
Abstract
The effects of Bacillus sp. SLS18, a plant-growth-promoting endophyte, on the biomass production and Mn/Cd uptake of sweet sorghum (Sorghum bicolor L.), Phytolacca acinosa Roxb., and Solanum nigrum L. were investigated. SLS18 displayed multiple heavy metals and antibiotics resistances. The strain also exhibited the capacity of producing indole-3-acetic acid, siderophores, and 1-aminocyclopropane-1-carboxylic acid deaminase. In pot experiments, SLS18 could not only infect plants effectively but also significantly increase the biomass of the three tested plants in the presence of Mn/Cd. The promoting effect order of SLS18 on the biomass of the tested plants was sweet sorghum > P. acinosa > S. nigrum L. In the presence of Mn (2,000 mg kg−1) and Cd (50 mg kg−1) in vermiculite, the total Mn/Cd uptakes in the aerial parts of sweet sorghum, P. acinosa, and S. nigrum L. were increased by 65.2%/40.0%, 55.2%/31.1%, and 18.6%/25.6%, respectively, compared to the uninoculated controls. This demonstrates that the symbiont of SLS18 and sweet sorghum has the potential of improving sweet sorghum biomass production and its total metal uptake on heavy metal-polluted marginal land. It offers the potential that heavy metal-polluted marginal land could be utilized in planting sweet sorghum as biofuel feedstock for ethanol production, which not only gives a promising phytoremediation strategy but also eases the competition for limited fertile farmland between energy crops and food crops.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Heavy metal contamination, caused by multiple human technogenic activities, poses a serious threat to human health and ecosystem (Abou-Shanab et al. 2006). Manganese (Mn) is an essential micronutrient and an activator for many enzymes (Xue et al. 2004), but exposure to excessive Mn leads to Mn poisoning, such as Parkinson symptoms (Gerber et al. 2002) and abnormalities of the immune system (Vartanian et al. 1999). Cadmium (Cd) is not only a non-essential element for biosynthetic metabolism but also toxic to living beings when accumulated. Therefore, the development of methods to lessen the harm of Mn and Cd in soil to public health and organisms has become a worldwide environmental concern. Because of less cost and being safer for human and environment, phytoremediation has more advantages than conventional technologies of removing heavy metals from soil (Krämer 2005; Dowling and Doty 2009; Glick 2010). In recent years, the concentration of heavy metals in lots of terrestrial plants has been researched (Xue et al. 2004). Some of those plants have been defined as hyperaccumulators that possess the ability to accumulate large quantities of heavy metals in their biomass without hindrance to their growth and development (Wu et al. 2006). Nevertheless, low biomass of hyperaccumulators and low bioavailability of heavy metals in soil limit the efficiency of phytoremediation by hyperaccumulators (Sheng and Xia 2006; Ma et al. 2011).
This has encouraged researchers to seek for the possible strategy to improve the efficiency of phytoremediation. Recent attention has been paid on the partnership between plants and plant-growth-promoting endophytes (PGPEs), which can effectively carry forward the application of phytoremediation by hyperaccumulators (Sheng et al. 2008b; Chen et al. 2010). The PGPEs assist their host plants in coping with contaminant-induced stress and improving plant growth through various mechanisms, such as indole-3-acetic acid (IAA), siderophores, and 1-aminocyclopropane-1-carboxylic acid (ACC) deaminase production, nitrogen fixation, and phosphates solution (Rajkumar et al. 2009; Ma et al. 2011). On the other hand, these mechanisms are of paramount importance for the use of plants as stocks of biofuels and carbon sequestration through biomass production (Taghavi et al. 2009).
It is well known that the competition of agricultural resources between energy crop and food crop is one of the critical socioeconomic issues with the increasing use of biofuels (Taghavi et al. 2009). This dilemma could be bypassed through producing biofuel feedstocks on marginal land (comprising soil that is infertile, arid, or has been contaminated by toxic heavy metals and organic pollutants), which is not suitable for agriculture (Weyens et al. 2009a). Bacterial endophytes, the most closely plant-associated bacteria colonizing the internal tissues of plants, could be of great value in enabling energy plants to grow better on marginal land for promoting the production of biofuels stocks. On account of recent economic analysis, sweet sorghum (Sorghum bicolor L.) is not only considered to be one of the most drought resistant energy crops but also equipped with higher biomass yield and photosynthetic efficiency and lower production costs than many other energy plants (Rubin 2008; Corredor et al. 2009; Li et al. 2010). Sweet sorghum contains quantities of soluble carbohydrates of glucose and sucrose and has been alternative as an important stock for the production of fuel ethanol (Jasberg et al. 1983; Mamma et al. 1995). Furthermore, sweet sorghum could also remove heavy metals such as Cd, Pb, and Cu from soil (Zhuang et al. 2009; Epelde et al. 2009). Therefore, as an energy crop (Bennett and Anex 2009; Linton et al. 2011) that can grow on marginal land contaminated by heavy metals, sweet sorghum is a good candidate for biofuel feedstock production and phytoremediation. However, little attention has been paid to the potential of heavy-metal-resistant PGPEs originating from hyperaccumulator to facilitate biomass production and heavy metal phytoremediation of sweet sorghum on heavy metals contaminated land. It is significant to explore whether excellent bacterial endophytic isolated from hyperaccumulator could successfully colonize and be beneficial to sweet sorghum.
In this study, an endophytic bacterial strain SLS18 with multiple heavy metals resistances and plant-growth-promoting traits was applied to inoculation with sweet sorghum for promoting biomass production and heavy metals phytoremediation. Two hyperaccumulators Phytolacca acinosa Roxb. (pokeberry) and Solanum nigrum L. (black nightshade) were also inoculated as comparison.
Materials and methods
Isolation of heavy-metal-resistant PGPE
Endophytic bacteria were isolated using a surface sterilization-trituration-plating technique from Mn-hyperaccumulator P. acinosa collected at a mine wasteland in Hengyang, Hunan Province, China, according to the previous protocol (Chen et al. 2010). An isolated bacterium from the stem of P. acinosa, designated as SLS18 with multiple heavy metals resistance and plant-growth-promoting traits, was selected for further studies.
PGP traits and carbon sources utilization of SLS18
The production of IAA by SLS18 was determined according to the method of Sheng et al. (2008a). Strain was cultured for 4 days at 30°C in the shaking sucrose-minimal salts (SMS) medium supplemented with 0.5 mg mL−1 of l-tryptophan. The concentration of IAA in culture was determined by the Salkowski’s reagent (Gordon and Weber 1951) and a calibration curve of pure IAA as a standard for linear regression analysis (Sheng et al. 2008a). Production of siderophores by strain SLS18 was detected according to the chrome azurol-S (CAS) analytical method (Schwyn and Neilands 1987). After incubation on CAS agar plate at 30°C for 4 days, appearance of orange halos around the colonies on plates demonstrated siderophores production. ACC deaminase activity was evaluated as the ability to grow in the sterile minimal medium, DF salts (Dworkin and Foster 1958) with ACC as the sole nitrogen source (Penrose and Glick 2003). The uninoculated DF medium was for control. After cultivation for 24 h, the bacterial growth was positive when the culture medium became turbid, demonstrating that the strain had ACC deaminase activity.
In order to examine autotrophy and nitrogen fixation, bacteria were grown in shaking Schatz minimal salt medium (Schatz and Bovell 1952) at 30°C for 3 days without any carbon or nitrogen sources, respectively. The bacterial growth was positive when the culture medium became turbid, proving that SLS18 had the capability of autotrophy and nitrogen fixation. Besides, to assess the potential of the strain SLS18 to thrive within the sweet sorghum interior, the utilization of carbon sources by SLS18 was tested in shaking Schatz medium containing different carbon nutrient sources (2 g L−1) at 30°C for 3 days. Two of these carbon sources, glucose and sucrose (soluble carbohydrates), were abundantly contained in sweet sorghum (Mamma et al. 1995). In this experiment, as positive control, strain was inoculated in Schatz minimal salt medium supplemented with C-mix (1 L of medium: 1.3 mL glucose 40%, 0.7 mL lactate 50%, 2.2 mL gluconate 30%, 2.7 mL fructose 20%, and 3 mL 1 M succinate) (Taghavi et al. 2009).
Heavy metal and antibiotic resistances of SLS18
Heavy metal and antibiotic resistances of SLS18 were evaluated by measuring the minimal inhibitory concentration of heavy metal ions and antibiotics. SLS18 was monitored for the growth under increasing concentrations of various metal ions and antibiotics, e.g., Cu2+ (0–300 mg L−1), Zn2+ (0–300 mg L−1), Cd2+ (0–50 mg L−1), Pb2+ (0–600 mg L−1), Mn2+ (0–3,000 mg L−1), kanamycin (0–50 μg mL−1), streptomycin (0–200 μg mL−1), rifampin (0–50 μg mL−1), tetracycline (0–50 μg mL−1), and ampicillin (0–100 μg mL−1) by sterile Luria–Betani (LB) agar medium. Plates were checked after incubation at 30°C for 7 days.
Molecular characterization of SLS18
DNA was extracted from the strain SLS18 after overnight cultivation in LB medium according to the method of Araújo et al. (2002). The obtained 16S rDNA, as a template, was amplified by PCR using universal bacterial 16S rDNA primers, 27f (5′-AGAGTTTGATCACTGGCTCAG-3′) and 1495r (5′-CTACGGCTACCTTGTTACGA-3′). The PCR was performed using an initial denaturing step performed at 94°C for 10 min, followed by 28 cycles of 50 s at 94°C, 50 s of annealing at 52°C, and a 65 s extension at 72°C, and a final polymerization step of 72°C for 10 min with a DNA Engine Thermal Cycler (PC-512, Bibby, USA). The PCR products of 16S rDNA fragments were purified with a DNA purification kit (KeyGEN, China) and sequenced at Invitrogen Company (Shanghai, China). The 16S rDNA sequence was compared against the GenBank database sequences with BLASTN tool in the National Center for Biotechnology Information (NCBI) website.
Pot experiments
The effects of SLS18 on plant growth and Mn/Cd uptake of sweet sorghum, P. acinosa, and S. nigrum L. were studied by pot experiments. Plastic pots were filled with 300 g sterile vermiculite containing Mn2+ (0, 1,000 and 2,000 mg kg−1) and Cd2+ (0 and 50 mg kg−1) and then moistened with sterile one-fourth Hoagland’s nutrient solution (Hoagland and Arnon 1950).
The seeds of sweet sorghum, P. acinosa, and S. nigrum L. were surface sterilized in 70% ethanol for 1 min, followed by another period of 10 min in 1% sodium hypochlorite, and then rinsed with sterile water. To confirm seed sterility, five seeds were incubated to LB agar for 3 days at 30°C. SLS18 was cultured in liquid LB medium for 24 h at 30°C. The resulting cells were collected through being centrifuged, washed, re-centrifuged, and re-suspended in sterile distilled water to get a density of 5 × 108 cfu mL−1. After that, surface-sterilized seeds were immersed in bacterial suspension for 4 h, while the seeds were soaked in sterile water as the control. Five seeds inoculated with strain SLS18 of each plant were sown in every pot, and three replicates were conducted for each treatment. Finally, all pots were deposited in a green house under controlled light (14 h photoperiod, PAR of 300 μmol m−2 s−1) and temperature (28/25°C day/night) and irrigated with sterile one-fourth Hoagland’s solution every 2 days.
After 2 weeks, the plants were thinned to one plant per pot, and all plants were carefully removed from pots after growing 8 weeks. The aerial part tissues and roots were separated, rinsed repeatedly, and dried at 105°C before surveying the dry weight of plant tissues. The concentrations of Mn and Cd in the aerial part and root of plants were detected according to the method of Sheng et al. (2008a) by an atomic absorption spectrometer (Z-2,000 Polarized Zeaman atomic absorption spectrophotometer, Japan). To assure the measurement accuracy and precision, blank test sample and duplicates were used.
Verification of inoculation
To verify whether the inoculation of SLS18 was successful or not, endophytic bacteria were re-isolated from the aerial parts and roots of sweet sorghum, P. acinosa, and S. nigrum L. Briefly, healthy plant tissues were surface sterilized according to the method of the above seeds surface sterilization. Aerial part materials (1 g) and root materials (0.5 g) were ground in 5 mL sterile 10 mM MgSO4 aqueous solution in a mortar with a pestle. The samples and their dilutions were spread on plates containing LB medium. After incubation for 4 days at 30°C, the re-isolated strains were identified by colony characteristics and sequenced against the 16S rDNA sequence of SLS18 inoculated to seeds.
Statistical analysis
All statistical analysis was conducted using SPSS 10.0 package. Analysis of variance, independent samples test, and the Student–Newman–Keuls test (p < 0.05) were used to compare treatment.
Results
Characterization of SLS18
The strain SLS18 was assayed for a number of properties that were important for plant-growth-promoting activity (Table 1). SLS18 possessed the ability to secrete IAA (13.7 ± 2.3 μg mL−1) after cultivation for 4 days at 30°C in the shaking SMS medium with L-tryptophane. The emergence of orange halos around the bacterial colonies of the strain SLS18 demonstrated siderophore production. Moreover, SLS18 was capable of utilizing ACC as the sole nitrogen source, demonstrating that the strain was also qualified with ACC deaminase activity. Additionally, the ability of SLS18 to utilize different carbon sources was tested. The result showed that SLS18 could not grow autotrophically and was unable to fix nitrogen. The bacterial strain SLS18 also exhibited different multiple heavy metals (Mn, Cd, Pb, Cu, and Zn) and antibiotics (kanamycin, streptomycin, rifampin, and ampicillin) resistant characteristics.
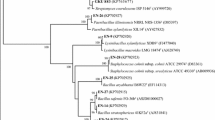
The partial sequence (1,437 bp) of 16S rDNA of SLS18 obtained was matched against nucleotide sequences present in GenBank using the BLASTn program. The highest sequence similarity (100%) clearly indicated that SLS18 belonged to Bacillus sp. The sequence has been deposited in the NCBI database under accession number HQ416900. In addition, the strain was deposited in the China Center for Type Culture Collection under accession number M2010359.
Plant growth promotion of SLS18
After inoculation of SLS18 or not, the three tested plants were subjected to different levels of Mn/Cd treatment for 10 weeks. The presence of 50 mg kg−1 Cd significantly (p < 0.05) decreased the aerial part dry weight of sweet sorghum and S. nigrum L.; the presence 2,000 mg kg−1 of Mn significantly decreased the aerial part dry weight of sweet sorghum (Fig. 1). However, the presence of 1,000 mg kg−1 Mn significantly increased the aerial part dry weight of sweet sorghum and the root dry weight of S. nigrum L. (Fig. 1). It might be attributed to the fact that Mn is an essential micronutrient, and appropriate amount of Mn ions is beneficial to plant growth. Meanwhile, the root dry weight of sweet sorghum and P. acinosa had no significant variation (p < 0.05) when exposed to Mn/Cd. The effect of SLS18 on the dry weight of the tested plants in the presence of Mn/Cd is shown in Fig. 1. In the absence of heavy metals and the presence of Mn (1,000 and 2,000 mg kg−1) and Cd (50 mg kg−1) in vermiculite, significant increases of aerial parts and root dry weights of all three plants were observed when the tested plants were inoculated with SLS18, compared with non-inoculated control (Fig. 1). Results revealed that the dry weights of aerial part and root for sweet sorghum increased by 46.8% and 77.7% (absent of metal), 37.0% and 74.4% (1,000 mg kg−1 Mn), 45.5% and 81% (2,000 mg kg−1 Mn), and 38.0% and 80.3% (50 mg kg−1 Cd) compared to non-inoculated control, respectively. The promoting effect order of SLS18 on biomass of the tested plants was sweet sorghum > P. acinosa > S. nigrum L.

Effects of SLS18 on the aerial part a and root b biomass (mg) of the tested plants in the presence of Mn (0, 1,000, and 2,000 mg kg−1) and Cd (0 and50 mg kg−1) in vermiculite. Each value is the mean of triplicates. Error bars represent standard deviation. An asterisk denotes that there is significant difference between the value with SLS18 and the value without SLS18. Containing any of the same letters for the same plant tissue indicates that the values have no significant difference within different heavy metal concentrations (p < 0.05, ANOVA; SNK test)
Effect of SLS18 on Mn/Cd uptake by plants
The effects of SLS18 on Mn/Cd concentration in sweet sorghum, P. acinosa, and S. nigrum L. tissues are shown in Table 2. Compared with non-inoculated control, no significant increase in Mn concentrations in S. nigrum L., but significant increases (p < 0.05) of Mn concentrations in sweet sorghum and P. acinosa were observed when the tested plants were inoculated with SLS18. However, no significant increase in Cd concentrations in all the tested plants was found when the tested plants were inoculated with SLS18, compared with control. The total Mn/Cd uptakes in aerial part of the symbionts of the plants and SLS18 were improved by 65.2%/40.0%, 55.2%/31.1%, and 18.6%/25.6% in sweet sorghum, P. acinosa, and S. nigrum L. in the presence of 2,000 mg kg−1 Mn or 50 mg kg−1Cd, respectively.
Establishment and survive of SLS18 in plant tissue
SLS18 was re-isolated to examine the survival rate of the colonized bacteria in the interior tissue of all the tested plants growing under different concentrations of heavy metals after seed inoculation with SLS18. The 16S rDNA sequence of the re-isolated strain that has the same colony characteristics of SLS18 was 100% identical with its parent strain. It was necessary to state that SLS18 was not detected in the non-inoculated plants. Colonization of SLS18 in the interior tissue of tested plants growing under different concentrations of heavy metals was determined (Table 3). The presence of Cd significantly inhibited the survival of SLS18 in the three tested plants. Except for the root of P. acinosa, the presence of 2,000 mg kg−1 Mn significantly decreased the growth of SLS18 in the three tested plants. No significant effect of 1,000 mg kg−1 Mn on the survival of SLS18 was found. In addition, bacterial survival was better in sweet sorghum and P. acinosa than in S. nigrum L., and the bacterial densities in aerial part were lower than those in root.
Discussion
During the long period of evolution with plants, some bacterial endophytes have formed a beneficial mutualistic symbiosis relationship with their host plants. Endophytes profit from host plants because of the enhanced availability of nutrients, and plants can also receive benefits from endophytes by growth enhancement or stress reduction (Hardoim et al. 2008; Weyens et al. 2009b). The SLS18 possessed a number of properties that are important for plant-growth-promoting activities, containing the ability of IAA, siderophore, and ACC deaminase production (Table 1). The presence of more than one PGP trait in this bacterial endophyte can facilitate the growth of plant by utilizing one or more above mechanisms at different times during the life cycle of host plant (Rajkumar et al. 2009). SLS18 seemed adapted to utilize a broad spectrum of plant-derived compounds as carbon sources, such as d-mannitol, lactose, sucrose, xylose, glucose, and fructose (Table 1). The strain SLS18 was able to utilize sucrose and glucose, which were typical soluble carbohydrates in sweet sorghum (Mamma et al. 1995). It has been reported that endophytes were able to utilize carbon sources in the host plant to thrive within the host plant (Taghavi et al. 2009), so SLS18 probably had the potential to survive within sweet sorghum interior. In natural environment, there is an abundance of endophyte species in the inter-tissue of plants. Some of the endophytes may secrete antibiotics against plant pathogen for protecting their host plants and against other endophytic fungi and bacteria for enhancing their own competitiveness (Strobel 2003). The bacterial strain SLS18 exhibited different multiple antibiotics as well as heavy-metal-resistant characteristics (Table 1). For this reason, the bacterial endophyte could survive in interior of Mn-hyperaccumulator P. acinosa, which accumulated a huge number of heavy metals and might contain antibiotics from other endophytes. Previous reports proved that the endophytic bacteria isolated from metal hyperaccumulator plants had the tolerance to high metal concentration in hyperaccumulators to adapt to such environmental conditions (Idris et al. 2004). Furthermore, if being considered as excellent plant-growth-promoting inoculants, bacteria must be capable of colonizing successfully in the interior tissue of plants (Sheng et al. 2008a). The effective re-isolation of SLS18 suggested that the strain could successfully colonize in both aerial part and root of the three tested plants (Table 3).
The association of endophytic bacteria with their hosts has beneficial effects for many different plant species (Taghavi et al. 2009). In this research, biomass of the three tested plants increased when being inoculated with SLS18 in the presence of Mn/Cd (Fig. 1). SLS18 had the capacity of facilitating the growth of plants, which might be relevant to its ability to produce IAA, siderophore, and ACC deaminase, in agreement with the reported results (Rajkumar et al. 2006; Rajkumar et al. 2009). The IAA produced by PGPE can enhance plant growth through altering the plant hormonal balance (Chen et al. 2010). It was also reported that PGPE could synthesize IAA to strengthen its host plant’s growth (Sheng et al. 2008b; Jha and Kumar 2009; Shi et al. 2010). Bacterial siderophores production, which may be stimulated by the presence of heavy metals, can help plant to reduce heavy metal toxicity by increasing the supply of iron to the plant (Burd et al. 2000) and also form some complexes with other bivalent heavy metal ions, which can be assimilated by the plant (Evers et al. 1989). Biomass increase was observed for mung plants inoculated with siderophore-producing Pseudomonas putida KNP9 in the presence of Cd (Tripathi et al. 2005). Bacteria with ACC deaminase activity could efficiently ease plant stress by decreasing excess ethylene biosynthesis, which might inhibit plant development and thus facilitate the growth and development of plant (Chen et al. 2010). For example, bacteria containing ACC deaminase significantly improved the growth of maize and tomato plants (Jiang et al. 2008). Meaningfully, the promoting effect order of SLS18 on biomass of the tested plants was sweet sorghum > P. acinosa > S. nigrum L. One possible explanation was that the detoxification ability of sweet sorghum was weaker than those of hyperaccumulators, so the growth-promoting ability of SLS18 played a more remarkable role in the growth of sweet sorghum in the presence of heavy metals. In addition, the strain SLS18 was an endophytic bacterium originally isolated from P. acinosa, so it might interact more effectively with P. acinosa than S. nigrum L. when re-colonizing to plant. Therefore, SLS18 has the potential to facilitate the growth of sweet sorghum on metal-contaminated marginal lands. The dry weight of sweet sorghum was higher than that of the two hyperaccumulators in the presence of Mn/Cd (Fig. 1).
After SLS18 inoculation, the phytoremediation efficiency of the tested plants could be promoted mainly through the increases in plant biomass (Fig. 1) and partly through the increases in metal uptake in plants (Table 2). Low production of plants biomass and low bioavailability of heavy metals in the soil restrict the efficiency of phytoremediation by hyperaccumulators. Some previous studies have confirmed the importance of the inoculation of heavy-metal-resistant bacteria for promoting plant growth and facilitating phytoremediation in heavy-metal-contaminated soils (Sheng et al. 2008a; Mastretta et al. 2009; Compant et al. 2010). In this study, SLS18 inoculation significantly facilitated phytoremediation efficiency of sweet sorghum. This reveals that SLS18 has the potential of practical application in increasing the biomass of sweet sorghum for biofuel stock production on marginal land, as well as the phytoremediation of heavy metal polluted marginal land.
In conclusion, heavy-metal-resistant PGPE Bacillus sp. SLS18 could not only successfully infect sweet sorghum, P. acinosa, and S. nigrum L. but also increase the growth and Mn/Cd accumulation of the three tested plants in the presence of Mn/Cd. Furthermore, the promoting effect order of SLS18 on biomass of the tested plants was sweet sorghum > P. acinosa > S. nigrum L. These results suggest that the symbiont of SLS18 and sweet sorghum has the potential of promoting the biomass production and heavy metal-polluted soil remediation effect of sweet sorghum on heavy metal polluted marginal land.
References
Abou-Shanab RAI, Angle JS, Chaney RL (2006) Bacterial inoculants affecting nickel uptake by Alyssum murale from low, moderate and high Ni soils. Soil Biol Biochem 38:2882–2889
Araújo WL, Marcon J, Maccheroni W Jr, van Elsas JD, van Vuurde JWL, Azevedo JL (2002) Diversity of endophytic bacterial populations and their interaction with Xylella fastidiosa in citrus plants. Appl Environ Microbiol 68:4906–4914
Bennett AS, Anex RP (2009) Production, transportation and milling costs of sweet sorghum as a feedstock for centralized bioethanol production in the upper Midwest. Bioresource Technol 100:1595–1607
Burd GI, Dixon DG, Glick BR (2000) Plant growth-promoting bacteria that decrease heavy metal toxicity in plants. Can J Microbiol 46:237–245
Chen L, Luo SL, Xiao X, Guo HJ, Chen JL, Wan Y, Li B, Xu TY, Xi Q, Rao C, Liu CB, Zeng GM (2010) Application of plant growth-promoting endophytes (PGPE) isolated from Solanum nigrum L. for phytoextraction of Cd-polluted soils. Appl Soil Ecol 46:383–389
Compant S, Clément C, Sessitsch A (2010) Plant growth-promoting bacteria in the rhizo- and endosphere of plants: their role, colonization, mechanisms involved and prospects for utilization. Soil Biol Biochem 42:669–678
Corredor DY, Salazar JM, Hohn KL, Bean S, Bean B, Wang D (2009) Evaluation and characterization of forage sorghum as feedstock for fermentable sugar production. Appl Biochem Biotech 158:164–179
Dowling DN, Doty SL (2009) Improving phytoremediation through biotechnology. Curr Opin Biotechnol 20:204–206
Dworkin M, Foster JW (1958) Experiments with some microorganisms which utilize ethane and hydrogen. J Bacteriol 75:592–603
Epelde L, Mijangos I, Becerril JM, Garbisu C (2009) Soil microbial community as bioindicator of the recovery of soil functioning derived from metal phytoextraction with sorghum. Soil Biol Biochem 41:1788–1794
Evers A, Hancock RD, Martell AE, Motekaitis RJ (1989) Metal ion recognition in ligands with negatively charged oxygen donor groups. Complexation of iron(III), gallium(III), indium(III), aluminum(III), and other highly charged metal ions. Inorg Chem 28:2189–2195
Gerber GB, Léonard A, Hantson P (2002) Carcinogenicity, mutagenicity and teratogenicity of manganese compounds. Crit Rev Oncol Hematol 42:25–34
Glick BR (2010) Using soil bacteria to facilitate phytoremediation. Biotechnol Adv 28:367–374
Gordon SA, Weber RP (1951) Colorimetric estimation of indoleacetic acid. Plant Physiol 26:192–195
Hardoim PR, van Overbeek LS, van Elsas JD (2008) Properties of bacterial endophytes and their proposed role in plant growth. Trends Microbiol 16:463–471
Hoagland DR, Arnon DI (1950) The water culture method for growing plants without soil. Calif Agr Exp Sta Circ 347:1–32
Idris R, Trifonova R, Puschenreiter M, Wenzel WW, Sessitsch A (2004) Bacterial communities associated with flowering plants of the Ni hyperaccumulator Thaspi goesingense. Appl Environ Microbiol 70:2667–2677
Jasberg BK, Montgomery RR, Anderson RA (1983) Preservation of sweet sorghum biomass. Biotechnol Bioeng Symp 13:113–120
Jha P, Kumar A (2009) Characterization of novel plant growth promoting endophytic bacterium Achromobacter xylosoxidans from wheat plant. Microb Ecol 58:179–188
Jiang CY, Sheng XF, Qian M, Wang QY (2008) Isolation and characterization of a heavy metal-resistant Burkholderia sp. from heavy metal-contaminated paddy field soil and its potential in promoting plant growth and heavy metal accumulation in metal-polluted soil. Chemosphere 72:157–164
Krämer U (2005) Phytoremediation: novel approaches to cleaning up polluted soils. Curr Opin Biotech 16:133–141
Li BZ, Balan V, Yuan YJ, Dale BE (2010) Process optimization to convert forage and sweet sorghum bagasse to ethanol based on ammonia fiber expansion (AFEX) pretreatment. Bioresource Technol 101:1285–1292
Linton JA, Miller JC, Little RD, Petrolia DR, Coble KH (2011) Economic feasibility of producing sweet sorghum as an ethanol feedstock in the southeastern United States. Biomass Bioenerg 35:3050–3057
Ma Y, Prasad MNV, Rajkumar M, Freitas H (2011) Plant growth promoting rhizobacteria and endophytes accelerate phytoremediation of metalliferous soils. Biotechnol Adv 29:248–258
Mamma D, Christakopoulos P, Koullas D, Kekos D, Macris BJ, Koukios E (1995) An alternative approach to the bioconversion of sweet sorghum carbohydrates to ethanol. Biomass Bioenerg 8:99–103
Mastretta C, Taghavi S, van der Lelie D, Mengoni A, Galardi F, Gonnelli C, Barac T, Boulet J, Weyens N, Vangronsveld J (2009) Endophytic bacteria from seeds of Nicotiana tabacum can reduce cadmium phytotoxicity. Int J Phytoremediat 11:251–267
Penrose DM, Glick BR (2003) Methods for isolating and characterizing ACC deaminase-containing plant growth-promoting rhizobacteria. Physiol Plantarum 118:10–15
Rajkumar M, Ae N, Freitas H (2009) Endophytic bacteria and their potential to enhance heavy metal phytoextraction. Chemosphere 77:153–160
Rajkumar M, Nagendran R, Lee KJ, Lee WH, Kim SZ (2006) Influence of plant growth promoting bacteria and Cr6+ on the growth of Indian mustard. Chemosphere 62:741–748
Rubin EM (2008) Genomics of cellulosic biofuels. Nature 454:841–845
Schatz A, Bovell C Jr (1952) Growth and hydrogenase activity of a new bacterium, Hydrogenomonas facilis. J Bacteriol 63:87–98
Schwyn B, Neilands JB (1987) Universal chemical assay for the detection and determination of siderophores. Anal Biochem 160:47–56
Sheng XF, Xia JJ (2006) Improvement of rape (Brassica napus) plant growth and cadmium uptake by cadmium-resistant bacteria. Chemosphere 64:1036–1042
Sheng XF, He LY, Wang QY, Ye HS, Jiang CY (2008a) Effects of inoculation of biosurfactant-producing Bacillus sp. J119 on plant growth and cadmium uptake in a cadmium-amended soil. J Hazard Mater 155:17–22
Sheng XF, Xia JJ, Jiang CY, He LY, Qian M (2008b) Characterization of heavy metal-resistant endophytic bacteria from rape (Brassica napus) roots and their potential in promoting the growth and lead accumulation of rape. Environ Pollut 156:1164–1170
Shi YW, Lou K, Li C (2010) Growth and photosynthetic efficiency promotion of sugar beet (Beta vulgaris L.) by endophytic bacteria. Photosynth Res 105:5–13
Strobel GA (2003) Endophytes as sources of bioactive products. Microbes Infect 5:535–544
Taghavi S, Garafola C, Monchy S, Newman L, Hoffman A, Weyens N, Barac T, Vangronsveld J, van der Lelie D (2009) Genome survey and characterization of endophytic bacteria exhibiting a beneficial effect on growth and development of poplar trees. Appl Environ Microbiol 75:748–757
Tripathi M, Munot HP, Shouche Y, Meyer JM, Goel R (2005) Isolation and functional characterization of siderophore-producing lead- and cadmium-resistant Pseudomonas putida KNP9. Curr Microbiol 50:233–237
Vartanian JP, Sala M, Henry M, Wain-Hobson S, Meyerhans A (1999) Manganese cations increase the mutation rate of human immunodeficiency virus type 1 ex vivo. J Gen Virol 80:1983–1986
Weyens N, van der Lelie D, Taghavi S, Newman L, Vangronsveld J (2009a) Exploiting plant–microbe partnerships to improve biomass production and remediation. Trends Microbiol 27:591–598
Weyens N, van der Lelie D, Taghavi S, Vangronsveld J (2009b) Phytoremediation: plant–endophyte partnerships take the challenge. Curr Opin Biotech 20:248–254
Wu CH, Wood TK, Mulchandani A, Chen W (2006) Engineering plant–microbe symbiosis for rhizoremediation of heavy metals. Appl Environ Microbiol 72:1129–1134
Xue SG, Chen YX, Reeves RD, Baker AJM, Lin Q, Fernando DR (2004) Manganese uptake and accumulation by the hyperaccumulator plant Phytolacca acinosa Roxb. (Phytolaccaceae). Environ Pollut 131:393–399
Zhuang P, Shu WS, Li Z, Liao B, Li JL, Shao JS (2009) Removal of metals by sorghum plants from contaminated land. J Environ Sci 21:1432–1437
Acknowledgments
This work was financially supported by a grant from National Science Fund for Distinguished Young Scholars (no. 50725825), the Key Program of National Natural Science Foundation of China (no. 50830301), and Hunan Province Natural Science Foundation of China (no.10JJ5051).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Luo, S., Xu, T., Chen, L. et al. Endophyte-assisted promotion of biomass production and metal-uptake of energy crop sweet sorghum by plant-growth-promoting endophyte Bacillus sp. SLS18. Appl Microbiol Biotechnol 93, 1745–1753 (2012). https://doi.org/10.1007/s00253-011-3483-0
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00253-011-3483-0