
Abstract
In this chapter the role played by H2S in the physiopathology of urogenital tract revising animal and human data available in the current relevant literature is discussed. H2S pathway has been demonstrated to be involved in the mechanism underlying penile erection in human and experimental animal. Both cystathionine-β synthase (CBS) and cystathionine-γ lyase (CSE) are expressed in the human corpus cavernosum and exogenous H2S relaxes isolated human corpus cavernosum strips in an endothelium-independent manner. Hydrogen sulfide pathway also accounts for the direct vasodilatory effect operated by testosterone on isolated vessels. Convincing evidence suggests that H2S can influence the cGMP pathway by inhibiting the phosphodiesterase 5 (PDE-5) activity. All these findings taken together suggest an important role for the H2S pathway in human corpus cavernosum homeostasis. However, H2S effect is not confined to human corpus cavernosum but also plays an important role in human bladder. Human bladder expresses mainly CBS and generates in vitro detectable amount of H2S. In addition the bladder relaxant effect of the PDE-5 inhibitor sildenafil involves H2S as mediator.
In conclusion the H2S pathway is not only involved in penile erection but also plays a role in bladder homeostasis. In addition the finding that it involved in the mechanism of action of PDE-5 inhibitors strongly suggests that modulation of this pathway can represent a therapeutic target for the treatment of erectile dysfunction and bladder diseases.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
- Bladder
- Corpus cavernosum
- Erectile dysfunction (ED)
- Benign prostatic hyperplasia (BPH)
- Lower urinary tract symptoms (LUTS)
- PDE-5 inhibitor
- CBS
- CSE
- 3MST
1 Introduction
Hydrogen sulfide (H2S) presence in mammalian tissues has been known since the 1980s, but H2S has been considered mainly as a metabolic waste product with no potential physiological activity. The first evidence, indicating H2S as an endogenous mediator was published in 1996 by Abe and Kimura (1996) that described a role for this mediator in the brain. Some years, after this first evidence was published, many research groups have focused their interest on this new pathway as described in the other chapters. H2S together with nitric oxide (NO) and carbon monoxide (CO) is now classified as gasotransmitter. Like the other two gasses, it has the ability to diffuse easily through the cellular membranes. Hydrogen sulfide can also interact with proteins such as ion channels or enzymes regulating their state either directly via chemical modification such as by sulfuration and sulfhydration or indirectly via second messengers.
The role of H2S in the homeostatic control of our body is now consistently supported by the literature, and in the other chapters of the present book, its synthesis, measurement, chemistry, and interaction have been reported. This chapter will deal with the role played by H2S in the physiopathology of urogenital tract revising animal and human data available in the current relevant literature.
2 Hydrogen Sulfide in Male Sexual Function
2.1 The Male Reproductive System
Male reproductive system consists of a number of sex organs that concur to the human reproductive process localized around the pelvic region. Briefly, the main male sex organs are the penis and the testicles important for producing semen and sperm. In particular, the penis and the scrotum represent the external genital organs. The penile erectile apparatus consists of two vascularized paired corpora cavernosa. The corpus spongiosum together with the urethra is related to the ventral area of the penis. The corpora cavernosa, the corpus spongiosum, and the glans penis are composed of septa of smooth muscle and erectile tissue that enclose vascular cavities. The tunica albuginea forms a thick fibrous coat to the spongy tissue of the corpora cavernosa and corpus spongiosum, and it consists of two layers. The muscles involved in penile function are the ischiocavernosum and bulbospongiosus. The ischiocavernous muscle inserts into the medial and inferior surface of the corpora and increases penile turgidity during erection. In the flaccid state, the smooth muscle fibers are tonically contracted by the sympathetic system. During erection, trabeculae smooth muscle is relaxed, and this allows a cascade of events. The arterioles of the penis are dilated while the veins are passively compressed between the tunica albuginea and the peripheral sinusoids reducing the venous outflow. This process is known as the venous-occlusive mechanism, and it leads to erection that is sustained by an increase of intra-cavernous blood pressure of about 100 mmHg. The contraction of the ischiocavernous muscle further increases the intra-cavernous pressure leading to the rigid erection phase.
The main role of the scrotum is to hold and protect the testes. It also contains numerous nerves and blood vessels. Between the male internal genital organs, there is the epididymis, a whitish mass of tightly coiled tubes cupped against the testicles. It acts as a maturation and storage for sperm before they pass into the vas deferens, which carry sperm to the ampullary gland and prostatic ducts. Testosterone is the most important sexual hormone in males and is released by the testes. This hormone has a crucial role in the development of sperm and is also responsible for the development of physical characteristics in men such as facial hair and deep voice. Moreover the accessory glands, such as the seminal vesicles and the prostate gland, provide fluids that lubricate the duct system and nourish the sperm cells. In particular, the prostate gland surrounds the ejaculatory ducts at the base of the male urethra, just below the bladder. The prostate gland is responsible for the proof semen, a liquid mixture of sperm cells, prostate fluid, and seminal fluid. This gland is also responsible for making the semen milky in appearance by mixing calcium to the semen coming from seminal vesicle. The semen remains cloudy and clumpy until the prostatic pro fibrinolysis is formed into fibrinolysis and lysis of the fibrinogen from the seminal vesicle fluids occurs.
2.2 Erectile Function and Dysfunction
Erection is a vascular event which relies upon interaction of neural and humoral mechanisms at various levels. Indeed, since the penis receives innervations from sacral parasympathetic (pelvic), thoracolumbar sympathetic, and somatic (pudenda) nerves, the erection phenomenon requires participation of these three systems. Thus, erection is a consequence of a complex integration of several signals. In simple words, erection is essentially a spinal reflex that can be initiated by recruitment of penile afferents, but also by visual, olfactory, and imaginary stimuli. All these stimuli may contribute to the increase in intra-cavernous pressure (Cirino et al. 2006) and thus to penile erection. Erectile dysfunction (ED) is defined as the consistent or recurrent inability of a man to attain and/or maintain a penile erection sufficient for sexual activity (2nd International Consultation on Sexual Dysfunction-Paris, June 28th–July 1st, 2003). Of note, ED shares many of the risk factors that contribute to the development and the progression of cardiovascular diseases such as age, hypercholesterolemia, obesity, diabetes, and smoking (Brunner et al. 2005). Several clinical studies have shown that ED is often associated with cardiovascular disturbance and therefore it could be taken in account as an early sign of cardiovascular diseases (Dong et al. 2011; Nehra et al. 2012). This hypothesis relies on the fact that penile artery size is smaller as compared to coronary arteries. Therefore, it is feasible that the same level of endothelial dysfunction can cause a more significant reduction of blood flow in erectile tissues compared to that elicited in coronary circulation. Therefore, ED could be considered a prognostic factor for possible cardiovascular problems (Gandaglia et al. 2014).
2.3 Hydrogen Sulfide and Erectile Function and Dysfunction
The corpora cavernosa, as briefly reported above, are composed of sinusoids bearing a single layer of endothelial cells surrounded by multiple layers of smooth muscle cells. Thus, the corpora cavernosa are vascular organs. Among the vasodilator agents, it is undisputed that nitric oxide (NO) is considered one of the most important endogenous factors since it is released not only by endothelial cells but also from plexus nerve. Recently hydrogen sulfide (H2S), as discussed in the other chapters, has been shown to possess an important role in the modulation of smooth muscle cell tone. In particular, it has been demonstrated that H2S relaxes smooth muscle cells, and this finding together with others have lead to study the role of this mediator in the penile physiology. To date, the role of this gas in corpus cavernosum function has been addressed in animal models as well as in the human tissues (d’Emmanuele di Villa Bianca et al. 2011; Qiu et al. 2012). The first evidence was published in 2006 by Srilatha and coworkers. These authors showed that the intra-cavernous injection of sodium hydrogen sulfide (NaHS) resulted in a significant increase in penile length and cavernous pressure in primates measured by using laser Doppler flow meter and cutaneous probe. To acquire further evidence on the role of H2S in penile erection, a study on rats was also performed. Administration of dl-propargylglycine [PAG, a cysthatione-γ lyase (CSE) inhibitor] to rats resulted in a significant reduction in cavernous nerve stimulation-evoked perfusion pressure. This study suggested a possible role for endogenous H2S in facilitating nerve-mediated penile tumescence (Srilatha et al. 2006). In 2009, d’Emmanuele di Villa Bianca and coworkers demonstrated the role and function of H2S in human. The authors showed that both cysthatione-β synthase (CBS) and CSE, the main enzymes involved in H2S synthesis, are expressed in the human corpus cavernosum (CC) (Fig. 1a, b). Moreover, by using human CC homogenates, as enzyme source, they confirmed that the enzymes present in human CC can efficiently convert l-Cys (the substrate) to H2S (Fig. 1c). CBS and CSE are localized within muscular trabeculae and smooth muscle component of the penile artery (Fig. 2). Interestingly, the enzymes appear to be differently distributed within the penile structure. In particular, CSE but not CBS is expressed in peripheral nerves (Fig. 2). Functional studies conducted by using human CC isolated strips demonstrated that both exogenous H2S and l-Cys cause a concentration-dependent relaxation of human CC strips. This relaxant effect was inhibited by the CBS inhibitor, aminoxyacetic acid (AOAA) and only slightly reduced by the endothelial NO-synthase inhibitor, l-NAME. Electrophysiological experiments performed using peripheral nerve electrical field stimulation of human penile tissue demonstrated that H2S pathway is involved in penile homeostasis. Indeed, EFS of human CC strips, under resting conditions, caused an increase in tension that was significantly potentiated by inhibiting CSE and/or CBS with the selective inhibitors PAG and/or AOAA (Fig. 3). The role of H2S pathway in erectile function was confirmed also in vivo by using an experimental animal model of penile erection. Using this model, it was demonstrated that either NaHS or l-Cys administration cause an increase in the intra-cavernous pressure in anesthetized rats (d’Emmanuele di Villa Bianca et al. 2009).

CBS and CSE: activity, Western blot analysis, and qRT-PCR of human penile tissue. (a) HCC-expressed mRNA for both CBS and CSE as determined by qRT-PCR. (b) Representative Western blot analysis for CBS and CSE. (c) HCC homogenate produced H2S under basal conditions (open bar). Incubation of HCC homogenate with 10 mM l-Cys caused a significant increase in the H2S production compared with basal values (**P < 0.001). PAG (10 mM), 1 mM AOAA, or 10 mM PAG plus 1 mM AOAA significantly inhibited the l-Cys-induced increase in H2S production († P < 0.01). Data represent the mean SEM from 3 or 4 different human specimens (Reprinted with permission from d’Emmanuele di Villa Bianca et al. 2009)

Immunochemistry for CBS and CSE in HCC. Immunohistochemical detection of CSE and CBS in HCC tissue. Immunoreactivity and nuclear staining appear brown (DAB) and blue (hematoxylin counterstain), respectively. CSE was detected in trabecular muscular tissue (a and b, black arrows) and vascular smooth muscle cells (c, white arrows). Immunoreactivity for CBS was mostly observed in trabecular muscular tissue (d, black arrows). Results illustrated are from a single experiment and are representative of three different specimens. (Original magnification, 200.) Immunohistochemical detection of CSE and CBS in HCC nerve fibers. Immunoreactivity and nuclear staining appear brown (DAB) and blue (hematoxylin counterstain), respectively. CSE was detected in nerve fibers in cryostat (e, arrows) and not in paraffin (g) sections. Both cryostat (f) and paraffin (h) sections lacked immunoreactivity for CBS. Results illustrated are from a single experiment and are representative of three different specimens. (Original magnification, 200.) (Reprinted with permission from d’Emmanuele di Villa Bianca et al. 2009)

EFS of HCC strips. EFS caused a frequency-related increase in basal tone. Incubation of HCC strips with 10 mM PAG for 30 or 60 min (a) or with 1 mM AOAA for 30 or 60 min (b) significantly increased the EFS-induced contraction [*P < 0.05; **P < 0.01 vs. control (CTR)]. Tissue responses to EFS are expressed as force in dynes per milligram of tissue. Experiments were performed on three different specimens (Reprinted with permission from d’Emmanuele di Villa Bianca et al. 2009)
These studies have revealed that there are important differences between the human and rat tissue for what concerns the H2S pathway. In fact, in rats, only CSE is expressed. This finding is also confirmed by the fact that CSE but not CBS inhibition significantly decreased H2S production when rat cavernosal tissue homogenates were used as enzyme source to generate H2S from l-cysteine in vitro. Concerning the role of CSE and H2S in rat corpus cavernosum, it has been also shown that CSE inhibition by PAG causes a significant increase in non-adrenergic non-cholinergic-induced relaxation leading to an enhanced neurogenic relaxation of rat corpus cavernosum induced by EFS. The authors hypothesize that, since H2S does not have a direct constrictor effect in corporal tissue, this effect might be due to either inhibition of NO synthesis by endogenous H2S or to a direct chemical reaction between H2S and NOS products (Ghasemi et al. 2012). Thus, while CSE in rats accounts for the production of H2S, in human, the H2S pathway is sustained by both CBS and CSE, which are also differently distributed in the human corpus cavernosum (Fig. 2) indicating that human and rat penile tissues are different for what concerns the H2S pathway involvement in penile erectile mechanism(s).
2.4 Mechanism(s) of Action that Sustain the Effect of H2S in Erection
2.4.1 Ion Channels and EDHF as H2S Target
Ion channels are intimately involved in the biochemical events associated with smooth muscle function, and their activation/inactivation is tightly associated with tumescence/detumescence function during erection. As already reported in the previous chapters, H2S activates within the vasculature the adenosine triphosphate (ATP)-activated potassium (KATP) channel. The mechanism of activation proposed involves S-sulfhydration by H2S of the cysteine residues located on a specific subunit of the extracellular loop of the KATP channel complex leading to opening of this channel (Jiang et al. 2010; Mustafa et al. 2011). In particular, potassium channels are important in mediating physiologically relevant relaxation responses in human, rabbit, and rat corpus cavernosum strips (Christ et al. 1993; Mirone et al. 2000; Karicheti and Christ 2001; Spektor et al. 2002; Ruiz Rubio et al. 2004). In this context, it has been proposed that KATP channels could have an important role as modulator of corporal smooth muscle tone (Christ et al. 1993). Indeed, in diabetes, the KATP channels-mediated relaxation is significantly reduced in human CC strips (Venkateswarlu et al. 2002). Interestingly, in human CC strips glybenclamide, a selective KATP channel inhibitor, significantly impairs H2S-induced relaxation (d’Emmanuele di Villa Bianca et al. 2009) indicating that these channels are involved in H2S effect. KATP channels may also act by increasing cyclic adenosine monophosphate (cAMP) and thus cause relaxation. In fact, KATP channels are a physiologically important target of the adenylate cyclase/cAMP/PKA signaling pathway (Nelson et al. 2011), confirming indirectly the possible involvement of this channel as proposed by Srilatha et al. in 2007. In fact, the effect of NaHS in rabbit CC can be blocked by using MDL 12,330A, an adenylate cyclase inhibitor (Srilatha et al. 2007). On this basis, it is feasible to hypothesize that in diabetes, where there is a reduced activity of these channels, the H2S pathways may be involved in ED associated with the diabetes. However, these issues need to be addressed more carefully in order to define the contribution of this mechanism to the physiology of erection as well as its involvement in ED.
Due to the ability of H2S to directly interact with proteins, another channel that seems to be its target is the calcium-dependent potassium (KCa) channel. The physiological role of this channel in human CCSM cells have been demonstrated (Christ et al. 1993). As mentioned before, NO is a key mediator in erectile function, and it derives from both peripheral nervous terminations and of course from endothelial cells. However, the existence of an unidentified endothelial factor that promotes smooth muscle hyperpolarization and relaxation, resistant to NO synthase (NOS) and cyclooxygenase (COX) inhibition, has been clearly established (Busse et al. 2002). This factor, named endothelial-derived hyperpolarizing factor (EDHF), has particular functional relevance in small arteries. This pathway involves the activation of two populations of endothelial potassium channels, the small/intermediate conductance and the large KCa channels (Grgic et al. 2009). It has been clearly demonstrated that in human penile resistance arteries, as opposite to human CC, there is a significant component in the relaxation response to acetylcholine that is resistant to NOS and COX inhibition attributed to EDHF-like activity (Angulo et al. 2003). This EDHF-like activity is impaired in human penile resistance arteries from diabetic patients. In this framework fits H2S since there are significant data supporting H2S as an EDHF (Wang 2003, 2009; d’Emmanuele di Villa Bianca et al. 2011; Tang et al. 2013).
Inhibitors of the phosphodiesterase-5 (PDE-5) are drugs largely used to treat men with ED to increase the half-life of cyclic guanosine monophosphate (cGMP) in the corpus cavernosum facilitating its relaxation and therefore penile erection. However, although sildenafil has been shown to be efficacious for treating ED, the percentage of efficacy in diabetic patients is clearly reduced when compared to nondiabetic men (Rendell et al. 1999; Price et al. 1998; Vickers and Satyanarayana 2002). Indeed, the presence of ED and diabetes in a patient represents a prognostic factor for a poor response to sildenafil but also with the other new PDE-5 inhibitors tadalafil (Saenz de Tejada et al. 2002) and vardenafil (Goldstein et al. 2003). The mechanism(s) responsible for this poor clinical outcome is not, as yet, understood. One hypothesis formulated is related to the impairment of EDHF in human penile resistance vessels of diabetes patients that is not restored by PDE-5 treatment. This hypothesis is supported by a study where it has been shown that dobesilate, a drug that specifically enhances endothelium-dependent relaxation attributed to EDHF, restores the endothelial function in penile arteries of diabetic patients (Angulo et al. 2003). Therefore, it is also feasible, and it needs to be explored the possibility, that the combination of an H2S donor and a PDE-5 inhibitor may overcome the poor outcome of PDE-5 inhibitors in diabetic patients and could be beneficial in the oral treatment of diabetic ED.
Other H2S target may be the calcium permeable channels. In fact, other authors have reported the ability of H2S to interfere not only with potassium channels but also with calcium homeostasis particularly in neurons, cardiomyocytes, and endothelial cells. Although some effects of H2S on calcium signals are secondary to KATP modulation, there is growing consensus about the existence of a direct effect of H2S on Ca2+-permeable channels (Munaron et al. 2013). The modulation/activation of these channels seems to be tightly related to the H2S concentration. At the present stage, there are no data available in the current literature on CCSM cells in both animal models and human tissues.
2.4.2 Testosterone, Gender Difference, and Biosynthesis of H2S
Testosterone (T), as already reported above, plays a critical role not only in human male sexual behavior, but several studies have indicated a relation between androgen levels and sexual interest, libido, and the frequencies of orgasm and nocturnal erections. Moreover, there is a growing body of evidence that low T levels are associated with an increased cardiovascular and cancer mortality (Hackett et al. 2014). Thus, T seems to have a protective effect in man as occurs for estrogen in woman. For example, T plasma levels negatively correlate with hypertension, diabetes, and severe coronary artery disease. While it is clear that T restoration to a physiological level is beneficial in hypogonadal subjects, the positive role of T treatment is questionable in men who are not clearly hypogonadal or eugonadal. Another important well-accepted concept is that T level correlates to ED (Isidori et al. 2014). In particular, patients with organic ED have lower free T levels than patients with psychogenic ED. Moreover, a strong positive correlation exists between free T levels and the degree of trabecular smooth muscle relaxation, as measured by resistive index at dynamic duplex ultrasound (Aversa et al. 2000). The role of T in regulating CCSM and penile arterial tone has been extensively explored in animal models. In this regards, a dual separate effect with or without the endothelium has been shown. The effect in presence of endothelium is related to NO; in particular, there are several evidences supporting a role for androgens in regulating the expression and activity of NOS isoforms in the corpus cavernosum in animal models (Traish et al. 2007). The increase in T plasma levels in response to sexual stimulation raised the question of its biological significance. To this aim, several indirect evidences support the presence of a local vascular effect of T in men. In fact, it has been demonstrated that penile erection in the healthy male is associated with a significant increase in T level in both circulating and in the cavernous plasma. Moreover, in the flaccid state, the T level in cavernous plasma is significantly lower when compared to the systemic concentration, and on this basis, the authors conclude that these differential evaluation could represent a diagnostic tool (Becker et al. 2000). Interestingly, during the penile tumescence and rigidity, T level in corpus cavernosum increases significantly in comparison to the flaccid state, and this increase is not observed in ED patients (Becker et al. 2001). These results further support the hypothesis that T, through the androgen receptor, has a direct effect on the cavernous smooth muscle. Androgen receptors are present in the human CC (Schultheiss et al. 2003), and T induces relaxation by activating smooth muscle KATP channels in human CC strips (Yildiz et al. 2009). This study was the first to report a direct non-genomic relaxant effect of T on human CC in vitro. The authors have demonstrated that T causes a rapid vasorelaxation partially mediated by an increase of potassium efflux through KATP channels, but not involving BKCa, voltage-dependent inward rectifier K channel, or voltage K channels. Thus, T-induced relaxation on smooth muscle CC seems to involve an endothelium-dependent (particularly trough NO pathway) and -independent (probably by K channels) mechanism (Aversa et al. 2000). It has been confirmed that these channels are also involved in T-induced relaxation within the vasculature and that T effect involves H2S pathway. In fact, it has been shown that T causes an increase in H2S level involving the l-Cys/H2S pathway in rat aorta (Bucci et al. 2009). The increase of H2S induced by T and its vasodilatory effect was prevented by the androgen receptor antagonist nilutamide in rat aorta rings indicating that the interaction with the androgen receptor is a key issue (Bucci et al. 2009). In this context, it has been recently shown the heat shock protein 90, which plays a role in the activation of androgen receptor, is also involved in CSE activation. In the same work, the presence of a similar mechanism for progesterone and 17-β-estradiol was ruled out. This latter observation well fit with the finding that H2S levels in human blood, collected from male healthy volunteers, were higher than those in female samples (Brancaleone et al. 2014).
Thus, if T level is related to ED, it could well be that an impairment of T-mediated H2S production plays a role in ED. On this specific subject, there is only one study available, performed using an in vivo rat model. In this work, the role of endogenous H2S in ED induced by androgen deficiency has been investigated. Serum T levels were significantly reduced after 2 and 4 weeks from castration, and in this condition, the H2S pathway was significantly impaired as well as there was a reduction in intra-cavernous pressure increase elicited by electrical stimulation. On the other hand, T replacement resulted efficacious to restore H2S release and to improve intra-cavernous pressure in rats. These in vivo data support the hypothesis that H2S pathway may be one of the mechanisms underlying androgen role in erection (Zuo et al. 2014). Moreover, it has been demonstrated that aging significantly impairs NO and H2S level both in plasma and corpus cavernosum tissue. A reduction of the intra-cavernous pressure is countered by NaHS or sildenafil after 10 weeks of treatment. The link between T and H2S was further confirmed by Srilatha and coauthors, who have shown a marked increase in T or estradiol levels after NaHS supplementation. These data support the idea that ED in aging may be also linked to a derangement in the H2S pathway accompanied by low T levels (Srilatha et al. 2012).
All these findings support the importance of androgens in regulating smooth muscle function in the penis. A possible clinical application could be the use of a combination of PDE-5, T, and/or H2S donors/substrate (such as l-Cysteine) to be used in patients nonresponder to PDE-5 therapy (Isidori et al. 2014).
2.4.3 Interaction Between H2S and NO
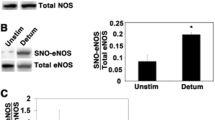
There is a deep discussion in literature concerning the role of NO/cGMP pathway in H2S effects, and this issue is under investigation in several anatomical district (see other chapters). For example, it has been demonstrated that CSE activity is upregulated by NO (Zhao et al. 2001), and it is partially inhibited by NOS blockade (Zhao and Wang 2002). The data available in the current literature demonstrated that H2S (1) inhibits eNOS activity partly through inhibition of eNOS phosphorylation by reducing Akt phosphorylation (Geng et al. 2007), (2) stimulates the activity of several upstream kinase such as Akt and in turn activates eNOS by phosphorylation (Cai et al. 2007), (3) acts as PDE inhibitor (Bucci et al. 2010), (4) directly causes eNOS phosphorylation (Altaany et al. 2014), (5) is mutually required with NO in order to elicit angiogenesis and vasodilatation (Coletta et al. 2012). Therefore, there are several evidence implying a cross talk at vascular level between NO and H2S. As opposite, few studies are available on human CC tissues. The relaxing effect of H2S on human CC strips has been demonstrated to be endothelium independent (d’Emmanuele di Villa Bianca et al. 2009), but this result does not exclude a possible indirect effect of H2S since, in presence of the endothelium blockade of NOS, l-NAME causes a significant inhibition of H2S-induced relaxation at higher concentration. This latter result is in line with a recent study by Meng and coworkers (Meng et al. 2013) demonstrating that incubation of rat CC tissues with NaHS leads to an increase in eNOS but not nNOS mRNA. In this study, the authors show that the increased mRNA expression of eNOS correlates with protein expression as well as with NO production. However, caveolin-1 expression, a dominant inhibitory interaction partner of eNOS, was not modified by H2S. On this basis, the authors conclude that H2S could be particularly useful in improving the clinical outcome of ED patients, whose erectile impairment involves a weakened function of endothelial-derived NO. However, whether H2S increases the soluble guanylil cyclase activity directly was not examined. The possible role of H2S on modulating the NO/cGMP pathway has been, instead, suggested in vascular studies, where it has been shown that H2S increases cGMP levels acting as a PDE inhibitor delaying cGMP degradation (Bucci et al. 2010; Coletta et al. 2012). Preliminary data indicate that an increase in cGMP level drive by PDE-5 can cause an increase in H2S production in mice corpus cavernosum (Dikmen et al. 2013). Many aspects concerning the interaction between these two gasses are still matter of debate in the current relevant literature. A recent review addresses in depth the possible role of this cross talk in erectile function/dysfunction (Yetik-Anacak et al. 2014).
2.5 Hydrogen Sulfide and Vas Deferens
The vas deferens transports sperm from the epididymis to the ejaculatory ducts in anticipation of ejaculation. Due to the role of H2S on erectile function, its presence and role in vas deferens have also been evaluated. Both CBS and CSE are functionally expressed in the vas deferens of rat, mice, and human. The endogenous H2S causes a smooth muscle relaxation of vas deferens (Li et al. 2011). The same group has also demonstrated that NaHS-induced effect is mediated by BKCa channel. Indeed, they observed a consistent reversion of the relaxant effect of rat vas deferens-induced relaxation by performing a pharmacological modulation with iberiotoxin or tetraethylammonium. The authors also demonstrated that H2S modulation of KCa channels requires a redox signaling. Indeed, N-ethylmaleimide, a sulfhydryl alkylation compound protecting thiols from oxidation, inhibited NaHS relaxation as opposite to DTT, a strong reducing agent, that did not affect the H2S response of vas deferens. Besides, the presence of the BKCa channels in rat vas deferens smooth muscle cells was also confirmed. Finally, the involvement of the NO pathway, the transient receptor potential (TRP) channels and of the KATP channels were excluded (Li et al. 2012).
2.6 Hydrogen Sulfide and Human Prostate
Regarding the role of H2S in the prostate, it has been reported by Guo and coworkers (Guo et al. 2012) that in human prostatic tissues (obtained from patients undergoing surgery for prostatic cancer) and cells, this pathway is physiologically present. The presence of H2S pathway has been also confirmed by using biopsy from cancer-free human prostate (Gai et al. 2013). Moreover, Guo and coworkers have demonstrated that both cell activity and CBS/CSE protein levels are higher in the androgen-dependent prostate cancer cell LNCaP than in all the other cell lines evaluated and that dihydrotestosterone downregulates this activity. Varying H2S levels, as well as CBS/CSE expression, in human prostate stromal and epithelial compartments have been also described. In particular, prostate epithelium expresses both CBS and CSE as opposite to stromal where only CSE is expressed. Overall, these data suggest that H2S pathway can be involved in prostate cancer and benign prostatic hyperplasia. These results also further confirm that a link between H2S and T exists. In physiological condition, T can modulate the H2S synthesis contributing to erectile function. In pathologic condition, such as androgen-dependent prostate cancer, where H2S pathway is over expressed, it may represent a potential therapeutic target (Guo et al. 2012). Indeed, H2S does not alter significantly androgen receptor expression or its phosphorylation but inhibits androgen receptor transactivation probably at the DNA-binding level. This effect may involve a posttranslational regulation of androgen receptor by S-sulfhydration leading to conformational change and alteration of protein function. Thus, H2S may decrease the genomic effect associated with androgen receptor activation exhibiting an anti-proliferating effect, data that are in agreement with the higher cell proliferating rate associated with aging in prostate tissues of CSE knock-out mice (Zhao et al. 2014).
3 Hydrogen Sulfide in Female Sexual Function
There is very little published on the possible involvement of the H2S pathway in female sexual physiology. In 2009, a pilot study has been published suggesting that H2S pathway plays a physiological role in female sexual apparatus. In particular, the authors have studied the effect of exogenous H2S in vaginal and clitoral cavernosal smooth muscle strips from New Zealand white female rabbits. By using H2S donors and several inhibitors towards different enzymes and channels, the authors have shown that H2S vasodilatory effect involves cAMP, (cyclic adenosine 3′5′-monophosphate), NO-cGMP (cyclic guanosine monophosphate), and KATP channels. Of particular interest is the finding that inhibition of H2S-induced relaxation was observed only when a combination of both adenylate- and guanylate- cyclase inhibitors was used indicating that both nucleotides may concur to H2S effect. Thus, H2S pathway appears to be involved also in female sexual responses. Moreover, these data further support the evidence that there exists an interplay between NO and H2S pathways (Srilatha et al. 2009).
4 Hydrogen Sulfide and Bladder
4.1 Bladder Function
The urinary bladder has two important functions: storage of urine and emptying. Storage of urine occurs at low pressure, which implies that the bladder relaxes during the filling phase. The wall of the bladder is composed of 3 layers:
-
1.
Outer layer of loose connective tissue, containing blood and lymphatic vessels and nerves.
-
2.
Middle layer, consisting of a mass of interlacing smooth muscle fibers and elastic tissue—this is called the detrusor muscle.
-
3.
Inner layer composed of transitional epithelium.
The storage and periodic elimination of urine are dependent upon the reciprocal activity of two functional units in the urinary tract: a reservoir (urinary bladder) and an outlet (bladder neck, smooth and striated muscle of the urethra). During urine storage, the outlet is closed, and the bladder is quiescent, allowing intravesical pressure to remain low over a wide range of bladder volumes. During voiding, the muscles of outlet relax and the bladder smooth muscles contract, raising intravesical pressure and inducing urine flow. These changes are coordinated by three sets of nerves (parasympathetic, sympathetic, and somatic) emerging from the sacral and thoracolumbar levels of spinal cord. Thus, when the bladder is filled, the relaxation of the wall stimulates the afferent fibers, and input is transmitted to the cortex eliciting the micturition reflex. During the micturition, the detrusor muscle and the longitudinal muscle of the neck and the urethral sphincter contract by activation of parasympathetic nerves. Contextually, an inhibitory input of the somatic nerves causes a relaxation of the external sphincter allowing the urine ejection. Different signaling molecules contribute to the fine tuning of the urinary bladder, acting both in autocrine and paracrine manner. The receptors involved are specifically distributed among different types of cells within the urinary bladder structure. Various neurotransmitters, including acetylcholine, norepinephrine, dopamine, serotonin, excitatory and inhibitory amino acids, adenosine thriphosphate, nitric oxide, and neuropeptides, are implicated in the neuronal regulation of micturition (de Groat and Yoshimura 2001). Several disorders can affect the bladder such as cystitis, urinary stones, bladder cancer, urinary incontinence, hematuria, urinary retention, cystocele, bed-wetting, dysuria, overactive bladder, and lower urinary tract symptoms (LUTS). Disturbances of storage function may result in LUTS, such as urgency, frequency, and urge incontinence, the components of the overactive bladder syndrome (Abrams et al. 2002). The overactive bladder syndrome, which may be due to involuntary contractions of the smooth muscle of the bladder (detrusor) during the storage phase, is a common problem (Milsom et al. 2001). H2S has been shown to be involved in the bladder function in different species such as trout, rat, pig, and human (Fusco et al. 2012; Gai et al. 2013; Fernandes et al. 2013a, b; Patacchini et al. 2004; Dombkowski et al. 2006).
4.2 Animal Studies
H2S is endogenously produced in urinary bladder of trout, mice, and rat implying that the contribution of this pathway to bladder function is conserved in different species. Similarly, to human tissues, homogenates of bladder can generate detectable amount of H2S in basal or stimulated conditions (Dombkowski et al. 2006; Matsunami et al. 2012; Gai et al. 2013). The expression of the enzymes responsible of H2S production has been reported in bladder of rat, pig, and mice by using different methods. CSE and CBS were detected by immunohistochemistry in nerve fibers and are widely distributed within the smooth muscle layer of the pig bladder neck (Fernandes et al. 2013a). CBS, CSE, and 3MST have been also shown to be present in rat bladder (Gai et al. 2013). In mouse bladder, only the presence of CSE has been reported so far, (Matsunami et al. 2012). However, the presence of CBS cannot be excluded since it has not been evaluated (Matsunami et al. 2012).
In the urinary tract, H2S donors caused smooth muscle relaxation and contraction depending on the species and experimental conditions used. NaHS (exogenous source of H2S) or l-cysteine (the substrate) relaxes rat bladder strips in a concentration-dependent manner (Gai et al. 2013). This effect can be inhibited by incubating tissues with glybenclamide, a KATP channel inhibitor, CSE, and CBS blockers (Gai et al. 2013). In trout bladder, NaHS, as well as Na2S (exogenous donor of H2S less used in the current literature), inhibits spontaneous contractions and relaxes precontracted strips (Dombkowski et al. 2006). In trout bladder, the H2S response does not appear to be mediated by KATP channels as occurs in human and rat since glybenclamide was ineffective. Similarly, the other types of K+ channels were not involved in H2S-induced relaxation, as well. The rat urinary bladder can be indirectly contracted through H2S stimulation of capsaicine-sensitive nerves as reported by Patacchini and coworkers (Patacchini et al. 2004, 2005). This discrepancy between rat and human could be due to the different derivation of the tissue (Kardong 2005). In rat, H2S has a contractile effect that is mediated by the activation of sensory neurons in a ruthenium red-sensitive but not capsazepine-dependent manner. Thus, it is feasible that in this case, the molecular target of H2S could be either a receptorial domain located on the transient receptor potential vanilloid receptor 1 (TRPV1) cation channel, independent from those bound by vanilloids and other TRPV1 activators, or another ruthenium red-sensitive TRP cation channels co-expressed on primary afferent neuron terminals (Patacchini et al. 2005). The super family of TRP includes the transient receptor potential ankyrin (TRPA) (Nilius et al. 2007), which has been found on capsaicine-sensitive primary sensory neurons (Story et al. 2003; Bautista et al. 2005). The TRPA1 activators allyl isothiocyanate and cinnamaldehyde contract rat urinary bladder through stimulation of the capsaicin-sensitive nerves (Andrade et al. 2006; Patacchini et al. 1990). Therefore, an action of H2S also on TRPA1 in urinary bladder has been suggested. This hypothesis is based upon the following findings i) co-localization of TRPA1 and TRPV1 in rat bladder afferents ii) H2S mimics the TRPA1 agonist cinnamaldehyde effect stimulating the micturition reflex after protamine sulfate pretreatment (Streng et al. 2008). Thus, TRPA1 represents a molecular target for H2S in rat bladder.
H2S-mediated O2 sensing has been demonstrated in a variety of O2-sensing tissues in vertebrate cardiovascular and respiratory systems, including smooth muscle in systemic and respiratory blood vessels and airways, carotid artery, adrenal medulla, and other peripheral as well as central chemoreceptors (Olson 2014). Concerning the possible involvement of this mechanism in urinary tract, it has been shown that H2S is involved in O2 sensing/signal transduction in the trout urinary bladder (Dombkowski et al. 2006) showing some similitude with mammalian tissue (Olson 2014).
An upregulation of CSE has been found in cyclophosphamide-induced cystitis in mouse, suggesting a role for the endogenous H2S in the pathogenesis of this experimental model (Matsunami et al. 2012). In addition, a link between H2S and Cav3.2 T-type channel has also been suggested in the cystitis-related nociceptive changes (Matsunami et al. 2012).
The control of the bladder neck tone is important during the voiding phase either in physiological or in pathological conditions. In this regard, CBS and CSE, as reported above, have been found expressed in nerve fibers in the pig bladder neck smooth muscle layer suggesting the involvement of H2S as mediator released following muscle distension (Fernandes et al. 2013a). Both electrical field or GYY4137, an H2S donor, relaxed the pig bladder neck strips through KATP channel activation as well as COX1-derived prostanoids (Fernandes et al. 2013a, b) indicating also a functional role at this level. These studies provided also evidence for a role of endogenous H2S released from nerves involvement in the inhibitory transmission of the outflow region (Fernandes et al. 2013a). Additionally, H2S promotes the release of inhibitory neuropeptide by the activation of TRPA1 and TRPV1 (Fernandes et al. 2013b). Therefore, at the present stage, KATP channels, TRPA1, and TRPV1 are considered feasible targets for H2S in the bladder function.
4.3 Human Studies
Human bladder expresses all three enzymes responsible for the H2S production namely CBS, CSE, and 3-MST (Fusco et al. 2012; Gai et al. 2013). Indeed, human bladder homogenate generates a basal level of production of H2S that can be enhanced by incubation with l-cysteine (the substrate). This effect is reversed by either PAG (CSE inhibitor) or AOAA (CBS inhibitor) (Fig. 4). Functional studies performed using isolated human bladder strips have demonstrated that both sodium hydrogen sulfide (NaHS) or l-cysteine relax human bladder strips precontracted by carbachol. Thus, H2S pathway is involved in the regulation of bladder homeostasis.

Cystathionine b-synthase (CBS) and cystathionine g-lyase (CSE) western blot, and activity in human bladder dome. (a) Representative western blot for CBS and CSE (upper) and densitometric analysis for CBS and CSE normalized to glyceraldehyde-3-phosphate dehydrogenase (GAPDH; lower) from nine patients; data were expressed as the mean plus or minus standard error of the mean (SE). (b) Production of hydrogen sulfide (H2S) in human tissue homogenates from five patients under basal condition (open bar) and after incubation with l-cysteine 10 mM (black bars). l-cysteine caused a significant increase in H2S production (***p < 0.001 vs. basal) that was significantly reverted by CSE inhibitor (d,l-propargylglycine [PPG]; 10 mM) and/or CBS inhibitor (aminooxyacetic acid [AOAA]; 1 mM) (°°p < 0.01 and °°°p < 0.001, respectively, vs. l-cysteine). Data were calculated as nanomoles per milligram of protein per minute and expressed as mean ± SE (Reprinted with permission from Fusco et al. 2012)
As discussed, KATP channels are activated by H2S. It is known that urinary bladder smooth muscles express KATP channels, and they have been shown to be involved in the regulation of bladder contractility (Andersson 1992; Bonev and Nelson 1993). The presence of mRNA for sulfonylurea receptors has been demonstrated in both pig and human detrusor (Buckner et al. 2000). The detrusor smooth muscle expresses a variety of K+ channels that are responsible for the detrusor smooth muscle excitability and contractility (Andersson and Arner 2004; Brading 2006; Christ and Hodges 2006; Gopalakrishnan and Shieh 2004). Interestingly, the density of KATP channels is significantly higher in detrusor smooth muscle (Petkov et al. 2001; Kajioka et al. 2008; Shieh et al. 2001). In this context, it has been shown that ~1 % of the functional KATP channels need to be activated in order to inhibit detrusor smooth muscle action potentials and related phasic contractions (Petkov et al. 2001). Studies on isolated detrusor muscle from human and several animal species have demonstrated that KATP channel openers reduce not only spontaneous contractions but also those induced by electrical stimulation, carbachol, and low external K+ concentrations (Andersson 1993). Interestingly, the relaxant effect elicited by H2S is inhibited by glybenclamide, a KATP channel inhibitor (Gai et al. 2013). Therefore, it is feasible to hypothesize that once released, H2S causes relaxation of the detrusor muscle that involves KATP channel activation thereby contributing to the tonic regulation of the bladder tone. Bladder tone regulation is a key issue in LUTS. LUTS markedly increase with age in both males and females, and it represents a major problem in the elderly population. Recently, the PDE-5 inhibitor tadalafil has been approved by FDA for LUTS treatment. In addition, it has been reported that sildenafil, a well-known PDE-5 inhibitor, can directly relax human bladder strips and that this effect involves the H2S pathway (Fusco et al. 2012). Indeed, sildenafil incubation of human bladder samples causes a time- and concentration-dependent increase in H2S production (Fig. 5). In conclusion, the human data, available in the current relevant literature, suggest that the H2S pathway is involved not only in the physiology of human bladder function but also in LUTS as well as in overactive bladder. However, further studies are necessary in order to better define the role of this pathway.

Sildenafil-induced hydrogen sulfide (H2S) production in human bladder dome. (a) Sildenafil 10 mM caused a time-dependent increase in H2S production (***p < 0.001 vs vehicle; **p < 0.01 vs. 30 min and 45 min). (b) Sildenafil incubation for 30 min caused a concentration-dependent increase in H2S production compared to vehicle (*p < 0.05 and **p < 0.001 vs. vehicle). (c) Combination of CBS and/or CSE inhibitors (dl-propargylglycine [PPG] 10 mM and/or aminooxyacetic acid [AOAA] 1 mM) significantly inhibited sildenafil-induced H2S production (°p < 0.05 and °°p < 0.01 vs sildenafil 10 mM; **p < 0.01 vs. vehicle). Data were calculated as nanomoles per milligram of protein per minute and expressed as mean ± SEM for five patients (Reprinted with permission from Fusco et al. 2012)
5 Hydrogen Sulfide and Urethra
The urethra together with smooth muscles in the bladder controls the storage and the voiding of urine. The presence as well as the ability of CBS, CSE, and 3-MST to convert l-cysteine into H2S has been reported both in human and rat urethra (Gai et al. 2013). On the functional side, H2S has been shown to have no effect on the basal tone but to relax the human urethra strips (Gratzke et al. 2009). The relaxing effect on human urethra strips has been proposed to involve the TRPA1 receptors that are expressed on nerves fibers, urothelium, and interstitial cells (Gratzke et al. 2009).
6 Conclusions and Future Directions
On the basis of the findings present in the current literature, from non-mammalian and mammalian, including human, it is clear that H2S is endogenously produced, and the enzymes responsible of its biosynthesis are constitutively present in the genitourinary tract. In addition, H2S appears to be a phylogenetically ancient and versatile regulatory molecule. Nevertheless, the H2S response in the genitourinary tract, as reported above, appears to be specie dependent. Therefore, more studies are needed in order to clearly define the role of this pathway in human since the translation of preclinical data to human is not always possible. For what concerns, drugs used in therapy that have been proposed to involve H2S as mediator, there are some considerations that can be made. It is now well established that patients’ response to a therapy is not equally effective, and who once were called nonresponders are now considered as a specific sub group. Indeed, it is now clear that genetics plays a great role in determining the nature of the drug responses. However, it is also true that, most likely, there are other therapeutic targets that have not as yet been defined within a specific pathology. In this regard, in both ED and in pathologies associated with bladder dysfunction, there is a need to find new therapies. Indeed, PDE-5 inhibitors often do not resolve the ED in diabetes patients or fail to act in a certain number of eligible patients with no comorbidities. In fact, the efficacy of PDE-5 inhibitor, which is a mainstay in the treatment of ED, is negatively associated with the nerve and endothelium damage. These latter features are associated with several pathologies such as cardiovascular disease, diabetes, obesity, and post-prostatectomy state and in turn lead to an impairment in the signaling of the NO/cGMP pathway. In this context, the H2S pathway represents an attractive target since a link between PDE-5 inhibitors and H2S pathway has been already shown. Indeed, tadalafil limits myocardial infarction through H2S signaling (Salloum et al. 2009), and sildenafil causes an increase in H2S production by CBS and CSE activities in human bladder (Fusco et al. 2012). A very preliminary attempt to develop a drug working on H2S and cGMP pathways has been taken by Shukla and coworkers who have synthesized and characterized an H2S-donating derivative of sildenafil (ACS6) (Shukla et al. 2009). For what concerns the bladder, it has to be stressed that the widely used antimuscarinic drugs often fails to alleviate the LUTS symptoms. Also in this case, by looking at the recent findings, highlighted in this chapter, the H2S pathway represents an attractive therapeutic target that may allow to develop new drugs. In Fig. 6 are shown the drugs for which the involvement of H2S in their mechanism of action has been proposed.

Drugs proposed to involve hydrogen sulfide (H2S) pathway in their mechanism of action. The cartoon reproduces a smooth muscle cell of the urogenital tract, where the involvement of H2S in testosterone and sildenafil mechanism of action is depicted. Adenosine triphosphate-activated potassium channel (KATP); Androgen receptor AR; calcium-dependent potassium channel (KCa); cyclic guanosine monophosphate (cGMP); Cysthatione-β synthase (CBS); Cysthatione-γ lyase (CSE), phosphodiesterase-5 (PDE-5)
Another possible therapeutic approach that rises from what has been discussed is the possibility to modulate H2S levels by exogenous supplementation. Intuitively, the easiest way should be to use H2S donors that release the gas slowly, in order to reproduce as much as possible, the physiological conditions. Alternatively, the H2S production could be enhanced by inducing H2S synthesis by using l-cysteine or other substrates. However, this approach could fail in condition where a downregulation of the enzyme CBS and CSE occurs. Several studies have addressed these issues in preclinical setting (Kashfi and Olson 2013). The activity of garlic-derived molecules, generally considered as H2S releasers following metabolization, such as diallyl disulfide, diallyl sulfide, diallyl trisulfide, and diallyl tetrasulfide, has been profusely investigated (Jacob et al. 2008). Garlic and its bioactive component, the S-allyl cysteine, have been shown to restore erectile function in diabetic rats by preventing ROS formation through modulation of NADPH oxidase subunit expression (Yang et al. 2013). In particular, anti-inflammatory and anticancer effects have been demonstrated. However, no data, concerning their efficacy, on urogenital tract are available. The GYY4137 is actually the most attracting synthetic H2S donor. It inhibits lipid accumulation exhibiting anti-atherosclerotic activity both in vitro and in vivo (Xu et al. 2014) and exerts anti-inflammatory (Li et al. 2013) and anticancer (Kashfi 2014) activity. Also in this case, there are no data available on GYY4137 effect in urogenital tract diseases.
References
Abe K, Kimura H (1996) The possible role of hydrogen sulfide as an endogenous neuromodulator. J Neurosci 16:1066–1071
Abrams P, Cardozo L, Fall M, Griffiths D, Rosier P, Ulmsten U, van Kerrebroeck P, Victor A, Wein A (2002) The standardization of terminology of lower urinary tract function: report from the Standardisation Sub-committee of the International Continence Society. Neurourol Urodyn 21:167–178
Altaany Z, Ju Y, Yang G, Wang R (2014) The coordination of S-sulfhydration, S-nitrosylation, and phosphorylation of endothelial nitric oxide synthase by hydrogen sulfide. Sci Signal 7:ra87
Andersson KE (1992) Clinical pharmacology of potassium channel openers. Pharmacol Toxicol 70:244–254
Andersson KE (1993) Pharmacology of lower urinary tract smooth muscles and penile erectile tissues. Pharmacol Rev 45:253–308
Andersson KE, Arner A (2004) Urinary bladder contraction and relaxation: physiology and pathophysiology. Physiol Rev 84:935–986
Andrade EL, Ferreira J, André E, Calixto JB (2006) Contractile mechanisms coupled to TRPA1 receptor activation in rat urinary bladder. Biochem Pharmacol 72:104–114
Angulo J, Cuevas P, Fernández A, Gabancho S, Allona A, Martín-Morales A, Moncada I, Videla S, Sáenz de Tejada I (2003) Diabetes impairs endothelium-dependent relaxation of human penile vascular tissues mediated by NO and EDHF. Biochem Biophys Res Commun 312:1202–1208
Aversa A, Isidori AM, De Martino MU, Caprio M, Fabbrini E, Rocchietti-March M, Frajese G, Fabbri A (2000) Androgens and penile erection: evidence for a direct relationship between free testosterone and cavernous vasodilation in men with erectile dysfunction. Clin Endocrinol (Oxf) 53:517–522
Bautista DM, Movahed P, Hinman A, Axelsson HE, Sterner O, Högestätt ED, Julius D, Jordt SE, Zygmunt PM (2005) Pungent products from garlic activate the sensory ion channel TRPA1. Proc Natl Acad Sci USA 102:12248–12252
Becker AJ, Uckert S, Stief CG, Truss MC, Machtens S, Scheller F, Knapp WH, Hartmann U, Jonas U (2000) Cavernous and systemic testosterone levels in different phases of human penile erection. Urology 56:125–129
Becker AJ, Uckert S, Stief CG, Scheller F, Knapp WH, Hartmann U, Jonas U (2001) Cavernous and systemic testosterone plasma levels during different penile conditions in healthy males and patients with erectile dysfunction. Urology 58:435–440
Bonev AD, Nelson MT (1993) ATP-sensitive potassium channels in smooth muscle cells from guinea pig urinary bladder. Am J Physiol Cell Physiol 264:C1190–C1200
Brading AF (2006) Spontaneous activity of lower urinary tract smooth muscles: correlation between ion channels and tissue function. J Physiol 570:13–22
Brancaleone V, Vellecco V, Matassa DS, d’Emmanuele di Villa Bianca R, Sorrentino R, Ianaro A, Bucci M, Esposito F, Cirino G (2014) Crucial role of androgen receptor in vascular H2S biosynthesis induced by testosterone. Br J Pharmacol 172(6):1505–1515
Brunner H, Cockcroft JR, Deanfield J, Donald A, Ferrannini E, Halcox J, Kiowski W, Lüscher TF, Mancia G, Natali A, Oliver JJ, Pessina AC, Rizzoni D, Rossi GP, Salvetti A, Spieker LE, Taddei S, Webb DJ, Working Group on Endothelins and Endothelial Factors of the European Society of Hypertension (2005) Endothelial function and dysfunction. Part II: association with cardiovascular risk factors and diseases. A statement by the working group on endothelins and endothelial factors of the european society of hypertension. J Hypertens 23:233–246
Bucci M, Mirone V, Di Lorenzo A, Vellecco V, Roviezzo F, Brancaleone V, Imbimbo C, Cirino G (2009) Hydrogen sulphide is involved in testosterone vascular effect. Eur Urol 56:378–383
Bucci M, Papapetropoulos A, Vellecco V, Zhou Z, Pyriochou A, Roussos C, Roviezzo F, Brancaleone V, Cirino G (2010) Hydrogen sulfide is an endogenous inhibitor of phosphodiesterase activity. Arterioscler Thromb Vasc Biol 30:1998–2004
Buckner SA, Milicic I, Daza A, Davis-Taber R, Scott VE, Sullivan JP, Brioni JD (2000) Pharmacological and molecular analysis of ATP-sensitive K channels in the pig and human detrusor. Eur J Pharmacol 400:287–295
Busse R, Edwards G, Félétou M, Fleming I, Vanhoutte PM, Weston AH (2002) EDHF: bringing the concepts together. Trends Pharmacol Sci 23:374–380
Cai WJ, Wang MJ, Moore PK, Jin HM, Yao T, Zhu YC (2007) The novel proangiogenic effect of hydrogen sulfide is dependent on Akt phosphorylation. Cardiovasc Res 76:29–40
Christ GJ, Hodges S (2006) Molecular mechanisms of detrusor and corporal myocyte contraction: identifying targets for pharmacotherapy of bladder and erectile dysfunction. Br J Pharmacol 147(Suppl 2):S41–S55
Christ GJ, Brink PR, Melman A, Spray DC (1993) The role of gap junctions and ion channels in the modulation of electrical and chemical signals in human corpus cavernosum smooth muscle. Int J Impot Res 5:77–96
Cirino G, Fusco F, Imbimbo C, Mirone V (2006) Pharmacology of erectile dysfunction in man. Pharmacol Ther 111:400–423
Coletta C, Papapetropoulos A, Erdelyi K, Olah G, Módis K, Panopoulos P, Asimakopoulou A, Gerö D, Sharina I, Martin E, Szabo C (2012) Hydrogen sulfide and nitric oxide are mutually dependent in the regulation of angiogenesis and endothelium-dependent vasorelaxation. Proc Natl Acad Sci USA 109:9161–9166
d’Emmanuele di Villa Bianca R, Sorrentino R, Maffia P, Mirone V, Imbimbo C, Fusco F, De Palma R, Ignarro LJ, Cirino G (2009) Hydrogen sulfide as a mediator of human corpus cavernosum smooth-muscle relaxation. Proc Natl Acad Sci USA 106:4513–4518
d’Emmanuele di Villa Bianca R, Sorrentino R, Coletta C, Mitidieri E, Rossi A, Vellecco V et al (2011) Hydrogen sulfide-induced dual vascular effect involves arachidonic acid cascade in rat mesenteric arterial bed. J Pharmacol Exp Ther 337:59–64
de Groat WC, Yoshimura N (2001) Pharmacology of the lower urinary tract. Annu Rev Pharmacol Toxicol 41:691–721
Dikmen A, d’Emmanuele di Villa Bianca R, Mitidieri E, Donnarumma E, Sevin G, Cirino G, Sorrentino R, Yetik-Anacak G (2013) New mechanism for the beneficial effect of sildenafil on erectile function: H2S. Nitric Oxide 31:S38
Dombkowski RA, Doellman MM, Head SK, Olson KR (2006) Hydrogen sulfide mediates hypoxia-induced relaxation of trout urinary bladder smooth muscle. J Exp Biol 209:3234–3340
Dong JY, Zhang YH, Qin LQ (2011) Erectile dysfunction and risk of cardiovascular disease: meta-analysis of prospective cohort studies. J Am Coll Cardiol 58:1378–1385
Fernandes VS, Ribeiro AS, Barahona MV, Orensanz LM, Martínez-Sáenz A, Recio P, Martínez AC, Bustamante S, Carballido J, García-Sacristán A, Prieto D, Hernández M (2013a) Hydrogen sulfide mediated inhibitory neurotransmission to the pig bladder neck: role of KATP channels, sensory nerves and calcium signaling. J Urol 190:746–756
Fernandes VS, Ribeiro AS, Martínez MP, Orensanz LM, Barahona MV, Martínez-Sáenz A, Recio P, Benedito S, Bustamante S, Carballido J, García-Sacristán A, Prieto D, Hernández M (2013b) Endogenous hydrogen sulfide has a powerful role in inhibitory neurotransmission to the pig bladder neck. J Urol 189:1567–1573
Fusco F, d’Emmanuele di Villa Bianca RD, Mitidieri E, Cirino G, Sorrentino R, Mirone V (2012) Sildenafil effect on the human bladder involves the L-cysteine/hydrogen sulfide pathway: a novel mechanism of action of phosphodiesterase type 5 inhibitors. Eur Urol 62:1174–1180
Gai JW, Wahafu W, Guo H, Liu M, Wang XC, Xiao YX, Zhang L, Xin ZC, Jin J (2013) Further evidence of endogenous hydrogen sulphide as a mediator of relaxation in human and rat bladder. Asian J Androl 15:692–696
Gandaglia G, Briganti A, Jackson G, Kloner RA, Montorsi F, Montorsi P, Vlachopoulos C (2014) A systematic review of the association between erectile dysfunction and cardiovascular disease. Eur Urol 65:968–978
Geng B, Cui Y, Zhao J, Yu F, Zhu Y, Xu G, Zhang Z, Tang C, Du J (2007) Hydrogen sulfide downregulates the aortic L-arginine/nitric oxide pathway in rats. Am J Physiol Regul Integr Comp Physiol 293:R1608–R1618
Ghasemi M, Dehpour AR, Moore KP, Mani AR (2012) Role of endogenous hydrogen sulfide in neurogenic relaxation of rat corpus cavernosum. Biochem Pharmacol 83:1261–1268
Goldstein I, Young JM, Fischer J, Bangerter K, Segerson T, Taylor T (2003) Vardenafil a new phosphodiesterase type 5 inhibitor, in the treatment of erectile dysfunction in men with diabetes: a multicenter double-blind placebo-controlled fixed-dose study. Diabetes Care 26:777–783
Gopalakrishnan M, Shieh CC (2004) Potassium channel subtypes as molecular targets for overactive bladder and other urological disorders. Expert Opin Ther Targets 8:437–458
Gratzke C, Streng T, Waldkirch E, Sigl K, Stief C, Andersson KE, Hedlund P (2009) Transient receptor potential A1 (TRPA1) activity in the human urethra—evidence for a functional role for TRPA1 in the outflow region. Eur Urol 55:696–704
Grgic I, Kaistha BP, Hoyer J, Köhler R (2009) Endothelial Ca + −activated K+ channels in normal and impaired EDHF-dilator responses–relevance to cardiovascular pathologies and drug discovery. Br J Pharmacol 157:509–526
Guo H, Gai JW, Wang Y, Jin HF, Du JB, Jin J (2012) Characterization of hydrogen sulfide and its synthases, cystathionine β-synthase and cystathionine γ-lyase, in human prostatic tissue and cells. Urology 79:483, e1-5
Hackett G, Kirby M, Sinclair AJ (2014) Testosterone deficiency, cardiac health, and older men. Int J Endocrinol 2014:143763
Isidori AM, Buvat J, Corona G, Goldstein I, Jannini EA, Lenzi A, Porst H, Salonia A, Traish AM, Maggi M (2014) A critical analysis of the role of testosterone in erectile function: from pathophysiology to treatment-a systematic review. Eur Urol 65:99–112
Jacob C, Anwar A, Burkholz T (2008) Perspective on recent developments on sulfur-containing agents and hydrogen sulfide signaling. Planta Med 74:1580–1592
Jiang B, Tang G, Cao K, Wu L, Wang R (2010) Molecular mechanism for H(2)S-induced activation of K(ATP) channels. Antioxid Redox Signal 12:1167–1178
Kajioka S, Nakayama S, Asano H, Seki N, Naito S, Brading AF (2008) Levcromakalim and MgGDP activate small conductance ATP-sensitive K + channels of K+ channel pore 6.1/sulfonylurea receptor 2A in pig detrusor smooth muscle cells: uncoupling of cAMP signal pathways. J Pharmacol Exp Ther 327:114–123
Kardong KV (2005) The urogenital system. In: Vertebrates: compartive anatomy, function, evolution. McGraw Hill Higher Education, New York, pp 577–578 (Chapter 14)
Karicheti V, Christ GJ (2001) Physiological roles for K+ channels and gap junctions in urogenital smooth muscle: implications for improved understanding of urogenital function, disease and therapy. Curr Drug Targets 2:1–20
Kashfi K (2014) Anti-cancer activity of new designer hydrogen sulfide-donating hybrids. Antioxid Redox Signal 20:831–846
Kashfi K, Olson KR (2013) Biology and therapeutic potential of hydrogen sulfide and hydrogen sulfide-releasing chimeras. Biochem Pharmacol 85:689–703
Li J, Li Y, Du Y, Mou K, Sun H, Zang Y, Liu C (2011) Endogenous hydrogen sulfide as a mediator of vas deferens smooth muscle relaxation. Fertil Steril 95:1833–1835
Li Y, Zang Y, Fu S, Zhang H, Gao L, Li J (2012) H2S relaxes vas deferens smooth muscle by modulating the large conductance Ca2 + −activated K+ (BKCa) channels via a redox mechanism. J Sex Med 9:2806–2813
Li L, Fox B, Keeble J, Salto-Tellez M, Winyard PG, Wood ME, Moore PK, Whiteman M (2013) The complex effects of the slow-releasing hydrogen sulfide donor GYY4137 in a model of acute joint inflammation and in human cartilage cells. J Cell Mol Med 17:365–376
Matsunami M, Miki T, Nishiura K, Hayashi Y, Okawa Y, Nishikawa H, Sekiguchi F, Kubo L, Ozaki T, Tsujiuchi T, Kawabata A (2012) Involvement of the endogenous hydrogen sulfide/Ca(v) 3.2 T-type Ca2+ channel pathway in cystitis-related bladder pain in mice. Br J Pharmacol 167:917–928
Meng J, Ganesan Adaikan P, Srilatha B (2013) Hydrogen sulfide promotes nitric oxide production in corpus cavernosum by enhancing expression of endothelial nitric oxide synthase. Int J Impot Res 25:86–90
Milsom I, Abrams P, Cardozo L, Roberts RG, Thuroff J, Wein AJ (2001) How widespread are the symptoms of an overactive bladder and how are they managed? A population-based prevalence study. BJU Int 87:760–766
Mirone V, Sorrentino R, di Villa BR, Imbimbo C, Palmieri A, Fusco F, Tajana G, Cirino G (2000) A standardized procedure for using human corpus cavernosum strips to evaluate drug activity. J Pharmacol Toxicol Methods 44:477–482
Munaron L, Avanzato D, Moccia F, Mancardi D (2013) Hydrogen sulfide as a regulator of calcium channels. Cell Calcium 53:77–84
Mustafa AK, Sikka G, Gazi SK, Steppan J, Jung SM, Bhunia AK, Barodka VM, Gazi FK, Barrow RK, Wang R, Amzel LM, Berkowitz DE, Snyder SH (2011) Hydrogen sulfide as endothelium-derived hyperpolarizing factor sulfhydrates potassium channels. Circ Res 109:1259–1268
Nehra A, Jackson G, Miner M, Billups KL, Burnett AL, Buvat J, Carson CC, Cunningham GR, Ganz P, Goldstein I, Guay AT, Hackett G, Kloner RA, Kostis J, Montorsi P, Ramsey M, Rosen R, Sadovsky R, Seftel AD, Shabsigh R, Vlachopoulos C, Wu FC (2012) The Princeton III Consensus recommendations for the management of erectile dysfunction and cardiovascular disease. Mayo Clin Proc 87:766–778
Nelson CP, Rainbow RD, Brignell JL, Perry MD, Willets JM, Davies NW, Standen NB, Challiss RA (2011) Principal role of adenylyl cyclase 6 in K+ channel regulation and vasodilator signalling in vascular smooth muscle cells. Cardiovasc Res 91:694–702
Nilius B, Owsianik G, Voets T, Peters JA (2007) Transient receptor potential cation channels in disease. Physiol Rev 87:165–217
Olson KR (2014) Hydrogen sulfide as an oxygen sensor. Antioxid Redox Signal 22(5):377–397
Patacchini R, Maggi CA, Meli A (1990) Capsaicin-like activity of some natural pungent substances on peripheral endings of visceral primary afferents. Naunyn Schmiedebergs Arch Pharmacol 342:72–77
Patacchini R, Santicioli P, Giuliani S, Maggi CA (2004) Hydrogen sulfide (H2S) stimulates capsaicin-sensitive primary afferent neurons in the rat urinary bladder. Br J Pharmacol 142:31–34
Patacchini R, Santicioli P, Giuliani S, Maggi CA (2005) Pharmacological investigation of hydrogen sulfide (H2S) contractile activity in rat detrusor muscle. Eur J Pharmacol 509:171–177
Petkov GV, Heppner TJ, Bonev AD, Herrera GM, Nelson MT (2001) Low levels of KATP channel activation decrease excitability and contractility of urinary bladder. Am J Physiol Regul Integr Comp Physiol 280:R1427–R1433
Price DE, Gingell JC, Gepi-Attee S, Warcham K, Yates P, Boolell M (1998) Sildenafil: study of a novel oral treatment for erectile dysfunction in diabetic men. Diabet Med 15:821–825
Qiu X, Villalta J, Lin G, Lue TF (2012) Role of hydrogen sulfide in the physiology of penile erection. J Androl 33:529–535
Rendell MS, Rajfer J, Wicker PA, Smith MD (1999) Sildenafil for treatment of erectile dysfunction in men with diabetes. A randomized controlled trial. JAMA 281:421–426
Ruiz Rubio JL, Hernández M, Arcos L, Benedito S, Recio P, García P, García-Sacristán A, Prieto D (2004) Role of ATP-sensitive K+ channels in relaxation of penile resistance arteries. Urology 63:800–805
Saenz de Tejada I, Anglin G, Knight JR, Emmick JT (2002) Effects of tadalafil on erectile dysfunction in men with diabetes. Diabetes Care 25:2159–2164
Salloum FN, Chau VQ, Hoke NN, Abbate A, Varma A, Ockaili RA, Toldo S, Kukreja RC (2009) Phosphodiesterase-5 inhibitor, tadalafil, protects against myocardial ischemia/reperfusion through protein-kinase g-dependent generation of hydrogen sulfide. Circulation 120:S31–S36
Schultheiss D, Badalyan R, Pilatz A, Gabouev AI, Schlote N, Wefer J, von Wasielewski R, Mertsching H, Sohn M, Stief CG, Jonas U (2003) Androgen and estrogen receptors in the human corpus cavernosum penis: immunohistochemical and cell culture results. World J Urol 21:320–324
Shieh CC, Feng J, Buckner SA, Brioni JD, Coghlan MJ, Sullivan JP, Gopalakrishnan M (2001) Functional implication of spare ATP-sensitive K+ channels in bladder smooth muscle cells. J Pharmacol Exp Ther 296:669–675
Shukla N, Rossoni G, Hotston M, Sparatore A, Del Soldato P, Tazzari V, Persad R, Angelini GD, Jeremy JY (2009) Effect of hydrogen sulphide-donating sildenafil (ACS6) on erectile function and oxidative stress in rabbit isolated corpus cavernosum and in hypertensive rats. BJU Int 103:1522–1529
Spektor M, Rodriguez R, Rosenbaum RS, Wang HZ, Melman A, Christ GJ (2002) Potassium channels and human corporeal smooth muscle cell tone: further evidence of the physiological relevance of the Maxi-K channel subtype to the regulation of human corporeal smooth muscle tone in vitro. J Urol 167:2628–2635
Srilatha B, Adaikan PG, Moore PK (2006) Possible role for the novel gasotransmitter hydrogen sulphide in erectile dysfunction–a pilot study. Eur J Pharmacol 535:280–282
Srilatha B, Adaikan PG, Li L, Moore PK (2007) Hydrogen sulphide: a novel endogenous gasotransmitter facilitates erectile function. J Sex Med 4:1304–1311
Srilatha B, Hu L, Adaikan GP, Moore PK (2009) Initial characterization of hydrogen sulfide effects in female sexual function. J Sex Med 6:1875–1884
Srilatha B, Muthulakshmi P, Adaikan PG, Moore PK (2012) Endogenous hydrogen sulfide insufficiency as a predictor of sexual dysfunction in aging rats. Aging Male 15:153–158
Story GM, Peier AM, Reeve AJ, Eid SR, Mosbacher J, Hricik TR, Earley TJ, Hergarden AC, Andersson DA, Hwang SW, McIntyre P, Jegla T, Bevan S, Patapoutian A (2003) ANKTM1, a TRP-like channel expressed in nociceptive neurons, is activated by cold temperatures. Cell 112:819–829
Streng T, Axelsson HE, Hedlund P, Andersson DA, Jordt SE, Bevan S, Andersson KE, Högestätt ED, Zygmunt PM (2008) Distribution and function of the hydrogen sulfide-sensitive TRPA1 ion channel in rat urinary bladder. Eur Urol 53:391–399
Tang G, Yang G, Jiang B, Ju Y, Wu L, Wang R (2013) H2S is an endothelium-derived hyperpolarizing factor. Antioxid Redox Signal 19:1634–1646
Traish AM, Goldstein I, Kim NN (2007) Testosterone and erectile function: from basic research to a new clinical paradigm for managing men with androgen insufficiency and erectile dysfunction. Eur Urol 52:54–70
Venkateswarlu K, Giraldi A, Zhao W, Wang HZ, Melman A, Spektor M, Christ GJ (2002) Potassium channels and human corporeal smooth muscle cell tone: diabetes and relaxation of human corpus cavernosum smooth muscle by adenosine triphosphate sensitive potassium channel openers. J Urol 168:355–361
Vickers MA, Satyanarayana R (2002) Phosphodiesterase type 5 inhibitors for the treatment of erectile dysfunction in patients with diabetes mellitus. Int J Impot Res 14:466–471
Wang R (2003) The gasotransmitter role of hydrogen sulfide. Antioxid Redox Signal 5:493–501
Wang R (2009) Hydrogen sulfide: a new EDRF. Kidney Int 76:700–704
Xu S, Liu Z, Liu P (2014) Targeting hydrogen sulfide as a promising therapeutic strategy for atherosclerosis. Int J Cardiol 172:313–317
Yang J, Wang T, Yang J, Rao K, Zhan Y, Chen RB, Liu Z, Li MC, Zhuan L, Zang GH, Guo SM, Xu H, Wang SG, Liu JH, Ye ZQ (2013) S-allyl cysteine restores erectile function through inhibition of reactive oxygen species generation in diabetic rats. Andrology 1:487–494
Yetik-Anacak G, Sorrentino R, Linder AE, Murat N (2014) Gas what: NO is not the only answer to sexual function. Br J Pharmacol 172(6):1434–1454
Yildiz O, Seyrek M, Irkilata HC, Yildirim I, Tahmaz L, Dayanc M (2009) Testosterone might cause relaxation of human corpus cavernosum by potassium channel opening action. Urology 74:229–232
Zhao W, Wang R (2002) H2S-induced vasorelaxation and underlying cellular and molecular mechanisms. Am J Physiol Heart Circ Physiol 283:H474–H480
Zhao W, Zhang J, Lu Y, Wang R (2001) The vasorelaxant effect of H2S as a novel endogenous gaseous KATP channel opener. EMBO J 20:6008–6016
Zhao K, Li S, Wu L, Lai C, Yang G (2014) Hydrogen sulfide represses androgen receptor transactivation by targeting at the second zinc finger module. J Biol Chem 289:20824–20835
Zuo C, Huang YM, Jiang R, Yang HF, Cheng B, Chen F (2014) Endogenous hydrogen sulfide and androgen deficiency-induced erectile dysfunction in rats. Zhonghua Nan Ke Xue 20:605–612
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2015 Springer International Publishing Switzerland
About this chapter
Cite this chapter
di Villa Bianca, R.d., Cirino, G., Sorrentino, R. (2015). Hydrogen Sulfide and Urogenital Tract. In: Moore, P., Whiteman, M. (eds) Chemistry, Biochemistry and Pharmacology of Hydrogen Sulfide. Handbook of Experimental Pharmacology, vol 230. Springer, Cham. https://doi.org/10.1007/978-3-319-18144-8_5
Download citation
DOI: https://doi.org/10.1007/978-3-319-18144-8_5
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-18143-1
Online ISBN: 978-3-319-18144-8
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)