
Abstract
Salinity along with drought is one of the leading abiotic stresses in agriculture which foster growth retardation, physiological abnormalities, and lower production output of field crops throughout the world. Salt stress remains a major growth-limitation factor mostly in arid and semiarid zones. The problem emerges when the concentration of Na+, Cl−, and ions of other allied salts exceeds in soil than standard levels, which primarily disturbs the osmotic functions and then consequently alters several metabolic activities required for normal growth and development of plants. Besides natural occurrences in some soils, excessive irrigation, climate change, and agricultural intensification are some of the contributing factors towards soil salinity. Although different plants differ in sensitivity to soil salinity, most of their growth and yield traits are negatively influenced as a result of disorders in respiration, photosynthesis, mineral uptake, and oxidative stress. Decreased crop productivity due to salinity—as expected to elevate in the coming decades—would pose severe threats to global food security in the future if the challenge is not properly directed. Sustainable agronomic practices, deployment of molecular and functional genomic approaches can boost our understanding of salinity stress and create salt-tolerant traits in major field crops which could potentially contribute to production and yield enhancement under elevated saline conditions.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
Introduction
Global food security is greatly reliant on agricultural crops and their products which need substantial increases for maintenance of the gap between production and consumption. The significance of enhancing crop productivity is even more emergent in recent times because of the growing population of humankind which currently stands at 7.6 billion and is projected to exceed 9.7 billion by the year 2050 (FAOSTAT 2018). Undoubtedly, the increase in population will exert pressure for production of more crops and food resources which seems a challenging task for plant biologists. Concurrently, climate change and several biotic and abiotic stresses challenge the growth and production of agricultural crops. Among abiotic stresses, salinity is considered as one of the leading limiting factors responsible for growth and production decline of agricultural crops throughout the world principally in arid and semiarid regions (Kaashyap et al. 2018). About 20–33% of the cultivated and irrigated land throughout the world is affected by salinity, and the adverse effects are expected to reach to 50% in the year 2050 (Soda et al. 2016; Machado and Serralheiro 2017).
The problem of salinity emerges when ion concentration of different salts (predominantly NaCl) elevates in soils beyond threshold level required for normal germination, growth, and physiological activities of plants in root zones. Standard agricultural soils which promote the growth of cultivated crops have salinity level ~4 dS m−1 which is equivalent to 40 mM NaCl generally determined as the electrical conductivity of the saturated extract (Shrivastava and Kumar 2015). Ionic concentration exceeding 4 dS m−1 in rhizosphere result in stress conditions which challenge the growth of crops in several ways. Leading causes of salinity may be the natural addition of salts from rocks and minerals through weathering process (primary salinity) or extensive human activities like irrigation, agricultural intensification, and deforestation (secondary salinity) (Athar and Ashraf 2009). In addition, high evaporation rate in tropics also seems to increase the level of salts in agricultural land. Salinity has drastic consequences on water availability in the soil for plants, the rate of transpiration and photosynthesis, stomatal opening and closures, and functional activities of plant roots (Khataar et al. 2018). The discrepancies triggered by salinity stress result in impaired growth, physiological functions, and low yield outputs of crops. Annual expenditures associated with crop losses as a result of salinity are documented as 27 Billion US dollars (Singh et al. 2016). Coupled with drought, salinity triggered crop losses range between 20 and 50% in documented studies (Shrivastava and Kumar 2015) while losses may even be much higher as a result of salinity stress in areas out of scientific investigations. From physiological aspects, almost all crops show sensitivity to the salinity stress albeit variation in responses to the imposed stress in different crops does exist based on their tolerance level. Major concerns about the adverse effects of growing salinity stress are associated with crops which have a substantial contribution to global food supply. Rice, maize, wheat, barley, sorghum, and potato are considered as key drivers in the fulfillment of food needs throughout the world, while their growth and productivity are considerably affected by salt stress in many parts.
To maintain a sustainable production of agricultural crops and to ensure future food sanctuary, efforts have been made over decades to address the adversities caused by salinity stress on crops. One of the basic approaches to prevent crop losses triggered by salinization is to induce salinity tolerance in salt-sensitive crops. Genomic and molecular approaches to understand the possibilities of salt resistance induction in crops have proven effective in further elaboration of making crops adaptable to saline conditions (Luo et al. 2017). In addition to molecular methods, breeding for salt tolerance, proper agronomic practices in irrigation, and search for cost-effective and feasible methods to reduce drastic effects of salinity on principal crops are crucially needed to be employed. This chapter presents an overview of the salinity imposed effects on crops, and employment of agronomic approaches, seed priming and plant growth promoting bacteria (PGPB) to induce tolerance in crops to salinity.
Causes of Salinity and General Effects
Root sphere is the zone of soil which provides water and minerals to plants in addition to harboring diverse beneficial microbial communities and plays a determining role in successful germination and the consequent establishment of growth and reproductive phases of plants. Imbalance in water contents and minerals in the rhizospheric soil can lead to deteriorated effects on plant growth and productive outputs. Soil salinity alters water and mineral contents and microbiota and results in poor germinability and growth of sensitive crops. Salinity is a physiological state of soil where the concentration of active ions of salts principally NaCl increases the threshold levels. It is established that salt concentration up to 4 dS m−1 (~40 mM) of NaCl in soils represent a suitable environment for most of the plant species; however, some plants known as halophytes can withstand salinity stress up to 100 mM (examples include Vesicaria, Atriplex, Chenopodium spp.) while most of the cultivated crops including cereals (known as glycophytes) show differential sensitivity to salinity stress and prefer to grow at ≤40 mM salt concentration (Rasool et al. 2013; Ismail and Horie 2017; Yang et al. 2018). Causes of salinity stress in the natural ecosystem and managed agriculture are both natural and anthropogenic. Naturally, the problem of salinity occurs when minerals and salty rocks are weathered and ion concentration accumulates in soil (Athar and Ashraf 2009). Transportation of salt contents to root growing zones from parent rocks (rich in salts) as a result of several types of weathering and from groundwater may result in an increased level of salt stress and could contribute to primary salinity (Daliakopoulos et al. 2016). Agricultural lands located near coastal regions are exposed to high salinity stress because of the presence of high concentrations of salts in seawater. In tropical regions, high temperature can stimulate evaporation rate and hence more salt ions in soils accumulate giving rise to salinity stress. Moreover, flooding and wind erosion may be regarded to some extent as natural causes of salinity due to the imbalanced flow of minerals and salts from one region to another. Shallow groundwater may also serve as a source of salinization when upward migration of salts occurs (Shrivastava and Kumar 2015). Land cover and climatic conditions are also thought to contribute to salinity (Fan et al. 2012). Anthropogenic salinity, representing a significant proportion of the overall global salinity, is caused by massive agricultural activities, poor irrigation, use of imbalanced fertilizers, soil degradation, and poor drainages (Shrivastava and Kumar 2015; Sandhu et al. 2017). Mining activities and the use of wastewater and industrial effluents enriched in diverse salt contents in addition to other hazardous materials may lead to agricultural salinity (Daliakopoulos et al. 2016). Deforestation is an imminent threat to changes in rainfall patterns and soil erosion which would lead to consequent salinity problems. Rhizospheric soils whether salinized by natural processes or human activities are generally classified on the basis of electrical conductivity (EC) as: (1) non-salinized (EC = 0–2 dS m−1), (2) marginally salinized (EC = 2–4 dS m−1), (3) moderately salinized (EC = 4–8 dS m−1), (4) strongly salinized (EC = 8–16 dS m−1), (5) rigorously salinized (EC =16–32 dS m−1), and (6) exceptionally salinized (EC >32 dS m−1) (Rasool et al. 2013). According to Munns and Gilliham (2015), the economic costs associated with salinity are variable in different regions which may range between 300 and 600 US dollars ton−1. Daliakopoulos et al. (2016) stated that soil salinity may impart an economic burden of up to 600 million euros in European countries due to reduced crop yields and agricultural degradation. According to FAO and ITPS, African continent is the most salt-affected region where salinized area exceeds 122 million hectares followed by north and central Asia which represent almost 91.5 Mha of salt-affected land (Table 1).
Effects of Salinity on Crops
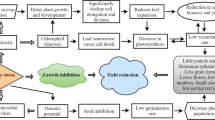
In general, plants are broadly categorized as glycophytes (sensitive to salinity) and halophytes (exhibit some degree of tolerance to salinity stress) on the basis of their tolerance in response to salinity stress. Halophytes are plants with typical examples Vesicaria, Atriplex, Chenopodium spp., and several others that can withstand high salt concentration while our major cultivated crops are glycophytes which show differential sensitivity to salinized soils (Borsani et al. 2003; Rasool et al. 2013; Yang et al. 2018). Differential responses of cultivated glycophytes depend on their growth phases, the soil characters, level of salinity, and agricultural practices (Daliakopoulos et al. 2016). Salinity stress primarily affects crops by causing water deficit condition in soils termed as “osmotic stress ” while secondarily due to the accumulation of toxic ions which consequently lead to a poor or delayed germination and post-germination growth abnormalities (Läuchli and Grattan 2007; Munns and Tester 2008). The primary osmotic stress is considered as hastened in its effects to which plants show abrupt responses while the secondary effects of salinity exposures are relatively slower and the adverse effects appear at later stages when sufficiently larger amounts of sodium ions gather in plant tissues and correspond to photosynthetic abnormalities, damage to cells, and many metabolic malfunctions (Munns and Tester 2008; Parihar et al. 2015; Hanin et al. 2016). Tuteja (2007) asserted that when Na ion accumulation in plant tissues increases 100 mM, enzymes’ functions, cell membrane structure, and cell division are severely influenced which result in a reduced growth. In leaves where transpiration proceeds, high level of ions may result in cell injuries and consequent retardation in growth (Parihar et al. 2015). Other drastic effects of salinity exposures on plants are misappropriated opening and closing of stomata, which lead to altered gaseous exchange, photosynthesis, and the rate of transpiration (Munns and Tester 2008; Hanin et al. 2016). Furthermore, it is strongly evident that higher concentration of salts’ ions in soil negatively affects the uptake of other necessary ions which plants require for several metabolic and enzymatic activities (Hanin et al. 2016). The high buildup of toxic ions in leaves outside threshold levels causes eventual death of many cells which are actively involved in photosynthesis with a net result in limited photosynthetic activity (Läuchli and Grattan 2007). The altered ratio of potassium and sodium ions significantly influence the entry of former into plant tissues and intercellular vicinities which lead to drastic consequences on ionic balance inside plants (Tuteja 2007). These abnormalities in physiological and biochemical processes of exposed plants are generally associated with salt stress symptoms which may range from reduced and/or delayed germination, plant growth, reproductive parameters, and yield outputs. In sum, osmotic stress developed due to higher salts concentration, accumulation of Na+ in plants, ion toxicity, altered ratio of Na+ and K+ results collectively in deficit water and nutrient uptake by germinating seeds and seedling roots, cell injuries, changes in vital enzymes and hormones, irregular stomatal opening and closing with disturbed gaseous exchange, rate of transpiration and photosynthesis; thus imposing stress environment for seed to properly germinate and for seedlings to properly grow. The overall result would be a reduction in growth and yields plants exposed to salinity.
In several studies, drastic consequences of salinity on germination, growth, yield, and physiological activities of major cultivated crops have been proven (Table 2). Cha-Um and Kirdmanee (2009) applied 100–400 mM NaCl stress to different genotypes of maize which responded to the imposed stress with reduced synthesis of chlorophylls while increased proline contents. Even lower salinity stress ranging between 3.5 and 8.5 dS m−1 caused a significant retardation in oil content and yield of safflower (Yeilaghi et al. 2012). In our previous study, we recorded a significant decline in germination and seedling length of wheat cultivars which were exposed to 8 dS m−1 NaCl stress (Muhammad et al. 2015). In okra, reduced germination and growth at 75 mM (Habib et al. 2016), while in chickpea arrested germination, growth, and biomass at 40 mM NaCl was observed (Atieno et al. 2017). In response to different level of salinity stress, reduced growth and biomass, leaf area, chlorophyll degradation, altered water status, defective stomatal functions, altered rate of transpiration and respiration, and imbalanced ion ratios in maize (Gul et al. 2017; Konuşkan et al. 2017), rice (Krishnamurthy et al. 2016); Islam et al. 2017; Shahzad et al. 2017; Radanielson et al. 2018), wheat (Gul et al. 2017; Fathi et al. 2017; Bajwa et al. 2018; Khataar et al. 2018), tomato (Rubio et al. 2017; Martinez et al. 2018), faba bean and common bean (Benidire et al. 2017; Hussein et al. 2017; Ahmad et al. 2018; Khataar et al. 2018), barley (Allel et al. 2018), tomato, cotton, and several other crops have been well documented (Meloni et al. 2003; Zhang et al. 2006; Sarabi et al. 2017; Sandhu et al. 2017; Martinez et al. 2018; Ahmadi et al. 2018).
Salinity Management Approaches
Modifications in Agricultural Practices
Rengasamy (2006) categorized three major types of salinity: (1) salinity associated with groundwater, (2) salinity associated with non-groundwater, and (3) salinity associated with irrigation. All the three types of salinity are manageable if preventive measures, depending on the types of salinity and location, are taken into account. Although it seems difficult to employ a particular method in agro farming systems for salinity management, a combination of several techniques would sufficiently help in reducing the drastic impacts of salinity on crops. As for natural factors responsible for salinity, man has little influence on bringing them under control. Since leading causes of anthropogenic salinity include land degradation, poor drainage, and use of substandard water and excessive irrigation, employment of appropriate practices will satisfactorily manage the salt menace. Foremost, increasing population pressure in urban areas stimulate migrations to rural areas—the main hub of agricultural activities—which consequently results in the utilization of agricultural land for dwelling purposes. In parts of fertile land destruction as a result of building constructions, the household outflow of water may cause disturbances in the quality of irrigated water. Thus, restoration and protection of fertile land would limit the potential secondary salinization. Rise in water table caused by irrigation practices, rainfall, and leaching of water which are perceived to be linked with the salinization process can be maintained by surface drainage. The exclusion of excess of water from rhizosphere is also an effective tool to maintain the permanent water table and avoid its rising (Tiwari and Goel 2017). Drainage can be achieved by making trenches in agricultural soils which are saturated with plenty of water or by installation of a drainage system although the latter one cannot be afforded by resource-poor farmers in many countries. Hanson and May (2004) stated that substrata drip irrigation could maintain water table in shallow water regions and correspond to better salinity management and improvement in crops in such adopted system. Ayars et al. (2006) proposed that subsurface drainage and management of water at specific depth in agricultural lands could significantly improve soil profile and salinization problems. They further suggested that proper design and post-installation maintenance of drainage system would be required for getting better results in arid soils with the saline profile. In a study conducted in Pakistan on the importance of different drainage projects in irrigated areas for salt, management revealed a 14–20% decrease in soil salinity (Azhar 2010). Valipour (2014), in a comprehensive review, presented the advantages of the drainage systems in reducing salinity problems in many countries where the problem prevails. He outlined that in Europe the drainage system was the most effective measure in controlling salinity stress; however, such systems were not recommended for areas where phosphorus deficiency remained a challenging problem. Hou et al. (2016) advocated the use of mulch-drip irrigation in soils enriched with salts to minimize salinity concentration and to promote plants’ growth. Besides artificial drainage systems, cultivation of deep-rooted trees such as Eucalyptus sp. may prevent rise in the water table and consequent salinization. Canal system in different countries, particularly in Pakistan and other developing countries, is extensively used for irrigation purposes. Inappropriate designs of canal systems and lack of protective lining in most of the developing countries due to high costs result in water seepage. The seeped water move through different soil zones and lead to the solubility of minerals and their accumulation in the fertile agricultural land which consequently result in soil salinity and water logging.
Another leading cause of salinization throughout the world is excessive irrigation and the use of substandard water for the stated purpose. In areas with arid and semiarid conditions, irrigation processes facilitate the mobility of salts to non-saline areas where secondary salinization occurs and disturb the growth parameters of cultivated plants (Smedema and Shiati (2002). Rietz and Haynes (2003) debated that irrigation with poor water coupled with unmanaged drainage results in water level rise and accumulation of salts in the soil surface layer where plants are grown. The scarcity of fresh water for irrigation and other purposes in many parts of the world coerce farmers to use low standard water which mainly contains municipal wastes and salts, thereby reducing soil quality and elevating salinity and sodicity levels (Qadir and Oster 2004). It is the prime priorities of agricultural stakeholders to stimulate scientific practices in irrigation. This may include drip irrigation (Hou et al. 2016; Ortega-Reig et al. 2017), installation of tube wells (Memon et al. 2017), sprinkler irrigation (Rudrapur et al. 2017), and proper drainage by making drenches and installing water removal equipment. Although it seems costly particularly in developing countries, comparisons with crops losses and land degradation as a result of salinity and poor irrigation can highlight the potential benefits of sustainable irrigation methods for the long term. To minimize energy consumption costs incurred by tube wells irrigation, deployment of solar panels can be specifically useful. Cultivation of deep-rooted trees such as Eucalyptus, Acacia, and Sesbania species can efficiently manage water table and consequently avoid salinity problems. Deforestation and overgrazing are also some of the important indirect contributing factors towards salinization because of changes in climatic condition, altered rainfall, elevated evaporation, and flooding; thus, maximum vegetation and protection of forests can be helpful in long-term salinity management.
Seed Priming and Plant Growth Promoting Bacteria (Bio-Priming) for Salinity Management
Pre-germination soaking of seeds with different solutions (or pure water) for the different duration is termed as “seed priming” (Song et al. 2017). Solutions of different compounds, both of natural and synthetic origins, are usually used as pre-soaking materials to prepare seeds for better performance under stressful environment. Seeds may be primed either with water, or salts, hormones, and natural metabolic substances although different priming agents have variable effects on treated seeds in terms of their responses to exposed stress. Soaking duration with priming agents is also a necessary factor which enables seeds to architect stress-combating machinery. In practice, halo priming (treatment of seeds with different concentration of salts) and hydropriming (water treatment) seem economical because of low cost and easy availability of soaking gents (Jisha et al. 2013) although other priming compounds such as polyethylene glycol (PEG), auxins, gibberellins, cytokinins, salicylic acid, jasmonic acid, kinetin, urea, and several natural metabolic substances have been well proven in inducing defense and stress tolerance to subjected seeds (Islam et al. 2015; Salah et al. 2015; Savvides et al. 2016). Primary mechanism underlying the priming-induced stress tolerance in crops is the activation of enzymes, hormones, homeostatic regulation, synthesis of new messenger RNA, improvement in imbibition potential and dormancy, detoxification of reactive oxygen species, enhanced antioxidant systems, and induction of “pre-stress memory ” (Bruce et al. 2007; Varier et al. 2010; Hussain et al. 2016) (Fig. 1). Primed seeds perform better than non-primed ones when sown either under normal conditions or in a stressed environment. Many studies suggest that priming cause better germination, seedling emergence, and vigor (Salah et al. 2015; Hussain et al. 2016). Ajouri et al. (2004) have documented that priming-induced salinity resistance in barley and better germination was achieved. Kaya et al. (2006) observed improved germination and growth of sunflower seedling in saline and drought conditions as a result of hydropriming. Under 100 mM NaCl stress, halopriming, and hydropriming invigorated wheat growth, yield, antioxidant activities, and ions regulation (Islam et al. 2015). Improved germination and reduced mean germination time of Nigella sativa were also recorded under 40 mM NaCl when seeds were treated with 1–2% NaCl and several other priming agents (Gholami et al. 2015). Khaliq et al. (2015) treated seeds of rice with 15–105 μmol L−1 concentrations of selenium and evaluated the primed seeds for germination, growth, and enzymatic activity under salinity stress. They observed the better performance of primed seed than control under stress conditions for all the studied parameters. Osmopriming of bread wheat seeds with CaCl2 resulted in enhanced tolerance to salinity (100 mM) resulting in improved leaf area, Na/K ion ratio, and grain yield (Tabassum et al. 2017). Pretreatment of broccoli sprout with methyl jasmonate and KCl induced salt-stress tolerance in tested plants which resulted in growth and biochemical parameters elevation under salinity stress (Hassini et al. 2017). Recently, Bajwa et al. (2018) effectively induced salt tolerance in wheat by pre-soaking seeds with sorghum extracts and benzyl amino-purine. Germination, growth, and biochemical attributes exhibited improved performance in salinity imposed stress.

In addition to wide applicability of seed priming in agriculture, plant growth promoting bacteria (PGPB) have also a potential role in improving crops’ stress responses. PGPB can manage drastic effects of salinity and other stress conditions on cultivated crops by several mechanisms. They may stimulate the production of specific proteins and osmoprotectants in crops exposed to stressful environment thereby reducing the adverse effects of stresses and protecting crops from stress injuries (Grover et al. 2011). Saleem et al. (2007) argued that one of the leading factors associated with stress is the elevated production of ethylene—a growth regulator produced indigenously by plants when they are challenged with stress conditions—is regulated by several strains of PGPB. It is established that many PGPB produce ACC deaminase and regulate the production and functions of cytokinins, antioxidants, and ethylene which not only help crops to take nutrient and water properly but also induce in them systemic resistance to salinity stress which imparts stimulatory effects on the growth and yield under such stresses (Yang et al. 2009). According to Glick (2012), plant growth suppression in fields is either caused by biotic (pathogenic interaction) or abiotic (salinity, intensive light, drought, temperature, etc.) stresses which are generally correlated with abnormal production of stress hormones, fluctuation in metabolic machinery. He provided evidence that many PGPB can manage the required production of ethylene, indole acetic acid (IAA), trehalose, cytokinin, and several other stress components. Shrivastava and Kumar (2015), in a comprehensive review, highlighted the role of PGPB in conferring salinity resistance to crops. They commented that PGPB induces systemic tolerance in crops besides their active role in the provision of nutrients, soil fertility, and disease suppression. Bharti et al. (2016) documented that Dietzia natronolimnaea, a plant growth promoting bacterium, was involved in regulating transcriptional factors which confer salt resistance to plants and protect them from salinity-induced injuries. Del Cerro et al. (2017) suggested that certain strains of Rhizobium tropici are involved in the production of nodulating factors under high salt concentration which promotes root nodulation in legume crops and helps in avoiding salinity stress. Some species of Streptomyces are recognized for their ability to produce 1-aminocyclopropane-1-carboxylate deaminase and promote the growth, ion uptake, and chlorophyll functions of rice under 150 mM NaCl stress (Jaemsaeng et al. 2018).
In the previous study, salinity tolerance and improved growth of tomato were recorded in response to PGPB (Achromobacter piechaudii) under 43 mM NaCl stress (Mayak et al. 2004). Tank and Saraf (2010) reported 50% increase in tomato growth and biomass under salt stress when plants were inoculated with different strains of Pseudomonas sp. under greenhouse conditions. Wheat seeds inoculated with Arthrobacter sp. exhibited efficient resistance to salt stress and yielded better than non-inoculated plants (Upadhyay and Singh 2015). Wheat inoculated with PGPB Klebsiella sp. (SBP8) and grown in 150–200 mM NaCl stress showed higher growth and chlorophyll activity than non-inoculated control (Singh et al. 2015). Edible peas grown under 70–130 mM NaCl stress after pretreatment with Variovorax paradoxus showed improved growth, photosynthetic performance and ion uptake (Wang et al. 2016). Bharti et al. (2016) reported that wheat inoculation with Dietzia natronolimnaea resulted in a significant elevation of plant height and dry biomass when salinity stress (150 mM NaCl) was imposed on test crop. Under similar salinity stress, Streptomyces sp. (strain GMKU 336) promoted plant growth, water status, chlorophyll and proline contents, and ion ratios of rice (Jaemsaeng et al. 2018). In other similar studies under high salt concentrations, Burkholderia and Enterobacter sp. stimulated the growth and physicochemical attributes of maize and wheat (Akhtar et al. 2015a, b), Bacillus cereus improved mung bean (Islam et al. 2016), and Enterobacter sp. promoted rice growth (Sarkar et al. 2018). These and dozens of other studies employing PGPB in salt stress environment indicate that several PGPB strains have a prospective role in inducing salt-stress tolerance to cultivated crops and enhancing their yield outputs.
Conclusions
Cultivated crops are challenged with several constraints among which salinity is an important one responsible for limited growth and crop production. Salinity stress imposes ion toxicity, water deficit conditions, imbalanced ion uptake, cellular damage, and degradation of chlorophylls, transpiration and respiration activities which definitely influence overall growth attributes of challenged plants. Both natural and man-triggered activities cause salinization in agriculture. Extensive irrigation, poor drainage, low-quality water for irrigation purposes are directly contributing to the problem of salinity. Employment of mechanisms and techniques to diminish the adverse effects of salinity and make crops adapted to withstand salinity would help in crop losses management. Thus, scientific approaches in agriculture such as sprinkler irrigation drench and pumping surface and groundwater, and lining of irrigating canals can significantly lower salinity levels in cultivated land. Seed priming with water, salts, hormones, and other chemicals induces stress adaptability in them and can prove effective tools to divert the adverse effects of salinity on crops. Bio-priming with plant growth promoting bacteria has also been well recognized for their ability to induce salinity tolerance in crops. To make the maximum use of seed priming agents and PGPB for salt management in cultivated crops, commercial availability, economic accessibility, and safety issues concerned with priming chemicals and PGPB strains must be ensured by stakeholders. Since farmers represent a basic component agricultural system, their awareness about the significance of seed priming and PGPB in salinity can contribute to further expansion of these techniques in agricultural areas which are challenged with salinity.
References
Ahanger MA, Agarwal RM (2017) Salinity stress induced alterations in antioxidant metabolism and nitrogen assimilation in wheat (Triticum aestivum L) as influenced by potassium supplementation. Plant Physiol Biochem 115:449–460
Ahmad P, Alyemeni MN, Ahanger MA, Egamberdieva D, Wijaya L, Alam P (2018) Salicylic acid (SA) induced alterations in growth, biochemical attributes and antioxidant enzyme activity in Faba Bean (Vicia faba L.) seedlings under NaCl toxicity. Russian J Plant Physiol 65(1):104–114
Ahmadi FI, Karimi K, Struik PC (2018) Effect of exogenous application of methyl jasmonate on physiological and biochemical characteristics of Brassica napus L. cv. Talaye under salinity stress. S Afr J Bot 115:5–11
Ajouri A, Asgedom H, Becker M (2004) Seed priming enhances germination and seedling growth of barley under conditions of P and Zn deficiency. J Plant Nutr Soil Sci 167:630–636
Akhtar SS, Andersen MN, Liu F (2015a) Residual effects of biochar on improving growth, physiology and yield of wheat under salt stress. Agric Water Manag 158:61–68
Akhtar SS, Andersen MN, Naveed M, Zahir ZA, Liu F (2015b) Interactive effect of biochar and plant growth-promoting bacterial endophytes on ameliorating salinity stress in maize. Funct Plant Biol 42:770–781
Allel D, Ben-Amar A, Abdelly C (2018) Leaf photosynthesis, chlorophyll fluorescence and ion content of barley (Hordeum vulgare) in response to salinity. J Plant Nutr 41:497–508
Athar HR, Ashraf M (2009) Strategies for crop improvement against salinity and drought stress: an overview. In: Ashraf M, Ozturk M, Athar HR (eds) Salinity and water stress. Springer, New York, pp 1–16
Atieno J, Li Y, Langridge P, Dowling K, Brien C, Berger B, Sutton T (2017) Exploring genetic variation for salinity tolerance in chickpea using image-based phenotyping. Sci Rep 7(1):1300
Ayars E, Christe EW, Hornbuckle JW (2006) Controlled drainage for improved water management in arid regions irrigated agriculture. Agric Water Manag 86:128–139
Azhar AH (2010) Impact of subsurface drainage on soil salinity in Pakistan. J Anim Plant Sci 20:94–98
Bacha H, Tekaya M, Drine S, Guasmi F, Touil L, Enneb H, Ferchichi A (2017) Impact of salt stress on morpho-physiological and biochemical parameters of Solanum lycopersicum cv. Microtom leaves. S Afr J Bot 108:364–369
Bajwa AA, Farooq M, Nawaz A (2018) Seed priming with sorghum extracts and benzyl aminopurine improves the tolerance against salt stress in wheat (Triticum aestivum L.). Physiol Mol Biol Plants 24:239-249
Benidire L, Lahrouni M, El Khalloufi F, Göttfert M, Oufdou K (2017) Effects of rhizobium leguminosarum inoculation on growth, nitrogen uptake and mineral assimilation in Vicia faba plants under salinity stress. J Agric Sci Technol 19:889–901
Bharti N, Pandey SS, Barnawal D, Patel VK, Kalra A (2016) Plant growth promoting rhizobacteria Dietzia natronolimnaea modulates the expression of stress responsive genes providing protection of wheat from salinity stress. Sci Rep 6:34768
Borsani O, Valpuesta V, Botella MA (2003) Developing salt tolerant plants in a new century: a molecular biology approach. Plant Cell Tissue Organ Cult 73:101–115
Bruce TJ, Matthes MC, Napier JA, Pickett JA (2007) Stressful “memories” of plants: evidence and possible mechanisms. Plant Sci 173:603–608
Cha-Um S, Kirdmanee C (2009) Effect of salt stress on proline accumulation, photosynthetic ability and growth characters in two maize cultivars. Pak J Bot 41:87–98
Daliakopoulos IN, Tsanis IK, Koutroulis A, Kourgialas NN, Varouchakis AE, Karatzas GP, Ritsema CJ (2016) The threat of soil salinity: a European scale review. Sci Total Environ 573:727–739
Del Cerro P, Pérez-Montaño F, Gil-Serrano A, López-Baena FJ, Megías M, Hungria M, Ollero FJ (2017) The Rhizobium tropici CIAT 899 NodD2 protein regulates the production of Nod factors under salt stress in a flavonoid-independent manner. Sci Rep 7:46712
Fan X, Pedroli B, Liu G, Liu Q, Liu H, Shu L (2012) Soil salinity development in the yellow river delta in relation to groundwater dynamics. Land Degrad Dev 23:175–189
FAOSTAT (2018) Food and Agricultural Organization Statistical database, Crop production. http://faostat3.fao.org/download /Q/QC/ E (accessed on March 20, 2018)
Fathi A, Zahedi M, Torabian S, Khoshgoftar A (2017) Response of wheat genotypes to foliar spray of ZnO and Fe2O3 nanoparticles under salt stress. J Plant Nutr 40:1376–1385
Gholami M, Mokhtarian F, Baninasab B (2015) Seed halopriming improves the germination performance of black seed (Nigella sativa) under salinity stress conditions. J Crop Sci Biotechnol 18:21–26
Glick BR (2012) Plant growth-promoting bacteria: mechanisms and applications. Scientifica 2012:1
Grover M, Ali SZ, Sandhya V, Rasul A, Venkateswarlu B (2011) Role of microorganisms in adaptation of agriculture crops to abiotic stresses. World J Microbiol Biotechnol 27:1231–1240
Gul H, Kinza S, Shinwari ZK, Hamayun M (2017) Effect of selenium on the biochemistry of Zea mays under salt stress. Pak J Bot 49:25–32
Habib SH, Kausar H, Saud HM (2016) Plant growth-promoting rhizobacteria enhance salinity stress tolerance in okra through ROS-scavenging enzymes. Biomed Res Int 2016:6284547
Hakim MA, Juraimi AS, Begum M, Hanafi MM, Ismail MR, Selamat A (2010) Effect of salt stress on germination and early seedling growth of rice (Oryza sativa L.). Afr J Biotechnol 9:1911–1918
Hanin M, Ebel C, Ngom M, Laplaze L, Masmoudi K (2016) New insights on plant salt tolerance mechanisms and their potential use for breeding. Front Plant Sci 7:1787
Hanson B, May D (2004) Effect of subsurface drip irrigation on processing tomato yield, water table depth, soil salinity, and profitability. Agric Water Manag 68:1–17
Hassini I, Martinez-Ballesta MC, Boughanmi N, Moreno DA, Carvajal M (2017) Improvement of broccoli sprouts (Brassica oleracea L. var. italica) growth and quality by KCl seed priming and methyl jasmonate under salinity stress. Sci Hortic 226:141–151
Hou M, Zhu L, Jin Q (2016) Surface drainage and mulching drip-irrigated tomatoes reduces soil salinity and improves fruit yield. PLoS One 11:e0154799
Hussain S, Khan F, Cao W, Wu L, Geng M (2016) Seed priming alters the production and detoxification of reactive oxygen intermediates in rice seedlings grown under sub-optimal temperature and nutrient supply. Front Plant Sci 7:439
Hussein M, Embiale A, Husen A, Aref IM, Iqbal M (2017) Salinity-induced modulation of plant growth and photosynthetic parameters in faba bean (Vicia faba) cultivars. Pak J Bot 49:867–877
Islam F, Yasmeen T, Ali S, Ali B, Farooq MA, Gill RA (2015) Priming-induced antioxidative responses in two wheat cultivars under saline stress. Acta Physiol Plant 37:153
Islam F, Yasmeen T, Arif MS, Ali S, Ali B, Hameed S, Zhou W (2016) Plant growth promoting bacteria confer salt tolerance in Vigna radiata by up-regulating antioxidant defense and biological soil fertility. Plant Growth Regul 80:23–36
Islam F, Farooq MA, Gill RA, Wang J, Yang C, Ali B, Zhou W (2017) 2, 4-D attenuates salinity-induced toxicity by mediating anatomical changes, antioxidant capacity and cation transporters in the roots of rice cultivars. Sci Rep 7:10443
Ismail AM, Horie T (2017) Genomics, physiology, and molecular breeding approaches for improving salt tolerance. Annu Rev Plant Biol 68:405–434
Jaemsaeng R, Jantasuriyarat C, Thamchaipenet A (2018) Molecular interaction of 1-aminocyclopropane-1-carboxylate deaminase (ACCD)-producing endophytic Streptomyces sp. GMKU 336 towards salt-stress resistance of Oryza sativa L. cv. KDML105. Sci Rep 8(1):1950
Jisha KC, Vijayakumari K, Puthur JT (2013) Seed priming for abiotic stress tolerance: an overview. Acta Physiol Plant 35:1381–1396
Kaashyap M, Ford R, Kudapa H, Jain M, Edwards D, Varshney R, Mantri N (2018) Differential regulation of genes involved in root morphogenesis and cell wall modification is associated with salinity tolerance in chickpea. Sci Rep 8:4855
Kaya MD, Okçu G, Atak M, Cıkılı Y, Kolsarıcı Ö (2006) Seed treatments to overcome salt and drought stress during germination in sunflower (Helianthus annuus L.). Eur J Agron 24:291–295
Kaya C, Ashraf M, Sonmez O, Polat T, Tuna AL (2017) The combined effects of nitric oxide and thiourea on plant growth and mineral nutrition of salt-stressed plants of two maize cultivars with differential salt tolerance. J Plant Nutr. https://doi.org/10.1080/01904167.2016.1270314
Khaliq A, Aslam F, Matloob A, Hussain S, Geng M, Wahid A, Rehman H (2015) Seed priming with selenium: consequences for emergence, seedling growth, and biochemical attributes of rice. Biol Trace Elem Res 166:236–244
Khataar M, Mohhamadi MH, Shabani F (2018) Soil salinity and matric potential interaction on water use, water use efficiency and yield response factor of bean and wheat. Sci Rep 8:2679
Konuşkan Ö, Gözübenli H, Atiş İ, Atak M (2017) Effects of salinity stress on emergence and seedling growth parameters of some maize genotypes (Zea mays L.). Turk J Agric Food Sci Technol 5:1668–1672
Krishnamurthy SL, Gautam RK, Sharma PC, Sharma DK (2016) Effect of different salt stresses on agro-morphological traits and utilisation of salt stress indices for reproductive stage salt tolerance in rice. Field Crops Res 190:26–33
Läuchli A, Grattan SR (2007) Plant growth and development under salinity stress. In: Jenks MA, Hasegawa PM, Mohan Jain S (eds) Advances in molecular breeding toward drought and salt tolerant crops. Springer, New York, pp 1–32
Li R, Shi F, Fukuda K, Yang Y (2010) Effects of salt and alkali stresses on germination, growth, photosynthesis and ion accumulation in alfalfa (Medicago sativa L.). Soil Sci Plant Nutr 56:725–733
Luo Y, Reid R, Freese D, Li C, Watkins J, Shi H, Song BH (2017) Salt tolerance response revealed by RNA-Seq in a diploid halophytic wild relative of sweet potato. Sci Rep 7:9624
Machado RMA, Serralheiro RP (2017) Soil salinity: effect on vegetable crop growth. Management practices to prevent and mitigate soil salinization. Horticulturae 3:30
Mahmood A, Turgay OC, Farooq M, Hayat R (2016) Seed biopriming with plant growth promoting rhizobacteria: a review. FEMS Microbiol Ecol 92(8):pii:fiw112
Martinez V, Nieves-Cordones M, Lopez-Delacalle M, Rodenas R, Mestre TC, Garcia-Sanchez F, Rivero RM (2018) Tolerance to stress combination in tomato plants: new insights in the protective role of melatonin. Molecules 23:535
Mayak S, Tirosh T, Glick BR (2004) Plant growth-promoting bacteria confer resistance in tomato plants to salt stress. Plant Physiol Biochem 42:565–572
Meloni DA, Oliva MA, Martinez CA, Cambraia J (2003) Photosynthesis and activity of superoxide dismutase, peroxidase and glutathione reductase in cotton under salt stress. Environ Exp Bot 49:69–76
Memon JA, Jogezai G, Hussain A, Alizai MQ, Baloch MA (2017) Rehabilitating traditional irrigation systems: assessing popular support for Karez rehabilitation in Balochistan, Pakistan. Hum Ecol 45:265–275
Muhammad Z, Hussain F, Rehmanullah, Majeed A (2015) Effect of halopriming on the induction of NaCl salt tolerance in different wheat genotypes. Pak J Bot 47: 1613-1620
Munns R, Tester M (2008) Mechanisms of Salinity Tolerance. Ann Rev Plant Biol 59:651-681
Munns R, Gilliham M (2015) Salinity tolerance of crops—what is the cost? New Phytol 208:668–673
Ortega-Reig M, Sanchis-Ibor C, Palau-Salvador G, García-Mollá M, Avellá-Reus L (2017) Institutional and management implications of drip irrigation introduction in collective irrigation systems in Spain. Agric Water Manag 187:164–172
Parihar P, Singh S, Singh R, Singh VP, Prasad SM (2015) Effect of salinity stress on plants and its tolerance strategies: a review. Environ Sci Pollut Res 22:4056–4075
Perveen S, Shahbaz M, Ashraf M (2010) Regulation in gas exchange and quantum yield of photosystem II (PSII) in salt-stressed and non-stressed wheat plants raised from seed treated with triacontanol. Pak J Bot 42:3073–3081
Qadir M, Oster JD (2004) Crop and irrigation management strategies for saline-sodic soils and waters aimed at environmentally sustainable agriculture. Sci Total Environ 323:1–19
Radanielson AM, Angeles O, Li T, Ismail AM, Gaydon DS (2018) Describing the physiological responses of different rice genotypes to salt stress using sigmoid and piecewise linear functions. Field Crops Res 220:46–56
Rasool S, Hameed A, Azooz MM, Siddiqi TO, Ahmad P (2013) Salt stress: causes, types and responses of plants. Ecophysiology and responses of plants under salt stress. Springer, New York, pp 1–24
Rengasamy P (2006) World salinization with emphasis on Australia. J Exp Bot 57:1017–1023
Rietz DN, Haynes RJ (2003) Effects of irrigation-induced salinity and sodicity on soil microbial activity. Soil Biol Biochem 35:845–854
Rivero RM, Mestre TC, Mittler RON, Rubio F, Garcia-Sanchez F, Martinez V (2014) The combined effect of salinity and heat reveals a specific physiological, biochemical and molecular response in tomato plants. Plant Cell Environ 37:1059–1073
Rubio MB, Hermosa R, Vicente R, Gómez-Acosta FA, Morcuende R, Monte E, Bettiol W (2017) The combination of Trichoderma harzianum and chemical fertilization leads to the deregulation of phytohormone networking, preventing the adaptive responses of tomato plants to salt stress. Front Plant Sci 8:294
Rudrapur S, Mundinamani SM, Kiresur VR, Hosamani SB, Manjunatha MV, Dolli SS (2017) Impact of sprinkler irrigation system on doubling Farmers’ income and water use efficiency of tur in northern Karnataka. In: Agricultural Economics Research Review
Salah SM, Yajing G, Dongdong C, Jie L, Aamir N, Qijuan H, Jin H (2015) Seed priming with polyethylene glycol regulating the physiological and molecular mechanism in rice (Oryza sativa L.) under nano-ZnO stress. Sci Rep 5:14278
Saleem M, Arshad M, Hussain S, Bhatti AS (2007) Perspective of plant growth promoting rhizobacteria (PGPR) containing ACC deaminase in stress agriculture. J Ind Microbiol Biotechnol 34(10):635–648
Sandhu D, Cornacchione MV, Ferreira JF, Suarez DL (2017) Variable salinity responses of 12 alfalfa genotypes and comparative expression analyses of salt-response genes. Sci Rep 7:42958
Sarabi B, Bolandnazar S, Ghaderi N, Ghashghaie J (2017) Genotypic differences in physiological and biochemical responses to salinity stress in melon (Cucumis melo L.) plants: prospects for selection of salt tolerant landraces. Plant Physiol Biochem 119:294–311
Sarkar A, Ghosh PK, Pramanik K, Mitra S, Soren T, Pandey S, Maiti TK (2018) A halotolerant Enterobacter sp. displaying ACC deaminase activity promotes rice seedling growth under salt stress. Res Microbiol 169:20–32
Savvides A, Ali S, Tester M, Fotopoulos V (2016) Chemical Priming of Plants Against Multiple Abiotic Stresses: Mission Possible?. Trends Plant Sci 21:329–340
Shahzad R, Khan AL, Bilal S, Waqas M, Kang SM, Lee IJ (2017) Inoculation of abscisic acid-producing endophytic bacteria enhances salinity stress tolerance in Oryza sativa. Environ Exp Bot 136:68–77
Shrivastava P, Kumar R (2015) Soil salinity: a serious environmental issue and plant growth promoting bacteria as one of the tools for its alleviation. Saudi J Biol Sci 22:123–131
Singh RP, Jha P, Jha PN (2015) The plant-growth-promoting bacterium Klebsiella sp. SBP-8 confers induced systemic tolerance in wheat (Triticum aestivum) under salt stress. J Plant Physiol 184:57–67
Singh VK, Mishra A, Haque I, Jha B (2016) A novel transcription factor-like gene SbSDR1 acts as a molecular switch and confers salt and osmotic endurance to transgenic tobacco. Sci Rep 6:31686
Smedema LK, Shiati K (2002) Irrigation and salinity: a perspective review of the salinity hazards of irrigation development in the arid zone. Irrig Drain Syst 16:161–174
Soda N, Sharan A, Gupta BK, Singla-Pareek SL, Pareek A (2016) Evidence for nuclear interaction of a cytoskeleton protein (OsIFL) with metallothionein and its role in salinity stress tolerance. Sci Rep 6:34762
Song GC, Choi HK, Kim YS, Choi JS, Ryu CM (2017) Seed defense biopriming with bacterial cyclodipeptides triggers immunity in cucumber and pepper. Sci Rep 7:14209
Tabassum T, Farooq M, Ahmad R, Zohaib A, Wahid A (2017) Seed priming and transgenerational drought memory improves tolerance against salt stress in bread wheat. Plant Physiol Biochem 118:362–369
Tank N, Saraf M (2010) Salinity-resistant plant growth promoting rhizobacteria ameliorates sodium chloride stress on tomato plants. J Plant Interact 5:51–58
Tiwari P, Goel A (2017) An overview of impact of subsurface drainage project studies on salinity management in developing countries. Appl Water Sci 7:569–580
Tuteja N (2007) Mechanisms of high salinity tolerance in plants. In: Methods in Enzymology. Academic Press, pp 419–438
Upadhyay SK, Singh DP (2015) Effect of salt-tolerant plant growth-promoting rhizobacteria on wheat plants and soil health in a saline environment. Plant Biol 17:288–293
Valipour M (2014) Drainage, waterlogging, and salinity. Arch Agron Soil Sci 60:1625–1640
Varier A, Vari AK, Dadlani M (2010) The subcellular basis of seed priming. Curr Sci 99:450–456
Wang Q, Dodd IC, Belimov AA, Jiang F (2016) Rhizosphere bacteria containing 1-aminocyclopropane-1-carboxylate deaminase increase growth and photosynthesis of pea plants under salt stress by limiting Na+ accumulation. Func Plant Biol 43:161–172
Yang J, Kloepper JW, Ryu CM (2009) Rhizosphere bacteria help plants tolerate abiotic stress. Trend Plant Sci 14:1–4
Yang A, Akhtar SS, Iqbal S, Qi Z, Alandia G, Saddiq MS, Jacobsen SE (2018) Saponin seed priming improves salt tolerance in quinoa. J Agron Crop Sci 204:31–39
Yeilaghi H, Arzani A, Ghaderian M, Fotovat R, Feizi M, Pourdad SS (2012) Effect of salinity on seed oil content and fatty acid composition of safflower (Carthamus tinctorius L.) genotypes. Food Chem 130:618–625
Zhang ZJ, Li HZ, Zhou WJ, Takeuchi Y, Yoneyama K (2006) Effect of 5-aminolevulinic acid on development and salt tolerance of potato (Solanum tuberosum L.) microtubers in vitro. Plant Growth Regul 49:27–34
Author information
Authors and Affiliations
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2019 Springer Nature Switzerland AG
About this chapter

Cite this chapter
Majeed, A., Muhammad, Z. (2019). Salinity: A Major Agricultural Problem—Causes, Impacts on Crop Productivity and Management Strategies. In: Hasanuzzaman, M., Hakeem, K., Nahar, K., Alharby, H. (eds) Plant Abiotic Stress Tolerance. Springer, Cham. https://doi.org/10.1007/978-3-030-06118-0_3
Download citation
DOI: https://doi.org/10.1007/978-3-030-06118-0_3
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-06117-3
Online ISBN: 978-3-030-06118-0
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)