
Abstract
From the 1960s onwards, the European wild boar population has grown and expanded dramatically, invading agricultural areas and causing increasing damage to croplands and economic losses. The problem is particularly weighty in protected areas, where hunting is forbidden. In these areas the main ways the problem is tackled is through shooting, compensation payments after claims and prevention using electric fences. The aim of this study, based on the case of Elba Island (Central Italy), was to apply a management model for wild boar populations in protected areas able to mitigate social and economic conflicts with farmers and hunters. We identified which types of crops and structures were sensitive to damage, and the months in which the events were concentrated. Leslie–Davis and Ricker models were used to estimate the wild boar population in every year. Population viability analyses (PVAs) were carried out to define the minimum control effort that can lead to a strong population decline. Finally, to identify the areas most at risk of damage, we built a predictive model, using an ensemble forecasting approach, averaging the prediction obtained following Resource Selection Probability Functions and the maximum entropy algorithm Maxent.
Damage was concentrated mainly on vineyards and dry walls. There was a significant negative trend of damage during the study period, whereas the number of wild boars shot increased significantly. Removal methods failed to provide a precise estimate of the population present on the Elba Island because of the high variability of the catch per unit effort. PVAs showed that only a heavy control effort concentrated on reproductive females is effective in markedly decreasing the population size and its survival probability. The model-average indicated that more than 45 % of the Elba Island is classified either in the high or in the maximum classes of risk, with areas most at risk of damage distributed particularly in the southern part of the island.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
Introduction
From the 1960s onwards, the wild boar ( Sus scrofa Linnaeus, 1758) has become a species of social and economic interest in Europe. In recent decades in fact, European populations have increased greatly (Apollonio et al. 2010), leading to an increase in problems, such as damage to croplands, collisions with vehicles, an impact on plant communities and ecosystems and transmission of diseases to livestock and humans (e.g. Gortázar et al. 2007; Bueno et al. 2009; Lagos et al. 2012; Puerta-Piñero et al. 2012; Li et al. 2013).
In Italy the population of Sus scrofa has dramatically increased in the last 40 years, growing from an estimate of 1900 individuals in 1977 to about 667,000 in 2004 (Meriggi et al. 2011). Moreover, the species has expanded its range, occupying all the hilly and mountainous areas of the Italian peninsula, and, more recently, several zones of the Alps and intensively cultivated plains (Merli and Meriggi 2006; Monaco et al. 2006; Carnevali et al. 2011). The wild boar is present even on many Italian islands, including Elba (Angelici et al. 2009).
This demographic explosion is due to various reasons: the extensive recovery of natural woodlands, the adaptability of the species to a wide range of environmental conditions, its great reproductive capacity, repeated releases of hand-reared animals for hunting and escapes of individuals from farms (Merli and Meriggi 2006; Apollonio et al. 2010; Ficetola et al. 2014; Massei et al. 2015).
The growth of the population has produced an increase of damage to croplands; at present, compensation payments for wild boar damage in Italy amount to about nine millions of Euros per year (Riga et al. 2011). The problem of damage is particularly important in protected areas (Parks, Natural Reserves, Wildlife Refuges). In these areas hunting is forbidden; moreover, hunting activity near boundaries induces the boar to concentrate within protected areas, with a consequent increase in damage to agricultural activities and natural habitats (Monaco et al. 2010), often unsustainable for the financial resources available. In Italy, the wild boar is the most important game ungulate; 200,000 wild boars were harvested in 2012, mainly in Northern Apennines and central Italy, but this number probably represents an underestimate of the true number of heads shot per year (Apollonio et al. 2010; Massei et al. 2015). All these factors demonstrate the need for effective management strategies that reconcile the objective of conservation in protected areas and the presence of the wild boar as a big game species in surrounding hunting districts.
In Italian protected areas, the response to damage is prevention, compensation and population control by capture, culling and shooting (Monaco et al. 2010). Shooting is used as a control method for wild boar in many parts of the world (Massei et al. 2011). It has been demonstrated to be very effective, particularly in high-density areas and with a great hunting effort, but its success can be limited by compensatory responses exhibited by boar, such as increased immigration and reproduction (Hanson et al. 2009). Therefore, effective numerical control of wild boar in protected areas could be economically unsustainable, because the reduction of density is only temporary, and control shooting may become so intense as to become the main activity of a protected area. Prevention with electrified fences can be very effective (e.g. Santilli and Mazzoni Della Stella 2006; Reidy et al. 2008), but, considering the high cost for buying and maintaining the materials, it is normally only recommended for fields of high economic value or for high-risk situations (Santilli and Mazzoni Della Stella 2006).
To improve cost-effective strategies for damage prevention, it is important to identify which factors affect the risk of damage caused by wild boar. Species distribution models allow identification of relationships between species occurrence (e.g. presence, abundance, damage) and environmental factors (e.g. land use, topography, landscape features, climate) (Elith and Leathwick 2009; Jiménez-Valverde et al. 2011).
Wild boar damage is affected by many factors, including both security and food availability. Safety factors include the distance to the edge of the nearest forests, roads, rivers, and the human presence (Calenge et al. 2004; Cocca et al. 2007; Honda and Sugita 2007; Thurfjell et al. 2009). Food factors include types, abundance, maturation time of crops and availability of agricultural lands (Herrero et al. 2006; Schley et al. 2008; Li et al. 2013).
Our aim is to define a management model for wild boar populations in protected areas able to mitigate social and economic problems. Moreover, we would like to (1) verify the effectiveness of shooting as a prevention method, (2) define the minimum control effort that could lead to a relevant population decline and (3) identify the areas most at risk of damage with a predictive model.
Study Area
The Elba Island
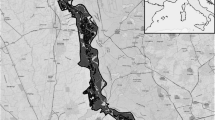
The study was conducted on Elba Island (in the Tuscan Archipelago, Central Italy), with an area of 223.2 km2 and a maximum altitude of 1016 m a.s.l. (Mount Capanne) (Fig. 11.1). The climate is Mediterranean, with a mean yearly temperature of 16.5 °C (minimum 10 °C in January, maximum 24.5 °C in July) and a mean yearly precipitation of 595 mm (minimum 13 mm in July, maximum 86 mm in November) (Chiatante et al. 2013). The island is covered by Mediterranean maquis with strawberry tree (Arbutus unedo), heather (Erica arborea), Cistus spp., rosemary (Rosmarinus officinalis) and lavender (Lavandula stoechas) (37.2 %), holm-oak (Quercus ilex) forests (26.2 %), urban areas (11.2 %), agricultural areas (10.2 %), pine plantations (7.0 %) and meadows (4.1 %). The surface of the island used for farming is partitioned among cereal crops, vineyards, orchards and olive groves; animal husbandry is based primarily on goats (Fifth National Census of Agriculture—Istituto Nazionale di Statistica 2000).

Location of Elba Island (Central Italy, Tyrrhenian Sea). The Tuscan Archipelago National Park is in grey
Over half of the island (127.4 km2) was designated as Tuscan Archipelago National Park (thereafter: TANP ) in 1996, the remaining part (95.8 km2) is a hunting district (ATC LI 10) (Fig. 11.1). In the National Park hunting is forbidden, although control of wild boar and mouflon (Ovis orientalis musimon) is carried out. Outside the National Park, hunting is mainly for wild boar, from October to January.
Hunting is performed by drives with hunting dogs. The hunt takes place in areas of approximately 50–120 ha; 25–30 hunters, armed and located in fixed positions, wait for the arrival of boars that are moved from their resting places by teams of dogs conducted by 6–8 persons.
History of Wild Boar on Elba Island
The presence of the wild boar on Elba Island has been reported for the first time at the beginning of the twentieth century (Damiani 1923). Then, some individuals were released for hunting in 1963 (Angelici et al. 2009). From the 1960s onwards, the presence of the species has caused a lot of problems: damage to the traditional agricultural system, the destruction of dry walls, negative effects on plant and animal biodiversity and collisions with vehicles (Giannini and Montauti 2010). Traffic accidents are particularly critical in summer, when Elba Island is visited by about two millions of tourists (Giannini and Montauti 2010).
Since 1997, to limit the negative impact of the wild boar, the administration of the TANP has developed a control programme, mostly based on trapping and hunting with hunting dogs.
The management of the wild boar on Elba Island is complicated because of the presence of different interests: the administration of the National Park wants to maintain the population at a low density, while the hunters want to maintain it at a high density. This leads to conflicts between the Park administration and the hunters, as a consequence of which Park trapping structures are often vandalized (about 30 events per year; Giannini and Montauti 2010).
Methods
General Description of Wild Boar Damages
The TANP Administration provided us data regarding all cases of crop damage caused by wild boar from 1999 to 2009 for which compensation was paid out. Any damage complained was refunded after a survey carried out by the technicians of the TANP, aimed to see if the damage was caused by wild boars and to estimate the extent of the economic damage. Each case was georeferred, and the amount of compensation and the type of crop/structure damaged (vineyards, orchards, vegetables, meadows and dry walls) were recorded.
To evaluate the trend of damage events and compensation payments during the study period, we performed regression analyses with curve estimation, putting damage variables (the number of events per year and compensation payments per year) as dependent variables and time as independent variable. A Spearman rank correlation test was performed to evaluate the relationship between (1) compensation payments and the number of damage events recorded and (2) the damage area and the number of damage events recorded. Finally, we looked for a seasonality of the distribution of damage events using a χ 2 goodness-of-fit test.
Relation between the Number of Wild Boars Removed and Damage
We collected hunting data (outside the National Park) and control data (inside the National Park) from 1998 to 2009. We analysed the trend of bags using regression analyses with curve estimation, putting the number of wild boars shot per year as dependent variable and time as independent variable.
Pearson’s correlations were used to evaluate (1) relations between damage events and the number of wild boars removed in the same year and (2) relations between damage events and the number of wild boars removed in the previous year. We performed correlation analyses considering the number of wild boars shot only in the National Park and those from the whole Elba Island.
Wild Boar Population Estimate
Hunting and control statistics were used to estimate the wild boar population in every year by removal methods: we used Leslie–Davis regression model (Leslie and Davis 1939) and Ricker model (Ricker 1954). Both methods require that three assumptions are satisfied: (1) the population is closed; (2) the probability of each individual being shot is constant throughout the experiment and (3) all individuals have the same probability of being shot in the time t. Tacking account that the area under study is an island and the hunting method adopted, we considered these assumptions realistic.
According to Leslie–Davis model, the population size at the time t (before starting to shoot boars) could be estimated with a linear regression between catch per unit effort and accumulated catch. In our case, we considered the accumulated harvest (x) and the harvest per unit effort (y). Ricker model presupposes a linear relationship between the natural logarithm of catch per unit effort (y) and the accumulated effort (x) (Krebs 1999).
Population Viability Analysis
Hunting data were useful to assess the demographic parameters of the wild boar on Elba Island (Meriggi et al. 2010) (Table 11.1). We used these parameters to run PVAs, which have been demonstrated to be an accurate tool for predicting population viability (Brook et al. 2000). The PVA is a quantitative approach useful to assess the effects of demographic, environmental and genetic stochasticity and different management practices on wildlife populations. It is largely used in conservation biology to predict extinction risks for threatened species and to compare alternative options for their management (e.g. Galimberti et al. 2001; Chilvers 2012; Carroll et al. 2013). In our case, we run PVA s with the main aim of determining the harvest effort necessary to cause population extinction in the TANP over a 30-year period.
We carried out PVAs starting with a population of 2500 wild boars (12.6 per km2) and estimating a carrying capacity of 3000 animals (15.1 per km2). We calculated the initial population size considering the number of boars harvested in 2009 (1500 heads) and a harvesting rate of 60 %; we estimated the carrying capacity considering the density range recorded for wild boar populations in Mediterranean protected areas in Italy (13–25 per km2, Massei et al. 1996; Massei and Genov 2000). We run PVAs simulating five different scenarios: (1) maintaining the present harvesting rate; (2) increasing the harvesting rate by 10 % in the first 5 years equally distributed on all age classes and on both sexes; (3) increasing the harvesting rate by 30 % in the first 5 years equally distributed on all age classes and on both sexes; (4) increasing the harvesting rate by 40 % in the first 5 years only on reproductive females and (5) increasing the harvesting rate by 40 % in the first 10 years only on reproductive females. PVAs were carried out with the software VORTEX (version 9.98) (Lacy 2000).
Factors Influencing Damage Distribution
In order to define the areas with the highest risk of wild boar damage, we applied a 1 km2-spaced grid to the study area that identified 278 individual cells. In all cells, using ArcMap v. 9.3 GIS software (ESRI, Redlands, USA), we measured the proportion of 17 habitat variables concerning vegetation cover, 11 variables of elevation (corresponding to 11 different classes of altitude), 7 variables of slope (corresponding to 7 different classes of slope) and 9 variables of exposure (Table 11.2). Data on habitat types were taken from a 1:25,000 digital vegetation map (Foggi et al. 1996), while elevation, slope and exposure data were derived from a Digital Terrain Model (DTM; cell size 20 m) produced by the Italian Military Geographic Institute.
We predicted the areas with the highest risk of damage following a model-averaging approach, combining the results obtained with two different methods: (1) RSPFs, following a use vs. availability approach obtained by Binary Logistic Regression Analysis (thereafter: BLRA) (Pearce and Ferrier 2000; Boyce et al. 2002; Keating and Cherry 2004) and (2) Maximum Entropy algorithm (MaxEnt version 3.3.3) (Phillips et al. 2006; Elith et al. 2011), which requires only presence data.
Resource Selection Probability Functions
We carried out BLRA comparing the features of damaged cells and of an equal number of cells randomly placed over the island. We defined the cells as “damaged” by the species if at least one damage event fell inside. We built models with all possible combinations of habitat variables. Inference from models was made according to the Information-Theoretic Approach (Anderson et al. 2000, 2001; Richards et al. 2011), using corrected Akaike Information Criterion (AICc) because of the small ratio of sample/parameters (Akaike 1973; Anderson et al. 2000, 2001; Johnson and Omland 2004). Models were ranked and scaled by the differences with minimum AICc (ΔAICc) and Akaike weights (ωi) for each i-model (Anderson et al. 2000; Burnham and Anderson 2002). Models with ΔAICc ≤ 2 were considered the best ones and used to develop model averaging (Burnham and Anderson 2002; Massolo and Meriggi 2007). The relative importance of predictor variables (ω) was measured, as resulted from the best models, by the sum of Akaike weights of the models in which each variable appeared (Burnham and Anderson 2002; Merli and Meriggi 2006).
Model validation was achieved through a k-fold crossvalidation process. We partitioned our data into two random subsets (Fielding and Bell 1997; Boyce et al. 2002; Chiatante et al. 2013); the former was used to calibrate the model and the latter was used to evaluate the result. We replicated this process two times, using each subset in turn for validation purpose, and we measured the model accuracy using the Boyce index (Boyce et al. 2002).
Maxent
Maxent is a general-purpose machine learning method (Phillips et al. 2006), widely used in species distribution modelling in a large range of taxons and across many disciplines (i.e. biological invasions, evolution, conservation; Ward 2007; Monterroso et al. 2009; Isaac et al. 2013; Ficetola et al. 2014), which follows a presence-only modelling approach.
Presence-only data consist of records describing known occurrences (presence) of species without information about known absences. The collection of real absence data is one of the major problems to solve; while collecting reliable data on animal presence is straightforward in most case studies, it is more difficult to verify the true absence of a species in a given habitat, especially when the target species occurs at low densities, is cryptic or elusive, thus resulting in a low detection probability (Santos et al. 2006; Isaac et al. 2013). For all these reasons, the absence of observations at a given location cannot be reliably interpreted as a true absence, thus it is necessary to rely only on presence data.
Maxent approach assesses the likelihood of presence in a given cell on the basis of the environmental features of that cell, establishing flexible relationships between the dependent and independent variables (Elith et al. 2011). It finds the most uniform species distribution with the constraint that the expected value for each variable should match the average value of a set of sample points taken from the target-species distribution (Phillips et al. 2006). The probability distribution is exponential, ranging from 0 to 1. This is achieved by dividing the sum of weighted probability values by a scaling constant.
The area under the Receiver Operating Characteristics curve (AUC) was used to examine model performance. AUC provides a measure of discrimination ability, varying from 0.5 for a model with discrimination ability no better than random to 1.0 for a model with perfect discriminatory ability.
A rough guide for classifying the accuracy of a diagnostic test is the traditional academic point system (Swets 1988): 0.91–1.00 = excellent; 0.81–0.90 = good; 0.71–0.80 = fair; 0.61–0.70 = poor; 0.51–0.60 = fail.
Damage Risk Map
For both methods, we computed a damage risk (DR) map, which indicates how the combination of the ecogeographical variables of a single cell determines the probability of risk of damage. All maps were classified in five risk groups: null (0.00–0.20), low (0.21–0.40), medium (0.41–0.60), high (0.61–0.80) and maximum (0.81–1.00).
Finally, to reduce the model-based uncertainty in range predictions, we used an ensemble forecasting approach, which represents one of the most efficient consensus methods (Araújo and New 2007), computing the average probability of damage predicted by the two methods.
Results
General Description of Wild Boar Damage
From 1999 to 2009 we recorded a total of 256 damage events in the TANP (mean per year ± SD: 23.3 ± 13.0 events). Damage was concentrated mainly on vineyards (46 % of total events) and dry walls (27 %), but was also reported on orchards (11 %), meadows (9 %) and vegetables (7 %).
There was a significant negative trend of damage during the study period; both the number of events and compensation payments decreased following a logarithmic model (number of events: y = 43.2 − 12.5 ln(x), r 2 = 0.462, F = 9.57, p = 0.013; compensation payments: y = 19.5 − 6.4ln(x), r 2 = 0.447, F = 9.09, p = 0.015).
Both the amount of compensation payments and the damage area were strongly correlated with the number of damage events (rho = 0.89, p = 0.001 and rho = 0.92, p = 0.001, respectively).
We observed a significant logarithmic reduction in the number of damage events to vineyards (y = 32.9 − 12.2ln(x), r 2 = 0.692, F = 23.50, p = 0.001), although for the other types of fields and for dry walls there were not significant trends (p > 0.05) (Fig. 11.2).

(a) Trend of wild boar damage (1999–2009), (b) trend of refunds paid for wild boar damage (1999–2009) and (c) trend of wild boar damage to vineyards (1999–2009)
There were strong monthly differences in the distribution of damage events (χ 2 = 400.03; df = 11; p < 0.001), with a peak recorded in August and September and a minimum in March, November and December. Damage to vineyards almost exclusively occurred in August and September, damage to orchards was concentrated from July to September, damage to vegetables occurred mainly in summer, damage to meadows was recorded throughout the year, while damage to dry walls almost exclusively occurred in spring and summer. Few events were recorded in autumn and winter (Fig. 11.3).

Monthly distribution of wild boar damage
Relation between the Number of Wild Boars Removed and Damage
The number of wild boars shot increased significantly from 1998 to 2009, following a linear model. This growth was observed cumulating data (y = 584.4 + 53.9x, r 2 = 0.409, F = 8.61, p = 0.015) and considering only the animals killed inside the TANP (y = 292.2 + 43.1x, r 2 = 0.282, F = 5.32, p = 0.044) and in the hunting district (y = 240.4 + 13.6x, r 2 = 0.329, F = 6.39; p = 0.030) (Fig. 11.4).

Trend in the number of wild boar killings on (a) the whole of Elba Island (1998–2009), (b) Tuscan Archipelago National Park (1998–2009) and (c) the hunting district of Elba Island (1998–2009)
No significant correlations were found between the amount of damage and the number of wild boars removed (Table 11.3).
Wild Boar Population Estimate
Considering harvest data, we obtained significant regressions only for 2004 and 2006 with both Leslie–Davis and Ricker method; considering control data, we obtained significant regressions for 1999 and 2002 with Leslie–Davis method and only for 2002 with Ricker method (Table 11.4). We did not find significant differences between the estimates obtained with the two methods.
Population Viability Analysis
Maintaining the current harvesting rate, the wild boar population decreased by about 30 % within the first 5 years. In the following years, the population still decreased, but more slowly, and stabilized after about 15 years. The survival probability decreased linearly over the simulation period, but it always maintained a high value (more than 0.6) (Fig. 11.5).

Population dimension and survival probability of the Elba Island wild boar population on the basis of a PVA simulating the current harvesting rate
Simulating an increased harvesting rate of 10 and 30 % on all the age classes and on both sexes, we observed a strong decrease in the population size only at the 30 % level. Nevertheless, the effect of this stronger control action had little long-term impact: the population started to grow up immediately at the end of the control period, returning to values of about 1000–1200 individuals in a few years. The survival probability was very similar comparing the two different scenarios: it decreased linearly during the simulation period, but it maintained a high value, of about 0.6 (Fig. 11.6).

Population size and survival probability of the Elba Island wild boar population on the basis of a PVA simulating an increased harvesting rate equally distributed on all age classes and on both sexes. The black line indicates an increased rate of 10 %, the grey line indicates an increased rate of 30 %
Simulating an increased harvesting rate by 40 % only on adult females, we observed a strong decrease of the population size; the population decreased to 437 individuals in the case of a control period of 5 years, and to only 97 individuals when the control was carried out for 10 years. In both cases, the population started to grow immediately at the end of the control period, returning to 980 and 573 individuals, respectively, at the end of the simulation period. In the case of a control period of 5 years, the survival probability decreased linearly reaching the value of 0.49 at the end of the simulation period, whereas with a control period protracted for 10 years there was a strong decrease of the survival probability, which reached the value of only 0.22 at the end of the simulation period (Fig. 11.7).

Population size and survival probability of the Elba Island wild boar population on the basis of a PVA simulating an increased harvesting rate by 40 % only on reproductive females. The black line indicates an increased rate for 10 years, the grey line indicates an increased rate for 5 years
Factors Influencing Damage Distribution
Resource Selection Probability Functions
In the study area we found a total of 56 cells with presence of wild boar damage. By multimodel inference we selected three models, which included a total of seven variables (Table 11.5). The most important variable positively affecting the probability of wild boar damage were the areas between 10° and 20° of slope, while for the other variables the effect was uncertain (Table 11.6). Crossvalidation results indicated an excellent predictive power of the model: calculating the Boyce Index we measured a Spearman rank coefficient of 0.99 (p < 0.001).
Maxent
The areas between 100 and 300 m a.s.l., European hop-hornbeam woods, shrublands, rocky areas, arable lands, grasslands and lentisk-dominated maquis were the most important variables in the prediction of the areas most at risk of damage. All these variables positively affected the probability of wild boar damages (Table 11.7).
The evaluation of the model performance with AUC indicated a fair predictive power of the model, with an area under the ROC curve of 0.78 (p < 0.001).
Damage Risk Map
Combining the two models more than 45 % of Elba Island was classified either in the high or maximum classes of risk (respectively, 29.1 % and 18.8 %). Of the remaining area, 29.1 % was classified in the medium risk, 19.4 % in the low risk and only 3.6 % in the null risk class. The areas with the highest risk of damage (medium, high or maximum) were distributed throughout the island (Fig. 11.8) but particularly in the south, in the central-eastern part.

Damage risk map for wild boar damages on Elba Island obtained following an ensemble forecasting approach
Discussion
Removal methods failed to provide a precise estimate of the population present on the Elba Island because of the high variability of the catch per unit effort; moreover, even when we obtained significant regressions, confidence intervals were very wide. The lack of an accurate measure of the population size could represent a great problem for wild boar management. Probably only with a heavy hunting and control pressure it is possible to use removal methods to obtain precise and accurate population estimates, otherwise it is necessary to adopt other census methods. But methods such as spotlight counts, aerial surveys and vantage point counts are difficult to carry out in Mediterranean environments, because the habitat is characterized by tall and dense vegetation. Thus, it is necessary to use different methods, like tracking plots and dung counts. These methods have been successfully used to monitor wild boar abundance (Engeman et al. 2001, 2013), a parameter that is often correlated with density (Hone 1995).
Hunting data are often used to assess density and relative abundance of wild boar populations (e.g. Boitani et al. 1994; Fernández-Llario et al. 2003; Bosch et al. 2012; Sarasa and Sarasa 2013). The application of this technique requires several assumptions; the existence of a linear relationship between catch and population abundance, the use of standardized methods of removal, the knowledge of all removals (Mitchell and Balogh 2007; Engeman et al. 2013). Unfortunately, most of these assumptions are difficult to meet or unknown, potentially biasing results.
To reduce bias, hunt indices require large samples (Siren et al. 2004). In our case, we collected systematically data regarding hunting bags over the 11 years of study, and the TANP administration did the same regarding claims for compensation. For these reasons, we are confident that these data could be used for the interpretation of our results.
Our results show a negative trend of wild boar damage on Elba Island from 1999 to 2009. Vineyards were the most frequently damaged crops. Damage monitoring in Europe mainly regards maize, other cereals and grasslands, while vineyards are less recorded, but comparisons with other countries are difficult because of the differences between their agricultural mosaics (Herrero et al. 2006; Schley et al. 2008; Novosel et al. 2012). On Elba Island, damage to vineyards almost exclusively occurred from August to October, in correspondence of the ripening period of grapes.
The number of boars shot increased during the study period, but the lack of correlation between wild boars removed and damage does not allow us to assume a positive effect of shooting in determining the reduction of damage events, damage area and compensation payments. In some cases, hunting has been demonstrated to be an effective solution in reducing damage to crops (Mazzoni Della Stella et al. 1995; Geisser and Reyer 2004). However, combination of shooting with prevention methods could increase the efficiency in damage reduction (McCann and Garcelon 2008; Massei et al. 2011).
Supplementary feeding (e.g. with maize) has been demonstrated to be a very effective tool to reduce the level of damage to vineyards in southeastern France (Calenge et al. 2004), which is very similar to Elba Island regarding the habitat composition. It is essential to distinguish between the “dissuasive” and the “attractive” food supply. In several areas, hunters spread maize throughout the year to attract boars to their hunting territories. This additional food may result in an increase in population size and leads to a long-term increase in the number of damage events (Geisser and Reyer 2004; Schley et al. 2008). On Elba Island, boars are not artificially fed. We strongly warn against this practice, and we stress that the dissuasive spreading of maize is different both in its aim and in its results. When used as a deterrent, the maize is spread only over a very short period, as long as the grapes are ripe. In fact, the dissuasive spreading of maize is a replacement food source rather than an additional food source (Calenge et al. 2004).
PVAs showed that only a heavy control effort concentrated on reproductive females is effective in markedly decreasing the population size and the survival probability of wild boar. Increasing the harvesting effort on reproductive females has been demonstrated to be the most efficient way to stop wild boar population growth (Bieber and Ruf 2005; Servanty et al. 2011). In our case, the effect is stronger simulating a control effort protracted for a longer period; we observed a reduction of the population size of 96 % after 10 years, and a reduction of the survival probability of 78 % after 30 years. This great result, however, is very difficult to reach in practice because of the great engagement and economic effort needed.
Our model identified landscape features that increase the risk of crop damage : the risk was highest in low altitude areas (100–300 m a.s.l.) with a high presence of arable lands, grasslands, European hop-hornbeam woods and shrublands. The high relative importance of arable lands and grasslands could be related to wild boar nutrition. Agricultural plants represent the main food resource for wild boar in several areas (Schley and Roper 2003; Herrero et al. 2006); wild boar damage to grasslands is caused by direct consumption and by rooting activity linked to the search for invertebrates, roots and bulbs (Baubet et al. 2004; Schley et al. 2008). Acorns and maize are higher in carbohydrates and fats, but have lower crude protein content than grasslands and earthworms (Massei et al. 1996; Baubet et al. 2004; Schley et al. 2008), so wild boars could have to counterbalance their diet with animal foods and graminoids. The positive relationship with shrublands and hop-hornbeam woods likely occurs because of the high suitability of this habitat for wild boars (Meriggi and Sacchi 2001; Schley et al. 2008; Honda and Kawauchi 2011). In fact, they can find alternative shelter areas in the garrigue and in the Mediterranean maquis, which are rife in the study area and, like woodlands, can provide a good cover.
Damage modelling produced a map of the risk with a satisfactory predictive power; the risk maps could be used to plan preventive measures with the aim of improving the effectiveness of management both in the short- and long-term perspectives. First, these maps can help to identify the areas more suitable for population monitoring schemes, which will allow a better knowledge of population dynamics, and can provide essential information for a more effective management of populations. Collecting standardized data and regularly updating the databases is necessary for an adaptive management of populations (Ficetola et al. 2014). Second, the maps could allow to identify crops most at risk that can be protected through fences (mechanical or electrified), feeding plots in forested areas or numerical shooting control.
Our ensemble forecasting approach will help to overcome reliance on a single modelling method. With recent advances in sophisticated species distribution modelling approaches (e.g. BIOMOD, Thuiller et al. 2009), wildlife ecologists will be able to construct robust ensemble models of up to a dozen different modelling approaches.
Conclusions
The goal of the wild boar management of the TANP is a drastic reduction of damage and of conflicts between the Park administration and the local population. This study, both with correlation analyses between killings and damages and with PVAs, highlighted the importance of a heavy control targeted on specific age or size classes as a powerful instrument in the reduction of damage. However, a sufficiently heavy numerical control is difficult to sustain in the long term, so control activity should be combined with damage prevention by stable or electrified fences and supplementary feeding, especially to protect vineyards, which represent the main crop damaged and, consequently, the main source of conflicts.
Models of damage risk can be an effective tool to address prevention effort with more effectiveness and lower costs. In particular, the ensemble forecasting approach, with its great robustness given by the use of different models which are based on different principles, seems to be an efficient method in predicting the areas most at risk of damage.
More research is needed to improve management strategies of wild boar on Elba Island; firstly, it is necessary to provide accurate density or abundance estimates, with drive counts, tracking plots or dung counts. Secondly, it is necessary to study the diet of the wild boar on Elba Island; only the knowledge of the foraging behaviour of the wild boar will allow understanding clearly the impact of the species on croplands and natural ecosystems. Besides the study of the feeding habits, it should be important monitoring the yearly availability of natural food resources (i.e. acorns and chestnuts), which often influences wild boar population dynamics and crop damage (Bieber and Ruf 2005; Servanty et al. 2009; Cutini et al. 2013).
References
Akaike H (1973) Information theory as an extension of the maximum likelihood principle. In: Petrov BN, Csaki F (eds) Second international symposium on information theory. Akademiai Kiado, Budapest, pp 267–281
Anderson DR, Burnham KP, Thompson WL (2000) Null hypothesis testing: problems, prevalence and an alternative. J Wildl Manag 64:912–923
Anderson DR, Link WA, Johnson DH, Burnham KP (2001) Suggestion for presenting the results of data analyses. J Wildl Manag 65:373–378
Angelici FM, Laurenti A, Nappi A (2009) A checklist of the mammals of small Italian islands. Hystrix 20:3–27
Apollonio M, Andersen R, Putman R (2010) European ungulates and their management in the 21st century. Cambridge University Press, Cambridge
Araújo MB, New M (2007) Ensemble forecasting of species distributions. Trends Ecol Evol 22:42–47
Baubet E, Bonenfant C, Brandt S (2004) Diet of the wild boar in the French Alps. Galemys 16:101–113
Bieber C, Ruf T (2005) Population dynamics in wild boar Sus scrofa: ecology, elasticity of growth rate and implications for the management of pulsed resource consumers. J Appl Ecol 42:1203–1213
Boitani L, Mattei L, Nonis D, Corsi F (1994) Spatial and activity patterns of wild boars in Tuscany. J Mammal 75:600–612
Bosch J, Peris S, Fonseca C, Martinez M, De la Torre A, Iglesias I, Muñoz MJ (2012) Distribution, abundance and density of the wild boar on the Iberian Peninsula, based on the CORINE program and hunting statistics. Folia Zool 61:138–151
Boyce MS, Vernier PR, Nielsen SE, Schmiegelow FKA (2002) Evaluating resource selection functions. Ecol Model 157:281–300
Brook BW, O’Grady JJ, Chapman AP, Burgman MA, Akçakaya HR, Frankham R (2000) Predictive accuracy of population viability analysis in conservation biology. Nature 404:385–387
Bueno CG, Alados CL, Gómez García D, Barrio IC, García-González R (2009) Understanding the main factors in the extent and distribution of wild boar rooting on alpine grasslands. J Zool 279:195–202
Burnham KP, Anderson DR (2002) Model selection and multimodel inference. A practical information—theoretic approach, 2nd edn. Springer, New York
Calenge C, Maillard D, Fournier P, Fouque C (2004) Efficiency of spreading maize in the garrigues to reduce wild boar (Sus scrofa) damage to Mediterranean vineyards. Eur J Wildl Res 50:112–120
Carnevali L, Di Luzio P, Riga F (2011) Ungulati in Italia, Distribuzione e consistenza. In: Riga F, Genghini M, Cascone C, Di Luzio P (eds) Impatto degli ungulati sulle colture agricole e forestali: proposta per linee guida nazionali. Manuali e Linee Guida ISPRA 68/2011, pp 3–19
Carroll C, Fredrickson RJ, Lacy RC (2013) Developing metapopulation connectivity criteria from genetic and habitat data to recover the endangered Mexican wolf. Conserv Biol 28:76–86
Chiatante G, Meriggi A, Giustini D, Baldaccini NE (2013) Density and habitat requirements of red-legged partridge on Elba Island (Tuscan Archipelago, Italy). Ital J Zool 80:402–411
Chilvers BL (2012) Population viability analysis of New Zealand sea lions, Auckland Islands, New Zealand’s sub-Antarctics: assessing relative impacts and uncertainty. Polar Biol 35:1607–1615
Cocca G, Sturaro E, Dal Compare L, Ramanzin M (2007) Wild boar Sus scrofa damages to mountain grassland: a case study in the Belluno province, Eastern Italian Alps. Ital J Anim Sci 6:845–847
Cutini A, Chianucci F, Chirichella R, Donaggio E, Mattioli L, Apollonio M (2013) Mast seeding in deciduous forests of the northern Apennines (Italy) and its influence on wild boar population dynamics. Ann For Sci 70:493–502
Damiani G (1923) La Fauna. In: L’Elba illustrata, pp 103–129
Elith J, Leathwick JR (2009) Species distribution models: ecological explanation and prediction across space and time. Annu Rev Ecol Evol Syst 40:677–697
Elith J, Phillips SJ, Hastie T, Dudík M, En Chee Y, Yates CJ (2011) A statistical explanation of MaxEnt for ecologists. Divers Distrib 17:43–57
Engeman RM, Constantin B, Nelson M, Woolard J, Bourassa J (2001) Monitoring changes in feral swine population and spatial distribution. Environ Conserv 28:235–240
Engeman RM, Massei G, Sage M, Gentle MN (2013) Monitoring wild pig populations: a review of methods. Environ Sci Pollut Res 20:8077–8091
Fernández-Llario P, Mateos-Quesada PM, Silvério A, Santos P (2003) Habitat effects and shooting techniques on two wild boar (Sus scrofa) populations in Spain and Portugal. Z Jagdwiss 49:120–129
Ficetola GF, Bonardi A, Mairota P, Leronni V, Padoa-Schioppa E (2014) Predicting wild boar damages to croplands in a mosaic of agricultural and natural areas. Curr Zool 60:170–179
Fielding AH, Bell JF (1997) A review of methods for the assessment of prediction errors in conservation presence/absence models. Environ Conserv 24:38–49
Foggi B, Cartei L, Pignotti L, Signorini MA, Viciani D, Dell’Olmo L, Menicagli E (1996) Il paesaggio vegetale dell’Isola d’Elba (Arcipelago Toscano): studio fitosociologico e cartografico. Fitosociologia 43:3–94
Galimberti F, Sanvito S, Boitani L, Fabiani A (2001) Viability of the southern elephant seal population of the Falkland Islands. Anim Conserv 4:81–88
Geisser H, Reyer HU (2004) Efficacy of hunting, feeding, and fencing to reduce crop damage by wild boars. J Wildl Manag 68:939–946
Giannini F, Montauti G (2010) Controllo del Cinghiale e conflitti sociali: il caso del Parco Nazionale Arcipelago Toscano. In: Monaco A, Carnevali L, Toso S (eds) Linee guida per la gestione del cinghiale (Sus scrofa) nelle aree protette, pp 86–87
Gortázar C, Ferroglio E, Höfle U, Frölich K, Vicente J (2007) Diseases shared between wildlife and livestock: a European perspective. Eur J Wildl Res 53:241–256
Hanson LB, Mitchell MS, Grand JB, Jolley DB, Sparklin BD, Ditchkoff SS (2009) Effect of experimental manipulation on survival and recruitment of feral hogs. Wildl Res 36:185–191
Herrero J, García-Serrano A, Couto S, Ortuño VM, García-González R (2006) Diet of wild boar Sus scrofa L. and crop damage in an intensive agroecosystem. Eur J Wildl Res 52:245–250
Honda T, Kawauchi N (2011) Methods for constructing a wild boar relative-density map to resolve human-wild boar conflicts. Mamm Stud 36:79–85
Honda T, Sugita M (2007) Environmental factors affecting damage by wild boars (Sus scrofa) to rice fields in Yamanashi Prefecture, central Japan. Mamm Stud 32:173–176
Hone J (1995) Spatial and temporal aspects of vertebrate pest damage with emphasis on feral hogs. J Appl Ecol 32:311–319
Isaac B, White J, Ieradiaconou D, Cooke R (2013) Response of a cryptic apex predator to a complete urban to a forest gradient. Wildl Res 40:427–436
Jiménez-Valverde A, Peterson AT, Soberón J, Overton JM, Aragón P, Lobo JM (2011) Use of niche models in invasive species risk assessments. Biol Invasions 13:2785–2797
Johnson JB, Omland KS (2004) Model selection in ecology and evolution. Trends Ecol Evol 19:101–108
Keating KA, Cherry S (2004) Use and interpretation of logistic regression in habitat-selection studies. J Wildl Manag 68:774–789
Krebs CJ (1999) Ecological methodology, 2nd edn. Addison-Wesley, Toronto
Lacy RC (2000) Structure of the VORTEX simulation model for population viability analysis. Ecol Bull 48:191–203
Lagos L, Picos J, Valero E (2012) Temporal pattern of wild ungulate-related traffic accidents in northwest Spain. Eur J Wildl Res 58:661–668
Leslie PH, Davis DHS (1939) An attempt to determine the absolute number of rats on a given area. J Anim Ecol 8:94–113
Li L, Shi J, Wang J, Gao Y, Wang L, Wang J, Ying X (2013) Factors influencing wild boar damage in Taohongling National Nature Reserve in China: a model approach. Eur J Wildl Res 59:179–184
Massei G, Genov PV (2000) Il Cinghiale. Calderini Edagricole, Bologna, 189pp
Massei G, Genov PV, Staines BW (1996) Diet, food availability and reproduction of wild boar in a Mediterranean coastal area. Acta Theriol 41:307–320
Massei G, Roy S, Bunting R (2011) Too many hogs? A review of methods to mitigate impact by wild boar and feral hogs. Hum Wildl Interact 5:79–99
Massei G, Kindberg J, Licoppe A, Gačić D, Šprem N, Kamler J, Baubet E, Hohmann U, Monaco A, Ozoliņš J, Cellina S, Podgorski T, Fonseca C, Markov N, Pokorny B, Rosellp C, Nahlikq A (2015) Wild boar populations up, numbers of hunters down? A review of trends and implications for Europe. Pest Manag Sci 71:492–500. doi:10.1002/ps.3965
Massolo A, Meriggi A (2007) I modelli di valutazione ambientale per le potenzialità faunistiche: aspetti metodologici e stato in Italia. In: Lucifero M, Genghini M (eds) Valorizzazione agroforestale e faunistica dei territori di collina e montagna. INFS, MIPAF, S.t.e.r.n.a. Ed. Grafiche 3B, Toscanella di Dozza (BO), pp 43–77
Mazzoni Della Stella R, Calovi F, Burrini L (1995) The wild boar management in a province of central Italy. J Mt Ecol 3:213–216
McCann BE, Garcelon DK (2008) Eradication of feral hogs from Pinnacles National Monument. J Wildl Manag 72:1287–1295
Meriggi A, Sacchi O (2001) Habitat requirements of wild boars in the northern Apennines (N Italy): a multi-level approach. Ital J Zool 68:47–55
Meriggi A, Milanesi P, Brangi A, Lamberti P (2010) Indagine sui danneggiamenti da cinghiale (Sus scrofa) nel Parco Nazionale dell’Arcipelago Toscano e sull’efficacia dei metodi di controllo della popolazione. Università di Pavia, 100 pp
Meriggi A, Brangi A, Schenone L, Signorelli D, Milanesi P (2011) Changes of wolf (Canis lupus) diet in relation to the increase of wild ungulate abundance. Ethol Ecol Evol 23:1–17
Merli E, Meriggi A (2006) Using harvest data to predict habitat-population relationship of the wild boar Sus scrofa in Northern Italy. Acta Theriol 51:383–394
Mitchell B, Balogh S (2007) Monitoring techniques for vertebrate pests—feral pigs. NSW Department of Agriculture, Orange
Monaco A, Carnevali L, Riga F, Toso S (2006) Il cinghiale sull’arco alpino: status e gestione delle popolazioni. Centro di Ecologia Alpina 38:5–23
Monaco A, Carnevali L, Toso S (2010) Linee guida per la gestione del cinghiale (Sus scrofa) nelle aree protette. Quaderni di Conservazione della Natura 34, Ministero dell’Ambiente—ISPRA
Monterroso P, Brito JC, Ferreras P, Alves PC (2009) Spatial ecology of the European wildcat in a Mediterranean ecosystem: dealing with small radio-tracking datasets in species conservation. J Zool 279:27–35
Novosel H, Piria M, Safner R, Kutnjak H, Sprem N (2012) The game damages on agricultural crops in Croatia. J Cent Eur Agric 13:631–642
Pearce J, Ferrier S (2000) Evaluating the predictive performance of habitat models developed using logistic regression. Ecol Model 133:225–245
Phillips SJ, Anderson RP, Schapire RE (2006) Maximum entropy modeling of species geographic distributions. Ecol Model 190:231–259
Puerta-Piñero C, Pino J, Gómez JM (2012) Direct and indirect landscape effects on Quercus ilex regeneration in heterogeneous environments. Oecologia 170:1009–1020
Reidy MM, Campbell TA, Hewitt DG (2008) Evaluation of electric fencing to inhibit feral pig movements. J Wildl Manag 72:1012–1018
Richards SA, Whittingham MJ, Stephens PA (2011) Model selection and model averaging in behavioural ecology: the utility of the IT-AIC framework. Behav Ecol Sociobiol 65:77–89
Ricker WE (1954) Stock and recruitment. J Fish Res Board Can 11:559–623
Riga F, Genghini M, Cascone C, Di Luzio P (2011) Impatto degli ungulati sulle colture agricole e forestali: proposta per linee guida nazionali. Manuali e Linee Guida ISPRA 68/2011
Santilli F, Mazzoni Della Stella R (2006) Electrical fencing of large farmland area to reduce crop damages by wild boars Sus scrofa. Agr Med 136:79–84
Santos X, Brito JC, Sillero N, Pleguezuelos JM, Llorente GA, Fahd S, Parellada X (2006) Inferring habitat-suitability areas with ecological modelling techniques and GIS: a contribution to assess the conservation status of Vipera latastei. Biol Conserv 130:416–425
Sarasa M, Sarasa J-A (2013) Intensive monitoring suggests population oscillations and migration in wild boar Sus scrofa in the Pyrenees. Anim Biodivers Conserv 36:79–88
Schley L, Roper T (2003) Diet of wild boar Sus scrofa in Western Europe, with particular reference to consumption of agricultural crops. Mammal Rev 33:43–56
Schley L, Dufrene M, Krier A, Frantz AC (2008) Patterns of crop damage by wild boar (Sus scrofa) in Luxembourg over a 10-year period. Eur J Wildl Res 54:589–599
Servanty S, Gaillard J-M, Toïgo C, Brandt S, Baubet E (2009) Pulsed resources and climate-induced variation in the reproductive traits of wild boar under high hunting pressure. J Anim Ecol 78:1278–1290
Servanty S, Gaillard J-M, Ronchi F, Focardi S, Baubet E, Gimenez O (2011) Influence of harvesting pressure on demographic tactics: implications for wildlife management. J Appl Ecol 48:835–843
Siren A, Hambäck P, Machoa J (2004) Including spatial heterogeneity and animal dispersal when evaluating hunting: a model analysis and empirical assessment in an Amazonian community. Conserv Biol 18:1315–1329
Swets KA (1988) Measuring the accuracy of diagnostic systems. Science 240:1285–1293
Thuiller W, Lafourcade B, Engler R, Araujo MB (2009) BIOMOD—a platform for ensemble forecasting of species distributions. Ecography 32:369–373
Thurfjell H, Ball JP, Åhlén PA, Kornacher P, Dettki H, Sjöberg K (2009) Habitat use and spatial patterns of wild boar Sus scrofa (L.): agricultural fields and edges. Eur J Wildl Res 55:517–523
Ward DF (2007) Modelling the potential geographic distribution of invasive ant species in New Zealand. Biol Invasions 9:723–735
Acknowledgements
The research was funded by the Tuscan Archipelago National Park. We thank the Hunting District n. 10 of Livorno province, that provided the hunting data from 1999 to 2009, and two anonymous referees that reviewed an early draft of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2016 Springer International Publishing Switzerland
About this chapter
Cite this chapter
Meriggi, A., Lombardini, M., Milanesi, P., Brangi, A., Lamberti, P., Giannini, F. (2016). Management of Wild Boar in Protected Areas: The Case of Elba Island. In: Angelici, F. (eds) Problematic Wildlife. Springer, Cham. https://doi.org/10.1007/978-3-319-22246-2_11
Download citation
DOI: https://doi.org/10.1007/978-3-319-22246-2_11
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-22245-5
Online ISBN: 978-3-319-22246-2
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)