properties of black soil humic acids from high altitude rock complexes in Brazil. Geoderma 127:104–113
Benites VM, Schaefer CEGR, Simas FNB, Santos HG (2007) Soils associated with rock outcrops in the Brazilian mountain ranges Mantiqueira and Espinhaço. Rev Bras Bot 30:569–577
Botrel RT, Oliveira-Filho AT, Rodrigues LA, Curi N (2002) Influencia do solo e topografia sobre as variações da composição florística e estrutura da comunidade arbóreo–arbustiva de uma floresta estacional semidecidual em Ingaí, Mg. Rev Bras Bot 25:195–213
Butcher R, Byrne M, Crayn DM (2007) Evidence for convergent evolution among phylogenetically distant rare species of Tetratheca (Elaeocarpaceae formerly Tremandraceae) from Western Australia. Aust Syst Bot 20:126–138
Campos NV, Pereira TAR, Machado MF, Guerra MBB, Tolentino GS, Araújo JS, Rezende MQ, Silva MCNA, Schaefer CEGR (2014) Evaluation of micro-energy dispersive X-ray fluorescence and histochemical tests for aluminium detection in plants from high altitude rocky complexes, Southeast Brazil. An Acad Bras Ciênc 86:285–296
Carmo FF, Jacobi CM (2012) As cangas do Quadrilátero Ferrífero. In: Jacobi CM, Carmo FF (eds) Diversidade florística nas cangas do Quadrilátero Ferrífero. IDM, Belo Horizonte, pp 1–13
Dias HCT, Fernandes EIF, Schaefer CEGR, Fontes LEF, Ventorim LB (2002) Geoambientes do Parque Estadual do Ibitipoca, Município de Lima Duarte-MG. R Árv 26:777–786
Diniz JMFDS, Dos Reis AA, Junior FWA, Gomide LR (2014) Detecção da expansão da área minerada no Quadrilátero Ferrífero, Minas Gerais, no período de 1985 a 2011 através de técnicas de sensoriamento remoto. Bol Cien Geod 20:683–700
Dossin IA, Garcia AJV, Uhlein A, Dossin TM (1987) Facies eólico na Formação Galho do Miguel, Supergrupo Espinhaço-MG. Bol Soc Bras Geol 6:85–96
EMBRAPA—Empresa Brasileira De Pesquisa Agropecuária (1997) Manual de métodos de análise de solo. Centro Nacional de Pesquisas de Solos, Rio de Janeiro
Gibson N (2004a) Flora and vegetation of the eastern Goldfields ranges: Part 6. mt manning range. J R Soc West Aust 87:35–47
Gibson N (2004b) Flora and vegetation of the eastern Goldfields ranges: Part 7. middle and south Ironcap, Digger Rock and Hatter Hill. J R Soc West Aust 87:49–62
Gibson N, Yates CJ, Dillon R (2010) Plant communities of the ironstone ranges of South Western Australia: Hotspots for plant diversity and mineral deposits. Biodiv Cons 19:3951–3962
Gibson N, Meissner R, Markey AS, Thompson WA (2012) Patterns of plant diversity in ironstone ranges in arid south Western Australia. J Arid Environ 77:25–31
Grime JP (2001) Plant strategies, vegetation processes, and ecosystem properties, 2nd edn. John Wiley, Sons Ltd, Chichester
Harley RM, Simmons NA (1986) Florula of Mucugê, Chapada Diamantina—Bahia, Brazil. Royal Botanical Garden, Kew
IBAMA—Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais Renováveis (2003) Plano de manejo para uso múltiplo da Floresta Nacional de Carajás. IBAMA, Brasília
IBAMA—Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais Renováveis (2006) Plano Operativo de prevenção e combate aos incêndios do Parque Nacional das Sempre-Vivas. IBAMA, Diamantina
Jacobi CM, Carmo FF (2011) Life–forms, pollination and seed dispersal syndromes in plant communities on ironstone outcrops, SE, Brazil. Act Bot Bras 25:395–412
Jacobi CM, Carmo FF, Vincent RC, Stehmann JR (2007) Plant communities on ironstone outcrops: A diverse and endangered Brazilian ecosystem. Biodiv Cons 16:2185–2200
Jacobi CM, Carmo FF, Vincent RC (2008) Estudo Fitossociológico de uma comunidade vegetal sobre canga com subsídio para a reabilitação de áreas mineradas no Quadrilátero Ferrífero, MG. Rev Arv 32:345–353
Jacobi CM, Carmo FF, Campos IC (2011) Soaring extinction threats to endemic plants in Brazilian metal-rich regions. Ambio 40:540–543
Lista de Espécies da Flora do Brasil (2015) Jardim Botânico do Rio de Janeiro. Available in. http://floradobrasil.jbrj.gov.br/. Accessed 29 Jan 2016
Markey AS, Dillon SJ (2008) Flora and vegetation of the banded iron formations of the Yilgarn Craton: The central tallering land system. Cons Sci West Aust 7:121–149
Markey AS, Dillon SJ (2009) Flora and vegetation of the banded iron formations of the Yilgarn Craton: Herbert Lukin Ridge (Wiluna). Conserv Sci West Aust 7:391–412
Markey AS, Dillon SJ (2010) Flora and vegetation of the banded iron formations of the Yilgarn Craton: Gullewa. Conserv Sci West Aust 7:531–556
Martinelli G (1989) Campos de, altitude edn. Index, Rio de Janeiro
Meirelles ST, Mattos EA, Silva AC (1997) Potential desiccation tolerant vascular plants from southeastern Brazil. Pol J Environ Stud 6:17–21
Meissner R, Caruso Y (2008) Flora and vegetation of banded iron formations of the Yilgam Craton: Koolanooka and Pernjori hills. Cons Sci West Aust 7:73–88
Meissner R, Wright J (2010) Flora and vegetation of banded iron formations on the Yilgarn Craton: South Illaara Greenstone Belt. Conse Sci West Aust 7:605–616
Meissner R, Owen G, Bayliss B (2009) Flora and vegetation of the banded iron formation of the Yilgarn Craton: Robinson Ranges and Mount Gould. Cons Sci West Aust 7:363–376
Mendonça MP, Lins LV (2000) Lista Vermelha das Espécies Ameaçadas de Extinção da Flora de Minas Gerais. Biodiversitas e Fundação ZooBotânica de Belo Horizonte, Belo Horizonte
Messias MCTB, Leite MGP, Meira-Neto JA, Kozovits AR, Tavares R (2013) Soil–vegetation relationship in quartzitic and ferruginous Brazilian rocky outcrops. Folia Geobot 48:509–521
Morellato PC, Rosa NA (1991) Caracterização de alguns tipos de vegetação na região amazônica, Serra dos Carajás. Pará Brasil Rev Bras Bot 14:1–14
Mourão A, Stehmann JR (2007) Levantamento da flora do campo rupestre sobre canga hematítica couraçada remanescente na Mina do Brucutu, Barão de Cocais, Minas Gerais, Brasil. Rodriguésia 58:775–785
Nunes JA, Schaefer CEGR, Ferreira-Júnior WG, NERI AV, Correa, GR, Enright NJ (2015) Soil-vegetation relationships on a banded ironstone island, Carajás Plateau, Brazilian Eastern Amazonia. An Acad Bras Cien 87:2097–2110
Porembski S, Barthlott W, Dorrstock S, Biedinger N (1994) Vegetation of rock outcrops in Guinea: Granite inselbergs, sandstone table mountains and ferricretes—remarks on species numbers and endemism. Flora 189:315–326
Porembski S, Seine R, Barthlott W (1997) Inselberg vegetation and biodiversity of granite outcrops. J Roy Soc West Austral 80:193–197
Porto ML, Silva MFF (1989) Tipos de vegetação metalófila em áreas da Serra de Carajás e de Minas Gerais, Brasil. Act Bot Bras 3:13–21
Queiroz LP, Sena TSN, Costa MJSL (1996) Flora vascular da Serra da Jibóia, Santa Terezinha. Bahia. I: O campo rupestre. Sitientibus 15:27–40
Rizzini CT (1979) Tratado de fitogeografia do Brasil: Aspectos sociológicos e florísticos. Editora da Universidade de São Paulo, São Paulo
Rodrigues PMS, Schaefer CEGR, Corrêa GR, Campos PV, Neri AV (2014) Soils, landform and vegetation determine the geoenvironments at a conservation unit in northern Minas Gerais, Brazil. Neotrop Biol Cons 10:31–42
Santos MDC, Varajão AF (2004) Sedimentation and pedogenic features in a clay deposit in Quadrilátero Ferrífero, Minas Gerais, Brazil. An Acad Bras Ciênc 76:147–159
Sarcinelli TS, Schaeffer CEGR, Lynch LS, Arato HD, Viana JHM, Albuquerque Filho MR Gonçalves TT (2009) Chemical, physical and micromorphological properties of termite mounds and adjacent soils along a toposequence in Zona da Mata, Minas Gerais State, Brazil. Catena 76:107–113
Schaefer CEGR (2001) Brazilian latosols and their B horizon microstructure as long-term biotic constructs. Soil Res 39:909–926
Schaefer CEGR (2013) Bases físicas da paisagem brasileira: estrutura geológica, relevo e solos. In: Araújo AP, Alves BJR (eds) Tópicos em ciência do solo. Sociedade Brasileira de Ciência do Solo, Viçosa, pp 1–69
Schaefer CEGR, Gilkes RJ, Fernandes RBA (2004) EDS/SEM study on microaggregates of Brazilian Latosols, in relation to P adsorption and clay fraction attributes. Geoderma 123:69–81
Schaefer CEGR, MendonçA BAF, Ribeiro ASS (2008) Solos desenvolvidos sobre canga ferruginosa no quadrilaÏtero ferriÏfero, Minas Gerais. Biodiversidade, Conservaçafio e Perspectivas de Sustentabilidade. UFMF/ ICB, Belo Horizonte, SimpoÏsio Afloramentos Ferruginosos no QuadrilaÏtero FerriÏfero
Schaefer CEGR, Simas FNB, Mendonça BAF, Saboya AS, Ferreira Júnior WG, Nunes JA, Correa GR (2009) Geodiversidade dos ambientes de canga na região de Carajás– Pará. Technical Report. part 2. Vale Brazilian Company
Schaefer CEGR, Cândido HG, Corrêa GR, Pereira A, Nunes JA, Souza OF, Marins A, Fernandes-Filho E, Ker JC (2015) Solos desenvolvidos sobre canga ferruginosa no Brasil: Uma revisão crítica e papel ecológico de termiteiros. In: Carmo FF, Kamino LHY (eds) Geossistemas ferruginosos do Brasil: Áreas prioritárias para conservação da diversidade geológica e biológica. Patrimônio cultural e serviços ambientais. 3i, Belo Horizonte, pp. 77–102
Schobbenhaus C, Brito-Neves BB (2003) A geologia do Brasil no contexto da Plataforma Sul-Americana. In: Schobbenhaus C, Vidotti RM, Gonçalves JH (eds) Geologia, tectônica e recursos minerais do Brasil. CPRM, Brasília, pp 5–25
Secco RS, Mesquita AL (1983) Nota sobre a vegetação de canga da Serra Norte. I. Boletim Mus Par Em Goeldi 59:1–13
Semir J (1991) Revisão taxonômica de Lychnophora Mart. Vernoniaceae: Compositae. Ph.D. Thesis, Universidade Estadual de Campinas, Campinas
Silva MFF (1992) Distribuição de metais pesados na vegetação metalófila de Carajás. Act Bot Bras 6:107–22
Silva MFF, Rosa N (1984) estudos botânicos na área do projeto ferro de Carajás, Serra Norte. I—Aspectos ecológicos e vegetacionais dos campos rupestres. In: XXXV Congresso Nacional de Botânica, Sociedade Brasileira Botânica, Manaus, pp. 367–379
Silveira FA, Negreiros D, Barbosa NP, Buisson E, Carmo FF, Carstensen DW, Lambers H (2016) Ecology and evolution of plant diversity in the endangered campo rupestre: a neglected conservation priority. Plant and Soil. doi: 10.1007/s11104-015-2637-8
Tardy Y, Nahon D (1985) Geochemistry of laterites, stability of Al-goethite, Al-hematite, and Fe3 + -kaolinite in bauxites and ferricretes: An approach to the mechanism of concretion formation. Am J Sci 285:865–903
Teixeira WA, Lemos-Filho JP (1998) Metais pesados em folhas de espécies lenhosas colonizadoras de uma área de mineração de ferro em Itabirito, Minas Gerais. Rev Árv 22:381–388
Teixeira WA, Lemos-Filho JP (2002) Fatores edáficos e a colonização de espécies lenhosas em uma cava de mineração de ferro em Itabirito, Minas Gerais. Rev Árv 26:25–33
Valentin C, Herbes JM, Poesen J (1999) Soil and water components of banded vegetation patterns. Catena 37:1–24
Veloso HP, Rangel Filho ALR, Lima JCA (1991) Classificação da vegetação brasileira, adaptada a um sistema universal. Instituto Brasileiro de Geografia e Estatística, Rio de Janeiro
Viana PL, Lombardi JA (2007) Florística e caracterização dos campos rupestres sobre canga na Serra da Calçada, Minas Gerais, Brasil. Rodriguesia 58:159–177
Vincent RC (2004) Florística, fitossociologia e relações entre a vegetação e o solo em área de campos ferruginosos no Quadrilátero Ferrífero, Minas Gerais. Ph.D. Dissertation, Universidade de São Paulo
Vincent RC, Meguro M (2008) Influence of soil properties on the abundance of plant species in ferruginous rocky soils vegetation, southeastern Brazil. Rev Bras Bot 31:377–388
Vincent RC, Jacobi CM, Antonini Y (2002) Diversidade na adversidade. Cien Hoje 31:64–67
Yates CJ, Ladd PG, Coates DJ, Mcarthur S (2007) Hierarchies of cause: Understanding rarity in an endemic shrub Verticordia staminosa (Myrtaceae) with a highly restricted distribution. Aust J Bot 55:194–205
Yates CJ, Gibson N, Pettit NE, Dillon R, Palmer R (2011) The ecological relationships and demography of restricted ironstone endemic plant species: Implications for conservation. Aust J Bot 59:692–700
Acknowledgments
Carlos Schaefer thanks Vale, CNPq and FAPEMIG for financing long-term studies on Carajás and Iron Quadrangle; thanks are due to Dr. Lilia Santos (Bioma), James Bockheim, and João Ker for many field trips and discussions on RGC ecology. Help for field work Sr. Alexander Castilho (Vale) is greatly appreciated.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Appendix 1
Appendix 1
Picture illustrations of phytophysiognomies and landscape features of Rupestrian Grassland Complex (RGC) on different lithologies in Brazil. The striking similarities between widely separate geographical sits, very contrasting parent-materials, indicate a notable convergence of edaphic/landscape features at different localities. (a) Itatiaia highlands at Campo Belo Valley (2400 m) on syenite (alkaline rocks) (mark 20 in Fig. 2.14). (b) Caparaó highlands at Upper Crystal Valley (above Terreirão Plateau), with Chusquea sp. on granitoid rocks (mark 17 in Fig. 2.14). (c) Serra do Brigadeiro Plateau Pedra do Pato, migmatites/granites Paraíba do Sul headwaters. (d) Shrubby rupestrian grassland in Serra do Espinhaço, Diamantina, Minas Gerais States, with Syagrus sp. (Palmeira da Serra), on micaceous quartzites (mark 12 in Fig. 2.14). (e) Sempre-Vivas National Park, shrubby rupestrian grassland on quartzite (mark 12 in Fig. 2.14). (f) Canastra National Park, open rupestrian grassland with Vellozia sp. on micaceaus quartzites/conglomerates (mark 19 in Fig. 2.14). (g) Ibitipoca State Park, with open rupestrian grassland on micaceous quartzites and metapelits (mark 23 in Fig. 2.14). (h) São Francisco River headwaters at Canastra National Park, with shrubby rupestrian grassland a quartzite plateau (mark 19 in Fig. 2.14). (i) Shrubby ferruginous rupestrian grassland on Fe-rich canga of Carajás Plateau, Santa Rita Durão, (see Caraça quartzite at background) (1000 m) (mark 8 in Fig. 2.14). (j) General view of ferruginous rupestrian grassland on the Fe-ich itabirite/canga on Serra Sul Plateau of Carajás (900 m) (mark 8 in Fig. 2.14). (k) Ferrugineous rupestrian grassland on the Fe-rich Itabirite/canga of Alto do Conta História (1600 m), Iron Quadrangle, Minas Gerais States (mark 16 in Fig. 2.14). (l) Exposed itabirite saprolite with overlying canga on the Moeda Plateau, with ferruginous rupestrian grassland (shrubby/open) (1380 m) (mark 16 in Fig. 2.14). (m) Open rupestrian savanna on metapelites from Serra de Grão Mogol (1200 m) (mark 12 in Fig. 2.14). (n) Open rupestrian grassland on conglomeratic quartzites of Diamantina Plateau, Minas Gerais States (mark 12 in Fig. 2.14) at 1420 m. (o) Sempre-Vivas National Park with open rupestrian grassland on quartzite with Lychnophora sp. (mark 12 in Fig. 2.14) at 1130 m. (p) Serra de São José Environmental Protected Area, Tiradentes, Minas Gerais States, with open rupestrian grassland on micaceous quartzite and schists at 1200 m (q) Pico da Neblina, the highest RG site in Brazil (close to 3000 m) on metarenites of Roraima Group, Amazonas State (mark 1 in Fig. 2.14). (r) Serra do Sol, at 2250 m altitude, on quartzites of the Roraima Group, Roraima State (mark 1 in Fig. 2.14). (s) Serra dos Pacaás Novos, with an altitude of 1050 m, an isolated spot of RG in the middle of the Amazon Forest domain, with metarenites to metapelitic rocks (mark 3 in Fig. 2.14). (t) (mark 1 in Fig. 2.14). (u) Serra Ricardo Franco, in the Brazil-Bolivia border, at Mato Grosso State, where RG occurs on metarenites (mark 4 in Fig. 2.14) at 1030 m. (v) Monte Roraima highest surface (mark 1 in Fig. 2.14), the largest Rupestrian Plateau in the Brazilian Amazonia (Roraima State), at 2850 m. (w) Monte Roraima top surface (2800 m), on Quartzite of the Roraima Group (mark 1 in Fig. 2.14). (x) Serra da Seringa, southeastern Pará State, an isolated RGC on Felsic Granites in the middle of the Caiapó Indigenous Land (mark 7 in Fig. 2.14).



Phytophysiognomies and landscape features of Rupestrian Grassland Complex (RGC) on different lithologies in Brazil
Rights and permissions
Copyright information
© 2016 Springer International Publishing Switzerland
About this chapter
Cite this chapter
Schaefer, C.E. et al. (2016). The Physical Environment of Rupestrian Grasslands (Campos Rupestres) in Brazil: Geological, Geomorphological and Pedological Characteristics, and Interplays. In: Fernandes, G. (eds) Ecology and Conservation of Mountaintop grasslands in Brazil. Springer, Cham. https://doi.org/10.1007/978-3-319-29808-5_2
Download citation
DOI: https://doi.org/10.1007/978-3-319-29808-5_2
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-29807-8
Online ISBN: 978-3-319-29808-5
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)