
Abstract
The pursuit of sustainable wheat production has significant economic, social, and environmental relevance. Yield levels and stability thereof are determined by continuously changing and fluctuating biotic and abiotic stresses. Achieving higher and more stable yields requires constant genetic improvement of numerous aspects of new wheat cultivars. Targeted traits may include wide adaptation, abiotic stress such as drought and salinity, tolerance, polygenic non-specific disease resistance, pyramided disease resistance, etc. Broadly defined breeding objectives such as these involve complex, polygenic, genetic mechanisms that pose formidable challenges to breeders. Fortunately, a diverse array of increasingly more sophisticated biotechnological tools is becoming available. Advances in understanding the mechanisms that determine sustainability traits, coupled with versatile and unambiguous genetic markers and generation acceleration methodologies, foster new opportunities for selection of such traits, yet breeding methodologies need to be adapted in parallel to fully capitalize on the new technology.
Recurrent selection applied to a self-pollinator provides for a powerful breeding tool. Continuous cross-hybridization maximizes heterogeneity and forges new linkage associations in genes. Subsequent inbreeding helps to weed out deleterious recessive genes, fixes desirable genes in the homozygous state, and allows for accurate progeny testing. In the past, the difficulty of randomly intercrossing large numbers of selected wheat plants has frustrated the application of the technique, however this problem has been solved through the use of genetic male sterility in conjunction with the hydroponic culture of tillers that are cut and pollinated at anthesis. Thus, it is possible to randomly intercross hundreds of selected genotypes to produce large F1 populations (upwards of 50,000 seeds).
A recurrent wheat mass selection program is being conducted at Stellenbosch University with the purpose of developing and testing the methodology. A highly heterogeneous base population was established and is being managed as a medium-sized breeding program. The experience we gained allowed us to streamline its execution, and herewith we review current methodology and progress made. Recurrent mass selection proved a simple, yet highly effective, technique that has major advantages compared to conventional wheat breeding methods; among these are reduced operational costs, accelerated selection progress, maximization of crossover and genetic recombination potential, and its suitability for broad breeding strategies.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
Introduction
Conventional wheat selection methods involve three distinct stages, starting with the making of planned crosses: normally single, triple, back, or double crosses (Stoskopf, 1999). The second phase involves development of inbred/homozygous lines through single plant selection in the early segregating generations (pedigree method); bulk propagation in the early generations followed by single plant and line selection in the later generations (bulk-population method); and random inbreeding (single seed descent method) or production of homozygotes through androgenesis or wide crosses (doubled haploid method). The final phase of progeny testing of inbred/homozygous lines is the same for the four methods. Annual crosses are normally planned based on the performance of advanced lines in the previous season, thus giving a cyclic long-term nature to the methodologies. New variations may be introduced regularly, normally through a process of prebreeding that limits cotransfer of undesired chromatin and preserves earlier selection gain. Objections to these methods include limited opportunities for genetic recombination and difficulty balancing simultaneous stringent selection of simple traits with high heritability and complex traits with low heritability. Backcross breeding, whereby single, desirable traits are entered into a commercially proven genetic background, is often employed in a supplementary manner in conventional breeding. The purpose may be to upgrade an otherwise superior genotype lacking a critical trait; to provide for prebreeding during introgression of a new trait; or to develop multilines or near-isogenic lines. Major drawbacks of the backcross approach are that it may impose selection ceilings and, if not used with discretion, may seriously narrow the genetic variability of a breeding population.
In conventional breeding of self-pollinating crops, genes are rapidly fixed through self-fertilization following the initial cross, and heterozygosity is halved with every successive filial generation (Stoskopf, 1999). This strongly reduces the opportunity for genetic recombination. In dealing with polygenic traits, it is therefore totally unrealistic to expect that the polygenic recombination potential of a cross can be adequately explored in a single cycle of crossing and selection. Recurrent mass selection is a well-established breeding technique for the genetic improvement of cross-pollinating species (Hallauer, 1981). It was developed primarily for the improvement of quantitatively inherited traits (controlled by numerous genes, each with small effect and modified by the environment), and the underlying objective is to systematically increase the frequency of desirable genes in a breeding population so that opportunities to extract superior genotypes are maximized. The inherently high level of heterozygosity in the breeding population coupled with a large number of cross combinations allow for more complete exploration of polygenic recombination potential (Jensen, 1970; Hallauer, 1981).
The principles underlying recurrent mass selection are equally applicable to autogamous crops, but the difficulty associated with intercrossing in each cycle, coupled with small amounts of seed produced, has discouraged its use. This led Hallauer (1981) to suggest that recurrent selection procedures should preferably be integrated with other selection methods, and that its products could not normally be expected to be directly useful for commercial cultivar development. Pilot studies that applied recurrent selection to self-pollinating small grain cereals generally had positive outcomes; however, these studies were mostly short-term (less than five cycles), were restricted in terms of the number of intercrosses that could be made, and pursued single traits (Wiersma et al., 2001). Wiersma et al. (2001), Díaz-Lago et al. (2002), and Liu et al. (2007) provided excellent overviews, highlighting the effectiveness of recurrent selection for genetic improvement of grain protein, kernel weight, grain yield, or disease resistance in numerous experiments with crops such as soybean, barley, wheat, and oat. Impressive selection progress was recorded for complex traits that are difficult to breed by conventional means—for example, grain yield of oat (De Koeyer and Stuthman, 1998), groat oil content of oat (Frey and Holland 1999), and partial resistance to oat crown rust (Díaz-Lago et al., 2002). In China, recurrent selection based on the Taigu source of male sterility was used to pyramid minor and major genes for scab resistance, improved salt tolerance, drought tolerance, and yield potential (He et al., 2001). However, the focus on single traits often resulted in undesirable correlated changes in unselected traits, thus reducing the usefulness of the strategy and highlighting the necessity for a holistic approach to its use in cultivar improvement (Wiersma et al., 2001).
Wallace and Yan (1998) stressed the importance of a systemic rather than focused approach to plant improvement. The living plant is a complex biological system driven and regulated by extensive and interdependent genetic, epigenetic, and environmental mechanisms. Photosynthate is apportioned for biomass accumulation and harvestable product in accordance with availability and the requirement of metabolic pathways, plant defense, and stress tolerance mechanisms. Selection for a specific trait is therefore likely to impact on interconnected pathways (Comeau et al., 2007), thus affecting overall plant performance. A well-adapted genotype appears to be better able to optimize its responses to the physiological demands of a particular environment, and to breed and select these genotypes, breeding should be less focused on narrowly defined objectives and should be more holistic and multidisciplinary based (Wallace and Yan, 1998; Comeau et al., 2007). During selection, a broad range of biotic and abiotic stresses relevant to the targeted production region should be applied. When done in conjunction with recurrent or convergent breeding strategies, this could aid the development of broadly adapted genotypes.
Jensen (1970) suggested the diallel selective mating system as a means to accommodate recurrent selection in self-pollinating species that are difficult to cross, and that produce few seeds per cross. This strategy integrates recurrent selection principles with conventional breeding strategies, and thus allows for simultaneous genetic input of a broader range of parents, breaking up linkage blocks, and freeing genetic variability by fostering genetic recombination. Jensen (1970) furthermore suggested the use of male sterility to facilitate crossing, and pointed out that the breeding material should be selected in specialized environments to maximize the genotypic expression of the traits being selected. McProud (1979) pointed out that the majority of established cereal-breeding programs around the world have a cyclic nature and may contain an element of recurrent selection. Such programs involve the generation of variability through crosses, followed by the derivation and evaluation of inbred lines. Superior lines are then intercrossed to sustain a next cycle of selection. While they have been successful, these programs are hampered by long selection cycles, and breeders need to guard against genetic bases that are too narrow and not enough introgressions of new variability. To maintain selection progress, it is necessary to introgress new useful variation in a controlled manner that would minimize the introduction of deleterious genes and preserve existing favorable gene combinations. Falk (2002) developed and tested a recurrent selection strategy called recurrent introgressive population enrichment (RIPE) in barley that was designed to address the above shortcomings of conventional breeding methodologies. The system is based on a recessive gene for male sterility (msg6) closely linked in a coupling phase with a recessive, xenia-expressing shrunken endosperm gene (sex1). F2 seeds with shrunken endosperm primarily develop into male-sterile (female) plants. Crosses and F1 multiplication are done in growth rooms during the off-season, the F2 is planted in the field in May of Year One; the F3 is grown in an off-season nursery; and the F4 is evaluated in an unreplicated field trial. Selected lines provide male parents in the next cycle of crosses and are also destined for advanced testing in multiple-location trials. Crosses are made annually and thus the duration of the female selection cycle (one year) differs from the male selection cycle, which extends over two years. A limited number of crosses (NC Design 1) are made each year, but the elite population is continuously enriched with new variation; this is achieved through eight generations (three years) of crosses with the elite population, resulting in the introgression of approximately 6% new alleles at a time. World Wide Wheat (W3) is a wheat breeding company that commercially employs male sterile facilitated recurrent selection (MSFRS). Compared to conventional pedigree breeding, the recurrent methodology claims to allow for more rapid and efficient cultivar development (http://www.worldwheat.com/company_overview.php).
The base population under recurrent selection should be assembled carefully to contain adequate genetic variability. It should provide scope for the genetic improvement of adaptation, production, and processing characteristics peculiar to the crop and targeted production region. A recurrent selection program aimed at increasing kernel weight in spring wheat was initiated at North Dakota State University in 1967 (Busch and Kofoid, 1982). Following a screening of about 100 cultivars and breeding lines for kernel weight, the ten best lines were intermated in the 45 possible cross combinations to form a base population. Subsequent cycles were generated through manual crosses of representative numbers of selected plants from the segregating generations.
Making use of the dominant male sterility gene, Ms2 (Taigu source), Huang and Deng (1988) established a recurrent selection breeding population segregating for male sterile (female) and male fertile plants. In their system, selected female plants were naturally (field) pollinated by selected male fertile plants. A small, nation-wide network of Chinese researchers pursued recurrent selection-based applications, and released a number of new cultivars (He et al. 2001). Cox et al. (1991) developed and registered a germplasm source segregating for the presence of the dominant male sterility gene, Ms3 (derived by Maan and Williams 1984). Over the course of several years, numerous sources of diverse disease- and insect-resistant, quality and yield genes were involved as parents to end up with a highly heterogeneous base population. Cross-pollination of male sterile spikes by fertile plants (greenhouse) was enhanced using fans and manual agitation. Marais et al. (2000), made use of the dominant male sterility gene Ms3, to establish a recurrent selection base population consisting of 50% male sterile plants. They developed a simple hydroponic system to achieve large-scale random intercrossing of the selected plants in a greenhouse. Liu et al. (2007) established a recurrent selection base population by pollinating a source with the D2-type of cytoplasmic male sterility, with 30 diverse elite cultivars and lines derived from six provinces in China.
Male sterility genes provide an easy means to obtain female plants for use in recurrent selection applications. However, numerous chemical hybridizing agents have been described (Chakraborty and Devakumar, 2006) that provide an alternative to the use of genetic or cytoplasmic male sterility for the implementation of a hybridization strategy.
The overall goal of recurrent selection is to increase the frequency of desirable genes in the base population, yet maintain genetic diversity while doing so (Wiersma et al. 2001). Continued selection progress depends on the amount of variation present, heritability of the trait, and the initial frequencies of desirable alleles. Past studies have suggested that for complex, quantitatively inherited traits, steady selection progress can be maintained for many cycles. If properly managed to avoid sampling effects, variation for unselected traits can similarly be maintained within the recurrent population (De Koeyer and Stuthman, 1998; Frey and Holland, 1999; Wiersma et al., 2001). It is also necessary to provide for continuous introgression of new, useful genes in the base population. This needs to be done in a manner that will not undo the selection progress already achieved, and therefore has to involve backcrosses to the base population (Marais et al., 2001a; Falk, 2002).
Sustainability of Rust Resistance
Commercial wheat production relies on genetically uniform, high-yielding cultivars that are grown over large areas of land, often for many years in succession. Resistance to the rusts is often based on single, major genes that exert strong selection pressure on the pathogen and, as a consequence, may be short-lived. “Arms races” result between wheat breeders and the pathogen that necessitate an ongoing search for new, effective rust resistant genes to employ and steer pathogen evolution in specific directions (Knott, 1989). This situation contrasts starkly with the high level of genetic diversity in natural grass populations, land races, and crop mixtures that historically serve to buffer the spread and evolution of a pathogen.
The desirability of reintroducing genotypic diversity of resistance in modern-day commercial wheat production has been advocated (Browning, 1988; Knott, 1989). Suggested ways in which this can be achieved include the use of species mixtures, cultivar mixtures, and clean and dirty multiple lines. A cultivar mixture is a comparatively simple strategy that effectively reduces the damage caused by rust diseases (Browning, 1988; Wolfe, 1988) but could compromise uniformity (Groenewegen and Zadoks 1979). Interfield diversity and regional deployment strategies to slow down pathogen progression are difficult to manage and are often not widely accepted by farmers (Frey et al., 1979; Knott, 1989). Multiple lines were developed in an attempt to improve the agronomic and technological disadvantages of cultivar mixtures (Groenewegen and Zadoks, 1979). The development of the near-isogenic or phenotypically similar components and continued maintenance of multiple lines are time- and effort-consuming, and the strategy is limiting as far as genetic progress with selection for nondisease traits is concerned (Frey et al., 1979; Groenewegen and Zadoks, 1979). However, multiple lines are reportedly effective in slowing down disease progression (Browning et al., 1979; Browning, 1988; Frey et al., 1979).
In cross-pollinators, recurrent selection can be used to develop and continuously improve open-pollinated cultivars. If the same principle is applied to a self pollinator, it is possible to breed cultivars that would be akin to modern land races. In such an approach, numerous diverse resistance genes may be introduced in the recurrent base population and selected to establish different gene frequencies depending on the nature of the pathogen population. The base population that is released as a “land-race” cultivar can be diversified continuously in terms of its resistance, and at the same time be selected for uniformity (agrotype and processing quality) and improved agronomic performance. To counter shifts in the resistance gene frequency that are due to natural selection, new foundation seed can regularly be derived from the base population. Compared to a multiple line, and as a result of the high level of heterogeneity and recombination in the base population, a “land-race” cultivar could include a much wider array of genotypic combinations of resistance genes that also exploit complementation, interaction, and additive effects of genes.
An alternative to intracultivar genotypic diversity is to pyramid universally effective resistance genes in a single genotype. This can also provide a more durable barrier to pathogen adaptation, which requires the pathogen to simultaneously mutate at a number of loci to be able to overcome the complex, polygenic resistance (Knott, 1989). Both the multiple line and gene pyramid strategies require a thorough knowledge of the dynamics of the pathogen population, and continuous adjustment with respect to genes that become ineffective and are complicated by the simultaneous use of the same genes in cultivars with single gene resistance (Browning 1988; Wolfe, 1988; Knott, 1989). Pyramiding of major resistance genes is complicated by the fact that the gene with the strongest effect masks the presence of genes with lesser phenotypic expression, and is thus best achieved with the use of markers, allowing all genes present to be known. Most attempts to pyramid resistance are based on backcrosses or convergent crosses that impose a yield ceiling and make it problematic to add onto existing gene pyramids. Recurrent selection, on the other hand, allows for continued pyramiding, without the sacrifice of selection progress for other traits (Pretorius et al., 2007).
The pursuit of durable resistance may be a more sensible approach towards achieving sustainable rust resistance. Polygenic, nonspecific resistance is conditioned by the presence of a number of genes, each with small effect on the total resistance phenotype. Should virulence develop for one of the components, this should result in only a small phenotypic effect. Polygenic, nonspecific resistance is postulated to be incomplete (yet this need not always be the case) and to exert only mild selection pressure on the pathogen (Parlevliet, 1988; Knott, 1989). Past experience has shown that partial or durable resistance is not necessarily polygenic (McIntosh, 1992; Rubiales and Niks, 1995). Similarly, while adult plant resistance is often partial (Bariana and McIntosh 1995) and likely to be durable (McIntosh, 1992), it can either be race-specific or race nonspecific (Kaur et al., 2000).
By definition, durable resistance remains effective after widespread use over a long period of time (Johnson and Law, 1975). Since durability is difficult to measure, it is problematic to breed for and generally entails selection for component traits. Breeding for durable, polygenic resistance requires the use of a pathotype that is virulent on all the parents in the seedling stage, or the prior elimination of cross progenies with genes for specific resistance. Selection then needs to focus on aspects such as incubation period, and number and size of uredia (Browning, 1988; Parlevliet, 1988; Knott, 1989). In areas where more than one rust disease is of importance, attempts to develop durable resistance may need to simultaneously address all, which may be difficult (Groenewegen and Zadoks, 1979; Frey et al., 1979).
Parlevliet and van Ommeren (1988) demonstrated that mild recurrent mass selection against susceptibility provided a powerful tool for the accumulation of partial resistance genes (in the absence of major race-specific genes and employing a defined pathogen population). The shadowing effect of race-specific resistance genes on phenotypic selection for durable resistance limits its integration in commercial breeding. This can be overcome by prior genetic analysis of appropriate segregating populations to identify and tag quantitative trait loci (QTL) for durable resistance that would allow marker-assisted breeding of the trait (Castro et al., 2003; Balakrishna et al., 2004).
Ultimately, the most appropriate strategy for achieving sustainable genetic control of rust diseases would be to combine single or multiple genes for durable resistance with major race-specific genes. This can be achieved by pyramiding the target genes in a single genotype or by combining genotypes with diverse resistances in a multiple line or a land-race-type cultivar. Such strategies will be technically challenging and will, of necessity, have to rely strongly on the availability of tightly linked molecular markers. While back- or convergent-crossing schemes are often employed in gene pyramiding and for the development of multiple lines, they impose yield ceilings. Recurrent selection, on the other hand, provides a highly effective alternative procedure of gene pyramiding and breeding for land-race cultivars that does not limit the genetic improvement of any other trait. Furthermore, recurrent selection is the breeding scheme best suited to holistic breeding objectives that encompass rust resistance, broader disease, and pest resistance, wide adaptation, yield, and quality. Because it is a population improvement strategy, it enables a breeder to focus on specific breeding targets at a given time without forfeiting the opportunity to subsequently select for other traits. Provided that the population is large enough and selection bottlenecks are avoided, variation for unselected and uncorrelated traits should remain unaffected, allowing for continued and intense recombination and exploitation.
Establishing a Recurrent Mass Selection Base Population at Stellenbosch
A genetically diverse base population, rich in genes for adaptation, quality, yield, and pest resistance, and segregating for male sterility was established. Accession KS87UP9 (Cox et al. 1991) of winter wheat segregating for the dominant male sterility gene, Ms3 (Maan and Williams, 1984), was obtained from the USDA-ARS, Dept of Agronomy at Kansas State University. A male sterile KS87UP9 plant was pollinated with the spring wheat, “Inia 66,” and sterile F1 plants were pollinated with a spring wheat breeding line. Male sterile F1 with a spring growth habit were then used in a multicross with seven spring wheats. Sterile male multicross F1 plants were subsequently randomly intercrossed with 60 wheat breeding lines with diverse disease resistance. In the following cycle (1999), 44 selections from a pedigree program and 157 selections from the recurrent program were used as male population. In 2000, the male population consisted of 64 selections from the pedigree breeding program and 157 selections from the recurrent population. In 2001–2004, the recurrent F1 was annually crossed with 60–120 selections from a pedigree breeding program. As will be shown in later sections, the selection cycle in this program extends over four years, and in each year the F1 of the previous year was used as a female parent. As a result, the genetic contributions of the various parental populations to the base population could be estimated as: (a) 2000 F1 female population : 12.5%; (b) 2001 male population : 12.5%; (c) 2002 male population : 23%; (d) 2003 male population : 24%; and (e) 2004 male population : 30%. In 2005, the base population was closed and male plants were only selected from within the recurrent mass selection base population. Subsequently, new variations would only be added once it had gone through a cycle of “recurrent backcrossing.”
The Recurrent Mass Selection Breeding Plan
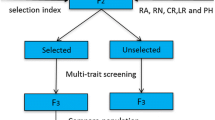
Marais et al. (2001a, b) proposed a recurrent mass selection strategy for wheat that is outlined in Fig. 1 . The duration of a breeding cycle is shown to be four years whereas the male and female components are handled differently. In this scheme, male selection (F6) is based on performance in an unreplicated single row at a single locality. This allows for a four-year breeding cycle that is an advantage in the initial stages of selection in a highly heterogeneous base population. Strict initial selection for simple, highly heritable traits (while maintaining large populations) means that the base population can be enriched for these traits without loss of heterogeneity for complex traits of low heritability. However, in the longer run, the breeding cycle may be extended to allow for better sampling of the target mega environment and to improve selection gain for quantitative traits of low heritability.

Common wheat recurrent mass selection scheme (abbreviations: DH = doubled haploid; MAS = marker-assisted selection; RMS = recurrent mass selection; SSD = single-seed descent)
Making of Crosses and Hydroponic Maintenance of Cut Tillers
To facilitate intercrossing, male sterile and male fertile spikes of selected plants are cut at the time of flowering and kept in hydroponic solution to effect pollination, where only the female tillers are maintained until the seeds ripen (Fig. 2 ). Galvanized iron trays with dimensions of 600 mm x 450 mm x 160 mm (and coated on the inside with black antifungal paint) are used that can each accommodate 230 female spikes. Up to 70 male spikes are placed into two narrower trays positioned on either side of the main tray and raised about 600 mm. The main tray is fitted with inlet and outlet holes for changing the nutrient solution without having to remove the female spikes. Enough spikes to fill 1–2 containers are collected twice a week over a period of six weeks. This results in a total of 12–15 trays (approximately 2,760–3,450 female spikes). Each container is filled with 40% nutrient solution and 0.01% “Jik” (household detergent containing 3.5% sodium hypochlorite; Reckitt and Colman South Africa Pty Ltd, Elandsfontein, South Africa) in tap water. The nutrient solution (pH neutral with electrical conductivity = 1.5–2.0 dSm–1) is made up of 164 g Sol-u-fert T3T (Kynoch Fertilizers Pty Ltd, Milnerton, South Africa), 2 g Microplex (Ocean Agriculture Pty Ltd, Muldersdrift, South Africa) and 77 ml potassium nitrate in 100 L H2O. The containers are kept in a growth chamber with 14 h day/10 h night cycles at 16oC/12oC for the duration of grain filling. An air pump is used to continuously aerate the solution. Female tillers are cut just below the second-to-last internode, and care is taken to keep the flag leaf intact. Florets on female tillers are cut open to facilitate pollination and, in the process, incompletely male sterile spikes are discarded. Male tillers are collected in buckets, stripped of their leaves, and arranged above the female tillers. As a result of the extensive handling, male tillers shed all their ripe pollen by the time they are arranged in the containers. This means that the next pollen shed is more or less synchronized among tillers. After allowing 5–6 days for pollination, the male tillers are discarded. Female tillers are trimmed and transferred to fresh nutrient solution once every two to three weeks.

Making of crosses: (a) F1 population segregating for male sterility; (b) male sterile (female = Msms) spike; (c) random female spikes being pollinated with random male spikes; and (d) hybrid seeds ripening on the female tillers
The Crossing Block
Each year, 9,000–12,000 F1 hybrid seeds from the preceding season are planted in a growth chamber and seedlings are screened with an inoculum mix consisting of 5–8 Puccinia triticina (leaf rust) and 3–5 P. graminis f. sp. tritici (stem rust) pathotypes (Fig. 1). Approximately 3,000 seedlings exhibiting the lowest levels of infection are transplanted to the crossing block. Fifty percent of these will be male sterile plants and can be used as females (the remaining 50% of male plants are used to initiate single-seed descent inbreeding). F6 inbred lines (100–120) that were field-selected in the previous season and originate from crosses made four years earlier are used as the male parents. In our system, seed set is about 90–95%, and we produce roughly 60,000–70,000, hybrid seeds per season. This is about five times the amount of seed we manage to screen. The 1,000-grain mass of seeds is about 16 g. While small, the seeds are well-developed and result in 85–90% germination.
The Breeding Cycle
Selection is done on both the male sterile and male fertile components. However, only the male fertile populations are field-tested. The male fertile plants are advanced rapidly from the F1 to the F5. Following the evaluation of F6 rows (unreplicated trial) for yield, agrotype, disease resistance (supplemented with marker-aided selection), and quality, the superior selections are used as male parents for hybridization. In conformance with funding conditions (Winter Cereal Trust), a set of lines with commercial potential is also identified at this time and distributed as an annual nursery to local wheat breeders (thus, the project also serves as a national prebreeding program). Had it not been the case, advanced progeny testing and selection of the F5-derived lines would have been done the same way, as in a regular breeding program for a self-fertilizing crop.
The F1 male plants are advanced to the F4 before field planting in May of the ensuing year (Fig. 1); growing two single seed descent populations in an uncooled greenhouse during the summer makes this possible. The F5 is grown under irrigation in the summer to yield F6 seed for unreplicated trials in May of the third year. Thus, generation acceleration comparable to doubled haploid technology is achieved through single-seed descent and summer planting. Advantages compared to doubled haploids are that it is much cheaper and larger numbers can be handled. Furthermore, it is possible to do seedling leaf and stem rust resistance and quality (sodium dodecyl sulphate (SDS) sedimentation) screens and selection at the start of each single-seed descent cycle. As a result, a large proportion of lines that are deficient in these respects can be discarded early on, making the process more cost-effective. Population size need not be compromised by strict selection, as larger numbers may simply be involved in the initial single-seed descent phases. Single-seed descent makes it possible to limit the total selection cycle to four years, which is a huge advantage in terms of realizable selection gain. The expected duration from the time a cross is made to the time a cultivar is released can be reduced accordingly (from 12–13 years to 7–8 years) and is similar to the length of the breeding cycle in doubled haploid breeding.
In a warm climate such as the Republic of South Africa, generation acceleration is ideally achieved through single-seed descent inbreeding. Short growing season cultivars are being bred, and by providing supplementary light during late fall, we manage to produce three generations per year. However, if resources are not limiting, or when dealing with intermediate or winter wheat, it would be preferable to produce doubled haploids instead. This alternative is also shown in Fig. 1. A general problem associated with the development of doubled haploids from heterogenous genotypes is the high number of lines that are subsequently discarded on the basis of highly heritable, simple traits, such as disease susceptibility (Kuchel et al., 2005). It would therefore be sensible to grow the male fertile F1 as a space-planted population in the field, and to select single plants on the grounds of agronomic and disease phenotype to be used for doubled haploid production in the third year. F2 segregates may again be seedling screened in a greenhouse before use. In the fourth year, a field-planted doubled haploid nursery can be grown and screened for agronomic, disease, and quality attributes. This population will then serve as a basis for the selection of male parents and genotypes destined for advanced testing. Whereas use of doubled haploids in recurrent selection may initially be complicated by genotypic variation for wide crossability or androgenetic response (Eudes and Amundsen, 2005), the frequency of genes that promote haploid production should increase over time in the recurrent mass selection population as a result of its indirect selection, thus facilitating doubled haploid production.
To facilitate selection for broad adaptation, it is possible to grow each of the F4 and F6 populations (single-seed descent option; Fig. 1) at different localities that will exert different environmental pressures. It is even possible to involve up to four localities in the course of a four-year cycle. Over time, the recurrent population is expected to become enriched with favorable alleles and it may then be advantageous to increase the selection cycle to five or six years, thereby creating an opportunity for more precise multiple locality testing of inbred/doubled haploid lines to improve the selection of quantitative traits.
Effective Population Size
Since the selection cycle extends over four years, the effective size of the base population is determined by the material handled within a four-year period. Involvement of comparatively large numbers of genotypes further buffers the effect that very strict selection (small groups of selected male parents) may have on the base population’s genetic background. Since the recurrent mass selection system can easily produce large numbers of progeny, it is advisable that very strict selection should always be offset by an increase in the size of the selected population.
Raising Allele Frequencies through Recurrent Mass Selection
Recurrent selection may be very effective in raising the frequency of single major genes of high heritability, such as those for wheat leaf-, stem-, and stripe rust resistance, as well as genes for which highly diagnostic markers are available. It will be equally effective to reduce the frequency of an undesirable gene. Figure 3 (a) shows the effect on allele frequency when the F1 only, the F6 only, or both the F1 and the F6 are selected (making use of the selection scheme outlined in Fig. 1 and with the single-seed descent option), assuming dominance and an initial allele frequency of 5%. It is evident from the graph that with selection in the F1, allele frequency changes gradually during four cycles and then increases sharply when the effect of selection is also being carried through the male parent. F6 selection (F5-derived inbred lines) is far more efficient in raising gene frequencies. One and two cycles of F6 selection is predicted to raise gene frequencies to higher levels than five and ten cycles of F1 selection, respectively. If the male population is inbred for five generations, resistant plants selected in the F6 are mostly homozygotes, primarily contributing gametes with the desired allele to the next generation (the process is therefore comparable to a backcross). Combined F1 and F6 selection is even more effective, but would be relatively costly.

(a) Expected change in the frequency of a target allele (initial frequency, p = 0.05) if selection is done in the F1 only; in the F6 only; or in both the F1 and the F6. (b), (c) & (d) Expected changes in target allele frequencies when selection is done in the F6 only. Also shown (broken lines) are the subsequent fluctuations in gene frequencies when selection is stopped after (b) one, two, three, or four years (initial p = 0.05); (c) three or four years (initial p = 0.40); and (d) two or three years (initial p = 0.60)
Clearly, strict selection of the male parent (F6 inbred lines) should only be done when gene frequencies are sufficiently high to prevent bottlenecks, or if large populations can be screened to ensure maintenance of genetic diversity for other characteristics. Should the initial frequency of an allele be very low, phenotypic or marker-assisted selection can be done at the onset of single-seed descent inbreeding to enrich the F6 field population in terms of the targeted genotypes, thus avoiding loss of heterogeneity through the use of small male populations.
If a base population is being selected with the purpose of deriving single genotypes with pyramided resistance, it may suffice to simply raise the frequencies of desirable resistance alleles to levels higher than 0.70. If the base population is being developed into a land race cultivar, it may be necessary to fix the frequencies of genes affecting phenotypic uniformity, and to raise the frequencies of resistance genes in accordance with the frequencies of virulence genes in the pathogen population (Frey et al., 1979). Figure 3b, c, d shows the effect of terminating selection (F6) at different stages for three different initial gene frequencies. If the frequency of the desired allele is initially very low, say 5% (Fig. 3b), then at least four seasons of selection are required to raise it to a level higher than 70%. When the initial frequency is about 0.40 (Fig. 3c), three seasons of selection will raise it to an average level exceeding 70%. When the initial frequency is 0.60 (Fig. 3d), two seasons of selection will raise it in excess of 70%.
Marker-Assisted Breeding
As gene marker technology continues to improve, it finds ever more application in routine breeding; however, compared to phenotypic measurement, the use of markers remains costly. It is therefore critically important to determine at which stage of a breeding program markers should be employed to ensure maximum genetic effectiveness and economic efficiency (Bonnett et al., 2003; Kuchel et al., 2005). In the present recurrent mass selection program, markers can be employed for F1 screening; for screening during single-seed descent inbreeding, or for screening of the F5-derived F6 inbred lines for use as parents. From Fig. 3, it appears that marker-assisted selection in the F6 would genetically and cost-wise be the most effective strategy. Marker selection done during single-seed descent inbreeding cannot be more effective than F6 selection in raising the frequency of the target allele, and of necessity will involve a higher volume of analyses.
Due to the high level of heterogeneity in the base population, markers should ideally be allele-specific. Such markers are normally easy to obtain for genes carried on alien translocation segments, yet are scarce in the case of genes that are of common wheat ancestry. However, if closely linked markers are available for a gene that is of common wheat origin, it may still be used effectively if a high level of linkage disequilibrium exists within the population. In this case, it will be necessary to determine the frequencies of the possible marker-trait associations that exist within the breeding population beforehand, making use of a random sample of genotypes. If a marker is positively linked to the target trait in a sufficiently high proportion of the population, it can still be used to raise frequencies to desired levels.
Use of Markers in Conjunction with Recurrent Mass Selection to Pyramid Rust Resistance Genes
In the recurrent mass selection scheme of Fig. 1 (single-seed descent option), the percentage of F6 inbred lines that can be expected to carry the target alleles for various numbers of (independent) genes and gene frequencies were calculated by Marais and Botes (2003). For example, when the frequencies of the target alleles are 0.7 with 10 genes targeted, about 2.6% of the inbred lines will carry all ten genes (in hetero- or homozygous form). With 15 genes targeted (allele frequencies 0.7), about 0.4% of the F6 can be expected to carry all of them. If the frequency of the desired allele at each of 20 loci is 0.80, about 1% of the inbred plants are expected to contain them all. Numerous other inbred lines will clearly have diverse combinations of fewer target genes that may still be very useful. Thus, if carefully managed, the recurrent population will in time become a sustainable and continuously improving source of diverse pyramided genotypes.
Evaluation of F6 Inbred Lines
About 1,500 new F6 inbred lines annually are field-tested in an unreplicated trial. Each line is planted as a single 3 m row, and is compared against the most successful commercial cultivars. Disease spreader rows are used to ensure that epidemic conditions will develop. Apart from natural infection, the spreaders are inoculated with prevalent pathotypes of Puccinia triticina (leaf rust), P. graminis f. sp. tritici (stem rust) and P. striiformis f. sp. tritici (stripe rust). Each F6 line is scored for Blumeria graminis f. sp. tritici (powdery mildew), leaf rust, stripe rust, stem rust, and Stagonospora nodorum (Septoria leaf and glume blotch) reaction, growth habit, plant type, straw strength, shattering, yield, quadrumat extraction, and mixograph characteristics. In addition, DNA extracts are made and lines screened by polymerase chain reaction (PCR) for the presence of targeted resistance genes. Marker-assisted selection is most cost-effective if done following the F6 field and quality evaluation of inbred lines. For this purpose, F7 seeds (± 4–5 seeds per inbred line) of selected lines are germinated in plastic dishes (25 compartments). The 1–2 mm long coleoptiles are harvested and pooled for DNA extraction.
Before the male parents are selected, it is possible to confirm that a targeted gene does not have negatively associated effects, and to abort its selection if it does. To do this, the total data set can be analyzed to confirm that there are no undesirable associations. Data contrasts may be derived to test whether the presence of the target gene may result in deleterious agronomic effects such as may be the case with some species-derived translocations. Following evaluation, the best 110–120 lines are selected to be used as male parents. Since 2006, the best material is also included in a nursery of prebred resistant genotypes distributed to local breeders who will evaluate it for direct release or use as parental material. The 2006 and 2007 nurseries were well received by breeders and were primarily used in crosses to improve the levels of rust resistance in their breeding populations.
Introgression of New Resistance Genes through Recurrent Backcrosses
To sustain genetic improvement, it is necessary to continuously introgress new and useful genes. To prevent degradation of the base population by the introduction of poorly adapted germplasm, new genes can be integrated via a series of backcrosses to the recurrent mass selection population. Currently, the genes Lr19-149 (lacking Sr25 as well as the yellow pigment genes; Marais et al., 2001b), Lr21, and the Sr31 complex (1BS38:9.1BL translocation that lacks Sec-1 yet has the Gli-1/Glu-3 loci; Lukaszewski, 2000) are being introgressed through recurrent backcrosses.
The germplasm carrying the targeted genes is used to pollinate male sterile spikes taken from the most recent F1 of the base population (Fig. 4 ). F1 carrying the target genes are then backcrossed to the most recent F1 base population. Such backcrosses need to involve large numbers of plants, particularly the final backcross. It appears that three backcrosses are adequate. Following the third backcross, the recurrent backcross population retains an average of 6.25% of the donor germplasm, and is substituted for the normal recurrent mass selection F1 population. The first male parents carrying the newly introgressed genes will be developed from this substituted population, and these will therefore only be available for crosses four years later. Due to this dela, the average percentage of foreign chromatin (in the F1) will continue to be diluted for another three years and then will raise slightly as the first male parents with introgressed genes come into use. The percentage of foreign chromatin introduced should eventually stabilize at between 2–2.5%.

Recurrent backcross strategy used to deliver new resistant genes to the recurrent mass selection base population (Abbreviations: RB = recurrent backcross; RMS = recurrent mass selection)
Recurrent backcrosses may also be used as an alternative to prevent bottlenecks arising from the initial selection of a gene that occurs at very low frequency in the base population. By initiating a subpopulation and continuously backcrossing to the latest F1 while involving large enough numbers, the frequency of the gene can be raised without skewing or loss of background diversity.
Conclusion
If properly managed, recurrent mass selection is very effective. In our program molecular marker-assisted selection for rust resistance was not introduced until 2005. Prior to that, selection was based on the resistance phenotype only. We practiced strict phenotypic selection for low incidence of susceptible pustules (following seedling inoculation with mixed stem rust and leaf rust spores) combined with adult plant leaf; stem, and stripe rust field resistance, and witnessed a rapid change in the average levels of resistance in the base population.
The recurrent mass selection procedure can readily be executed on a large scale and with a modest budget. The program is surprisingly cost-effective, and in our opinion more so than conventional wheat breeding techniques such as pedigree, bulk, and doubled haploid. The labor required for producing 60,000–70,000 F1 hybrid seeds is roughly equivalent to making about 150–200 planned crosses by hand in a medium-sized pedigree breeding program. The number of F1 produced was always well beyond the numbers we could utilize in a season. In this program, there are no pedigrees to keep and inbred lines are simply numbered consecutively in the F6. The strong emphasis that is placed on early generation selection means that lines with obvious defects are eliminated early on, making it possible to carry larger initial populations. Since only the male parents are field-tested, selection response for adaptation, quality, and yield will be halved; however, much of this effect will be offset by the inbreeding steps.
Application of the technique can be very varied—for example, it may be used as a breeding strategy in its own right, employing either a single base population or several different base populations. Such populations may be used to derive either inbred lines or land race cultivars. On the other hand, it may be used to supplement a conventional pedigree breeding program with the aim to optimally exploit and pyramid beneficial genes within a small group of well-adapted elite lines. Recurrent mass selection applications may also differ with respect to the composition of the base population. The base population may be genotypically highly variable and assembled from diverse and not necessarily adapted germplasm. This would typically be the case, where a new breeding program is being initiated and the breeder wishes to experiment with a wider range of variation. On the other hand, it may be possible to construct the base population in such a way that it will be uniform in some aspects, yet variable for other traits—for example, by combining near-isogenic lines carrying a range of diverse resistance genes in a genetic background that has good processing quality and agrotype. In such a population, screening for quality and agrotype is negated and the breeder can focus all his effort on the development of lines with complex resistance.
References
Balakrishna P.V., Bariana H.S., Singh R.P., Verbyla A., Park R.F. (2004) Identification of genomic regions associated with durable stripe rust resistance in wheat line 11IBWSN50, in: Fisher, T. et al. (Eds.), Proc. 4th Int. Crop Science Congress, Brisbane, Australia.
Bariana H.S., McIntosh R.A. (1995) Genetics of adult plant rust resistance in four Australian wheats and the French cultivar “Hybride-de-Bersée”. Plant Breeding 114, 485–491.
Bonnett D.G., Spielmeyer W., Rebetzke G.J., Ellis M.H., Richards R.A. (2003) A holistic approach to marker implementation, in: Pogna N.E., Romanò M., Pogna E.A., Galterio G., Proc. 10th Int. Wheat Genet. Symp., SIMI, Via N Nisco 3/A-00179, Rome, Italy, pp 105–108.
Browning J.A., Frey K.J., McDaniel M.E., Simons M.D., Wahl I. (1979) The bio-logic of using multilines to buffer pathogen populations and prevent disease loss, Indian J. Genet. Plant Breed. 39, 3–9.
Browning J.A. (1988) Current thinking on the use of diversity to buffer small grains against highly epidemic and variable foliar pathogens: problems and future prospects, in: Simmonds N.W., Rajaram S. (Eds.), Breeding strategies for resistance to the rusts of wheat. Mexico, D.F. CIMMYT, pp 76–90.
Busch R.H., Kofoid K. (1982) Recurrent selection for kernel weight in spring wheat, Crop Sci. 22, 568–572.
Castro A.J., Capettini F., Corey A.E., Filichkina T., Hayes P.M., Kleinhofs A., Kudrna D., Richardson K., Sandoval-Islas S., Rossi C., Vivar H. (2003) Mapping and pyramiding of qualitative and quantitative resistance to stripe rust in barley, Theor. Appl. Genet. 107, 922–930.
Chakraborty K., Devakumar C. (2006) Evaluation of chemical compounds for induction of male sterility in wheat (Triticum aestivum L.), Euphytica 147, 329–335.
Comeau A., Langevin F., Gilbert J., Voldeng H., Savard M., Dion Y., Rioux S., Martin R.A., Haber S., Somers D. (2007) A multi-stress selection approach with better biodiversity of resistance mechanisms achieves good results for the development of Fusarium resistant germplasm – the example of FL62R1 wheat, in: Clear R. (Ed.), 5th Canadian Workshop on Fusarium Head Blight, Canadian Grain Commission, Winnipeg MB, Canada.
Cox T.S., Sears R.G., Gill B.S. (1991) Registration of KS87UP9, a winter wheat germplasm segregating for a dominant male sterility gene, Crop Sci. 31, 247.
De Koeyer D.L., Stuthman D.D. (1998) Continued response through seven cycles of recurrent selection for grain yield in oat (Avena sativa L.), Euphytica 104, 67–72.
Díaz-Lago J.E., Stuthman D.D., Abadie T.E. (2002). Recurrent selection for partial resistance to crown rust in oat, Crop Sci. 42, 1475–1482.
Eudes F., Amundsen E. (2005) Isolated microspore culture of Canadian 6x triticale cultivars, Plant Cell, Tissue Org. Cult. 82, 233–241.
Falk D.E. (2002) The theory, methods, and results of using recurrent selection in breeding barley, in: Mare, Faccioli and Stanca (Eds.), Proc. EUCARPIA, Cereal Sec., Salsomaggiore, Italy, pp 83–88.
Frey K.J., Browning J.A., Simons M.D. (1979) Management systems for host genes to control disease loss, Indian J. Genet. Plant Breed. 39, 10–21.
Frey K.J., Holland J.B. (1999) Nine cycles of recurrent selection for increased groat-oil content in oat, Crop Sci. 39, 1636–1641.
Groenewegen L.J.M., Zadoks J.C. (1979) Exploiting within-field diversity as a defence against cereal diseases: A plea for “poly-genotype” varieties, Indian J. Genet. Plant Breed. 39, 81–94.
Hallauer A.R. (1981) Selection and breeding methods, in: Frey K.J. (Ed.), Plant Breeding II. Iowa State University Press, Ames, Iowa 50010, USA, pp 3–55.
He Z.H., Rajaram S., Xin Z.Y., Huang G.Z. (2001) A history of wheat breeding in China, CYMMIT, Mexico, D.F.
Huang Y.Y., Deng J.Y. (1988) Preliminary analyses of the effectiveness of utilization of Taigu genetic male-sterile wheat in recurrent selection and complex crossing, in: Miller T.E., Koebner R.M.D. (Eds.), Proc. 7th Int. Wheat Genet. Symp., Cambridge, pp 1105–1108.
Jensen E.F. (1970) A diallel selective mating system for cereal breeding, Crop Sci. 10, 629–635.
Johnson R., Law C.N. (1975) Genetic control of durable resistance to yellow rust (Puccinia striiformis) in the wheat cultivar Hybride de Bersée, Ann. Appl. Biol. 81, 385–391.
Kaur M., Saini R.G., Preet K. (2000) Adult plant leaf rust resistance from 111 wheat (Triticum aestivum L.) cultivars, Euphytica 113, 235–243.
Knott D.R. (1989) The wheat rusts – Breeding for resistance, Springer-Verlag, Berlin.
Kuchel H., Ye G., Fox R., Jefferie S. (2005) Genetic and economic analysis of a targeted marker-assisted wheat breeding strategy, Mol. Breeding 16, 67–78.
Liu J., Liu L., Hou N., Zhang A., Liu L. (2007) Genetic diversity of wheat gene pool of recurrent selection assessed by microsatellite markers and morphological traits, Euphytica 155, 249–258.
Lukaszewski A.J. (2000) Manipulation of the 1RS.1BL translocation in wheat by induced homoeologous recombination, Crop Sci. 40, 216–225.
Maan S.S., Williams N.D. (1984) An EMS-induced dominant allele for male sterility transferred to euplasmic wheat, Crop Sci. 23:1097–852.
Marais G.F., Botes W.C. Louw J.H. (2000) Recurrent selection using male sterility and hydroponic tiller culture in pedigree breeding of wheat. Plant Breeding 119, 440–442.
Marais G.F., Botes W.C., Louw J.H. (2001a) Wheat breeding based on recurrent mass selection, Cereal Res. Commun. 29, 339–342.
Marais G.F., Marais A.S., Groenewald J.Z. (2001b) Evaluation and reduction of Lr19-149, a recombined form of the Lr19 translocation of wheat, Euphytica 121, 289–295.
Marais G.F., Botes W.C. (2003) Recurrent mass selection as a means to pyramid major genes for pest resistance in spring wheat, in: Pogna N.E., Romanò M., Pogna E.A., Galterio G. (Eds.), Proc. 10th Int. Wheat Genet. Symp. Paestrum, Italy, pp. 757–759.
McIntosh R.A. (1992) Close genetic linkage of genes conferring adult-plant resistance to leaf rust and stripe rust in wheat, Plant Pathol. 41, 523–527.
McProud W.L. (1979) Repetitive cycling and simple recurrent selection in traditional barley breeding programs, Euphytica 28, 473–480.
Parlevliet J.E. (1988) Strategies for the utilization of partial resistance for the control of cereal rusts, in: Simmonds N.W., Rajaram S. (Eds.), Breeding strategies for resistance to the rusts of wheat, Mexico, D.F. CIMMYT, pp 48–62.
Parlevliet J.E., van Ommeren A. (1988) Accumulation of partial resistance in barley to barley leaf rust and powdery mildew through recurrent selection against susceptibility, Euphytica 37, 261–274.
Pretorius Z.A., Pakendorf K.W., Marais G.F., Prins R., Komen J.S. (2007) Challenges for sustainable cereal rust control in South Africa, Austr. J. Agric. Res. 58, 1–9.
Rubiales D., Niks R.E. (1995) Characterization of Lr34, a major gene conferring nonhypersensitive resistance to wheat leaf rust, Plant Dis. 79, 1208–1212.
Stoskopf N.C. (1999) Plant breeding: Theory and practice, Scientific Publishers, PO Box 91, Jodhpur, India.
Wallace D.H., Yan W. (1998) Plant breeding and whole-system crop physiology – improving crop maturity, adaptation and yield, CAB International, NY, USA. 390 pp.
Wiersma J.J., Busch R.H., Fulcher G.G., Hareland G.A. (2001) Recurrent selection for kernel weight in spring wheat, Crop Sci. 41, 999–1005.
Wolfe M.S. (1988) The use of variety mixtures to control diseases and stabilize yield, in: Simmonds N.W., Rajaram S. (Eds.), Breeding strategies for resistance to the rusts of wheat. Mexico, D.F. CIMMYT, pp 91–100.
Acknowledgments
We gratefully acknowledge the financial support of the Winter Cereal Trust and National Research Foundation THRIP program.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2009 Springer Science+Business Media B.V.
About this chapter
Cite this chapter
Marais, G., Botes, W. (2009). Recurrent Mass Selection for Routine Improvement of Common Wheat: A Review. In: Lichtfouse, E. (eds) Organic Farming, Pest Control and Remediation of Soil Pollutants. Sustainable Agriculture Reviews, vol 1. Springer, Dordrecht. https://doi.org/10.1007/978-1-4020-9654-9_6
Download citation
DOI: https://doi.org/10.1007/978-1-4020-9654-9_6
Published:
Publisher Name: Springer, Dordrecht
Print ISBN: 978-1-4020-9653-2
Online ISBN: 978-1-4020-9654-9
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)