
Abstract
It is well established that the highly nonlinear and anisotropic mechanical behaviors of soft tissues are an emergent behavior of the underlying tissue microstructure. Numerical solutions form the cornerstone in the application of constitutive models in contemporary biomechanics. Herein, a structural constitutive model into a finite element framework specialized for membrane tissues. Multiple deformation modes were simulated, including strip biaxial, planar biaxial with two attachment methods, and membrane inflation. Detailed comparisons with experimental data were undertaken to insure faithful simulations of both the macro-level stress–strain insights into adaptations of the fiber architecture under stress, such as fiber reorientation and fiber recruitment. Results indicated a high degree of fidelity and demonstrated interesting microstructural adaptions to stress and the important role of the underlying tissue matrix.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
- Uniaxial Tension
- Strain Energy Function
- Fiber Recruitment
- Fiber Orientation Distribution
- Stretch Direction
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
17.1 Introduction
Traditionally, soft tissues are modeled as pseudo-hyperelastic materials using either phenomenological or structural approaches (Criscione etal. 2003; Holzapfel and Ogden 2009; Sacks 2000). A common phenomenological model is the Fung-type (Fung 1993; Tong and Fung 1976), in which the strain energy function is a quadratic exponential function of the Green-Lagrange strain tensor. The original form was based on the observed linear relation between tissue stiffness and stress under uniaxial conditions (Fung 1993). However, phenomenological models lack physical interpretation and cannot, in general, be used for simulations beyond the strain range utilized in parameter estimation. This effect has been shown to be the case even when the strain magnitudes did not exceed the maximum values measured but where substantially far from the available experimental data (Sun etal. 2003). While the underlying reasons for this still need to be elucidated, models which possess greater links to the underlying physical mechanisms appear to be the next step.
Like any biological or synthetic biomaterial, the complex mechanical behavior of soft tissues results from the deformations and interactions of the constituent phases. For most soft tissues, these include collagen, elastin, muscular, and related matrix components such as glycosaminoglycans and proteoglycans. The idea of accounting for tissue structure into mechanical models of soft tissues goes back to at least the work on leather mechanics in 1945 by Mitton. More contemporary work on structural approaches followed, with growing popularity in the 1970s (Beskos and Jenkiins 1975), with the concept of stochastic constituent fiber recruitment developed about the same time (Soong and Huang 1973) based on related structural studies (Kenedi etal. 1965). In part a result of the availability of the first planar biaxial data for soft tissues, Lanir developed the first comprehensive, multidimensional structural constitutive model formulation (Lanir 1979). With various modifications, Lanir etal. applied this approach to many soft tissues such as lung (Lanir 1983) and myocardium (Horowitz etal. 1988).
By linking tissue deformation at macroscopic scale and microscopic (fiber) scale through affine deformation assumption, the structural constitutive model can be considered a statistical multi-scale approach. Above all, structural constitutive modeling approaches can, in principle, provide valuable insight into tissue function. For example, Billiar and Sacks (2000a, b) demonstrated for aortic valve leaflets that, using a simplified leaflet structure, angular rotation of the fibers account for such important features such as pronounced mechanical anisotropy, axial coupling, and very large strains (>80 %) even though the tissue is composed of collagen fibers that fail at less than ∼12 % strain. Later, Sacks demonstrated that with the use of only an equi-biaxial test and the experimentally measured fiber orientation distribution, the complete in-plane biaxial response could be simulated (Sacks 2003). More recently, structural approaches have been used for a wide range of native and engineered tissue applications, such as elastomeric tissue engineering scaffolds (Courtney etal. 2006), urinary bladder wall (Wognum etal. 2009), and many others (Fata etal. 2014; Hansen etal. 2009; Hollander etal. 2011; Kao etal. 2011).
Due to the need to solve soft tissue problems that involve complex anatomical geometries and boundary conditions, many constitutive models for soft tissues in various forms have been implemented into a computational framework (Driessen etal. 2007; Holzapfel etal. 1996; Prot etal. 2007; Sun and Sacks 2005; Hariton etal. 2007). Yet, robust evaluation and rigorous validation of structural constitutive models remain quite limited. Moreover, studies on structural model have mainly focused on either material parameter estimation (Jor etal. 2011) or comparison with different constitutive models (Bischoff 2006; Cortes etal. 2010; Tonge etal. 2013). Structural models that incorporate fiber recruitment have rarely been used, largely due to computational demands of the additional integration. Recent ability to get detailed fiber recruitment data (e.g., Chen etal. 2011; Fata etal. 2014; Hansen etal. 2009) makes this approach all the more relevant. The deep insights that structural models can provide, such as the role of fiber structure and kinematics, still have yet to be fully explored by simulation.
Since many soft tissues are relatively thin, they can be modeled using shell or membrane elements in FE analysis, greatly speeding up the simulations. In the present study, we implemented a planar structural constitutive model into the commercial finite element (FE) package ABAQUS. By numerical simulation of one single element subjected to uniaxial tension, we first revealed that matrix must be present to prevent unrealistic tissue deformations. Flexural simulations were utilized to estimate the matrix modulus, since the underlying collagen fibers remain undulated due to the small extensional strains, and thus have little effect on tissue stress development. Strip biaxial strain and equi-biaxial tension simulations were also performed and compared with experimental collagen fiber measurements to demonstrate the effects of initial fiber orientation distribution on fiber reorientation. Simulation of membrane inflation tests were also applied to further test the structural model. In addition to prediction of macroscopic mechanical response of soft tissues, we demonstrate how the structural model can provide insights into tissue micro-structural events.
17.2 Methods
17.2.1 Theoretical Formulation
Soft biological tissues primarily have two major load-bearing components: the fibrous network and the nonfibrous (i.e., amorphous) ground matrix. Based on Fung (1993), we idealize the elastic behavior of soft tissues as pseudo-hyperelastic composite materials. Thus, the total strain energy function Ψ of soft tissue at a represent volume element (RVE) is defined using
where Ψ f and Ψ m are the strain energy functions for the fiber and matrix phases, respectively, ϕ if and ϕ m are the volume fraction of fiber and matrix, respectively, with \( {\displaystyle \sum_{i=1}^n{\phi}_{\mathrm{f}}^i}+{\phi}_{\mathrm{m}}=1 \), \( J= \det \left(\tilde{\mathbf{F}}\right) \), and p the Lagrange multiplier to enforce incompressibility due to soft tissue’s high water content. The contributions of the nonfibrous components and fluid phases are assumed to be responsible for the incompressibility of the tissue. Based on previous results (Buchanan and Sacks 2013), we model the matrix phase (which compromises all nonfibrous components) as a single isotropic hyperelastic Neo-Hookean material with the strain energy function \( {\varPsi}_{\mathrm{m}}={C}_1\left({I}_{\mathrm{C}}-3\right) \). Here, I C is the first invariant of the right Cauchy-Green tensor \( \tilde{\mathbf{C}}={\tilde{\mathbf{F}}}^{\mathrm{T}}\tilde{\mathbf{F}} \), and to be consistent with linear elasticity \( {C}_1=\mu /2 \), where μ is the shear modulus. Using \( \mathbf{S}=\partial \varPsi /\partial \mathbf{E} \), the resulting second-Piola Kirchhoff stress is given by
Next, without loss of generality, we focus on a single, undulated Type I collagen fiber type with planar structures and a plane stress state. As in related work on collagenous tissues (Lanir 1983; Sacks 2003), we assume a linear \( {S}_{\mathrm{f}}=\eta {E}_{\mathrm{f}} \) relationship for the individual collagen fibers
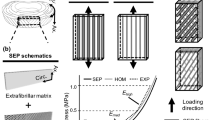
where η is the elastic modulus of individual straight collagen fibers (Fig. 17.1a). Due to their crimped structure, we express individual fiber’s true fiber strain using \( {E}_{\mathrm{t}}=\frac{E_{\mathrm{f}}-{E}_{\mathrm{s}}}{1+2{E}_{\mathrm{s}}} \) where E s is the fiber slack strain (Fig. 17.1a). The resulting individual collagen fiber strain energy is thus

(a) Assumed stress–strain response of a single undulated fiber. Graphical depiction of a (b) fiber ensemble
To develop the first level homogenization, we define a fiber ensemble as the collection of all fibers within the RVE with a common direction \( \widehat{\mathbf{N}}=\left[\begin{array}{ccc} \cos \left(\theta \right) & \sin \left(\theta \right) & 0 \end{array}\right] \) (Fig. 17.1b). The collective mechanical contribution from the ensemble is represented by its strain energy Ψ ens. Assuming affine deformation (Lanir 1983; Sacks 2003), the fiber ensemble strain E ens in the direction \( \widehat{\mathbf{N}} \) is related to the macroscopic tissue-level Green-Lagrange strain tensor \( \tilde{\mathbf{E}}=\frac{1}{2}\left(\tilde{\mathbf{C}}-\tilde{\mathbf{I}}\right) \) by
We make the distinction between fiber and ensemble strains here since individual collagen fibers will have a different strain levels due to their undulations. Note that the nonlinearity of the tissue evolves from the gradual recruitment of the linearly elastic collagen fibers (Lanir 1983), and is thus a structural as opposed to a material property.
To stochastically account for the gradual recruitment of the collagen fiber in each fiber ensemble with strain, we define the function D(E s) over the ensemble strain range \( {E}_{\mathrm{ens}}\in \left[{E}_{\mathrm{lb}},{E}_{\mathrm{ub}}\right] \). Here, E lb and E ub represent the lower and upper bounds of collagen fiber ensemble recruitment strain levels, with E ub > E lb > 0 and \( {\displaystyle {\int}_{E_{\mathrm{lb}}}^{E_{\mathrm{ub}}}D(x)dx=1} \). The ensuing fiber ensemble strain energy and stress–strain relation are then described as the sum of individual fiber strain energies of the ensemble weighted by the distribution of slack strains D, so that
D was represented as a Beta distribution B defined over \( {E}_{\mathrm{s}}\in \left[{E}_{\mathrm{lb}},{E}_{\mathrm{ub}}\right] \),
where α and β are the shape factors. Note that for simplicity we chose \( {E}_{\mathrm{lb}} = 0 \), although generally it is not (Fata etal. 2014).
In situations where computational demands are very high, we also present an alternative formulation for the ensemble stress–strain relation using simplified exponential form that emulates the recruitment behavior at both low and high strains. The novel aspect here is that the terminal stiffness of the fiber ensemble is reproduced for ensemble strains above E ub. This simple but important modification helps avoid unrealistically high fiber stresses at high strains when using an exponential model alone. For an exponential model, this becomes
where A and B are material constants. Note that the tangent modulus is continuous at \( E={E}_{\mathrm{ub}} \).
In the final step, we homogenize the ensemble response to the tissue level by defining the tissue strain energy as the sum of the strain energy of fiber ensembles, weighted by the orientation distribution function (ODF) Γ(θ). Thus, we have
with the normalization constraint \( {\displaystyle {\int}_{-\pi /2}^{\pi /2}\varGamma \left(\theta \right)d\theta }=1 \). In summary, the total strain energy function of soft tissue in the RVE is expressed as
For the plane stress case, the out-of-plane stress component \( {S}_{33}=2\partial \varPsi /\partial {C}_{33}=0 \), so that the Lagrange multiplier p can be determined directly from
The total second Piola-Kirchhoff stress can be thus written as
for the recruitment model, and
for the simplified model (17.8).
17.2.2 Finite Element Implementation
The structural model was implemented into commercial finite element software ABAQUS/Standard (Dassault Systèmes Simulia Corp., Providence, RI) via user-defined material subroutine UMAT. The stress tensor components utilized in UMAT is defined in a co-rotational coordinate system in which the local material axes defined in the initial configuration rotates with the material (ABAQUS 2011). Using polar decomposition theorem (Marsden and Hughes 1983) we have \( \tilde{\mathbf{F}}=\tilde{\mathbf{R}}\tilde{\mathbf{U}} \), where \( \tilde{\mathbf{R}} \) is the rigid body rotation tensor and Ũ is the right symmetric stretch tensor. Also, the rotated Cauchy stress can be determined using \( \tilde{\mathbf{t}}={\mathrm{J}}^{-1}\tilde{\mathbf{U}}\tilde{\mathbf{S}}\tilde{\mathbf{U}} \) and the fourth-rank material elasticity tensor ℂSE are updated in the UMAT code. In the actual implementation, \( \tilde{\mathbf{S}} \) and ℂSE require integration over \( \theta \in \left[-\pi /2,\pi /2\right] \) as well as \( {E}_{\mathrm{ens}}\in \left[{E}_{\mathrm{lb}},{E}_{\mathrm{ub}}\right] \). Since a closed form solutions are not available in general, a numerical integration scheme was used as follows. During implementation, the angle domain and the fiber strain domain were separated into twenty segments with equal size. In each segment, Gaussian quadrature integration rule (Hughes 2000) was performed with five integration points.
17.2.3 Further Model Modifications and Material ParameterEstimation
For the present work we merged (without loss of generality) the material parameters ϕ m and μ m for matrix component into μ m, as well as ϕ f and η for the fiber component were also combined into η. Due to its high collagen Type I content, generally planar tissue architecture, well-characterized structure and mechanical properties, and previous use in structural models (Sacks 2003) made native bovine pericardium natural choice for the representative tissue for simulations. To obtain the value for μ m, we utilized flexural data from native bovine pericardium (Mirnajafi etal. 2005). In that study, a nearly linear moment-curvature relation has been observed. This suggested that the collagen fibers have little effect in flexure, which is consistent with the very low strains that occur in this deformation mode (so that the collagen fibers remain fully undulated and only the matrix contributes). We thus obtained μ m by fitting the moment-curvature curve (Mirnajafi etal. 2005), using methods described in the next section.
The total fiber angular distribution function is expressed as a linear combination of Gaussian distribution and uniform distribution
Equation (17.14) was chosen to allow graduations in aligned and isotropic fiber distributions to be simulated easily. Here σ denotes the standard deviation of the Gaussian distribution function, and the error function erf() is introduced so that the integration of the Gaussian distribution function over angle domain \( \theta \mathit{\in}\left[-\pi /2,\pi /2\right] \) is equal to unity. The fiber angular distribution function was obtained previous measurements and fitting the experimental data with d = 1 (Billiar and Sacks 1997).
One way to evaluate the robustness and accuracy of the FE implementation is to examine applications where very large strains are known to occur, which induce large fiber rotations and stretches. Previous experimental results have revealed that the mechanical behavior of soft collagenous tissue is strongly dependent on gripping methods (Waldman and Michael Lee 2002). In particular, we noted in that study that clamps induced large rotations in the corner regions between the clamps. Thus, the material parameters from both model forms were obtained by fitting stress–strain curve from the equi-biaxial loading stress–strain data with suturing arrangement from Waldman and Michael Lee (2002). This allowed us to directly compare the FE results to the experimental findings from that study.
17.2.4 Finite Element Simulations
We start with a basic simulation of a single element under uniaxial tension to investigate the effects of matrix. For this example, a square element was subjected to uniaxial strain in X 1 direction (Fig. 17.2a). Nodes 1 and 2 were constrained in X 2 direction, and nodes 1 and 4 were constrained in X 1 direction, with uniform displacements applied to nodes 2 and 3 in the X 1 direction. The preferred fiber orientation coincided with the X 1 direction. Next, to verify minimal fiber recruitment occurred during flexure, simulation of a bending test was performed. The “The length and width of the specimen…” length and width of the specimen used for bending simulation is 20.0 and 3.0 mm, respectively, with the thickness of the tissue as 0.4 mm and the span 16.0 mm (Fig. 17.2b). The loading was applied at the center of the tissue through the middle post, with the three posts being considered as rigid bodies. The friction coefficient was assumed to be zero for the tissue in contact with the left and right posts.

Effective deformations of fibers on soft tissues under three-point bending showing (a) the moment-curvature curve relation obtained using a Neo-Hookean model with matrix only and structural model with fiber and matrix, (b) Green Strain E 11 contour, (c) Fractional ensemble fiber recruitment contour in X 1 direction
To investigate the effects of both boundary conditions (as both localized point and distributed loads) on fiber reorientation, we simulated native bovine pericardium using sutures under strip biaxial tension using data from Billiar and Sacks (1997) and equi-biaxial tension using clamped boundary conditions using data from Waldman etal. (2002). To simulate both high and low orientations, we utilized two levels of d (d = 1.0 and d = 0.25). For the first test, the dimensions of the specimen were 19.2 mm × 19.2 mm and the thickness was 0.4 mm. As in the original experiment, uniform displacements were applied on the seven suture attachment points along each side the specimen, with the initial preferred fiber orientation set to 27° from the X 1 axis (Fig. 17.3). Two loading cases were considered; 30 % along X 1 direction/0 % for the X 2, and 30 % along the X 2 direction/0 % along the X 1. For the clamped equi-biaxial tension test, the dimensions of the specimen were 22.0 mm × 22.0 mm (Fig. 17.4) and the thickness 0.4 mm. The tissue was stretched 10 % in X 1 and X 2 directions. The initial fiber orientation was assumed to be the X 1 direction (Fig. 17.3d).

Preferred fiber reorientation and standard deviation contour of fiber angular distribution function from simulation under strip biaxial stretch (a) in X 1 direction with d = 1.0, (b) in X 1 direction with d = 0.25, (c) in X 2 direction with d = 1.0, (d) in X 2 direction with d = 0.25. Inset—measured SALS data from the Billiar and Sacks 1997 study, showing very good agreement

(a) Azimuthal plot of the fractional ensemble fiber recruitment under different stretch ratios, (b) total percentage of fiber recruitment contour after stretch in X 1 direction, (c) total percentage of fiber recruitment along the line X 1 and X 2 in (b)
As a final test, we simulated a fetal membrane (FM) inflation test using data from Joyce etal. (2009). Uniform pressure was applied on the top surface of a circular membrane and only half of the tissue was modeled due to symmetry. The radius of the circular membrane was 21.0 mm and the thickness 0.228 mm. The tube was modeled as rigid body with an inner radius of 15.0 mm and the edge of the tissue fixed. SALS measurements of the intact FM (Joyce etal. 2009) revealed that the tissue contains no preferred collagen direction, therefore a uniform fiber angular distribution function \( \varGamma \left(\theta \right)=1/\pi \) was utilized. The friction coefficient was assumed to be zero for the contact interaction of tissue with the rigid tube. Note that for flexural simulations, four-node quadrilateral shell elements were used, and for all the other simulations four-node quadrilateral membrane elements were used.
17.2.5 Simulation Post-processing
To provide insights into the deformations of soft tissue microstructure under strain, we implemented the following post-processing procedures. The following two-dimensional form
where J 2D is the determinate of the in-plane deformation gradient. Note that Γt(β) should be plotted against the deformed fiber angle \( \beta ={ \tan}^{-1}\left(\frac{F_{21} \cos \left(\theta \right)+{F}_{22} \sin \left(\theta \right)}{F_{11} \cos \left(\theta \right)+{F}_{12} \sin \left(\theta \right)}\right) \) since it refers to the deformed (convected) fiber direction. In addition to its ability to provide high fidelity simulations of the stress–strain behaviors, the structural model also provides a great deal of information structural adaptation to within the RVE. These are usually overlooked in the literature. Thus, in the present study we defined the fractional ensemble fiber recruitment (FEFR) structural metric for a given direction as
In addition, the total fiber recruitment (TFR) structural metric for a given RVE was defined as
Note that both metrics are expressed using a percentile scale.
17.3 Results
17.3.1 Uniaxial Tension Simulation
Overall, we determined that the matrix had a signification effect on the simulated deformation of soft tissue under uniaxial tension. When the tissue is modeled with fibers only, the lateral deformation of the tissue may be unrealistic. Specifically, since under uniaxial tension the stress component in the X 2 direction is \( {S}_{22}={\displaystyle {\int}_{-\pi /2}^{\pi /2}\varGamma \left(\theta \right){S}_{\mathrm{ens}}\left({E}_{\mathrm{ens}}\right){ \sin}^2\left(\theta \right)d\theta } \), with \( {S}_{\mathrm{ens}}=D(x)=0 \) in this direction since the fibers cannot carry any load when compressed. Since \( \varGamma \left(\theta \right)\ge 0 \) and \( { \sin}^2\left(\theta \right)\ge 0 \), S 22 must be greater than zero, yet under uniaxial tension \( {S}_{22}=0 \), so that an equilibrium state cannot be achieved. For example, when the element is stretched 6 % (Fig. 17.3a) the Green-Lagrange strain E 11 = 0.618 and E 22 = −0.0434 with a ratio of –E 22/E 11 of 0.702. When the element is stretched 12 % (Fig. 17.3b), the strain ratio –E 22/E 11 increased to 2.049. The deformation in X 2 direction is larger than that in X 1 direction, and the element collapses to a single line when the stretch ratio is greater than 15 %. However, when the matrix is included, the strain ratio –E 22/E 11 is 0.459 and 0.424 as the element is stretched 6 % and 12 %, respectively, so that the deformation of the element is acceptable. Thus for the present model a matrix should be present and in sufficient quantity in the structural model to prevent unphysical characterization of the mechanical behavior of soft tissues under uniaxial tension.
17.3.2 Flexural Simulations
The moment-curvature curves from FE simulation using only the isotropic neo-Hookean model were in good agreement with the published experimental results (Mirnajafi etal. 2005) (Fig. 17.2a). The simulation results also confirmed that collagen fiber contributions were negligible; the moment-curvature curves from FE simulations are the same using Neo-Hookean model and the structural model with both fiber and matrix (Fig. 17.2). The maximum tensile strain (Green-Lagrange strain) under bending was 0.0346 located at the bottom surface. Virtually all (>99 %) of the fibers were still undulated in this loading configuration (Fig. 17.2), supporting our use of these studies to determine ground matrix mechanical behavior.
17.3.3 Biaxial Test Simulations: Sutured Boundary Conditions
For the strip biaxial test stretched in the X 1 direction, fiber orientation from SALS measurements (Billiar and Sacks 1997) revealed that the overall preferred fiber direction was reoriented towards the direction of stretch and the degree of fiber alignment was increased. When d = 1.0 in the fiber angular density function, the preferred fiber direction rotated only about 4° towards the stretch direction (Fig. 17.3a). However, for d = 0.25, the preferred fiber direction rotated about 15° towards the stretch direction (Fig. 17.3b), in agreement with SALS measurements (Billiar and Sacks 1997). Around the suture points, the simulation results with d = 0.25 (Fig. 17.3b) demonstrated that the preferred fiber direction rotated towards the suture points, also in agreement with the SALS data (Billiar and Sacks 1997). When the tissue was stretched in the X 2 direction with d = 1.0, the overall preferred fiber direction reoriented only about 4° towards the stretch direction (Fig. 17.3c). While for d = 0.25, the overall preferred fiber direction reoriented about 35.0o towards the stretch direction (Fig. 17.3d).
When the tissue was stretched in the X 1 (preferred) direction, more fibers are recruited in the X 1 direction than those in the X 2 direction. For d = 0.25, the polar FEFR plot (Fig. 17.4a) under different stretch ratios revealed the increasing FEFR in all directions. At \( \lambda =1.18 \) the FEFR in all directions was less than 20 %. As the stretch increased to 1.21, more than 40 % of fibers were recruited in the X 1 direction. All the fibers in the X 1 direction were straightened when the strain in X 1 direction just reaches the upper bound strain at \( \uplambda =1.24 \), with all fibers within 24° from the X 1 direction straightened by a stretch of 1.4. The TFR is 40 % uniformly distributed in the center region of the soft tissue (Figs. 17.4c and 17.6b). The maximum TFR occurs at the suture points (Fig. 17.4b, c) due to stress concentration.
17.3.4 Biaxial Test Simulations: Clamped BoundaryConditions
For the tissue with clamped boundary conditions, SALS measurements (Waldman etal. 2002) indicated that fibers between grip faces were highly aligned due to shearing (Fig. 17.5a). The preferred fiber orientation rotated about 37.0° at the corner regions from simulation with d = 0.25. The distribution of the standard deviation of fiber density from simulation is similar to the distribution of orientation index (OI) from SALS measurements. The structural model provided deep insights into the deformation of fibers. All the fibers initially undulated were straightened gradually with increasing load. Less than 1 % of fibers were recruited in the center region of the tissue specimen and more than 70 % of fibers in the corner regions were recruited. Along the diagonal line, the TFR is uniformly distributed around the center region of the tissue specimen. The TFR increased dramatically at the corner regions due to large shear strain.

(a) SALS data (Waldman etal. 2002), (b) preferred fiber orientation and standard deviation contour of fiber angular distribution function from simulation with d = 0.25, (c) total percentage of fiber recruitment contour, (d) total percentage of fiber recruitment along the diagonal line in (c)
17.3.5 Membrane Inflation Simulations
Simulation results revealed that all fibers around the center dome region were straightened (Fig. 17.6a). The preferred fiber direction in the deformed shape was rotated to the radial direction of the tissue (Fig. 17.6a). The TFR decreased gradually from 100 % in the center region to 32 % at the edge (Fig. 17.6a). For element A (Fig. 17.6a) at the center of tissue, the strain is the same in each direction. Therefore the fractional ensemble fiber recruitment is equal in each direction (Fig. 17.6b). However for location B (Fig. 17.6a) at the tissue edge, the strain in the radial direction is larger than that in the circumferential direction. More fibers were recruited in the radial direction (Fig. 17.6c).

Results from the membrane inflation simulations showing (a) preferred fiber orientation and total percentage of fiber recruitment contour from simulation, and the azimuthal plot of the fractional ensemble fiber recruitment of the elements located at (b) element A and (c) element B. Note here that while increased pressure loading predictably increased the total amount of fiber recruitment at both locations, substantial angular variations with pressure was observed at elementB
17.4 Discussion
A framework for the implementation of a structural constitutive model for soft tissues into a finite element framework was developed and validated. Simulation of a single element under uniaxial tension revealed that when using fiber systems with angular dispersion, a matrix phase must be present to prevent nonphysiological deformations. This finding may shed insight into how the non-collagenous components of soft tissues play an important role guiding the overall mechanical responses. For example, Lake and Barocas (2011) studied the effects of simulated non-fibrillar matrix using an agarose analog on the behavior of a collagen-agarose co-gel in uniaxial tension. They reported that the Poisson’s ratio of co-gel decreased from a range of 1.5–3.0 (with a large volume decrease) with no agarose to ∼0.5 (i.e., nearly incompressible) with high concentration of agarose. Thus, both the experimental results and the present simulation results suggest that matrix phase may have significant effects on the mechanical behavior of soft tissues. Biaxial tension simulations demonstrated that the presence of an isotropically oriented fiber phase can significantly affect the overall fiber orientation in the deformed configuration. For d = 1.0, the preferred fiber orientations were far from the SALS measurement for both strip biaxial tests and equi-biaxial tests. However, simulation results with d = 0.25 are in good agreement with the SALS measurement. Not surprisingly, this observation revealed that accurate measurement of the fiber ODF is critical to the structural model. By incorporating fiber orientation distribution and fiber recruitment distribution into the strain energy function, the structural model can predict not only the mechanical behavior of soft tissues at the macroscopic scale, but also fiber deformations patterns (in a statistical sense) at the microscopic scale. The new indices introduced herein may be particularly useful in understanding structural adaptations and could easily be used in structural optimization studies in engineered tissue design. Moreover, the present results underscore the importance of architecture in tissue modeling development and application.
References
ABAQUS. Abaqus user subroutines reference manual. 2011.
Beskos DE, Jenkiins JT. A mechanical model for mammalian tendon. J Appl Mech. 1975;42:755.
Billiar KL, Sacks MS. A method to quantify the fiber kinematics of planar tissues under biaxial stretch. J Biomech. 1997;30:753–6.
Billiar KL, Sacks MS. Biaxial mechanical properties of the native and glutaraldehyde-treated aortic valve cusp: Part II–A structural constitutive model. J Biomech Eng. 2000a;122:327–35.
Billiar KL, Sacks MS. Biaxial mechanical properties of the natural and glutaraldehyde treated aortic valve cusp–Part I: experimental results. J Biomech Eng. 2000b;122:23–30.
Bischoff JE. Continuous versus discrete (invariant) representations of fibrous structure for modeling non-linear anisotropic soft tissue behavior. Int J Non Linear Mech. 2006;41:167–79.
Buchanan RM, Sacks MS. Interlayer micromechanics of the aortic heart valve leaflet. Biomech Model Mechanobiol. 2013;3(4):1–4.
Chen H, Liu Y, Slipchenko MN, Zhao X, Cheng JX, Kassab GS. The layered structure of coronary adventitia under mechanical load. Biophys J. 2011;101:2555–62.
Cortes DH, Lake SP, Kadlowec JA, Soslowsky LJ, Elliott DM. Characterizing the mechanical contribution of fiber angular distribution in connective tissue: comparison of two modeling approaches. Biomech Model Mechanobiol. 2010;9:651–8.
Courtney T, Sacks MS, Stankus J, Guan J, Wagner WR. Design and analysis of tissue engineering scaffolds that mimic soft tissue mechanical anisotropy. Biomaterials. 2006;27:3631–8.
Criscione JC, Sacks MS, Hunter WC. Experimentally tractable, pseudo-elastic constitutive law for biomembranes: I. Theory. J Biomech Eng. 2003;125:94–9.
Driessen NJ, Mol A, Bouten CV, Baaijens FP. Modeling the mechanics of tissue-engineered human heart valve leaflets. J Biomech. 2007;40:325–34.
Fata B, Zhang W, Amini R, Sacks MS. Insights into regional adaptations in the growing pulmonary artery using a meso-scale structural model: effects of ascending aorta impingement. J Biomech Eng. 2014;136:021009.
Fung YC. Biomechanics: mechanical properties of living tissues. 2nd ed. New York: Springer; 1993.
Hansen L, Wan W, Gleason RL. Microstructurally motivated constitutive modeling of mouse arteries cultured under altered axial stretch. J Biomech Eng. 2009;131:101015.
Hariton I, de Botton G, Gasser TC, Holzapfel GA. Stress-driven collagen fiber remodeling in arterial walls. Biomech Model Mechanobiol. 2007;6:163–75.
Hollander Y, Durban D, Lu X, Kassab GS, Lanir Y. Experimentally validated microstructural 3D constitutive model of coronary arterial media. J Biomech Eng. 2011;133:031007.
Holzapfel GA, Eberlein R, Wriggers P, Weizascker HW. Large strain analysis of soft biological membranes: formulatin and finite element analysis. Comput Methods Appl Mech Eng. 1996;132:45–61.
Holzapfel GA, Ogden RW. Constitutive modelling of passive myocardium: a structurally based framework for material characterization. Philos Transact A Math Phys Eng Sci. 2009;367:3445–75.
Horowitz A, Lanir Y, Yin FC, Perl M, Sheinman I, Strumpf RK. Structural three-dimensional constitutive law for the passive myocardium. J Biomech Eng. 1988;110:200–7.
Hughes TJR. The finite element method: linear static and dynamic finite element analysis. New York: Dover; 2000.
Jor JW, Nash MP, Nielsen PM, Hunter PJ. Estimating material parameters of a structurally based constitutive relation for skin mechanics. Biomech Model Mechanobiol. 2011;10:767–78.
Joyce EM, Moore JJ, Sacks MS. Biomechanics of the fetal membrane prior to mechanical failure: review and implications. Eur J Obstet Gynecol Reprod Biol. 2009;144 Suppl 1:S121–7.
Kao PH, Lammers S, Tian L, Hunter K, Stenmark KR, Shandas R, Qi HJ. A microstructurally-driven model for pulmonary artery tissue. J Biomech Eng. 2011;133:051002.
Kenedi RM, Gibson T, Daly CH. Biomechanics and related bio-engineering topics. In: Kenedi RM, editor. Bioengineering studies of human skin. Oxford: Pergamon; 1965. p. 147–58.
Lake SP, Barocas VH. Mechanical and structural contribution of non-fibrillar matrix in uniaxial tension: a collagen-agarose co-gel model. Ann Biomed Eng. 2011;39:1891–903.
Lanir Y. A structural theory for the homogeneous biaxial stress-strain relationships in flat collagenous tissues. J Biomech. 1979;12:423–36.
Lanir Y. Constitutive equations for fibrous connective tissues. J Biomech. 1983;16:1–12.
Marsden JE, Hughes TJR. Mathematical foundations of elasticity. Don Mills: Dover; 1983.
Mirnajafi A, Raymer J, Scott MJ, Sacks MS. The effects of collagen fiber orientation on the flexural properties of pericardial heterograft biomaterials. Biomaterials. 2005;26:795–804.
Mitton R. Mechanical properties of leather fibers. J Soc Leather Trades’ Chem. 1945;29:169–94.
Prot V, Skallerud B, Holzapfel G. Transversely isotropic membrane shells with application to mitral valve mechanics. Constitutive modelling and finite element implementation. Int J Numer Methods Eng. 2007;71:987–1008.
Sacks M. Biaxial mechanical evaluation of planar biological materials. J Elast. 2000;61:199–246.
Sacks MS. Incorporation of experimentally-derived fiber orientation into a structural constitutive model for planar collagenous tissues. J Biomech Eng. 2003;125:280–7.
Soong TT, Huang WN. A stochastic model for biological tissue elasticity in simple elongation. JBiomech. 1973;6:451–8.
Sun W, Sacks MS. Finite element implementation of a generalized Fung-elastic constitutive model for planar soft tissues. Biomech Model Mechanobiol. 2005;4(2-3):190–9.
Sun W, Sacks MS, Sellaro TL, Slaughter WS, Scott MJ. Biaxial mechanical response of bioprosthetic heart valve biomaterials to high in-plane shear. J Biomech Eng. 2003;125:372–80.
Tong P, Fung YC. The stress-strain relationship for the skin. J Biomech. 1976;9:649–57.
Tonge TK, Voo LM, Nguyen TD. Full-field bulge test for planar anisotropic tissues: part II–a thin shell method for determining material parameters and comparison of two distributed fiber modeling approaches. Acta Biomater. 2013;9:5926–42.
Waldman SD, Michael Lee J. Boundary conditions during biaxial testing of planar connective tissues. Part 1: dynamic behavior. J Mater Sci Mater Med. 2002;13:933–8.
Waldman SD, Sacks MS, Lee JM. Boundary conditions during biaxial testing of planar connective tissues: Part II: Fiber orientation. J Mater Sci Lett. 2002;21:1215–21.
Wognum S, Schmidt DE, Sacks MS. On the mechanical role of de novo synthesized elastin in the urinary bladder wall. J Biomech Eng. 2009;131:101018.
Acknowledgements
Funding for this work was supported by FDA contract HHSF223201111595P and NIH/NHLBI Grant NHLBI R01 HL108330 and R01 HL119297-01.
Conflict of Interests
The authors have no conflict of interests to report in this work.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2016 Springer Science+Business Media, LLC
About this chapter
Cite this chapter
Sacks, M.S. (2016). Finite Element Implementation of Structural Constitutive Models. In: Kassab, G., Sacks, M. (eds) Structure-Based Mechanics of Tissues and Organs. Springer, Boston, MA. https://doi.org/10.1007/978-1-4899-7630-7_17
Download citation
DOI: https://doi.org/10.1007/978-1-4899-7630-7_17
Publisher Name: Springer, Boston, MA
Print ISBN: 978-1-4899-7629-1
Online ISBN: 978-1-4899-7630-7
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)