
Abstract
Secondary plant metabolites are natural bioactive compounds which are an important income for the pharmaceutical, food, cosmetic, agriculture, and other sectors due to their health-promoting properties and prevention and treatment of some diseases. The secondary metabolites can be classified into three main groups: phenolic compounds, terpenoids, and nitrogen compounds. The secondary metabolism in plants is a mechanism of adaptation and evolution as a defense to harsh environmental factors that induce stress. According to the hormesis curve of each plant model, the stress can be divided into distress (bad stress that leads to damage and ultimately plant death) or eustress (good stress that leads to activation of secondary metabolism). The environmental factors can be divided into biotic and abiotic which can be artificially induced to activate plant defense responses leading to the production of secondary metabolites. Several approaches to this process called elicitation have been proposed in the last decades with different types of metabolism-inducing factors or elicitors. Novel elicitation using abiotic factors includes electromagnetic waves (including several wavelengths of the light spectra, and electric and magnetic fields), acoustic waves, nanostructures, volatile compounds, nutrient deprivation, and several metals and salt soil pollutants. In the same order, novel elicitation using biotic factors include new bacteria consortium, fungi, phytohormones, and miRNA solutions. In general, the purpose of elicitation is to interact with the biochemical routes in order to produce secondary metabolites in high quantities, usually with negative effects in biomass production or morphology, but increasing plant quality in terms of aroma, taste or color. The metabolic profile and general response of elicitation vary greatly depending on the plant model, the elicitor level or concentration, and the stimulation time. Considering these facts, this chapter clearly and concisely discusses the most current strategies of elicitation for the increase of secondary metabolites production in plants.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
5.1 Introduction
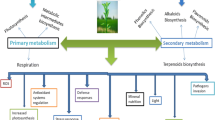
A variety of substances or compounds called secondary metabolites are mainly responsible for the adaptation of plants to environmental changes. Plants synthesize a wide range of secondary metabolites such as alkaloids, flavonoids, phenolic, steroids, anthocyanins between others, which are used as pharmaceuticals, agrochemicals, biopesticides, color, additives, etc. Secondary metabolites are not considered to be part of fundamental life processes of plants, however, they play a significant role in protection from insect, pest, herbivores, phytopathogens, and other harsh environmental variables (Thakur et al. 2019). Therefore, the synthesis of secondary metabolites depends on internal and external factors (stressors and stress factors, respectively) that influence plant metabolism, and can affect plant reproduction and productivity (Kranner et al. 2010). The stress factors, that for their nature can be biotic or abiotic, modify positively or negatively the plant metabolism (Cheynier et al. 2013). Biotic stress is the result of the interaction between plant and viral, bacterial, fungi, pheromones, phytohormones, and nucleic acids among others. Meanwhile, abiotic stress can be physical factors (Light spectra, temperature, water stress, acoustic waves, others) and chemical factors (Nanostructures, gasses, nutriment, others) (Vázquez-Hernández et al. 2019).
According to data provided by Agathokleous et al. (2019), exist 109,821 publications that review the topic of “plant stress” only in the period 2000 to 2018. The results indicated that the dose–response is not always linear, observing a biphasic response when the doses allow it. This balance of response between the plant and stress factors provides information on a positive (eustress) or a negative effect (distress), a phenomenon called “Hormesis” (Agathokleous et al. 2019; Vázquez-Hernández et al. 2019). The concept of hormesis is a term used in medicine for the application of toxins in low doses (Calabrese 2004). Paracelsus (1493–1541) defined it as “All things are poison and nothing is without poison, only the dose permits something not to be poisonous” (cited by (Vázquez-Hernández et al. 2019)). Currently, the term is applied in horticultural and agricultural practices as a biphasic response in which doses of a toxic agent could cause inhibition (distress) or can cause stimulation (eustress) (Vargas-Hernandez et al. 2017).
Plants are sessile organisms susceptible to the interaction between various types of stress, which has resulted in an evolved defense system that increases the synthesis of secondary metabolites (Ghorbanpour et al. 2014). The variety of stress factors together can affect the plant physiology, plant-plant interaction, defense type, reproductivity, among others. For example, salinity and low/high-temperature are conditions which restrict plant growth and productivity (Akula and Ravishankar 2011). In the signaling response to pathogens or herbivorous insects, several response pathways are invoked, some of these are induced by infection and some are performed regardless of the antimicrobial nature (Zaynab et al. 2018). Another example is the interaction between plants and herbivory insects that causes the plant to emit volatile organic compounds which influence the plant-to-plant communication, pollinators, and other insects, and increase fluidity of cell membranes for thermo-tolerance and leaf tissue protection from atmospheric oxidants within and around leaves (Faiola and Taipale 2020).
The foregoing indicates that plants can react in various ways in the presence of one or more stress factors and that, in the same way, the response to the stimulus may be the activation of a synthesis pathway of only one metabolite or a series of secondary metabolites. For this reason, this work will focus on stress factors and the production of secondary metabolites in plants.
5.2 Abiotic Stress
“Any unfavorable condition or substance that affects or blocks a plant's metabolism, growth or development” is the definition of plant stress suggested by Lichtenthaler (1996). Currently, this definition has been modified depending on the stimulus origin, defining as stress factors those stimuli that are external to the plant, biotic (fungi, insects, etc.) or abiotic (temperature, luminosity, nanoparticles, metals and polluting salts, water, etc.) (Kranner et al. 2002; Thakur et al. 2019). Abiotic stress origin is not biological and can be divided into chemical or physical (Vázquez-Hernández et al. 2019). For example, the adaptation to cold environments in some plants is the result of an increase in the synthesis of flavonoids due to acclimatization processes at low temperatures or by the application of UV radiation (Samanta et al. 2011; Nakabayashi et al. 2014). In Vitis vinifera, it has been observed that stimulation with heavy metals such as Cadmium (Cd2+), Cobalt (Co2+) and Silver (Ag+), can increase the synthesis of Resveratrol (Cai et al. 2013), while the application of UV-C irradiation induces the synthesis of stilbene (Wang et al. 2010; Liu et al. 2010). This indicates that the synthesis of secondary metabolites will depend on various factors such as the type of stimulus, the concentration, and the form of application. These same observations are appreciated by Feregrino-Perez et al. (2018), where the effect of nanomaterials on germination, development of plants, and synthesis of secondary metabolites is reviewed, concluding that the stress level will depend on the used nanomaterial, the dose and the time of exposition. Low/high-temperature, relative humidity in air, drought, microelements shortage, and CO2 reduction are classical abiotic factors for plants elicitation. The role of novel abiotic stress factors on the production of secondary metabolites is described below.
5.2.1 Electromagnetic Sources
Electromagnetic phenomena can be seen as an abiotic stress elicitor to affect plants. In this context, many reports have demonstrated more advantages than disadvantages when strong or weak electric fields, magnetic fields were applied to plants (Dannehl 2018). Electromagnetic sources have been studied as another possibility to increase plant growth and development due to the alteration in the electrostatic balance of the plant system at the cell membrane level (Radhakrishnan 2019). An electromagnetic field is produced by a distribution of electric current and charge. An electric field (EF) can occur under high-voltage lines and the units in the SI are newtons per coulomb or, equivalently volts per meter (V/m); in the same way, a magnetic field (MF) is usually measured in terms of its magnetic flux density whose unit is expressed as Tesla (T) (Dannehl 2018).
Pulsed electric field PEF technology consists of the application of short, high power electrical pulses to products placed in a treatment chamber, confined between electrodes (Soliva-Fortuny et al. 2017). A high electric field can cause cell membrane disruption, whereby inner secondary metabolites are released from intracellular cell compartments, resulting in a high content of bioactive compounds (Odriozola-Serrano et al. 2009; Janositz and Knorr 2010). In the case of MF, several studies have used small boxes with coils, iron bars, a function generator, and a power amplifier. The application of magnetized water to plants is a novel area of indirect application of MF that the scientific community is currently researching (Dannehl 2018).
Light is an electromagnetic wave within the visible spectrum, however, that definition depends upon the sense of sight involving the response of individuals (Koshel 2004), therefore, the UV and infrared parts of the electromagnetic spectrum are roughly included, but will be considered in this chapter due to their importance in the plant production of secondary metabolites. Plants sense light through specific molecules called photoreceptors that trigger specific signals for photomorphogenesis or other defense systems. Depending on the dose rate and exposure time, either insufficient or excess levels, light can become a type of eustress, producing several effects, from damage to cellular components to triggering of defense systems for secondary metabolite production (Alvarado et al. 2019; Muller-Xing et al. 2014; Akula and Ravishankar 2011). The next sections of this chapter will discuss the effect of electromagnetic sources in the production of natural bioactive compounds.
5.2.1.1 Light
Light is one of the most important and obvious requirements for plant growth and development, where the energy of sunlight and artificial light sources is mainly used for photosynthesis. However, light is not only involved in the photosynthesis process but also in the production of natural bioactive compounds, gene expression, and synchronization of the circadian clock in the light/dark cycle (Larner et al. 2018). Changes in the light intensity, quality, direction, and duration are sensed by specialized photoreceptors which are specially designed proteins that sense light, triggering chain reactions that have been studied in terms of photomorphogenesis and primary and secondary metabolites production (Alvarado et al. 2019). Photoreceptors perceive specific light wavelengths of over a continuous spectral range through a small cofactor or chromophore molecule (Burgie et al. 2014). Five photosensory systems have been identified: phytochromes perceiving red (660–700 nm) and far-red (700–750 nm), cryptochromes, phototropins, and members of the Zeitlupe family perceiving blue (495–400 nm) and UV-A (400–315 nm), and UV Resistance Locus 8 (UVR8) perceiving (315–280 nm) (Bantis et al. 2018; Alvarado et al. 2019).
Recent investigation has focused on the effect of light technology in plant growth, developmental traits, and primary and secondary metabolites by using one or more light wavelengths, intensities, and photoperiods. It has been reported that blue light increases phenolic compounds by promoting the production of malonyl CoA and coumaroyl CoA, participating in the synthesis of phenolic compounds (Qian et al. 2016). In addition, red and far-red wavelengths are perceived by the phytochromes photoreceptors, which regulates biosynthetic pathways involved in the synthesis of anthocyanins, molecules that belong to the phenolic compounds known as flavonoids and have many functions in plants including pigmentation (Alokam et al. 2002). In the same way, plants produce secondary metabolites such as flavonoids and anthocyanins to cope with cell damage produced by UV radiation (Jiang et al. 2017b). Serious damage to DNA, membrane, and proteins can be caused by UV-B radiation, whereas UV-A induces DNA damage less efficiently because of the activation of photoreactions forming reactive oxygen species (ROS) (Hideg and Strid 2017; Häder et al. 2015).
Supplemental lighting has been accepted for improving horticultural crops. Light-emitting diode (LED) technology has been linked to controlled environments in horticulture for achieving crop yield, phytochemical content, nutritional value, flowering control, transplant success, pre-harvest and postharvest product quality, and production of regeneration material (Bantis et al. 2018; Alvarado et al. 2019). LEDs have allowed a sustainable and highly efficient use of energy and reproduce true spectral composition of blue, green, red, and far-red wavelengths that matches with plant-specific photoreceptors (Singh et al. 2015). Other light technologies, as high sodium pressure (HSP) and other high-intensity discharge (HID) lamps are still used in greenhouse and plant experimentation, however, LED technology is replacing these devices due to the various advantages LEDs offer. Table 5.1 summarizes some examples of the application of supplemental light on plants or foods with a commercial interest and presents the effect on the production of natural bioactive compounds.
5.2.1.2 Electric and Magnetic Fields
Magnetic fields (MFs) are considered an abiotic factor that can induce eustress with significant effects on the growth and development of plants. The effect of light, gravity, mechanical damage, and electrical signaling on plants has been studied and documented over the past years concluding strong facts relating to phototropism, gravitropism, and thigmotropism (Maffei 2014). The geomagnetic field (GMF) is a natural component of our environment, however, its impact on plant growth and development is not well-understood, moreover, the effects of artificial magnetic fields on plants have been poorly studied (Maffei 2014). Several experiments with lower and higher values than the GMF has been conducted with predominantly positive effects depending on the plant, time of exposure and intensity. For example, an increase in germination or subsequent seedling growth barley, corn, beans, wheat, hornwort, mung bean, pea, chickpea, tomato, and okra, but it was reduced in seeds of rice. In a similar way, the effect on roots, shoots, gravitropism, photosynthesis, and lipid composition present a similar pattern (Maffei 2014).
Several theories and studies about the biological effect on MF have been proposed. A polar structure in various chemical bonds in the organic material may be linked to the polar water molecules and dissociated ions of mineral salts conferring magnetic properties (Chepets et al. 1985). A MF can decrease the disease index of plants due to the modulation of calcium signaling, and proline and polyamines pathways (Radhakrishnan 2019). The plant cells contain about 4500 iron atoms in the ferritin molecules involved in growth and metabolism. The magnetic rotator moment of ultimate iron atoms creates an external MF which collectively generates an atom re-positioning in the direction of MF that leads to an increase of the plant temperature (Vaezzadeh et al. 2006). Photoreceptors have been also proposed to be potential magnetoreceptors since cryptochromes and phytochromes produce radical pairs after the exposure to their corresponding light wavelength triggers (Maffei 2014; Dhiman and Galland 2018). Cryptochrome-dependent responses such as blue-light-dependent anthocyanin accumulation and blue-light-dependent degradation of CRY2 protein were enhanced at higher magnetic intensities in Arabidopsis mutants lacking cryptochromes (Ahmad and Jones 1979). Limited information is available on the molecular basis and the function of the MF receptors and their activation by physiological signals, therefore, their involvement in directing the overall response in different plant organs is yet to be determined (Radhakrishnan 2019).
Static magnetic field (SMF) exposition in plants has been found to be an effective and emerging tool to control diseases and increase tolerance against the adverse environment (Radhakrishnan 2019). However, a small number of studies have been attempted to determine the role of MF on plant tolerance against various stress conditions (Radhakrishnan 2019). The effect on secondary metabolites production has been scarcely studied; presenting an opportunity area in this field.
Electrostatic fields (EF) has also been used in the horticultural industry. Most of the applications of this technology are for germination and seedlings improvement due to its importance in the supply chain. For example, drought resistance and removal of free radicals in maize seedlings with electric field intensity 200 kV/m, pulse width 80 ms and frequency 1 Hz were analyzed by He et al. (2017), where the growth of root, the ability of self-organization, and the respiration metabolism of root cells was improved. Another example with potato tubers subjected to pulsed electric field (PEF) through a treatment voltage across the potatoes of 5 kV and a discharge capacitance of 450 pF prior to planting had an increase in the yield of 22–29% (Gachovska et al. 2015). In the same way, winter wheat seeds increased the germination energy up to 32.41% and weight up to 23.8% under PEF (Starodubtseva et al. 2018). In the experiment of Yan et al. (2017), PEF through a high-voltage pulse power supply and arc electrode was studied on cotton seeds vigor with different frequencies of 1, 5, 10, 20, and 50 Hz at the voltage of 16 and 20 kV, and the treatment time was 40 s, finding that when the frequency of the electric field increased, the effects increased and reached the maximum at 10 Hz, and after 10 Hz, as the electric field frequency increased, the effects began to decrease. This last experiment exemplifies the hormetic curve in a vigor treatment where there is a maximal dose at which a maximum response value is reached and then vigor starts decreasing.
In several studies, PEF pretreatments in plants, whole fruits, or other food sources result in an increase in the natural bioactive compounds. When biological cells are exposed to an EF, the charge accumulates along the plasma membrane causing electroporation, a transmembrane potential difference which causes porosity, and thus the diffusion of intracellular components in cellular juice increasing the extractability of natural bioactive compounds by the release of solutes into the solvent (El Kantar et al. 2018; Vicaş et al. 2017; Barba et al. 2015; Hendrawan et al. 2019). The time exposure and intensity of the EF are critical since a lower EF may form smaller pores allowing the ions to pass through, but large molecules may not get out of the cell, however, higher EF are suspected to damage antioxidant compounds due to long exposure to high-voltage electric current (Hendrawan et al. 2019). Table 5.2 summarizes some examples of the application of magnetic or electric fields on plants or foods with a commercial interest and presents the effect on the production of natural bioactive compounds.
5.2.2 Acoustic Emissions
Acoustic emissions (AE) stimulus is one of the recent physical abiotic factors whose beneficial effects on plant growth, development, and health have been discussed. AE from ecological conditions or artificially applied can initiate diverse signals that trigger transduction cascades, similar to other abiotic stress factors (Alvarado et al. 2019). From a bioacoustics perspective, chewing serves as an alarm signal to plants and has been demonstrated that applying recorded insect chewing sounds caused an increase of phytochemical production (Appel and Cocroft 2014). In the same way, Jeong et al. (2014), reported an improvement of natural protection responses in rice plants caused by amplification at 100 decibels of a wide range of frequencies between 0 and 1.5 kHz. Moreover, Hassanien et al. (2014), found a higher disease resistance in pepper, cucumber, and tomato after AE treatments.
The biological mechanism of how sound affects plants is still under discussion. A mechano-stimuli perception of waves has been proposed, but a reliable explanation of sound-specific structure for recognition by plants has not been completely elucidated (Alvarado et al. 2019). This mechanism consists of the second messenger of calcium ion (Ca2+) signals. The channels that mediate Ca2+ flux are possibly located in the plasmatic membrane where Ca2+ is sensed possibly through various Ca2+ sensors and/or CDPKs (Calcium-dependent protein kinase), which pass the message through phosphorylation/dephosphorylation to different signaling proteins or to transcription factors (Mishra et al. 2016). In that way, it is strongly suggested that AE can influence the synthesis of secondary metabolites. The most common acoustic emission utilized for the stimulation of bioactive compounds is ultrasound (US). Several mechanisms of how the US interacts within the cell have been proposed. When the US is applied, cavitation bubbles creates a pressure zone change that occurs and increases up to 400 km h−1, causing higher porosity, rupture or removal of cell membranes, facilitating the mass transfer from the cells’ interior when imploding (Toma et al. 2001; Vinatoru 2001). In that way, the increase of natural bioactive compounds may be caused by better extractability due to rupture of membranes of cell organelles, however, when a decrease is presented, it could be explained by the creation of reactive forms of oxygen (ROS) during cavitation, and that the collapsing bubbles release high doses of energy, raising the temperature (>5000 K) enough to decompose polyphenols (Witrowa-Rajchert et al. 2014; Kentish and Ashokkumar 2011). The second possible explanation is an enhancement of enzymes activity when the US is applied by contact, leading to phenolic compounds’ reduction, more significantly after longer treatment time (Wiktor et al. 2016). Ampofo and Ngadi (2020) established that elicitation of common beans with the US, increased the accumulation of stress markers from the onset until the process was arrested, signifying a demand for sprout protection, resulting in an elevated stimulation of defense phenolic triggering enzymes (PAL and TAL), and final biosynthesis of phenolic compounds. In that way, application in food or plants of US treatment should be studied to find the optimal time, frequency, and intensity in order to optimize the production of secondary metabolites. Table 5.3 summarizes some examples of the application of acoustic emissions on plants or foods with a commercial interest and presents the effect on the production of natural bioactive compounds.
5.2.3 Nanoparticles
Nanoparticles (NPs) vary in size from 1 to 100 nm and have physicochemical properties, due to their dimensions, which generate a high added value for the nanotechnology industry (Yokel and MacPhail 2011). Nanoscale materials can be found on medical imaging, drug delivery, personal care products, cosmetics, clothing, electronics, agrochemicals, motor vehicles, among other products and applications (Vance et al. 2015; Yokel and MacPhail 2011). The metal-based NPs most commonly studied and found in industrial products are Cd (cadmium) in various complexes, GaAs (gallium arsenide), Au (gold), Ni (nickel), Pt (platinum), Ag (silver), Al2O3 (aluminum oxide or alumina), CeO2 (cerium dioxide or ceria), SiO2 (Silicon dioxide or silica), TiO2 (titanium dioxide or titania), ZnO (zinc oxide), CuO (copper oxide), and Fe3O4/Fe2O3 (iron oxides) (Khot et al. 2012; Yokel and MacPhail 2011). Among the carbon-based nanomaterials often studied are fullerene, single-walled carbon nanotubes (SWCNTs), and multiwalled carbon nanotubes (MWCNTs) (Balbus et al. 2007).
In contrast to its benefits, the release of nanomaterial-containing wastes has become a threat, since they cause pollution to air, water, and soil (Oberdörster et al. 2005). Their size, equivalent to that of cellular components, allows them to easily permeate cells, causing adverse biological effects (plants and animals) (Shang et al. 2014; Vecchio et al. 2012). In plant cells, NPs can enter from the apoplast and cross the plasma membrane towards cytosol or other organelles via endocytosis, specific membrane-bound transporter proteins or through induction of new pores by using ion carrier substances; subsequently, they can be transported between cells through symplastic flow (Anjum et al. 2019; Marslin et al. 2017). Despite its potential of toxicity, studies have reported positive effects on plant development and physiology, which are dependent on the nature of the nanomaterial, dose and time of exposure, the plant species, and growth conditions (Cox et al. 2016).
Positive physiological effects using carbon-based NPs include increased water uptake, and enhanced assimilation of CO2 in broccoli (Martínez-Ballesta et al. 2016), promotion of seed germination and root growth in rice (Jiang et al. 2014), enhanced germination and seedling growth in sweet corn, barley, rice soybean, switchgrass, and tomato (Lahiani et al. 2015; Tiwari et al. 2014), increase fruit yield in tomato and bitter melon (Khodakovskaya et al. 2013; Kole et al. 2013), among many others. Studies using metallic nanoparticles have reported similar effects. In wheat, CeO2 particles improved plant growth, shoot biomass, and grain yield (Rico et al. 2014). Au nanoparticles in chinese mustard (Brassica juncea) had positive effects on growth parameters and seed yield (Arora et al. 2012). The response of maize exposed to ZnO nanoparticles showed enhanced germination, seedling vigor, and zinc biofortification of grains (Subbaiah et al. 2016).
The effect of NPs on secondary plant metabolism is still largely unknown compared to physiological and phenotypic responses. However, studies have shown that a constant response between species is the induction of reactive oxygen species (ROS) (Marslin et al. 2017). Studies reporting NPs-elicitation of specialized metabolites often report a reduction in the photosynthetic rate and inhibition of growth. These phytotoxic effects have been linked to the inhibition of Ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubisco) activity and decreased photo-protective capacity of PSII (Jiang et al. 2017a; Wang et al. 2016). When NPs permeate the cells, damage on the photosynthetic apparatus is done because of its accumulation in chloroplasts, at the same time, when they cross the plasma membrane they probably dissociate into ions (rather than stay as intact particles) and bound to NADPH oxidases, causing the production of ROS at the apoplast (Jiang et al. 2017a; Sosan et al. 2016). Besides oxidative burst, it has been reported that NPs also induce reactive nitrogen species (*NO, nitric oxide) (Marslin et al. 2017).
Initial responses also include calcium ion (Ca2+) spikes, Ca2+ flux movements, and upregulation/phosphorylation of mitogen-activated protein kinase (MAPK) cascades that together with ROS production, ultimately lead to the activation of the pathways of specialized metabolites biosynthesis (Anjum et al. 2019; Marslin et al. 2017). As expected, plants exposed to stressful concentrations of NPs have shown to cope with the oxidative stress and lipid peroxidation through the upregulation of enzymatic antioxidants such as superoxide dismutase (SOD), ascorbate peroxidase (APX), glutathione-S-transferase (GST), and catalase (CAT) (Dimkpa et al. 2012; Fu et al. 2014; Mirzajani et al. 2014; Zhao et al. 2012).
Through these findings, the concept of “nano-elicitors” have recently emerged as a novel alternative to stimulate the production of valuable bioactive compounds that might be used as additives in food, cosmetics, and pharmaceutical products. The most widely used nano-elicitors with this purpose are carbon nanotubes, silver, gold, copper, zinc oxide, and titanium dioxide (Anjum et al. 2019). The way to supply NPs to plants can be carried out through foliar spray, directly in the soil or through the nutrient solution applied. Table 5.4 summarizes some published studies in recent years reporting the enhanced production of commercially important specialized metabolites using metallic-, metal oxide-, and carbon related-NPs.
5.2.4 Metals and Salt Metals
Metals, at high concentrations, act as stress agents to plants, therefore, they can induce changes in the secondary metabolism causing an elicitation effect. Exposure of plants to metals, such as Ni, Ag, Fe, and Co, has shown increased production of secondary metabolites in a variety of plants (Zhao et al. 2001). For instance, cadmium (Cd2+) and copper (Cu2+) are known for their toxicity and for not having any value for plants (Das et al. 1997). However, Cd and Cu treatments resulted in enhanced phenolic accumulation on the medicinal plant Gynura procumbens (Ibrahim et al. 2017). Several factors influence the response of plants to metal exposure, mainly depending on the chemical metal species and concentration, the plant species, climate conditions, growth stage, among others (Lajayer et al. 2017).
The use of nonfood crops with the capacity of absorbing and accumulating heavy metals is an alternative for remediation of contaminated environments. It has been shown in certain medicinal and aromatic plants that this practice can lead to the accumulation of secondary metabolites, which can be phytoextracted to obtain high-value compounds (Lajayer et al. 2017). Metabolic changes by the action of heavy metals can lead to inhibition of enzymes involved in the production of photosynthetic pigments, sugars, proteins, and nonprotein thiols (Naik and Al-Khayri 2016; Nasim and Dhir 2010). To date, many studies have shown increases in medicinal plant performance following exposure to heavy metal stress. For example, in a study where garden mint (Mentha crispa L., Lamiaceae) was used for phytoaccumulation of lead (Pb), the chemical composition of the essential oil of the plant was affected by improving the production of carvone, a major component of essential oils (Sá et al. 2015).
Heavy metals have also shown to have a role in stress amelioration through changes in antioxidant balance which often comes hand in hand with increased secondary metabolites. A study subjecting Camellia sinensis (L) plants to drought stress was performed to understand the role of Zn in modulating stress conditions. Results showed decreases in hydrogen peroxide (H2O2) and lipid peroxidation, and at the same time increases in phenolics content and differential expression of antioxidant enzymes, such as superoxide dismutase (SOD), catalase (CAT), peroxidase (POX), polyphenol peroxidase (PPO), glutathione reductase (GR), and ascorbate peroxidase (APX) (Upadhyaya et al. 2013). Similar results were obtained in Brassica napus exposed to cadmium (Cd) stress, exogenous application of low concentrations of selenium (Se) increased the tolerance of plants meanwhile concentrations of ascorbic acid and reduced glutathione were increased (Hasanuzzaman et al. 2012).
Metallic salts have also shown enhanced production of secondary metabolites during in vitro root cultures treatments such as two tropane alkaloids, scopolamine and hyoscyamine, by eliciting with silver nitrate (AgNO3) and cadmium chloride (CdCl2) in Brugmansia candida (Angelova et al. 2006), increases in tanshinone contents using AgNO3 in Perovskia abrotanoides (Zaker et al. 2015) and sesquiterpenoid–defensive compounds using cadmium salts in Datura stramonium (Furze et al. 1991).
5.2.5 Volatile Organic Compounds
Volatile organic compounds (VOCs) are low-molecular weight compounds that are emitted in the atmosphere in vapors or gaseous form. VOCs are produced as secondary metabolites by micro- (bacteria and fungi) and macro-organisms (animals and plants) and play vital roles, such as regulation of physiological processes and inter-organismal communication (Fincheira and Quiroz 2018; Rakshit et al. 2020). Plants can emit VOCs constitutively to attract pollinators and seed dispersers, or in response to a stimulus as a defense against insects or predators, plant-to-plant communication, thermo-tolerance, and environmental stress adaptation (Vivaldo et al. 2017). Plant VOCs can be classified into terpenoids, fatty acid derivatives, phenylpropanoids/benzenoids, and amino acid derivatives (Dudareva et al. 2013). Under-ground VOC’s are emitted by plants through their roots (Rakshit et al. 2020), where they also interact with bacteria and fungi in the rhizosphere zone giving rise to a deep symbiotic plant-microorganisms relation (Dessaux et al. 2016). Microorganisms benefit from root’s exudates while they produce nonvolatile metabolites that affect the plant’s nutrient assimilation and benefits plant growth (Dotaniya and Meena 2015).
A new plant–microbe interaction involving microbial volatile organic compounds (mVOCs) was discovered by (Ryu et al. 2003). In their study, they identified that volatile compounds from Bacillus subtilis act as strong promoters of growth in Arabidopsis thaliana. Since then, studies focusing on mVOCs as potential compounds with practical applications on regulating characteristics of agronomic importance have emerged. Bacterial and fungal volatile compounds may activate defense responses against biotic and abiotic stress, induce systemic resistance, promote growth, and enhance health processes in plants (Kanchiswamy et al. 2015; Piechulla and Degenhardt 2014). There are approximately 1000 mVOCs produced by bacteria and fungi reported in the literature, a few examples include 3-hydroxy-2-butanone (acetoin), 2,3-butanediol, 2-pentylfuran, or dimethylhexadecylmine (Fincheira and Quiroz 2018; Piechulla and Degenhardt 2014).
The idea of using VOCs to elicit secondary metabolites in plants is novel and still very little studied. There are cases of success in the literature to this purpose, which are summarized in Table 5.5, most of them have shown that different volatile compounds from bacterial can increase commercially valued components of essential oils, such as monoterpenes, pulegone, menthone, menthol, limonene, menthyl acetate, terpineol, and eugenol (Banchio et al. 2009; Santoro et al. 2011, 2016; Zhou et al. 2016). These studies are often performed in sterile plastic boxes or petri dishes with divided into two compartments by a physical barrier so that microbial and plant cultures interact without physical contact.
5.2.6 Nutrient Deficiency
The soil provides water and nutrients to plants. Fourteen mineral nutrients are required for plant correct growth and development which are divided into macronutrients (N, P, K, Ca, Mg, and S) and micronutrients (Cl, Fe, B, Mn, Zn Cu, Ni, and Mo). Macronutrients form structural and energy compounds in plants. On the other hand, microelements are related to enzymatic responses. For example, Zn, Fe, Mn, and Cu are components of enzymatic antioxidants, which regulate oxidation processes in the plant (Hajiboland 2012; Nath and Tuteja 2016). Due to the indispensable role of nutrients, plant roots have developed an efficient sensing and signaling system to maintain nutrient homeostasis. Low availability of nutrients in the soil is detected by roots, and in response, chemical signaling and chain reactions are produced. Plants employ signaling players as phytohormones, reactive oxygen species (ROS), sugars, and transcription factors to maintain nutrients homeostasis within the plant (Nath and Tuteja 2016; Isah 2019).
Nutrient deficiency can produce metabolic responses that cause an increased accumulation of secondary metabolites. Natural bioactive compounds are sought in bioproduction processes and improvement of nutritional quality of vegetables and fruits, therefore a nutrient deficiency of specific macro or micronutrients may be an alternative. However, this stress can cause a decrease in crop yields (Hawkesford et al. 2012). To produce secondary metabolites of interest without a considerable loss of growth and biomass, it is necessary to generate eustress in the plant (El-Nakhel et al. 2019). Currently, nutritional eustress is a strategy used in protected production systems, where soilless crops allow greater control of nutrient supply through nutrient solutions.
Nitrogen is a macronutrient constituent of primary metabolites (e.g., protein, peptides, amino acids, and nucleic acids), phytohormones and secondary metabolites. Plants can uptake nitrogen as nitrate and ammonium (mineral form) (Isah 2019). Research has shown an inverse relationship between low nitrogen availability and the synthesis of phenolic compounds. According to this hypothesis, low nitrogen availability increases synthesis of metabolites that contain C, H, and O in their structure. Therefore, terpenes and phenolic compounds synthesis will be favored. On the contrary, metabolites that contain N in its structure such as, alkaloids, nonprotein amino acids, and cyanogenic compounds, will decrease its synthesis (Nath and Tuteja 2016). For example, growing lettuce (Lactuca sativa) increases its content of phenolic compounds and antioxidant capacity in the presence of nitrogen deficiency and drought (Galieni et al. 2015). Table 5.6 shows more examples of the effect on phenolic compounds of experiments where nitrogen deficiency was the stress factor on plants or food with commercial interest.
Phosphorus (P) is a component of molecules such as nucleic acids, lipids and nucleotides with an energy function (ATP and ADP). Plants uptake P in the form of inorganic orthophosphate (Pi, HPO42−, and H2PO4−) which deficiency promotes anthocyanin synthesis (Jezek et al. 2016). Peng et al. (2019) proposed a model of com-modulation (miR399d) and epigenetic modification as a regulatory mechanism of anthocyanin synthesis that depends on the P availability. Sulfur is a structural component of amino acid precursors of secondary metabolites. Therefore, its deficiency negatively affects the biosynthesis of lycopenes and carotenoids (Mohammed et al. 2015). Micronutrient deficiencies are shown to have a negative impact on the synthesis of phenolic compounds and terpenes. Micronutrients such as Cu, Fe, Mo, and Mn, act as factors for the synthesis of secondary metabolites.
5.3 Biotic Stress
Biological stressors are those considered within living organisms (plants or pathogens) including bacteria, insect or herbivores, fungi, phytohormones, and miRNA, among others, that results in biotic stress (Patel and Krishnamurthy 2013). The action mechanism of this factor includes activation or inactivation of enzymes, interaction with receptors, ion channels, stimulation of bioactive compounds, and so forth (Joshi et al. 2019). Some biotic stress factors and their role in the synthesis of secondary metabolites in plants are described below.
5.3.1 Bacteria and Viruses
Plants are exposed to interactions with other living things. The interaction between microorganisms and plants can have positive effects. Microorganism colonization (pathogens and non-pathogens) triggers the resistance mechanism of the plant, conferring resistance against other stressors (Nejat and Mantri 2017; Choudhary et al. 2016). Nonpathogenic microorganisms act as plant biostimulants. A plant biostimulant is defined as any substance or microorganism applied to plants in order to improve nutritional efficiency, tolerance to biotic and abiotic stress, and quality (Van Oosten et al. 2017; Du Jardin 2015). Arbuscular mycorrhizal fungi, Trichoderma, and plant growth-promoting rhizobacteria are biostimulant microorganisms used in crops.
Microorganisms can confer a certain degree of tolerance against abiotic stress conditions. Colonized plants produce a wide range of enzymes and metabolites that allow them to generate tolerance to stress (Miliute et al. 2015). Some genera of bacteria like Rhizobium, Bacillus, Pseudomonas, Pantoea, Paenibacillus, Burkholderia, Achromobacter, Azospirillum, Microbacterium, Methylobacterium, variovorax, Enterobacter, have been shown to induce tolerance to abiotic stress (Choudhary et al. 2016; Naveed et al. 2014; Gururani et al. 2013). Tolerance generated by pathogen attack can induce resistance to abiotic stress factors. The biochemical response generated by the attack of the pathogen is similar to the response generated by abiotic factors. Plants attacked by Verticillium dahliae (pathogenic fungus) develop tolerance to drought due to the formation of xylem but reducing the growth rate (Tani et al. 2018).
Viruses are considered symbiotes. They can behave as pathogens or mutualists depending on the environmental conditions where the host is (Roossinck 2015). Research suggests that the mutualistic behavior of a virus occurs when the titer virus is low and the environmental disturbance is low (Bao and Roossinck 2013). Plant viruses can have a positive effect like other pathogens. The presence of the virus in the plant can increase its ability to cope with biotic and abiotic stress factors because of the activation of the plant defense system. Metabolomic studies in infected plants have shown a significant increase in the quantity and diversity of secondary metabolites. This metabolic effect allows the plant to cope with the stress caused by the infection, as well as other stressors present in the environment. For example, Sade et al. (2015), reported a significant impact on the metabolome in tomato plants infected with Tomato yellow leaf curl virus (TYLCV) where resistant and susceptible cultivars showed a major expression of the phenylpropanoid pathway which is related to the production of antioxidant compounds, among others. In the same research, the expression in resistant cultivars was more significant in terms of the production of flavonoids and other antioxidants. On the other hand, rice plants infected with Brome mosaic virus (BMV) and beet plants (Beta vulgaris) infected with Cucumber mosaic virus (CMV) increased the accumulation of osmoprotectants and antioxidant compounds, conferring drought tolerance to both crops (Xu et al. 2008).
5.3.2 Fungi
Plants have a strong symbiosis relationship with some fungi and bacteria present in the substrate where they are grown. These microorganisms, endophytes or exogenous, can induce eustress to the crop, increasing the production of specialized metabolites, e.g.,Aspergillus sp. applied as an elicitor in Artemisia annua L. callus culture, enhanced the production of artemisinin, an endoperoxide sesquiterpene lactone and an effective antimalarial agent (Yuliani et al. 2018). Soil-borne beneficial microbes have shown a protecting potential against pathogens and herbivores via the elicitation of plant responses e.g. plant growth-promoting fungi (PGPF) and arbuscular mycorrhizal fungi (AMF) (Pappas et al. 2018). Fungal elicitation (including yeas extract) is one of the most used to enhance the production of secondary metabolites (Singh et al. 2018).
Fungi with a beneficial effect on plant development that associate to plant roots are called PGPF and are considered the first prevention mechanism in the pathogen infection. Plants need to detect PGPFs and take advantage of the presence of microbe-associated molecular patterns (MAMPs) that can be recognized by pattern recognition receptors (PRRs). PGPFs can stimulate the defense system of plants involving the modification of cell walls by the accumulation of lignin, callose, phenols, etc., preventing the growth and proliferation of pathogens. In addition, elicitors such as chitin, chitosan, and β-glucan that are part of the fungal cell wall have been researched (Naziya et al. 2020; Fesel and Zuccaro 2016; Li et al. 2016).
Recent studies have been performed in different plants, e.g., fungal elicitation in Leguminosae increased the accumulation of isoflavonoids and stilbenoids (Araya-Cloutier et al. 2017). Moola and Diana (2019) used Aspergillus niger, Penicillium notatum, and Rhizopus oligosporus as elicitors on hairy root culture of Beta vulgaris to enhance betalain synthesis. Trichoderma spp. is one of the most widely used microorganisms as a pathogen biocontrol agent and elicitor. In addition, this fungus has been shown to colonize roots and can also induce systemic resistance (ISR), which favors plant growth, increases nutrient availability and enhances disease resistance. The mechanisms employed by Trichoderma spp. are the modulation of plant hormonal mechanisms and the production of secondary metabolites (Nandini et al. 2020; Guzmán-Guzmán et al. 2019; Silva et al. 2019; Martínez‐Medina et al. 2017). PGPFs can be used as bio-fertilizers, improving the quality and quantity of products, and reducing the contamination of the agricultural environment by lowering the use of chemical fertilizers (Pereira et al. 2019; Zhou et al. 2018). Table 5.7 shows some of the fungi used to induce the production of secondary metabolites in plants.
5.3.3 Phytohormones
Phytohormones are molecules synthesized by defined organs that regulate plant growth and have a prominent impact on plant metabolism. Additionally, they play a vital role in the stimulation of response mechanisms of plant defense against stress. Auxins, gibberellins, cytokinins (CK), abscisic acid (ABA), jasmonates (jasmonic acid (JA), methyl jasmonate (MeJA)), salicylic acid (SA), brassinosteroids, strigolactones, cinnamic acid (CA), among others, are examples of phytohormones. Auxins and the auxin índole-3-acetic acid (IAA) act as promoters of growth and developmental events in plants (cell división, elongation, and differentiation). CKs are involved in the maintaining of cellular proliferation, differentiation, and prevention of senescence. ABA has an important role in the plant response to stress and adaptation. Gibberellic acid (GA) is a plant growth regulator and has a vital role in seed dormancy, formation of floral organs, and lateral shoot growth (Egamberdieva et al. 2017). SA has an important role in plant stress tolerance through modulation of antioxidative enzyme activities. JA is a lipid-derived compound synthesized via the octadecanoid pathway and is important in the development, structure, and flowering of plants (Złotek et al. 2020). CA is a bioprecursor of podophyllotoxin and a huge number of plant substances, including tannins, flavonoids, etc. (Kašparová et al. 2018).
Currently, the use of phytohormones in agriculture and research has been implemented as a strategy to induce the production of secondary metabolites (Egamberdieva et al. 2017). The exogenous application of phytohormones by spraying has resulted in increased production of various significant bioactive compounds in plants (Akram et al. 2020, Wang et al. 2018a). For example, Garcia-Ibañez et al. (2019), applied MeJA, SA, and SA + MeJA to Bimi® plants resulting in a differentiated response to each elicitor, all treatments showed an increased content of GLs in leaves and inflorescences. Table 5.8 shows the result of plant elicitation with phytohormones in the production of bioactive compounds of recent studies.
5.3.4 miRNA
MicroRNAs (miRNAs - length of 18 to 28 nucleotides) are noncoding RNAs. The main function of these molecules is to participate in gene expression and regulation at the post-transcriptional level by degrading mRNA or at the translational level by blocking protein biosynthesis at different stages (Fig. 5.1), resulting in regulation of plant development, metabolism, and response to biotic or abiotic stress (Tripathi et al. 2019). In plants miRNAs genes are transcribed by RNA polymerase II producing primary miRNA (Pri-miRNA) which are important for the regulation of genome integrity, primary and secondary metabolism, development, signal transduction, signaling pathways, homeostasis, innate immunity, and environmental stress responses (Vargas-Hernández et al. 2019; Wang et al. 2018b; Samad et al. 2019).

Clasification of plant noncoding RNAs (ncRNAs). Linear noncoding RNAs (Linear ncRNAs), circular noncoding RNAs (cncRNAs), HouseKeeping noncoding RNAs (hk ncRNAs), Regulatory noncoding RNAs (Regulatory ncRNAs), long noncoding RNAs (lncRNAs), small RNAs (sRNAs), small interfering RNAs (siRNAs), micro RNAs (miRNAs)
The regulation mechanism of gene expression by miRNAs is via RNA interference. These small molecules are transcribed as longer precursors in the nucleus and are then further processed into their mature forms (Wang et al. 2018b; Samad et al. 2017). Most plant miRNA genes are located inside intergenic regions between two adjacent genes and are transcriptionally regulated by their promoters and terminators (Hossain et al. 2019). miRNAs were reported to be involved in plant secondary metabolite regulation such as terpenoid, phenolic, fatty acid, flavonoids, phenolic, and nitrogen-containing compound biosynthesis (Liu et al. 2017; Samad et al. 2019).
There is a very close relationship between miRNAs and the transcription factors (TFs), either to be switched “on” or switched “off”. Current research trends have focused on knowing the regulation of miRNA in response to environmental stress and how they interact with transcription factors (Fig. 5.2) (Samad et al. 2019).

Stress mechanism and interaction between transcription factors and miRNAs
The main goal of functional studies on miRNAs has been to understand the biological processes in which the miRNAs are involved. Different technologies have been developed to characterize the function and action mechanisms of these small molecules in various plant materials (Liu et al. 2017). Research on the regulation of miRNAs in Taxus callus cells has been conducted, concluding that miRNAs are capable of direct regulation of secondary metabolism by modulating transcriptional factors (Chen et al. 2020). The expresión level of miRNAs is mostly governed by temperature and radiation (Tripathi et al. 2019). Studies have demonstrated that miRNAs may act as master regulators of flavonoid biosynthesis, e.g., miR156-SPL9 network directly influences anthocyanin production, miR163 targets S-adenosyl-Met-dependent methyltransferases that methylates secondary metabolites and signaling molecules, and miR397 regulates lignin biosynthesis in Arabidopsis and Populus spp (Sharma et al. 2016).
Studies carried out on different plant species indicate that the cellular levels of miRNA have a high regulation control for optimal spatiotemporal regulation of target genes, adding an additional layer of complexity to the signaling processes (Gupta et al. 2017; Sharma et al. 2016). These results suggest that a strategy to induce the production of secondary metabolites may be the use of miRNAs that promote the expression of genes involved in plant biosynthetic pathways.
5.4 Future Perspectives
The use of AE stimuli as a new elicitor in plants has huge sustainable potential. However, this “environmentally friendly” agricultural technology is still under discussion and more studies should be encouraged. Currently, the optimal sound therapy is not known since the effect could differ depending on several factors such as plant model, amplitude, frequency or time and duration of treatment, and application distance among others that are not explained in most studies (Alvarado et al. 2019).
PEF and MF in food, horticulture, and biotechnology asthe postharvest processes have increased substantially during the last few years (Xi-ran and Ting 2017; Gilani et al. 2017; Rusakova et al. 2017). Generally, it can be noted that the application of PEF and MF as pretreatment is a novel method to improve plant development due to a higher production of ROS and an associated activation of antioxidant defense systems, such as POD, SOD, and CAT. However, the effect of electromagnetic sources is plant and treatment specific. It is difficult to have a strong conclusion in the analyzed experiments in this chapter since the experimental details often were incomplete, including an insufficient description of equipment and treatment conditions. It is recommended to consider the description of parameters, such as electric field strength (V/m), frequency (Hz), magnetic flux density (T), electric current (A), PPFD, distance of light source to plant, light decay among the experimental source, photoperiod, light wavelength, the duration of application, and plant or food complete physical description (Dannehl 2018; Alvarado et al. 2019).
Nanoparticles in agriculture have shown benefits related to physiological and growth parameters. Moreover, the new knowledge related to its effect on secondary metabolism has opened a field of possibilities in the production of bioactive compounds with high commercial value using NPs. However, as with many elicitors, there is still a lack of knowledge about the type and size of nanoparticle and appropriate concentrations to use depending on the species of interest. Furthermore, a large part of the existing nanoparticles has not yet been studied and it is also necessary to continue generating knowledge to understand the molecular mechanisms of elicitation with NPs.
Metal ions have been proposed as suitable elicitors of secondary metabolism in cell cultures (Rudrappa et al. 2004), since they can make more efficient the tissue culture techniques during the obtention of valuable secondary metabolites. Many studies have proposed the use of different chemical metal species to enhance bioactive compounds in plants. This tool is very attractive in the sense that, in addition to producing these types of compounds, it can become an environmental remediation technique. Also, the idea of producing specialized metabolites for later extraction eliminates the risks associated with the potential health risks to the consumers.
Among the study of new agricultural tools in the last decade, VOCs stands out for being considered as an eco-friendly, cheap, and effective alternative. Even genetically modified plants with altered VOC emission and synthetic formulations of plant VOCs are been developed as a promising technology for agriculture and horticulture (Rakshit et al. 2020). However, there are still many unknowns, especially about the mode of action of VOCs and about the molecular and biochemical mechanisms related to eliciting responses of interest in plants.
The application of phytohormones in different phenological stages of plants, including in the postharvest stage, induces the production of secondary metabolites and is an effective strategy that uses defense mechanisms to mimic stress caused by various environmental factors. This method can be used at the agronomic level to improve the quality of plant products by increasing the content of bioactive molecules. Defense plant response due to different types of stress depends on the type of crosstalk (positive or negative) between the hormone signaling pathways rather than on the individual contributions of each hormone (Verma et al. 2016). This suggests that for future perspectives it should be taken into account that the results of elicitation will depend on the synergy of the used phytohormones, the concentration, the application conditions, and the type of cultivation. For example, there is a balance between SA and JA to regulate biotic stress in tomato (Verma et al. 2016). In the same way, SA and gibberellins have been used as elicitor and biostimulant to enhance the production of steviol glycosides in stevia, producing tall plants with a greater number of leaves and a larger stem diameter (Vazquez-Hernandez et al. 2019).
Endophytic microbes have been shown to be able to promote plant growth, induce tolerance and production of bioactive compounds (Lata et al. 2018). Endophytic microbes generally reside in tissues and plants without causing symptoms. However, they activate plant defense system and induce secondary metabolites production with potential pharmaceutical use (Jalgaonwala and Mahajan 2014). Therefore, the inoculation of plants with this type of microorganism is a strategy that can be applied to induce and/or increase the production of natural bioactive compounds in plants of commercial interest.
Fungi elicitation is a popular practice among horticultural producers due to the demonstrated increase in crop yield and the production of bioactive compounds. The ability of plant roots to uptake nutrients from the substrate is enhanced by this type of elicitation, with a positive influence on phytohormone production and gene expression reprogramming. Fungi are used as biocontrol treatments, bioremediation agents, and as biostimulants, which can contribute to the development of sustainable agriculture. In the same way, mineral nutrients play an important role in the growth, development, and yield of crops (Nath and Tuteja 2016). Nutrient management is a common agricultural practice, especially in soilless cultivation and controlled systems in order to guarantee high crop yield. On the other hand, consumers demand natural products with high nutritional quality. Both objectives are feasible to be induced by the nutrition of the crops. Therefore, the management of nutrients during the production cycle allows a balance between growth and accumulation of bioactive compounds.
The use of specific miRNAs to enhance the production of metabolites is a novel technique that is still under research and discussion among the scientific community. Research has focused on the identification of the regulatory mechanisms of these molecules that will lead to the design of strategies for direct manipulation, identification, and understanding of the spatial and temporal expression scheme of miRNAs. Innovative tools (e.g., bioinformatics) can be used to predict, by means of algorithms, the modulation of miRNA molecules which are part of a very complex regulatory network of secondary metabolites. Research in this area could help to understand how biosynthetic pathways are modified by these small molecules that have particular target genes and that can influence metabolic plant bioengineering, generating technology to induce the production of secondary metabolites.
References
Abdollahi F, Amiri H, Niknam V, Ghanati F, Mahdigholi K (2019) Effects of static magnetic fields on the antioxidant system of almond seeds. Russ J Plant Physl 66(2):299–307
Agathokleous E, Kitao M, Calabrese EJ (2019) Hormesis: a compelling platform for sophisticated plant science. Trends Plant Sci 24(4):318–327
Ahmad N, Jones RW (1979) Glycinebetaine, proline and inorganic ion levels in barley seedlings following transient stress. Plant Sci Lett 15(3):231–237
Akhgari A, Laakso I, Maaheimo H, Choi YH, Seppänen-Laakso T, Oksman-Caldentey K-M, Rischer H (2019) Methyljasmonate elicitation increases terpenoid indole alkaloid accumulation in rhazya stricta hairy root cultures. Plants 8(12):534
Akram W, Saeed T, Ahmad A, Yasin NA, Akbar M, Khan WU, Ahmed S, Guo J, Luo W, Wu T (2020) Liquiritin elicitation can increase the content of medicinally important glucosinolates and phenolic compounds in Chinese kale plants. J Sci Food Agric 100(4):1616–1624
Akula R, Ravishankar GA (2011) Influence of abiotic stress signals on secondary metabolites in plants. Plant Signal Behav 6(11):1720–1731
Alokam S, Chinnappa C, Reid DM (2002) Red/far-red light mediated stem elongation and anthocyanin accumulation in Stellaria longipes: differential response of alpine and prairie ecotypes. Can J Bot 80(1):72–81
Alvarado AM, Aguirre-Becerra H, Vázquez-Hernández MC, Magaña-Lopez E, Parola-Contreras I, Caicedo-Lopez LH, Contreras-Medina LM, Garcia-Trejo JF, Guevara-Gonzalez RG, Feregrino-Perez AA (2019) Influence of elicitors and eustressors on the production of plant secondary metabolites. In: Natural bio-active compounds. Springer, pp 333–388
Ampofo JO, Ngadi M (2020) Ultrasonic assisted phenolic elicitation and antioxidant potential of common bean (Phaseolus vulgaris) sprouts. Ultrason Sonochem 64:104974
Andini S, Dekker P, Gruppen H, Araya-Cloutier C, Vincken J-P (2019) Modulation of glucosinolate composition in brassicaceae seeds by germination and fungal elicitation. J Agric Food Chem 67(46):12770–12779
Angelova Z, Georgiev S, Roos W (2006) Elicitation of plants. Biotechnol Biotechnol Equip 20(2):72–83
Anjum S, Anjum I, Hano C, Kousar S (2019) Advances in nanomaterials as novel elicitors of pharmacologically active plant specialized metabolites: current status and future outlooks. RSC Adv 9(69):40404–40423
Appel HM, Cocroft R (2014) Plants respond to leaf vibrations caused by insect herbivore chewing. Oecologia 175(4):1257–1266
Araya-Cloutier C, den Besten HM, Aisyah S, Gruppen H, Vincken J-P (2017) The position of prenylation of isoflavonoids and stilbenoids from legumes (Fabaceae) modulates the antimicrobial activity against Gram positive pathogens. Food Chem 226:193–201
Arora S, Sharma P, Kumar S, Nayan R, Khanna P, Zaidi M (2012) Gold-nanoparticle induced enhancement in growth and seed yield of Brassica juncea. J Plant Growth Regul 66(3):303–310
Azad MOK, Kim WW, Park CH, Cho DH (2018) Effect of artificial LED light and far infrared irradiation on phenolic compound, isoflavones and antioxidant capacity in soybean (Glycine max L.) sprout. Foods 7(10):174
Azimi N, Majd A, Nejadsattari T, Ghanati F, Arbabian S (2018) Effects of magnetically treated water on physiological characteristics of Lens culinaris L. Iran J Sci Technol A 42(2):331–337
Azizi SMY, Sarghein SH, Majd A, Peyvandi M (2019) The effects of the electromagnetic fields on the biochemical components, enzymatic and non-enzymatic antioxidant systems of tea Camellia sinensis L. Physiol Mol Biol Plants 25(6):1445–1456
Balbus JM, Maynard AD, Colvin VL, Castranova V, Daston GP, Denison RA, Dreher KL, Goering PL, Goldberg AM, Kulinowski KM (2007) Meeting report: hazard assessment for nanoparticles—report from an interdisciplinary workshop. Environ Health Perspect 115(11):1654–1659
Banchio E, Xie X, Zhang H, Pare PW (2009) Soil bacteria elevate essential oil accumulation and emissions in sweet basil. J Agric Food Chem 57(2):653–657
Bantis F, Smirnakou S, Ouzounis T, Koukounaras A, Ntagkas N, Radoglou K (2018) Current status and recent achievements in the field of horticulture with the use of light-emitting diodes (LEDs). Sci Hortic-Amsterdam 235:437–451
Bao X, Roossinck MJ (2013) A life history view of mutualistic viral symbioses: quantity or quality for cooperation? Curr Opin Microbiol 16(4):514–518
Barba FJ, Galanakis CM, Esteve MJ, Frigola A, Vorobiev E (2015) Potential use of pulsed electric technologies and ultrasounds to improve the recovery of high-added value compounds from blackberries. J Food Eng 167:38–44
Becker C, Urlić B, Špika MJ, Kläring H-P, Krumbein A, Baldermann S, Ban SG, Perica S, Schwarz D (2015) Nitrogen limited red and green leaf lettuce accumulate flavonoid glycosides, caffeic acid derivatives, and sucrose while losing chlorophylls, β-carotene and xanthophylls. PloS one 10 (11)
Burgie ES, Bussell AN, Walker JM, Dubiel K, Vierstra RD (2014) Crystal structure of the photosensing module from a red/far-red light-absorbing plant phytochrome. Proc Natl Acad Sci U S A 111(28):10179–10184
Cai Z, Kastell A, Speiser C, Smetanska I (2013) Enhanced resveratrol production in Vitis vinifera cell suspension cultures by heavy metals without loss of cell viability. Appl Biochem Biotechnol 171(2):330–340
Calabrese EJ (2004) Hormesis: a revolution in toxicology, risk assessment and medicine. EMBO Rep 5(S1):S37–S40
Chen Y, Zhang M, Jin X, Tao H, Wang Y, Peng B, Fu C, Yu L (2020) Transcriptional reprogramming strategies and miRNA-mediated regulation networks of Taxus media induced into callus cells from tissues. BMC Genomics 21(1):1–15
Chepets A, Morozova Z, Tkacheva N (1985) Wheat and sorghum yield and grain quality as affected by magnetic field.
Cheynier V, Comte G, Davies KM, Lattanzio V, Martens S (2013) Plant phenolics: recent advances on their biosynthesis, genetics, and ecophysiology. Plant Physiol Biochem 72:1–20
Choudhary DK, Kasotia A, Jain S, Vaishnav A, Kumari S, Sharma KP, Varma A (2016) Bacterial-mediated tolerance and resistance to plants under abiotic and biotic stresses. J Plant Growth Regul 35(1):276–300
Cox A, Venkatachalam P, Sahi S, Sharma N (2016) Silver and titanium dioxide nanoparticle toxicity in plants: a review of current research. Plant Physiol Biochem 107:147–163
Dannehl D (2018) Effects of electricity on plant responses. Sci Hortic-Amsterdam 234:382–392
Das P, Samantaray S, Rout G (1997) Studies on cadmium toxicity in plants: a review. Environ Pollut 98(1):29–36
Denoya GI, Pataro G, Ferrari G (2020) Effects of postharvest pulsed light treatments on the quality and antioxidant properties of persimmons during storage. Postharvest Biol Technol 160:111055
Dessaux Y, Grandclément C, Faure D (2016) Engineering the rhizosphere. Trends Plant Sci 21(3):266–278
Dhiman SK, Galland P (2018) Effects of weak static magnetic fields on the gene expression of seedlings of Arabidopsis thaliana. J Plant Physiol 231:9–18
Dimkpa CO, McLean JE, Latta DE, Manangón E, Britt DW, Johnson WP, Boyanov MI, Anderson AJ (2012) CuO and ZnO nanoparticles: phytotoxicity, metal speciation, and induction of oxidative stress in sand-grown wheat. J Nanopart Res 14(9):1125
Dotaniya M, Meena V (2015) Rhizosphere effect on nutrient availability in soil and its uptake by plants: a review. P NATL A SCI INDIA B 85(1):1–12
Dou H, Niu G, Gu M (2019) Pre-harvest UV-B radiation and photosynthetic photon flux density interactively affect plant photosynthesis, growth, and secondary metabolites accumulation in basil (Ocimum basilicum) plants. Agronomy 9(8):434
Du Jardin P (2015) Plant biostimulants: definition, concept, main categories and regulation. Sci Hortic-Amsterdam 196:3–14
Dudareva N, Klempien A, Muhlemann JK, Kaplan I (2013) Biosynthesis, function and metabolic engineering of plant volatile organic compounds. New Phytol 198(1):16–32
Egamberdieva D, Wirth SJ, Alqarawi AA, Abd_Allah EF, Hashem A (2017) Phytohormones and beneficial microbes: essential components for plants to balance stress and fitness. Front Microbiol 8:2104
El Kantar S, Boussetta N, Lebovka N, Foucart F, Rajha HN, Maroun RG, Louka N, Vorobiev E (2018) Pulsed electric field treatment of citrus fruits: improvement of juice and polyphenols extraction. Innov Food Sci Emerg 46:153–161
El-Nakhel C, Pannico A, Kyriacou MC, Giordano M, De Pascale S, Rouphael Y (2019) Macronutrient deprivation eustress elicits differential secondary metabolites in red and green-pigmented butterhead lettuce grown in a closed soilless system. J Sci Food Agric 99(15):6962–6972
Faiola C, Taipale D (2020) Impact of insect herbivory on plant stress volatile emissions from trees: a synthesis of quantitative measurements and recommendations for future research. Atmos Environ X 5:100060
Fazal H, Abbasi BH, Ahmad N, Ali M, Shujait Ali S, Khan A, Wei D-Q (2019) Sustainable production of biomass and industrially important secondary metabolites in cell cultures of selfheal (Prunella vulgaris L.) elicited by silver and gold nanoparticles. Artif Cells Nanomed Biotechnol 47(1):2553–2561
Feregrino-Perez AA, Magaña-López E, Guzmán C, Esquivel K (2018) A general overview of the benefits and possible negative effects of the nanotechnology in horticulture. Sci Hortic-Amsterdam 238:126–137
Fesel PH, Zuccaro A (2016) β-glucan: Crucial component of the fungal cell wall and elusive MAMP in plants. Fungal Genet Biol 90:53–60
Fincheira P, Quiroz A (2018) Microbial volatiles as plant growth inducers. Microbiol Res 208:63–75
Fu PP, Xia Q, Hwang H-M, Ray PC, Yu H (2014) Mechanisms of nanotoxicity: generation of reactive oxygen species. J Food Drug Anal 22(1):64–75
Furze JM, Rhodes MJ, Parr AJ, Robins RJ, Withehead IM, Threlfall DR (1991) Abiotic factors elicit sesquiterpenoid phytoalexin production but not alkaloid production in transformed root cultures of Datura stramonium. Plant Cell Rep 10(3):111–114
Gachovska TK, Armyanov N, Palov I (2015) Pre-planting pulsed electric field treatment of potato tubers. In: 2015 IEEE Canada international humanitarian technology conference (IHTC2015). IEEE, pp 1–4
Galieni A, Di Mattia C, De Gregorio M, Speca S, Mastrocola D, Pisante M, Stagnari F (2015) Effects of nutrient deficiency and abiotic environmental stresses on yield, phenolic compounds and antiradical activity in lettuce (Lactuca sativa L.). Sci Hortic-Amsterdam 187:93–101
Garcia-Ibañez P, Moreno DA, Nuñez-Gomez V, Agudelo A, Carvajal M (2019) The use of elicitation in the cultivation of Bimi® for food and ingredients. J Sci Food Agric 100(5):2099–2109
Ghorbanpour M, Khavazi K, Hatami M (2014) Chemical compositions and antimicrobial activity of Salvia officinalis L. essential oil under rhizobacteria (Pseudomonas fluorescens and Putida) inoculation. Eur J Soil Biol
Gilani A, Kermanshahi H, Gholizadeh M, Golian A (2017) Agricultural water managenemt through magnetizatio of irrigation and drinking water: a review. J Aridland Agric 3:23–27
Gupta OP, Karkute SG, Banerjee S, Meena NL, Dahuja A (2017) Contemporary understanding of miRNA-based regulation of secondary metabolites biosynthesis in plants. Front Plant Sci 8:374
Gururani MA, Upadhyaya CP, Baskar V, Venkatesh J, Nookaraju A, Park SW (2013) Plant growth-promoting rhizobacteria enhance abiotic stress tolerance in Solanum tuberosum through inducing changes in the expression of ROS-scavenging enzymes and improved photosynthetic performance. J Plant Growth Regul 32(2):245–258
Gutiérrez-Gamboa G, Garde-Cerdán T, Gonzalo-Diago A, Moreno-Simunovic Y, Martínez-Gil AM (2017) Effect of different foliar nitrogen applications on the must amino acids and glutathione composition in Cabernet Sauvignon vineyard. LWT 75:147–154
Guzmán-Guzmán P, Porras-Troncoso MD, Olmedo-Monfil V, Herrera-Estrella A (2019) Trichoderma species: versatile plant symbionts. Phytopathology 109(1):6–16
Häder D-P, Williamson CE, Wängberg S-Å, Rautio M, Rose KC, Gao K, Helbling EW, Sinha RP, Worrest R (2015) Effects of UV radiation on aquatic ecosystems and interactions with other environmental factors. Photoch Photobio SCI 14(1):108–126
Hajiboland R (2012) Effect of micronutrient deficiencies on plants stress responses. In: Abiotic stress responses in plants. Springer, pp 283–329
Halder M, Sarkar S, Jha S (2019) Elicitation: A biotechnological tool for enhanced production of secondary metabolites in hairy root cultures. Eng Life Sci 19(12):880–895
Hasan M, Alharby H, Hajar A, Hakeem K, Alzahrani Y, Arabia S (2018) Effects of magnetized water on phenolic compounds, lipid peroxidation and antioxidant activity of moringa species under drought stress. J Anim Plant Sci 28(3):1–6
Hasanuzzaman M, Hossain MA, Fujita M (2012) Exogenous selenium pretreatment protects rapeseed seedlings from cadmium-induced oxidative stress by upregulating antioxidant defense and methylglyoxal detoxification systems. Biol Trace Elem Res 149(2):248–261
Hassanien RH, Hou T, Li Y, Li B (2014) Advances in effects of sound waves on plants. J Integr Agric 13(2):335–348
Hawkesford M, Horst W, Kichey T, Lambers H, Schjoerring J, Møller IS, White P (2012) Functions of macronutrients. In: Marschner's mineral nutrition of higher plants. Elsevier, pp 135–189
He R, Xi G, Liu K (2017) Alleviating effect of extremely low frequency pulsed electric field on drought damage of maize seedling roots. J Lumin 188:441–447
Hendrawan Y, Sabrinauly S, Hawa LC, Argo BD, Rachmawati M (2019) Analysis of the phenol and flavonoid content from basil leaves (Ocimum Americanum L) extract using pulsed electric field (PEF) pre-treatment. Agric Eng Int CIGR J 21(2):149–158
Hideg É, Strid Å (2017) The effects of UV-B on the biochemistry and metabolism in plants. In: CABI (ed) UV-B radiation and plant life. Molecular biology to ecology. pp 90–110
Hossain MS, Hoang NTH, Yan Z, Tóth K, Meyers BC, Stacey G (2019) Characterization of the spatial and temporal expression of two soybean miRNAs identifies SCL6 as a novel regulator of soybean nodulation. Front Plant Sci 10:475
Ibrahim MH, Chee Kong Y, Zain M, Amalina N (2017) Effect of cadmium and copper exposure on growth, secondary metabolites and antioxidant activity in the medicinal plant Sambung Nyawa (Gynura procumbens (L.) Merr). Molecules 22(10):1623
Isah T (2019) Stress and defense responses in plant secondary metabolites production. Biol Res 52(1):39
Jalgaonwala R, Mahajan R (2014) A review on microbial endophytes from plants: a treasure search for biologically active metabolites. Glob J Res Med Plants Indig Med 3(6):263
Janositz A, Knorr D (2010) Microscopic visualization of pulsed electric field induced changes on plant cellular level. Innov Food Sci Emerg 11(4):592–597
Jeong M-J, Cho J-I, Park S-H, Kim K-H, Lee SK, Kwon T-R, Park S-C, Siddiqui ZS (2014) Sound frequencies induce drought tolerance in rice plant. Pak J Bot 46:2015–2020
Jezek M, Zörb C, Merkt N, Mühling K-H, Geilfus C-M (2016) The Relationship between Plant nutrition and Anthocyanins and their significance for agriculture. In: Magnesium deficiency in maize and effectiveness of nutrient supply through MgSO4 leaf-application. Kiel, pp 78–118
Jiang HS, Yin LY, Ren NN, Zhao ST, Li Z, Zhi Y, Shao H, Li W, Gontero B (2017) Silver nanoparticles induced reactive oxygen species via photosynthetic energy transport imbalance in an aquatic plant. Nanotoxicology 11(2):157–167
Jiang N, Grundy S, Bian Z, Lu C (2017) Investigation of LED light effects on plant growth in improved protected horticulture system. Greensys 2017—international symposium on new technologies for environment control. Energy-Saving and Crop Production in Greenhouse and Plant Factory, Beijing, China, pp 20–24
Jiang Y, Hua Z, Zhao Y, Liu Q, Wang F, Zhang Q (2014) The effect of carbon nanotubes on rice seed germination and root growth. In: Proceedings of the 2012 international conference on applied biotechnology (ICAB 2012). Springer, pp 1207–1212
Joshi R, Paul M, Kumar A, Pandey D (2019) Role of calreticulin in biotic and abiotic stress signalling and tolerance mechanisms in plants. Gene 714:144004
Kanchiswamy CN, Malnoy M, Maffei ME (2015) Bioprospecting bacterial and fungal volatiles for sustainable agriculture. Trends Plant Sci 20(4):206–211
Kašparová M, Pilařová P, Tůmová L, Siatka T (2018) Effect of precursor and phytohormones on podophyllotoxin production in juniperus virginiana suspension cultures. Nat Prod Commun 13(11):1934578X1801301129
Kentish S, Ashokkumar M (2011) The physical and chemical effects of ultrasound. In: Ultrasound technologies for food and bioprocessing. Springer, pp 1–12
Khodakovskaya MV, Kim BS, Kim JN, Alimohammadi M, Dervishi E, Mustafa T, Cernigla CE (2013) Carbon nanotubes as plant growth regulators: effects on tomato growth, reproductive system, and soil microbial community. Small 9(1):115–123
Khot LR, Sankaran S, Maja JM, Ehsani R, Schuster EW (2012) Applications of nanomaterials in agricultural production and crop protection: a review. Crop Protect 35:64–70
Kole C, Kole P, Randunu KM, Choudhary P, Podila R, Ke PC, Rao AM, Marcus RK (2013) Nanobiotechnology can boost crop production and quality: first evidence from increased plant biomass, fruit yield and phytomedicine content in bitter melon (Momordica charantia). BMC Biotechnol 13(1):37
Koshel RJ (2004) Illumination engineering. Opt Eng 43(7):1478–1480
Kováčik J, Klejdus B (2014) Induction of phenolic metabolites and physiological changes in chamomile plants in relation to nitrogen nutrition. Food Chem 142:334–341
Kranner I, Beckett RP, Wornik S, Zorn M, Pfeifhofer HW (2002) Revival of a resurrection plant correlates with its antioxidant status. Plant J 31(1):13–24
Kranner I, Minibayeva FV, Beckett RP, Seal CE (2010) What is stress? Concepts, definitions and applications in seed science. New Phytol 188(3):655–673
Lahiani MH, Chen J, Irin F, Puretzky AA, Green MJ, Khodakovskaya MV (2015) Interaction of carbon nanohorns with plants: uptake and biological effects. Carbon 81:607–619
Lajayer BA, Ghorbanpour M, Nikabadi S (2017) Heavy metals in contaminated environment: destiny of secondary metabolite biosynthesis, oxidative status and phytoextraction in medicinal plants. Ecotoxicol Environ Saf 145:377–390
Larner VS, Franklin KA, Whitelam GC (2018) Photoreceptors and light signalling pathways in plants. Annual Plant Rev Online 107–131
Lata R, Chowdhury S, Gond SK, White JF Jr (2018) Induction of abiotic stress tolerance in plants by endophytic microbes. Lett Appl Microbiol 66(4):268–276
Li P, Linhardt RJ, Cao Z (2016) Structural characterization of oligochitosan elicitor from Fusarium sambucinum and its elicitation of defensive responses in Zanthoxylum bungeanum. Int J Mol Sci 17(12):2076
Lichtenthaler HK (1996) Vegetation stress: an introduction to the stress concept in plants. J Plant Physiol 148(1–2):4–14
Lilia C, Abdelkader A, Karima A-KA, Tarek B (2018) The effect of ultrasound pre-treatment on the yield, chemical composition and antioxidant activity of essential oil from wild Lavandula stoechas L. J Essent Oil-Bear 21(1):253–263
Liu J, Yuan Y, Wang Y, Jiang C, Chen T, Zhu F, Zhao Y, Zhou J, Huang L (2017) Regulation of fatty acid and flavonoid biosynthesis by miRNAs in Lonicera japonica. RSC Adv 7(56):35426–35437
Liu W, Liu C, Yang C, Wang L, Li S (2010) Effect of grape genotype and tissue type on callus growth and production of resveratrols and their piceids after UV-C irradiation. Food Chem 122(3):475–481
Lu C, Ding J, Park HK, Feng H (2020) High intensity ultrasound as a physical elicitor affects secondary metabolites and antioxidant capacity of tomato fruits. Food Control 113:107176
Maffei ME (2014) Magnetic field effects on plant growth, development, and evolution. Front Plant Sci 5:445
Marslin G, Sheeba CJ, Franklin G (2017) Nanoparticles alter secondary metabolism in plants via ROS burst. Front Plant Sci 8:832
Martínez-Ballesta MC, Zapata L, Chalbi N, Carvajal M (2016) Multiwalled carbon nanotubes enter broccoli cells enhancing growth and water uptake of plants exposed to salinity. J Nanobiotechnol 14(1):42
Martínez-Medina A, Van Wees SC, Pieterse CM (2017) Airborne signals from Trichoderma fungi stimulate iron uptake responses in roots resulting in priming of jasmonic acid-dependent defences in shoots of Arabidopsis thaliana and Solanum lycopersicum. Plant Cell Environ 40(11):2691–2705
Medina-Pérez V, López-Laredo AR, Sepúlveda-Jiménez G, Zamilpa A, Trejo-Tapia G (2015) Nitrogen deficiency stimulates biosynthesis of bioactive phenylethanoid glycosides in the medicinal plant Castilleja tenuiflora Benth. Acta Physiol Plant 37(5):93
Miliute I, Buzaite O, Baniulis D, Stanys V (2015) Bacterial endophytes in agricultural crops and their role in stress tolerance: a review. Zemdirbyste 102(4):465–478
Mirzajani F, Askari H, Hamzelou S, Schober Y, Römpp A, Ghassempour A, Spengler B (2014) Proteomics study of silver nanoparticles toxicity on Oryza sativa L. Ecotoxicol Environ Saf 108:335–339
Mishra RC, Ghosh R, Bae H (2016) Plant acoustics: in the search of a sound mechanism for sound signaling in plants. J Exp Bot 67(15):4483–4494
Moghimi M, Farzaneh V, Bakhshabadi H (2018) The effect of ultrasound pretreatment on some selected physicochemical properties of black cumin (Nigella Sativa). Nutrire 43(1):18
Mohammed KA, Hellal F, SAA E-S, (2015) Influence of sulfur deprivation on biomass allocation, mineral composition and fruit quality of tomato plants. Middle East J 4(1):42–48
Moola AK, Diana RKB (2019) Elicitation as a means for enhanced plant secondary metabolites through hairy root system. J Adv Sci Res 10(3)
Muller-Xing R, Xing Q, Goodrich J (2014) Footprints of the sun: memory of UV and light stress in plants. Front Plant Sci 5:474. https://doi.org/10.3389/fpls.2014.00474
Naik PM, Al-Khayri JM (2016) Abiotic and biotic elicitors–role in secondary metabolites production through in vitro culture of medicinal plants. Abiotic and biotic stress in plants-recent advances and future perspectives: 247–277
Nakabayashi R, Yonekura-Sakakibara K, Urano K, Suzuki M, Yamada Y, Nishizawa T, Matsuda F, Kojima M, Sakakibara H, Shinozaki K (2014) Enhancement of oxidative and drought tolerance in Arabidopsis by overaccumulation of antioxidant flavonoids. Plant J 77(3):367–379
Nandini B, Puttaswamy H, Prakash HS, Adhikari S, Jogaiah S, Nagaraja G (2020) Elicitation of novel trichogenic-lipid nanoemulsion signaling resistance against Pearl Millet Downy Mildew disease. Biomolecules 10(1):25
Nasim SA, Dhir B (2010) Heavy metals alter the potency of medicinal plants. In: Reviews of environmental contamination and toxicology. Springer, pp 139–149
Nath M, Tuteja N (2016) NPKS uptake, sensing, and signaling and miRNAs in plant nutrient stress. Protoplasma 253(3):767–786
Naveed M, Hussain MB, Zahir ZA, Mitter B, Sessitsch A (2014) Drought stress amelioration in wheat through inoculation with Burkholderia phytofirmans strain PsJN. J Plant Growth Regul 73(2):121–131
Naziya B, Murali M, Amruthesh KN (2020) Plant growth-promoting fungi (PGPF) instigate plant growth and induce disease resistance in Capsicum annuum L. upon infection with Colletotrichum capsici (Syd.) Butler & Bisby. Biomolecules 10(1):41
Naznin M, Lefsrud M, Gravel V, Hao X (2016) Different ratios of red and blue LED light effects on coriander productivity and antioxidant properties. In: VIII international symposium on light in horticulture 1134. pp 223–230
Nejat N, Mantri N (2017) Plant immune system: crosstalk between responses to biotic and abiotic stresses the missing link in understanding plant defence. Curr Issues Mol Biol 23:1–16
Oancea S, Ghincevici D, Ketney O (2014) The effect of ultrasonic pretreatment and sample preparation on the extraction yield of antioxidant compounds and activity of black currant fruits. Acta Chim Slov 62(1):242–248
Oberdörster G, Oberdörster E, Oberdörster J (2005) Nanotoxicology: an emerging discipline evolving from studies of ultrafine particles. Environ Health Perspect 113(7):823–839
Odriozola-Serrano I, Soliva-Fortuny R, Hernández-Jover T, Martín-Belloso O (2009) Carotenoid and phenolic profile of tomato juices processed by high intensity pulsed electric fields compared with conventional thermal treatments. Food Chem 112(1):258–266
Panjai L, Noga G, Fiebig A, Hunsche M (2017) Effects of continuous red light and short daily UV exposure during postharvest on carotenoid concentration and antioxidant capacity in stored tomatoes. Sci Hortic-Amsterdam 226:97–103
Pappas ML, Liapoura M, Papantoniou D, Avramidou M, Kavroulakis N, Weinhold A, Broufas GD, Papadopoulou KK (2018) The beneficial endophytic fungus Fusariumsolani strain K alters tomato responses against spider mites to the benefit of the plant. Front Plant Sci 9:1603
Patel H, Krishnamurthy R (2013) Elicitors in plant tissue culture. Int J Pharmacogn Phytochem 2(2):60–65
Peng Z, Tian J, Luo R, Kang Y, Lu Y, Hu Y, Liu N, Zhang J, Cheng H, Niu S (2019) MiR399d and epigenetic modification comodulate anthocyanin accumulation in Malus leaves suffering from phosphorus deficiency. Plant Cell Environ 43(5):1148–1159
Pereira M, Morais L, Marques E, Martins A, Cavalcanti V, Rodrigues F, Gonçalves W, Blank A, Pasqual M, Dória J (2019) Humic substances and efficient microorganisms: elicitation of medicinal plants—a review. J Agric Sci 11(7)
Pérez-Ambrocio A, Guerrero-Beltrán J, Aparicio-Fernández X, Ávila-Sosa R, Hernández-Carranza P, Cid-Pérez S, Ochoa-Velasco C (2018) Effect of blue and ultraviolet-C light irradiation on bioactive compounds and antioxidant capacity of habanero pepper (Capsicum chinense) during refrigeration storage. Postharvest Biol Technol 135:19–26
Piechulla B, Degenhardt J (2014) The emerging importance of microbial volatile organic compounds. Plant Cell Environ 37(4):811–812
Qi X, Fang H, Yu X, Xu D, Li L, Liang C, Lu H, Li W, Chen Y, Chen Z (2018) Transcriptome analysis of JA signal transduction, transcription factors, and monoterpene biosynthesis pathway in response to methyl jasmonate elicitation in Mentha canadensis L. Int J Mol Sci 19(8):2364
Qian H, Liu T, Deng M, Miao H, Cai C, Shen W, Wang Q (2016) Effects of light quality on main health-promoting compounds and antioxidant capacity of Chinese kale sprouts. Food Chem 196:1232–1238
Răcuciu M, Oancea S (2018) Impact of 50 Hz magnetic field on the content of polyphenolic compounds from blackberries. Bulg Chem Commun 50(3):393–397
Radhakrishnan R (2019) Magnetic field regulates plant functions, growth and enhances tolerance against environmental stresses. Physiol Mol Biol Plants 1–13
Raei M, Angaji SA, Omidi M, Khodayari M (2014) Effect of abiotic elicitors on tissue culture of Aloe vera. Int J Biosci 5(1):74–81
Rahmani N, Radjabian T, Soltani BM (2020) Impacts of foliar exposure to multi-walled carbon nanotubes on physiological and molecular traits of Salvia verticillata L. as a medicinal plant. Plant Physiol Biochem 150:27–38
Rakshit A, Singh HB, Singh AK, Singh US, Fraceto L (2020) New frontiers in stress management for durable agriculture. Springer
Ramirez-Estrada K, Vidal-Limon H, Hidalgo D, Moyano E, Golenioswki M, Cusidó RM, Palazon J (2016) Elicitation, an effective strategy for the biotechnological production of bioactive high-added value compounds in plant cell factories. Molecules 21(2):182
Ribas-Agustí A, Martín-Belloso O, Soliva-Fortuny R, Elez-Martínez P (2019) Enhancing hydroxycinnamic acids and flavan-3-ol contents by pulsed electric fields without affecting quality attributes of apple. Food Res Int 121:433–440
Rico CM, Lee SC, Rubenecia R, Mukherjee A, Hong J, Peralta-Videa JR, Gardea-Torresdey JL (2014) Cerium oxide nanoparticles impact yield and modify nutritional parameters in wheat (Triticum aestivum L.). J Agric Food Chem 62(40):9669–9675
Roossinck MJ (2015) Plants, viruses and the environment: ecology and mutualism. Virology 479:271–277
Routray W, Orsat V, Lefsrud M (2018) Effect of postharvest LED application on phenolic and antioxidant components of blueberry leaves. Chem Eng 2(4):56
Rudrappa T, Neelwarne B, Aswathanarayana RG (2004) In situ and ex situ adsorption and recovery of betalains from hairy root cultures of Beta vulgaris. Biotechnol Prog 20(3):777–785
Rusakova A, Nosachev I, Lysenko V, Guo Y, Logvinov A, Kirichenko E, Varduny T, Cherednikov S, Chugueva O (2017) Impact of high strength electromagnetic fields generated by Tesla transformer on plant cell ultrastructure. Inf Process Agric 4(3):253–258
Ryu C-M, Farag MA, Hu C-H, Reddy MS, Wei H-X, Paré PW, Kloepper JW (2003) Bacterial volatiles promote growth in Arabidopsis. Proc Natl Acad Sci U S A 100(8):4927–4932
Sá RA, Sá RA, Alberton O, Gazim ZC, Laverde A Jr, Caetano J, Amorin AC, Dragunski DC (2015) Phytoaccumulation and effect of lead on yield and chemical composition of Mentha crispa essential oil. Desalin Water Treat 53(11):3007–3017
Sade D, Shriki O, Cuadros-Inostroza A, Tohge T, Semel Y, Haviv Y, Willmitzer L, Fernie AR, Czosnek H, Brotman Y (2015) Comparative metabolomics and transcriptomics of plant response to Tomato yellow leaf curl virus infection in resistant and susceptible tomato cultivars. Metabolomics 11(1):81–97
Saeed S, Ali H, Khan T, Kayani W, Khan MA (2017) Impacts of methyl jasmonate and phenyl acetic acid on biomass accumulation and antioxidant potential in adventitious roots of Ajuga bracteosa Wall ex Benth., a high valued endangered medicinal plant. Physiol Mol Biol Plants 23(1):229–237
Samad AF, Sajad M, Nazaruddin N, Fauzi IA, Murad A, Zainal Z, Ismail I (2017) MicroRNA and transcription factor: key players in plant regulatory network. Front Plant Sci 8:565
Samad AFA, Rahnamaie-Tajadod R, Sajad M, Jani J, Murad AMA, Noor NM, Ismail I (2019) Regulation of terpenoid biosynthesis by miRNA in Persicaria minor induced by Fusarium oxysporum. BMC Genomics 20(1):586
Samanta A, Das G, Das SK (2011) Roles of flavonoids in plants. Carbon 100(6):12–35
Sánchez-López ÁM, Baslam M, De Diego N, Muñoz FJ, Bahaji A, Almagro G, Ricarte-Bermejo A, García-Gómez P, Li J, Humplík JF (2016) Volatile compounds emitted by diverse phytopathogenic microorganisms promote plant growth and flowering through cytokinin action. Plant Cell Environ 39(12):2592–2608
Santoro MV, Bogino PC, Nocelli N, Cappellari LdR, Giordano WF, Banchio E (2016) Analysis of plant growth-promoting effects of fluorescent Pseudomonas strains isolated from Mentha piperita rhizosphere and effects of their volatile organic compounds on essential oil composition. Front Microbiol 7:1085
Santoro MV, Zygadlo J, Giordano W, Banchio E (2011) Volatile organic compounds from rhizobacteria increase biosynthesis of essential oils and growth parameters in peppermint (Mentha piperita). Plant Physiol Biochem 49(10):1177–1182
Shahhoseini R, Azizi M, Asili J, Moshtaghi N, Samiei L (2020) Effects of zinc oxide nanoelicitors on yield, secondary metabolites, zinc and iron absorption of Feverfew (Tanacetum parthenium (L.) Schultz Bip.). Acta Physiol Plant 42:1–18
Shang L, Nienhaus K, Nienhaus GU (2014) Engineered nanoparticles interacting with cells: size matters. J Nanobiotechnol 12(1):5
Sharma D, Tiwari M, Pandey A, Bhatia C, Sharma A, Trivedi PK (2016) MicroRNA858 is a potential regulator of phenylpropanoid pathway and plant development. Plant Physiol 171(2):944–959
Silva RN, Monteiro VN, Steindorff AS, Gomes EV, Noronha EF, Ulhoa CJ (2019) Trichoderma/pathogen/plant interaction in pre-harvest food security. Fungal Biol 123(8):565–583
Singh A, Dwivedi P (2018) Methyl-jasmonate and salicylic acid as potent elicitors for secondary metabolite production in medicinal plants: a review. J Pharmacogn Phytochem 7(1):750–757
Singh D, Basu C, Meinhardt-Wollweber M, Roth B (2015) LEDs for energy efficient greenhouse lighting. Renew Sust Energ Rev 49:139–147
Singh NR, Rath SK, Behera S, Naik SK (2018) In vitro secondary metabolite production through fungal elicitation: an approach for sustainability. In: Fungal nanobionics: principles and applications. Springer, pp 215–242
Soliva-Fortuny R, Vendrell-Pacheco M, Martín-Belloso O, Elez-Martínez P (2017) Effect of pulsed electric fields on the antioxidant potential of apples stored at different temperatures. Postharvest Biol Technol 132:195–201
Sosan A, Svistunenko D, Straltsova D, Tsiurkina K, Smolich I, Lawson T, Subramaniam S, Golovko V, Anderson D, Sokolik A (2016) Engineered silver nanoparticles are sensed at the plasma membrane and dramatically modify the physiology of Arabidopsis thaliana plants. Plant J 85(2):245–257
Starodubtseva GP, Livinskiy SA, Gabriyelyan SZ, Lubaya SI, Afanacev MA (2018) Process control of Pre-sowing seed treatment by Pulsed electric field. Acta Technol Agric 21(1):28–32
Subbaiah LV, Prasad TNVKV, Krishna TG, Sudhakar P, Reddy BR, Pradeep T (2016) Novel effects of nanoparticulate delivery of zinc on growth, productivity, and zinc biofortification in maize (Zea mays L.). J Agric Food Chem 64(19):3778–3788
Taghizadeh M, Nasibi F, Kalantari KM, Ghanati F (2019) Evaluation of secondary metabolites and antioxidant activity in Dracocephalum polychaetum Bornm. cell suspension culture under magnetite nanoparticles and static magnetic field elicitation. Plant Cell Tiss Org 136(3):489–498
Tani E, Kizis D, Markellou E, Papadakis I, Tsamadia D, Leventis G, Makrogianni D, Karapanos I (2018) Cultivar-dependent responses of eggplant (Solanum melongena L.) to simultaneous Verticillium dahliae infection and drought. Front Plant Sci 9:1181
Thakur M, Bhattacharya S, Khosla PK, Puri S (2019) Improving production of plant secondary metabolites through biotic and abiotic elicitation. J Appl Res Med Aroma 12:1–12
Tian H, Ghorbanpour M, Kariman K (2018) Manganese oxide nanoparticle-induced changes in growth, redox reactions and elicitation of antioxidant metabolites in deadly nightshade (Atropa belladonna L.). Ind Crop Prod 126:403–414
Tiwari D, Dasgupta-Schubert N, Cendejas LV, Villegas J, Montoya LC, García SB (2014) Interfacing carbon nanotubes (CNT) with plants: enhancement of growth, water and ionic nutrient uptake in maize (Zea mays) and implications for nanoagriculture. Appl Nanosci 4(5):577–591
Toma M, Vinatoru M, Paniwnyk L, Mason TJ (2001) Investigation of the effects of ultrasound on vegetal tissues during solvent extraction. Ultrason Sonochem 8(2):137–142
Tripathi AM, Singh A, Singh R, Verma AK, Roy S (2019) Modulation of miRNA expression in natural populations of A. thaliana along a wide altitudinal gradient of Indian Himalayas. Sci Rep 9(1):1–16
Upadhyaya H, Dutta BK, Panda SK (2013) Zinc modulates drought-induced biochemical damages in tea [Camellia sinensis (L) O Kuntze]. J Agric Food Chem 61(27):6660–6670
Vaezzadeh M, Noruzifar E, Faezeh G, Salehkotahi M, Mehdian R (2006) Excitation of plant growth in dormant temperature by steady magnetic field. J Magn Magn Mater 302(1):105–108
Van Oosten MJ, Pepe O, De Pascale S, Silletti S, Maggio A (2017) The role of biostimulants and bioeffectors as alleviators of abiotic stress in crop plants. Chem Biol Technol Agric 4(1):5
Vance ME, Kuiken T, Vejerano EP, McGinnis SP, Hochella MF Jr, Rejeski D, Hull MS (2015) Nanotechnology in the real world: redeveloping the nanomaterial consumer products inventory. Beilstein J Nanotechnol 6(1):1769–1780
Vargas-Hernandez M, Macias-Bobadilla I, Guevara-Gonzalez RG, Romero-Gomez SdJ, Rico-Garcia E, Ocampo-Velazquez RV, Alvarez-Arquieta LdL, Torres-Pacheco I (2017) Plant hormesis management with biostimulants of biotic origin in agriculture. Front Plant Sci 8:1762
Vargas-Hernández M, Vázquez-Marrufo G, Aguilar-Ruiz Carlos A, González-Márquez MA, Rocha O, Cerna-Pantoja D, Cruz-Hernández A (2019) MicroRNAs associated with secondary metabolites production. In: Plant physiological aspects of phenolic compounds. IntechOpen
Vazquez-Hernandez C, Feregrino-Perez AA, Perez-Ramirez I, Ocampo-Velazquez RV, Rico-García E, Torres-Pacheco I, Guevara-Gonzalez RG (2019) Controlled elicitation increases steviol glycosides (SGs) content and gene expression-associated to biosynthesis of SGs in Stevia rebaudiana B. cv. Morita II. Ind Crop Prod 139:111479
Vázquez-Hernández M, Parola-Contreras I, Montoya-Gómez L, Torres-Pacheco I, Schwarz D, Guevara-González R (2019) Eustressors: Chemical and physical stress factors used to enhance vegetables production. Sci Hortic-Amsterdam 250:223–229
Vecchio G, Galeone A, Brunetti V, Maiorano G, Rizzello L, Sabella S, Cingolani R, Pompa PP (2012) Mutagenic effects of gold nanoparticles induce aberrant phenotypes in Drosophila melanogaster. Nanomed Nanotechnol Biol Med 8(1):1–7
Verma V, Ravindran P, Kumar PP (2016) Plant hormone-mediated regulation of stress responses. BMC Plant Biol 16(1):86
Vicaş SI, Bandici L, Teuşdea AC, Turcin V, Popa D, Bandici GE (2017) The bioactive compounds, antioxidant capacity, and color intensity in must and wines derived from grapes processed by pulsed electric field. CYTA-J Food 15(4):553–562
Villarreal-García D, Nair V, Cisneros-Zevallos L, Jacobo-Velázquez DA (2016) Plants as biofactories: Postharvest stress-induced accumulation of phenolic compounds and glucosinolates in broccoli subjected to wounding stress and exogenous phytohormones. Front Plant Sci 7:45
Vinatoru M (2001) An overview of the ultrasonically assisted extraction of bioactive principles from herbs. Ultrason Sonochem 8(3):303–313
Vivaldo G, Masi E, Taiti C, Caldarelli G, Mancuso S (2017) The network of plants volatile organic compounds. Sci Rep 7(1):1–18
Wang W, Li Y, Dang P, Zhao S, Lai D, Zhou L (2018) Rice secondary metabolites: structures, roles, biosynthesis, and metabolic regulation. Molecules 23(12):3098
Wang W, Tang K, Yang H-R, Wen P-F, Zhang P, Wang H-L, Huang W-D (2010) Distribution of resveratrol and stilbene synthase in young grape plants (Vitis vinifera L. cv. Cabernet Sauvignon) and the effect of UV-C on its accumulation. Plant Physiol Biochem 48(2–3):142–152
Wang X, Yang X, Chen S, Li Q, Wang W, Hou C, Gao X, Wang L, Wang S (2016) Zinc oxide nanoparticles affect biomass accumulation and photosynthesis in Arabidopsis. Front Plant Sci 6:1243
Wang X, Zhang D, Cui N, Yu Y, Yu G, Fan H (2018) Transcriptome and miRNA analyses of the response to Corynespora cassiicola in cucumber. Sci Rep 8(1):1–16
Wiktor A, Sledz M, Nowacka M, Rybak K, Chudoba T, Lojkowski W, Witrowa-Rajchert D (2015) The impact of pulsed electric field treatment on selected bioactive compound content and color of plant tissue. Innov Food Sci Emerg 30:69–78
Wiktor A, Sledz M, Nowacka M, Rybak K, Witrowa-Rajchert D (2016) The influence of immersion and contact ultrasound treatment on selected properties of the apple tissue. Appl Acoust 103:136–142
Witrowa-Rajchert D, Wiktor A, Sledz M, Nowacka M (2014) Selected emerging technologies to enhance the drying process: A review. Dry Technol 32(11):1386–1396
Xi-ran L, Ting F (2017) The application of high pulsed electric field technology in extracting natural compounds. Food Ind 1:69
Xu P, Chen F, Mannas JP, Feldman T, Sumner LW, Roossinck MJ (2008) Virus infection improves drought tolerance. New Phytol 180(4):911–921
Yan Y, Zhou S, Song Z, Li F, Zhang C, Zhang X, Song H (2017) Effects of frequency and voltage of high voltage pulsed electric field on improving vigor of aged cotton seed. Trans Chinese Soc Agric Eng 33(13):310–314
Yokel RA, MacPhail RC (2011) Engineered nanomaterials: exposures, hazards, and risk prevention. J Occup Med Toxicol 6(1):7
Yu J, Engeseth NJ, Feng H (2016) High intensity ultrasound as an abiotic elicitor—effects on antioxidant capacity and overall quality of romaine lettuce. Food Bioprocess Tech 9(2):262–273
Yuliani F, Dewi WS, Yunus A, Siswanto U (2018) The study of artemisinin content in callus Artemisia annua L. cultures elicited with endophytic fungi Aspergillus sp. Molekul 13(2):155–161
Zaker A, Sykora C, Gössnitzer F, Abrishamchi P, Asili J, Mousavi SH, Wawrosch C (2015) Effects of some elicitors on tanshinone production in adventitious root cultures of Perovskia abrotanoides Karel. Ind Crop Prod 67:97–102
Zareei E, Zaare-Nahandi F, Oustan S, Hajilou J (2019) Effects of magnetic solutions on some biochemical properties and production of some phenolic compounds in grapevine (Vitis vinifera L.). Sci Hortic-Amsterdam 253:217–226
Zaynab M, Fatima M, Abbas S, Sharif Y, Umair M, Zafar MH, Bahadar K (2018) Role of secondary metabolites in plant defense against pathogens. Microb Pathog 124:198–202
Zhao J, Zhu W-H, Hu Q (2001) Selection of fungal elicitors to increase indole alkaloid accumulation in Catharanthus roseus suspension cell culture. Enzyme Microb Technol 28(7–8):666–672
Zhao L, Huang Y, Hu J, Zhou H, Adeleye AS, Keller AA (2016) 1H NMR and GC-MS based metabolomics reveal defense and detoxification mechanism of cucumber plant under nano-Cu stress. Environ Sci Technol 50(4):2000–2010
Zhao L, Peng B, Hernandez-Viezcas JA, Rico C, Sun Y, Peralta-Videa JR, Tang X, Niu G, Jin L, Varela-Ramirez A (2012) Stress response and tolerance of Zea mays to CeO2 nanoparticles: cross talk among H2O2, heat shock protein, and lipid peroxidation. ACS Nano 6(11):9615–9622
Zhou J-Y, Li X, Zheng J-Y, Dai C-C (2016) Volatiles released by endophytic Pseudomonas fluorescens promoting the growth and volatile oil accumulation in Atractylodes lancea. Plant Physiol Biochem 101:132–140
Zhou LS, Tang K, Guo SX (2018) The plant growth-promoting fungus (PGPF) Alternaria sp. A13 markedly enhances Salvia miltiorrhiza root growth and active ingredient accumulation under greenhouse and field conditions. Int J Mol Sci 19(1):270
Złotek U, Szymanowska U, Rybczyńska-Tkaczyk K, Jakubczyk A (2020) Effect of jasmonic acid, yeast extract elicitation, and drying methods on the main bioactive compounds and consumer quality of lovage (Levisticum Officinale Koch). Foods 9(3):323
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2021 The Author(s), under exclusive license to Springer Nature Switzerland AG
About this chapter

Cite this chapter
Aguirre-Becerra, H. et al. (2021). Role of Stress and Defense in Plant Secondary Metabolites Production. In: Pal, D., Nayak, A.K. (eds) Bioactive Natural Products for Pharmaceutical Applications. Advanced Structured Materials, vol 140. Springer, Cham. https://doi.org/10.1007/978-3-030-54027-2_5
Download citation
DOI: https://doi.org/10.1007/978-3-030-54027-2_5
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-54026-5
Online ISBN: 978-3-030-54027-2
eBook Packages: Chemistry and Materials ScienceChemistry and Material Science (R0)