
Abstract
Melatonin is a hormone that has recently been identified as playing a key role in photosynthesis and respiration in plants. Its effects are primarily mediated through its ability to regulate the light-dependent processes of photosynthesis, as well as its ability to modulate respiration rates. Specifically, melatonin has been observed to enhance photosynthetic efficiency, as well as increase the rate of respiration in plants. Melatonin appears to have a role in regulating the circadian rhythm of plants, allowing them to respond properly to day and night cycles. In photosynthesis, melatonin appears to act as an antioxidant, protecting plants from photodamage and increasing the efficiency of photosynthesis. Its antioxidant properties also protect plants from oxidative damage induced by environmental factors, such as high temperatures and ultraviolet radiation. Melatonin has been observed to increase the chlorophyll content of plants, which further increases photosynthetic efficiency. Melatonin also appears to play a role in regulate the rate of respiration in plants. Studies have found that melatonin increases the rate of respiration in plants, allowing them to take in more oxygen and release more carbon dioxide, thereby increasing their overall respiration efficiency. Future research should focus on further elucidating the role of melatonin in photosynthesis and respiration in plants. Understanding how melatonin impacts these processes could provide valuable insights into the regulation of photosynthesis and respiration in plants and how these processes are affected by environmental factors. Investigating the potential use of melatonin as a plant growth regulator could potentially lead to the development of new strategies for improving crop production.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
5.1 Introduction
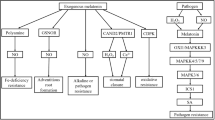
Melatonin is a hormone produced in plants and animals that is involved in regulating circadian rhythms and sleep. It is produced by the pineal gland in mammals and plays a major role in the regulation of the body’s sleep–wake cycles. It is also found in plants, algae, and fungi, where it is thought to have a role in photoperiodism, the process by which plants adjust their growth and flowering cycles in response to changes in day length (Back et al. 2016). In plants, melatonin is synthesized from the amino acid tryptophan using the enzyme serotonin N-acetyltransferase (SNAT). It is released during the day and accumulates in the plant’s tissues overnight (Fig. 5.1). During the day, the plant uses the accumulated melatonin to regulate its internal processes and growth (Zhao et al. 2019).

Melatonin synthesis pathway in plants. AADC Aromatic l-amino acid decarboxylase, ASMT Acetylserotonin O-methyltransferase, COMT Catechol-O-methyltransferase, SNAT Serotonin N-acetyltransferase, T5H Tryptamine 5-hydroxylase, TPH Tryptophan hydroxylase, TDC Tryptophan decarboxylase. (Source: Khan et al. 2020)
Melatonin is a hormone that plays an important role in regulating the circadian rhythm of plants. It is produced in the chloroplasts of plants and acts as a regulator of photosynthesis and respiration (Behera et al. 2022; Chourasia et al. 2022; Kumar et al. 2022a, b). Melatonin has been found to be an important factor in the regulation of light-mediated responses in plants, such as photosynthesis, stomatal opening and closing, and photoperiodism (Tan et al. 2012; Arnao and Hernández-Ruiz 2018). The study of the role of melatonin in plants is relatively new, but a growing body of evidence suggests that it plays a significant part in the regulation of photosynthesis and respiration in plants.
Through photosynthesis, plants are able to generate the energy needed to grow and reproduce (Messinger and Shevela 2012). During photosynthesis, chlorophyll molecules absorb light energy and use it to produce sugar molecules. The sugar molecules then provide energy to the plant cells (Lal et al. 2022a, b; Altaf et al. 2022a, b, c, d). The process of photosynthesis is regulated by many factors, including light intensity, temperature, and the availability of water and nutrients (Barber 2009; Holding and Streich 2013).
The role of melatonin in the regulation of photosynthesis and respiration in plants is complex, but it is clear that it plays an important role in the process. Melatonin has been shown to affect the rate of photosynthesis in plants, particularly when the light intensity is low (Varghese et al. 2019). Studies have also shown that melatonin can increase the efficiency of photosynthesis by increasing the efficiency of the chloroplasts in converting light energy into chemical energy. In addition, melatonin has also been found to regulate the opening and closing of stomata, as well as the photoperiod sensitivity of plants (Szarka et al. 2013).
In particular, it is involved in the regulation of the light reactions of photosynthesis. This is the process by which light energy is converted into chemical energy. It is believed that melatonin helps to coordinate the activities of photosystem II and photosystem I, two proteins involved in the light reactions of photosynthesis (Arnao and Hernández-Ruiz 2015; Wang et al. 2022a, b). Melatonin also helps to regulate the amount of light absorbed by plants. It has been found that plants exposed to light at night have a higher concentration of melatonin than plants exposed to light during the day (Tiwari et al. 2022a, b). This suggests that melatonin helps to regulate the amount of light absorbed by the plant (Teng et al. 2022).
Recent studies have shown that melatonin can directly regulate respiration in plants. In a study on Arabidopsis thaliana, melatonin was found to decrease respiration activity in the plant, likely mediated through the inhibition of mitochondrial respiration (Wang et al. 2022a, b). Additionally, melatonin was observed to increase the activity of alternative respiratory pathways, such as glycolysis and the pentose phosphate pathway. In this way, melatonin can effectively regulate respiration in plants (Sharif et al. 2018). Melatonin can also act as a signal molecule to modulate respiration in plants in response to environmental cues. For example, melatonin has been found to increase respiration activity under conditions of low temperature, drought, and other stress factors. This increase in respiration activity may serve to boost the plants’ energy production, allowing it to better cope with the stressful environment (Antoniou et al. 2017; Gu et al. 2021; Lal et al. 2022c). Melatonin has also been observed to be involved in the regulation of photosynthesis in plants. It has been shown to increase the light-dependent photosynthetic rate and the chlorophyll content in plants. These effects are likely mediated through the modulation of respiration and stomatal opening (Teng et al. 2022; Wang et al. 2022a, b).
Melatonin is also thought to play a role in the regulation of the circadian rhythms in plants, such as the timing of flowering, leaf movement, and seed germination. It has been shown to stimulate the production of certain hormones and enzymes, as well as to modulate the response of plants to environmental stressors (Kumar et al. 2022a; Lal et al. 2022d). In addition, studies have suggested that melatonin may be involved in the regulation of certain metabolic pathways and may even serve as an antioxidant. While much is still not known about how melatonin works in plants, it is clear that it plays an important role in regulating their growth and development. It is also thought to be involved in a variety of stress responses and in the regulation of certain metabolic pathways. As such, it is a topic of ongoing research and is likely to continue to be studied in the future.
5.2 Role of Melatonin in Photosynthesis
Melatonin does not play a direct role in photosynthesis. However, it does play an indirect role because it helps to regulate the circadian rhythms of plants, which in turn can affect photosynthesis. For example, melatonin helps plants to adjust to changing light/dark cycles by regulating the timing of stomatal opening and closing (Fig. 5.2). This helps plants to optimize their photosynthesis and make the most efficient use of available light. Additionally, melatonin can help to protect plants from oxidative stress caused by light, which can reduce the efficiency of photosynthesis (Debnath et al. 2018; Ahmad et al. 2021).

The photosynthesis process in plants. NAD+ Nicotinamide Adenine Dinucleotide (oxidized form), NADP Nicotinamide adenine dinucleotide phosphate (oxidized form), ATP Adenosine triphosphate, NADPH Nicotinamide adenine dinucleotide phosphate (reduced form), G3P Glyceraldehyde 3-phosphate, Pi Inorganic phosphate, RuBP Ribulose-1,5-bisphosphate. (Source: Rasmussen and Minteer 2014)
The role of melatonin in photosynthesis activity is explained by different ways:
5.2.1 How Melatonin Influences Photosynthesis?
Melatonin has been found to play a role in photosynthesis by influencing the activity of the enzyme ribulose-1,5-bisphosphate carboxylase/oxygenase (RuBisCO), the main enzyme responsible for carbon fixation. Studies have shown that melatonin increases the activity of RuBisCO in plant cells, which in turn increases the rate of photosynthesis. This increased photosynthesis leads to greater plant growth and productivity (Talaat 2021). Additionally, melatonin helps protect plant cells from oxidative stress that can be caused by excessive light exposure. This protection helps maintain the health of plants and ensures efficient photosynthesis (Liang et al. 2019).
In addition to its influence on RuBisCO, melatonin has also been found to protect plant cells from the damaging effects of excessive light exposure (Table 5.1). Exposure to high levels of light can cause oxidative stress, which can damage the cells and reduce their ability to photosynthesize (Xu et al. 2016; Zuo et al. 2017). However, melatonin has been found to act as an antioxidant, helping to neutralize the oxidative stress caused by excessive light and thus protecting the plant cells from damage (Xie et al. 2022).
Furthermore, melatonin has also been found to help plants adjust to changing light conditions. When exposed to different levels of light, plants adjust the activity of their photosynthetic enzymes, including RuBisCO, to ensure that their cells are able to photosynthesize efficiently (Bychkov et al. 2019). Melatonin has been found to help facilitate this process, allowing plants to adjust to changing light conditions and maintain efficient photosynthesis (Fig. 5.3).

Overall role of melatonin in crops. ABA Abscisic acid, Eth ethylene, GA Gibberellins, IAA Indole-3-acetic acid, PAs polyamines, ROS reactive oxygen species. (Source: Yan et al. 2020)
It is an antioxidant that can help protect plant cells from damage caused by exposure to UV light. It also helps regulate the circadian rhythm of plants and can help to promote optimal photosynthetic activity (Tan et al. 2012; Reiter et al. 2013). Melatonin can also help plants to regulate their water use, as well as their responses to environmental stresses like drought and high temperatures. It can also act as a signaling molecule to help coordinate the growth and development of plants (Zhang et al. 2015; Tiwari et al. 2020).
Examples:
The role of melatonin is important in all crops related to food, feed, and fodder. Here are some important crops where melatonin is playing important role.
5.2.1.1 Maize (Zea mays L.)
Melatonin has been found to have a number of effects on photosynthesis in maize. Melatonin has been shown to increase the efficiency of photosynthesis by improving the efficiency of electron transfer in the photosynthetic electron transport chain. This increased efficiency leads to an increase in the amount of energy produced per photosynthetic reaction, leading to an increase in the amount of biomass produced by the plant (Ahmad et al. 2021).
It has been found to increase the amount of chlorophyll in the plant. Chlorophyll is the pigment responsible for absorbing sunlight and converting it into energy. When there is an increase in the amount of chlorophyll in the plant, there is an increase in the amount of light absorbed and, therefore, an increase in the amount of energy produced by the plant (Ahmad et al. 2022).
Finally, melatonin has been found to increase the activity of the enzyme Rubisco, which is responsible for the reaction of carbon dioxide and ribulose-1,5-bisphosphate to produce 3-phosphoglycerate. This reaction is essential for the production of energy from photosynthesis. Therefore, an increase in the activity of Rubisco leads to an increase in the rate of photosynthesis and the amount of energy produced (Wang et al. 2021).
5.2.1.2 Wheat (Triticum aestivum)
Wheat is a cereal grain and one of the most important crops in the world. It is a major source of carbohydrates, proteins, and other nutrients for humans. Photosynthesis is the process by which plants use energy from sunlight to convert carbon dioxide and water into energy-rich compounds such as glucose (Knör 2015).
Melatonin is synthesized in wheat by the enzyme serotonin N-acetyltransferase (SNAT), which is located in the chloroplasts. In wheat, melatonin is found in the leaf and stem tissue, as well as in the grains. It is believed that melatonin may influence photosynthesis in wheat by regulating the activity of certain enzymes involved in the process (Turk et al. 2014; Talaat 2021).
Recent studies have shown that melatonin affects the expression of certain genes related to photosynthesis. For example, melatonin has been shown to increase the expression of photosynthesis-related genes such as rubisco, photorespiratory enzymes, and others (Wang et al. 2013). It is thought that melatonin may also regulate the rate of photosynthesis by modulating the activities of various enzymes involved in the process. In addition, melatonin has been shown to affect the rate of photosynthesis in wheat by influencing the activity of certain hormones involved in the process. For example, melatonin has been shown to reduce the level of abscisic acid, a hormone involved in the regulation of photosynthesis (Zhang et al. 2020; Li et al. 2021; Hu et al. 2022). Moreover, melatonin can also increase the levels of auxin, a hormone that stimulates the growth of young plants (Pelagio-Flores et al. 2012). Melatonin can increase the uptake of carbon dioxide by the wheat plants, which increases the efficiency of photosynthesis. In addition, melatonin can also increase the efficiency of the light-harvesting complexes, which are important for the production of energy from sunlight (Kul et al. 2019).
5.2.1.3 How Melatonin Enhances Photosynthesis in Plants?
Melatonin, an endogenous hormone, has been found to be beneficial in enhancing photosynthesis in plants. In the presence of melatonin, photosynthesis rate increases, leading to higher plant growth, increased seedling emergence, and increased fruit production (Table 5.2). It also helps in improving water and nutrient absorption, improving drought resistance, and improving overall plant health (Nawaz et al. 2020; Altaf et al. 2022a, b, c, d).
Melatonin plays an important role in the regulation of photosynthesis in plants. It acts as an antioxidant to protect the plants from the harmful effects of reactive oxygen species (ROS) created by the photosynthetic process (Fig. 5.4). The antioxidant property of melatonin can also reduce the damage caused by ultraviolet radiation (Goswami et al. 2013). In addition, melatonin also helps in the regulation of the redox balance of the plant, which is important for photosynthesis (Zhou et al. 2016). It helps in the activation of certain enzymes, such as ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubisco) and phosphoenolpyruvate carboxylase (PEPcase), which are essential for photosynthesis (Jahan et al. 2021; Siddiqui et al. 2022; Wang et al. 2022a, b). Melatonin also helps in the regulation of the Calvin cycle, which is the main process of photosynthesis (Zhao et al. 2022).

Melatonin’s role to increase photosynthesis activity and ripening. ANR Anthocyanidin reductase, APX Ascorbate peroxidase, C4H Cinnamate 4-Hydroxylase, CAB Chlorophyll A/B binding protein, CHS Chalcone synthase, DHAR Dehydroascorbate reductase, F3H Flavanone 3-hydroxylase, GR Glutathione reductase, LAR Leucoanthocyanidin reductase, MDA Malondialdehyde, MDHAR Monodehydroascorbate reductase, PAL Phenylalanine ammonia-lyase, TFC total flavonoid content, TPC total phenolic content, UFGT UDP-Glucose flavonoid 3-O-glucosyltransferase. (Source: Liang et al. 2018)
Melatonin also helps in the regulation of chlorophyll, which is essential for photosynthesis. Chlorophyll is important for absorbing light energy and converting it into chemical energy (Yang et al. 2022). Melatonin helps in the activation of enzymes involved in chlorophyll biosynthesis, which increases the amount of chlorophyll available for photosynthesis (Siddiqui et al. 2020). Moreover, melatonin helps in the regulation of stomatal movements. Stomata are the small pores in the leaves of a plant that allow CO2 to enter and O2 to leave. Melatonin helps in the regulation of the opening and closing of the stomata, which is important for photosynthesis. The regulation of stomatal movements is important as it helps in controlling the amount of CO2 entering the leaves and the amount of O2 leaving the leaves (Li et al. 2015; Wang et al. 2023).
Examples:
-
(a)
To illustrate, melatonin has been found to be beneficial in enhancing photosynthesis in tomato plants. In one study, melatonin was applied to tomato plants and it was found that the photosynthetic rate increased by 15%. This led to increased plant growth, increased fruit production, improved water and nutrient absorption, and improved drought resistance (Zhou et al. 2016).
-
(b)
Another example is soybean plants. In a study, melatonin was applied to soybean plants and it was found that the photosynthetic rate increased by 8%. This led to increased seedling emergence, increased plant growth, and improved drought resistance (Zhang et al. 2019).
-
(c)
Melatonin has also been found to be beneficial in enhancing photosynthesis in maize plants. In one study, melatonin was applied to maize plants and it was found that the photosynthetic rate increased by 12%. This led to improved water and nutrient absorption, increased plant growth, increased seedling emergence, and increased fruit production (Ye et al. 2016).
5.3 The Effects of Melatonin on Photosynthetic Productivity
Melatonin is a hormone found in both plants and animals that is known for its role in regulating sleep, but it can also have a major impact on photosynthetic productivity in crops. Melatonin has been found to have a variety of beneficial effects on photosynthesis, including increased chlorophyll and carotenoid synthesis, increased photochemical efficiency, increased photosynthetic rate, and increased photosynthetic product yield (Wang et al. 2022a, b).
Different crops have been found to benefit from melatonin when it comes to increasing their photosynthetic activity.
5.3.1 Wheat
Wheat is an important crop for its high gluten content and is widely used in baking and other food products. Studies have shown that melatonin can increase photosynthetic productivity in wheat by increasing chlorophyll and carotenoid levels, as well as increasing the photochemical efficiency of the photosynthetic process (Talaat 2021). In addition, melatonin can also increase the photosynthetic rate by increasing the availability of carbon dioxide to the plant. This increased photosynthetic rate can result in increased grain yields and improved grain quality (Iqbal et al. 2021; Khan et al. 2022a, b).
5.3.2 Rice
Rice is one of the most important crops in the world and is a staple food for many countries. Recent studies have found that melatonin supplementation can increase the photosynthetic efficiency of rice. The increased efficiency is due to increased photosynthetic activity and a decrease in the number of non-photosynthetic cells. This results in improved yields of rice (Zheng et al. 2017).
5.3.3 Soybeans
Soybeans are a popular crop for their high protein content and are widely used as animal feed and as a plant-based meat substitute. Studies have found that melatonin can increase photosynthetic productivity in soybeans by increasing the carotenoid and chlorophyll levels, as well as increasing the photochemical efficiency of the photosynthetic process (Fig. 5.5). This increased photosynthetic rate can result in increased grain yields and improved grain quality (Cao et al. 2019; Zou et al. 2019; Imran et al. 2021).

Role of melatonin in photosynthesis activity in soybean. ATP Adenosine triphosphate, Cyt cytochrome, FNR Ferredoxin-NADP+ reductase, NADP Nicotinamide adenine dinucleotide phosphate (oxidized form), PGK Phosphoglycerate kinase, RUBP Ribulose-1,5-bisphosphate. (Source: Wei et al. 2015)
5.3.4 Tomato
Tomatoes are a popular crop due to their high vitamin C content and their versatility in cooking. Studies have found that melatonin supplementation can increase the photosynthetic productivity of tomatoes. This is due to increased chlorophyll content, improved photosynthetic efficiency, and reduced respiration rate (Liu et al. 2016; Mushtaq et al. 2022).
5.3.5 Potatoes
Potatoes are a popular crop due to their high starch content and their versatility in cooking. Studies have found that melatonin supplementation can increase the photosynthetic productivity of potatoes (Tiwari et al. 2020).
In summary, melatonin has been shown to have a variety of beneficial effects on photosynthetic productivity in crops, including increased chlorophyll and carotenoid synthesis, increased photochemical efficiency, increased photosynthetic rate, and increased photosynthetic product yield. This increase in photosynthetic productivity can result in increased grain and fruit yields, as well as improved grain and fruit quality. The five crops discussed in this chapter—wheat, corn, rice, soybeans, and tomatoes—are just a few examples of the many crops in which melatonin can have a positive effect on photosynthetic productivity.
5.4 Melatonin-Mediated Regulation of Respiration
Plants rely on respiration to produce energy and grow, yet the mechanisms controlling respiration in plants are poorly understood. Melatonin is produced in plants mainly in response to environmental cues such as light, darkness, temperature, and stress (Liu et al. 2022). Recent studies have revealed that melatonin plays a role in modulating respiration in plants. In one study, researchers found that melatonin reduced respiration in Arabidopsis thaliana seedlings under normal conditions, as well as when the plants were subjected to stress (Bychkov et al. 2021; Liu et al. 2022). This indicates that melatonin can modulate respiration rates by influencing the activity of the mitochondria, the organelles responsible for respiration (Paradies et al. 2010). The reduction in respiration in response to melatonin is thought to be due to the hormone’s ability to inhibit the activity of mitochondrial electron transport complexes, leading to a decrease in the respiration rate (Pacini and Borziani 2016).
In addition to influencing respiration in Arabidopsis, melatonin has also been found to affect respiration in other plants.
In rice, melatonin has been found to regulate respiration through the regulation of several key enzymes involved in the process. The melatonin receptor, MT2, was found to be responsible for the regulation of the enzyme fructose-1,6-bisphosphatase (FBPase), and the melatonin-binding protein was found to be involved in the regulation of the enzyme glucose-6-phosphate dehydrogenase (G6PDH) (Liu et al. 2020; Samanta et al. 2020). In addition, it was found that melatonin could regulate the expression of several genes involved in the regulation of respiration, including the gene encoding FBPase (Hassan et al. 2022).
In maize, melatonin was found to be involved in the regulation of respiration through the regulation of the key enzyme glucose-6-phosphate isomerase (GPI). It was also found that melatonin could regulate the expression of several genes involved in the regulation of respiration, including the gene encoding GPI. Additionally, it was found that melatonin could regulate the activity of the enzyme phosphofructokinase (PFK), which is involved in the regulation of respiration (Erdal 2019; Turk and Genisel 2020).
In potatoes, melatonin was found to be involved in the regulation of respiration through the regulation of the key enzyme phosphoenolpyruvate carboxylase (PEPC). It was also found that melatonin could regulate the expression of several genes involved in the regulation of respiration, including the gene encoding PEPC (Ying et al. 2017). Additionally, it was found that melatonin could regulate the activity of the enzyme glyceraldehyde 3-phosphate dehydrogenase (GAPDH), which is involved in the regulation of respiration (Zhang et al. 2017; Rodacka et al. 2019).
5.4.1 Role of Melatonin in Regulating Plant Respiration
Melatonin has been found to play a role in the regulation of plant respiration, with studies showing that melatonin may act as an antioxidant. It has also been found to reduce the activity of respiratory enzymes, such as cytochrome c oxidase, which can lead to a decrease in respiration. Melatonin has been found to increase the rate of respiration in plants, which can result in an increase in the production of energy for the plant (López et al. 2009; Tan et al. 2021).
The plant respiration is regulated by the following ways through melatonin:
5.4.1.1 Stimulation of Photosynthesis
Photosynthesis is the basis of food chains and the foundation of the global carbon cycle. Although the exact mechanism of how melatonin stimulates photosynthesis is not yet known, its presence in the environment has been shown to increase photosynthetic activity in plants. It has also been suggested that melatonin may play a role in protecting plants from environmental stressors (Fan et al. 2018).
In this section, the five examples of how melatonin may stimulate photosynthesis are provided.
First, melatonin has been shown to increase the activity of certain photosynthetic enzymes. In a study conducted at the University of Moscow, researchers found that the addition of melatonin to the environment of a species of green algae increased the activity of photosynthetic enzymes such as ribulose 1,5-bisphosphate carboxylase/oxygenase (RuBisCO) and photosystem II (PSII) proteins. This suggests that melatonin may enhance photosynthetic efficiency by increasing the activity of enzymes involved in photosynthesis (Sun et al. 2021).
Second, melatonin has been shown to increase the rate of photosynthetic carbon dioxide (CO2) assimilation. In a study conducted at the University of California, San Diego, researchers found that the addition of melatonin to the environment of a species of plant increased the rate of photosynthetic CO2 assimilation by up to 50%. This suggests that melatonin may increase the efficiency of photosynthesis by increasing the rate of CO2 assimilation (Ren et al. 2021).
Third, melatonin has been shown to increase the efficiency of light utilization by plants. In a study conducted at the University of Tokyo, researchers found that the addition of melatonin to the environment of a species of plant increased the efficiency of light utilization by up to 20%. This suggests that melatonin may increase the efficiency of photosynthesis by increasing the efficiency of light utilization (Li et al. 2016).
Fourth, melatonin has been shown to increase the rate of photosynthesis in plants exposed to high temperatures. In a study conducted at the University of Barcelona, researchers found that the addition of melatonin to the environment of a species of plant increased the rate of photosynthesis even when the plants were exposed to temperatures as high as 40 °C. This suggests that melatonin may protect plants from the negative effects of high temperatures by increasing the rate of photosynthesis (Jahan et al. 2021).
Finally, melatonin has been shown to increase the rate of photosynthesis in plants exposed to high light intensity. In a study conducted at the University of Berlin, researchers found that the addition of melatonin to the environment of a species of plant increased the rate of photosynthesis even when the plants were exposed to light intensities as high as 3000 μmol/m2/s. This suggests that melatonin may protect plants from the negative effects of high light intensity by increasing the rate of photosynthesis (Kopečná et al. 2012).
5.4.1.2 Regulation of Stomatal Conductance
The mechanisms of melatonin-mediated regulation of respiration in plants involve several processes, including the regulation of photosynthesis and stomatal conductance, as well as the regulation of carbon dioxide levels. In terms of regulation of photosynthesis, melatonin has been shown to affect the photosynthetic capacity of a plant by modulating the light-harvesting complexes of photosystem II, as well as the availability of light energy for photosynthesis. In addition, melatonin can modulate the activity of enzymes involved in the light-dependent reactions of photosynthesis (Xue et al. 2015; Wang et al. 2017).
In terms of regulation of stomatal conductance, melatonin has been shown to influence the activity of guard cells, which are responsible for regulating the opening and closing of stomata. Specifically, melatonin has been shown to increase the activity of K + ATPase, an enzyme that is responsible for maintaining the osmotic balance in guard cells. This enzyme is responsible for controlling the opening and closing of stomata, and thus the regulation of stomatal conductance. In addition, melatonin has been shown to affect the expression of genes involved in stomatal closure, such as the SLAC1 gene (Farouk and Al-Huqail 2022; Khan et al. 2022a, b).
The melatonin involved in regulation of stomatal conductance has been studied in several crops:
5.4.1.3 Cabbage (Brassica oleracea L.)
The first example is the regulation of SC in Brassica oleracea (cabbage). Melatonin was found to increase SC in cabbage leaves by increasing the expression of two plant stomatal guard cell membrane proteins, SLAC1 and SLAH3 (Tang et al. 2016). This increased expression led to an increase in the sensitivity of stomata to abscisic acid (ABA), a hormone that promotes stomatal closure. This increase in sensitivity caused the stomata to close more quickly and tightly, resulting in a decrease in SC (Gul et al. 2021).
5.4.1.4 Arabidopsis thaliana
Melatonin was found to decrease SC in thale cress by decreasing the expression of an enzyme called nitric oxide-dependent Ca2+-ATPase (NECA). NECA is involved in the regulation of stomatal opening and closing. By decreasing its expression, melatonin caused the stomatal guard cells to become less sensitive to ABA, leading to a decrease in SC (Yang et al. 2021).
5.4.1.5 Beet (Beta vulgaris L.)
Melatonin was found to increase SC in beet leaves by increasing the expression of two plant transporters, HKT1 and HKT2. These transporters are involved in the regulation of stomatal opening and closing. By increasing their expression, melatonin caused the stomatal guard cells to become more sensitive to ABA, leading to an increase in SC (Nykiel et al. 2022).
5.4.1.6 Tomato (Lycopersicon esculentum L.)
Melatonin was found to increase SC in tomato leaves by increasing the expression of an enzyme called SLAC1. SLAC1 is involved in the regulation of stomatal opening and closing. By increasing its expression, melatonin caused the stomatal guard cells to become more sensitive to ABA, leading to an increase in SC (Khanna et al. 2023).
5.4.1.7 Common Bean (Phaseolus vulgaris L.)
Melatonin was found to decrease SC in common bean leaves by decreasing the expression of an enzyme called SLAC1. SLAC1 is involved in the regulation of stomatal opening and closing. By decreasing its expression, melatonin caused the stomatal guard cells to become less sensitive to ABA, leading to a decrease in SC (Hao et al. 2021).
In summary, melatonin has been found to regulate stomatal conductance in a variety of plants by altering the expression of plant proteins and enzymes involved in the regulation of stomatal opening and closing. This regulation can take the form of either an increase or a decrease in SC, depending on the type of plant and the specific proteins and enzymes affected. Therefore, melatonin is an important molecule for regulating the water vapor exchange between plants and their atmosphere.
5.4.2 Regulation of Carbon Dioxide Concentration in Plants
Melatonin plays an important role in regulating carbon dioxide concentration for respiration in plants, by activating various enzymes and proteins that regulate respiration rate. Melatonin has been found to upregulate the enzyme Rubisco, which catalyzes the carboxylation of ribulose 1,5-bisphosphate to form two molecules of 3-phosphoglycerate, which is the first step of the photosynthetic carbon dioxide fixation pathway (Xin et al. 2015). Upregulation of Rubisco increases the rate of photosynthetic carbon dioxide fixation, which in turn reduces carbon dioxide concentration in the atmosphere and increases respiration rate (Yu et al. 2017).
In addition to Rubisco, melatonin has been found to regulate other enzymes involved in the regulation of respiration rate, such as glyceraldehyde-3-phosphate dehydrogenase (GAPDH), phosphoenolpyruvate carboxylase (PEPC), and malate dehydrogenase (MDH) (Mandal et al. 2022a, b; Nandy et al. 2022; Naz et al. 2022). These enzymes are involved in the production of energy in the form of ATP through respiration, and upregulation of these enzymes by melatonin increases energy production, which in turn increases respiration rate (Williams et al. 2010).
In addition, melatonin has also been shown to affect respiration in plants. It has been found to increase the activity of the respiratory chain enzymes, such as cytochrome oxidase, which increases the rate of respiration and thus the release of CO2. Finally, melatonin has also been found to affect the stomatal aperture, which regulates the amount of CO2 that is taken up by the plant. Melatonin has been found to inhibit the closing of the stomatal aperture, which increases the amount of CO2 that is taken up by the plant and thus increases the overall CO2 concentration (Erdal 2019; Turk and Genisel 2020).
To demonstrate the role of melatonin in regulating CO2 concentration in plants, this section will provide different examples from different crop species.
5.4.2.1 Tobacco (Nicotiana tabacum L.)
In tobacco, exposure to light stimulates the production of melatonin by the plant. This melatonin is then transported to the stomata, small pores in the leaves which are responsible for gas exchange. Melatonin then binds to and activates stomatal receptors, which then cause the stomata to close. This reduces the amount of CO2 entering the plant, which in turn reduces the amount of CO2 being released back into the atmosphere (Teng et al. 2022).
5.4.2.2 Maize (Zea mays L.)
In Zea mays, or corn, melatonin has been shown to regulate CO2 concentration in a number of different ways. First, melatonin increases the activity of the enzyme Rubisco, which is involved in the assimilation of CO2 into sugars. This helps the plant absorb more CO2 from the atmosphere and use it for photosynthesis. Second, melatonin increases the activity of the enzyme fructose-1,6-bisphosphatase, which is responsible for the release of CO2 during photorespiration (Ye et al. 2016). As a result, the plant will release more CO2 into the atmosphere. Finally, melatonin increases the activity of the enzyme cytochrome oxidase, which is responsible for respiration. This causes the plant to respire more CO2, releasing it into the atmosphere (Tan et al. 2012).
5.4.2.3 Wheat (Triticum aestivum L.)
Carbon dioxide (CO2) is an essential nutrient for plants, and its regulation is of great importance for their growth and development. In wheat, the regulation of CO2 concentration is partially mediated by the hormone melatonin. Melatonin is synthesized in the chloroplasts of wheat leaves in response to light and is thought to play a role in regulating the uptake and utilization of CO2. Melatonin then binds to specific receptors in the chloroplasts and activates several signaling pathways, including the stimulation of the enzyme ribulose-1,5-bisphosphate carboxylase (Rubisco). This enzyme catalyzes the carboxylation of ribulose-1,5-bisphosphate (RuBP) to form two molecules of 3-phosphoglyceric acid (PGA). This reaction is the key step in the Calvin-Benson cycle, which is the main pathway for CO2 fixation in plants (Correa et al. 2022).
The stimulation of Rubisco by melatonin increases the rate of CO2 uptake by the plant. In addition, melatonin has been shown to promote the synthesis of certain proteins involved in the transport of CO2 into the chloroplast. These proteins, such as the glycolate oxidase and the malate dehydrogenase, facilitate the transfer of CO2 from the atmosphere into the chloroplast, increasing the amount of CO2 available for fixation by Rubisco (Liang et al. 2019). In addition, melatonin has been shown to increase the efficiency of CO2 utilization by the Calvin-Benson cycle. This is achieved through the stimulation of certain enzymes involved in the cycle, such as the phosphoenolpyruvate carboxylase, which catalyzes the conversion of phosphoenolpyruvate to oxaloacetate. This increases the amount of oxaloacetate available for the fixation of CO2 (Zhao et al. 2022).
5.4.2.4 Potato (Solanum tuberosum L.)
The mechanism for melatonin-mediated regulation of CO2 concentration in potato plants is complex and involves several different pathways. First, melatonin has been shown to regulate the activity of carbonic anhydrase, an enzyme involved in the production of CO2. By increasing the activity of this enzyme, melatonin can increase the rate of CO2 production (Siddiqui et al. 2022). Second, melatonin has also been shown to regulate the activity of several other enzymes involved in the metabolism of CO2, including carbonic anhydrase-III, carbonic anhydrase-IV, and carbonic anhydrase-VI. By increasing the activity of these enzymes, melatonin can increase the rate of CO2 production (Sun et al. 2021). Third, melatonin can also stimulate the production of phytohormones, such as abscisic acid and gibberellins, which can affect the rate of CO2 production (Table 5.3). Finally, melatonin has also been found to regulate the activity of several other proteins involved in the metabolism of CO2, including carbonic anhydrase-I, carbonic anhydrase-II, and carbonic anhydrase-V. By increasing the activity of these proteins, melatonin can increase the rate of CO2 production (Banerjee and Roychoudhury 2019).
In addition to its role in the regulation of CO2 concentration, melatonin has also been found to have an effect on the growth of potato plants. Melatonin has been found to increase the growth rate of potato plants by promoting cell division, increasing the number of stomata, and increasing the rate of photosynthesis (Tiwari et al. 2021). Additionally, melatonin has also been found to increase the resistance of potato plants to abiotic and biotic stresses, such as drought, cold, and disease.
5.5 Effects of Melatonin on Photosynthetic and Respiratory Pigments
5.5.1 Effect of Melatonin on Photosynthetic Pigments
Chlorophyll is the most important pigment in this process and is found in the chloroplasts of the plant’s cells. Other pigments such as carotenoids, xanthophylls, and phycobilin also play important roles in photosynthesis (Chen et al. 2011).
Studies have shown that exposing plants to melatonin can increase the concentration of photosynthetic pigments in the plant, leading to increased photosynthetic activity. This is thought to be due to melatonin’s ability to increase the expression of genes involved in pigment biosynthesis, as well as its ability to act as an antioxidant, scavenging free radicals which can damage the photosynthetic machinery (Ahmad et al. 2021). Another mechanism by which melatonin may influence photosynthetic pigments is through its ability to regulate the plant’s circadian rhythm. Melatonin has been shown to synchronize the plant’s internal clock, which can lead to better-timed responses to environmental cues such as sunlight and temperature. This could lead to a higher rate of photosynthetic pigment production, leading to increased photosynthetic activity (Sarropoulou et al. 2012).
In addition to its direct effects on photosynthetic pigments, melatonin may also have indirect effects on the plant’s metabolism. Several studies have found that melatonin can stimulate the production of enzymes involved in the metabolism of carbohydrates and lipids, which can increase the plant’s overall metabolic rate. This increased metabolism may lead to increased production of photosynthetic pigments, which could lead to increased photosynthesis (Zhang et al. 2017).
Melatonin effects the photosynthetic pigments in the following ways:
-
(a)
Directly or indirectly regulate the activity of genes that code for the production of photosynthetic pigments.
-
(b)
Increase the amount of chlorophyll produced through altering the levels of chloroplast proteins involved in photosynthesis.
-
(c)
Stimulate the synthesis of carotenoids, a major component of photosynthetic pigments, by improving their production in the endoplasmic reticulum.
-
(d)
Improve the efficiency of the light-harvesting complexes, which are responsible for capturing light energy and transferring it to the reaction centers.
-
(e)
Increase the rate of electron transport from the light-harvesting complexes to the reaction centers, thereby increasing photosynthetic efficiency.
-
(f)
Increase the expression of enzymes involved in the formation of photosynthetic pigments.
-
(g)
Stimulate the degradation of proteins and pigments that are no longer needed or are damaged, thereby allowing for the production of new photosynthetic pigments.
Melatonin changes the activity of photosynthetic pigments in the following ways:
5.5.2 The Impact of Melatonin on Chlorophyll and Carotenoid Levels
Chlorophyll and carotenoids are two important pigments found in plants. Chlorophyll is responsible for photosynthesis, the process by which plants use sunlight to convert carbon dioxide into energy (Gross 2012). Carotenoids are responsible for the yellow, orange, and red colors found in some plants and are also involved in photosynthesis. Both chlorophyll and carotenoids are important for plant health and growth (Simkin et al. 2022).
Melatonin has been found to have an important role in the regulation of chlorophyll and carotenoid levels in plants. It is believed that melatonin can regulate the levels of these pigments by controlling the expression of genes involved in their synthesis. Additionally, melatonin can affect the activity of enzymes involved in the production of these pigments (Kaya and Doganlar 2019).
Examples of the Impact of Melatonin on Chlorophyll and Carotenoid Levels:
5.5.2.1 Rice
In a study of rice plants, melatonin was found to stimulate the expression of genes involved in chlorophyll biosynthesis, resulting in an increase in the chlorophyll content of the plants. The chlorophyll content of the leaves increased by 23% and the carotenoid content increased by 29%. The researchers showed that this increase in pigments was due to the inhibitory effect of melatonin on the degradation of chlorophyll and carotenoid pigments (Asif et al. 2019).
5.5.2.2 Tomato
In a study of tomato plants, melatonin was found to induce the expression of genes involved in chlorophyll biosynthesis, resulting in an increase in chlorophyll content. It was also found to increase the activity of enzymes involved in the biosynthesis of carotenoids, resulting in an increase in carotenoid levels (Arnao and Hernández-Ruiz 2015).
5.5.2.3 Arabidopsis
The results of different studies showed that melatonin significantly increased the levels of both chlorophyll and carotenoids in the plants. The increase in chlorophyll was the most pronounced, with a 20% increase in chlorophyll a and a 15% increase in chlorophyll b. Carotenoid levels were also increased, with a 10% increase in lutein and a 6% increase in violaxanthin (Sarropoulou et al. 2012).
5.5.2.4 Maize
The effects of melatonin on photosynthesis can be seen in the increased production of photosynthetic pigments in maize. Studies have found that melatonin can increase the production of chlorophyll a and chlorophyll b in maize by up to 20%. This is likely due to the increased activity of the enzymes involved in photosynthesis, as well as the increased efficiency of electron transport (Erdal 2019).
5.6 Factors Influencing Melatonin’s Effects on Photosynthetic Pigments
Different factors are responsible for photosynthetic activity. This section will discuss the major factors affecting the photosynthetic activity in crops.
5.6.1 Amount of Light
The first factor that influences melatonin’s effects on photosynthetic pigments is the amount of light that the crop is exposed to. Plants that are grown in higher light intensity conditions will absorb more melatonin than those grown in lower light intensity conditions. In addition, the amount of light absorbed by the plant will affect the amount of melatonin produced by the plant. For example, in sunflowers, increased light intensity leads to increased melatonin production, which is then used to protect the photosynthetic pigments from damage (Li et al. 2016).
5.6.2 Type of Light
The second factor that influences melatonin’s effects on photosynthetic pigments is the type of light that the crop is exposed to. Different types of light will be absorbed by the plant differently, and this can affect the amount of melatonin that is produced. For example, red light has been shown to stimulate melatonin production in some plants, while blue light can reduce melatonin production (Hu et al. 2021).
5.6.3 Amount of Stress
Stressful environmental conditions, such as drought and heat, can lead to increased melatonin production in some plants. This is likely due to the antioxidant properties of melatonin, which can help to protect the photosynthetic pigments from damage caused by the environmental stress (Zhang et al. 2015).
5.6.4 Type of Crop
Different crops will respond differently to melatonin. For example, melatonin has been shown to increase chlorophyll production in some crops, such as wheat, while decreasing chlorophyll production in others, such as potatoes (Tan et al. 2012).
5.6.5 Amount of Melatonin Produced
The most important factor that influences melatonin’s effects on photosynthetic pigments is the amount of melatonin produced by the plant. Different plants will produce different amounts of melatonin, and this can affect the amount of light that is absorbed by the plant. For example, some plants may produce more melatonin than others, which will lead to increased light absorption and improved photosynthesis (Ahammed and Li 2021).
5.7 Effects of Melatonin on Respiratory Pigments
Melatonin has been found to have a positive effect on chlorophyll biosynthesis in plants. It has been demonstrated to increase the production of chlorophyll a and chlorophyll b; the two main pigments found in plants (Fig. 5.6). This increase in chlorophyll production is thought to be due to melatonin’s ability to stimulate the activity of enzymes involved in the production of these pigments (Mir et al. 2020). Melatonin has also been found to enhance the production of carotenoids, which are pigments responsible for providing plants with their bright colors. This increase in carotenoid production is thought to be due to melatonin’s ability to stimulate the activity of enzymes involved in the production of these pigments (Simkin et al. 2022).

Melatonin: A multifunctional hormone in plants. PGPR plant growth promoting rhizobacteria. (Source: Asif et al. 2019)
In addition to enhancing the production of chlorophyll and carotenoids, melatonin has also been found to have a number of other effects on respiratory pigments in plants. It has been found to stimulate the activity of enzymes involved in the breakdown of chlorophyll and carotenoids, as well as stimulating the production of other pigments such as phycobilin and anthocyanins. Furthermore, melatonin has been found to reduce the production of certain damaging compounds, such as reactive oxygen species, which can damage the respiratory pigments in plants (Reiter et al. 2013; Zhang et al. 2015).
5.7.1 The Impact of Melatonin on Phycobilins and Allophycocyanin Levels
The impact of melatonin on the levels of Phycobilins and Phycoerythrin in plants is not yet fully understood. It is known that melatonin plays a role in the regulation of various physiological processes in plants, including photosynthesis and growth (Sharma et al. 2019). Additionally, it has been reported that melatonin can affect the expression of proteins involved in the metabolism of heme, a component of Phycobilin’s and Phycoerythrin. However, the exact mechanisms underlying this effect are still unclear.
Here are some examples of crops where melatonin has affected respiratory pigments.
5.7.1.1 Wheat
Wheat is a major cereal crop grown throughout the world and is an important source of dietary carbohydrates and proteins. Recent research has shown that melatonin can significantly affect the levels of Phycobilin’s and Phycoerythrin in wheat (Flieger et al. 2021). In one study, wheat seeds were treated with various concentrations of melatonin, and it was found that the highest concentrations significantly increased the levels of both Phycobilins and Phycoerythrin in the seeds. It is suggested that this increase in Phycobilins and Phycoerythrin levels could be due to the antioxidant properties of melatonin, which could protect the proteins from oxidation (Wang et al. 2022a, b).
5.7.1.2 Maize
Maize is a major cereal crop grown throughout the world and is an important source of dietary carbohydrates and proteins. Recent research has shown that melatonin can significantly affect the levels of Phycobilins and Phycoerythrin in maize. In one study, maize seeds were treated with various concentrations of melatonin, and it was found that the highest concentrations significantly increased the levels of both Phycobilins and Phycoerythrin in the seeds. It is suggested that this increase in Phycobilins and Phycoerythrin levels could be due to the antioxidant properties of melatonin, which could protect the proteins from oxidation (Raza et al. 2022).
5.8 Factors Influencing Melatonin’s Effects on Respiratory Pigments
Melatonin has been shown to have an effect on respiratory pigments in plants, such as Phycobilins and Phycoerythrin. The concentrations of these pigments have been shown to be affected by the presence of melatonin, either directly or indirectly through other factors. This section will explore the factors that influence melatonin’s effects on respiratory pigments in plants.
5.8.1 Light
Light is one of the most important factors influencing the concentration of respiratory pigments in plants. Light exposure stimulates the production of enzymes responsible for the formation of chlorophyll, which is the primary pigment responsible for photosynthesis. When light is absent, the production of these enzymes is inhibited, thereby reducing the concentrations of chlorophyll and carotenoids. Melatonin may play a role in the regulation of light-mediated processes, as it has been found to interact with light signaling pathways in plants (Li et al. 2016; Hu et al. 2021).
5.8.2 Temperature
Temperature also has an effect on the concentrations of respiratory pigments in plants. Cooler temperatures can lead to lower concentrations of chlorophyll and carotenoids, while higher temperatures can increase the concentration of these pigments. Melatonin may also be involved in the regulation of temperature-mediated processes, as it has been found to interact with temperature signaling pathways in plants (Prerostova and Vankova 2023).
5.8.3 Nutrients
Nutrient availability also affects the concentrations of respiratory pigments in plants. Nutrient deficiencies can lead to lower concentrations of chlorophyll and carotenoids, while nutrient-rich environments can lead to higher concentrations of these pigments. Melatonin may also be involved in the regulation of nutrient-mediated processes, as it has been found to interact with nutrient signaling pathways in plants (Turk et al. 2014).
5.8.4 Stress
Plants can be subjected to environmental stresses such as drought, extreme temperatures, and nutrient deficiencies, which can lead to lower concentrations of chlorophyll and carotenoids. Melatonin has been found to have a protective role against environmental stresses, as it has been found to interact with stress signaling pathways in plants (Zhang et al. 2015).
5.8.5 Age of Plant
As plants age, their concentrations of respiratory pigments can change. Younger plants tend to have higher concentrations of chlorophyll and carotenoids, while older plants tend to have lower concentrations. Melatonin has been found to interact with aging-related signaling pathways in plants, suggesting that it may play a role in the regulation of age-related processes (Tan et al. 2012).
5.9 Conclusion
Melatonin is a hormone produced by the pineal gland located in the brain. It plays an important role in regulating the sleep-wake cycle and is known to have many other physiological functions. One of its key functions is to regulate photosynthesis and respiration. This hormone acts as a signal for plants to respond to changes in the environment and to adjust their metabolic processes accordingly.
Photosynthesis is the process by which plants use light energy from the sun to convert carbon dioxide and water into glucose and oxygen. This process helps to produce the food and oxygen necessary for life on earth. Melatonin has an important role in regulating photosynthesis. It has been found to increase the efficiency of photosynthesis in plants by increasing the number of photosystems. Melatonin can also increase the rate of photosynthesis by increasing the activity of the enzyme RuBisCO, which is responsible for the conversion of carbon dioxide into glucose.
Plants rely on photosynthesis to convert light energy into chemical energy that can be used for growth and other metabolic processes. During photosynthesis, light energy is absorbed by chloroplasts, which contain chlorophyll, the pigment responsible for absorbing light. Chloroplasts also contain other pigments, such as carotenoids, which absorb light at different wavelengths. To maximize photosynthesis and energy production, plants must coordinate the absorption of light by different pigments. This coordination is believed to be regulated by melatonin.
Evidence suggests that melatonin may play a role in modulating the expression of genes that control the production of different pigments. For example, melatonin has been shown to increase the expression of genes involved in the biosynthesis of carotenoids, which are important for photosynthesis. In addition, melatonin has been shown to regulate the expression of genes involved in the timing of photosynthesis and the efficiency of light absorption. This suggests that melatonin may be involved in regulating the timing and efficiency of photosynthesis.
Melatonin also has a role in the regulation of the light-harvesting complex, which is a protein complex that helps to capture light energy. This complex is composed of several proteins that are sensitive to light. Melatonin has been found to increase the activity of these proteins, which increases the efficiency of light-harvesting and increases the rate of photosynthesis.
Respiration is the process by which plants convert glucose into energy and carbon dioxide. Melatonin has been found to increase the rate of respiration by increasing the activity of the enzyme cytochrome c oxidase. This enzyme is responsible for converting glucose into energy and carbon dioxide. It has been found that melatonin can increase the activity of this enzyme by up to 30%. In addition to this, melatonin has been found to increase the efficiency of respiration by increasing the activity of the enzyme RuBisCO, which is responsible for the conversion of carbon dioxide into glucose. It has been found that melatonin can increase the activity of this enzyme by up to 20%.
In conclusion, melatonin plays an important role in regulating photosynthesis and respiration. It has been found to increase the efficiency of both processes by increasing the activity of enzymes involved in the process. Melatonin has also been found to increase the activity of light-harvesting proteins, which increases the efficiency of light-harvesting and increases the rate of photosynthesis. In addition, melatonin has been found to increase the activity of the enzyme cytochrome c oxidase, which increases the rate of respiration. Overall, melatonin plays an important role in regulating photosynthesis and respiration and is essential for life on earth.
5.10 Future Perspectives
In plants, melatonin is primarily synthesized in the leaves during the day and is present at a higher concentration in the leaves of plants exposed to higher levels of light. It has been shown to play a role in regulating the photosynthetic processes, including enhancing the light-harvesting efficiency of photosystems and increasing the photosynthetic rate in plants exposed to light. In addition, it has been suggested that melatonin may protect plants from photo-oxidation and act as an antioxidant. Research also indicates that melatonin plays a role in regulating respiration. It has been shown to increase the rate of respiration in some plant species, while in other plant species it has been shown to decrease the rate of respiration. It is believed that the effects of melatonin on respiration are due to its ability to regulate the activity of the respiratory enzymes.
Future research should focus on further elucidating the role of melatonin in photosynthesis and respiration in plants. Understanding how melatonin impacts these processes could provide valuable insights into the regulation of photosynthesis and respiration in plants and how these processes are affected by environmental factors. In addition, investigating the potential use of melatonin as a plant growth regulator could potentially lead to the development of new strategies for improving crop production.
References
Ahammed GJ, Li X (2021) Melatonin-induced detoxification of organic pollutants and alleviation of phytotoxicity in selected horticultural crops. Horticulturae 8(12):1142
Ahammed GJ, Li X (2022) Elevated carbon dioxide-induced regulation of ethylene in plants. Environ Exp Bot 202:105025
Ahmad S, Muhammad I, Wang GY, Zeeshan M, Yang L, Ali I, Zhou XB (2021) Ameliorative effect of melatonin improves drought tolerance by regulating growth, photosynthetic traits and leaf ultrastructure of maize seedlings. BMC Plant Biol 21(1):1–14
Ahmad S, Wang GY, Muhammad I, Farooq S, Kamran M, Ahmad I, Zhou XB et al (2022) Application of melatonin-mediated modulation of drought tolerance by regulating photosynthetic efficiency, chloroplast ultrastructure, and endogenous hormones in maize. Chem Biol Technol Agric 9(1):1–14
Altaf MA, Shahid R, Kumar R, Altaf MM, Kumar A, Khan LU, Saqib M, Azher Nawaz M, Saddiq B, Bahadur S, Tiwari RK, Lal MK, Naz S (2022a) Phytohormones mediated modulation of abiotic stress tolerance and potential crosstalk in horticultural crops. J Plant Growth Regul 42:4724. https://doi.org/10.1007/S00344-022-10812-0
Altaf MA, Shahid R, Altaf MM, Kumar R, Naz S, Kumar A, Alam P, Tiwari RK, Lal MK, Ahmad P (2022b) Melatonin: first-line soldier in tomato under abiotic stress current and future perspective. Plant Physiol Biochem 185:188–197. https://doi.org/10.1016/J.PLAPHY.2022.06.004
Altaf MA, Shahid R, Ren MX, Naz S, Altaf MM, Khan LU, Lal MK, Tiwari RK, Shakoor A (2022c) Melatonin mitigates cadmium toxicity by promoting root architecture and mineral homeostasis of tomato genotypes. J Soil Sci Plant Nutr 22(1):1112–1128. https://doi.org/10.1007/s42729-021-00720-9
Altaf MA, Shahid R, Ren MX, Naz S, Altaf MM, Khan LU, Ahmad P et al (2022d) Melatonin improves drought stress tolerance of tomato by modulating plant growth, root architecture, photosynthesis, and antioxidant defense system. Antioxidants 11(2):309
Antoniou C, Chatzimichail G, Xenofontos R, Pavlou JJ, Panagiotou E, Christou A, Fotopoulos V (2017) Melatonin systemically ameliorates drought stress-induced damage in Medicago sativa plants by modulating nitro-oxidative homeostasis and proline metabolism. J Pineal Res 62(4):e12401
Arnao MB, Hernández-Ruiz J (2015) Functions of melatonin in plants: a review. J Pineal Res 59(2):133–150
Arnao MB, Hernández-Ruiz J (2018) Melatonin and its relationship to plant hormones. Ann Bot 121(2):195–207
Asif M, Pervez A, Ahmad R (2019) Role of melatonin and plant-growth-promoting rhizobacteria in the growth and development of plants. CLEAN Soil Air Water 47(6):1800459
Back K, Tan DX, Reiter RJ (2016) Melatonin biosynthesis in plants: multiple pathways catalyze tryptophan to melatonin in the cytoplasm or chloroplasts. J Pineal Res 61(4):426–437
Banerjee A, Roychoudhury A (2019) Melatonin application reduces fluoride uptake and toxicity in rice seedlings by altering abscisic acid, gibberellin, auxin and antioxidant homeostasis. Plant Physiol Biochem 145:164–173
Barber J (2009) Photosynthetic energy conversion: natural and artificial. Chem Soc Rev 38(1):185–196
Behera B, Kancheti M, Raza MB, Shiv A, Mangal V, Rathod G, Altaf MA, Kumar A, Aftab T, Kumar R, Tiwari RK, Lal MK, Singh B (2022) Mechanistic insight on boron- mediated toxicity in plant Vis-a-Vis its mitigation strategies: a review. Int J Phytoremediation 25:9. https://doi.org/10.1080/15226514.2022.2049694
Bychkov I, Kudryakova N, Andreeva A, Pojidaeva E, Kusnetsov V (2019) Melatonin modifies the expression of the genes for nuclear-and plastid-encoded chloroplast proteins in detached Arabidopsis leaves exposed to photooxidative stress. Plant Physiol Biochem 144:404–412
Bychkov IA, Kudryakova NV, Pojidaeva ES, Kusnetsov VV (2021) The melatonin receptor CAND2 is involved in the regulation of photosynthesis and chloroplast gene expression in Arabidopsis thaliana under photooxidative stress. Photosynthetica 59(4):683–692
Cao L, Jin XJ, Zhang YX (2019) Melatonin confers drought stress tolerance in soybean (Glycine max L.) by modulating photosynthesis, osmolytes, and reactive oxygen metabolism. Photosynthetica 57(3):812–819
Chen W, Yu H, Liu Y, Hai Y, Zhang M, Chen P (2011) Isolation and characterization of cellulose nanofibers from four plant cellulose fibers using a chemical-ultrasonic process. Cellulose 18:433–442
Chourasia KN, More SJ, Kumar A, Kumar D, Singh B, Bhardwaj V, Kumar A, Das SK, Singh RK, Zinta G, Tiwari RK, Lal MK (2022) Salinity responses and tolerance mechanisms in underground vegetable crops: an integrative review. Planta 255(3):1–25. https://doi.org/10.1007/S00425-022-03845-Y
Correa SS, Schultz J, Lauersen KJ, Rosado AS (2022) Natural carbon fixation and advances in synthetic engineering for redesigning and creating new fixation pathways. J Adv Res 47:75
Debnath B, Hussain M, Irshad M, Mitra S, Li M, Liu S, Qiu D (2018) Exogenous melatonin mitigates acid rain stress to tomato plants through modulation of leaf ultrastructure, photosynthesis and antioxidant potential. Molecules 23(2):388
Erdal S (2019) Melatonin promotes plant growth by maintaining integration and coordination between carbon and nitrogen metabolisms. Plant Cell Rep 38:1001–1012
Fan J, Xie Y, Zhang Z, Chen L (2018) Melatonin: a multifunctional factor in plants. Int J Mol Sci 19(5):1528
Farouk S, Al-Huqail AA (2022) Sustainable biochar and/or melatonin improve salinity tolerance in borage plants by modulating osmotic adjustment, antioxidants, and ion homeostasis. Plan Theory 11(6):765
Flieger J, Flieger W, Baj J, Maciejewski R (2021) Antioxidants: classification, natural sources, activity/capacity measurements, and usefulness for the synthesis of nanoparticles. Materials 14(15):4135
Goswami S, Sharma S, Haldar C (2013) The oxidative damages caused by ultraviolet radiation type C (UVC) to a tropical rodent Funambulus pennanti: role of melatonin. J Photochem Photobiol B Biol 125:19–25
Gross J (2012) Pigments in vegetables: chlorophylls and carotenoids. Springer Science & Business Media, Berlin
Gu X, Xue L, Lu L, Xiao J, Song G, Xie M, Zhang H (2021) Melatonin enhances the waterlogging tolerance of Prunus persica by modulating antioxidant metabolism and anaerobic respiration. J Plant Growth Regul 40:2178–2190
Gul M, Khan FA, Wani SA, Bhat SA, Mir SA, Malik A, Lone SA et al (2021) Effects of foliar application of melatonin on gas exchange and certain biochemical characteristics broccoli cv. Palam Samridhi. J Appl Nat Sci 13(2):791–797
Hao S, Wang Y, Yan Y, Liu Y, Wang J, Chen S (2021) A review on plant responses to salt stress and their mechanisms of salt resistance. Horticulturae 7(6):132
Hassan MU, Ghareeb RY, Nawaz M, Mahmood A, Shah AN, Abdel-Megeed A, Qari SH et al (2022) Melatonin: a vital pro-tectant for crops against heat stress: mechanisms and prospects. Agronomy 12(5):1116
Holding DR, Streich AM (2013) Plant growth processes: transpiration, photosynthesis, and respiration. The Board of Reagents of the University of Nebraska, Lincoln, NE, pp 1–10
Hu W, Zhang J, Yan K, Zhou Z, Zhao W, Zhang X, Pu Y, Yu R (2021) Beneficial effects of abscisic acid and melatonin in overcoming drought stress in cotton (Gossypium hirsutum L.). Physiol Plant 173(4):2041–2054
Hu W, Zhang J, Wu Z, Loka DA, Zhao W, Chen B, Gao L et al (2022) Effects of single and combined exogenous application of abscisic acid and melatonin on cotton carbohydrate metabolism and yield under drought stress. Ind Crop Prod 176:114302
Imran M, Aaqil Khan M, Shahzad R, Bilal S, Khan M, Yun BW, Lee IJ et al (2021) Melatonin ameliorates thermotolerance in soybean seedling through balancing redox homeostasis and modulating antioxidant defense, phytohormones and polyamines biosynthesis. Molecules 26(17):5116
Iqbal N, Fatma M, Gautam H, Umar S, Sofo A, D’ippolito I, Khan NA (2021) The crosstalk of melatonin and hydrogen sulfide determines photosynthetic performance by regulation of carbohydrate metabolism in wheat under heat stress. Plan Theory 10(9):1778
Jahan MS, Guo S, Sun J, Shu S, Wang Y, Abou El-Yazied A, Hasan MM et al (2021) Melatonin-mediated photosynthetic performance of tomato seedlings under high-temperature stress. Plant Physiol Biochem 167:309–320
Kaya A, Doganlar ZB (2019) Melatonin improves the multiple stress tolerance in pepper (Capsicum annuum). Sci Hortic 256:108509
Khan A, Numan M, Khan AL, Lee IJ, Imran M, Asaf S, Al-Harrasi A (2020) Melatonin: awakening the defense mechanisms during plant oxidative stress. Plan Theory 9(4):407
Khan S, Sehar Z, Fatma M, Mir IR, Iqbal N, Tarighat MA, Khan NA et al (2022a) Involvement of ethylene in melatonin-modified photosynthetic-N use efficiency and antioxidant activity to improve photosynthesis of salt grown wheat. Physiol Plant 174(6):e13832
Khan TA, Saleem M, Fariduddin Q (2022b) Melatonin influences stomatal behavior, root morphology, cell viability, photosynthetic responses, fruit yield, and fruit quality of tomato plants exposed to salt stress. J Plant Growth Regul 42:1–25
Khanna K, Bhardwaj R, Alam P, Reiter RJ, Ahmad P (2023) Phytomelatonin: a master regulator for plant oxidative stress management. Plant Physiol Biochem 196:260
Knör G (2015) Recent progress in homogeneous multielectron transfer photocatalysis and artificial photosynthetic solar energy conversion. Coord Chem Rev 304:102–108
Kopečná J, Komenda J, Bučinská L, Sobotka R (2012) Long-term acclimation of the Cyanobacterium synechocystis sp. PCC 6803 to high light is accompanied by an enhanced production of chlorophyll that is preferentially channeled to trimeric photosystem I. Plant Physiol 160(4):2239–2250
Kul R, Esringü A, Dadasoglu E, Sahin Ü, Turan M, Örs S, Yildirim E et al (2019) Melatonin: role in increasing plant tolerance in abiotic stress conditions. In: Abiotic and biotic stress in plants, vol 1. IntechOpen, London, p 19
Kumar A, Sahoo U, Lal MK, Tiwari RK, Lenka SK, Singh NR, Gupta OP, Sah RP, Sharma S (2022a) Biochemical markers for low glycemic index and approaches to alter starch digestibility in rice. J Cereal Sci 106:103501. https://doi.org/10.1016/j.jcs.2022.103501
Kumar R, Kaundal P, Tiwari RK, Siddappa S, Kumari H, Lal MK, Naga KC, Sharma S, Sagar V, Kumar M (2022b) Establishment of a one-step reverse transcription recombinase polymerase amplification assay for the detection of potato virus S. J Virol Methods 307:114568. https://doi.org/10.1016/j.jviromet.2022.114568
Lal MK, Sharma N, Adavi SB, Sharma E, Altaf MA, Tiwari RK, Kumar R, Kumar A, Dey A, Paul V, Singh B (2022a) From source to sink: mechanistic insight of photoassimilates synthesis and partitioning under high temperature and elevated [CO2]. Plant Mol Biol 110:1–20. https://doi.org/10.1007/s11103-022-01274-9
Lal MK, Singh B, Tiwari RK, Kumar S, Gopalakrishnan S, Gaikwad K, Kumar A, Paul V, Singh MP (2022b) Interactive effect of retrogradation and addition of pulses, cooking oil on predicted glycemic index and resistant starch of potato. Starch-Stärke 74:2100221. https://doi.org/10.1002/star.202100221
Lal MK, Tiwari RK, Jaiswal A, Luthra SK, Singh B, Kumar S, Gopalakrishnan S, Gaikwad K, Kumar A, Paul V, Singh MP (2022c) Combinatorial interactive effect of vegetable and condiments with potato on starch digestibility and estimated in vitro glycemic response. J Food Measure Characterization 16:1–13. https://doi.org/10.1007/s11694-022-01354-w
Lal MK, Sharma E, Tiwari RK, Devi R, Mishra UN, Thakur R, Gupta R, Dey A, Lal P, Kumar A, Altaf MA, Sahu DN, Kumar R, Singh B, Sahu SK (2022d) Nutrient-mediated perception and signalling in human metabolism: a perspective of nutrigenomics. Int J Mol Sci 23:11305. https://doi.org/10.3390/ijms231911305
Li C, Tan DX, Liang D, Chang C, Jia D, Ma F (2015) Melatonin mediates the regulation of ABA metabolism, free-radical scavenging, and stomatal behaviour in two malus species under drought stress. J Exp Bot 66(3):669–680
Li C, Liang B, Chang C, Wei Z, Zhou S, Ma F (2016) Exogenous melatonin improved potassium content in malus under different stress conditions. J Pineal Res 61(2):218–229
Li Z, Su X, Chen Y, Fan X, He L, Guo J, Yang Q et al (2021) Melatonin improves drought resistance in maize seedlings by enhancing the antioxidant system and regulating abscisic acid metabolism to maintain stomatal opening under PEG-induced drought. J Plant Biol 64:299–312
Liang D, Shen Y, Ni Z, Wang Q, Lei Z, Xu N, Xia H et al (2018) Exogenous melatonin application delays senescence of kiwifruit leaves by regulating the antioxidant capacity and biosynthesis of flavonoids. Front Plant Sci 9:426
Liang D, Ni Z, Xia H, Xie Y, Lv X, Wang J, Luo X et al (2019) Exogenous melatonin promotes biomass accumulation and photosynthesis of kiwifruit seedlings under drought stress. Sci Hortic 246:34–43
Liu J, Zhang R, Sun Y, Liu Z, Jin W, Sun Y (2016) The beneficial effects of exogenous melatonin on tomato fruit properties. Sci Hortic 207:14–20
Liu J, Shabala S, Zhang J, Ma G, Chen D, Shabala L, Zhao Q et al (2020) Melatonin improves rice salinity stress tolerance by NADPH oxidase-dependent control of the plasma membrane K+ transporters and K+ homeostasis. Plant Cell Environ 43(11):2591–2605
Liu G, Hu Q, Zhang X, Jiang J, Zhang Y, Zhang Z (2022) Melatonin biosynthesis and signal transduction in plants in response to environmental conditions. J Exp Bot 73(17):5818–5827
López A, García JA, Escames G, Venegas C, Ortiz F, López LC, Acuña-Castroviejo D (2009) Melatonin protects the mitochondria from oxidative damage reducing oxygen consumption, membrane potential, and superoxide anion production. J Pineal Res 46(2):188–198
Mandal S, Ghorai M, Anand U, Roy D, Kant N, Mishra T, Mane AB, Jha NK, Lal MK, Tiwari RK, Kumar M, Radha, Ghosh A, Bhattacharjee R, Proćków J, Dey A (2022a) Cytokinins: a genetic target for increasing yield potential in the CRISPR era. Front Genet 13:26. https://doi.org/10.3389/fgene.2022.883930
Mandal S, Ghorai M, Anand U, Samanta D, Kant N, Mishra T, Rahman MH, Jha NK, Jha SK, Lal MK, Tiwari RK, Kumar M, Radha, Prasanth DA, Mane AB, Gopalakrishnan AV, Biswas P, Proćków J, Dey A (2022b) Cytokinin and abiotic stress tolerance -what has been accomplished and the way forward? Front Genet 13:1916. https://doi.org/10.3389/FGENE.2022.943025/BIBTE
Messinger J, Shevela D (2012) Principles of photosynthesis. In: Fundamentals of materials and energy and environmental sustainability, pp 302–314
Mir AR, Faizan M, Bajguz A, Sami F, Siddiqui H, Hayat S (2020) Occurrence and biosynthesis of melatonin and its exogenous effect on plants. Acta Soc Bot Pol 89(2)
Muhammad I, Yang L, Ahmad S, Mosaad IS, Al-Ghamdi AA, Abbasi AM, Zhou XB (2022) Melatonin application alleviates stress-induced photosynthetic inhibition and oxidative damage by regulating antioxidant defense system of maize: a meta-analysis. Antioxidants 11(3):512
Mukherjee S, Corpas FJ (2020) Crosstalk among hydrogen sulfide (H2S), nitric oxide (NO) and carbon monoxide (CO) in root-system development and its rhizosphere interactions: a gaseous interactome. Plant Physiol Biochem 155:800–814
Mushtaq N, Iqbal S, Hayat F, Raziq A, Ayaz A, Zaman W (2022) Melatonin in micro-tom tomato: improved drought tolerance via the regulation of the photosynthetic apparatus, membrane stability, Osmoprotectants, and root system. Life 12(11):1922
Nandy S, Mandal S, Gupta SK, Anand U, Ghorai M, Mundhra A, Rahman MH, Ray P, Mitra S, Ray D, Lal MK, Tiwari RK, Nongdam P, Pandey DK, Shekhawat MS, Jha NK, Jha SK, Kumar M, Radha, Dey A (2022) Role of polyamines in molecular regulation and cross-talks against drought tolerance in plants. J Plant Growth Regul 2022:1–17. https://doi.org/10.1007/S00344-022-10802-2
Nawaz K, Chaudhary R, Sarwar A, Ahmad B, Gul A, Hano C, Anjum S et al (2020) Melatonin as master regulator in plant growth, development and stress alleviator for sustainable agricultural production: current status and future perspectives. Sustainability 13(1):294
Naz S, Mushtaq A, Ali S, Muhammad HMD, Saddiq B, Ahmad R, Zulfiqar F, Hayat F, Tiwari RK, Lal MK, Altaf MA, Naz S, Mushtaq A, Ali S, Muhammad HMD, Saddiq B, Ahmad R, Zulfiqar F, Hayat F, Altaf MA (2022) Foliar application of ascorbic acid enhances growth and yield of lettuce (Lactuca sativa) under saline conditions by improving antioxidant defence mechanism. Funct Plant Biol. https://doi.org/10.1071/FP22139
Nykiel M, Gietler M, Fidler J, Prabucka B, Rybarczyk-Płońska A, Graska J, Labudda M et al (2022) Signal transduction in cereal plants struggling with environmental stresses: from perception to response. Plan Theory 11(8):1009
Pacini N, Borziani F (2016) Oncostatic-cytoprotective effect of melatonin and other bioactive molecules: a common target in mitochondrial respiration. Int J Mol Sci 17(3):341
Paradies G, Petrosillo G, Paradies V, Reiter RJ, Ruggiero FM (2010) Melatonin, cardiolipin and mitochondrial bioenergetics in health and disease. J Pineal Res 48(4):297–310
Pelagio-Flores R, Muñoz-Parra E, Ortiz-Castro R, López-Bucio J (2012) Melatonin regulates Arabidopsis root system architecture likely acting independently of auxin signaling. J Pineal Res 53(3):279–288
Prerostova S, Vankova R (2023) Phytohormone-mediated regulation of heat stress response in plants. In: Plant hormones and climate change. Springer Nature Singapore, Singapore, pp 167–206
Rasmussen M, Minteer SD (2014) Photobioelectrochemistry: solar energy conversion and biofuel production with photosynthetic catalysts. J Electrochem Soc 161(10):H647
Raza A, Charagh S, García-Caparrós P, Rahman MA, Ogwugwa VH, Saeed F, Jin W (2022) Melatonin-mediated temperature stress tolerance in plants. GM Crops Food 13(1):196–217
Reiter RJ, Tan DX, Rosales-Corral S, Manchester LC (2013) The universal nature, unequal distribution and antioxidant functions of melatonin and its derivatives. Mini Rev Med Chem 13(3):373–384
Ren J, Yang X, Ma C, Wang Y, Zhao J (2021) Melatonin enhances drought stress tolerance in maize through coordinated regulation of carbon and nitrogen assimilation. Plant Physiol Biochem 167:958–969
Rodacka A, Strumillo J, Puchala M, Serafin E, Bartosz G (2019) Comparison of protective properties of resveratrol and melatonin in the radiation inactivation and destruction of glyceraldehyde-3-phosphate dehydrogenase and lactate dehydrogenase. Int J Radiat Biol 95(11):1472–1483
Samanta S, Singh A, Banerjee A, Roychoudhury A (2020) Exogenous supplementation of melatonin alters representative organic acids and enzymes of respiratory cycle as well as sugar metabolism during arsenic stress in two contrasting indica rice cultivars. J Biotechnol 324:220–232
Sarropoulou V, Dimassi-Theriou K, Therios I, Koukourikou-Petridou M (2012) Melatonin enhances root regeneration, photosynthetic pigments, biomass, total carbohydrates and proline content in the cherry rootstock PHL-C (Prunus avium× Prunus cerasus). Plant Physiol Biochem 61:162–168
Sharif R, Xie C, Zhang H, Arnao MB, Ali M, Ali Q, Li Y et al (2018) Melatonin and its effects on plant systems. Molecules 23(9):2352
Sharma A, Shahzad B, Kumar V, Kohli SK, Sidhu GPS, Bali AS, Handa N, Kapoor D, Bhardwaj R, Zheng B (2019) Phytohormones regulate accumulation of osmolytes under abiotic stress. Biomolecules 9(7):285
Sharma A, Wang J, Xu D, Tao S, Chong S, Yan D, Zheng B et al (2020) Melatonin regulates the functional components of photosynthesis, antioxidant system, gene expression, and metabolic pathways to induce drought resistance in grafted Carya cathayensis plants. Sci Total Environ 713:136675
Siddiqui MH, Alamri S, Alsubaie QD, Ali HM (2020) Melatonin and gibberellic acid promote growth and chlorophyll biosynthesis by regulating antioxidant and methylglyoxal detoxification system in tomato seedlings under salinity. J Plant Growth Regul 39:1488–1502
Siddiqui MH, Mukherjee S, Kumar R, Alansi S, Shah AA, Kalaji HM, Raza A et al (2022) Potassium and melatonin-mediated regulation of fructose-1, 6-bisphosphatase (FBPase) and sedoheptulose-1, 7-bisphosphatase (SBPase) activity improve photosynthetic efficiency, carbon assimilation and modulate glyoxalase system accompanying tolerance to cadmium stress in tomato seedlings. Plant Physiol Biochem 171:49–65
Simkin AJ, Kapoor L, Doss CGP, Hofmann TA, Lawson T, Ramamoorthy S (2022) The role of photosynthesis related pigments in light harvesting, photoprotection and enhancement of photosynthetic yield in planta. Photosynth Res 152(1):23–42
Sun C, Liu L, Wang L, Li B, Jin C, Lin X (2021) Melatonin: a master regulator of plant development and stress responses. J Integr Plant Biol 63(1):126–145
Szarka A, Bánhegyi G, Asard H (2013) The inter-relationship of ascorbate transport, metabolism and mitochondrial, plastidic respiration. Antioxid Redox Signal 19(9):1036–1044
Talaat NB (2021) Co-application of melatonin and salicylic acid counteracts salt stress-induced damage in wheat (Triticum aestivum L.) photosynthetic machinery. J Soil Sci Plant Nutr 21(4):2893–2906
Tan DX, Hardeland R, Manchester LC, Korkmaz A, Ma S, Rosales-Corral S, Reiter RJ (2012) Functional roles of melatonin in plants, and perspectives in nutritional and agricultural science. J Exp Bot 63(2):577–597
Tan XL, Fan ZQ, Zeng ZX, Shan W, Kuang JF, Lu WJ, Zhao YT et al (2021) Exogenous melatonin maintains leaf quality of postharvest Chinese flowering cabbage by modulating respiratory metabolism and energy status. Postharvest Biol Technol 177:111524
Tang Y, Li J, Zhang E, Li H (2016) Effects of exogenous melatonin on photosynthetic characteristics of Chinese cabbage seedlings under aluminum stress. In: 2016 2nd international conference on advances in energy, environment and chemical engineering (AEECE 2016). Atlantis Press, Amsterdam, pp 60–63
Teng Z, Zheng W, Jiang S, Hong SB, Zhu Z, Zang Y (2022) Role of melatonin in promoting plant growth by regulating carbon assimilation and ATP accumulation. Plant Sci 319:111276
Tiwari RK, Lal MK, Naga KC, Kumar R, Chourasia KN, Subhash S, Sharma S et al (2020) Emerging roles of melatonin in mitigating abiotic and biotic stresses of horticultural crops. Sci Hortic 272:109592
Tiwari RK, Lal MK, Kumar R, Chourasia KN, Naga KC, Kumar D, Zinta G et al (2021) Mechanistic insights on melatonin-mediated drought stress mitigation in plants. Physiol Plant 172(2):1212–1226
Tiwari RK, Kumar R, Lal MK, Kumar A, Altaf MA, Devi R, Mangal V, Naz S, Altaf MM, Dey A, Aftab T (2022a) Melatonin- polyamine interplay in the regulation of stress responses in plants. J Plant Growth Regul 2022:1–17. https://doi.org/10.1007/S00344-022-10717-Y
Tiwari RK, Kumar R, Lal MK, Kumar A, Altaf MA, Devi R, Mangal V, Naz S, Altaf MM, Dey A, Aftab T (2022b) Melatonin- polyamine interplay in the regulation of stress responses in plants. J Plant Growth Regul 42:1–17. https://doi.org/10.1007/s00344-022-10717-y
Turk H, Genisel M (2020) Melatonin-related mitochondrial respiration responses are associated with growth promotion and cold tolerance in plants. Cryobiology 92:76–85
Turk H, Erdal S, Genisel M, Atici O, Demir Y, Yanmis D (2014) The regulatory effect of melatonin on physiological, biochemical and molecular parameters in cold-stressed wheat seedlings. Plant Growth Regul 74:139–152
Varghese N, Alyammahi O, Nasreddine S, Alhassani A, Gururani MA (2019) Melatonin positively influences the photosynthetic machinery and antioxidant system of Avena sativa during salinity stress. Plan Theory 8(12):610
Wang P, Sun X, Chang C, Feng F, Liang D, Cheng L, Ma F (2013) Delay in leaf senescence of Malus hupehensis by long-term melatonin application is associated with its regulation of metabolic status and protein degradation. J Pineal Res 55(4):424–434
Wang K, Tu W, Liu C, Rao Y, Gao Z, Yang C (2017) 9-cis-neoxanthin in light harvesting complexes of photosystem II regulates the binding of violaxanthin and xanthophyll cycle. Plant Physiol 174(1):86–96
Wang YF, Guo YY, Zhao CF, Li HJ, Zhang RH (2021) Exogenous melatonin achieves drought tolerance by improving photosynthesis in maize seedlings leaves. Russ J Plant Physiol 68:718–727
Wang G, Xing M, Hu T, Ji M, Li X, Amombo E, Fu J et al (2022a) Photosystem II photochemical adjustment of tall fescue against heat stress after melatonin priming. J Plant Physiol 275:153758
Wang N, Fang H, Yang Q, Liu Z, Feng H, Ji S (2022b) Exogenous melatonin alleviated leaf yellowing via inhibiting respiration and ethylene biosynthesis during shelf life in Pakchoi. Plan Theory 11(16):2102
Wang Z, Mu Y, Zhang L, Liu Z, Liu D, Jin Z, Pei Y (2023) Hydrogen sulfide mediated the melatonin induced stoma closure by regulating the K+ channel in Arabidopsis thaliana. Environ Exp Bot 205:105125
Wei W, Li QT, Chu YN, Reiter RJ, Yu XM, Zhu DH, Chen SY et al (2015) Melatonin enhances plant growth and abiotic stress tolerance in soybean plants. J Exp Bot 66(3):695–707
Williams TC, Poolman MG, Howden AJ, Schwarzlander M, Fell DA, Ratcliffe RG, Sweetlove LJ (2010) A genome-scale metabolic model accurately predicts fluxes in central carbon metabolism under stress conditions. Plant Physiol 154(1):311–323
Xie Q, Zhang Y, Cheng Y, Tian Y, Luo J, Hu Z, Chen G (2022) The role of melatonin in tomato stress response, growth and development. Plant Cell Rep 41(8):1631–1650
Xin CP, Tholen D, Devloo V, Zhu XG (2015) The benefits of photorespiratory bypasses: how can they work? Plant Physiol 167(2):574–585
Xu W, Cai SY, Zhang Y, Wang Y, Ahammed GJ, Xia XJ, Zhou J et al (2016) Melatonin enhances thermotolerance by promoting cellular protein protection in tomato plants. J Pineal Res 61(4):457–469
Xue H, Tokutsu R, Bergner SV, Scholz M, Minagawa J, Hippler M (2015) PHOTOSYSTEM II SUBUNIT R is required for efficient binding of LIGHT-HARVESTING COMPLEX STRESS-RELATED PROTEIN3 to photosystem II-light-harvesting supercomplexes in Chlamydomonas reinhardtii. Plant Physiol 167(4):1566–1578
Yan Y, Shi Q, Gong B (2020) Review of melatonin in horticultural crops. In: Melatonin—the hormone of darkness and its therapeutic potential and perspectives. IntechOpen, London
Yang X, Pan H, Shaheen SM, Wang H, Rinklebe J (2021) Immobilization of cadmium and lead using phosphorus-rich animal-derived and iron-modified plant-derived biochars under dynamic redox conditions in a paddy soil. Environ Int 156:106628
Yang S, Zhao Y, Qin X, Ding C, Chen Y, Tang Z, Yuan M et al (2022) New insights into the role of melatonin in photosynthesis. J Exp Bot 73(17):5918–5927
Ye J, Wang S, Deng X, Yin L, Xiong B, Wang X (2016) Melatonin increased maize (Zea mays L.) seedling drought tolerance by alleviating drought-induced photosynthetic inhibition and oxidative damage. Acta Physiol Plant 38(2):48
Ying S, Hill AT, Pyc M, Anderson EM, Snedden WA, Mullen RT, Plaxton WC et al (2017) Regulatory phosphorylation of bacterial-type PEP carboxylase by the Ca2+−dependent protein kinase RcCDPK1 in developing castor oil seeds. Plant Physiol 174(2):1012–1027
Yu J, Li R, Fan N, Yang Z, Huang B (2017) Metabolic pathways involved in carbon dioxide enhanced heat tolerance in Bermudagrass. Front Plant Sci 8:1506
Zhang N, Sun Q, Zhang H, Cao Y, Weeda S, Ren S, Guo YD (2015) Roles of melatonin in abiotic stress resistance in plants. J Exp Bot 66(3):647–656
Zhang N, Zhang HJ, Sun QQ, Cao YY, Li X, Zhao B, Guo YD et al (2017) Proteomic analysis reveals a role of melatonin in promoting cucumber seed germination under high salinity by regulating energy production. Sci Rep 7(1):1–15
Zhang M, He S, Zhan Y, Qin B, Jin X, Wang M, Wu Y et al (2019) Exogenous melatonin reduces the inhibitory effect of osmotic stress on photosynthesis in soybean. PloS One 14(12):e0226542
Zhang JY, Cun Z, Chen JW (2020) Photosynthetic performance and photosynthesis-related gene expression coordinated in a shade-tolerant species Panax notoginseng under nitrogen regimes. BMC Plant Biol 20:1–19
Zhao D, Yu Y, Shen Y, Liu Q, Zhao Z, Sharma R, Reiter RJ (2019) Melatonin synthesis and function: evolutionary history in animals and plants. Front Endocrinol 10:249
Zhao H, Zhang Z, Zhang Y, Bai L, Hu X, Li X, Wang Y et al (2022) Melatonin reduces photoinhibition in cucumber during chilling by regulating the Calvin-Benson cycle. Sci Hortic 299:111007
Zheng X, Tan DX, Allan AC, Zuo B, Zhao Y, Reiter RJ, Kong J et al (2017) Chloroplastic biosynthesis of melatonin and its involvement in protection of plants from salt stress. Sci Rep 7(1):1–12
Zhou X, Zhao H, Cao K, Hu L, Du T, Baluška F, Zou Z (2016) Beneficial roles of melatonin on redox regulation of photosynthetic electron transport and synthesis of D1 protein in tomato seedlings under salt stress. Front Plant Sci 7:1823
Zou JN, Jin XJ, Zhang YX, Ren CY, Zhang MC, Wang MX (2019) Effects of melatonin on photosynthesis and soybean seed growth during grain filling under drought stress. Photosynthetica 57(2):512–520
Zuo Z, Sun L, Wang T, Miao P, Zhu X, Liu S, Li X et al (2017) Melatonin improves the photosynthetic carbon assimilation and antioxidant capacity in wheat exposed to nano-ZnO stress. Molecules 22(10):1727
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2023 The Author(s), under exclusive license to Springer Nature Singapore Pte Ltd.
About this chapter

Cite this chapter
Shahani, A.A.A., Saad, M., Shakoor, A., Gonzalez, N.C.T., Farooq, T.H. (2023). Regulatory Role of Melatonin in Photosynthesis and Respiration. In: Kumar, R., Altaf, M.A., Lal, M.K., Tiwari, R.K. (eds) Melatonin in Plants: A Regulator for Plant Growth and Development. Springer, Singapore. https://doi.org/10.1007/978-981-99-6745-2_5
Download citation
DOI: https://doi.org/10.1007/978-981-99-6745-2_5
Published:
Publisher Name: Springer, Singapore
Print ISBN: 978-981-99-6744-5
Online ISBN: 978-981-99-6745-2
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)