
Abstract
Phosphorus is one of the most important elements for plants but limiting with time due to extensive cropping and changing environments. Optimum crop productions are on the verge of decline due to phosphorus deficiency worldwide. Phosphorus use efficiency can be improved by improving phosphorus uptake, transportation, and metabolism. In this chapter we will discuss Phosphorus as a plant nutrient, Phosphorous use efficiency (PUE) plant responses to phosphorous deficiency and will unfold nature mysteries by focusing on genetic and molecular factors involved in phosphorous deficiency, and propose transgenic approaches to improve PUE.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
Introduction
Phosphorus (P) is a crucial plant macronutrient for plant growth and development. It cannot be substituted by any other element, and provision of adequate amount is necessary for plant survival. Phosphorus fertilizers have increase crop production over the past 50 years and help in ensuring food security for increasing population (Pavinato et al., 2020). Phosphorus is an integral part of plant cells and performs key functions in plant metabolism such as photosynthesis, sugar metabolism and translocation, and transformation of metabolites. The normal phosphorus concentrations in plants are from 0.1% to 0.5% (Gagnon et al., 2020).
Phosphorus is an important phytonutrient complicated in several biochemical pathways and biosynthesis of biomolecules like lipids, nucleic acids, and cellular membranes. Phosphorus also promotes the signal cascade of cells by acting as a signal mediator and is an important for metabolism. Due to its extensive use in agriculture, phosphorus resources are limited. Therefore, it is vital to formulate scientific strategies to improve the efficiency of phosphorus utilization and recycle in the future. The soluble, bioavailable form of phosphorus (phosphate; Pi) used for absorption is readily removed from the topsoil, which can cause serious havoc. In addition to this, scouring Pi from the topsoil requires a constant supply of pi to preserve satisfactory fertilizer levels, which worsen the condition. To overcome phosphorus stress, plants are recognized to undergo fundamental cellular changes in physiology, metabolism, and hormone balance.
Better understanding to these govern changes can result in improving agronomic traits, resource conservation and step forward for sustainable agriculture as well as the use of biotechnology strategies to improve crop phosphorus utilization efficiency.
Phosphorus Uptake and Transport
As soil is the primary source, Pi enters through root hairs, root tips, or outermost layers of root cells (Zahra et al., 2020). The uptake can be increased by some soil microbes especially mycorrhizal fungi that grow as a symbiont with plants. Phosphorus is taken up by plant roots as the primary orthophosphate ion (H2PO4−), which is the only possible available form, but in some cases, it is can also be absorbed as (HPO4−2) secondary orthophosphate (Amin et al., 2017; Ceulemans et al., 2017). Once entered into the plant roots, P may be accumulated for the time in root tissues or transported to the aerial parts of the plant. Through several biochemical reactions, it is incorporated to organic compounds, like DNA and RNA (nucleic acids), phosphoproteins and lipids, various antioxidant enzymes, and energy-rich phosphate compound, that is, adenosine triphosphate (ATP). Phosphorus in inorganic as well as in organic form moved throughout the plant body and is available for different metabolic processes (Galatro et al., 2020).
Phosphorus Use Efficiency
There are numerous definitions for PUE (Brown et al., 2012). P absorption efficiency means the ability of plants to extract phosphorus from the soil, and PUE stands for biomass production capacity of the absorbed phosphorus (Wang et al., 2010). The PUE is improved either by improving intake or optimizing its internal usage (Reymond et al., 2006). Importance of each measure depends on the environment in which the crop grows. In agricultural ecosystems with high phosphorus consumption, the importance of phosphorus absorption is reduced because of the higher availability of phosphorus. Under these conditions, increasing the efficiency of phosphorus utilization and reducing the amount of phosphorus recovered during harvesting will bring agronomic benefits. In contrast, in environment with low P availability, both optimum acquisition and utilization are of benefit. Given the collective importance of this important macronutrient, we must deepen our understanding of the phosphorus-deficiency response due to cellular and physiological changes to ensure that plants with higher phosphorus efficiency are developed to increase agricultural productivity.
Plant Physiological Response to Phosphorus
When exposed to phosphorus-deficient conditions, plants exhibit several characteristics and physiological responses. Due to the lack of phosphorus, the development of buds and flowers is greatly slowed down, resulting in delayed plant growth and reduced bud branches. Maintaining root stem cells under phosphorus-deficient conditions requires (ER) endoplasmic reticulum to reside PDR2 (phosphate deficiency response 2, P5-like ATPase), which interacts genetically with low phosphate root 1, a multi-copper oxidase that is functionally important for phosphorus sensing at the root tip (Reymond et al., 2006: Wang et al., 2010). The accumulation of anthocyanin pigments usually occurs in leaf tissues to protect chloroplasts and nucleic acids from strong light or ultraviolet rays (Bustos et al., 2010). In addition, the growth of the taproot is strongly inhibited, while the development of lateral roots and hairy roots is strongly stimulated, which ultimately leads to a higher root/bud ratio (Brown et al., 2012).
Roots are an important entry point for phosphorus. In the case of phosphorus deficiency, changes in root structure are essential to maximize the effective nutrient absorption for plant survival (Niu et al., 2012). When phosphorus is lacking to utilize phosphorus in the form of phospholipids, organic Pi transporters are also induced (Ramaiah et al., 2011). Although plants have very limited ability to obtain phosphorus from other organic sources outside the roots, they can directly use phosphorus from phytate (Richardson et al., 2001). In addition, genes that respond to phosphorus deficiency, such as genes induced by phosphorus deficiency (IPS1), RNS1, phospholipase DZ2 (PLDZ2), Pht1;4, and At4, are strongly induced under phosphorus-free conditions (Jain et al., 2009). Due to the low availability of phosphorus in the soils, plants are often deficient in phosphorus. Plants that are prone to phosphorus (Pi) deficiency may suffer from inhibition of primary root growth, reduced phosphorus concentration in various tissues, and reduced photosynthesis rate, but the utilization of photosynthetic phosphorus is higher (López-Arredondo et al., 2014). The plant response to pi deficiency based mainly on experimental data obtained by many scientists on herb model plants like Arabidopsis thaliana has a different phosphorus feeding strategy than wood plants (Rennenberg & Herschbach, 2013) (Table 16.1).
Molecular and Genetic Responses to Phosphorus Deficiency
In phosphorus-rich and low phosphorus conditions, different transcription factor groups perform regulatory functions to maintain phosphorus in plant, which is essential for normal plant metabolism. The first transcription factor described as related to phosphorus deficiency is phosphorus deficiency response 1 (PHR1), which is a transcription factor TF (MYB) that responds to phosphorus deficiency at posttranscriptional level (Rubio et al., 2001). PHR1 regulates genes that respond to phosphorus deficiency, including genes encoding AtIPS1 and ribonuclease. Responsive genes, such as AtIPS1, (AtPHT1) phosphate transporter1, (ribonuclease1) AtRNS1, (PHO1) phosphate 1, (SQD1) UDP-sulfoquinovose synthase 1, and (DGD2) digalactosyldiacylglycerol synthase 2, have common motifs (GNATATNC). It was also found that transcription factor responds to low phosphorus levels at the transcription level, which is the zinc finger C2H2 is in the nucleus (Devaiah et al., 2007). The main helix TF (bHLH) was found to be responsible for the resistance of rice to phosphorus stress (Yi et al., 2005). PHO1 gene family has been involved in phosphorus transportation from roots to shoots in Arabidopsis and rice (Secco et al., 2010). In phosphorus-rich conditions, the (AtIPS1) and GUS reporter line showed significant GUS activity, which was limited by the endoderm of the mutant pho1., while no GUS activity was perceived in wild plants, PHO1 evolved as Pi exporter (Hamburger et al., 2002).
Phosphate monoesters are hydrolyzed by purple acid phosphatase (PAP) to Pi. The genes encoding PAP are strongly induced by phosphorus deficiency (Li et al., 2012). The resistance of plants to phosphorus deficiency is partly due to the release of acid phosphatase. In Arabidopsis, AtPAP26 is one of 29 acid phosphatases. It exhibits significant phosphatase activity during phosphorus deficiency and leaf aging and has dual functions of secretion or local vacuole form (Robinson et al., 2012). Posttranslational regulation clearly involves signal transduction for phosphorus deficiency. AtSIZ1 is a small plant, while ubiquitin-like modified E3 ligase is a regulator of plant stress response (Miura et al., 2011). When phosphorus is deficient, phosphorus can be recovered from the nucleic acid source. The lack of phosphorus strongly induces ribonuclease outside and inside the cell. Although AtRNS3 is expressed in different tissues and remains relatively stable in the case of phosphorus deficiency, AtRNS1 and AtRNS2 are strongly induced by phosphorus deficiency. AtRNS1 is highly expressed in flowers, and the protein it encodes is secreted outside the cell, while AtRNS2 remains inside the cell (Deal et al., 2007).
Noncoding RNA has become a key regulator of phosphorus deficiency signals. In Arabidopsis, selection of microRNAs (miRNAs) that specifically respond to phosphorus deficiency not to other nutrients has identified miR399 family as the mediator of the phosphorus deficiency response (Abbas et al., 2022; Fujii et al., 2005). There is increasing evidence that miR399s regulate signaling in response to phosphorus deficiency. The posttranscriptional closure of the AtPHO2 gene is precisely controlled and regulated by other noncoding RNAs of the AtIPS1 family, which are strongly induced in phosphorus-deficient conditions (Franco-Zorrilla et al., 2007). Cytokinins regulate the responses to environmental stress, including strong interactions and cross talks with ABA (Ha et al., 2012). More and more evidences show that cytokinins can act as negative mediators in response to phosphorus deficiency. Many independent studies have shown that the cytokinin signal pathway affects the phosphorus signal pathway. P. vulgaris response regulator 1 (PvRR1) transcript increases in the absence of P, N, and K (Camacho et al., 2008). Although treatment with exogenous cytokinin will increase the expression of PvRR1 under sufficient conditions, it will decrease the expression level of PvRR1 under conditions of phosphorus deficiency (Franco-Zorrilla et al., 2007). Ethylene also regulates the phosphorus deficiency responses, especially in the development of hair roots (Li et al., 2011). Overexpressed Pht1; Compared with wild plants under sufficient phosphorus conditions, 5 exhibits altered phosphorus redistribution, with lower phosphorus content in buds and higher phosphorus content in roots and pods, leading to premature senescence (Nagarajan et al., 2011). Numerous new hps mutants, namely, the hps3 and hps4, have been characterized. Hps3 is highly sensitive to phosphorus deficiency; however, the accumulation of anthocyanins is one of the typical reactions of phosphorus deficiency, but it accumulates less with phosphorus deficiency (Wang et al., 2012). The analysis of differential gene expression among the root tips of low-sensitivity mutants to phosphorus 4 (lpi4) and wild plant under low phosphorus conditions revealed that a large group of genes belonging to the family of jasmonic acid and ethylene synthesis and signal pathways have been changed (Chacon-Lopez et al., 2011).
A growing body of evidence also shows that gibberellin plays a role in regulating the phosphorus deficiency response. The proteins involved in gibberellin signal in the DELLA domain are composed of gibberellin-responsive repressor proteins ga13 (RGA), RGAlike (RGL) 1, RGL2, and RGL3, which are key mediators of gibberellin signaling in Arabidopsis (Sheerin et al., 2011). In the case of phosphorus deficiency, the lipid composition of the membrane will change dynamically. Phospholipids are the main component of membrane lipids and can also be regarded as storage of phosphorus. Genes such as MGD2 and MGD3 involved in membrane lipid remodeling are regulated by AUX and AUX signal. The expression of MGD2 and MGD3 is inhibited by AUX transport inhibitors (Kobayashi et al., 2009). Genes such as DGD2, SQD1, NPC4, and NPC5 are inhibited in the expression of slr and arf7 arf19 mutants, resulting in a significant decrease in DGDG and SQDG levels (Narise et al., 2010). Although the expression of these genes related to lipid metabolism is controlled by auxin signals and is mainly caused by phosphorus deficiency, this auxin-mediated regulation may be an indirect effect, because the elements that respond to auxin are not existing in their composition. Promoter sequences and genes that respond to phosphorus deficiency are not removed in transactivation systems. Various methods and strategies have been tried to produce effective transgenic plants, such as the high expression of ZmPTF1, resulting in better root development, increased ear yield, and the largest grain size in transgenic corn plants (Zhou et al., 2008). Phosphorus starvation tolerance gene 1 (PSTOL1) encodes a phosphorus-specific protein kinase, which has been shown to be important for increasing grain yield in low-phosphorus soils and promoting early root growth of various rice varieties, which is the tolerance to phosphorus deficiency (Gamuyao et al., 2012). This gene is constantly missing in the phosphorus-deficiency genome of modern rice varieties. Therefore, PSTOL1 is an important gene to be engineered to improve plant resistance to low-phosphorus conditions. Target genes include genes encoding transport proteins PAP, TF, Pi, protein kinase, oxidoreductase, and genes involved in the production of organic acids. Although the Pi transporter genes seem to be suitable and promising candidates for increasing phosphorus uptake, these genes can cause growth retardation due to excessive phosphorus levels (Li et al., 2011). Overexpression of PAP or excretion of malic acid improves Pi absorption and increases biomass growth and production (Lu et al., 2012).
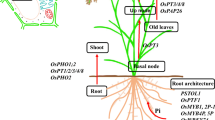
In Arabidopsis, PHT1, PHT2, PHO1, and PHO2 are gene families encoding pi transporters, PHT1 and AtPHT1 that are transporters, which mainly work at low Pi concentrations (López-Arredondo et al., 2014). AtPHT1;5, it participates in the translocation of Pi from the source to the receiving organ according to the development signal and the level of phosphorus (Nagarajan et al., 2011). AtPHT1;9 is responsible for the absorption of Pi at the root-soil interface and the movement of Pi from root to shoot (Lapis-Gaza et al., 2014; Remy et al., 2012). AtPHT2;1 is situated in the membrane of the inner shell of the chloroplast and acts as a low-affinity Pi transporter to ensure the distribution of Pi in plants (Versaw & Harrison, 2002). Also it plays role in the transfer of Pi to root xylem vessels (Stefanovic et al., 2007). AtPHO2 encodes E2-bound ubiquitin and prevents excessive Pi accumulation in shoots (Kant et al., 2011). According to the internal and external states of phosphorus, these genes show different transcriptional expressions in Arabidopsis (López-Arredondo et al., 2014). Homologs of AtPHO2 have been identified in the genome of Populus trichocarpa, and PHT1 phosphate transporter gene family is identified and characterized in mycorrhiza P. trichocarpa (Loth-Pereda et al., 2011; Tuskan et al., 2006). However, little is known about their expression profile in response to phosphorus deficiency in poplar species (Fig. 16.1).

Factors affecting phosphorus use efficiency
Role of Phosphorus in Plant Metabolism
Photosynthesis
Photosynthesis is an important biochemical reaction in nature. Plant converts light energy into sugars and oxygen with the energy being captured in ATP (Liang et al., 2020). This ATP is further available as a source of energy for many other metabolic processes that occur in plant. The carbohydrate produced is helpful in building other cell organelles and storge components. Most of the converted carbohydrate within plant body involve phosphorylated forms. Phosphorus is involved in the photosynthesis at a crucial point at which energy is transformed from light into sugar, so in order to achieve optimal growth, the phosphorus availability in soil solution should be ensure (Feder et al., 2020; Shah et al., 2021).
Plant Energy Reactions
P is critical in energy transfer mechanisms of plant. Phosphorus in the form of adenosine diphosphate (ADP) and ATP is a source of energy that drives various biochemical reactions within plant (Zhao & Yang, 2020). Phosphorus in plants transfers high energy phosphate molecules to the other molecules and provides the activation energy to photosynthesis, respiration, carbohydrates formation, and many other chemical reactions to occur (Fu & Zhang, 2020).
Genetic Transfer
P is a major component of various substances that constitute genes and chromosomes, and sugar phosphate is an important structural constituent of DNA and RNA (Cu et al., 2020). The biochemical structures of DNA and RNA are linked together with the help of phosphorus. An adequate phosphorus nutrition is essential for genetic transfer code from one generation to the next and is essential for the development of new cells. A significant concentration of P in the form of phytin is found in seeds of different agricultural and garden crops, and P is also believed to be essential for seed germination and development (Xu et al., 2018). About 60–70% of P in cereals grain and about 50% in legumes are stored as phytin or similar compounds. It is noted that an improper supply of P can retard seed number, size, and seed viability.
Nutrient Transport
Nutrient availability to crops depends on nutrient supply in the rhizosphere and root surface area. Roots are able to absorb nutrients when they come into contact with active cells, so P uptake is highly dependent on plant’s root system (Suleman et al., 2018). Nutrients contact the root cells mainly by three mechanisms: diffusion, mass flow, and root interception. Root interception is a major pathway for P uptake, and better root growth provides additional root surface area for P uptake. Legume crops help in fixing nitrogen through their roots in the soil, but this fixation can only be possible with adequate phosphorous, which ensure root development (Ferrol et al., 2019). Movement of essential nutrients within the plant depends chiefly on transport through plasma membrane, which required energy to suppress the force of osmosis (Palmgren, 2001). High energy P compounds and ATP provides the needed energy for transport of macro- and micronutrients from root to aerial parts of plant (Bashir et al., 2019).
Transgenic Approaches to Improve Phosphorus Use Efficiency
For sustainable agriculture with the changing environment, it is very important to improve phosphorus use efficiency using latest available resources, and it can only be done by improving the intake capability and internal efficient use of phosphorous; this can be done by efficient breeding and proper selection as all the phosphorus sources are finite (Heuer et al., 2017; Thudi et al., 2021). Within this new era of modern agriculture, PUE is improved either by opting latest cultivation practices like hydroponic systems, using new fertigation systems like drip irrigation and fumigation, and the second way is to improve plant varieties (Lakhiar et al., 2018; Sambo et al., 2019). With the advancement in the field of plant genetics and plant biotechnology, it is possible now to engineer plants according to needs and according to immerging issues. With the reduction in sequencing costs, more and more genomes are available now, which is revolutionizing agriculture by revolution in fields of omics. New techniques in biotechnology like RNA-induced gene silencing, CRISPR-mediated gene editing, and TALENS-mediated gene editing equipped plant scientists with the enough capability to alter plant genetics and plant physiology (Mutezo et al., 2021; Steinwand & Ronald, 2020). Plant internal use efficiency can be improved by overexpressing phosphorus transporters and genes involved in phosphorus assimilation (Shen et al., 2011).
Future Prospects
Earth climate is changing rapidly, and element depletion is happening at the rapid pace, which was not noticed before, and this may also be due to consecutive cultivation of crops to secure food security. Alike other nutrients, soil is the only source of phosphorus, and supplies are shrinking. Phosphorus deficiency is very common in all our major crops, minimized by fertilizer supplement, which is not a permanent solution to this problem. This problem will increase with time and cause substantial decrease in yields, so it needs to be tackled in time. For sustainable agriculture and for the sake of future generations, we need to retain potential sources and develop varieties with increased phosphorus use efficiency.
References
Abbas, A., Shah, A. N., Tanveer, M., Ahmed, W., Shah, A. A., Fiaz, S., Waqas, M. M., & Ullah, S. (2022). MiRNA fine tuning for crop improvement: using advance computational models and biotechnological tools. Molecular Biology Reports, 1–14. https://doi.org/10.1007/s11033-022-07231-5
Amin, A., Nasim, W., Mubeen, M., Nadeem, M., Ali, L., Hammad, H. M., Sultana, S. R., Jabran, K., ur Rehman, M. H., Ahmad, S., Awais, M., Shah, A. N., et al. (2017). Optimizing the phosphorus use in cotton by using CSM-CROPGRO-cotton model for semi-arid climate of Vehari-Punjab, Pakistan. Environmental Science and Pollution Research, 24(6), 5811–5823. https://doi.org/10.1007/s11356-016-8311-8
Bashir, K., Seki, M., & Nishizawa, N. K. (2019). The transport of essential micronutrients in rice. Molecular Breeding, 39(12), 1–17.
Brown, L. K., George, T. S., Thompson, J. A., et al. (2012). What are the implications of variation in root hair length on tolerance to phosphorus deficiency in combination with water stress in barley (Hordeum vulgare). Annals of Botany, 110, 319–328.
Bustos, R., Castrillo, G., Linhares, F., et al. (2010). A central regulatory system largely controls transcriptional activation and repression responses to phosphate starvation in Arabidopsis. PLoS Genetics, 6, e1001102.
Camacho, Y., Martinez-Castilla, L., Fragoso, S., et al. (2008). Characterization of a type A response regulator in the common bean (Phaseolus vulgaris) in response to phosphate starvation. Physiologia Plantarum, 132, 272–282.
Ceulemans, T., Bodé, S., Bollyn, J., Harpole, S., Coorevits, K., Peeters, G., et al. (2017). Phosphorus resource partitioning shapes phosphorus acquisition and plant species abundance in grasslands. Nature Plants, 3(2), 1–7.
Chacon-Lopez, A., Ibarra-Laclette, E., Sanchez-Calderon, L., et al. (2011). Global expression pattern comparison between low phosphorus insensitive 4 and WT Arabidopsis reveals an important role of reactive oxygen species and jasmonic acid in the root tip response to phosphate starvation. Plant Signaling & Behavior, 6, 382–392.
Chen, J., Xu, L., Cai, Y., & Xu, J. (2008). QTL mapping of phosphorus efficiency and relative biologic characteristics in maize (Zea mays L.) at two sites. Plant and Soil, 313, 251–266.
Chen, J., Xu, L., Cai, Y., & Xu, J. (2009). Identification of QTLs for phosphorus utilization efficiency in maize (Zea mays L.) across P levels. Euphytica, 167, 245–252.
Chen, J., et al. (2011). Identification of QTLs for biomass production in maize (Zea mays L.) under different phosphorus levels at two sites. Frontiers of Agriculture in China, 5, 152–161.
Cu, S. T., Guild, G., Nicolson, A., Velu, G., Singh, R., & Stangoulis, J. (2020). Genetic dissection of zinc, iron, copper, manganese and phosphorus in wheat (Triticum aestivum L.) grain and rachis at two developmental stages. Plant Science, 291, 110338.
Deal, R. B., Topp, C. N., McKinney, E. C., & Meagher, R. B. (2007). Repression of flowering in Arabidopsis requires activation of FLOWERING LOCUS C expression by the histone variant H2A.Z. Plant Cell, 19, 74–83.
Devaiah, B. N., Nagarajan, V. K., & Raghothama, K. G. (2007). Phosphate homeostasis and root development in Arabidopsis are synchronized by the zinc finger transcription factor ZAT6. Plant Physiology, 145, 147–159.
Feder, D., McGeary, R. P., Mitić, N., Lonhienne, T., Furtado, A., Schulz, B. L., et al. (2020). Structural elements that modulate the substrate specificity of plant purple acid phosphatases: avenues for improved phosphorus acquisition in crops. Plant Science, 294, 110445.
Ferrol, N., Azcón-Aguilar, C., & Pérez-Tienda, J. (2019). Arbuscular mycorrhizas as key players in sustainable plant phosphorus acquisition: an overview on the mechanisms involved. Plant Science, 280, 441–447.
Franco-Zorrilla, J. M., Valli, A., Todesco, M., et al. (2007). Target mimicry provides a new mechanism for regulation of microRNA activity. Nature Genetics, 39, 1033–1037.
Fu, W., & Zhang, X. (2020). Global phosphorus dynamics in terms of phosphine. npj Climate and Atmospheric Science, 3(1), 1–6.
Fujii, H., Chiou, T. J., Lin, S. I., et al. (2005). A miRNA involved in phosphatestarvation response in Arabidopsis. Current Biology, 15, 2038–2043.
Gagnon, B., Ziadi, N., Bélanger, G., & Parent, G. (2020). Validation and use of critical phosphorus concentration in maize. European Journal of Agronomy, 120, 126147.
Galatro, A., Ramos-Artuso, F., Luquet, M., Buet, A., & Simontacchi, M. (2020). An update on nitric oxide production and role under phosphorus scarcity in plants. Frontiers in Plant Science, 11, 413.
Gamuyao, R., Chin, J. H., Pariasca-Tanaka, J., et al. (2012). The protein kinase Pstol1 from traditional rice confers tolerance of phosphorus deficiency. Nature, 488, 535–539.
Ha, S., Vankova, R., Yamaguchi-Shinozaki, K., et al. (2012). Cytokinins: metabolism and function in plant daptation to environmental stresses. Trends in Plant Science, 17, 172–179.
Hamburger, D., Rezzonico, E., MacDonald-Comber Petetot, J., et al. (2002). Identification and characterization of the Arabidopsis PHO1 gene involved in phosphate loading to the xylem. Plant Cell, 14, 889–902.
Heuer, S., et al. (2017). Improving phosphorus use efficiency: a complex trait with emerging opportunities. The Plant Journal, 90, 868–885.
Jain, A., Poling, M. D., Smith, A. P., et al. (2009). Variations in the composition of gelling agents affect morphophysiological and molecular responses to deficiencies of phosphate and other nutrients. Plant Physiology, 150, 1033–1049.
Kaeppler, S. M., Parke, J. L., Mueller, S. M., Senior, L., Stuber, C., & Tracy, W. F. (2000). Variation among maize inbred lines and detection of quantitative trait loci for growth at low phosphorus and responsiveness to arbuscular mycorrhizal fungi. Crop Science, 40, 358–364.
Kant, S., Peng, M., & Rothstein, S. J. (2011). Genetic regulation by NLA and microRNA827 for maintaining nitrate-dependent phosphate homeostasis in Arabidopsis. PLoS Genetics, 7, e1002021. https://doi.org/10.1371/journal.pgen.1002021
Kobayashi, K., Awai, K., Nakamura, M., et al. (2009). Type-B monogalactosyldiacylglycerol synthases are involved in phosphate starvation-induced lipid remodeling, and are crucial for low-phosphate adaptation. The Plant Journal, 57, 322–331.
Lakhiar, I. A., Jianmin, G., Syed, T. N., Chandio, F. A., Buttar, N. A., & Qureshi, W. A. (2018). Monitoring and control systems in agriculture using intelligent sensor techniques: a review of the aeroponic system. Journal of Sensor, 2018, 8672769.
Lapis-Gaza, H. R., Jost, R., & Finnegan, P. M. (2014). Arabidopsis PHOSPHATE TRANSPORTER1 genes PHT1; 8 and PHT1;9 are involved in root-to-shoot translocation of orthophosphate. BMC Plant Biology, 14, 334.
Li, M., Guo, X., Zhang, M., Wang, X., Zhang, G., Tian, Y., & Wang, Z. (2010). Mapping QTLs for grain yield and yield components under high and low phosphorus treatments in maize (Zea mays L.). Plant Science, 178, 454–462.
Li, Z., Gao, Q., Liu, Y., et al. (2011). Overexpression of transcription factor ZmPTF1 improves low phosphate tolerance of maize by regulating carbon metabolism and root growth. Planta, 233, 1129–1143.
Li, C., Gui, S., Yang, T., et al. (2012). Identification of soybean purple acid phosphatase genes and their expression responses to phosphorus availability and symbiosis. Annals of Botany, 109, 275–285.
Liang, X., Zhang, T., Lu, X., Ellsworth, D. S., Bassiri Rad, H., You, C., et al. (2020). Global response patterns of plant photosynthesis to nitrogen addition: a meta-analysis. Global Change Biology, 26(6), 3585–3600.
López-Arredondo, D. L., Leyva-González, M. A., González-Morales, S. I., López-Bucio, J., & Herrera-Estrella, L. (2014). Phosphate nutrition: improving low-phosphate tolerance in crops. Annual Review of Plant Biology, 65, 95–123. https://doi.org/10.1146/annurev-arplant-050213-035949
Loth-Pereda, V., Orsini, E., Courty, P. E., et al. (2011). Structure and expression profile of the phosphate Pht1 transporter gene family in mycorrhizal Populus trichocarpa. Plant Physiology, 156, 2141–2154. https://doi.org/10.1104/pp.111.180646
Miura, K., Lee, J., Gong, Q., et al. (2011). SIZ1 regulation of phosphate starvation-induced root architecture remodeling involves the control of auxin accumulation. Plant Physiology, 155, 1000–1012.
Mutezo, W., Sedibe, M. M., Mofokeng, A., Shargie, N., & Soko, T. (2021). The application of CRISPR/Cas9 technology in the management of genetic and nongenetic plant traits. International Journal of Agronomy, 2021, 9993784.
Nagarajan, V. K., Jain, A., Poling, M. D., Lewis, A. J., Raghothama, K. G., & Smith, A. P. (2011). Arabidopsis Pht1;5 mobilizes phosphate between source and sink organs and influences the interaction between phosphate homeostasis and ethylene signaling. Plant Physiology, 156, 1149–1163. https://doi.org/10.1104/pp.111.174805
Narise, T., Kobayashi, K., Baba, S., et al. (2010). Involvement of auxin signaling mediated by IAA14 and ARF7/19 in membrane lipid remodeling during phosphate starvation. Plant Molecular Biology, 72, 533–544.
Niu, Y. F., Chai, R. S., Jin, G. L., et al. (2012). Responses of root architecture development to low phosphorus availability: a review. Annals of Botany, 118, 1–18. https://doi.org/10.1093/aob/mcs285
Palmgren, M. G. (2001). Plant plasma membrane H+-ATPases: powerhouses for nutrient uptake. Annual Review of Plant Biology, 52(1), 817–845.
Pavinato, P. S., Cherubin, M. R., Soltangheisi, A., Rocha, G. C., Chadwick, D. R., & Jones, D. L. (2020). Revealing soil legacy phosphorus to promote sustainable agriculture in Brazil. Scientific Reports, 10(1), 1–11.
Ramaiah, M., Jain, A., Baldwin, J. C., et al. (2011). Characterization of the phosphate starvation-induced glycerol-3-phosphate permease gene family in Arabidopsis. Plant Physiology, 157, 279–291.
Reiter, R. S., Coors, J. G., Sussman, M. R., & Gabelman, W. H. (1991). Genetic analysis of tolerance to low-phosphorus stress in maize using restriction fragment length polymorphisms. Theoretical and Applied Genetics, 82, 561–568.
Remy, E., Cabrito, T. R., Batista, R. A., Teixeira, M. C., Sá-Correia, I., & Duque, P. (2012). The Pht1;9 and Pht1; 8 transporters mediate inorganic phosphate acquisition by the Arabidopsis thaliana root during phosphorus starvation. The New Phytologist, 195, 356–371. https://doi.org/10.1111/j.1469-8137.2012.04167.x
Rennenberg, H., & Herschbach, C. (2013). Phosphorus nutrition of woody plants: many questions—few answers. Plant Biology, 15, 785–788. https://doi.org/10.1111/plb.12078
Reymond, M., Svistoonoff, S., Loudet, O., et al. (2006). Identification of QTL controlling root growth response to phosphate starvation in Arabidopsis thaliana. Plant, Cell & Environment, 29, 115–125.
Richardson, A. E., Hadobas, P. A., & Hayes, J. E. (2001). Extracellular secretion of aspergillus phytase from Arabidopsis roots enables plants to obtain phosphorus from phytate. The Plant Journal, 25, 641–649.
Robinson, W. D., Carson, I., Ying, S., et al. (2012). Eliminating the purple acid phosphatase AtPAP26 in Arabidopsis thaliana delays leaf senescence and impairs phosphorus remobilization. The New Phytologist, 196, 1024–1029.
Rubio, V., Linhares, F., Solano, R., et al. (2001). A conserved MYB transcription factor involved in phosphate starvation signaling both in vascular plants and in unicellular algae. Genes & Development, 15, 2122–2133.
Sambo, P., et al. (2019). Hydroponic solutions for soilless production systems: issues and opportunities in a smart agriculture perspective. Frontiers in Plant Science, 10, 923.
Secco, D., Baumann, A., & Poirier, Y. (2010). Characterization of the rice PHO1 gene family reveals a key role for OsPHO1; 2 in phosphate homeostasis and the evolution of a distinct clade in dicotyledons. Plant Physiology, 152, 1693–1704.
Shah, A. N., Wu, Y., Iqbal, J., Tanveer, M., Bashir, S., Rahman, S. U., Hafeez, A., Ali, S., Ma, X., Alotaibi, S. S., El-Shehawi, A., & Yang, G. (2021). Nitrogen and plant density effects on growth, yield performance of two different cotton cultivars from different origin. Journal of King Saud University-Science, 33(6), 101512. https://doi.org/10.1016/j.jksus.2021.101512
Sheerin, D. J., Buchanan, J., Kirk, C., et al. (2011). Inter- and intra-molecular interactions of Arabidopsis thaliana DELLA protein RGL1. Biochemical Journal, 435, 629–639.
Shen, J., et al. (2011). Phosphorus dynamics: from soil to plant. Plant Physiology, 156, 997–1005.
Stefanovic, A., Ribot, C., Rouached, H., Wang, Y., Chong, J., Belbahri, L., Delessert, S., & Poirier, Y. (2007). Members of the PHO1 gene family show limited functional redundancy in phosphate transfer to the shoot, and are regulated by phosphate deficiency via distinct pathways. The Plant Journal, 50, 982–994. https://doi.org/10.1111/j.1365-313X.2007.03108.x
Steinwand, M. A., & Ronald, P. C. (2020). Crop biotechnology and the future of food. Nature Food, 1, 273–283.
Suleman, M., Yasmin, S., Rasul, M., Yahya, M., Atta, B. M., & Mirza, M. S. (2018). Phosphate solubilizing bacteria with glucose dehydrogenase gene for phosphorus uptake and beneficial effects on wheat. PLoS One, 13(9), e0204408.
Thudi, M., et al. (2021). Novel genes and genetic loci associated with root morphological traits, phosphorus-acquisition efficiency and phosphorus-use efficiency in chickpea. Frontiers in Plant Science, 12, 10.
Tuskan, G. A., DiFazio, S., Jansson, S., et al. (2006). The genome of black cottonwood, Populus trichocarpa (Torr. & Gray). Science, 313, 1596–1604. https://doi.org/10.1126/science.1128691
Versaw, W. K., & Harrison, M. J. (2002). A chloroplast phosphate transporter, PHT2;1, influences allocation of phosphate within the plant and phosphate-starvation responses. Plant Cell, 14, 1751–1766. https://doi.org/10.1105/tpc.002220
Wang, X., Du, G., Meng, Y., et al. (2010). The function of LPR1 is controlled by an element in the promoter and is independent of SUMO E3 ligase SIZ1 in response to low pi stress in Arabidopsis thaliana. Plant & Cell Physiology, 51, 380–394.
Wang, L., Dong, J., Gao, Z., & Liu, D. (2012). The Arabidopsis gene hypersensitive to phosphate starvation 3 encodes ethylene overproduction 1. Plant & Cell Physiology, 53, 1093–1105.
Xu, C., Zhang, H., Sun, J., Guo, Z., Zou, C., Li, W. X., et al. (2018). Genome-wide association study dissects yield components associated with low-phosphorus stress tolerance in maize. Theoretical and Applied Genetics, 131(8), 1699–1714.
Yi, K., Wu, Z., Zhou, J., et al. (2005). OsPTF1, a novel transcription factor involved in tolerance to phosphate starvation in rice. Plant Physiology, 138, 2087–2096.
Zahra, Z., Arshad, M., Ali, M. A., Farooqi, M. Q. U., & Choi, H. K. (2020). Phosphorus phytoavailability upon nanoparticle application. In Sustainable agriculture reviews (Vol. 41, pp. 41–61). Springer.
Zhao, Y., Li, Y., & Yang, F. (2020). Critical review on soil phosphorus migration and transformation under freezing-thawing cycles and typical regulatory measurements. Science of the Total Environment, 751, 141614.
Zhou, J., Jiao, F., Wu, Z., et al. (2008). OsPHR2 is involved in phosphatestarvation signaling and excessive phosphate accumulation in shoots of plants. Plant Physiology, 146, 1673–1686.
Zhu, J., Kaeppler, S. M., & Lynch, J. P. (2005a). Mapping of QTL controlling root hair length in maize (Zea mays L.) under phosphorus deficiency. Plant and Soil, 270, 299–310.
Zhu, J., Kaeppler, S. M., & Lynch, J. P. (2005b). Mapping of QTLs for lateral root branching and length in maize (Zea mays L.) under differential phosphorus supply. Theoretical and Applied Genetics, 111, 688–695.
Zhu, J., Mickelson, S. M., Kaeppler, S. M., & Lynch, J. P. (2006). Detection of quantitative trait loci for seminal root traits in maize (Zea mays L.) seedlings grown under differential phosphorus levels. Theoretical and Applied Genetics, 113, 1–10.
Author information
Authors and Affiliations
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2023 The Author(s), under exclusive license to Springer Nature Switzerland AG
About this chapter

Cite this chapter
Shah, A.N., Abbas, A., Waqas, M.M., Nawaz, M., Ali, M., Fiaz, S. (2023). Genetic and Molecular Factors Modulating Phosphorus Use Efficiency in Plants. In: Prakash, C.S., Fiaz, S., Nadeem, M.A., Baloch, F.S., Qayyum, A. (eds) Sustainable Agriculture in the Era of the OMICs Revolution. Springer, Cham. https://doi.org/10.1007/978-3-031-15568-0_16
Download citation
DOI: https://doi.org/10.1007/978-3-031-15568-0_16
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-031-15567-3
Online ISBN: 978-3-031-15568-0
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)