
Abstract
Soluble sugars are metabolic substrates that play a fundamental role in controlling several processes in diverse plant developmental phases. Sugars such as glucose, sucrose and fructose are reported throughout the plant kingdom and have a dominant role in structure and metabolism at the cellular and whole-plant level. They also act as vital signals in the plant developmental regulation and are highly sensitive to environmental stresses. Abiotic stresses such as drought, heat, salinity, freezing or chilling stresses affect the movement of carbohydrates from source to sink tissues. Plants under abiotic stresses usually accumulate an excess of reactive oxygen species (ROS) which have damaging effects at tissue and cellular levels.
Soluble sugars acts as nutrient, osmoprotectant and metabolite signalling molecules and are involved in a variety of abiotic stress responses. Some soluble sugars have dual functions in gene regulation as shown by the upregulation of growth-related genes and downregulation of stress-related genes through hexokinase-dependent and/or hexokinase-independent cascades. This chapter aims at reviewing topical evidences on what method plants can sense and strategically respond to abiotic stresses via sugar-sensing mechanisms. The current role of antioxidants during abiotic stress, with a specific focus on the primary function of sugars as ROS scavengers, is also discussed. Yet, a complete biochemical characterization using advanced molecular biology methodology of the signalling pathways and molecules involved would be essential to regulate specificity and crosstalk in soluble sugar-regulated abiotic stress signalling cascades. This would facilitate the advancement of approaches for crop tolerance to abiotic stress conditions.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
14.1 Introduction
Soluble sugars (mono- and disaccharides) play a main role in the assembly and functioning of all living cells. Their origin gives the impression to be firmly connected with prebiotic and primary biotic evolution (Hazra et al. 2019). In plants, sugars are the metabolic substrates involved in actively modulating several metabolic processes at different stages of growth and development (Alagbe et al. 2020). Synthesis of sugar is the crucial process taking place in a plant through photosynthesis by carbon dioxide reduction while the solar energy is exploited for oxidation of water molecule (Taiz and Zeiger 2002). Glucose (Glc) has been reported to affect germination, floral transitioning, cell wall development and senescence (Dekkers et al. 2004). Glc promoted cell division, root architecture and elongation. It also controls the secondary metabolite synthesis (glucosinolates and phenolic compounds) (Wei et al. 2011). On the other hand, sucrose (Suc) assists in cell expansion and starch synthesis (Gibson 2005). The plant sugar status regulates the innate and ecological (external) factors that affect the overall progress (Sedigheh 2019). Sugar starvation in plants affects various pathways such as it increases the synthesis of α-amylase (Gill et al. 2003). On the contrary, it also leads to decline in respiration rate, protein synthesis, sugar metabolism and nitrate assimilation (Rolland and Sheen 2005). Also, the enzyme activity and synthesis increase for catabolic processes, viz. fatty acids, amino acids and proteins (Scott 2008).
The recent rise in global population, decrease in freshwater levels and continuous increase in cultivable land deterioration pose severe environmental stresses. Cumulatively, the aforementioned factors cause threats to worldwide agricultural production and food security (Dutta et al. 2018). Songstad et al. (2017) reported up to 70% reductions in global production due to abiotic stresses. As sessile organisms, plants have to tolerate drastic environmental changes as well as a diverse array of abiotic stressors (Singh et al. 2018). These abiotic stresses consist of temperature changes, ultraviolet-B radiation (UVBR), high light intensity, soil salinity, drought, flooding, chilling, freezing, heavy metals and nutrient deficiency or excess that discretely or combined negatively impacts both crop yield and quality (Khan et al. 2019). Abiotic stresses directly or indirectly impact plant growth, photosynthetic cycle, carbon partitioning, carbohydrate and lipid homeostasis, osmotic metabolism, protein synthesis and gene expression (Rosa et al. 2009). Therefore, plants have developed an extensive range of adaptative strategies to evade the impacts of environmental stresses. Plants in general are regularly subjected to a combination of diverse abiotic stresses (Gratão et al. 2005). Hence, plant responses to combined stresses are exclusive and cannot be applied independently to each stress. Plants’ responses to abiotic stresses are complicated phenomena as it can be synergistically or antagonistically adapted (Chaves et al. 2002).
The molecular mechanisms underlying these responses are barely identified. The lack of necessary progress is due to the fact that tolerance to abiotic stress is affected by coordinated and differential expression of gene networks. Plant strategies to deal with abiotic stresses typically involve either stress avoidance mechanism or tolerance mechanism. A common response to abiotic stresses is the accumulation of sugars and other compatible solutes (Sami et al. 2016). Soluble sugars also act as signalling molecules and help in regulating several processes of plant development (Loreti et al. 2001). The sugar signalling pathways may interact with stress pathways to form a complex network for regulating plant metabolic responses (Tran et al. 2007). Soluble sugars might act as negative modulators of plant sensitivity and cell responses to stress-induced distant signals. Numerous stress reactive genes have been described to be activated by Glc, thus signifying the role of soluble sugars in such responses (Price et al. 2004). The response of soluble sugars in plants to salt stress is complex. It is attained through a series of biochemical reactions and molecular mechanisms, ultimately leading to differential expression of stress-related genes (Espasandin et al. 2018; Lu et al. 2018). The differential regulation of 31 genes corresponded to carbohydrate content as well as metabolic enzymes in Arabidopsis under salt, cold and drought stresses (Loreti et al. 2005). Soluble sugars by acting as osmoprotectants can alleviate the adverse effects of salinity on plants. Rosa et al. (2009) reported a substantial rise in Glc, Suc and Fru levels in plants during salinity stress. The exogenous application of Glc can decrease Na+ accumulation and increase K+ uptake, thus continuing ion homeostasis under stress. Sugars played a dynamic role in carbon storage, osmotic defence and free radical scavenging (Nemati et al. 2011). Trehalose (T6P) is a non-reducing disaccharide of Glc, which plays an important physiological role as an abiotic stress protectant (Paul et al. 2018). By acting as an osmoprotectant, it can possibly stabilize biomolecules (dehydrated enzymes, proteins and lipid membranes) under abiotic stress (Denver and Ullah 2019).
Sugar sensing can be defined as the interaction amid a sugar moiety and a sensor protein in such a way that a signal can be produced, which in turn can stimulate signal transduction cascades resulting in various cellular responses. The sugar level in plant cells can be sensed by the sensor proteins. Hexokinase (HXK) enzyme is reported to have a functional role of a Glc sensor (Moore et al. 2003). Different Glc sensing processes are reported that regulate different genes and pathways in plants. These are the HXK-dependent and HXK-independent systems. In the former system, HXK is an active signalling mediator, whereas in the latter system, the receptor is unknown. There is also a metabolite-reliant system which is dependent on glycolysis pathway downstream of HXK (Rolland et al. 2006). As signalling molecules, sugar impacts the plants at all growth phases beginning from germination. As hormones, it can act as prime messenger and control signals, hence regulating gene expression.
Reactive oxygen species (ROS) are partially reduced forms of oxygen and act as both signalling and damaging entities in plants (Møller and Sweetlove 2010). ROS such as hydroxyl radical (•OH), superoxide (O2•-), hydrogen peroxide (H2O2) and singlet oxygen (1O2) are continually generated as by-product of aerobic metabolism in different plant organelles (viz. mitochondria, peroxisomes and chloroplasts) (Miller et al. 2010). Under abiotic stress, plants tend to accumulate higher ROS, and this might be due to a disruption of equilibrium between ROS generation and scavenging at different cellular locations (Bolouri-Moghaddam et al. 2010). ROS can act as signals triggering stress (by oxidizing proteins, lipids and nucleic acids) and defence pathways. In plants, ROS generation and photosynthetic mechanism are interlinked (Peshev and Van den Ende 2013). Stress-induced ROS accretion in plants is counteracted by two processes: firstly, inhibition of ROS development and, secondly, ROS scavenging by enzymatic or non-enzymatic antioxidants (Gill and Tuteja 2010). Soluble sugars appear to be involved in ROS-generating metabolic pathways. However, specifically, those sugars that interact with membranes can act as innate ROS scavengers (antioxidant) in plants (Peshev and Van den Ende 2013). They can correspondingly feed on NADPH-generating metabolic pathways such as the oxidative pentose phosphate (OPP) pathway and in so doing contribute to the ROS scavenging cascade (Khan et al. 2019).
The objective of this chapter is to review topical evidences on what method plants can sense and strategically respond to abiotic stresses via sugar-sensing mechanisms. The different functions of Glc, Suc and other sugars will be deliberated. We have also assessed the dual position of soluble sugars on the basis of our current knowledge on ROS production in plants. ROS evasion and scavenging will be discussed with respect to abiotic stress in plants with specific consideration on the role of sugars as antioxidants. Sugar responses to different abiotic stresses and biochemical and genetic evidences for diverse sugar-sensing mechanisms are analysed and summarized. Finally, this could provide a background for the developmental strategies for crop tolerance enhancement to abiotic stress conditions.
14.2 Physiological Role of Sugars in Plants
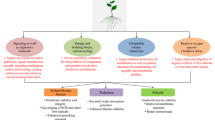
Soluble sugars guarantee an adequate flow of energy and building materials to carry out specific developmental activities in plants (Fig. 14.1). Glc (C6H12O6) is a simple sugar and the most abundant monosaccharide. Fru (fruit sugar) is a ketonic monosaccharide, where it is frequently bonded to a Glc molecule to form the disaccharide Suc in plants. Suc (C12H22O11) is a common sugar produced naturally in plants. Mannitol is a six-carbon acyclic sugar alcohol that can act as osmoprotectant to counter ROS and as a carbon storing compound. T6P is a naturally occurring non-reducing disaccharide. It has a crucial role in plant metabolic processes and acts as a reserve carbohydrate in plants (Stephanie et al. 2010). Sink formation, carbon metabolism and sugar build-up have significant functions in vegetative plant development, possibly through sugar signalling cascade.

A diagrammatic representation of the role of soluble sugar in plant growth and development. See text for details. SnRK1 Snf1-related kinase, T6P trehalose, ABA abscisic acid
14.2.1 Germination and Seedling Development
In the process of seed germination , the chief reserved compounds (carbohydrates, lipids and proteins) are mobilized by hydrolytic enzymes which in turn are used in seedling growth (Soltani et al. 2006). The utilization of these reserve substances continues till the seed turn into autotrophic seedling (Pritchard et al. 2002). The high activity of cell wall invertase creates high hexose concentration that helps embryo advancement by promoting rapid cell division. This has been observed by high-resolution histographical mapping in emerging cotyledons (Borisjuk et al. 1998). Exogenous treatment of Glc negatively affects germination in a concentration-dependent (1.5 < 2.5 < 5%) manner (Zhao et al. 2009). The Glc might function as a “morphogen” possibly due to sugar-mediated regulation of cyclin D gene (Riou-Khamlichi et al. 2000). In the transition phase, the embryo shifts from a predominant mitotic phase to differentiation determined by cellular enlargement due to Suc uptake surge and the creation of embryo sink strength. Elevated concentrations can lead to obstruction in germination as well as can inhibit the early stages of seedling growth (Price et al. 2003). However, the process is reversible, as the growth continues when the Glc concentration decreases (Gibson 2005). The effect is seen in Vigna radiata where with increase in Glc content, the germination rate and height of sprouts are decreased (Dewi 2015).
Arabidopsis seed growth shows a similar pattern, but the only difference is that its storage material is mainly lipids, and T6P metabolism also has a crucial role during embryogenesis. The inhibitory action of Glc at higher level on the development of seedling is also well studied. Arabidopsis seedlings at higher Glc concentration are incapable of developing green colour and also fail to develop expanded cotyledons or leaves (To et al. 2003). Researchers reported that a loss-of-function mutation in the putative APETALA2 (AP2)-type TF intensified the seed mass and yield. The increase in cell size and count in the mutant embryos is linked with the elevated hexose to Suc ratio during embryo development (Jofuku et al. 2005). Hypocotyl elongation is inversely proportional to the Glc concentration (Kushwah and Laxmi 2014). Glc intermediated regulation of seed germination might be an outcome of its capacity to mitigate the inhibitory impacts of abscisic acid (ABA) on germination in a concentration-dependent way. Additionally, few ABA and Glc signalling mutants demonstrate standard germination kinetics (Price et al. 2003), signifying the role of specific signalling pathways in germination and differential reaction to sugars conditional on the developmental phase. An essential role of ABA in plant sugar metabolism was validated by the description of Glc insensitive5 and Glc insensitive6/Suc uncoupling6/sugar insensitive5 as mutant alleles of ABA3 and the gene that codes the AP2-type TF ABI4, respectively (Leon and Sheen 2003). ABA response element (ABF2) is an essential component of sugar signalling, and overexpression of ABF3–4 also upsurges Glc sensitivity (Kim et al. 2004). Exogenous Glc explicitly improved the expression of ABA synthesis and signalling genes along with the endogenous levels (Cheng et al. 2002). Hence, it is proposed that Glc-specific build-up of ABA is essential for sugar signalling in early seedling growth .
14.2.2 Carbon and Nitrogen Metabolism
Glc is a minor photosynthetic product; however, its exogenous treatment can modify the photosynthetic cycle and its associated traits. In the presence of Glc, the rate of electron allocation to photosystem I (PSI) increases. In turn, Glc hydrolysis to NADH and NADPH (reducing powers) also increases which are vital for several biosynthetic reactions (Rineau et al. 2013). The rate of chlorophyll molecule synthesis also increases in the incidence of Glc. It was reported that excised, etiolated bean leaves showed enhanced chlorophyll synthesis on external application of Glc (Prasanna et al. 2004). The effect of Glc on the photosynthetic process is concentration dependent. With increasing concentration of Glc, the chloroplast development is suppressed and finally inhibited. This inhibition is due to Glc- and Suc-mediated repressed transcription of photosynthetic genes (Sinha and Roitsch 2002). The rate of oxygen evolution and electron transport also declines in Glc presence. RuBisCO is the vital enzyme in photosynthesis that increases with increasing Glc concentration (Spreitzer and Salvucci 2002). Carotenoids are photoprotective pigments that also act as precursors for ABA biosynthesis in plants. It is reported that carotenoid synthesis is decreased by the Glc treatment probably due to repression of genes coding for carotenoid pathway enzymes (Stephanie et al. 2010). β-Carotene is the amplest form of vitamin A present in fruits and vegetables. On the contrary, the concentration of β-carotene rises in plants after Glc application (Krinsky and Johnson 2005). Exogenous application of Glc can increase the expression level of glycolysis, pyruvate metabolism, Krebs cycle (energy-generating mechanisms) and respiration (energy-consuming mechanism). It is reported that the presence of Glc can increase respiration but decreases starch in potato (Stephanie et al. 2010). This response is similar to overexpression of invertase enzyme (Alagbe et al. 2020). Plant assimilates nitrates into other organic compounds, and their reduction into nitrite is catalysed by nitrate reductase. It is reported that Glc could affect enzyme nitrate reductase by enhancing the accumulation of nitrate in roots of corn (Oaks 1994). NRT2.1 is a nitrate transporter whose protein levels and transferring capacity are augmented in the presence of Glc. However, the augmentation is independent of HXK1-mediated stimulation (Jong et al. 2014).
14.2.3 Plant Growth and Development
Soluble sugars are capable of regulating overall plant growth and development. Genes that translate proteins like AGPase and Snf1-related kinases (SnRK) in apical meristem assist as markers for initial leaf expansion (Pien et al. 2001). Shoot maturation is separable into juvenile, adult nonreproductive and reproductive phases. By repression of the small subunit of RuBisCO, shoot morphogenesis is delayed, whereas the shoot to root ratio is increased (Tsai et al. 1997). Glc suppresses miR156 (transition of juvenile phase to adult phase) expression up to 50% (through HXK1-dependent pathway) due to a decrease in miR156 genes (Yang et al. 2013). Sugars are also reported to regulate the development of organs. Arabidopsis seedlings (dark-grown) showed adventitious root formation initiation by exogenous Suc application (Takahashi et al. 2003). Hence, it is established that Glc treatment may result in suspending the transition process in plants. In the root meristematic zone, Glc concentration increases the rate of meristematic cell differentiation along with its transport to the elongation and differentiation region. The effect is that the Glc concentration is inversely proportional to the root length (Lorenz et al. 2003). Additionally, initiation of flowering is allied with starch utilization and elevation in leaf carbohydrate transfer to the shoot apical meristem, signifying that sugars in the phloem are critical components. It is reported that Suc accessibility boosts morphogenesis and flowering in Arabidopsis even in dark conditions. Exogenous Suc (1%) treatment induces the late-flowering phenotype of different mutants, whereas at higher concentration of sugars, the transition is delayed by extension of vegetative phase (Roitsch and Gonzalez 2004). Arabidopsis sugar-insensitive mutant (glz2) showed late flowering, entirely sterile gynoecium and abnormal flowers and fruits (Chen et al. 2004). The essential function of precise carbon allocation in reproductive advancement arises from antisense SnRK transgenic barley and tobacco male sterility phenotypes with tissue explicit antisense repression of a cell wall invertase (Ruuska et al. 2002). An associated delay in stimulation of LEAFY advocates that sugars can regulate the expression of floral meristem identity genes (Ohto et al. 2005).
14.2.4 Senescence
Leaf senescence is the last step in the plant’s life cycle in which the metabolites are remobilized and transported into the developing organs from old leaves (Fig. 14.1). Senescence can be stimulated by different conditions, and numerous signalling pathways are intricate in the regulation. Possibly, it is the result of integration of environmental and sugar signalling. Leaf senescence was induced in Arabidopsis by Glc application (2%) in combination with low nitrogen supply (Wang et al. 2008). Amino acid amount decreased in senescing leaves, but the sugar content was increased in numerous plant species (Jongebloed et al. 2004). In Arabidopsis, the contents of Glc and Fru (hexoses) upsurge, whereas Suc content remains stable in senescing leaves (Wingler 2007). Data analysis of Affymetrix GeneChip assay confirmed that variations in gene expression during Glc-stimulated senescence are representative of developmental senescence. Sugars can be supposedly synthesized by gluconeogenesis from fatty acids. Senescence during drought stress is beneficial as it not only reutilizes nutrients but also averts excessive water loss, thus improving plant water balance (Lim et al. 2007). On the contrary, in another study, researchers reported that delayed ageing can enhance drought tolerance in tobacco plants (Rivero et al. 2007). Stessman et al. (2002) reported hexose accumulation in senescing leaves of Arabidopsis. Glc, Fru, Suc and other osmolytes are reported to get accumulated during cold treatment (Kaplan et al. 2007). In another report, Jongebloed et al. (2004) displayed that phloem obstruction due to callose deposition may perhaps result in an age-dependent sugar accumulation. The ABA is an imperative signal in plant retort to stresses like cold, drought and salinity. The phytohormone synthesis and signalling has identified a vital component in sugar signalling. Arabidopsis mutants in ABA synthesis or signalling did not show developmental seizure in response to high-sugar levels (Rolland et al. 2006). A few cold response genes can be induced by ABA and sugar signalling; therefore, a feedforward loop might occur enhancing stress acclimation by sugar aggregation (Masclaux-Daubresse et al. 2007). Furthermore, exogenous Glc application did not induce senescence in cold-grown Arabidopsis as in higher temperatures. This controlling interaction might be significant in the prevention of premature senescence .
14.3 Sugar as a Signalling Molecule: Signal Transduction Cascades
In order to coordinate the metabolic processes in plants and respond appropriately to the altering environmental changes and energy demands, plants have developed an array of mechanisms to sense diverse sugar signals (Gupta and Kaur 2005). The signal then begins the signalling transduction cascade that leads to different cellular responses. Plant HXK (HXK) has been proposed as a dual-functioning enzyme with both regulatory and catalytic functions (Xiao et al. 2000). Plants have two different HXK-Glc sensing and signalling pathways, which catalyse via the phosphorylation of Glc, Fru and other molecules (Wei et al. 2020). First is the HXK-dependent pathway and the second is the HXK-independent pathway. The HXK-dependent signalling pathway requires phosphorylation of sugars, whereas the HXK-independent signalling pathway does not require phosphorylation. Sugar analogues that are phosphorylated by HXK were able to activate downregulation of photosynthetic genes. Additional breakdown of sugar phosphates was not important to cause suppression, as 2-deoxyglucose (2-DG) and mannose cannot undergo further processing after phosphorylation and this might likewise cause downregulation. Moreover, mannoheptulose, a competitive inhibitor of HXK , can inhibit the suppression caused by 2-DG (Gupta and Kaur 2005). Moreover, 3-oxy-methyl-glucose (3-o-MG) cannot diminish the suppression by 2-DG since it cannot also be phosphorylated by HXK (Gupta and Kaur 2005). The epimer of Glc is mannose, which is phosphorylated by HXK to mannose-6-phosphate, which only gradually enters glycolytic metabolism (Smeekens 2000). Arabidopsis plant has Glc analogue such as 2-DG and mannose which inhibits seed germination through HXK signalling cascade. HXK-independent substrates such as 3-o-MG and 6-deoxyglucose have no effects on seed germination, whereas the HXK inhibitor mannoheptulose impedes the mannose-induced inhibition of seed germination.
In an experiment, the role of Arabidopsis HXK (AtHXK1 and AtHXK2) in the sugar-induced repression of photorespiration (gene) and early seedling development is shown (Xiao et al. 2000). The metabolic enzyme (AtHXK1) acts as a Glc sensor by modifying the HXK levels or with a heterologous yeast, viz. YHXK that give a primary catalytic function. This concept is supported by a five times increase in hexose phosphorylation activity, but no inhibition of seedling development in plants ectopically expressing YHXK2. Plants overexpressing AtHXK1 show both an increased hexose phosphorylation activity and an inhibition of germination and early seedling development. It was reported that AtHXK1 had an important signalling function which was not altered by YHXK2 (Jang et al. 1997). Some genes, involved in carbon and nitrogen metabolism, were reported to be independent of HXK (Sheen et al. 1999).
The standard definition of HXK is that it acts as a dimeric cytosolic enzyme crucial for glycolysis. HXK is a Glc sensor, where intracellular sugar (Glc) signals are sensed, or it has regulatory and catalytic functions or whether the ATP/AMP ratio is the actual signal (Bolouri-Moghaddam et al. 2010). HXK activity could be involved in cell expansion, cell division and ROS signalling in plant organelles such as mitochondria and chloroplasts (Alagbe et al. 2020). Mitochondrial HXK (mtHXK) activity has an important role in producing ADP to maintain oxidative phosphorylation, thereby controlling ATP generation-associated limitation of oxidative respiration and successive H2O2 generation. In maize roots, mtHXK is supposed to be connected to sugar synthesis that is needed for phenylpropanoid, flavonoid and cellulose biosynthesis (Bolouri-Moghaddam et al. 2010). The mtHXK enzyme in plant performs an antioxidant action in mitochondria. Glc signalling through HXK downregulates a branch of the ethylene-signalling pathways that induce germination and cotyledon and leaf development (Smeekens 2000). HXK and SnRK are already regarded as conserved sugar signalling components regulating energy homeostasis, survival, abiotic stress resistance and longevity. Additionally, invertase-related sugar signals appear to be very important during plant defence reactions (Bolouri-Moghaddam et al. 2010). A significant role for HXK-dependent sugar signalling in leaf senescence has also been reported (Bolouri-Moghaddam et al. 2010). Figure 14.2 shows a diagrammatic depiction of sugar-sensing mechanisms in plants.

A diagrammatic representation of sugar-sensing mechanisms in plants. Sucrose and glucose are sensed by transporter homologues at the plasma membrane. The HXK glucose sensor is linked with the outer membrane of plastids (mitochondria and chloroplast) and cytoplasm. HXK is also found in the nucleus with high-molecular-weight complexes; here it regulates transcription and proteasome-mediated breakdown of the EIN3 transcription factor. G-protein-coupled receptor signalling by RGS1 and GCR1 is intricate in glucose-mediated plant development, but by the hexokinase-independent pathway. SnRK1 protein in plant has an imperative function in sugar and starvation cascade; however, its regulation by sugars is still uncertain. Downstream of SnRK1, significant regulatory effects for T6P (trehalose) are reported. Lastly, several transcription factors controlling the sugar-regulated transcription are located in the nucleus. See text for additional details
14.4 Abiotic Stresses
Since the last few decades, the existence of plants under abiotic stress is a wide area of research. These abiotic stresses directly or indirectly distress the concentration, metabolism, transport and storage of sugars as well as the sugar signalling pathways (Cramer et al. 2011). Abiotic stress in plants can also reduce the growth and yield below the optimum levels. Sugars are firstly involved in several metabolic activities; it controls numerous genes specially those of Suc metabolism and photosynthesis. Secondly, accumulation of soluble sugars enhances plant survival under abiotic stresses, such as salt, heat, chilling and drought (Mohammadkhani and Heidari 2008). Other forms of carbohydrate such as raffinose family oligosaccharides (RFOs), T6P and fructans also function as protectants against different types of abiotic stresses. A set of genes involved in sugar metabolism influenced under glucose concentration and abiotic stresses in plants are shown in Table 14.1. The role of sugars under diverse abiotic stresses has been described further in details.
14.4.1 Water Stress
Drought is a major limiting factor in plant development and productivity. In a study conducted with a variety of plants, it was validated that drought or flooding induced conversion of hexoses, for instance, Suc and starch into polyols (sugar alcohols) (Mohammadkhani and Heidari 2008). Former reports stated that during drought stress, sugars protect the cells by two mechanisms. Primarily, the -OH groups of sugars may replace water (H2O) to sustain hydrophilic interactions in proteins and membranes during dehydration. Thus, sugars make hydrogen bonding with proteins and membranes and inhibit protein denaturation. Secondly, sugars are an extensive contributing factor to vitrification, i.e. the synthesis of a biological glass in the cytoplasm of dehydrated cells. In numerous plant species, it was reported that raffinose (Raf), verbascose and stachyose accumulated at the time of desiccation of seeds as well as accumulated in the leaves when the plant was exposed to abiotic stress (Sami et al. 2016). These sugars have a significant function during stress in membrane protection and ROS scavenging. Soluble sugars also prevent dehydration of membranes and maintain the turgidity of leaves. Drought is amongst the major factors that significantly downregulate photosynthesis (Sami et al. 2016). Glc induces stomatal closure and enhances plant’s adaptability under drought stress (Osakabe et al. 2014). Another significant component of carbohydrate metabolism is sucrose-phosphate synthase (SPS) present in plant leaves . Gupta and Kaur (2005) reported that SPS activity was decreased in leaves of plants subjected to drought stress. During meiosis water stress induced male sterility and prevent starch accumulation but enhanced the accumulation of soluble sugars (Pressman et al. 2002).
14.4.2 Salt Stress
Salt stress not only modifies nitrogen assimilation, carbohydrate levels, cell division and plant growth but also modifies the types of carbohydrates synthesized and transferred by the source tissues (Sami et al. 2016). Osmotic stress has an effect on the carbohydrate source (photosynthesis) as well as the mobilization of carbohydrate stock (Henry et al. 2015). Increased concentration of salt leads to ionic imbalance, declines stomatal conductance and causes reduction of leaf growth (Sami et al. 2016). Salinity induces alterations in processes such as energy metabolism, protein synthesis, ion transport and protein fate (Cramer et al. 2011). Osmotic imbalance subsequent from salinized soils or drought can be calamitous for plants like maize, chiefly due to its effect on early kernel development (Henry et al. 2015). Osmotic stress induces kernel abortion by diminishing photosynthesis and evapotranspiration in maize. Osmotic stress also reduces Suc-sugar levels, transcription levels, Suc-degrading enzyme activity and starch in the kernels. The accumulation of sugar serves as carbon storage and osmoprotectant, maintains osmotic homeostasis as well as scavenges free radicals to mitigate the damaging effects of salt stress (Sami et al. 2016). Plants try to reduce their osmotic potential by enhancing mineral ion content and synthesis of compatible solutes to enhance water uptake under stress (Nemati et al. 2011). The synthesized and accumulated total soluble carbohydrates in the cytosol are also important solutes during salinity stress. Total soluble carbohydrates induce better osmotic adjustment and maintain turgor for growth under salinity. Plants and bacteria are reported to produce fructans (polyfructose molecules) that have a crucial part in adaptation to osmotic tension due to their high soluble characteristic.
14.4.3 Cold Stress
Cold stress causes substantial dysfunctions at the cellular level; these comprise disruption of membranes, generation of ROS, protein denaturation and accretion of toxic by-products (Yuanyuan et al. 2009). Development of cold tolerance in higher plants enhanced by the accumulation of a diverse variety of soluble sugars such as Glc, Fru, Suc, stachyose and Raf (Sami et al. 2016). It has been reported that these sugars function as osmoprotectants and also protect the cellular membrane from damage, induced by dehydration and freezing (abiotic stress) through interaction with the lipid bilayer. In addition, T6P sugar also serves as osmolyte and stabilizes protein, lipid and membrane. In rice, higher concentration of T6P is detected under cold stress. Some oligosaccharides like fructans and Suc induce resistance from crystallization and then enable glass formation within the cell and protection of membrane phospholipids under freezing. In other words, the accumulation of soluble sugars contributes to the enhancement of cryo-stability of cell membranes. This enhanced membrane stability is a prerequisite for dehydration or cold tolerance because freezing causes destabilization of cellular membranes which is the initial cause of damage in plants. Polyols like glycerol, sorbitol, Suc and mannitol are osmoprotectants in insects , exposed to freezing algae and certain halophytic plants (Rathinasabapathi 2000).
14.4.4 Heat Stress
In heat stress , the accumulation of soluble sugar and starch in the pollen grains was markedly lesser than that under normal conditions (Pressman et al. 2002). Abiotic factors such as high temperature and water stress affect SPS activity (Lafta and Lorenzen 1995). The enhancement in SPS activity is transferred to the entire plant level from source leaves at high temperature (heat stress). In this experiment, the changes in SPS activity at high temperature coincided with altered enzyme concentrations of carbohydrate metabolism in developing tissues. It was also associated with increased carbohydrate partitioning in young leaves and stems. High temperature reduced the plant growth and leaf expansion. Heat stress affects more the tuber growth than the shoot growth (Feil and Lunn 2018). High-temperature-treated leaves reported higher levels of Suc at the edge of the photoperiod as compared to the leaves from the cold treatment (low temperature), whereas starch concentration was reported to decline in the plant leaves after high-temperature treatment (Feil and Lunn 2018). A paper proposes that heat stress-induced depletion in sugar transport to reproductive tissue leads to loss of gametophyte growth (Pressman et al. 2002).
14.5 Abiotic Stress-Induced Oxidative Stress and Scavenging in Plants
Abiotic stress at cellular and tissue levels commonly induces an oxidative stress. It is defined as a disproportion between ROS synthesis and scavenging via antioxidants like Glc (Keunen et al. 2013). Under physiological conditions, ROS such as 1O2, O2•-, •OH and H2O2 are generated as by-products of oxidative metabolism in different plant organelles. ROS plays a dual role as signalling and damaging compounds (Sami et al. 2016). Increased ROS accumulation under abiotic stress is life-threatening as it leads to oxidization of proteins, lipids and nucleic acids (Hu et al. 2012). In addition, it affects the gene expression, plant cell growth, cell membrane integrity, hormone signalling and programmed cell death (Fraire-Velázquez and Balderas-Hernández 2013). Salt stress affects the plant’s metabolism in several ways such as ion toxicity, production of ROS and osmotic stress (Hu et al. 2012). ROS are highly reactive and can seriously impair normal metabolism causing photooxidative damage and leading to lipid peroxidation. Drought stress leads to an induction of high accumulation of ROS, which damages the biomolecules. Researchers stated that the shortage of water in cucumber cells caused lipoxygenase activity, membrane injury and lipid peroxidation affecting normal growth (Morsy et al. 2007; Hu et al. 2012). Soluble sugars are already testified to improve the antioxidative mechanisms as it is involved in ROS scavenging, thus contributing to an overall environmental stress tolerance (Keunen et al. 2013). In another way, soluble sugars can also funnel NADPH-producing metabolic pathways (i.e. OPP pathway), which can scavenge the ROS. Moreover, sugars and sugar alcohols have been widely accepted as an antioxidant system quenching ROS against various kinds of stresses, regulating the osmotic alteration and providing membrane protection (Sami et al. 2016; Hu et al. 2012).
14.5.1 ROS Scavenging in Plant Chloroplast
Under standard as well as stressed conditions, ROS is generated as the by-product of high photophosphorylation (oxidizing metabolism) or rapid electron flow in organelles such as chloroplasts and peroxisomes (Keunen et al. 2013). In the thylakoid membrane, photosystem I and II reaction centres account for high levels of ROS in the chloroplast. Soluble sugars can be involved in ROS producing photorespiratory pathway which is a major producer of H2O2. Environmental stresses, for instance, salinity, excess light, drought and metal exposure, enhance chloroplast ROS production. It can also utilize the OPP pathway producing reduced power for glutathione (GSH) production and contribute to ROS quenching (Cramer et al. 2011). Glucose-6-phosphate dehydrogenase (G6PDH) enzyme catalyses the first reaction in this pathway. This enzyme activity can be vital in regulating the ROS decontamination capacity along with redox potential in the chloroplast. Moreover, limited sugar supply may interrupt respiratory metabolism, thereby increasing ROS creation at the level of the electron transport chain (ETC). Sugars such as galactinol (Gol), RFOs, fructans and sugar alcohols (inositol, mannitol, sorbitol) are suggested to function as antioxidants and possess ROS quenching capacities (Keunen et al. 2013). Commonly, they have higher scavenging affinity for •OH as compared to 1O2. Nishizawa et al. (2008) analysed that Gol and Raf had •OH scavenging ability and showed similar antioxidant capabilities to GSH. Moreover, the researchers also reported that their concentrations are appropriately ranged to protect plant cells from oxidative damage. Interestingly, Raf concentration can directly scavenge •OH radicals as compared to ascorbate (AsA) and GSH in chloroplasts of stressed plants. In chloroplast stroma, ascorbate-glutathione (AsA-GSH) cycle is present that is fully functional to neutralize H2O2 produced by the difference of 1O2 catalysed by superoxide dismutase (SOD) (detoxifying ROS).
Some water-soluble antioxidants such as carotenoids and tocopherols are highly abundant in chloroplasts. Tocopherols efficiently reduce 1O2 and are located in the thylakoid membranes. They are also reported to scavenge lipid peroxides precisely under abiotic stress conditions (Keunen et al. 2013). On the other hand, carotenoid pigments protect the photosynthetic apparatus from photo-oxidation. Raf was shown to protect chloroplast from photorespiration and chloroplast membranes against desiccation, high temperature and freezing stress (Keunen et al. 2013). The oxidized RFO radicals might be regenerated by reducing antioxidants such as AsA or flavonoids. The overexpression of galactinol and raffinose synthase in transgenic Arabidopsis plants enhanced the Gol and Raf concentrations and showed effective oxidative stress tolerance and ROS scavenging capacity (Keunen et al. 2013). Peroxiredoxins (Prxs) are thiol-based peroxide reductases that are capable of reducing several substrates specifically ranging from peroxynitrite to H2O2 (Van den Ende and Valluru 2009). Fructans can protect plants against water stress by stabilizing and activating other specific antioxidative defence mechanisms. Additionally, sugar availability can increase AsA biosynthesis, possibly due to an enhanced respiration rate (Bolouri-Moghaddam et al. 2010). In tobacco, mannitol is reported to protect ferredoxin, GSH, thioredoxin and the thiol-regulated enzymes against •OH (Keunen et al. 2013).
14.5.2 ROS Scavenging in Plant Mitochondria
In non-photosynthetic cells , mitochondria are the core sites of ROS generation. This might be due to electron outflow at complexes I and III in the ETC (Keunen et al. 2013). Proactive energy-dissipating systems are present in plant mitochondria, which are able to considerably reduce the rates of ROS generation at the ETC (Hussain et al. 2019). In this situation, the function of the alternative oxidase (AOX) bypassing ETC III and IV complexes is confirmed to reduce ROS creation (Møller 2001). Plant uncoupling proteins are another proactive energy-dissipating structure that reduces the ROS production through fine-tuning the membrane potential of plant mitochondria (Keunen et al. 2013). Soluble sugars showing protective effects against oxidative or abiotic stress have been mostly owed to signalling cross-talks that could stimulate explicit ROS scavenger (Singh et al. 2019). However, sugars, AOX, uncoupling proteins and their allied metabolic enzymes may perhaps also act as ROS scavengers at the level of the mitochondrial ETC. However, it was recently proposed that the synergistic interaction of sugars and phenolic compounds is part of an integrated redox scheme, contributing to ROS scavenging and stress tolerance. The mitochondria-associated HXK (mtHXK) have catalytic activity which regulates detoxification of ROS and consecutive signalling pathways, thus generating antioxidant defence systems in plants (Keunen et al. 2013). The inner mitochondrial membrane contains galactono-glactone dehydrogenase enzyme which catalyses AsA synthesis. In the biosynthesis of AsA, SOD and sucrosyl oligosaccharides (SOS) act as antioxidants and detoxify ROS. In plant mitochondria, GSH levels could also cause cell survival during environmental stress, as GSH protects mitochondrial DNA and proteins from being oxidized. The enzyme mtHXK can play an important role in producing ADP to support oxidative phosphorylation (Bolouri-Moghaddam et al. 2010).
14.5.3 ROS Scavenging in Plant Peroxisomes
Similar to other plant organelles described earlier, ROS is also generated in peroxisomes as a by-product of its oxidative metabolism (Keunen et al. 2013). In the peroxisomal matrix, O2•- is produced by xanthine oxidase enzyme, and in membranes, it is produced via NADPH-dependent ETC reactions. In peroxisome, APX and MDAR are located on the membrane facing the cytosol. Peroxisomes are subcellular single-membrane organelles where H2O2 is generated by the activities of flavin oxidases and catalase (CAT) enzymatic components (Keunen et al. 2013). Peroxisomal ROS are also produced by various factors such as metals, xenobiotics, soil salinity and abiotic stresses. The presence of CAT in peroxisomes has been unequivocally recognized. The CAT enzyme dismutases H2O2 into O2 and H2O however, no external sources of reducing equivalents are needed for the reaction catalyzed by this enzyme. Three isoforms of enzyme i.e. CAT1–3 are reported to be present in the peroxisomal matrix of Arabidopsis. Correspondingly, the presence of Prx in plant peroxisomes could furthermore enhance the H2O2 quenching system in peroxisomes (Corpas et al. 2003). Experimental information suggests that only APX3 could have a significant role in the peroxisomal ASC-GSH cycle. Numerous components of the AsA-GSH cycle have already been reported to be present in the peroxisomal membrane and matrix. The CAT enzyme was found to coordinate with the AsA-GSH cycle . Until now, there are no reports that could clearly indicate the involvement of soluble sugars such as Glc and Suc in peroxisomal antioxidant defence.
14.5.4 Emerging ROS Scavenging Systems in Plant Vacuoles
Abiotic stresses can cause an enhancement of cytosolic H2O2 concentration that can enter the plant vacuole via aquaporin or diffusion throughout the tonoplastic membrane (Singh et al. 2017). Class III peroxidases are reported to be associated closely with the inner membrane of tonoplast. Lipid peroxidases can attack H2O2 and convert it into •OH radicals via the hydrolytic cycle, under stress conditions. Vacuolar sugars or sugar-like compounds can deeply insert itself amid the hydrophilic headgroups of the tonoplast membranes, thus stabilizing them under abiotic stress and playing a crucial role in scavenging these radicles. NADPH oxidases are enzymes present at the tonoplastic membrane which might be the major producer of ROS (cytosolic NADPH), as reported by some proteomic studies (Keunen et al. 2013). The initial product of this enzyme is 1O2, which is detected in the tonoplast. This 1O2 could be converted to the less toxic H2O spontaneously or by the activity of tonoplast-associated SOD (Van den Ende and Valluru 2009). Vacuoles accumulate a complex of anthocyanins, phenolics and malate (strong antioxidant compounds), which impacts the potential to buffer the redox state inside the cells (Keunen et al. 2013). Flavonoids such as anthocyanins and flavonols concentrate in the plant vacuoles and play versatile roles in plant metabolism. Phenolic compounds and fructans might function in a synergistic way to detoxify the excess vacuolar H2O2 and redox regulation processes. Vacuolar sugars and sugar alcohols are crucial players in antioxidative stress defence (Stoyanova et al. 2011). In addition, fructans were suggested to be acting directly as ROS quencher in the vicinity of the tonoplastic membrane (Singh et al. 2015). GSH, RFOs, fructans and SOS are supposed to protect biological membranes by detoxifying ROS in vacuoles and chloroplast or indirectly activating the antioxidative defence system (Van den Ende and Valluru 2009).
14.6 Possible Mechanisms of Sugar-Mediated Abiotic Stress Tolerance in Plants
Carbohydrates synthesized by photosynthesis are the building block and energy generator that supports plant growth. Under abiotic stress conditions, major plant growth is prevented, but photosynthesis and accumulation of fructans, disaccharides (Suc, T6P) and RFOs are slightly permitted. RFOs such as Raf, verbascose and stachyose are produced in the cytosol (Keunen et al. 2013). In addition to RFO accumulation, the activity of enzymes and gene expression involved in the metabolism are strongly connected with abiotic stress responses (Afzal et al. 2019). Under stress conditions, fructans can interact with apoplast cell membranes by directly forming hydrogen bonds (Livingston et al. 2009). RFOs and fructans are sucrosyl oligosaccharides, and their associated enzymes together interact with ROS scavenging cascades (Keunen et al. 2013). Carbohydrates and carbohydrate-containing biomolecules (soluble sugars) play a leading role in the cellular redox equilibrium due to their proximate associations with mitochondrial respiration, photosynthesis and fatty acid β-oxidation (Keunen et al. 2013). Small water-soluble sugars and the enzymes linked with their synthesis and signalling pathways are generally believed to be allied with oxidative stress and are capable of scavenging ROS. Any imbalance in the redox equilibrium of plant cells requires induction of specific antioxidant or antioxidant enzymes which represses ROS. The possible role and mechanism of sugar-mediated abiotic stress tolerance in plants have been described further in details (Table 14.2).
14.6.1 The Defending Role of Sugar and its Associated Metabolic Enzymes Under Abiotic Stress
Fructans are Suc-derived Fru polymers supposed to be produced in the plant vacuoles. The presence of fructans is reported in only about ≤15% of all angiosperms. Fructans such as inulin show a stronger interaction with apoplast or tonoplast membranes as compared to levan-type fructans. Fructans can inhibit lipid condensation and cessation of the transition phase by decreasing the molecular motions of the lipid headgroups. RFOs show analogous physiological characteristics in plants and were reported to be involved in dehydration and desiccation stress tolerance in seeds. Thus, both RFOs and fructans are supposed to protect cellular membranes under stress maintaining cellular integrity (Keunen et al. 2013). Some soluble carbohydrates function as osmoprotectant carbohydrates (e.g. Suc, T6P, fructans, Raf and Gol) (Van den Ende and Peshev 2013). Suc is supposed to function as an osmoprotectant, stabilizing/protecting biological membranes and maintaining cell turgor pressure (Morsy et al. 2007). T6P can function also as an osmolyte and keep stabilizing membranes and proteins (Hernandez-Marin and Martínez 2012).
Soluble sugar may contribute to the enhancement in cryo-stability of cellular membranes by maintaining membranes in their appropriate phase which is essential for survival under abiotic stress (Krasensky and Jonak 2012). Sugars keep membrane surfaces “hydrated” and inhibit membrane fusion by inserting into the space between phospholipid biomolecules (Van den Ende and Peshev 2013). Hence, soluble sugars can function as cryoprotectant for protecting the membrane in the dry state or dehydrated state. In the glass forming process, T6P not only crystallizes but also undergoes vitrification. It is the only sugar that can remain in glasslike state during dehydration stress (Fernandez et al. 2010). Raf may be more effective in membrane stabilization than disaccharide or monosaccharide (Morsy et al. 2007).
ROS accumulation has been shown to be inhibited by dysfunction of G6PDH, suggesting that the protective role of sugars was dependent on OPP activity. Soluble sugars can replenish NADPH, required for monodehydroascorbate reductase and glutathione reductase activities (Singh et al. 2016). Keunen et al. (2013) reported the roles of Gol and RFOs in ROS homeostasis that have a potential function as neutral invertase and HXK functioning in chloroplasts to prevent ROS build-up in stressed plants. Oligogalacturonides were reported to stimulate the metabolic and enzymatic antioxidative defence system in alfalfa roots, which can be potentially connected with sugar signalling in plant defence against oxidative stress (Keunen et al. 2013). In addition, sugars can stimulate the synthesis of auxins that places sugars at a pivotal position in the regulatory processes driving plant development, under normal as well as under environmental stress conditions (Van den Ende and Peshev 2013).
14.6.2 Sugars as Antioxidants in Plants
Small water-soluble sugars such as Glc, Suc, sugar alcohols and Gol are proven to function as antioxidants in plants (Keunen et al. 2013). Bolouri-Moghaddam et al. (2010) hypothesized that sugars or sugar-like complexes such as Suc (present in the vacuoles of sugar beet and sugar cane plants) might function as an antioxidative compound inhibiting lipid peroxidation. This might be due to high concentration of H2O2 produced under stress (Van den Ende and Peshev 2013) or by directly quenching the •OH and •OOH radicals that are formed around the membranes (Hernandez-Marin and Martínez 2012). Mannitol could not reduce •OH radical generation in the chloroplast, but it could enhance the scavenging ability, thus defending the cells against oxidative damage. In addition, mannitol accumulation had no toxic effects on plants (Van den Ende and Peshev 2013).
In an experiment, it was reported that in Arabidopsis plants, oligosaccharides’ concentration such as Gol and Raf was increased in stressed conditions (Krasensky and Jonak 2012). Accordingly, α-galactosidase is reported to be inhibited in Petunia plant, causing increased Raf synthesis and freezing tolerance (Pennycooke et al. 2003). The Arabidopsis plant contains seven Gol (GolS) genes, one of which is induced by cold (GolS3) and the other two by drought (GolS1 and GolS2). In addition, overexpression of drought caused GolS1 and GolS2 genes in the plant to enhance Gol and Raf levels, as well as showed decreased transpiration rates to improve drought stress tolerance. These plants exhibit more effective oxidative stress tolerance and ROS scavenging capacity than wild-type plants (Van den Ende and Peshev 2013). Nishizawa et al. (2008) evaluated the •OH scavenging ability of oligosaccharides (Raf and Gol) in vitro and indicated similar antioxidant scavenging abilities for both metabolites as compared to GSH. Interestingly, Raf levels in stressed plants are comparable with AsA and GSH, signifying that this water-soluble sugar can unequivocally neutralize •OH radicals in chloroplasts (Moller et al. 2008).
Production of •OH radical is highly dangerous in the vicinity of vacuolar membrane (Stoyanova et al. 2011). Thus, the presence of sugar metabolites can intensively interact with vacuolar membranes, such as fructans, and this could be critical for stabilization of membrane under stress (freezing tolerance) (Van den Ende and Peshev 2013). The accumulation of water-soluble sugar can be considered as true antioxidant capable of quenching ROS. It is found that inulin and stevioside are superior scavengers of both O2•- and •OH and are more effective than Suc and mannitol (Stoyanova et al. 2011). T6P also plays a role in ROS scavenging in vitro (Van den Ende and Peshev 2013). Krasensky and Jonak (2012) analysed some genetically engineered plants and reported that increased T6P metabolism can positively influence stress tolerance, but the exact role of T6P during abiotic stress remains unclear. Suc-treated plantlets were able to sense altering ROS levels and in response could trigger efficient scavenging and antioxidant systems (Ramel et al. 2009).
14.6.3 Effect of Sugars on Proline Accumulation
Under abiotic stress, amino acid metabolism plays an essential role in stimulation of stress tolerance in plants. As a result, different amino acids (proline (Pro), nonprotein amino acids, minor amino acids and branched-chain amino acids) tend to accumulate in plants (Kaur and Asthir 2015). Pro is a proteinogenic amino acid reported to function both in plant metabolism (as molecular chaperone) and defence (osmoprotectant) (Woodrow et al. 2017). In plants, Pro synthesis takes place by using two pathways, primarily glutamate pathway and secondarily ornithine pathway (activated in chloroplasts) (Suprasanna et al. 2016). Synthesis of Pro in the cytoplasm or chloroplast during unfavourable conditions stabilizes the low NADPH to NADP+ ratio, normalizes the redox balance by maintaining the electron flow between photosynthetic excitation centres and overall protects the photosynthetic apparatus from photoinhibition and damage (Dar et al. 2016). Accumulation of Pro has been described in various studies in response to different types of abiotic stresses, viz. salinity, drought, high temperature and metal toxicity (Vives-Peris et al. 2017). Surekha et al. (2014) reported a four times rise in Pro accretion in Cajanus cajan (transformed with V. aconitifolia P5CSF129A) as a response to salinity stress as compared to the wild type. In another report, the transformation of P5CS gene from V. aconitifolia in transgenic rice and chickpea plants showed a higher increase in Pro content (up to five times) as compared to non-transformed ones (Karthikeyan et al. 2011). In a topical study, Li et al. (2019) reported a transgenic Arabidopsis expressing salt-tolerant IbRAP2–12 gene from sweet potato. The plant was found to accumulate higher Pro content with lesser ROS generation under abiotic stresses as compared to the wild counterpart. It is still not clear whether accumulation of Pro is an adaptive strategy, a stress response or just an indication. Pro is reported to stabilize membranes and proteins, scavenge ROS, act as an antioxidant and induce expression of salt stress-responsive genes (Carillo 2018). Pro metabolism in the mitochondria under unfavourable conditions can contribute to additional oxidative respiration that may produce energy for continuing plant development (Kaur and Asthir 2015). It also functions as a sugar remobilization signal for restabilizing plant growth, thus causing beneficial effects on the growth (Woodrow et al. 2017). Nevertheless, there are species explicit variances in Pro accumulation strategies and sugar signalling crosstalk in plants under abiotic stress (Dar et al. 2016).
14.6.4 Sugar and Phytohormone Interaction
Sugars can likewise crosstalk with prevailing phytohormone signalling systems to control significant developmental processes in plants like germination and seedling and tuber growth (Zouari et al. 2016). Plants malfunctioned in ABA or ethylene signalling and sensitivity show altered sugar retort phenotypes. Hence, an enormous overlay between sugar, ethylene, ABA signalling and regulating metabolic processes has been reported (Gibson 2005). ABA biosynthesis and ABA-insensitive mutants are also reported to be insensitive to elevated Glc concentrations (Dekkers et al. 2008). Identification of mutants during sugar response screening also displayed defect in ABA metabolism functioning, thus proving a connection amid them. Ethylene signalling pathways are also thoroughly associated with the sugar and ABA sensing interactions. Ethylene mutants, ethylene insensitive 2, ethylene receptor 1 and ethylene insensitive 3 are Glc hypersensitive, whereas constitutive triple response 1, which is a negative regulator of ethylene signalling, is reported to be Glc insensitive (Araújo et al. 2010). Glc and ABA act provocatively during the process of germination and seedling growth. Here exogenous Glc allows germination of wild-type seeds on otherwise inhibitory ABA concentration (Leon and Sheen 2003). On the other hand, ABA upsurges the Suc stimulation and expression of starch biosynthetic genes (Rook et al. 2001). In another study, a splicing factor (SR45) acts as a negative regulator of sugar signalling during initial development. It is involved in the repression of Glc-induced ABA build-up and downregulation of ABA biosynthesis genes (Carvalho et al. 2010). In maize seeds, the signal stimulus is regulated by soluble sugars in the emerging embryo, and the stimulus binds to regulatory elements for ABA and sugars equally (Niu et al. 2002). ABA concentration is improved in transformed ethylene precursor 1-aminocyclopropane-1-carboxylic acid phenocopy gin mutants and wild-type seedlings (Dahiya et al. 2017).
14.7 Conclusions and Future Perspectives
Sugars are acknowledged to partake vast and diversified functions in plants. Sugars are universally present in plants as an essential carbon and energy source. Sugar moieties act as nutrients and regulate vital functions, viz. growth, metabolism and stress tolerance responses. It overall participates in the development of plant from embryogenesis to senescence. In addition to being an imperative energy source and primary metabolite, the focus of investigation has now been shifted to the role of sugar as a regulatory and signalling molecule. Genes are differentially expressed depending upon the plant sugar status. The work in this specific area of research highlights the diversity of developments associated with sugars and their necessity in plants. The regulatory networks to which it is associated with physiological processes like photosynthesis, germination, reproduction and senescence are also explained. Fascinatingly, sugar signalling functions at the gene level in relation with different phytohormones. The role of sugars as signalling moieties is clear when the plant is under environmental stress. Under abiotic stress, plants produce sugars (osmoprotectants) for scavenging ROS and preservation of homeostasis. The sensor proteins of plant cells help in identifying status of sugar in the cell.
Sugar signalling in circadian rhythm and developmental mechanisms has also been conferred. The molecular specifics of signal transduction cascades and their interaction with other mechanisms can be discovered by using an amalgamation of genomic proteomic and genetic methods. Novel genomic tools such as microarray and clustering analysis offer a comprehensive view on the transcript aspects regulated by diverse sugar responses that can help in identification of new regulatory components and target genes. The signal created by signal transduction cascades can stimulate Ca2+, calmodulins, mitogen-activated protein kinases and protein phosphatases and lead to apt gene expression. In spite of the prevailing research on this subject, there remains a lack of information, as countless key problems would be addressed if we might recognize the controlling complexity and the mechanisms involved in sugar homeostasis, cellular and subcellular distribution as well as long-distance translocation. New molecular sensors and FRET (fluorescence resonance energy transfer)-based imaging have the capability to picture and quantify the accurate location and level of different sugar moieties as well as metabolites in alive cells. Sugar signalling study under abiotic stress will remain to be a fascinating research area for many years to come.
References
Afzal S, Sirohi P, Yadav AK et al (2019) A comparative screening of abiotic stress tolerance in early flowering rice mutants. J Biotechnol 302:112–122
Alagbe EE, Amlabu YS, Daniel EO et al (2020) Effect of varying drying temperature on the soluble sugar and nutritional content of banana. Open Chem Eng J 21(1):14
Araújo WL, Ishizaki K, Nunes-Nesi A et al (2010) Identification of the 2-hydroxyglutarate and isovaleryl CoA dehydrogenases as alternative electron donors linking lysine catabolism to the electron transport chain of Arabidopsis mitochondria. Plant Cell 22:1549–1563. https://doi.org/10.1105/tpc.110.075630
Bolouri-Moghaddam MR, Le Roy K, Xiang L et al (2010) Sugar signalling and antioxidant network connections in plant cells. FEBS J 277(9):2022–2037
Borisjuk L, Walenta S, Weber H et al (1998) High-resolution histographical mapping of glucose concentrations in developing cotyledons of Vicia faba in relation to mitotic activity and storage processes: glucose as a possible developmental trigger. Plant J 15:583–591
Carillo P (2018) GABA shunt in durum wheat. Front Plant Sci 9:100. https://doi.org/10.3389/fpls.2018.00100
Carvalho RF, Carvalho SD, Duque P (2010) The plant-specific SR45 protein negatively regulates glucose and ABA signaling during early seedling development in Arabidopsis. Plant Physiol 154:772–783
Chaves MM, Pereira JS, Maroco J et al (2002) How plants cope with water stress in the field. Photosynthesis and growth. Ann Bot 89(7):907–916
Chen M, Xia X, Zheng H et al (2004) The GAOLAOZHUANGREN2 gene is required for normal glucose response and development of Arabidopsis. J Plant Res 117:473–476
Cheng WH, Endo A, Zhou L et al (2002) A unique short-chain dehydrogenase/reductase in Arabidopsis glucose signaling and abscisic acid biosynthesis and functions. Plant Cell 14:2723–2743
Chiang YJ, Stushnoff C, McSay AE (2005) Overexpression of mannitol-1-phosphate dehydrogenase increases mannitol accumulation and adds protection against chilling injury in petunia. J Am Soc Hortic Sci 130:605–610
Corpas FJ, Pedrajas JR, Sandalio LM, León et al (2003) Localization of peroxiredoxin in peroxisomes from pea leaves. Free Radic Res 37:19–20. Taylor, Francis Ltd
Cortina C, Culiáñez-Macià FA (2005) Tomato abiotic stress enhanced tolerance by trehalose biosynthesis. Plant Sci 169:75–82
Cramer GR, Urano K, Delrot S et al (2011) Effects of abiotic stress on plants: a systems biology perspective. BMC Plant Biol 11(1):163
Dahiya A, Saini R, Saini HS et al (2017) Sucrose metabolism: controls the sugar sensing and generation of signalling molecules in plants. J Pharmacogn Phytochem 6:1563–1572
Dar MI, Naikoo MI, Rehman F et al (2016) Proline accumulation in plants: roles in stress tolerance and plant development. In: Iqbal N, Nazar R, Khan NA (eds) Osmolytes and plants acclimation to changing environment: emerging omics technologies. Springer, New Delhi, pp 155–166. https://doi.org/10.1007/978-81-322-2616-1_9
Dekkers BJW, Schuurmans JAMJ, Smeekens SCM (2004) Glucose delays seed germination in Arabidopsis thaliana. Planta 218:579–588
Dekkers BJ, Schuurmans JA, Smeekens SC (2008) Interaction between sugar and abscisic acid signalling during early seedling development in Arabidopsis. Plant Mol Biol 67:151–167
Denver JB, Ullah H (2019) MiR393s regulate salt stress response pathway in Arabidopsis thaliana through scaffold protein RACK1A mediated ABA signalling pathways. Plant Signal Behav 14(6):1600394. https://doi.org/10.1080/15592.324.2019.16003.94
Dewi K (2015) The effect of glucose and light quality on seed germination, vitamin E and chlorophyll content of green bean sprout (Vigna radiata (L.) Wilcz.). KnE Life Sci 2:317
Dutta T, Neelapu NR, Wani SH et al (2018) Compatible solute engineering of crop plants for improved tolerance toward abiotic stresses. In: Wani SH (ed) Biochemical, physiological and molecular avenues for combating abiotic stress tolerance in plants, 1st edn. Academic Press, Cambridge, MA, pp 221–254. https://doi.org/10.1016/B978-0-12-81306.6-7.00012-7
Espasandin FD, Calzadilla PI, Maiale SJ et al (2018) Overexpression of the arginine decarboxylase gene improves tolerance to salt stress in Lotus tenuis plants. J Plant Growth Regul 37(1):156–165
Feil R, Lunn JE (2018) Quantification of soluble sugars and sugar alcohols by LC-MS/MS. In: Plant Metabolomics. Humana Press, New York, pp 87–100
Fernandez O, Béthencourt L, Quero A et al (2010) Trehalose and plant stress responses: friend or foe? Trends Plant Sci 7:409–417
Fraire-Velázquez S, Balderas-Hernández VE (2013) Abiotic stress in plants and metabolic responses. In: Abiotic stress-plant responses and applications in agriculture. In Tech, Rijeka, pp 25–48
Gibson SI (2005) Control of plant development and gene expression by sugar signaling. Curr Opin in Plant Biol 8:93–102
Gill SS, Tuteja N (2010) Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol Biochem 48:909–930
Gill PK, Sharma AD, Singh P et al (2003) Changes in germination, growth and soluble sugar contents of Sorghum bicolor (L.) Moench seeds under various abiotic stresses. Plant Growth Regul 40(2):157–162
Gratão P, Polle A, Lea PJ et al (2005) Making the live of heavy metal-stressed plants a little easier. Funct Plant Biol 32(6):481–494
Gupta AK, Kaur N (2005) Sugar signalling and gene expression in relation to carbohydrate metabolism under abiotic stresses in plants. J Biosci 30(5):761–776
Han SE, Park SR, Kwon HB et al (2005) Genetic engineering of drought-resistant tobacco plants by introducing the trehalose phosphorylase (TP) gene from Pleurotus sajor-caju. Plant Cell Tissue Organ Cult 82:151–158
Hazra A, Dasgupta N, Sengupta C et al (2019) MIPS: functional dynamics in evolutionary pathways of plant kingdom. Genomics 111(6):1929–1945. https://doi.org/10.1016/j.ygeno.2019.01.004
Henry C, Bledsoe SW, Griffiths CA et al (2015) Differential role for trehalose metabolism in salt-stressed maize. Plant Physiol 169(2):1072–1089
Hernandez-Marin E, Martínez A (2012) Carbohydrates and their free radical scavenging capability: a theoretical study. J Phys Chem B 116(32):9668–9675
Hu M, Shi Z, Zhang Z et al (2012) Effects of exogenous glucose on seed germination and antioxidant capacity in wheat seedlings under salt stress. Plant Growth Regul 68(2):177–188
Hussain I, Singh A, Singh NB et al (2019) Plant-nanoceria interaction: toxicity, accumulation, translocation and biotransformation. S Afr J 121:239–247
Jang JC, León P, Zhou L et al (1997) Hexokinase as a sugar sensor in higher plants. Plant Cell 1:5–19
Jofuku KD, Omidyar PK, Gee Z et al (2005) Control of seed mass and seed yield by the floral homeotic gene APETALA2. Proc Natl Acad Sci USA 102:3117–3122
Jong F, Thodey K, Lejay LV et al (2014) Glucose elevates nitrate transporter 2.1 protein levels and nitrate transport activity independently of its hexokinase1 mediated stimulation of nitrate transporter 2.1 expression. Plant Physiol 164:308–320
Jongebloed U, Szederke’nyi J, Hartig K et al (2004) Sequence of morphological and physiological events during natural ageing of a castor bean leaf: sieve tube occlusions and carbohydrate back-up precede chlorophyll degradation. Physiol Plant 120:338–346
Kaplan F, Kopka J, Sung DY et al (2007) Transcript and metabolite profiling during cold acclimation of Arabidopsis reveals an intricate relationship of cold-regulated gene expression with modifications in metabolite content. Plant J 50:967–981
Karthikeyan A, Pandian SK, Ramesh M (2011) Transgenic Indica rice cv. ADT 43 expressing a Δ1-pyrroline-5-carboxylate synthetase (P5CS) gene from Vigna aconitifolia demonstrates salt tolerance. Plant Cell Tissue Organ Cult 107:383–395. https://doi.org/10.1007/s1124.0-011-9989-4
Kaur G, Asthir B (2015) Proline: a key player in plant abiotic stress tolerance. Biol Plant 59:609–619. https://doi.org/10.1007/s1053.5-015-0549-3
Kawakami A, Sato Y, Yoshida M (2008) Genetic engineering of rice capable of synthesizing fructans and enhancing chilling tolerance. J Exp Bot 59:793–802
Keunen ELS, Peshev D, Vangronsveld J et al (2013) Plant sugars are crucial players in the oxidative challenge during abiotic stress: extending the traditional concept. Plant Cell Environ 36(7):1242–1255
Khan MIR, Reddy PS, Ferrante A et al (2019) Plant signalling molecules: role and regulation under stressful environments. Woodhead Publishing Limited, Cambridge
Kim S, Kang JY, Cho DI et al (2004) ABF2, an ABRE-binding bZIP factor, is an essential component of glucose signaling and its overexpression affects multiple stress tolerance. Plant J 40:75–87
Krasensky J, Jonak C (2012) Drought, salt and temperature stress-induced metabolic rearrangements and regulatory networks. J Exp Bot 63(4):1593–1608
Krinsky NI, Johnson EJ (2005) Carotenoid actions and their relation to health and disease. Mol Asp Med 26:459–516
Kushwah S, Laxmi A (2014) The interaction between glucose and cytokinin signal transduction pathway in Arabidopsis thaliana. Plant Cell Environ 37:235–253
Lafta AM, Lorenzen JH (1995) Effect of high temperature on plant growth and carbohydrate metabolism in potato. Plant Physiol 109(2):637–643
Leon P, Sheen J (2003) Sugar and hormone connections. Trends Plant Sci 8:110–116
Li Y, Lee KK, Walsh S et al (2006) Establishing glucose-and ABA-regulated transcription networks in Arabidopsis by microarray analysis and promoter classification using a relevance vector machine. Genome Res 16(3):414–427
Li HJ, Yang AF, Zhang XC et al (2007) Improving freezing tolerance of transgenic tobacco expressing sucrose: Sucrose-1-fructosyltransferase gene from Lactuca sativa. Plant Cell Tissue Organ Cult 89:37–48
Li Y, Zhang H, Zhang Q et al (2019) An AP2/ERF gene, IbRAP2-12 from sweet potato is involved in salt and drought tolerance in transgenic Arabidopsis. Plant Sci 281:19–30. https://doi.org/10.1016/j.plant.sci.2019.01.009
Lim PO, Kim HJ, Nam HG (2007) Leaf senescence. Annu Rev Plant Biol 58:115–136
Liu HL, Dai XY, Xu YY et al (2007) Over-expression of OsUGE-1 altered raffinose level and tolerance to abiotic stress but not morphology in Arabidopsis. J Plant Physiol 164:1384–1390
Livingston DP, Hincha DK, Heyer AG (2009) Fructan and its relationship to abiotic stress tolerance in plants. Cell Mol Life Sci 66(13):2007–2023
Lorenz S, Tintelnot S, Reski R et al (2003) Cyclin D-knockout uncouples developmental progression from sugar availability. Plant Mol Biol 53:227–236
Loreti E, de Bellis L, Alpi A et al (2001) Why and how do plant cells sense sugars? Ann Bot 88:803–812
Loreti E, Poggi A, Novi G et al (2005) A genome-wide analysis of the effects of sucrose on gene expression in Arabidopsis seedlings under anoxia. Plant Physiol 137:1130–1138
Lu P, Magwanga RO, Lu H et al (2018) A novel G-protein-coupled receptors gene from upland cotton enhances salt stress tolerance in transgenic Arabidopsis. Genes 9(4):209
Masclaux-Daubresse C, Purdy S, Lemaitre T et al (2007) Genetic variation suggests interaction between cold acclimation and metabolic regulation of leaf senescence. Plant Physiol 143:434–446
Miller G, Suzuki N, Ciftci-Yilmaz S et al (2010) Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant Cell Environ 33:453–467
Miranda JA, Avonce N, Suárez R et al (2007) A bifunctional TPS-TPP enzyme from yeast confers tolerance to multiple and extreme abiotic-stress conditions in transgenic Arabidopsis. Planta 226:1411–1421
Mohammadkhani N, Heidari R (2008) Drought-induced accumulation of soluble sugars and proline in two maize varieties. World Appl Sci J 3(3):448–453
Møller IM (2001) Plant mitochondria and oxidative stress: Electron transport, NADPH turnover and metabolism of reactive oxygen species. Annu Rev Plant Biol 52(1):561–591
Møller IM, Sweetlove LJ (2010) ROS signalling – specificity is required. Trends Plant Sci 15:370–374
Moller A, Yabuta Y, Shigeoka S (2008) Galactinol and raffinose constitute a novel function to protect plants from oxidative damage. Plant Physiol 147(3):1251–1263
Moore B, Zhou L, Rolland F et al (2003) Role of the Arabidopsis glucose sensor HXK1 in nutrient, light, and hormonal signalling. Science 300:332–336
Morsy MR, Jouve L, Hausman JF et al (2007) Alteration of oxidative and carbohydrate metabolism under abiotic stress in two rice (Oryza sativa L.) genotypes contrasting in chilling tolerance. J Plant Physiol 164(2):157–167
Nemati I, Moradi F, Gholizadeh S et al (2011) The effect of salinity stress on ions and soluble sugars distribution in leaves, leaf sheaths and roots of rice (Oryza sativa L.) seedlings. Plant Soil Environ 57(1):26–33
Nishizawa A, Yabuta Y, Shigeoka S (2008) Galactinol and raffinose constitute a novel function to protect plants from oxidative damage. Plant Physiol 147(3):1251–1263
Niu X, Helentjaris T, Bate NJ (2002) Maize ABI4 binds coupling element1 in abscisic acid and sugar response genes. Plant Cell 14:2565–2575
Oaks A (1994) Primary nitrogen assimilation in higher plants and its regulation. Can J Bot 72:739–750
Ohto M, Fischer RL, Goldberg RB et al (2005) Control of seed mass by APETALA2. Proc Natl Acad Sci USA 102:3123–3128
Osakabe Y, Yamaguchi-Shinozaki K, Shinozaki K et al (2014) ABA control of plant macroelement membrane transport systems in response to water deficit and high salinity. New Phytol 202(1):35–49
Parvanova D, Popova A, Zaharieva I et al (2004) Low temperature tolerance of tobacco plants transformed to accumulate proline, fructans, or glycine betaine. Variable chlorophyll fluorescence evidence. Photosynthetica 42:179–185
Paul MJ, Gonzalez-Uriarte A, Griffiths CA et al (2018) The role of trehalose-6-phosphate in crop yield and resilience. Plant Physiol 177(1):12–23
Pennycooke JC, Jones ML, Stushnoff C (2003) Down-regulating α-galactosidase enhances freezing tolerance in transgenic petunia. Plant Physiol 133(2):901–909
Peshev D, Van den Ende W (2013) Sugars as antioxidants in plants. In: Tuteja N, Gill SS (eds) Crop improvement under adverse conditions. Springer-Verlag, Heidelberg, pp 285–308
Pien S, Wyrzykowska J, Fleming AJ (2001) Novel marker genes for early leaf development indicate spatial regulation of carbohydrate metabolism within the apical meristem. Plant J 25:663–674
Prasanna R, Pabby A, Singh PK (2004) Effect of glucose and light/dark environment on pigmentation profiles in Calothrix elenkenii. Folia Microbiol 49:26–30
Pressman E, Peet MM, Pharr DM et al (2002) The effect of heat stress on tomato pollen characteristics is associated with changes in carbohydrate concentration in the developing anthers. Ann Bot 90(5):631–636
Price J, Li TC, Kang SG et al (2003) Mechanisms of glucose signalling during germination of Arabidopsis. Plant Physiol 132:1424–1438
Price J, Laxmi A, Martin SKS et al (2004) Global transcription profiling reveals multiple sugar signal transduction mechanisms in Arabidopsis. Plant Cell 16:2128–2150
Pritchard SL, Charlton WL, Baker A et al (2002) Germination and storage reserve mobilization is regulated independently in Arabidopsis. Plant J 31:639–647
Pujni D, Chaudhary A, Rajam MV (2007) Increased tolerance to salinity and drought in transgenic indica rice by mannitol accumulation. J Plant Biochem Biot 16:1–7
Ramel F, Sulmon C, Bogard M et al (2009) Differential patterns of reactive oxygen species and antioxidative mechanisms during atrazine injury and sucrose-induced tolerance in Arabidopsis thaliana plantlets. BMC Plant Biol 9(1):28
Rathinasabapathi B (2000) Metabolic engineering for stress tolerance: installing osmoprotectants synthesis pathways. Ann Bot 86(4):709–716
Rineau F, Shah F, Smits MM et al (2013) Carbon availability triggers the decomposition of plant litter and assimilation of nitrogen by an ectomycorrhizal fungus. ISME J 7(10):2010–2022
Riou-Khamlichi C, Menges M, Healy JM et al (2000) Sugar control of the plant cell cycle: differential regulation of Arabidopsis D-type cyclin gene expression. Mol Cell Biol 20:4513–4521
Rivero RM, Kojima M, Gepstein A et al (2007) Delayed leaf senescence induces extreme drought tolerance in a flowering plant. Proc Natl Acad Sci USA 104:19631–19636
Roitsch T, Gonzalez MC (2004) Function and regulation of plant invertases: sweet sensations. Trends Plant Sci 9:606–613
Rolland F, Sheen J (2005) Sugar sensing and signalling networks in plants. Biochem Soc 33:269–271
Rolland F, Baena-Gonzales E, Sheen J (2006) Sugar sensing and signalling in plants: conserved and novel mechanisms. Annu Rev Plant Biol 57:675–709
Rook F, Corke F, Card R et al (2001) Impaired sucrose-induction mutants reveal the modulation of sugar-induced starch biosynthetic gene expression by abscisic acid signalling. Plant J 26:421–433
Rosa M, Prado C, Podazza G et al (2009) Soluble sugars: metabolism, sensing and abiotic stress – a complex network in the life of plants. Plant Signal Behav 4(5):388–393
Ruuska SA, Girke T, Benning C et al (2002) Contrapuntal networks of gene expression during Arabidopsis seed filling. Plant Cell 14:1191–1206
Sami F, Yusuf M, Faizan M et al (2016) Role of sugars under abiotic stress. Plant Physiol Biochem 109:54–61
Scott P (2008) Physiology and behaviour of plants, 1st edn. Wiley J and Sons Ltd, Chichester
Sedigheh N (2019) Morphological, biochemical and soluble sugars characters of Aloe vera subjected to saline condition. Egypt Acad J Biol Sci, H Botany 10(1):13–23
Seki M, Ishida J, Narusaka M et al (2002) Monitoring the expression pattern of around 7. Funct Integr Genomics 2(6):282–291
Sheen J, Zhou L, Jang JC (1999) Sugars as signalling molecules. Curr Opin Plant Biol 2(5):410–418
Shen B, Jensen RC, Bohnert HJ (1997) Increased resistance to oxidative stress in transgenic plants by targeting mannitol biosynthesis to chloroplasts. Plant Physiol 113:1177–1183
Singh M, Kumar J, Singh S et al (2015) Roles of osmoprotectants in improving salinity and drought tolerance in plants: a review. Rev Environ Sci Biotechnol 14(3):407–426
Singh A, Singh NB, Hussain I et al (2016) Green synthesis of nano zinc oxide and evaluation of its impact on germination and metabolic activity of Solanum lycopersicum. J Biotechnol 233:84–94
Singh A, Singh NB, Hussain I et al (2017) Effect of biologically synthesized copper oxide nanoparticles on metabolism and antioxidant activity to the crop plants Solanum lycopersicum and Brassica oleracea var. botrytis. J Biotechnol 262:11–27
Singh A, Singh NB, Afzal S et al (2018) Zinc oxide nanoparticles: a review of their biological synthesis, antimicrobial activity, uptake, translocation and biotransformation in plants. J Mater Sci 53(1):185–201
Singh A, Hussain I, Singh NB et al (2019) Uptake, translocation and impact of green synthesized nanoceria on growth and antioxidant enzymes activity of Solanum lycopersicum L. ecotoxicology and environmental safety. Ecotox Environ Safe 182:109410
Sinha AK, Roitsch T (2002) Effect of different sugars on photosynthesis and chlorophyll fluorescence in photoautotrophic tomato suspension cultures. Photosynthetica 39:611–644
Smeekens S (2000) Sugar-induced signal transduction in plants. Annu Rev Plant Biol 51(1):49–81
Soltani A, Gholipoor M, Zeinali E et al (2006) Seed reserve utilization and seedling growth of wheat as affected by drought and salinity. Environ Exp Bot 55:195–200
Songstad DD, Petolino JF, Voytas DF et al (2017) Genome editing of plants. Crit Rev Plant Sci 36:1–23. https://doi.org/10.1080/07352.689.2017.12816.63
Spreitzer RJ, Salvucci ME (2002) RUBISCO: structure, regulatory interactions, and possibilities for a better enzyme. Annu Rev Plant Biol 53:449–475
Stephanie C, McClure MD, Katherine FACP et al (2010) Vitamin a use today and its potential toxicities. Clin Geriatr 18(2):41–45
Stessman D, Miller A, Spalding M et al (2002) Regulation of photosynthesis during Arabidopsis leaf development in continuous light. Photosynthesis Res 72:27–37
Stoyanova S, Geuns J, Hideg E et al (2011) The food additives inulin and stevioside counteract oxidative stress. Int J Food Nutr 62(3):207–214
Suprasanna P, Nikalje GC, Rai AN (2016) Osmolyte accumulation and implications in plant abiotic stress tolerance. In: Iqbal N, Nazar R, Khan NA (eds) Osmolytes and plants acclimation to changing planta environment. Emerging Omics Technologies/Springer, New Delhi, pp 1–12. https://doi.org/10.1007/978-81-322-2616-1_1
Surekha CH, Kumari KN, Aruna LV et al (2014) Expression of the Vigna aconitifolia P5CSF129A gene in transgenic pigeon pea enhances proline accumulation and salt tolerance. Plant Cell Tissue Organ Cult 116:27–36. https://doi.org/10.1007/s1124.0-013-0378-z
Taiz L, Zeiger E (2002) Plant physiology, 3rd edn. Sinauer Associates, Sunderland
Takahashi F, Sato-Nara K, Kobayashi K et al (2003) Sugar-induced adventitious roots in Arabidopsis seedlings. J Plant Res 116:83–91
To JP, Reiter WD, Gibson SI (2003) Chloroplast biogenesis by Arabidopsis seedlings is impaired in the presence of exogenous glucose. Physiol Plant 118:456–463
Tran LS, Nakashima K, Shinozaki K et al (2007) Plant gene networks in osmotic stress response: from genes to regulatory networks. Methods Enzymol 428:109–128
Tsai CH, Miller A, Spalding M et al (1997) Source strength regulates an early phase transition of tobacco shoot morphogenesis. Plant Physiol 115:907–914
Van den Ende W, Peshev D (2013) Sugars as antioxidants in plants. In: Crop improvement under adverse conditions. Springer, New York, pp 285–307
Van den Ende W, Valluru R (2009) Sucrose, sucrosyl oligosaccharides, and oxidative stress: scavenging and salvaging? J Exp Bot 60(1):9–18
Vives-Peris V, Gómez-Cadenas A, Pérez-Clemente RM (2017) Citrus plants exude proline and phytohormones under abiotic stress conditions. Plant Cell Rep 36:1971–1984. https://doi.org/10.1007/s0029.9-017-2214-0
Wang ZY, Li FM, Xiong YC et al (2008) Soil-water threshold range of chemical signals and drought tolerance was mediated by ROS homeostasis in winter wheat during progressive soil drying. J Plant Growth Regul 27:309–319
Wei J, Miaoa H, Wang Q (2011) Effect of glucose on glucosinolates, antioxidants and metabolic enzymes in Brassica sprouts. Sci Hortic 129:535–540
Wei K, Ma C, Sun K et al (2020) Relationship between optical properties and soluble sugar contents of apple flesh during storage. Postharvest Biol Tec 159:111021
Wingler A (2007) Transcriptional or post-transcriptional regulation – how does a plant know when to senesce? New Phytol 175:7–9
Woodrow P, Ciarmiello LF, Annunziata MG et al (2017) Durum wheat seedling responses to simultaneous high light and salinity involve a fine reconfiguration of amino acids and carbohydrate metabolism. Physiol Plant 159:290–312. https://doi.org/10.1111/ppl.12513
Xiao W, Sheen J, Jang JC (2000) The role of hexokinase in plant sugar signal transduction and growth and development. Plant Mol Biol 44(4):451–461
Yang L, Xu M, Koo Y, Poethig RS (2013) Sugar promotes vegetative phase change in Arabidopsis thaliana by repressing the expression of MIR156A and MIR156C. elife 2:e00260
Yuanyuan M, Yali Z, Jiang L et al (2009) Roles of plant soluble sugars and their responses to plant cold stress. Afr J Biotechnol 8(10):2004–2010
Zhang SZ, Yang BP, Feng CL et al (2005) Genetic transformation of tobacco with the trehalose synthase gene from Grifola frondosa Fr. enhances the resistance to drought and salt in tobacco. J Integr Plant Biol 47:579–587
Zhao MG, Liu RJ, Chen L et al (2009) Glucose-induced inhibition of seed germination in Lotus japonicus is alleviated by nitric oxide and spermine. J Plant Physiol 166:213–218
Zhifang G, Loescher WH (2003) Expression of a celery mannose-6-phosphate reductase in Arabidopsis thaliana enhances salt tolerance and induces biosynthesis of both mannitol and a glucosyl-mannitol dimer. Plant Cell Environ 26:275–283
Zouari M, Ahmed CB, Elloumi N et al (2016) Impact of proline application on cadmium accumulation, mineral nutrition and enzymatic antioxidant defense system of Olea europaea L. cv. Chemlali exposed to cadmium stress. Ecotoxicol Environ Saf 128:195–205. https://doi.org/10.1016/j.ecoen.v.2016.02.024
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2021 The Author(s), under exclusive license to Springer Nature Switzerland AG
About this chapter

Cite this chapter
Afzal, S., Chaudhary, N., Singh, N.K. (2021). Role of Soluble Sugars in Metabolism and Sensing Under Abiotic Stress. In: Aftab, T., Hakeem, K.R. (eds) Plant Growth Regulators. Springer, Cham. https://doi.org/10.1007/978-3-030-61153-8_14
Download citation
DOI: https://doi.org/10.1007/978-3-030-61153-8_14
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-61152-1
Online ISBN: 978-3-030-61153-8
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)