
Abstract
Endophytic microbes including bacteria and fungi inhabiting in seed tissues have recently gained significant importance owing to a diversity of roles that they play and eventually resulting in improved plant growth as well as plant fitness. Some of the major roles played by seed endophytic microbes include plant growth promotion by enhanced nutrient acquisition or production of growth hormones, nitrogen fixation, phosphate solubilisation, and protection against pathogens as well as abiotic stresses. Since, rice is one of the important staple crop across the globe, there is a great need to explore and decipher the roles of the endophytic community present inside it. This chapter focuses on the diversity and distribution of rice seed endophytes, their transmission along with the various functional roles that they play inside the plants with an aim to provide deep insights on rice seed endophytes as plant growth promoting and biocontrol agents.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
1 Introduction
The term ‘endophyte’ originally was coined by de Bary in 1866 for all those microbes which live inside plants. Endophytes are often bacteria and fungi, which grow and reside for all or part of their life cycles inside the living host plant without showing disease symptoms or negative effect to the plant tissues (Wilson 1995). Bacterial and fungal endophytes have been found to be associated with nearly all plants that have been examined. It is important to discover the various impacts and roles that endophytes have on the plant development (Arnold et al. 2000). Endophytes have been isolated from roots (Murphy et al. 2015), shoots (Rojas et al. 2016), flowers, leaves (Yumlembam and Borkar 2014), fruits (Krishnan et al. 2012), seeds (Truyens et al. 2015; Verma et al. 2017a, b) and meristematic tissues of the plants (Pirttilä et al. 2003). The bacterial community of seeds, either endophytic or an epiphytic, has been shown to contribute to rhizospheric community development (Johnston-Monje et al. 2016). Seed endophytes have been reported from numerous crops including rice (Verma et al. 2018), wheat (Larran et al. 2016), cotton (Lopez 2015; Irizarry and White 2017) and corn (Gond et al. 2015; Shehata et al. 2016). Seed-vectored endophytic bacteria are indigenous in plants and have been shown to be transmitted vertically from generation to generation. Endophytes are known to have positive impacts on plants starting from germination and seedling establishment, since they play important roles in nutrient mobilisation (Li et al. 2012; White et al. 2015), nitrogen fixation (Elbeltagy et al. 2001), phosphate solubilisation (Barrow and Osuna 2002; Verma et al. 2017a, b) and plant growth promotion (Compant et al. 2010) and provide tolerance against biotic and abiotic stresses (Kuldau and Bacon 2008; Gond et al. 2015). Bacterial seed endophytes are also found to reduce fungal disease infection during seedling development (Verma et al. 2017b; Verma and White 2018; Verma et al. 2018). The potential application of the bacterial and fungal endophytes in important food crops has attracted the scientific community and stimulated research to gain a better characterisation and understanding of endophytes and their functional roles in plant development and ecology (Mitter et al. 2013).
Rice is a staple crop of nearly half of the world’s total population and one of the most important food crops of the world (Gyaneshwar et al. 2001). A major problem is the burgeoning human population with limited cultivable land; hence, efforts are needed to increase rice productivity. On the other hand, modern agricultural practices employed in improving rice production include plant breeding programmes which involve years of breeding, as well as use of chemical fertilisers and pesticides that pose threats to our already suffering environment (Mano and Morisaki 2008). The proper management and application of seed endophytic bacteria in promoting plant growth are gaining importance due to endophyte beneficial effects and their environmentally friendly nature. Studies have reported that certain seed-derived endophytic bacteria play important roles in modulating seedling development and defence against pathogens (Verma et al. 2017a, b; Ma et al. 2011; Glick 2005). Similarly, other experiments conducted on seed endophytes in rice have shown growth promotional activity including nitrogen fixation, hormonal modulation, siderophore formation and phosphate solubilisation (Walitang et al. 2017). Seed endophytic microbes are shown to be important in studies of plant-endophyte interactions (Hardoim et al. 2008). Seed-borne endophytes are better positioned to interact with young developing seedlings, when compared to the bulk of soil bacteria (Rosenblueth and Martínez-Romero 2006). In addition, seed endophytes are transferred vertically and hence remain associated with plants for many generations, thereby establishing a closely adapted and strong association with plants (Johnston-Monje and Raizada 2011).
This book chapter provides a comprehensive review of the diversity of seed endophytes inhabiting rice and their possible functional roles in development and growth of rice plants, along with other contributions in various processes such as in providing resistance against biotic and abiotic stresses. This chapter also includes recent studies on functional metagenomics of seeds which tells about the putative functional role of non-cultivable endophytes residing in rice seeds. Future prospects for understanding the basic science of rice seed endophytes as well as their potential applications for sustainable agriculture have also been considered.
2 Diversity and Distribution of Rice Seed Endophytes
Research on the presence of endophytic bacteria inside plant tissues originated in the 1870s by Pasteur and group (Hollis 1949), but there was a general tendency that these microorganisms inside plants were pathogenic in nature, causing harmful effects. It was Perotti who firstly reported the incidence of non-pathogenic bacterial isolates inside the root tissues of plants (Perotti 1926). Currently, these endophytic microorganisms are present in all the plant species and have been isolated from many varieties of plants (Lodewyckx et al. 2002). Besides being present in vascular plants (Zhang and Yao 2015), endophytes have been found to colonise marine algae (Mathan 2016), mosses and ferns (Verma et al. 2017a, b; Hoysted et al. 2018). Endophyte communities are affected by host plant species and the environmental conditions under which the host is growing (Rosenblueth and Martínez-Romero 2006). Generally, it is believed that endophytic microbes originate from microbes of the rhizosphere, gaining entrance into the plant through natural openings or wounds or penetrating the plant tissues through enzymes such as cellulases and pectinases (Hallmann et al. 1997). However, the first asymptomatic endophytic fungus of genus Epichloë in grasses was discovered at the end of the nineteenth century in Lolium temulentum (Freeman 1904). Some studies suggest that interactions between fungi and host could result from pathogen-host antagonism (Schulz et al. 1999), while other studies suggest a closely coadapted mutualistic symbiotic association.
To obtain information on endophytic bacterial and fungal diversity, culture-dependent as well as culture-independent approaches can be employed, but culture-independent molecular approaches such as 16S rDNA sequencing techniques, denaturing gradient gel electrophoresis (DGGE), etc. are gaining more importance because of the unknown conditions for growth of many endophytic microorganisms (Tholozan et al. 1999).
Rice is a gramineous plant and has a deep fibrous root system for drawing for water and nutrient absorption. Various bacteria have been reported from rice plants, including Pantoea from the seeds, Rhizobium and Burkholderia from the roots, Methylobacterium from the shoots, etc. (Mano and Morisaki 2008). Researchers isolated Pantoea ananas from the seeds of Oryza alta, Herbaspirillum seropedicae and Methylobacterium sp. from Oryza meridionalis and Klebsiella oxytoca from Oryza sativa (Elbeltagy et al. 2000). Similarly, Bacillus cereus, Pantoea ananatis, Sphingomonas echinoides and Sphingomonas parapaucimobilis were isolated from Oryza sativa (Okunishi et al. 2005). Various bacterial endophytes such as Pseudomonas fluorescens, Xanthomonas sacchari and Staphylococcus sp. were also identified from milled rice seeds using ribosomal intergenic spacer analysis (RISA). A study was conducted on the diversity of endophytic community of rice seeds of salt-tolerant and salt-sensitive cultivars (Walitang et al. 2017). In this study, it was found that the cultivable seed endophytes were dominated by Proteobacteria notably class Gammaproteobacteria. Extremely identical type strains were isolated that were Flavobacterium sp., Mycobacterium sp. and Xanthomonas sp. from the salt-tolerant as well as salt-sensitive cultivars. The study showed how selective screening of endophytes with physiological characteristics related with tolerance to osmotic stress or ability to detoxify ROS can be done by rice seeds (Walitang et al. 2017). Fungal endophytes such as Alternaria alternata, Epicoccum purpurascens, Fusarium equiseti, Nigrospora oryzae, etc. were isolated from seeds of several different cultivars of rice (Fisher and Petrini 1992).
The endophytic bacteria and fungi isolated from seeds of rice plants are listed in Table 11.1.
3 Transmission of Seed Endophytes
The transmission of endophytes can take place either from the vegetative parts of the plant to the seed or endosperm via chalaza, funiculus and micropyle, i.e. plant maternal vascular connections, or directly through the ovules, thereby colonising the endosperm and the newly formed embryo (Truyens et al. 2015). The transmission of endophyte directly through the ovules results in vertical transmission of endophytes. The transformation of shoot meristems in which an endophyte grows into reproductive meristems followed by endophytic colonisation is the likely route of vertical transmission as these will give rise to ovules and finally seed (Pirttilä et al. 2000). Vertical transmission of bacterial endophytes likely occurs in most species of plants. In one study, it was concluded that seeds contributed the major portion of endophytes to mature wheatgrass plants since the same species of endophytes were recovered from seeds and mature tissues (Ringelberg et al. 2012). Similarly, a study was conducted on vertical transmission of seed endophytes in switchgrass plants, and it was found that the same Bacillus sp. and Microbacterium spp. which inhabited the seeds in first year were present in the switchgrass plant in the second year (Gagne-Bourgue et al. 2013). Moreover, another study reported continuity of the endophytic bacterial community of maize from parent to offsprings. Additionally, genetically connected maize hybrids contained similar species and genera of endophytic bacteria (Liu et al. 2012, 2013).
Fungal endophytes may be transmitted through varied routes in the case of transmission from one host to another, whereas seed transmission is employed when transmission takes place from one generation to another (Shearin et al. 2018; Vujanovic and Germida 2017). In a study conducted on seed endophytes in forbs, it was proposed that vertical transmission is a widespread phenomenon (Hodgson et al. 2014).
4 Functional Role of Rice Seed Endophytes
Seeds contain a diversity of microflora consisting of bacteria and fungi which play various physiological roles, but little work has been done to determine the roles that endophytes play in plant development and ecology. Major functions include plant growth promotion, biocontrol activity, and many other unknown benefits. Some of these functions are described below.
4.1 Seed Endophytes as Plant Growth-Promoting Agents
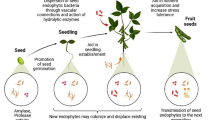
Plant growth promotion is one of the major roles that endophytes play in plants. Plant growth promotion activity in plants can be the result of either direct or indirect stimulation of plant growth. Direct stimulation of plant growth involves facilitation of acquisition of essential nutrients or modulation of growth hormones within plants by the bacteria or fungi; indirect growth promotion may involve inhibition of plant pathogens by the endophytes, thereby reducing the level of infection in the plant tissues (Doty et al. 2009; Luo et al. 2012; Sziderics et al. 2007) (Figs. 11.1 and 11.2).

Scheme depicting functional roles, mechanism of action of seed endophytic bacteria during seedling developments and plant protection

Effect of seed-inhabiting bacteria on seedling development. Where (a) and (d) are control, disinfected with 4% NaOCl and streptomycin (100 μg ml−1), (b) treated with Pantoea dispersa (B2), (c) treated with Bacillus amyloliquefaciens (M4), (e) treated with Pseudomonas sp. and (f) treated with Pantoea dispersa (B2) (From our previous studies; Verma and White 2018; Verma et al. 2017a, b)
4.2 Nutrient Acquisition Facilitation
The mode of action of many endophytic bacteria involves increasing the availability of nutrients for the plant in the rhizosphere (Glick 1995). The essential nutrients involved in uptake are mainly nitrogen, phosphorus and iron. Enhanced plant growth was observed when rice was inoculated with diazotrophic endophyte Azoarcus sp. strain BH72, with stimulation seen even in Nif− mutants (Hurek et al. 1994). Therefore, it was hypothesised that the observed plant growth might be the result of enhanced nutrient-water uptake associated with the colonisation of Azoarcus and not nitrogen fixation (Hurek et al. 1994). Phosphate solubilisation and siderophore formation are the two major methods by which endophytes facilitate nutrient acquisition by plants.
4.2.1 Phosphate Solubilisation
Phosphorus is one of the most important minerals limiting the growth of terrestrial plants. Although large reserves of phosphorus are present in soils, most of the phosphorus remains unavailable to plants. The reason behind this unavailability is that most of the phosphorus in soils is present in insoluble forms (Stevenson and Cole 1999). Plants absorb two basic soluble forms of phosphorus, monobasic (H2PO4−) and dibasic (HPO42−) ions (Glass 1989). Secretion of organic acids and phosphatases are commonly utilised by endophytic bacteria for the facilitation of phosphorus solubilisation.
A study was conducted to determine the diversity of endophytic bacteria residing inside the seeds of a deepwater rice variety, and mineral phosphate solubilisation activity of the isolates was assessed to judge their growth promotional potentials. Four out of seven isolates showed phosphate solubilisation activity (Verma et al. 2001). Similarly, a diazotrophic bacterium isolated from wild rice (Porteresia coarctata) was tested for its phosphate mineralisation activity and found positive based on clearing zones in the tricalcium phosphate solubilisation test (Loganathan and Nair 2003).
4.2.2 Siderophore Formation
Iron is an essential element required for the growth of microorganisms; however, most of the iron remains unavailable to microbes since it is present in hard-to-solubilise mineral form in the soil. In order to sequester iron from the environment, many microorganisms secrete low molecular weight, iron-binding compounds known as siderophores. Siderophores have strong affinity to bind with the Fe+3 form of iron. This soluble form of iron is now transported back inside the microbial cell and is utilised for its growth. Acremonium and Fusarium sp. isolated from seeds of a rice variety from Sri Lanka improved growth of rice, and it was hypothesised that siderophore formation and nitrogen scavenging by the endophytic fungal species could be the reasons behind the improved growth of the plant (Wijesooriya and Deshappriya 2016).
Siderophores are produced by bacteria as well as fungi, but bacterial siderophores have been found to be more effective than fungal siderophores, and thus they scavenge more effectively than fungi, effectively limiting the growth of fungi. Some plants have the ability to release the iron from iron-siderophore complexes by binding to them and releasing iron for plant growth. In this way, plant growth is boosted in two ways, i.e. by growth suppression of pathogenic fungi and enhanced iron nutrition (Bashan and De-Bashan 2005). A study was conducted on endophytic bacteria producing siderophores in rice in Uruguayan soils, and it was proposed that most of the heterotrophic bacteria were siderophore producing in mature plants. Additionally, in vitro inhibition assays showed that siderophore-producing bacteria of the genus Burkholderia were good antagonists of disease-causing fungi (Loaces et al. 2011).
4.2.3 Transference of Nutrients to Plants from Bacteria via the Rhizophagy Cycle Symbiosis
The rhizophagy cycle is a process whereby plants may obtain nutrients from bacteria that alternate between a root intracellular endophytic phase and a free-living soil phase (Paungfoo-Lonhienne et al. 2010; Kandel et al. 2017; Prieto et al. 2017; Verma et al. 2018). Bacteria acquire soil nutrients in the free-living soil phase; nutrients are extracted from bacteria oxidatively in the intracellular endophytic phase (White et al. 2018, 2019). We conducted experiments on seed-vectored pseudomonad endophytes from Phragmites australis using Poa annua as surrogate host. We found that initially the symbiotic pseudomonads grow on the rhizoplane in the exudate zone behind the root meristem. Bacteria enter root tip meristem cells—locating within the periplasmic space between cell wall and plasma membrane. In the periplasmic spaces of root cells, bacteria convert to wall-less L-forms (White et al. 2018, 2019; Verma and White 2018; Verma et al. 2017a, b, 2018). As root cells mature, bacteria are exposed to reactive oxygen (superoxide) produced by NADPH oxidases (NOX) on the root cell plasma membranes. Reactive oxygen degrades some of the intracellular bacteria—effectively extracting nutrients from them likely in the form of oxidised macro- and micronutrients. Surviving bacteria in root epidermal cells trigger root hair elongation (Verma et al. 2017a, b, 2018), and as hairs elongate bacteria exit at the hair tips, reforming cell walls and rod shapes as they emerge into the rhizosphere where they may obtain additional nutrients. Release of bacteria from elongating root hairs ensures that symbiotic bacteria are deposited into the rhizosphere where additional nutrients may be acquired rather than on the rhizoplane where soil nutrients are scarce. Later attraction of bacteria to the root exudate zone behind the root tip meristem again places bacteria in position to enter root meristem cells. Plants appear to manipulate symbiotic bacteria in the rhizophagy cycle by (1) stimulating bacterial growth around root tip meristems of seedlings by secretion of root exudates around the root tip (Lareen et al. 2016); (2) triggering bacteria to enter into periplasmic spaces in root cells at the root tip meristem by absorbing bacterial fermentation products including butyric acid, causing bacteria to upregulate virulence/endoparasitism genes (White 2017; White et al. 2019; Tramontano and Scanlon 1996; Sun and O’Riordan 2013); (3) subjecting bacteria in periplasmic spaces to superoxide formed on root cell plasma membranes to extract nutrients from bacteria (Verma et al. 2017a, b, 2018); and (4) depositing surviving intracellular bacteria back into the rhizosphere from the tips of elongating root hairs to maximise new nutrient acquisition by bacteria (White et al. 2019). Through the rhizophagy symbiosis, plants appear to ‘farm’ symbiotic microbes. Isotope-tracking experiments where plantlets of Agave tequilana were inoculated with 15N-labelled bacteria demonstrated that nitrogen in bacteria was transferred to the plant likely via rhizophagy symbiosis (Beltran-Garcia et al. 2014). Experiments involving grass seedlings with and without endophytic bacteria grown on 15N-labelled proteins suggest that the rhizophagy cycle could account for 30% of the nutrients absorbed by grass roots (White et al. 2015). Evidence to date suggests that all vascular plants engage in rhizophagy symbiosis to some extent (White et al. 2019). Rhizophagy symbiosis may be a critical means by which plants extract nutrients from symbiotic bacteria.
4.2.4 Modulation of Hormonal Levels
The mechanism of hormonal modulation is most commonly exploited by endophytic microbes which synthesise several hormones, including auxins, cytokinins and gibberellins. The synthesis of these hormones can be done either singly or in combinations by microbes (Narula et al. 2013). It is believed that phytohormones produced by endophytic microbes have the ability to stimulate plant growth resulting in bigger and more branched roots with greater surface area (Vessey 2003).
IAA (indole-3-acetic acid) is a type of auxin generally synthesised by endophytic bacteria and is known to stimulate root initiation, cell division and cell enlargement (Salisbury 1994). Most of the endophytic IAA-producing bacteria are thought to promote root growth and increase root length, thereby increasing the root surface area and enabling the plant to absorb more water and nutrients from soil. Indole acetic acid production by endophytic bacteria was estimated in seeds of deepwater rice varieties, and all seven isolates were found to produce indole acetic acid (Verma et al. 2001). In one study 576 isolates of endophytic bacteria were isolated from different parts of 10 Korean rice cultivars (Ji et al. 2014). From these isolates 12 were identified as diazotrophic, and their growth-promoting activities were assessed by their application to seeds of rice cultivars. It was found that inoculated seeds showed improved plant growth and increased height and dry weight. Ten strains showed higher auxin production activity, and it was proposed that auxin production was one of the possible mechanisms of plant growth promotion (Ji et al. 2014). Similarly, a study was conducted on seed-vectored endophytic bacteria in modulation of development of rice seedlings, and IAA production by three isolates (VWB1, VWB2 and VWB3) was determined. The isolates VWB1 and VWB2 were found to produce IAA, and it was proposed that endophyte-produced IAA may be involved in triggering root hair development (Verma et al. 2017a, b).
Cytokinins are also a class of plant hormones known to be involved in cell division promotion, cell enlargement and tissue expansion in specific plant parts. Other groups of phytohormones include gibberellins, abscisic acids and ethylene. Gibberellins are known to be associated with the extension of stem tissues of plants, thereby modifying the plant morphology (Salisbury 1994). Evidence of gibberellin production by endophytic bacteria is rare; however, a study has reported the production of gibberellic acid along with IAA, abscisic acid and cytokinin by a diazotrophic rice endophyte Pantoea agglomerans YS19. The phytohormones were detected by ELISA and finally confirmed by gas chromatography (Feng et al. 2006). An endophytic bacteria RWL isolated from seeds of rice and later identified as Bacillus amyloliquefaciens by sequencing and phylogenetic analysis of 16S rRNA was assessed for its phytohormone production ability. It was reported that the endophytic bacteria secreted various forms of bioactive and inactive gibberellic acids which colonised the roots and promoted rice plant growth (Shahzad et al. 2016).
4.2.5 ACC Deaminase Activity
In response to pathogenic attack or stress, higher amounts of the hormone ethylene are generally secreted by plants which further stimulates senescence, inhibits plant growth and triggers cell death near the infection site. Hence, lowering the level of ethylene after infection could prove very useful to plants. The bacterial enzyme ACC (1-aminocyclopropane-1-carboxylate) deaminase has been found to have an active role in plant growth promotion and was first characterised by Honma and Shimomura in 1978 (Honma and Shimomura 1978). The enzyme catalyses the conversion of ACC (1-aminocyclopropane-1-carboxylate), the immediate precursor of ethylene synthesis in plants into ammonia and α-ketobutyrate. The seeds or plant roots exude the ACC which is metabolised by the bacteria resulting in stimulation of efflux of ACC, thereby decreasing the root ACC concentration as well as root ethylene evolution ultimately increasing root growth (Glick et al. 1998). Several studies have reported that promotion of plant growth by some ACC deaminase-producing bacteria is done under various stressful conditions like flooding, saline conditions and drought (Holguin and Patten 1999). Isolation of ACC deaminase-producing bacteria from seeds of non-cultivated plants and then inoculating them onto crop plants could result in improved growth, even in harsh environmental conditions.
5 Seed Endophytes as Biocontrol Agents
Different types of biotic stresses such as diseases and pests decrease the agricultural productivity of crop plants. Such pests and diseases are controlled by using pesticides and chemical fertilisers resulting in ill-effects on human health and ecotoxicity. The use of endophytic microflora could solve the toxicity issue and help in improving human health (Boddey et al. 1995). Various seed endophytic fungi and bacteria have been reported to possess anti-pathogenic properties making them key prospects to act as biocontrol agents. In rice seed endophytes, two Bacillus strains were found to show maximum antifungal properties against Rhizoctonia solani, Pythium myriotylum, Gaeumannomyces graminis and Heterobasidion annosum, and it was believed that the antifungal activity was due to the production of volatile antifungal compound ammonia or N-acetyl-β-d-glucosaminidase or both (Mukhopadhyay et al. 1996). A study suggested the presence of antifungal activity in numerous strains of Microbacterium, Pseudomonas, Pantoea, Paenibacillus and Curtobacterium, all isolated from rice seeds and were found to be effective against Curvularia sp., Fusarium oxysporum and Pythium ultimum (Ruiza et al. 2011). Similar study on rice seed endophytes reported in vitro antifungal activity against Rhizoctonia solani and Pyricularia grisea (Cottyn et al. 2001). We also found that presence of seed endophytes protects seedlings from fungal infection during seedling development (Fig. 11.3).

Effect of seed endophytes Pantoea dispersa on root hair formations and protection from fungal infection. Where (a, b, c) are control and (d, e, f) are treatment with Pantoea dispersa; (a) very few and short root hairs, (b) clean root parenchyma colonise with fungal mycelium (arrows), (c) clean root hairs colonise with fungal mycelium (arrows), (d) very long and dense root hairs (arrows), (e) intracellular bacteria in root parenchyma and no fungal infection (arrows) and (f) internalisation of bacteria in root hairs (from our previous studies; Verma et al. 2017b, 2018)
Studies have reported that toxin production by endophytic fungi gives them the capacity to deter insects, inducing weight loss and also causing an increase in pest death rates (Azevedo et al. 2000). Several studies suggest that a number of fungi have completely different mode of action and that they render the plant uneatable for various kinds of pests like grasshopper, aphids, etc. (Clay 1989; Carroll 1991). A study was done on two classical rice varieties (Suwandel and Kaluheenati) of Sri Lanka, and it was reported that rice seed-borne fungi Absidia and Cylindrocladium showed high inhibition to pathogen (Magnaporthe grisea), and it was found that these fungi controlled the pathogen by coiling the hyphae around it, forming clamps and loops (Atugala and Deshappriya 2015).
The mechanisms behind inhibition of plant pathogens by endophytes are still not very clear, but a number of explanations have been given in this context. Reports suggest that some endophytes inhibit pathogens by direct mechanism involving the secretion of lytic enzymes and hence directly suppressing them by antibiosis. This kind of interaction between fungal endophytes and pathogens can be complex as well as very specific (Arnold et al. 2003). Another mechanism of pathogen inhibition by fungal endophytes involves the production of organic compounds such as alkaloids by the plant, and these alkaloids help in growth suppression of pathogens (Cheplick et al. 1989). Systemic acquired resistance (SAR) mediated by salicylic acid, as well as pathogenesis-related (PR) proteins, and induced systemic resistance (ISR) mediated by jasmonic acid or ethylene are other modes of action by which growth of pathogens may be suppressed in plants by endophytes (Vallad and Goodman 2004).
5.1 Synthesis of Allelochemicals
Allelochemicals are bioactive compounds, including antibiotics, siderophores, lytic enzymes, etc. that may act defensively against invading competitors, pathogens or pests, while allelopathy refers to the defensive action of allelochemicals (Saraf et al. 2014). These naturally synthesised chemical compounds can be utilised in crop disease management programmes in place of chemical pesticides. As natural pesticides, allelochemicals may solve health-related problems and other soil- and environmental pollution-related issues leading to climate change caused by the use of agrochemicals (Farooq et al. 2011). Allelopathic effects frequently depend on a number of environmental conditions such as presence of water, nutrition, bacterial density and soil structure as well as its texture (Barazani and Friedman 2001). Studies suggest that allelopathic rice can have a positive influence on the plant population density as well as soil microbial community (Kong et al. 2008). For allelopathy to be more effective, either improved environmental conditions favouring the biological agent could be provided or the microbe’s allelochemicals may be enhanced/increased through genetic transformation (Hornok 2000). Genetic enhancement of the biocontrol agent can be achieved by various means such as mutation (physical or chemical), development of sexual hybrids, homokaryon production or by genetic manipulations like protoplast fusion, site-directed mutagenesis, recombination, transformation, etc. (Palumbo et al. 2005). Several different methods of allelopathy are described below.
5.2 Antibiotic Production
Production of antibiotics is one of the most powerful mechanisms utilised by endophytes against the phytopathogens. Antibiotics refer to those naturally synthesised chemical compounds released by microorganisms such as bacteria that inhibit the growth and metabolic activities of other microorganisms, and this phenomenon of suppression of pathogen is known as antibiosis (Fravel 1988). In one study, Pseudomonas fluorescens strain 7–14 isolated from the rice rhizosphere was found to produce antibiotics that were effective against Pyricularia sp., Rhizoctonia solani, Pythium ultimum and Gaeumannomyces graminis var. tritici. The data also showed that Pseudomonas fluorescens strain 7–14 was as effective as the fungicide tricyclazole in controlling several diseases of rice (Chatterjee et al. 1996). The experiments conducted under laboratory conditions have shown production of several antibiotics, as well as their effectiveness against various pathogens, but antibiotic availability under field conditions remains an area of concern. The antibiotic activity of endophytes may be increased as the genes involved in the synthesis of some of the antibiotics are well known, and hence, although theoretical, the suppression of phytopathogens can also be enhanced (Holguin and Patten 1999).
5.3 Lytic Enzyme Production
Cell wall lysis is one of the potential mechanisms by which endophytic microbes can control growth of pathogens. Several bacteria have the ability to produce various enzymes that can easily hydrolyse chitin, cellulose, hemicellulose and proteins, thereby directly suppressing the growth of pathogenic agents. In a study, Stenotrophomonas maltophilia strain F-81, isolated from the rhizosphere of sugar beet, showed the production of extracellular chitinase and protease enzymes that restricted the growth of the phytopathogenic fungus Pythium ultimum in vitro (Dunne et al. 1997). Similarly, endophytic Bacillus cereus strain 65 was found to produce and excrete the enzyme chitinase that suppressed the growth of Rhizoctonia solani (Pleban et al. 1997). A study was done on endophytic Bacillus sp. isolated from the roots of balloon flower (Platycodon grandiflorus) which showed strong antifungal activity against Rhizoctonia solani, Pythium ultimum and Fusarium oxysporum. The bacterium synthesised iturin A along with cellulase and xylanase (Cho et al. 2003). Most of the studies in this field are focused on rhizospheric bacteria, and hence very little is known about mechanism of cell lysis resulting in suppression of pathogen growth by endophytic microorganisms.
5.4 Quorum Sensing
Quorum sensing is a process of bacterial cell-cell communication and can be defined as the regulation of gene expression in response to chemical signalling molecules called autoinducers secreted by bacteria with respect to fluctuations in cell density (Miller and Bassler 2001). The accumulation of autoinducers in the environment denotes the increase in cell density of bacteria, and bacteria use this information to monitor changes in their cell numbers and alter gene expression accordingly (Rutherford and Bassler 2012). Quorum sensing controls several important activities of bacteria, including sporulation, antibiotic production, biofilm production, secretion of virulence factors, etc. (Williams and Cámara 2009).
A study was done on ToxR regulator of toxoflavin biosynthesis and transport in Burkholderia glumae, which causes rice grain rot and seedling rot in rice, and it was shown that the expression of ToxJ, a transcriptional activator, was controlled by the process of quorum sensing (Kim et al. 2004). Further, the study was extended by a group of scientists that engineered the Burkholderia glumae with an N-acyl-homoserine lactonase (aiiA) gene from Bacillus thuringiensis. The results of the study revealed that the introduced gene restricted the production of quorum sensing signals by Burkholderia glumae in vitro and reduced the occurrence of rice seedling rot caused by the pathogenic bacterium Burkholderia glumae in situ (Cho et al. 2007). A recent study also establishes the fact that quorum sensing is an important cell-cell communication process required for effective colonisation and establishment of a beneficial interaction by using a mutant strain of Burkholderia phytofirmans PsJN (Zúñiga et al. 2013).
5.5 Induced Systemic Resistance
Increasing resistance of plants to disease-causing microbes can help in protecting plants against a wide range of pathogens and for a sustained duration of time. When a plant is exposed to a pathogen (virulent or avirulent) or to metabolites secreted by pathogens, it results in the stimulation of the plant’s natural defence mechanisms resulting in immunisation of plants against bacterial, fungal or viral infections, even before the establishment of pathogenic infection (Bashan and De-Bashan 2005). Induced systemic resistance is a type of generalised resistance already present in plants but is induced or enhanced by plant-associated non-pathogenic bacteria (Van Loon et al. 1998). ISR depends on pathways governed by jasmonic acid or ethylene (Yan et al. 2002). In a study, induced systemic resistance was assessed using two Pseudomonas fluorescens strains (PF1 and PF7) against Rhizoctonia solani in rice. It was found that the treatment of rice with Pseudomonas led to the induction of induced systemic resistance against Rhizoctonia solani as a result of increase in chitinase and peroxidase activity. Also, 35 kDa chitinase along with three isozymes of peroxidases (PO3–PO5) was found to be responsible in the induction and establishment of ISR (Nandakumar et al. 2001). Similarly, six fluorescent Pseudomonas strains were used for the induction of induced systemic resistance in rice against Rhizoctonia solani possessing 2,4-diacetlyphloroglucinol (2,4-DAPG) antibiotic genes. The results of the study showed that 2,4-DAPG-positive strains of fluorescent Pseudomonas sp. produced peroxidase, polyphenol oxidase, phenylalanine ammonia lyase and transcinnamic acid that resulted in the induction of induced systemic resistance in rice against the rice sheath blight pathogen Rhizoctonia solani (Reshma et al. 2018). In a study, ISR was also induced in Oryza sativa using Pseudomonas fluorescens WCS374r against the leaf blast causing pathogen Magnaporthe oryzae. This study showed that induction of ISR was dependent on pseudobactin-mediated priming for a salicylic acid-repressible multifaceted defence response (De Vleesschauwer et al. 2008).
6 Seed Endophytes for Improving Phytoremediation of Soils
Phytoremediation may be defined as the removal of toxic contaminants from soil through the use of plants (White et al. 2003). Rhizo-degradation is an aspect of phytoremediation that deals with the use of plants to stimulate the microbial community present at the root-soil interface to augment the degradation of toxic contaminants in the soil (Newman and Reynolds 2005). The potential role of seed endophytes in the process of phytoremediation merits study as it is believed that some endophytes contain genetic machinery capable of degrading toxic molecules in the plant rhizospheric region and other contaminated sites (Siciliano et al. 2001). Although endophytes can play an important role in toxin degradation, not all the endophytic microbes have the ability to degrade all toxic compounds as well as have the potential to thrive in the given contaminant site; hence, genetically engineered microbes have been used which can degrade the toxins present and also survive in the contaminated site (Menn et al. 2008). The possible advantage of using endophytic microorganisms in phytoremediation strategies is that these organisms are easier to manipulate and take less time to genetically engineer. In addition to this, the assessment of the efficiency of phytoremediation process can easily be monitored by quantitative gene expression of pollutant catabolic genes within endophytic populations (Newman and Reynolds 2005). At Brookhaven National Laboratory, a genetically engineered plant endophyte was developed using genetic engineering to degrade toluene, a chemical that is highly toxic to plants as well as humans (Barac et al. 2004). Similarly, a transformed Burkholderia cepacia was produced using a conjugation process that was able to reduce toluene levels in the inoculated soils (Shields et al. 1995). However, the use of genetically engineered endophytic organisms requires resolution of social and ethical issues, and field trials must be conducted to determine the actual state of microbial action on the toxins, the survival of the engineered microbe, etc.
7 Metagenomic Studies on Rice Endophytes
A fraction of the endophytic microbes in seeds may be cultured from the seed-vectored community; therefore, there is a gap in knowledge about activities that these endophytes perform inside the seeds and seedlings (Tsurumaru et al. 2015). In addition to this, the mechanism of interaction between microorganisms is also not clearly understood. The recent advancements in genetic engineering tools such as rapid and inexpensive DNA sequencing have propelled large-scale genomic and metagenomic projects providing insights into the diversity of endophytes in seeds as well as the complexity of the community of endophytes within rice seeds (Brader et al. 2017). Metagenomic analysis relies on culture-independent molecular approaches like 16S rRNA libraries and thus helps in the study of many novel endophytic organisms which were previously impossible to culture in media or were present in a non-viable state (Lucas et al. 2013). A study was conducted in which six independent Gram-negative, facultatively anaerobic, non-spore-forming, nitrogen-fixing, rod-shaped isolates were isolated from the endosphere of rice and were characterised using genomic DNA-DNA hybridisation. The results established the existence of two new endophytic species of Enterobacter that were able to enhance plant growth by supplementing nitrogen and phosphorus (Hardoim et al. 2013). In another study, a culture-independent analysis was performed using 16S rDNA amplicons of the bacterial community of rice endosphere that provided important information on bacterial diversity in the endosphere (Bertani et al. 2016).
8 Conclusions
The bacterial and fungal species present within, and on the surface of, rice seeds form an important group of microflora. Seed endophytes play important roles in seed germination, seedling development and plant growth. These microbes possess plant growth promotion, biocontrol and phytoremediation properties. Many of the seed endophytes are cultivable, but others have unknown cultivable conditions or are uncultivable; additional metagenomic studies on rice seed endophytes must be done to develop a more basic understanding about the diversity of total endophytic community. The functions of seed-vectored endophytes are still not fully understood. In this respect experiments are needed to develop an understanding of the functions and mechanisms of activity of endophytic microbes. Seed endophytes possess characteristics that can be exploited in various biotechnological applications owing to unique seed environment as well as the transmission to successive generations. Seed endophytes of rice are worthwhile studying for their potential to improve plant growth and development, stress tolerance, disease resistance and bioremediation. The potential for increasing rice productivity to meet the demand of the ever-increasing world population makes studies on endophytes an attractive area. The application of seed endophytes in industries, agriculture and environment requires a more detailed study to elucidate the functions of endophytes during seed germination and plant development, the mechanisms through which they control and modulate plant growth and their interactions with other endophytes.
References
Arnold AE, Maynard Z et al (2000) Are tropical fungal endophytes hyperdiverse? Ecol Lett 3(4):267–274
Arnold AE, Mejía LC et al (2003) Fungal endophytes limit pathogen damage in a tropical tree. Proc Natl Acad Sci USA 100(26):15649–15654
Atugala DM, Deshappriya N (2015) Effect of endophytic fungi on plant growth and blast disease incidence of two traditional rice varieties. J Natl Sci Found 43(2):173
Azevedo JL, Maccheroni W Jr et al (2000) Endophytic microorganisms: a review on insect control and recent advances on tropical plants. Electron J Biotechnol 3(1):15–16
Barac T, Taghavi S et al (2004) Engineered endophytic bacteria improve phytoremediation of water-soluble, volatile, organic pollutants. Nat Biotechnol 22(5):583
Barazani O, Friedman J (2001) Allelopathic bacteria and their impact on higher plants. Crit Rev Microbiol 27(1):41–55
Barrow JR, Osuna P (2002) Phosphorus solubilization and uptake by dark septate fungi in fourwing saltbush, Atriplex canescens (Pursh) Nutt. J Arid Environ 51(3):449–459
Bashan Y, De-Bashan LE (2005) Plant growth-promoting. In: Encyclopedia of soils in the environment, vol 1. Elsevier, Oxford, pp 103–115
Beltran-Garcia MJ, White JF Jr et al (2014) Nitrogen acquisition in Agave tequilana from degradation of endophytic bacteria. Sci Rep 4:6938
Bertani I, Abbruscato P et al (2016) Rice bacterial endophytes: isolation of a collection, identification of beneficial strains and microbiome analysis. Environ Microbiol Rep 8(3):388–398
Boddey RM, Oliveira OC et al (1995) Biological nitrogen fixation associated with sugar cane and rice: contributions and prospects for improvement. Plant Soil 174(1):195–209
Brader G, Corretto E et al (2017) Metagenomics of plant microbiomes. In: Functional metagenomics: tools and applications. Springer, Cham, pp 179–200
Carroll GC (1991) Fungal associates of woody plants as insect antagonists in leaves and stems. In: Microbial mediation of plant-herbivore interactions. Wiley, New York, pp 253–271
Chatterjee A, Valasubramanian R et al (1996) Isolation of ant mutants of Pseudomonas fluorescens strain Pf7-14 altered in antibiotic production, cloning of ant+ DNA, and evaluation of the role of antibiotic production in the control of blast and sheath blight of rice. Biol Control 7(2):185–195
Cheplick GP, Clay K et al (1989) Interactions between infection by endophytic fungi and nutrient limitation in the grasses Lolium perenne and Festuca arundinacea. New Phytol 111(1):89–97
Cho SJ, Lim WJ et al (2003) Endophytic colonization of balloon flower by antifungal strain Bacillus sp. CY22. Biosci Biotechnol Biochem 67(10):2132–2138
Cho HS, Park SY et al (2007) Interference of quorum sensing and virulence of the rice pathogen Burkholderia glumae by an engineered endophytic bacterium. FEMS Microbiol Ecol 60(1):14–23
Clay K (1989) Clavicipitaceous endophytes of grasses: their potential as biocontrol agents. Mycol Res 92(1):1–12
Compant S, Clément C et al (2010) Plant growth-promoting bacteria in the rhizo-and endosphere of plants: their role, colonization, mechanisms involved and prospects for utilization. Soil Biol Biochem 42(5):669–678
Cottyn B, Regalado E et al (2001) Bacterial populations associated with rice seed in the tropical environment. Phytopathology 91(3):282–292
De Vleesschauwer D, Djavaheri M et al (2008) Pseudomonas fluorescens WCS374r-induced systemic resistance in rice against Magnaporthe oryzae is based on pseudobactin-mediated priming for a salicylic acid-repressible multifaceted defense response. Plant Physiol 148(4):1996–2012
Doty SL, Oakley B et al (2009) Diazotrophic endophytes of native black cottonwood and willow. Symbiosis 47(1):23–33
Dunne C, Crowley JJ et al (1997) Biological control of Pythium ultimum by Stenotrophomonas maltophilia W81 is mediated by an extracellular proteolytic activity. Microbiology 143(12):3921–3931
Elbeltagy A, Nishioka K et al (2000) Isolation and characterization of endophytic bacteria from wild and traditionally cultivated rice varieties. Soil Sci Plant Nutr 46(3):617–629
Elbeltagy A, Nishioka K et al (2001) Endophytic colonization and in planta nitrogen fixation by a Herbaspirillum sp. isolated from wild rice species. Appl Environ Microbiol 67(11):5285–5293
Farooq M, Jabran K et al (2011) The role of allelopathy in agricultural pest management. Pest Manag Sci 67:493–506
Feng Y, Shen D et al (2006) Rice endophyte Pantoea agglomerans YS19 promotes host plant growth and affects allocations of host photosynthates. J Appl Microbiol 100(5):938–945
Fisher PJ, Petrini O (1992) Fungal saprobes and pathogens as endophytes of rice (Oryza sativa L.). New Phytol 120(1):137–143
Fravel DR (1988) Role of antibiosis in the biocontrol of plant diseases. Annu Rev Phytopathol 26(1):75–91
Freeman EM (1904) The seed-fungus of Lolium temulentum L., the darnel. Philos Trans R Soc Lond B 196(214–224):1–27
Gagne-Bourgue F, Aliferis KA et al (2013) Isolation and characterization of indigenous endophytic bacteria associated with leaves of switchgrass (Panicum virgatum L.) cultivars. J Appl Microbiol 114(3):836–853
Glass AD (1989) Plant mineral nutrition. An introduction to current concepts. Jones and Bartlett, Boston
Glick BR (1995) The enhancement of plant growth by free-living bacteria. Can J Microbiol 41(2):109–117
Glick BR (2005) Modulation of plant ethylene levels by the bacterial enzyme ACC deaminase. FEMS Microbiol Lett 251(1):1–7
Glick BR, Penrose DM et al (1998) A model for the lowering of plant ethylene concentrations by plant growth-promoting bacteria. J Theor Biol 190(1):63–68
Gond SK, Bergen MS, Torres MS, White Jr JF (2015) Endophytic bacillus spp. produce antifungal lipopeptides and induce host defence gene expression in maize. Microbiol Res 172:79–87
Gyaneshwar P, James EK et al (2001) Endophytic colonization of rice by a diazotrophic strain of Serratia marcescens. J Bacteriol 183:2634–2645
Hallmann J, Quadt-Hallmann A et al (1997) Bacterial endophytes in agricultural crops. Can J Microbiol 43(10):895–914
Hardoim PR, van Overbeek LS et al (2008) Properties of bacterial endophytes and their proposed role in plant growth. Trends Microbiol 16(10):463–471
Hardoim PR, Nazir R et al (2013) The new species Enterobacter oryziphilus sp. nov. and Enterobacter oryzendophyticus sp. nov. are key inhabitants of the endosphere of rice. BMC Microbiol 13(1):164
Hodgson S, Cates C et al (2014) Vertical transmission of fungal endophytes is widespread in forbs. Ecol Evol 4(8):1199–1208
Holguin G, Patten CL (1999) Biochemical and genetic mechanisms used by plant growth promoting bacteria. World Scientific, London
Hollis J (1949) Location of bacteria in healthy potato tissue. Phytopathology 39(1):9–10
Honma M, Shimomura T (1978) Metabolism of 1-aminocyclopropane-1-carboxylic acid. Agric Biol Chem 42(10):1825–1831
Hornok L (2000) Genetically modified microorganisms in biological control. Növényvédelem 36(5):229–237
Hoysted GA, Kowal J et al (2018) A mycorrhizal revolution. Curr Opin Plant Biol 44:1–6
Hurek T, Reinhold-Hurek B et al (1994) Root colonization and systemic spreading of Azoarcus sp. strain BH72 in grasses. J Bacteriol 176(7):1913–1923
Irizarry I, White JF (2017) Application of bacteria from non-cultivated plants to promote growth, alter root architecture and alleviate salt stress of cotton. J Appl Microbiol 122(4):1110–1120
Ji SH, Gururani MA et al (2014) Isolation and characterization of plant growth promoting endophytic diazotrophic bacteria from Korean rice cultivars. Microbiol Res 169(1):83–98
Johnston-Monje D, Raizada MN (2011) Conservation and diversity of seed associated endophytes in Zea across boundaries of evolution, ethnography and ecology. PLoS One 6(6):e20396
Johnston-Monje D, Lundberg DS et al (2016) Bacterial populations in juvenile maize rhizospheres originate from both seed and soil. Plant Soil 405(1–2):337–355
Kaga H, Mano H, Tanaka F, Watanabe A, Kaneko S, Morisaki H (2009) Rice seeds as sources of endophytic bacteria. Microbes Environ 24(2):154–162
Kandel SL, Joubert PM et al (2017) Bacterial endophyte colonization and distribution within plants. Microorganisms 5(4):77
Kim J, Kim JG et al (2004) Quorum sensing and the LysR-type transcriptional activator ToxR regulate toxoflavin biosynthesis and transport in Burkholderia glumae. Mol Microbiol 54(4):921–934
Kong CH, Wang P et al (2008) Impact of allelochemical exuded from allelopathic rice on soil microbial community. Soil Biol Biochem 40(7):1862–1869
Krishnan P, Bhat R et al (2012) Isolation and functional characterization of bacterial endophytes from Carica papaya fruits. J Appl Microbiol 113(2):308–317
Kuldau G, Bacon C (2008) Clavicipitaceous endophytes: their ability to enhance resistance of grasses to multiple stresses. Biol Control 46(1):57–71
Lareen A, Burton F, Schäfer P (2016) Plant root-microbe communication in shaping root microbiomes. Plant Mol Biol 90:575–587
Larran S, Simon MR et al (2016) Endophytes from wheat as biocontrol agents against tan spot disease. Biol Control 92:17–23
Li HY, Wei DQ et al (2012) Endophytes and their role in phytoremediation. Fungal Divers 54(1):11–18
Liu Y, Zuo S et al (2012) Study on diversity of endophytic bacterial communities in seeds of hybrid maize and their parental lines. Arch Microbiol 194(12):1001–1012
Liu Y, Zuo S et al (2013) Investigation on diversity and population succession dynamics of endophytic bacteria from seeds of maize (Zea mays L., Nongda108) at different growth stages. Ann Microbiol 63(1):71–79
Loaces I, Ferrando L et al (2011) Dynamics, diversity and function of endophytic siderophore-producing bacteria in rice. Microb Ecol 61(3):606–618
Lodewyckx C, Vangronsveld J et al (2002) Endophytic bacteria and their potential applications. Crit Rev Plant Sci 21(6):583–606
Loganathan P, Nair S (2003) Crop-specific endophytic colonization by a novel, salt-tolerant, N2-fixing and phosphate-solubilizing Gluconacetobacter sp. from wild rice. Biotechnol Lett 25(6):497–501
Lopez DC (2015) Ecological roles of two entomopathogenic endophytes: Beauveria bassiana and Purpureocillium lilacinum in cultivated cotton. Texas A&M University
Lucas JA, García-Villaraco A et al (2013) Structural and functional study in the rhizosphere of Oryza sativa L. plants growing under biotic and abiotic stress. J Appl Microbiol 115(1):218–235
Luo S, Xu T et al (2012) Endophyte-assisted promotion of biomass production and metal-uptake of energy crop sweet sorghum by plant-growth-promoting endophyte Bacillus sp. SLS18. Appl Microbiol Biotechnol 93(4):1745–1753
Ma Y, Prasad MNV et al (2011) Plant growth promoting rhizobacteria and endophytes accelerate phytoremediation of metalliferous soils. Biotechnol Adv 29(2):248–258
Mano H, Morisaki H (2008) Endophytic bacteria in the rice plant. Microbes Environ 23:109–117
Mathan S (2016) Isolation of endophytic fungi from marine algae and its bioactivity. Int J Res Pharm Biomed Sci 4(1):45–49
Menn FM, Easter JP et al (2008) Genetically engineered microorganisms and bioremediation, 2nd ed. Biotechnology Set, pp 441–463
Miller MB, Bassler BL (2001) Quorum sensing in bacteria. Annu Rev Microbiol 55(1):165–199
Mitter B, Petric A et al (2013) Comparative genome analysis of Burkholderia phytofirmans PsJN reveals a wide spectrum of endophytic lifestyles based on interaction strategies with host plants. Front Plant Sci 4:120
Mukhopadhyay K, Garrison NK et al (1996) Identification and characterization of bacterial endophytes of rice. Mycopathologia 134(3):151–159
Murphy BR, Doohan FM et al (2015) Fungal root endophytes of a wild barley species increase yield in a nutrient-stressed barley cultivar. Symbiosis 65(1):1–7
Nandakumar R, Babu S et al (2001) Induction of systemic resistance in rice against sheath blight disease by Pseudomonas fluorescens. Soil Biol Biochem 33(4–5):603–612
Narula S, Anand RC et al (2013) Beneficial traits of endophytic bacteria from field pea nodules and plant growth promotion of field pea. J Food Legumes 26(3 and 4):73–79
Newman LA, Reynolds CM (2005) Bacteria and phytoremediation: new uses for endophytic bacteria in plants. Trends Biotechnol 23(1):6–8
Okunishi S, Sako K et al (2005) Bacterial flora of endophytes in the maturing seed of cultivated rice (Oryza sativa). Microbes Environ 20(3):168–177
Palumbo JD, Yuen GY et al (2005) Mutagenesis of β-1,3-glucanase genes in Lysobacter enzymogenes strain C3 results in reduced biological control activity toward Bipolaris leaf spot of tall fescue and Pythium damping-off of sugar beet. Phytopathology 95(6):701–707
Paungfoo-Lonhienne C, Rentsch D et al (2010) Turning the table: plants consume microbes as a source of nutrients. PLoS One 5:e11915
Perotti R (1926) On the limits of biological enquiry in soil science. Proc Int Soc Soil Sci 2:146–161
Pirttilä AM, Laukkanen H et al (2000) Detection of intracellular bacteria in the buds of Scotch pine (Pinus sylvestris L.) by in situ hybridization. Appl Environ Microbiol 66(7):3073–3077
Pirttilä AM, Pospiech H et al (2003) Two endophytic fungi in different tissues of Scots pine buds (Pinus sylvestris L.). Microb Ecol 45(1):53–62
Pleban S, Chernin L et al (1997) Chitinolytic activity of an endophytic strain of Bacillus cereus. Lett Appl Microbiol 25(4):284–288
Prieto KR, Echaide-Aquino F et al (2017) Endophytic bacteria and rare earth elements; promising candidates for nutrient use efficiency in plants. In: Plant macronutrient use efficiency: molecular and genomic perspectives in crop plants. Elsevier, pp 285–306
Reshma P, Naik MK et al (2018) Induced systemic resistance by 2,4-diacetylphloroglucinol positive fluorescent Pseudomonas strains against rice sheath blight. Indian J Exp Biol 56(3):207–212
Ringelberg D, Foley K et al (2012) Bacterial endophyte communities of two wheatgrass varieties following propagation in different growing media. Can J Microbiol 58(1):67–80
Rojas GJ et al (2016) Infection with a shoot-specific fungal endophyte (Epichloë) alters tall fescue soil microbial communities. Microb Ecol 72(1):197–206
Rosenblueth M, Martínez-Romero E (2006) Bacterial endophytes and their interactions with hosts. Mol Plant-Microbe Interact 19(8):827–837
Ruiza D, Agaras B et al (2011) Characterization and screening of plant probiotic traits of bacteria isolated from rice seeds cultivated in Argentina. J Microbiol 49(6):902–912
Rutherford ST, Bassler BL (2012) Bacterial quorum sensing: its role in virulence and possibilities for its control. Cold Spring Harb Perspect Med 2(11):a012427
Salisbury FB (1994) The role of plant hormones. Plant-environment interactions. Dekker, New York, pp 39–81
Saraf M, Pandya U et al (2014) Role of allelochemicals in plant growth promoting rhizobacteria for biocontrol of phytopathogens. Microbiol Res 169(1):18–29
Schulz B, Römmert AK et al (1999) The endophyte-host interaction: a balanced antagonism? Mycol Res 103(10):1275–1283
Shahzad R, Waqas M et al (2016) Seed-borne endophytic Bacillus amyloliquefaciens RWL-1 produces gibberellins and regulates endogenous phytohormones of Oryza sativa. Plant Physiol Biochem 106:236–243
Shearin ZR, Filipek M et al (2018) Fungal endophytes from seeds of invasive, non-native Phragmites australis and their potential role in germination and seedling growth. Plant Soil 422(1–2):183–194
Shehata HR, Ettinger CL et al (2016) Genes required for the anti-fungal activity of a bacterial endophyte isolated from a corn landrace grown continuously by subsistence farmers since 1000 BC. Front Microbiol 7:1548
Shields MS, Reagin MJ et al (1995) TOM, a new aromatic degradative plasmid from Burkholderia (Pseudomonas) cepacia G4. Appl Environ Microbiol 61(4):1352–1356
Siciliano SD, Fortin N et al (2001) Selection of specific endophytic bacterial genotypes by plants in response to soil contamination. Appl Environ Microbiol 67(6):2469–2475
Stevenson FJ, Cole MA (1999) Cycles of soils: carbon, nitrogen, phosphorus, sulfur, micronutrients. Wiley, New York
Sun Y, O’Riordan MX (2013) Regulation of bacterial pathogenesis by intestinal short-chain fatty acids. Adv Appl Microbiol 85:93–118
Sziderics AH, Rasche F et al (2007) Bacterial endophytes contribute to abiotic stress adaptation in pepper plants (Capsicum annuum L.). Can J Microbiol 53(11):1195–1202
Tholozan JL, Cappelier JM et al (1999) Physiological characterization of viable-but-nonculturable Campylobacter jejuni cells. Appl Environ Microbiol 65(3):1110–1116
Tramontano WA, Scanlon C (1996) Cell cycle inhibition by butyrate in legume root meristems. Phytochemistry 41:85–88
Truyens S, Weyens N et al (2015) Bacterial seed endophytes: genera, vertical transmission and interaction with plants. Environ Microbiol 7(1):40–50
Tsurumaru H, Okubo T et al (2015) Metagenomic analysis of the bacterial community associated with the taproot of sugar beet. Microbes Environ 30(1):63–69
Vallad GE, Goodman RM (2004) Systemic acquired resistance and induced systemic resistance in conventional agriculture. J Trop Crop Sci 44(6):1920–1934
Van Loon LC, Bakker PAHM et al (1998) Systemic resistance induced by rhizosphere bacteria. Annu Rev Phytopathol 36(1):453–483
Verma SC, Ladha JK et al (2001) Evaluation of plant growth promoting and colonization ability of endophytic diazotrophs from deep water rice. J Biotechnol 91(2–3):127–141
Verma SK, Gond SK et al (2017a) Fungal endophytes representing diverse habitats and their role in plant protection. In: Developments in fungal biology and applied mycology. Springer, Singapore, pp 135–157
Verma SK, Kingsley K et al (2017b) Seed vectored endophytic bacteria modulate development of rice seedlings. J Appl Microbiol 122(6):1680–1691
Verma SK, White JF (2018) Indigenous endophytic seed bacteria promote seedling development and defend against fungal disease in browntop millet (L.). J Appl Microbiol 124(3):764–778
Verma SK, Kingsley K et al (2018) Bacterial endophytes from rice cut grass (Leersia oryzoides L.) increase growth, promote root gravitropic response, stimulate root hair formation, and protect rice seedlings from disease. Plant Soil 422(1–2):223–238
Vessey JK (2003) Plant growth promoting rhizobacteria as biofertilizers. Plant Soil 255(2):571–586
Vujanovic V, Germida JJ (2017) Seed endosymbiosis: a vital relationship in providing prenatal care to plants. Can J Plant Sci 97(6):972–981
Walitang DI, Kim K et al (2017) Characterizing endophytic competence and plant growth promotion of bacterial endophytes inhabiting the seed endosphere of Rice. BMC Microbiol 17(1):209
White JF (2017) Syntrophic imbalance and the etiology of bacterial endoparasitism diseases. Med Hypotheses 107:14–15
White JC, Wang X et al (2003) Subspecies-level variation in the phytoextraction of weathered p,p′-DDE by Cucurbita pepo. Environ Sci Technol 37(19):4368–4373
White JF, Chen Q et al (2015) Collaboration between grass seedlings and rhizobacteria to scavenge organic nitrogen in soils. AoB Plants 7
White J F, Kingsley K L et al (2018) Reactive oxygen defense against cellular endoparasites and the origin of eukaryotes. Transformative paleobotany: papers to commemorate the life and legacy of Thomas N. Taylor. Elsevier, Amsterdam
White JF, Torres MS et al (2019) Evidence for widespread microbivory of endophytic bacteria in roots of vascular plants through oxidative degradation in root cell periplasmic spaces. In: PGPR amelioration in sustainable agriculture: food security and environmental management. Elsevier, London
Wijesooriya WADK, Deshappriya N (2016) An inoculum of endophytic fungi for improved growth of a traditional rice variety in Sri Lanka. Trop Plant Res 3(3):470–480
Williams P, Cámara M (2009) Quorum sensing and environmental adaptation in Pseudomonas aeruginosa: a tale of regulatory networks and multifunctional signal molecules. Curr Opin Microbiol 12(2):182–191
Wilson D (1995) Endophyte: the evolution of a term, and clarification of its use and definition. Oikos 73:274–276
Yan Z, Reddy MS et al (2002) Induced systemic protection against tomato late blight elicited by plant growth-promoting rhizobacteria. Phytopathology 92(12):1329–1333
Yumlembam RA, Borkar SG (2014) Assessment of antibacterial properties of medicinal plants having bacterial leaf endophytes against plant pathogenic Xanthomonads. Indian Phytopathol 67(4):353–357
Zhang T, Yao YF (2015) Endophytic fungal communities associated with vascular plants in the high arctic zone are highly diverse and host-plant specific. PLoS One 10(6):e0130051
Zúñiga A, Poupin MJ et al (2013) Quorum sensing and indole-3-acetic acid degradation play a role in colonization and plant growth promotion of Arabidopsis thaliana by Burkholderia phytofirmans PsJN. Mol Plant-Microbe Interact 26(5):546–553
Acknowledgement
The authors are grateful to the Department of Plant Biology, Rutgers University, NJ, for research facilities. SKV thanks to UGC, India, for providing a Raman Post Doctoral Fellowship No. F 5-11/2016(IC) for the year 2016–2017 to work in the USA and support as grant, Project-UGC-BSR startup-M14-26. The SKV and RNK are also grateful to the Head and Coordinator of CAS and DST-FIST and PURSE of Botany, BHU, Varanasi, for providing facilities and leave to pursue endophyte research. The authors are also thankful for support from the John E. and Christina C. Craighead Foundation, USDA-NIFA Multistate Project W3147, and the New Jersey Agricultural Experiment Station.
Author information
Authors and Affiliations
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2019 Springer Nature Switzerland AG
About this chapter

Cite this chapter
Pal, G., Kumar, K., Verma, A., White, J.F., Verma, S.K. (2019). Functional Roles of Seed-Inhabiting Endophytes of Rice. In: Verma, S., White, Jr, J. (eds) Seed Endophytes. Springer, Cham. https://doi.org/10.1007/978-3-030-10504-4_11
Download citation
DOI: https://doi.org/10.1007/978-3-030-10504-4_11
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-10503-7
Online ISBN: 978-3-030-10504-4
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)