
Abstract
The analysis of gene function in many monocotyledonous species has been held back to a certain extent by the difficulties of stable gene transformation. This chapter discusses the increasing use of transient in planta expression systems available for both RNA interference (RNAi)-mediated downregulation and overexpression of target genes of interest in monocots, as well as the advantages and disadvantages or limitations associated with each approach. In particular, existing virus-induced gene-silencing (VIGS) systems, virus-mediated overexpression (VOX) systems and other common transient expression systems based on microprojectile particle bombardment or Agrobacterium-mediated delivery are reviewed, as well as some of the more specialized expression systems that have been developed recently for functional gene analyses in monocots.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
- Virus-induced gene silencing (VIGS)
- Virus-mediated overexpression (VOX)
- RNAi
- Transient-induced gene silencing (TIGS)
- Barley stripe mosaic virus (BSMV)
- Brome mosaic virus (BMV)
- Rice tungro bacilliform virus (RTBV)
- Cymbidium mosaic virus (CymMV)
- Bamboo mosaic virus (BaMV)
- Small interfering RNAs (siRNAs)
- Bacterial type III secretion system (T3SS)
- Microprojectile particles
- Biolistic bombardment
- Agrobacterium tumefaciens
- Wheat
- Cereals
Introduction
Advances in genome and transcriptome sequencing technologies have led to ever-increasing volumes of data being generated, with a corresponding explosion in the numbers of candidate genes of interest being identified from plants and plant-associating organisms. Identification of the functions of these genes poses a major challenge. The more traditional approaches to determining gene function have included abolishing or highly reducing gene expression using chemical or irradiation mutation, or transfer DNA (T-DNA) insertion mutation. Alternatively, the more recent approach has been to generate stable transgenic plants that overexpress a fragment of a target gene in order to trigger silencing of the corresponding endogenous plant gene, using the process of post-transcriptional gene silencing (also known as RNA interference, RNAi). Stable transformation of plants to overexpress genes of interest to elucidate their function is also common, particularly in model organisms.
In non-model plants and, in particular in many monocotyledonous species, the often large genome sizes, low transformation efficiency and long life-cycles may mean that these stable technologies may be at best, laborious and time-consuming, and at worst, non-applicable. Even in monocots such as wheat (Triticum aestivum) and barley (Hordeum vulgare), for which stable transformation has become a relatively routine technique, transformation protocols have been optimized for only a handful of cultivars in each species.
As a consequence of these drawbacks, there has been increasing interest in transient in planta expression systems that allow expression of double-stranded RNAs (dsRNAs) that trigger silencing of endogenous plant genes and, hence, reduce the expression of proteins of interest. These systems allow relatively rapid prescreening of candidate genes for specific traits or cellular responses. This chapter focuses mainly on viral-based transient gene silencing and protein expression systems available for monocots and recent advances therein. However, some common nonviral-based transient systems are also discussed for comparison along with their associated advantages and limitations.
RNAi-Based Downregulation of Endogenous Plant Genes
Virus-Induced Gene Silencing (VIGS)
Virus-induced gene silencing (VIGS) has been used during the past two decades to study the function of plant genes, first in model dicotyledonous species, but now increasingly in commercially important monocotyledonous species. The technique exploits a plant antiviral defence mechanism called post-transcriptional gene silencing (Waterhouse et al. 2001) in which viral RNA is targeted for degradation in a sequence homology-based manner. The genome of a viral vector is engineered to express a short fragment of a transcribed sequence of a plant gene of interest. When the modified virus infects and spreads throughout inoculated test plants, it triggers silencing both of itself and of the endogenous gene corresponding to the plant gene sequence inserted into the vector. This results in the reduction of target gene transcript levels and, subsequently, target gene activity (Lee et al. 2012) .
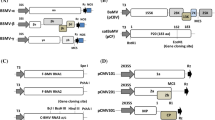
VIGS is particularly useful for prescreening large numbers of candidate genes, and for studying essential genes whose silencing would have a embryonic lethal phenotype. Whilst nearly 30 viruses have been adapted for VIGS in numerous dicot species (Huang et al. 2012), to date there have only been 5 monocot-infecting viruses reported to have been adapted for VIGS: Barley stripe mosaic virus (BSMV) , Brome mosaic Virus (BMV) , Rice tungro bacilliform virus (RTBV) , Cymbidium mosaic virus (CymMV) and, most recently, Bamboo mosaic virus (BaMV) and its associated satellite RNA satBaMV. Details of the VIGS systems reported to date are summarized in Table 14.1. A brief description of each is given below .
BSMV-Based VIGS Systems
BSMV, the type member of the Hordeiviridae , was the first monocot-infecting virus to be adapted for VIGS. It has a tripartite genome which comprises the positive-sense RNAα, RNAβ and RNAγ. The first described BSMV-based VIGS system (Holzberg et al. 2002) was based on complementary DNA (cDNA) clones of the ND18 strain of BSMV under the control of the bacteriophage T7 promoter (Petty et al. 1989). Holzberg et al. (2002) added PacI and NotI sites downstream of the γb gene in the plasmid representing BSMV RNAγ, for the insertion of foreign sequences. A stop codon was introduced between the γb open reading frame (ORF) and the PacI site to prevent translation of the inserted sequences. In addition, the majority of the βa gene, which encodes the viral coat protein (CP), was deleted from the genomic RNAβ. Deletion of the CP appeared to enhance silencing efficiency, although it also had the less desirable effect of increasing viral-induced symptom severity.
There are now a number of variant BSMV–VIGS vectors available, the main details of which are summarized in Table 14.1. In most of these systems, foreign gene fragments are introduced into RNAγ using cloning sites downstream of the γb ORF, as in the original BSMV–VIGS vector developed by Holzberg et al. (2002). An exception is the variant system developed by Tai et al. (2005), in which the start codon for the γb gene sequence was modified to create a BamHI site for the insertion of a foreign sequence. This modification also blocked translation of the BSMV γb silencing suppressor. Kawalek et al . (2012) introduced a cloning site for foreign sequences at the end of the βc ORF in the plasmid representing BSMV RNAβ. When combined with wild-type RNAα and the RNAγ VIGS vector developed by Bruun-Rasmussen et al. (2007; see also Table 14.1 for details) this facilitated simultaneous silencing of two unrelated genes when fragments targeting different genes were inserted into each of the cloning sites on RNAβ and RNAγ. However, delivery of a foreign gene fragment from RNAβ appeared to induce weaker silencing than when gene fragments were inserted into RNAγ (Kawalek et al. 2012) .
Other modifications to the BSMV–VIGS vectors have been made in attempts to increase the throughput and ease of use of this research tool. A DNA-based BSMV–VIGS system developed by Meng et al. (2009) eliminated the costly in vitro transcription step by cloning the BSMV cDNA sequences from the vectors described by Holzberg et al. (2002) into separate binary vectors (whilst retaining the βa gene in the RNAβ). The T7 promoter was replaced with the 35S promoter from Cauliflower mosaic virus and a ribozyme sequence added downstream of each viral cDNA sequence to generate the correct 3ʹ-end after transcription. Particle bombardment could then be used to introduce the plasmids containing the BSMV genomes into barley seedlings (Meng et al. 2009). Another variant retained the T7 promoter but replaced the original cloning site downstream of the CP ORF with a ligation-independent cloning (LIC) site, to facilitate efficient insertion of foreign gene sequences (Pacak et al. 2010a). Yuan et al. (2011) combined these two approaches by cloning the BSMV genomes into binary vectors under the control of a double 35S promoter and introducing a LIC site downstream of the γb ORF. These plasmids were then delivered via agroinfiltration into the leaves of an intermediate host, Nicotiana benthamiana, and the sap from these plants used to rub-inoculate leaves of a large number of monocot plants (Yuan et al. 2011) .
BMV-Based VIGS Systems
BMV is a positive-strand RNA virus of the genus Bromovirus, with a tripartite genome comprising RNA1, RNA2 and RNA3. Ding et al. (2006) created and modified BMV clones to produce a hybrid BMV strain (H-BMV)-based VIGS vector. RNA1 and RNA2 from the rice-infecting fescue strain of BMV (F-BMV) were used in combination with RNA3 of the Russian strain of BMV (R-BMV). This allowed foreign gene sequences to be inserted downstream of the CP ORF using a unique HindIII site in the cDNA clone of R-BMV RNA3 (the F-BMV cDNA clone contained two HindIII sites). Successful silencing of the phytoene desaturase (PDS) gene using the H-BMV vector was demonstrated in barley, rice (Oryza sativa) and maize (Zea mays). However, in order to reduce the severity of visual symptoms induced by the virus itself, Ding et al. (2006) proceeded to modify the cDNA clone representing F-BMV RNA3 by replacing a portion of the intergenic sequence between the movement and CP genes with the corresponding fragment from the R-BMV RNA3 cDNA clone. This intergenic region from R-BMV RNA3 had been shown previously to be responsible for a higher accumulation of R-BMV RNA3 and subgenomic RNA4 during plant infection (Hema and Kao 2004). The resulting BMV vector containing the chimeric RNA3, and RNA1 and RNA2 from F-BMV accumulated to higher levels than the parental F-BMV strain and induced fewer disease symptoms than R-BMV or H-BMV in infected rice plants (Ding et al. 2006) .
The chimeric BMV VIGS system has been modified further more recently by transferring each of the BMV cDNA clones into a binary vector between a double 35S promoter and a ribozyme sequence (Ding et al. 2010; Sun et al. 2013; Table 14.1). In addition, a new multiple cloning site has been added at the 3ʹ-end of the CP ORF to allow directional cloning of foreign gene fragments into the chimeric RNA3 clone. This DNA-based BMV vector can be introduced into rice plants via Agrobacterium-mediated vacuum infiltration (Ding et al. 2010) .
Pacak et al. (2010b) used a different R-BMV-based vector for VIGS studies in rice. In this vector, which was originally developed for studying viral RNA recombination (Alejska et al. 2005), foreign gene sequences are also inserted at a cloning site downstream of the CP ORF. This cloning site comprises SpeI and BamHI sites separated by a 337-nt spacer sequence which allows expression of either separated inverted repeats or a single-gene fragment.
RTBV-Based VIGS System
RTBV is a member of the genus Pararetrovirus, with a monopartite double-stranded DNA genome that contains four ORFs . ORF III encodes a polyprotein which is post-translationally processed into the CP, an aspartate protease and a reverse transcriptase enzyme with ribonuclease H activity. It is possible to remove ORF I, ORF II and part of ORF IV (leaving only ORF III and the remainder of ORF IV) from the virus without removing its ability to replicate and spread systemically in host plants (Purkayastha et al. 2010). These authors generated a RTBV-derived VIGS vector by cloning a 6.1-kb fragment, representing ORF III and truncated ORF IV from an Indian isolate of RTBV, as a partial dimer into the T-DNA of a binary vector. This fragment also incorporated the constitutively expressed maize ubiquitin promoter, a transfer RNA (tRNA)-binding site and the Kozak sequences at the 5ʹ-end, whilst a multiple-cloning site for the insertion of foreign gene fragments was introduced at the 3ʹ-end of the fragment. VIGS of the marker gene PDS using this RTBV vector was demonstrated in rice (Purkayastha et al. 2010) .
CymMV-Based VIGS System
The potexvirus CymMV , which infects many species within the Orchidaceae, is the first virus to have been adapted specifically for VIGS in non-grass monocots (Lu et al. 2007). CymMV has a monopartite positive-sense single-strand RNA genome of approximately 6200 nt, encoding an RNA-dependent RNA polymerase; triple gene block movement proteins 1, 2 and 3; and CP. A symptomless strain of CymMV was modified to function as a VIGS vector by inserting a duplicated fragment (60-nt long) of the subgenomic promoter of the CP gene immediately upstream to the original CP subgenomic promoter. Foreign gene fragments are inserted downstream of this duplicated promoter which subsequently drives transcription of the foreign sequence during viral infection. VIGS of the marker gene PDS and the orchid floral organ identity gene PeMADS6 were demonstrated in the native orchid species Phalaenopsis amabilis (moon orchid) and in a commercial cultivar Phalaenopsis Sogo Musadium (Fig. 14.1) .

Cymbidium mosaic virus (CymMV)-mediated VIGS of the floral organ identity gene PeMADS6 in floral tissue of Phalaenopsis spp. (reproduced with permission from Lu et al. 2007). Plants were inoculated with buffer (a, d and g), an empty virus control construct (b, e and h) or with a CymMV VIGS construct targeting PeMADS6 for silencing (c, f and i). a–c and g–i , Phalaenopsis Sogo Musadium. d–f , Phalaenopsis amabilis var. formosa. Green streaks developed on the sepals, petals and lips of flowers of plants in which PeMADS6 had been silenced (indicated by black arrows), but not on flowers of buffer treated or virus control-infected plants. VIGS virus-induced gene silencing
BaMV-Based VIGS System
BaMV, another member of the Potexvirus genus , also has a monopartite positive-sense single-strand RNA genome that contains five conserved ORFs (Lin et al. 1994; Yang et al. 1997). It is the most recent monocot-infecting virus to have been modified to function as a VIGS vector and the only one for which a satellite RNA associated with the virus has also been modified for VIGS application (Liou et al. 2013). The modified BaMV VIGS vector was generated by the duplication of the subgenomic promoter for the BaMV CP gene, which was inserted upstream of the original CP subgenomic promoter. A multiple cloning site was introduced downstream of the duplicated promoter to allow insertion of foreign gene fragments (Table 14.1).
The BaMV-based VIGS system differs from other VIGS systems for monocots in that the satellite RNA associated with BaMV, satBaMV, can be modified to express a second gene fragment during virus infection unrelated genes can then be silenced simultaneously when fragments from different genes are inserted into the BaMV and satBaMV genomic sequences (Liou et al. 2013; see also Sect. 2.2.3) . Satellite RNAs are dependent on their associated (helper) virus for replication, encapsidation and cell-to-cell movement (Roossinck et al. 1992). Therefore, satBaMV replicates and induces silencing only when it is co-inoculated onto a host plant together with BaMV (Liou et al. 2013) .
satBaMV is a linear RNA molecule of 836 nucleotides, encoding a 20-kDa protein (P20) which is flanked by 5ʹ- and 3ʹ-untranslated regions (Lin and Hsu 1994). Foreign gene fragments can be inserted into satBaMV at an EcoNI cloning site present towards the 3ʹ-end of the P20 ORF. Silencing of single-gene targets using either the BaMV-based vector or a modified satBaMV with an unmodified (i.e. not carrying a target gene fragment) BaMV helper virus has been demonstrated in two model species, namely N. benthamiana and Brachypodium distachyon (Liou et al. 2013). However, simultaneous silencing of two target genes by expressing a fragment of one gene from BaMV, and a fragment of the second gene from satBaMV has only been demonstrated in N. benthamiana (Liou et al. 2013) .
Comparison of the Infection Biology of the Viral Vectors Currently Adapted for VIGS
One of the first considerations when assessing the applicability of a VIGS vector is the viral vector host range (see Table 14.2). However, of equally high importance is the biology of the host–virus interaction. For example, BSMV can be transmitted through the seed and pollen of infected plants in many species, and BSMV-mediated VIGS has been demonstrated in wheat ears, pollen and developing grain and even in the progeny of infected plants (Bennypaul et al. 2012; Lee et al. 2012; Ma et al. 2012b). BSMV-mediated VIGS has also been reported in wheat roots (Bennypaul et al. 2012) and wheat stems (Fig. 14.2), as well as in the leaves of many monocot species (Holzberg et al. 2002; Scofield et al. 2005; Renner et al. 2009; Demircan and Akkaya 2010; Wang et al. 2010; Pacak et al. 2010a; Martin et al. 2013). By contrast, BMV, CymMV, RBTV and BaMV have not been reported to be seed or pollen transmitted (Description of Plant Viruses database, www.dpvweb.net), and it is unlikely that these viruses would be able to mediate VIGS in pollen or developing grain. Nonetheless, the CymMV-based VIGS system has been used to silence gene expression in orchid floral tissue (Lu et al. 2007; Fig. 14.1). Currently, there are little data available on the applicability of BMV-, RBTV- or BaMV-mediated VIGS to plant tissues other than leaves.

Barley stripe mosaic virus-mediated VIGS of phytoene desaturase (PDS) gene in wheat leaves, leaf sheaths and ears. Silencing of PDS results in a photobleached phenotype in aerial tissues of BSMV:asPDS-infected wheat plants (a). Upper uninoculated leaves from control (BSMV:asGFP-infected; top) or BSMV:asPDS-infected (bottom) plants. B–C. Leaf sheaths of b BSMV:asPDS-infected and c BSMV:asGFP-infected wheat plants. D–E. Ears of d BSMV:asGFP-infected or e BSMV:asPDS-infected plants. VIGS virus-induced gene silencing
The Potential for Improving Existing VIGS Systems
VIGS was first deployed as a reverse genetics tool in monocot species for the functional analysis of host genes involved in plant–fungal pathogen interactions (Hein et al. 2005; Scofield et al. 2005). In recent years, there has been an increased application of VIGS to disciplines beyond fungal plant pathology, such as those relating to the study of plant cell wall biosynthesis (Oikawa et al. 2007; Held et al. 2008), insect–wheat interactions (van Eck et al. 2010), nutrient uptake and translocation (Pacak et al. 2010a; Li et al. 2014), plant–virus interactions (Shi et al. 2011; Cao et al. 2012), abiotic stress tolerance (Liang et al. 2012; Rong et al. 2014), leaf development (Wang et al. 2011; Ma et al. 2012a), grain development (Bennypaul et al. 2012), and flower development (Lu et al. 2007), amongst others. As the application of VIGS in monocots has increased, naturally so has interest in developing and modifying existing VIGS systems for enhanced deployment. In particular, there have been significant advances in improving BSMV- and BMV-based VIGS. However, it is evident that there are still many limitations associated with these systems. The approaches that have been used to reduce or overcome these limitations are discussed later, together with potential methods for further improving existing VIGS systems in monocots. As the aim of this chapter is to provide an overview of existing transient expression systems in monocotyledonous species, the potential of modifying other viruses as VIGS vectors is not discussed here.
Assessing and Reducing Interference from Virus-Induced Symptoms
An important consideration when utilizing VIGS to investigate plant gene function is that infection of host plants by the virus vector itself may influence host developmental and cellular processes. Varying degrees of chlorosis and stunting are common symptoms induced by viral infections, and may make the phenotype due to silencing of target genes difficult to analyze when symptoms are severe. It is therefore important to include appropriate controls during VIGS experiments, to determine the effect or influence of virus infection itself on the phenotype under investigation. This is particularly true when studying responses to abiotic or biotic stresses, as virus infection may activate certain host defence signalling pathways that could affect plant responses to other pathogens or stresses. For example, infection of wheat plants with BSMV was reported to enhance resistance to the blast fungus Magnaporthe oryzae, although the interaction with Blumeria graminis, the causal agent of powdery mildew disease , did not appear to be affected (Tufan et al. 2011).
The severity of symptoms induced by BSMV- and BMV-silencing vectors has been shown to be variable between different varieties or genotypes within the same species (reviewed by Ramanna et al. 2013). Careful selection of plant varieties for VIGS experimentation can help to mitigate the problem of severe VIGS vector-induced symptoms that may obscure or confuse gene function analyses. However, silencing efficiency is also variable and the choice of host genotype may therefore require a compromise between varietal differences in silencing efficiency and severity of symptoms induced by the VIGS vector (see the section ‘Silencing Stability and Efficiency’).
Another possible strategy to minimize undesirable side effects of infection by the viral vector on host development and metabolic processes is to develop a VIGS vector that induces mild or no visible symptoms. The CymMV-based VIGS vector was developed using a specially selected symptomless strain of the virus (Lu et al. 2007). The RTBV-based VIGS vector also infects rice plants without inducing visible systems (Purkayastha et al. 2010). However, in the RTBV-VIGS system, this was suggested to be due to the replacement of the original RTBV promoter by the maize ubiquitin promoter, as it is thought that the symptoms normally associated with RTBV infection in rice are related to the sequestration of two host transcription factors via interactions with a Box II element in the viral promoter (Dai et al. 2008). Thus, it may be possible to minimize symptom severity by utilizing different viral strains, or by modifying or removing elements associated with symptom induction (if these are known) in the viral vector . It must be noted though that the absence of visible virus-induced symptoms does not indicate that there are no effects on host metabolism and signalling at the cellular level. A complete lack of virus symptoms on experimental plants could also mean that plant infection would have to be verified by enzyme-linked immunosorbent assay (ELISA)- or reverse transcription polymerase chain reaction (RT-PCR)-aided detection of the virus vector, rather than by monitoring the appearance of viral symptoms.
Silencing Stability and Efficiency
A range of factors affects the stability and efficiency of silencing that can be achieved in a VIGS experiment. A problem associated particularly with VIGS is the question of foreign gene fragment insert stability in the virus vector. In small grain cereal species in particular, VIGS phenotypes are generally observed only in two or three systemically infected leaves and in the intermediate stem tissue when young vegetative stage plants are inoculated (Holzberg et al. 2002; Scofield et al. 2005; Ding et al. 2006). Usually, the silencing phenotype in newer leaves becomes increasingly patchy and incomplete until it appears to fade entirely. This tends to be associated with the loss of the foreign gene insert from the virus vector through recombination (Bruun-Rasmussen et al. 2007). Both the size and possibly the actual sequence of the insert appear to affect insert stability and, therefore the efficiency of silencing. Studies carried out with fragments of different lengths inserted into the BSMV vector indicated that fragments larger than 500 bp are often unstable, whilst fragments smaller than 120 bp may not efficiently induce silencing (Scofield et al. 2005; Bruun-Rasmussen et al. 2007; Yuan et al. 2011).
The orientation and design of foreign gene fragments in the vector may also have a significant effect on silencing efficiency. Fragments inserted into the BSMV or BMV vectors in antisense orientation generally induce more efficient silencing (and never less efficient silencing) when compared to fragments inserted in sense orientation (Lacomme et al. 2003; Pacak et al. 2010b). The expression of a short direct-inverted repeat from a BSMV vector was suggested to induce stronger and more stable silencing than expression of an antisense fragment for the same target gene (Lacomme et al. 2003). However, later studies showed that short inverted repeats were in some cases highly unstable and less efficient than antisense fragments inserted into BSMV and BMV VIGS vectors (Pacak et al. 2010a; b). Therefore, it may be that the stability of short inverted repeats is dependent on their specific sequence.
VIGS construct design can be aided by the use of predictive tools such as the siRNA-Finder (si-Fi) software developed at IPK-Gatersleben in Germany (http://labtools.ipk-gatersleben.de/). This software allows the researcher to pinpoint regions of a target gene sequence that are most likely to generate a large number of small interfering RNAs (siRNAs) efficient in inducing gene silencing. si-Fi can also be used to help design constructs for stable RNAi or VIGS with no or minimal off-target silencing effect (provided the complete genome or the transcriptome sequences from the host plant species are available). Whereas the in silico predictions are not a substitute for in planta validation of the efficiency of VIGS constructs, si-Fi can be very useful when designing VIGS constructs (Lee et al. 2014).
The choice of host genotype is another important factor that influences silencing efficiency. Several studies have shown that there can be a substantial variation in the level and persistence of silencing induced by the same VIGS construct in different varieties or genotypes within the same host species (Hein et al. 2005; Meng et al. 2009; Cakir and Tör 2010; Bennypaul et al. 2012). In some wheat genotypes inoculated at the two-leaf stage, BSMV-induced silencing of PDS may persist throughout the developing plant into the flag leaf and spike, and may even be inherited in a percentage of the progeny of these plants, whilst in others the photobleaching phenotype associated with PDS silencing is no longer visible in the sixth leaf (Bruun-Rasmussen et al. 2007; Bennypaul et al. 2012; Lee et al. 2012).
An additional consideration related to VIGS is that of the ambient temperature. Generally, daytime temperatures of 20–24 °C appear to yield better BSMV-induced gene silencing in wheat and barley than temperatures of 16–18 °C or 26–28 °C (Scofield et al. 2005; Bruun-Rasmussen et al. 2007; Cakir and Tör 2010). Similarly, Purkayastha et al. (2010) observed RTBV-mediated silencing of PDS in the majority (around 80 %) of inoculated rice plants when the plants were kept at 27 °C, but not in plants in glasshouses maintained at 30 °C, although there were also other differences in the growth conditions that may have affected silencing efficiency. In addition to the effect of temperature, it has been found that light intensities greater than 300 μmol m−2 s−1 appear to induce more severe symptoms in wheat cultivars in response to BSMV infection in specific growth room conditions (i.e. 23 °C day/20° C night, 16 h light), perhaps because the higher light levels compound the stress induced by virus infection (Lee WS, Kanyuka K, unpublished).
Silencing of Multiple Genes
As VIGS, like all other RNAi-based techniques, involves silencing of target genes based on their sequence homology to the silencing inducer dsRNA expressed inside plant cells, it is possible using one VIGS construct to knock-down expression of related or even families of genes by targeting sequences conserved between these genes. What is more difficult is to silence two unrelated genes in the same plant cell. Limited success has been achieved by inserting fragments of two individual genes joined in cis into BSMV RNAγ (Cakir and Scofield 2008; Cakir and Tör 2010).This enabled simultaneous silencing of the two target genes although the degree of silencing for each gene was highly variable between replicate experiments, and was always less efficient than silencing induced by constructs targeting only one plant gene. Interestingly, the order the fragments were inserted into the virus vector appeared to affect the degree of silencing observed for each gene (Cakir and Tör 2010). Mixed infections in which BSMV RNAα and RNAβ were combined with two BSMV RNAγ sequences, each carrying a fragment of a different target gene, also induced limited silencing of both genes in the same area of leaf tissue. However, silencing efficiency again was highly variable, and it was not possible to confirm whether both genes were silenced in the same plant cell (Cakir and Scofield 2008; Cakir and Tör 2010). In this respect, the BaMV/satBaMV combined VIGS system described in Sect. 2.1.5 differs because the helper virus and satellite RNA replicate together in the same host cell. Hence, when BaMV and satBMV have been modified to carry different silencing constructs, any observed silencing of target genes is likely to have been activated in the same plant cell as discussed further later.
More success was observed in a recent study in which two individual fragments targeting different genes were inserted into BSMV RNAβ and RNAγ, respectively (Kawalek et al. 2012). Simultaneous silencing of the two target genes was observed consistently, although silencing from RNAγ was always stronger than from RNAβ, probably because BSMV RNAγ accumulates to much higher levels than that of RNAβ during plant infection. However, foreign inserts in BSMV RNAβ were more stable than sequences inserted into RNAγ (Kawalek et al. 2012). Interestingly, when fragments targeting the same gene were inserted into both RNAβ and RNAγ, the overall level of silencing was much better than when fragments were expressed from one genomic RNA component alone, suggesting that this could be an attractive strategy for inducing more efficient and possibly more stable silencing of a single target gene or gene family (Kawalek et al. 2012).
In a different approach, simultaneous silencing of two genes in the same plant cell using BaMV and its associate satellite RNA satBaMV was reported in N. benthamiana by Liou et al. (2013). In this study, green fluorescent protein gene (GFP)-transgenic N. benthamiana plants were co-infected with BaMV modified to express a fragment of the sulphur gene and satBaMV carrying a fragment of GFP. Although simultaneous silencing of two genes has not yet been demonstrated in a monocot species with this system, successful silencing of single-gene targets by expressing a gene fragment from either BaMV or satBaMV has been reported in B. distachyon (Liou et al. 2013), indicating that this may be a viable system for multiple gene silencing in monocots. However, recombinant BaMV and satBaMV from which gene sequence inserts had been partially or fully lost (see the section ‘Silencing Stability and Efficiency’) could be detected already in both the directly inoculated and systemically infected leaves of B. distachyon plants by 16 days post inoculation (Liou et al. 2013). Further studies and investigation into the stability and duration of silencing induced by this BaMV-/satBaMV-based system are required to assess applicability of the system to other monocotyledonous species.
Expanding the Host Range of Existing VIGS Systems
To date, most monocot VIGS studies have been carried out predominantly in the crop species barley (Hordeum vulgare), hexaploid wheat (Triticum aestivum) and, to a lesser extent, maize (Zea mays) and rice (Oryza sativa). However, the successful application of VIGS has been reported for a growing number of other, mostly small grain cereal crops (Table 14.2), although in many of these only silencing of the visual marker PDS has been demonstrated (Renner et al. 2009; Demircan and Akkaya 2010; Pacak et al. 2010a; Martin et al. 2013). Both BSMV and BMV have a broad experimental host range, particularly within the Poaceae (Table 14.2), and there is considerable potential for the application of VIGS to other grass and non-grass species using the vectors currently available. For example, in a recent publication (Ramanna et al. 2013), there was mention of an unpublished but successful use of the BMV binary VIGS vector for gene silencing in foxtail millet (Setaria italica) and switchgrass (Panicum virgatum). Apparently, this was done by first delivering the BMV plasmids into N. benthamiana leaves via agroinfiltration and using the sap to rub-inoculate leaves of monocots. Similarly, CymMV has a broad host range within the Orchidaceae, one of the two largest families of angiosperms. Therefore, future applications for this VIGS vector may be expected to extend beyond the orchid species within the genus Phalaenopsis.
When testing the applicability of existing VIGS systems in other monocotyledonous species, multiple varieties or accessions should be tested in order to identify genotypes in which efficient silencing is coupled with moderate or mild virus-induced symptoms (see sections ‘Assessing and Reducing Interference from Virus-Induced Symptoms’ and ‘Silencing Stability and Efficiency’). In addition, certain genotypes may be resistant to the VIGS vector, whilst other accessions of the same species may be susceptible. For example, of 150 barley accessions and landraces tested at Rothamsted Research with the ND18 wild-type strain of BSMV, 89 were potentially suitable hosts for further VIGS studies, 32 developed overly severe symptoms and there was poor or no infection in 29 genotypes (Lee WS, Ruiz O, Kanyuka K, unpublished).
The currently available BMV-VIGS vectors are based on either the R-BMV strain or a hybrid virus comprising sequences from the two different strains, R-BMV and F-BMV (see the section ‘BMV-Based VIGS Systems’) . Similarly, almost all of the BSMV-VIGS vectors described to date are based on the ND18 strain of BSMV. The one exception is a vector system described by Pacak et al. (2010a), comprising RNAα and RNAβ from the oat-infecting strain CV42 in combination with RNAγ modified for insertion of foreign sequences from the ND18 strain. The ND18 strain does not infect oat. Using this hybrid BSMV, Pacak et al. (2010a) were able to observe limited photobleaching due to silencing of PDS in diploid oat (Avena strigosa) and hexaploid oat (Avena sativa) cultivars. Whilst the silencing phenotype observed was not very robust in this study, these results demonstrate the potential for utilizing hybrid or alternative viral strains to expand the VIGS target host range.
Nonviral Transient-Induced Gene-Silencing Systems
There are a number of nonviral transient gene-silencing systems that utilize the innate plant defence mechanism of post-transcriptional gene silencing . As with VIGS, these systems involve the delivery of dsRNA fragments to plant cells that activate the host RNA silencing machinery, leading to sequence-specific degradation of a target RNA. Two of these systems, namely microprojectile bombardment and Agrobacterium-mediated delivery, have been applied for gene function studies in monocots. The advantages and disadvantages associated with each system are discussed below.
Microprojectile/Biolistic-Bombardment-Based Expression
Microprojectile bombardment , otherwise known as biolistic bombardment, has been used extensively for single-cell transient overexpression of DNA in tissues of monocot plants since the early 1990s (see section ‘Microprojectile/Biolistic Bombardment’). In 2000, microprojectile bombardment of dsRNA into leaf epidermal cells was used to trigger transient-induced gene silencing (TIGS) of target genes in maize, barley and wheat (Schweizer et al. 2000). Microprojectile bombardment-mediated TIGS has also been demonstrated in rice protoplasts and leaf cells (Miki and Shimamoto 2004). Since then, this technique in monocots has been applied most extensively to the identification and study of wheat and barley genes involved in resistance or susceptibility to different formae speciales of the powdery mildew fungus Blumeria graminis (Azevedo et al. 2002; Christensen et al. 2004; Douchkov et al. 2005; Dong et al. 2006; Zimmermann et al. 2006). Powdery mildews only attack the outermost epidermal cell layer, thus making this single-cell TIGS system highly amenable to the study of cereal–powdery mildew interactions. As TIGS is triggered within 2–3 days after the target tissues have been bombarded with tungsten or gold microparticles (of approximately 0.4- or 2-μm diameter, respectively) coated with dsRNA (Schweizer et al. 2000) this technique has the advantage of being relatively rapid. Furthermore, the development of high-throughput cloning methods for the generation of inverted-repeat RNAi constructs for microprojectile bombardment, such as one based on the Gateway cloning system and reported by Douchkov et al. (2005), means that libraries of gene sequences can be generated and screened in a high-throughput manner.
Another advantage of microprojectile bombardment-mediated TIGS is that plasmids carrying silencing constructs can be co-bombarded with plasmids containing reporter gene constructs. As co-bombarded plasmids have a high co-expression rate, this aids the identification of dsRNA construct transformed cells as reporter gene expression can be used as an indicator of successful transformation (Schweizer et al. 1999; see also section ‘Microprojectile/Biolistic Bombardment’) . Indeed, microprojectile bombardment can be used to deliver multiple plasmid vectors into the same cell, as the gold or tungsten particles can be coated with plasmid DNA comprising a mixture of different vectors. This was demonstrated by Marzin and colleagues (2008), who co-bombarded barley leaf epidermal tissue with three plasmids; two encoding GFP and Discosoma sp. red fluorescent protein (DsRed), respectively, and one carrying an RNAi construct in order to trigger TIGS against a candidate gene of interest. This combination was used to test candidate barley genes for their involvement in cell-autonomous responses to dehydration stress by monitoring the effect of TIGS of target genes on DsRed fluorescence. The fluorescence of DsRed is reduced under denaturing conditions, such as those imposed by drought stress (Marzin et al. 2008). Although TIGS of multiple gene targets using co-bombardment of two or more dsRNA constructs has yet to be reported in monocots, this is a distinct possibility.
A notable disadvantage of microprojectile bombardment-mediated TIGS is that silencing is only triggered in directly bombarded cells. Thus, although this technique is useful for studying genes involved in cell-autonomous processes or single-cell interactions in leaf or potentially root epidermal cell layers (see the section ‘Microprojectile/Biolistic Bombardment’), it is not suitable for studying genes involved in developmental processes or plant–pathogen interactions which involve multiple cells or whole tissues (Marzin et al. 2008). The set-up costs of establishing a microprojectile bombardment system can also be expensive.
Agrobacterium-Mediated Delivery
Agrobacterium tumefaciens-mediated transient gene-silencing assays have been used increasingly to study gene function in a number of dicotyledonous species (Kapila et al. 1997; Johansen and Carrington 2001; Wroblewski et al. 2005; Bhaskar et al. 2009) . A. tumefaciens delivers RNAi constructs into plant cells where they trigger TIGS. Unlike microprojectile bombardment, Agrobacterium-mediated TIGS does not require specialized equipment, and silencing of target genes is induced throughout agrobacteria-infiltrated tissue, allowing the study of genes involved in multicellular processes. However, many monocotyledonous species are considered to be either a non-host or a poor host for A. tumefaciens. It is for this reason that Agrobacterium-mediated transformation of monocotyledonous cereal plants is relatively inefficient. As a consequence, cereal transformation groups as well as specialist laboratories tend to work with only 1–2 cereal genotypes and 1–2 compatible A. tumefaciens strains capable of transferring the T-DNA into the host plant’s nuclear DNA (Sparks et al. 2014). Similarly, transient Agrobacterium-mediated expression in non-embryogenic tissues, such as leaves, is especially problematic in cereal species. This is because the standard pressure infiltration of Agrobacterium suspensions into these tissues using a needleless syringe is very difficult due to several intrinsic structural features, such as extensive epidermal cuticular waxes, considerable silica content and the low volume of intercellular space. In various dicotyledonous species, this procedure, known as ‘agroinfiltration’, is used routinely to explore gene function (Vaghchhipawala et al. 2011) .
In spite of the difficulty of agroinfiltration into cereal leaves, a method for Agrobacterium-mediated TIGS in leaves through transient gene expression in rice (O. sativa) has recently been reported (Andrieu et al. 2012). Leaves of japonica and indica rice plants were mechanically wounded using 600-μm diameter needles mounted onto a custom-made apparatus. The wounded leaves were then incubated for 30–60 min in a suspension of Agrobacterium (OD600 nm of 0.5–0.8) containing the surfactant Silwet L-77. Andrieu et al. (2012) were able to introduce hairpin RNAi constructs targeting the phytoene desaturase (OsPDS) and SLENDER 1 (OsSLR1) genes into rice leaves, and demonstrated siRNA accumulation in the agroinfiltrated leaf area for both constructs. This was associated with a reduction of the targeted rice mRNA concentrations in both the agroinfiltrated and adjacent tissues, indicating that the gene-silencing mechanism had been activated. The same procedure was also used to overexpress transiently the β-glucuronidase (gus) reporter gene in rice leaves (Andrieu et al. 2012; see also the section ‘Agrobacterium-Mediated Expression). It remains to be determined whether other rice genotypes and, indeed, other cereal species such as wheat and barley are equally amenable to transient transformation via this new procedure .
Transient Expression Assays
Microprojectile/Biolistic Bombardment
The initial concept of transient expression was first developed using electroporation of isolated plant protoplasts (Fromm et al. 1985) . Microprojectile bombardment for DNA delivery directly into live plant cells was developed soon after (Klein et al. 1987). This method remains one of the most frequently used techniques for stable genetic transformation of various monocotyledonous cereals (Sparks and Jones 2014). In microprojectile bombardment-mediated transformation , the construct designed for in planta protein expression typically contains a complete gene or just the coding sequence (CDS) of a gene of interest flanked by an appropriate promoter on its 5ʹ-end, with an appropriate terminator sequence on its 3ʹ-end. The gene construct is coated onto gold or tungsten microparticles (approximately 0.4–2 µm in size), which are then bombarded using high pressure from a helium gun into cells that exhibit embryogenic competence, such as immature embryos, scutella, immature inflorescences, and shoot tips. Finally, genetically transformed plants are regenerated from these bombarded tissues following labour-intensive tissue culture procedures.
Due to its relative simplicity, speed and ability to deliver DNA into various plant organs, tissue and cell types, microprojectile bombardment is also often used for transient gene expression. For instance, this method is particularly useful for the identification of novel promoters and promoter elements and for analyses of their activity, e.g. level and timing of expression or tissue and organ specificity. Most frequently, the constructs used in these analyses have novel promoter sequences upstream of genes that encode reporter proteins, such as GFP, β-glucuronidase (GUS) or luciferase, expression of which can be visualized easily by microscopy-based techniques and can be quantified (Fig. 14.3) .

Effects of developmental age of caryopses on transient expression of the GUS chimeric constructs under the control of cereal seed storage protein gene promoters (reproduced with permission from Hwang et al. 2001). Promoters: rice glutelin gene Gt3, rice glutelin gene GluB-1, rice glutelin gene GluB-2, rice prolamin gene PG5a, rice prolamin gene RP6, rice globulin gene Glb, and wheat glutenin gene B×7. Rice immature endosperm was isolated from three different stages of caryopses and used as a target material for biolistic bombardment. Histochemical analysis of bombarded endosperm was carried out following 24-h incubation at 25 °C
Microparticle bombardment has also proved to be useful for analyzing the localization of proteins to particular cellular compartments, such as the nucleus, vacuole, plasma membrane, endoplasmic reticulum (ER) and Golgi bodies, amongst others. Expression constructs used in these studies typically contain a gene of interest fused in frame to GFP or another fluorescent protein reporter, e.g. yellow fluorescent protein (YFP), DsRed, mCherry fluorescent protein or monomeric red fluorescent protein (mRFP) and placed under the control of a strong constitutively active promoter such as that found in the rice actin (Act-1) or maize ubiquitin (Ubi-1) genes. Subcellular localization of the fluorescent protein tag can be analyzed using confocal laser scanning microscopy. Peels of onion epidermal cells and wheat leaf sheath epidermal cells are frequently used for these studies, as these cells are relatively large, translucent and lack chloroplasts (Fig. 14.4) .

The overlay projections of confocal stacks spanning onion epidermal cells simultaneously transformed with the GFP-tagged barley sucrose synthase genes and the mCherry-tagged subcellular markers (reproduced with permission from Barrero-Sicilia et al. 2011). Barley sucrose synthases: HvSS1: a, e, i, m, q; HvSS2: b, f, j, n, r; HvSS3: c, g, k, o, s; HvSS4: d, h, l, p, t. Five different organelle markers specific for (a–d) endoplasmic reticulum, (e–h) plasma membrane, (i–l) Golgi apparatus, (m–p) plastids and (q–t) mitochondria. GFP green fluorescent protein
In monocots, genes for anthocyanin biosynthesis are commonly used reporters for transient expression assays, and have been exploited to demonstrate the applicability of microprojectile bombardment-mediated gene expression in maize aleurone and tassel primordial tissues (Klein et al. 1989; Dupuis and Pace 1993), wheat inflorescences (Leduc et al. 1994) and barley coleoptile and leaf epidermal cells (Nelson and Bushnell 1997). GUS reporter protein fusions have also been used to demonstrate the viability of this transient expression technology in the leaves, coleoptiles and roots of rice, barley and perennial ryegrass (Lolium perenne; Hensgens et al. 1993), and in co-bombardment studies as a marker of transformed cells in many monocots. Since the late 1990s to early 2000s, transient expression and/or gene silencing of candidate genes using particle bombardment of leaf cells in monocots has also been applied in the field of molecular plant pathology. This technology has revolutionized research on the cell biology and molecular analyses of cereal–powdery mildew (Blumeria graminis f. sp. tritici and B. graminis f. sp. hordei) interactions . These obligate biotrophic fungal plant pathogens, mainly known to infect wheat and barley, invade exclusively epidermal leaf cells, which are ideal for bioimaging studies. In addition, defences mounted by the plant in response to powdery mildew appear to operate cell autonomously, and can be dissected using a biolistic single-cell transformation approach. The short life cycle of powdery mildew (3–5 days from the initial spore germination to the production of the next generation of asexual spores) also permits the use of detached leaves, and the cutting out of ‘windowpanes’ of only epidermal cells, which simplifies the pathoassays. In this approach (Panstruga 2004; Hückelhoven and Panstruga 2011), detached leaves are bombarded with microprojectiles carrying at least two different constructs, one for overexpression or silencing of a candidate gene, and another for expression of a reporter protein, e.g. GUS or GFP, and then inoculated with fungal conidiospores. Co-bombardment is known to result in a high frequency of construct co-integration in plant cell nuclear DNA, such that the majority of single-leaf epidermal cells that express the reporter protein is also likely to be expressing a test gene construct. It is therefore possible to determine whether the introduced test construct has had any impact on the outcome of the particular cereal genotype–powdery mildew isolate interaction by assessing the success of infection or monitoring fungal development throughout the infection time course in the reporter protein-expressing plant cells (Fig. 14.5) . This type of experimentation can also be supplemented by using various compounds that stain specifically fungal structures and/or certain cellular compartments, providing additional valuable insights and better understanding of cell biology of fungal–plant interactions (Weis et al. 2013). This remains a very intensive area of research, which continues to provide exciting novel discoveries.

Micrograph of successful fungal penetration on a biolistically transformed barley epidermal cell (reproduced with permission from Panstruga 2004). A barley leaf was bombarded with a GUS reporter construct. Subsequently, the leaf was inoculated with powdery mildew conidia and, at 48 h post inoculation, stained for GUS activity. Fungal structures were highlighted by Coomassie Brilliant Blue for microscopic evaluation. Successful penetration is indicated by the presence of a haustorium and elongating secondary hyphae. c, conidiospore; h, haustorium; s, secondary hyphae. Scale bar = 20 μm
Agrobacterium-Mediated Expression
A. tumefaciens-mediated transfer and transient expression of binary Ti plasmid-based vector constructs in leaf cells has been used extensively for testing gene function in a number of dicotyledonous species . In contrast to the microprojectile bombardment-mediated method used to introduce constructs into plant tissues, which allows heterologous protein expression or gene silencing in single cells, A. tumefaciens is able to deliver a binary T-DNA vector to the genome of most of the plant cells in the infiltration zone of the leaf, and also allows experiments to be carried out using non-sterile glasshouse-raised plants.
Unfortunately, the standard methods of agroinfiltration developed for dicots do not work well in many monocotyledonous species (see discussion in the section ‘Agrobacterium-Mediated Delivery’). Therefore, several strategies have been used to improve this procedure for monocots, including mechanical wounding of tissues prior to or during agroinfiltration (e.g. by sonication, mixing by vortex with carborundum, or wounding of plant tissue with a needle), and the incorporation of additives to the agroinfiltration medium. Surfactants, such as Li700 and Silwet-L77, and/or various thiol compounds including L-cysteine and dithiothreitol, are thought to inhibit wound- and pathogen-induced responses (Chen et al. 2010) .
In a typical example, Requesens et al. (2010) developed an efficient and reliable transient A. tumefaciens-mediated assay for testing the functionality of endosperm-preferred promoters in maize. In this method, kernels were isolated from maize ears by cutting the kernels at the base with a scalpel, and dissecting out and discarding the embryos. The pericarp was peeled away exposing approximately three quarters of the endosperm surface, thus increasing the area available for contact with the agrobacteria. The remaining developing endosperm was co-cultivated with Agrobacterium suspensions for 3–5 days following vigorous mixing by vortexing and subsequent sonication for 30 s.
Wounding of plants in these bioassays may interfere with functional analyses of stress-related genes. Vacuum infiltration has been used as an alternative method for introducing Agrobacterium into otherwise unwounded leaves in harvested switchgrass (Panicum virgatum; VanderGheynst et al. 2008). Detached leaves were vortexed with a suspension of A. tumefaciens carrying a GUS reporter construct and non-ionic surfactant added before the application of a low-pressure vacuum (25 kPa). The infiltrated leaves were incubated on moistened sterile filter paper at 22 °C in the dark for 3–6 days. Although GUS expression could be detected in the switchgrass leaf cells after 3 days incubation, leaf decay was observed by 6 days of incubation (VanderGheynst et al. 2008). In a more recent report, a method was described for Agrobacterium-mediated transient expression in several cereals (namely rice, barley, maize, oats, rye, sorghum and wheat), which allows functional analyses of genes involved in various abiotic stress responses and which eliminates the need for wounding of plants (Dhadi et al. 2012). In this procedure, young 12–15-day--old plantlets were uprooted, cleaned, co-cultivated with Agrobacterium harbouring the expression construct in half strength Murashige and Skoog (0.5 MS) basal salt medium supplemented with acetosyringone, a natural phenolic wound response product, at 28 °C for 15 h on a rotating platform. After co-cultivation, the plantlets were incubated for 8–12 h with 0.5 MS medium supplemented with the antibiotic carbenicillin to prevent bacterial contamination .
In almost all the published work to date, A. tumefaciens-mediated protein expression has been shown to be maintained only transiently, usually during the first 5–7 days after agroinfiltration or cocultivation. However, a new method involving cocultivation of mature seeds with agrobacteria appears to enable a longer period of transient expression (Fursova et al. 2012). The seeds were trimmed with scissors to remove about the uppermost one third of each seed, leaving the intact embryo and a ‘sufficient’ portion of the cotyledon. Trimmed seeds were incubated with A. tumefaciens in the presence of acetosyringone, Silwett-L77 and plant extracts from Nicotiana tabacum leaves (these are known to contain metabolites inducing Agrobacteria to initiate gene transfer) for 30 h at 21 °C (Fursova et al. 2012). Seed trimming appeared to aid Agrobacterium penetration through the intracellular spaces of the seed tissues and allowed efficient transformation of the embryonic cells. On average, 10 % of the cocultivated seeds were able to germinate and the resulting plantlets maintained active expression of proteins for up to 12 weeks. This allowed analyses of the effects of gene expression at different stages throughout plant development. Currently, this method has been described only for the model grass species B. distachyon. However, this method may prove to be very useful in functional genomics studies if it can be applied to other monocots, including cereal crops .
Bacterial Type III Secretion System-Based Expression
As discussed earlier, Agrobacterium-mediated transient expression is not very efficient in many monocots, especially in wheat and barley , whereas microprojectile bombardment-mediated gene expression is technically challenging, with expensive set-up costs. This technology also only permits protein expression in individual cells scattered among many untransformed cells. These drawbacks of the ‘conventional’ technologies necessitate development of novel transient expression systems for cereals that allow higher throughput analyses and expression in all or most cells in the target tissue or region.
Many pathogenic bacteria use the type III secretion system (T3SS), a complex protein assembly that is said to resemble a syringe with a needle, to inject small secreted proteins (effector proteins) into the cytoplasm of targeted plant cells to initiate and aid infection (Block et al. 2008). During the past decade, T3SS of various plant pathogenic bacteria have been utilized for protein delivery into cells of dicots, mainly Arabidopsis thaliana and Nicotiana benthamiana. In this system, a heterologous protein is fused to the N-terminus of one of the well-characterized bacterial effector proteins, such as AvrRps4 or AvrRpm1, that contain signal peptides for T3SS-mediated delivery . The main focus of many of these studies has been functional analyses of cytoplasmic effector proteins predicted in the genome sequences of various bacterial, oomycete and fungal pathogens of plants (Sohn et al. 2007; Whisson et al. 2007; Rentel et al. 2008; Fabro et al. 2011; Goritschnig et al. 2012).
Very recently, a similar approach has been applied for delivery of pathogen effector proteins into leaf cells of monocots. The bacterial pathogen of rice, Burkholderia glumae, was used in a rice leaf sheath inoculation assay for in planta expression, and subcellular localization analyses of effector proteins of the rice blast fungus Magnaporthe oryzae (Sharma et al. 2013). In the same study, the B. glumae system was shown to deliver fluorescently labelled fungal effectors to wheat and barley leaf sheath cells and, therefore, this system may be applied for effector discovery. However, the pathogenic nature of this bacterium may restrict its use .
Another system utilizes the soil-dwelling bacterium Pseudomonas fluorescens. This benign species is non-pathogenic on plants and currently seems to have greater potential to become used widely for studies on the detailed characterization of effectors from different plant-pathogenic organisms. The P. fluorescens strain Pf0–1 naturally lacks an endogenous T3SS-encoding region as well as candidate type-III effector genes. Thomas et al. (2009) integrated the entire T3SS-encoding region from a plant pathogenic bacterium, Pseudomonas syringae pv. syringae strain 61, into the genome of P. fluorescens Pf0–1. This engineered strain, known as EtHAn (Effector-to-Host Analyzer), is capable of delivering individual bacterial type III effector proteins, or effector proteins of a nonbacterial origin (as C-terminal fusions to the T3SS signal), directly into the cells of different dicotyledonous plants. The EtHAn-mediated effector delivery system has been shown to work moderately well in wheat (Yin and Hulbert 2011) . A modified and improved version of this system has been used recently in a relatively high-throughput screen of stem rust fungus Puccinia graminis f. sp. tritici effectors in wheat. This resulted in identification of a candidate avirulence effector capable of inducing genotype-specific hypersensitive response that depended on the presence of the Sr22 resistance gene (Upadhyaya et al. 2014) .
Virus-Mediated Overexpression
Transient expression using plant virus vectors is known as VOX , virus-mediated overexpression, and can provide rapid and high-level production of a recombinant protein. Since the mid-1990s, many plant viruses (mainly those with positive-sense single-stranded RNA genomes) have been cloned and modified to express foreign peptides and proteins in planta. Full-length virus vectors can spread systemically within a plant to colonize the majority of the phloem sink tissue, and are easily transmitted to new plants when scale-up is required. Therefore, this technology proved to be very useful, especially for moderate-to-large scale production of a variety of biopharmaceutical proteins (Hefferon 2012). Other important uses of plant virus vectors include the investigation and manipulation of metabolic pathways, monitoring virus trafficking and defining function of virus-encoded proteins (through expression of reporter proteins such as GFP or GUS), functional characterization of host disease resistance genes and pathogen effector proteins, and cellular protein localization studies.
For cereals , only two plant RNA virus vectors have been described, namely BSMV and Wheat streak mosaic virus (WSMV), to be capable of systemic expression of heterologous proteins (Tatineni et al. 2011; Lee et al. 2012) . The most widely used in wheat and barley is the BSMV vector system, initially developed for investigations of viral cell-to-cell and long distance movement (Haupt et al. 2001; Lawrence and Jackson 2001). The most commonly used BSMV vector variants are those designed to express recombinant proteins as fusions to the C-terminus of the small (17 kDa) viral γb protein. Using this type of vector, for example, Manning et al. (2010) achieved expression of a functionally active fungal (Pyrenophora tritici-repentis) ToxA effector protein in wheat. However, for many applications, it is advantageous to be able to produce a free heterologous protein without a viral γb protein, which may have a negative impact on protein activity, function or its intracellular localization. This can be achieved, for example, by introducing an autoproteolytic 2A peptide of picornaviruses between the fused sequences providing co-translational cleavage and release of free proteins. Using this approach, Lee et al. (2012) successfully expressed a number of relatively small proteins, including iLOV, a flavin-based fluorescent reporter protein and the necrosis-inducing secreted effector protein Nip1 from the fungal pathogen Rhynchosporium commune, in barley and wheat. In these experiments, Nip1 elicited systemic necrosis specifically in barley genotypes carrying the cognate Rrs1 resistance gene (Fig. 14.6). In another BSMV vector design, GFP and 42 rice coding gene sequences (CDS), between 200- and 1800-nt in size, were expressed as N-terminal fusions to the haemagglutinin epitope (HA) tag linked to the 2A self-cleaved peptide sequence followed by BSMV γb (Pogue and Holzberg 2012) . Western blot analyses of systemically infected barley tissue using anti-HA antibodies confirmed expression of 38 out of 43 recombinant protein products (Fig. 14.7). Reassuringly, 2A cleavage rates of 60–95 % were observed. Apparently, this vector was also capable of expressing proteins requiring maturation through the plant secretory pathway, which demonstrates the ability of the 2A cleavage system to deliver functionally active proteins to distinct subcellular fates. Moreover, the expression was shown to be relatively stable and at least in the case of GFP, its expression was observed regularly in leaves 1–4 above the inoculated leaf and maintained for up to 18 days post inoculation (Pogue and Holzberg 2012) .

BSMV::RcNip1 induces necrosis (arrows) specifically in barley ‘Atlas 46’ carrying the cognate resistance gene Rrs1 photographs taken at 8 days after virus inoculation. BSMV Barley stripe mosaic virus

BSMV-mediated expression of free GFP or heterologous proteins (reproduced with permission from Pogue and Holzberg 2012). Rice coding gene sequences (CDS) are arranged according to size. CDS whose predicted proteins were detected by Western blot analysis are unshaded, while blue shaded bars indicate the lack of detectable HA-tagged protein. Green bar represents the HA-tagged GFP control. BSMV Barley stripe mosaic virus, GFP green fluorescent protein
Concluding Remarks
The difficulty of obtaining stable transformants of most monocotyledonous species has contributed to the lag in gene function analyses in comparison to dicotyledonous species. However, as discussed in this chapter, there have been considerable advances in recent years in both the range and reliability of transient in planta expression systems available for both RNAi-mediated downregulation and overexpression of target genes of interest in monocots. In combination with the ongoing explosion of genomic and transcriptomic data being generated for many monocotyledonous species, major advances are foreseen in understanding gene function in these plants, and processes involved in interactions between monocots and associated organisms will continue to accelerate. In future, the existing technologies described in this chapter are likely to be improved further, thereby lowering costs whilst simultaneously increasing efficiency.
References
Alejska M, Figlerowicz M, Malinowska N, Urbanowicz A (2005) A universal BMV-based RNA recombination system—how to search for general rules in RNA recombination. Nucleic Acids Res 33:e105
Andrieu A, Breitler J, Siré C, Meynard D, Gantet P, Guiderdoni E (2012) An in-planta, Agrobacterium-mediated transient gene expression method for inducing gene silencing in rice (Oryza sativa L.) leaves. Rice 5:1–13
Azevedo C, Sadanandom A, Kitagawa K, Freialdenhoven A, Shirasu K, Schulze-Lefert P (2002) The RAR1 interactor SGT1, an essential component of R gene-triggered disease resistance. Science 295:2073–2076
Barrero-Sicilia C, Hernando-Amado S, Gonzalez-Melendi P, Carbonero P (2011) Structure, expression profile and subcellular localisation of four different sucrose synthase genes from barley. Planta 234:391–403
Bennypaul H, Mutti J, Rustgi S, Kumar N, Okubara P, Gill K (2012) Virus-induced gene silencing (VIGS) of genes expressed in root, leaf, and meiotic tissues of wheat. Funct Integr Genomics 12:143–156
Bhaskar PB, Venkateshwaran M, Wu L, Ané J-M, Jiang J (2009) Agrobacterium-mediated transient gene expression and silencing: a rapid tool for functional gene assay in potato. PLoS ONE 4:e5812
Block A, Li G, Fu AQ, Alfano JR (2008) Phytopathogen type III effector weaponry and their plant targets. Curr Opin Plant Biol 11:396–403
Bruun-Rasmussen M, Madsen CT, Jessing S, Albrechtsen M (2007) Stability of Barley stripe mosaic virus-induced gene silencing in barley. Mol Plant Microbe Interact 20:1323–1331
Cakir C, Scofield SR (2008) Evaluating the ability of the Barley stripe mosaic virus-induced gene silencing system to simultaneously silence two wheat genes. Cereal Res Commun 36:217–222
Cakir C, Tör M (2010) Factors influencing Barley stripe mosaic virus-mediated gene silencing in wheat. Physiol Mol Plant Pathol 74:246–253
Cao Y, Shi Y, Li Y, Cheng Y, Zhou T, Fan Z (2012) Possible involvement of maize Rop1 in the defence responses of plants to viral infection. Mol Plant Pathol 13:732–743
Chen X, Equi R, Baxter H, Berk K, Han J, Agarwal S, Zale J (2010) A high-throughput transient gene expression system for switchgrass (Panicum virgatum L.) seedlings. Biotechnol Biofuels 3:9
Christensen AB, Thordal-Christensen H, Zimmermann G, Gjetting T, Lyngkjær MF, Dudler R (2004) The germinlike protein GLP4 exhibits superoxide dismutase activity and is an important component of quantitative resistance in wheat and barley. Mol Plant Microbe Interact 17:109–117
Dai S, Wei X, Alfonso AA, Pei L, Duque UG, Zhang Z, Babb GM, Beachy RN (2008) Transgenic rice plants that overexpress transcription factors RF2a and RF2b are tolerant to Rice tungro virus replication and disease. Proc Natl Acad Sci USA 105:21012–21016
Demircan T, Akkaya MS (2010) Virus induced gene silencing in Brachypodium distachyon, a model organism for cereals. Plant Cell Tissue Organ Cult 100:91–96
Dhadi SR, Deshpande A, Ramakrishna W (2012) A novel non-wounding transient expression assay for cereals mediated by Agrobacterium tumefaciens. Plant Mol Biol Rep 30:36–45
Ding XS, Schneider WL, Chaluvadi SR, Mian MA, Nelson RS (2006) Characterization of a Brome mosaic virus strain and its use as a vector for gene silencing in monocotyledonous hosts. Mol Plant Microbe Interact 19:1229–1239
Ding XS, Ballard K, Nelson RS (2010) Improving virus induced gene silencing (VIGS) in rice through Agrobacterium infiltration. In: Antoun H, Avis T, Brisson L, Prevost D, Trepanier M (eds) Biology of molecular plant-microbe interactions. St. Paul, MN, USA International Society For Molecular Plant-Microbe Interactions, vol 7, Paper 59
Dong W, Nowara D, Schweizer P (2006) Protein polyubiquitination plays a role in basal host resistance of barley. Plant Cell 18:3321–3331
Douchkov D, Nowara D, Zierold U, Schweizer P (2005) A high-throughput gene-silencing system for the functional assessment of defense-related genes in barley epidermal cells. Mol Plant Microbe Interact 18:755–761
Dupuis I, Pace G (1993) Gene transfer to maize male reproductive structure by particle bombardment of tassel primordia. Plant Cell Rep 12:607–611
Fabro G, Steinbrenner J, Coates M, Ishaque N, Baxter L, Studholme DJ (2011) Multiple candidate effectors from the oomycete pathogen Hyaloperonospora arabidopsidis suppress host plant immunity. PLoS Pathog 7:e1002348
Fromm M, Taylor LP, Walbot V (1985) Expression of genes transferred into monocot and dicot plant cells by electroporation. Proc Natl Acad Sci U S A 82:5824–5828
Fursova O, Pogorelko G, Zabotina OA (2012) An efficient method for transient gene expression in monocots applied to modify the Brachypodium distachyon cell wall. Ann Bot 110:47–56
Goritschnig S, Krasileva KV, Dahlbeck D, Staskawicz BJ (2012) Computational prediction and molecular characterization of an oomycete effector and the cognate Arabidopsis resistance gene. PLoS Genet 8:e1002502
Haupt S, Duncan GH, Holzberg S, Oparka KJ (2001) Evidence for symplastic phloem unloading in sink leaves of barley. Plant Physiol 125:209–218
Hefferon KL (2012) Plant virus expression vectors set the stage as production platforms for biopharmaceutical proteins. Virology 433:1–6
Hein I, Barciszewska-Pacak M, Hrubikova K, Williamson S, Dinesen M, Soenderby IE (2005) Virus-induced gene silencing-based functional characterization of genes associated with powdery mildew resistance in barley. Plant Physiol 138:2155–2164
Held MA, Penning B, Brandt AS, Kessans SA, Yong W, Scofield SR, Carpita NC (2008) Small-interfering RNAs from natural antisense transcripts derived from a cellulose synthase gene modulate cell wall biosynthesis in barley. Proc Natl Acad Sci U S A 105:20534–20539
Hema M, Kao CC (2004) Template sequence near the initiation nucleotide can modulate brome mosaic virus RNA accumulation in plant protoplasts. J Virol 78:1169–1180
Hensgens LM, Bakker EHM, Os-Ruygrok E, Rueb S, Mark F, Maas H (1993) Transient and stable expression of gusA fusions with rice genes in rice, barley and perennial ryegrass. Plant Mol Biol 23:643–669
Holzberg S, Brosio P, Gross C, Pogue GP (2002) Barley stripe mosaic virus-induced gene silencing in a monocot plant. Plant J 30:315–327
Huang C, Qian Y, Li Z, Zhou X (2012) Virus-induced gene silencing and its application in plant functional genomics. Sci China Life Sci 55:99–108
Hückelhoven R, Panstruga R (2011) Cell biology of the plant-powdery mildew interaction. Curr Opin Plant Biol 14:738–746
Hwang Y-S, McCullar C, Huang N (2001) Evaluation of expression cassettes in developing rice endosperm using a transient expression assay. Plant Sci 161:1107–1116
Johansen LK, Carrington JC (2001) Silencing on the spot. Induction and suppression of RNA silencing in the Agrobacterium-mediated transient expression system. Plant Physiol 126:930–938
Kapila J, De Rycke R, Van Montagu M, Angenon G (1997) An Agrobacterium-mediated transient gene expression system for intact leaves. Plant Sci 122:101–108
Kawalek A, Dmochowska-Boguta M, Nadolska-Orczyk A, Orczyk W (2012) A new BSMV-based vector with modified β molecule allows simultaneous and stable silencing of two genes. Cell Mol Biol Lett 17:107–123
Klein TM, Roth BA, Fromm ME (1989) Regulation of anthocyanin biosynthetic genes introduced into intact maize tissues by microprojectiles. Proc Natl Acad Sci U S A 86:6681–6685
Klein TM, Wolf ED, Wu R, Sanford JC (1987) High-velocity microprojectiles for delivering nucleic acids into living cells. Nature 327:70–73
Lacomme C, Hrubikova K, Hein I (2003) Enhancement of virus-induced gene silencing through viral-based production of inverted-repeats. Plant J 34:543–553
Lawrence DM, Jackson AO (2001) Requirements for cell-to-cell movement of Barley stripe mosaic virus in monocot and dicot hosts. Mol Plant Pathol 2:65–75
Leduc N, Iglesias V, Bilang R, Gisel A, Potrykus I, Sautter C (1994) Gene transfer to inflorescence and flower meristems using ballistic micro-targeting. Sex Plant Reprod 7:135–143
Lee W-S, Hammond-Kosack KE, Kanyuka K (2012) Barley stripe mosaic virus-mediated tools for investigating gene function in cereal plants and their pathogens: Virus-induced gene silencing, host-mediated gene silencing, and vrus-mediated overexpression of heterologous proteins. Plant Physiol 160:582–590
Lee W-S, Rudd JJ, Hammond-Kosack KE, Kanyuka K (2014) Mycosphaerella graminicola LysM effector-mediated stealth pathogenesis subverts recognition through both CERK1 and CEBiP homologues in wheat. Mol Plant Microbe Interact 27:236–243
Li H, Fan R, Li L, Wei BO, Li G, Gu L, Wang X, Zhang X (2014) Identification and characterization of a novel copper transporter gene family TaCT1 in common wheat. Plant Cell Environ. doi:10.1111/pce.12263
Liang J, Deng G, Long H, Pan Z, Wang C, Cai P, Xu D, Nima Z-X, Yu M (2012) Virus-induced silencing of genes encoding LEA protein in Tibetan hulless barley (Hordeum vulgare ssp. vulgare) and their relationship to drought tolerance. Mol Breeding 30:441–451
Lin N-S, Hsu Y-H (1994) A satellite RNA associated with bamboo mosaic potexvirus. Virology 202:707–714
Lin N-S, Lin B-Y, Lo N-W, Hu C-C, Chow T-Y, Hsu Y-H (1994) Nucleotide sequence of the genomic RNA of bamboo mosaic potexvirus. J Gen Virol 75:2513–2518
Liou M-R, Huang Y-W, Hu C-C, Lin N-S, Hsu Y-H (2013) A dual gene-silencing vector system for monocot and dicot plants. Plant Biotechnol J. doi:10.1111/pbi.12140
Lu HC, Chen HH, Tsai WC, Chen WH, Su HJ, Chang DC, Yeh HH (2007) Strategies for functional validation of genes involved in reproductive stages of orchids. Plant Physiol 143:558–569
Ma H-Z, Liu G-Q, Li C-W, Kang G-Z, Guo T-C (2012a) Identification of the TaBTF3 gene in wheat (Triticum aestivum L.) and the effect of its silencing on wheat chloroplast, mitochondria and mesophyll cell development. Biochem Biophys Res Commun 426:608–614
Ma M, Yan Y, Huang L, Chen M, Zhao H (2012b) Virus-induced gene-silencing in wheat spikes and grains and its application in functional analysis of HMW-GS-encoding genes. BMC Plant Biol 12:141
Manning VA, Chu AL, Scofield SR, Ciuffetti LM (2010) Intracellular expression of a host-selective toxin, ToxA, in diverse plants phenocopies silencing of a ToxA-interacting protein, ToxABP1. New Phytol 187:1034–1047
Martin T, Biruma M, Fridborg I, Okori P, Dixelius C (2011) A highly conserved NB-LRR encoding gene cluster effective against Setosphaeria turcica in sorghum. BMC Plant Biol 11:151
Martin R, Glover-Cutter K, Martin R, Dombrowski J (2013) Virus induced gene silencing in Lolium temulentum. Plant Cell Tissue Organ Cult 113:163–171
Marzin S, Mihaly R, Pauk J, Schweizer P (2008) A transient assay system for the assessment of cell-autonomous gene function in dehydration-stressed barley. J Exp Bot 59:3359–3369
Meng Y, Moscou MJ, Wise RP (2009) Blufensin1 negatively impacts basal defense in response to barley powdery mildew. Plant Physiol 149:271–285
Miki D, Shimamoto K (2004) Simple RNAi vectors for stable and transient suppression of gene function in rice. Plant Cell Physiol 45:490–495
Nelson A, Bushnell W (1997) Transient expression of anthocyanin genes in barley epidermal cells: potential for use in evaluation of disease response genes. Transgenic Res 6:233–244
Oikawa A, Rahman A, Yamashita T, Taira H, Kidou S (2007) Virus-induced gene silencing of P23k in barley leaf reveals morphological changes involved in secondary wall formation. J Exp Bot 58:2617–2625
Pacak A, Geisler K, Jorgensen B, Barciszewska-Pacak M, Nilsson L, Nielsen TH (2010a) Investigations of barley stripe mosaic virus as a gene silencing vector in barley roots and in Brachypodium distachyon and oat. Plant Methods 6:26
Pacak A, Strozycki P, Barciszewska-Pacak M, Alejska M, Lacomme C, Jarmołowski A (2010b) The Brome mosaic virus-based recombination vector triggers a limited gene silencing response depending on the orientation of the inserted sequence. Arch Virol 155:169–179
Panstruga R (2004) A golden shot: how ballistic single cell transformation boosts the molecular analysis of cereal-mildew interactions. Mol Plant Pathol 5:141–148
Petty ITD, Hunter BG, Wei N, Jackson AO (1989) Infectious Barley stripe mosaic virus RNA transcribed in vitro from full-length genomic cDNA clones. Virology 171:342–349
Pogue GP, Holzberg S (2012) Transient virus expression systems for recombinant protein expression in dicot- and monocotyledonous plants. In: Dhal NK, Sahu SC (eds) Plant science. InTech, Croatia, pp 191–216. doi:10.5772/54187
Purkayastha A, Mathur S, Verma V, Sharma S, Dasgupta I (2010) Virus-induced gene silencing in rice using a vector derived from a DNA virus. Planta 232:1531–1540
Ramanna H, Ding X, Nelson R (2013) Rationale for developing new virus vectors to analyse gene function in grasses through virus-induced gene silencing. In: Becker A (ed) Virus-induced gene silencing. Humana, New York, (pp 15–32). (Methods Mol Biol 975. doi:10.1007/978-1-62703-278-0_2)
Renner T, Bragg J, Driscoll HE, Cho J, Jackson AO, Specht CD (2009) Virus-induced gene silencing in the culinary ginger (Zingiber officinale): an effective mechanism for down-regulating gene expression in tropical monocots. Mol Plant 2:1084–1094
Rentel MC, Leonelli L, Dahlbeck D, Zhao B, Staskawicz BJ (2008) Recognition of the Hyaloperonospora parasitica effector ATR13 triggers resistance against oomycete, bacterial, and viral pathogens. Proc Natl Acad Sci U S A 105:1091–1096
Requesens DV, Egelkrout E, Devaiah S, Hood EE (2010) A method for transient expression in maize endosperm. In Vitro Cell Dev Biol Plant 46:485–490
Rong W, Qi L, Wang A, Ye X, Du L, Liang H, Xin Z, Zhang Z (2014) The ERF transcription factor TaERF3 promotes tolerance to salt and drought stresses in wheat. Plant Biotechnol J. doi:10.1111/pbi.12153
Roossinck MJ, Sleat D, Palukaitis P (1992) Satellite RNAs of plant viruses: structures and biological effects. Microbiol Rev 56:265–279
Schweizer P, Pokorny J, Abderhalden O, Dudler R (1999) A transient assay system for the functional assessment of defense-related genes in wheat. Mol Plant Microbe Interact 12:647–654
Schweizer P, Pokorny J, Schulze-Lefert P, Dudler R (2000) Double-stranded RNA interferes with gene function at the single-cell level in cereals. Plant J 24:895–903
Scofield SR, Huang L, Brandt AS, Gill BS (2005) Development of a virus-induced gene-silencing system for hexaploid wheat and its use in functional analysis of the Lr21-mediated leaf rust resistance pathway. Plant Physiol 138:2165–2173
Sharma S, Hirabuchi A, Yoshida K, Fujisaki K, Ito A, Uemura A (2013) Deployment of the Burkholderia glumae type III secretion system as an efficient tool for translocating pathogen effectors to monocot cells. Plant J 74:701–712
Shi Y, Qin Y, Cao Y, Sun H, Zhou T, Hong Y, Fan Z (2011) Influence of an m-type thioredoxin in maize on potyviral infection. Eur J Plant Pathol 131:317–326
Sohn KH, Lei R, Nemri A, Jones JDG (2007) The downy mildew effector proteins ATR1 and ATR13 promote disease susceptibility in Arabidopsis thaliana. Plant Cell 19:4077–4090
Sparks CA, Jones HD (2014) Genetic transformation of wheat via particle bombardment. In: Henry R, Furtado A (eds) Cereal genomics: methods and protocols. Humana, New York, (pp 201–218). (Methods Mol Biol 1099. doi:10.1007/978-1-62703-715-0_17)
Sparks CA, Doherty A, Jones HD (2014) Genetic transformation of wheat via Agrobacterium-mediated DNA delivery. In: Henry R, Furtado A (eds) Cereal genomics: methods and protocols. Humana, New York, (pp 235–250) (Methods Mol Biol 1099. doi:10.1007/978-1-62703-715-0_17)
Sun L, Zhang H, Li D, Huang L, Hong Y, Ding XS (2013) Functions of rice NAC transcriptional factors, ONAC122 and ONAC131, in defense responses against Magnaporthe grisea. Plant Mol Biol 81:41–56
Tai Y-S, Bragg J, Edwards MC (2005) Virus vector for gene silencing in wheat. Biotechniques 39:310–312
Tatineni S, McMechan AJ, Hein GL, French R (2011) Efficient and stable expression of GFP through Wheat streak mosaic virus-based vectors in cereal hosts using a range of cleavage sites: Formation of dense fluorescent aggregates for sensitive virus tracking. Virology 410:268–281
Thomas WJ, Thireault CA, Kimbrel JA, Chang JH (2009) Recombineering and stable integration of the Pseudomonas syringae pv. syringae 61 hrp/hrc cluster into the genome of the soil bacterium Pseudomonas fluorescens Pf0-1. Plant J 60:919–928
Tufan HA, Stefanato FL, McGrann GR, MacCormack R, Boyd LA (2011) The Barley stripe mosaic virus system used for virus-induced gene silencing in cereals differentially affects susceptibility to fungal pathogens in wheat. J Plant Physiol 168:990–994
Upadhyaya NM, Mago R, Staskawicz BJ, Ayliffe MA, Ellis JG, Dodds PN (2014) A bacterial type III secretion assay for delivery of fungal effector proteins into wheat. Mol Plant Microbe Interact 27:255–264
Vaghchhipawala Z, Rojas CM, Senthil-Kumar M, Mysore KS (2011) Agroinoculation and agroinfiltration: simple tools for complex gene function analyses. In: Pereira A (ed) Plant reverse genetics: methods and protocols. Humana, New York, (pp 65–76) (Methods Mol Biol 678. doi: 10.1007/978-1-60761-682-5_6)
van Eck L, Schultz T, Leach JE, Scofield SR, Peairs FB, Botha AM (2010) Virus-induced gene silencing of WRKY53 and an inducible phenylalanine ammonia-lyase in wheat reduces aphid resistance. Plant Biotechnol J 8:1023–1032
Vandergheynst JS, Guo HY, Simmons CW (2008) Response surface studies that elucidate the role of infiltration conditions on Agrobacterium tumefaciens-mediated transient transgene expression in harvested switchgrass (Panicum virgatum). Biomass Bioenerg 32:372–379
Wang X, Cao A, Yu C, Wang D, Chen P (2010) Establishment of an effective virus induced gene silencing system with BSMV in Haynaldia villosa. Mol Biol Rep 37:967–972
Wang G-F, Wei X, Fan R, Zhou H, Wang X, Yu C (2011) Molecular analysis of common wheat genes encoding three types of cytosolic heat shock protein 90 (Hsp90): functional involvement of cytosolic Hsp90s in the control of wheat seedling growth and disease resistance. New Phytol 191:418–431
Waterhouse PM, Wang MB, Lough T (2001) Gene silencing as an adaptive defence against viruses. Nature 411:834–842
Weis C, Hückelhoven R, Eichmann R (2013) LIFEGUARD proteins support plant colonization by biotrophic powdery mildew fungi. J Exp Bot 64:3855–3867
Whisson SC, Boevink PC, Moleleki L, Avrova AO, Morales JG, Gilroy EM (2007) A translocation signal for delivery of oomycete effector proteins into host plant cells. Nature 450:115–118
Wroblewski T, Tomczak A, Michelmore R (2005) Optimization of Agrobacterium-mediated transient assays of gene expression in lettuce, tomato and Arabidopsis. Plant Biotechnol J 3:259–273
Yang CC, Liu JS, Lin CP, Lin NS (1997) Nucleotide sequence and phylogenetic analysis of a bamboo mosaic potexvirus isolate from common bamboo (Bambusa vulgaris McClure). Bot Bull Acad Sin 38:77–84
Yin C, Hulbert S (2011) Prospects for functional analysis of effectors from cereal rust fungi. Euphytica 179:57–67
Yuan C, Li C, Yan L, Jackson AO, Liu Z, Han C, Yu J, Li D (2011) A high throughput Barley stripe mosaic virus vector for virus induced gene silencing in monocots and dicots. PLoS ONE 6:e26468
Zimmermann G, Bäumlein H, Mock H-P, Himmelbach A, Schweizer P (2006) The multigene family encoding germin-like proteins of barley. Regulation and function in basal host resistance. Plant Physiol 142:181–192
Acknowledgments
All the authors are supported by the Biotechnology and Biological Sciences Research Council of the UK (BBSRC) through the Institute Strategic Program 20:20® Wheat.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2015 Springer Science+Business Media, LLC
About this chapter
Cite this chapter
Lee, WS., Hammond-Kosack, K., Kanyuka, K. (2015). In Planta Transient Expression Systems for Monocots. In: Azhakanandam, K., Silverstone, A., Daniell, H., Davey, M. (eds) Recent Advancements in Gene Expression and Enabling Technologies in Crop Plants. Springer, New York, NY. https://doi.org/10.1007/978-1-4939-2202-4_14
Download citation
DOI: https://doi.org/10.1007/978-1-4939-2202-4_14
Published:
Publisher Name: Springer, New York, NY
Print ISBN: 978-1-4939-2201-7
Online ISBN: 978-1-4939-2202-4
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)