
Abstract
The postindustrial age radically altered global climate conditions, posing a difficult task for plants and crops to thrive under stress environments like high temperatures, salt, waterlogging, heavy metals, drought, and so on. A small period of poor weather had a substantial impact on the development and growth of plants, eventually influencing crop quality, yield, and agricultural sustainability as a whole. Plant cells produce free oxygen (O2) radicals and their derivatives, known as reactive oxygen species (ROS), as by-products of other reactions in such hostile environments. Furthermore, these ROS molecules are used as signaling molecules in plants for signal transduction in response to changing environmental conditions. The cytoplasmic balance that triggers the antioxidant defense mechanisms is disrupted as a result of the excessive accumulation of ROSs inside the cell. Plants have developed a complicated ROS scavenging system to avoid sensitive cellular components from being damaged by reactive oxygen species. Enzymatic antioxidants, like catalase (CAT), peroxidase (POD), superoxide dismutase (SOD), glutathione peroxidases (GPX), and ascorbate peroxidases (APX), and nonenzymatic antioxidants, like glutathione, ascorbate, tocopherols, and phenolic compounds, are important antioxidants that play key roles in eliminating superoxide (O2−) and (H2O2). The antioxidant capacity of plants is the sum of the activities of all enzymatic and nonenzymatic antioxidant systems. This chapter seeks to provide fundamental information on enzymatic and nonenzymatic antioxidants, their occurrence, characteristics, and the antioxidant defense system involved in reactive oxygen species (ROS) detoxification under various stresses, as well as their interactions with cellular components.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
1.1 Introduction
Based on their biochemical nature, antioxidants are divided into two categories: enzymatic and nonenzymatic. While substantial anabolic and catabolic reactions are occurring, these chemicals are engaged in the detoxification of free radicals or reactive oxygen species (ROS). Both classes of antioxidants are capable of efficiently neutralizing ROS and converting them into relatively stable nontoxic molecules, preventing oxidative damage to cellular apparatuses. As a result, antioxidants are the most important first line of defense against the oxidative stress-induced cell damage. Antioxidants, both enzymatic and nonenzymatic, are electron-rich compounds that readily share electrons with highly energetic ROS and free radicals, stabilizing cellular randomness.
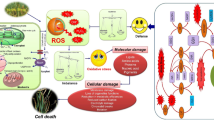
Furthermore, they may interfere with the oxidizing chain reaction in order to reduce free radical damage (Apel and Hirt 2004). Antioxidants are also known as ROS scavengers because they use dynamic and synergistic processes to keep the intracellular concentration of ROS in check. It (antioxidant) is a substance that may scavenge reactive oxygen species (ROS) without being converted into a harmful radical (Noctor and Foyer 1998). As a result, antioxidant enzymes are crucial for sustaining good cellular and systemic health and well-being. All highly reactive, oxygen-containing molecules, including the free radicals, are referred to as ROS. The hydroxyl radical (OH−), singlet oxygen, superoxide anion radical, H2O2 (hydrogen peroxide), hypochlorite radicals, lipid peroxides, and nitric oxide radical are all examples of reactive oxygen species. All have the ability to react with membrane lipids, enzymes, and other molecules, resulting in the loss of critical cellular structures and functions and a variety of negative consequences for plants and animals. As previously stated, free radicals or ROS are highly reactive compounds that are released directly or as a by-product during normal metabolic processes in various cellular compartments like mitochondria, chloroplast, peroxisomes, and apoplast (Panieri and Santoro 2015), but their synthesis is accelerated during extreme conditions. Because these compounds have a lone pair of electrons, they are very unstable and thus highly reactive (Del Río and López-Huertas 2016). ROS-induced oxidative stress is a well-controlled process, and the balance in-between the ROS and its quenching define a plant’s and any other organism’s well-being. If antioxidants reach a condition of disequilibrium, meaning they are unable to destroy free radicals, the cell and tissue suffer oxidative damage. The degree of oxidative stress caused by free radicals/ROS is determined by their concentration, kind, synthesis site, and developmental stage (Møller et al. 2007). Plants exposed to ROS for a long time period can suffer considerable damage to their cell machinery and biomolecules, including protein oxidation, lipid peroxidation (Mittler 2002), PCD (programmed cell death), and inhibition of the enzymes and also damages nucleic acid, which can lead to tissue necrosis or plant death (Fig. 1.1) (Pérez-Pérez et al. 2012).

Illustrative representation of different agents generating free radicals and reactive oxygen species in plants and different antioxidants scavenging agents
1.2 Enzymatic Antioxidants
Several enzymes, like GPX, SOD, glutathione reductase, APX, and CAT (catalase), are the enzymatic components of the antioxidant defense system found in diverse subcellular compartments. In plants under oxidative stress, these enzymatic antioxidant molecules are critical for maintaining cellular homeostasis.
1.2.1 SOD (Superoxide Dismutases)
SODs are enzymes which can catalyze the dismutation/partitioning of O2− to H2O2 and molecular oxygen (O2). Because O2− is a typical consequence of oxygen metabolism, SOD is thought to be the first line of defense against the reactive O2 species-induced damage. By neutralizing the superoxide radical, the Haber–Weiss reaction eliminates the possibility of hydroxyl ion production. These enzymes are classed as metalloenzymes based on metal cofactors, subcellular distribution, and protein folds. Cu-/Zn-containing SODs have been found in prokaryotic and eukaryotic organisms, and in plant cells, they can be found in the cytoplasm, extracellular space, or chloroplasts and can also be found in lysosomes, cytoplasm, and nuclear compartments in mammalian cells. Plant cytoplasm and chloroplasts have been found to have Fe-containing SODs. MnSODs are found in all kingdoms but particularly in eukaryotic mitochondria, where they play a critical role in preserving mitochondria by scavenging ROS (Pilon et al. 2011) and initiating cellular differentiation (Moller 2001, 2012). According to Feng et al. (2016), SODs are found in different organisms, such as mammals, yeast, bacteria, and plants. Multiple genes producing SODs exist in plants, and their expression can be influenced by developmental stage, tissue type, and environmental cues (Scandalios 2005; Menezes-Benavente et al. 2004). There are nine SOD genes in tomatoes, which are unevenly distributed across 12 chromosomes and include four Cu/ZnSODs, one MnSOD, and three FeSODs (Feng et al. 2016). Various investigations revealed that different types of SOD genes had varied levels of expression under harsh environmental conditions. Underwater stress, for example, the expression patterns of the banana genes like MaCSD1B and MaMSD1A, which are involved in SOD production, were utterly incompatible (Feng et al. 2015).
Furthermore, SODs with the same cofactor may not necessarily have the same role in various species. MnSOD expression was not affected by oxidative stress in Arabidopsis, but it was affected significantly by drought and cold stress in wheat and salt stress in pea (Baek and Skinner 2003). Additionally, alternative splicing and miRNAs have been implicated in the regulation of SOD gene expression (Lu et al. 2010). Various plant species, such as Populus trichocarpa, Sorghum bicolor, Musa acuminata, and Arabidopsis thaliana, have been found to contain the SOD genes (Srivastava et al. 2009).
1.2.2 CAT (Catalases)
These are tetrameric enzymes, with iron as a prosthetic group attached to each monomer; they catalyze the energy-efficient disproportionation of H2O2 into water molecules (H2O) and molecular oxygen (Regelsberger et al. 2002; Zamocky et al. 2008). In contrast to H2O2, it has a lower affinity for R-O-O-R (organic peroxides) and also has a higher turnover rate among antioxidant enzymes and is unique among antioxidant enzymes because it does not need a reducing counterpart. Unfavorable conditions necessitate increased energy generation, and expenditure by plants and other organisms leads to an increase in catabolic reactions, which yields H2O2. CATs are predominantly attached to peroxisomes, where they execute β-oxidation of fatty acid, photorespiration, and purine catabolism, all of which produce a considerable amount of H2O2 (Mittler 2002). Hydrogen peroxide (H2O2) is also a key signaling molecule in plant development and plant response to the environment (Mhamdi et al. 2010). Plants have numerous CATs, each produced by a separate gene, that respond differently to different stressors that are known to cause ROS. Recent investigations reveal that CAT is also prevalent in other subcellular compartments like the mitochondria, chloroplast, and cytosol, albeit no evidence of substantial CAT activity has been observed (Mhamdi et al. 2010). CAT1 is mostly expressed in plant pollen and seeds; on the other side, CAT2 is not only expressed in photosynthetic tissues (mostly) but also expressed in seeds and roots, while CAT3 is predominantly expressed in vascular tissues and leaf in angiosperms (McClung 1997; Frugoli et al. 1996).
1.2.3 APX (Ascorbate Peroxidases)
Ascorbate peroxidases are the heme peroxidase superfamily that is involved in the recycling of AsA and the response to environmental stress in plants (Ishikawa and Shigeoka 2008; Lazzarotto et al. 2011). These enzymes catalyze the conversion of H2O2 to water and monodehydroascorbate (MDHA) using ascorbate (AsA) as an electron donor (Caverzan et al. 2012). Plants have five different isoforms of APX, which are found in diverse subcellular compartments like the cytosol, mitochondria, peroxisomes, and chloroplast (Sharma and Dubey 2005). These enzymes are divided into groups based on the amino acids they contain and the cell compartments in which they are found. Several abiotic stressors in plants have varied effects on the APX genes (Rosa et al. 2010; Caverzan et al. 2014). APXs, CATs, and SODs must be in balance to determine the effective intracellular level of H2O2 and oxygen, and variations in this equilibrium appear to trigger compensation processes (Apel and Hirt 2004; Scandalios 2002, 2005). Under a normal and stressful environment, APX is a key component of the ASC–GSH (ascorbate–glutathione) cycle, which eliminates excess H2O2 from plant cells (Mittler and Zilinskas 1991). Because APX is more broadly distributed and also has a higher affinity for hydrogen peroxide than CAT, it is a more effective H2O2 scavenger during stressful situations. APX is a chloroplastic isoenzyme expressed by a single gene in higher plants (apx1).
1.2.4 GPx (Glutathione Peroxidases)
The term glutathione peroxidases collectively describe the group of phylogenetically related nonheme and thiol-containing peroxidase enzymes. It was called after GPx-1, the mammalian tetrameric seleno enzyme, which was the first to be defined and reported. More than 700 members of the GPx family have been identified so far, overall domains of life. G. C. Mills discovered its activity in red blood cells in 1957, where it protected hemoglobin from oxidative degradation. The biological function of GPxs is to catalyze the H2O2 reduction or lipid hydroperoxides into H2O or the corresponding alcohols by using GSH (glutathione) as a reducing agent. The catalytic moiety of selenium was later identified as selenocysteine, the 21st naturally occurring amino acid. Selenocysteine was recommended as a way to ensure a rapid reaction with the hydroperoxide and glutathione reducibility. GPx1 is found in the mitochondria, chloroplasts, and cytosol of a wide range of plants and animals, and it serves as an antioxidant in several cellular compartments. In human beings, there are eight distinct glutathione peroxidase isoforms (GPx1-8) that have been found. The mechanism of reaction comprises oxidation of the selenol of a selenocysteine residue by H2O2. The RSeOH (selenenic acid) group is formed as a result of this action. A two-step mechanism converts selenenic acid back to selenol, starting with a reaction with GSH to create the GS-SeR and H2O. The GS-SeR intermediate is reduced back to selenol by a second GSH molecule, releasing a by-product GS-SG.
1.3 Nonenzymatic Antioxidants
Antioxidants that are generally nonenzymatic in nature are found in all subcellular parts of plants. These antioxidants help to detoxify reactive O2 species and free radicals and can also help to reduce the substrates from antioxidant enzymes (Mittler 2002). The primary cellular redox buffers GSH and ascorbate, along with carotenoids, tocopherol, and polyphenolic compounds, are different forms of nonenzymatic defense pathways (Scandalios 2002).
1.3.1 Vitamin C (Ascorbic Acid)
Ascorbate is a well-known vitamin having anti-oxidizing properties that have been found in a variety of organelles and even apoplast. It occurs in different reduced and oxidized forms, as ascorbic acid (90% of the ascorbate pool) and mono- and dehydroascorbic acid (Smirnoff 2011). The ratio of oxidized to reduced ascorbate is a key element that influences plant oxidative stress resistance (Conklin et al. 2000; Cruz-Rus et al. 2012). Glutathione reductase, dehydroascorbate reductase, and monodehydroascorbate reductase are among the NAD(P)H-dependent enzymes that keep AsA in its reduced state (Mittler 2002; Foyer and Noctor 2011). According to the Horemans et al. (2000), the mitochondria are the major site for ascorbate production, from which it is transferred to other cell organs via a proton-electron gradient. Due to its ability to transfer electrons in enzymatic and nonenzymatic processes, AsA is a critical component for ROS detoxification in the aqueous phase. AsA can quickly remove O2−, hydroxide ions, and 1O2, and therefore, it can reduce H2O2 to H2O by the ascorbate peroxidase mechanism, protecting membranes (Blokhina et al. 2003). In the ascorbate–glutathione cycle, APX utilizes two AsA molecules to decrease H2O2 to water, with the intermediate monodehydroascorbate, which is a short-lived radical that is further disproportionated into dehydroascorbate (DHA) and AsA. Furthermore, it plays a crucial role in the dynamic and control of the cell cycle, the advancement of the G1 to S stage of the cell division pathway, and cellular elongation, as well as many photosynthetic functions (Smirnoff 2011). Although the exact biosynthetic process for ascorbate is unknown, it is speculated that d-glucose may be used as a precursor.
1.3.2 Glutathione
Glutathione (GSH) oxidation by ROS results in the formation of GSSG, which is present in all plant cell compartments. In cellular compartments, GSH, together with its oxidized counterpart, GSSG, maintains a redox balance. It has been revealed that the GSH/GSSH pair is involved in gene regulation and cell cycle pathways (Mittler 2002). The antioxidant glutathione and ascorbic acid are prolific and stable, and they have the right redox potential to react with different varieties of substrate and compounds. Glutathione is a tripeptide molecule made up of three amino acids, glutamine, cysteine, and glycine, that are found in all plant cell parts, including the cytoplasm, vacuoles, chloroplasts, mitochondria, and endoplasmic reticulum (Millar et al. 2003). In most plant cells, glutathione is the important source of nonprotein thiols. Glutathione is appropriate for a wide range of metabolic actions in all organisms due to the presence of a thiol group and its reactivity. The reduced form of glutathione (GSH) is found at higher concentrations in chloroplasts. Unlike the ascorbate biogenesis system, the glutathione biosynthetic pathway is well-established and identical throughout all domains of life. The amino acids are linked to produce the full tripeptide in two ATP-dependent stages catalyzed by GSHS and γ-ECS. These events take place in chloroplastic and non-chloroplastic segments, and glutathione concentrations and redox status play a crucial role in different pathways. GSH is used to reduce DHA both in an enzymatic and nonenzymatic way in the ascorbate–glutathione pathway, and it is then oxidized to GSSG. Glutathione reductase catalyzes the regeneration of GSH from GSSG, with NADPH as the reducing agent. The cysteine residue in GSH’s tripeptide has a high reactivity potential. GSH scavenges harmful H2O2 by reacting nonenzymatically with O2−, OH−, and 1O2. The ability of GSH to replenish another potent antioxidant, ascorbic acid, via the ascorbate–glutathione cycle gives it a crucial role in antioxidative defense (Millar et al. 2003). It has been observed that the antioxidant property of glutathione was utilized to achieve transgenic lines of tobacco (Foyer and Noctor 2005), which can withstand oxidative stress (Del Río and López-Huertas 2016). GSH has been involved in sensing alterations in redox equilibrium and transferring these alterations to appropriate target proteins, in addition to being a co-substrate and reducing agent in defense against reactive oxygen species.
1.3.3 Vitamin E
All types of tocopherols are methylated fat-soluble phenols that have similar to vitamin E (Sharma et al. 2012). They primarily function as antioxidants in plants, algae, and mammals, but they may also have additional functions. In lipid bilayers, α-tocopherol is the most effective for the removal of peroxyl radicals. Tocopherols are the most powerful scavengers of free radicals. At the energy of 323 kJ mol−1, the hydroxyl bond present in vitamin E becomes weaker than in the majority of phenols and readily liberates hydrogen atom and thereby helps in quenching peroxyl radicals and other free radicals, decreasing their harmful effect (Lide 2006). The produced tocopheryl radical is generally nonreactive, and through redox interaction with a hydrogen donor such as ascorbate or other antioxidants, it reverts to tocopherol (Igamberdiev and Hill 2004; Traber and Stevens 2011). Tocopherols are integrated into cell membranes and thereby protect the chloroplast membrane from oxidative damage due to their fat-soluble nature (Blokhina et al. 2003). α-Tocopherols are significantly bioactive and prominent antioxidants in the chloroplast lamina and are primarily utilized for defending them against the photooxidative effect. It has been observed that a single α-tocopherol molecule may neutralize one 20 singlet oxygen and also act as reusable chain reaction terminators for PUFA radicals synthesized during oxidation of lipids (Hare et al. 1998; Wu and Tang 2004; Ledford and Niyogi 2005). Photosynthesis and other metabolic processes in chloroplasts produce ROS, which causes lipid peroxidation in plant cells. To cope up with a range of abiotic stress conditions, the content of α-tocopherol in photosynthetic plant tissues increases dramatically (Noctor 2006). The ability of α-tocopherols to scavenge and quench ROS aids in the modulation of signal transduction and the stabilization of membranes (Kruk et al. 2005; Noctor 2006). They work as a free radical capturing process by preventing the chain extension stage in lipid autooxidation. Plants respond to oxidative stress by expressing genes involved in tocopherol production (Table 1.1) (Giacomelli et al. 2007; Wu and Tang 2004).
1.3.4 Carotenoids
Carotenoids, also known as tetra terpenoids, are pigments that naturally occur in plants and microorganisms (Otles and Cagindi 2008). To date, more than 750 natural carotenoids have been discovered. These chemicals give different vegetables and fruits their distinct colors. Xanthophylls and carotenes are the two broad categories of carotenoids. Carotenoids are classified into two broad subclasses, xanthophylls and carotenes; the former class contains oxygen, whereas the latter class is purely hydrocarbons and contains no oxygen (Ngamwonglumlert et al. 2017). Carotenoids found in the human diet may help to prevent cancer, age-related muscle degeneration, atherosclerosis, and other disorders. These pigments are lipid-soluble and are absorbed together with fats through the gut tract. Carotenoids have a variety of beneficial activities in plants, including attracting pollinators, indicating fruit development, assisting in photosynthesis, and protecting cells from light-induced damage in plants and in photosynthetic bacteria and algae (Lerfall 2016). The ability of β-carotene to quench free radicals such as O2−, OH−, and 1O2 without undergoing any changes or degradation is largely due to its highly delocalized polyene backbone or conjugated double bond structure, which is primarily responsible for its antioxidant behavior. Carotenoids found in higher concentrations in particular tissues and organs can protect lipids against oxidative damage.
1.3.5 Phenolic Compounds
Tannins, flavonoids, lignins, and stilbenes are examples of phenolic compounds, which constitute a varied group of naturally occurring secondary metabolites common in plants. Multiple phenol rings distinguish these compounds, making them suited for free radical scavenging in both de novo and in vitro conditions. Over 8000 phenolic bioactive substances have been detected in different plant families. Phenylalanine/shikimic acid is the common intermediate precursor for all plant phenolic compounds. Under in vitro conditions, polyphenols have been shown to be more potent antioxidants than ascorbate and tocopherols. Phenols are the most significant dietary elements for humans, providing bitterness, color, astringency, flavor, odor, and oxidative stability in food (Schroeter et al. 2002). Antioxidative activities of polyphenols are characterized by the presence of readily available donor hydrogen or electron (Rice-Evans et al. 1997). In another way, phenols alter the peroxidation kinetics of lipid membrane and packaging, thereby protecting the membrane integrity (Schroeter et al. 2002). Furthermore, phenolics have been implicated in the H2O2 scavenging cascade in plant cells. Polyphenols play different functions in plants, which include pigmentation to plants; increase and decrease of plant growth regulators, for example, auxin; UV protectants against ionizing light; deterrence to herbivores; phytoalexins; and signaling compounds in ripening and other plant developmental activities (Huber et al. 2003; Lattanzio et al. 2006).
1.4 Conclusion
Antioxidants are created normally, but they are triggered and upregulated in stressful situations, which help to retain the structural firmness of cell organelles while probably reducing oxidative damage. Plant defense is aided by a number of antioxidant enzymes. The production and activation of ROS scavenging enzyme systems in transgenic plants to increase their tolerance to a variety of stress conditions. Furthermore, because numerous enzymes and their different isoforms are involved and reactive oxygen species is only one of the major factors of plant resistance to unfavorable environmental and biotic stimuli, further research is needed in this field. The increasing number of articles addressing superoxide dismutase, common antioxidant enzyme, ascorbate peroxidase, glutathione peroxidase enzyme, and glutathione reductase enzymes demonstrates these enzymes’ favorable responses to biotic and abiotic stressors. These findings highlight the need to investigate these enzymes in order to better understand their role in the scavenging of hazardous cell products in a variety of species and the relationship between biological processes and oxidative stress.
References
Apel K, Hirt H (2004) Reactive oxygen species: Metabolism, oxidative stress and signal transduction. Annu Rev Plant Biol 55:373–399
Baek KH, Skinner DZ (2003) Alteration of antioxidant enzyme gene expression during cold acclimation of near-isogenic wheat lines. Plant Sci 165(6):1221–1227
Blokhina O, Virolainen E, Fagerstedt KV (2003) Antioxidants, oxidative damage and oxygen deprivation stress: A review. Ann Bot 91(2):179–194
Caverzan A, Passaia G, Rosa SB, Ribeiro CW, Lazzarotto F, Margis-Pinheiro M (2012) Plant responses to stresses: role of ascorbate peroxidase in the antioxidant protection. Genet Mol Biol 35(4):1011–1019
Caverzan A, Bonifacio A, Carvalho FEL, Andrade CMB, Passaia G, Schünemann M, Maraschin FS, Martins MO, Teixeira FK, Rauber R, Margis R, Silveira JAG, Margis-Pinheiro M (2014) The knockdown of chloroplastic ascorbate peroxidases reveals its regulatory role in the photosynthesis and protection under photo-oxidative stress in rice. Plant Sci 214:74–87
Conklin PL, Saracco SA, Norris SR, Last RL (2000) Identification of ascorbic acid-deficient Arabidopsis thaliana mutants. Genetics 154(2):847–856
Cruz-Rus E, Amaya I, Valpuesta V (2012) The challenge of increasing vitamin C content in plant foods. Biotechnol J 7(9):1110–1121
Del Río LA, López-Huertas E (2016) ROS generation in peroxisomes and its role in cell signaling. Plant Cell Physiol 57(7):1364–1376
Feng X, Lai Z, Lin Y, Lai G, Lian C (2015) Genome-wide identification and characterization of the superoxide dismutase gene family in Musa acuminata cv. Tianbaojiao (AAA group). BMC Genomics 16:823
Feng K, Yu J, Cheng Y, Ruan M, Wang R, Ye Q, Zhou G, Li Z, Yao Z, Yang Y, Zheng Q, Wan H (2016) The SOD gene family in tomato: Identification, phylogenetic relationships, and expression patterns. Front Plant Sci 7:1279
Foyer CH, Noctor G (2005) Redox homeostasis and antioxidant signaling: a metabolic interface between stress perception and physiological responses. Plant Cell 17(7):1866–1875
Foyer CH, Noctor G (2011) Ascorbate and glutathione: the heart of the redox hub. Plant Physiol 155(1):2–18
Frugoli JA, Zhong HH, Nuccio ML, McCourt P, McPeek MA, Thomas TL, McClung CR (1996) Catalase is encoded by a multigene family in Arabidopsis thaliana (L.) Heynh. Plant Physiol 112(1):327–336
Giacomelli L, Masi A, Ripoll DR, Lee MJ, van Wijk KJ (2007) Arabidopsis thaliana deficient in two chloroplast ascorbate peroxidases shows accelerated light-induced necrosis when levels of cellular ascorbate are low. Plant Mol Biol 65(5):627–644
Hare PD, Cress WA, Van Staden JV (1998) Dissecting the roles of osmolyte accumulation during stress. Plant Cell Environ 21(6):535–553
Horemans N, Foyer CH, Asard H (2000) Transport and action of ascorbate at the plant plasma membrane. Trends Plant Sci 5(6):263–267
Huber B, Eberl L, Feucht W, Polster J (2003) Influence of polyphenols on bacterial biofilm formation and quorum-sensing. Zeitschrift Fur Naturforschung. C. J Biosci 58(11–12):879–884
Igamberdiev AU, Hill RD (2004) Nitrate, NO and haemoglobin in plant adaptation to hypoxia: An alternative to classic fermentation pathways. J Exp Bot 55(408):2473–2482
Ishikawa T, Shigeoka S (2008) Recent advances in ascorbate biosynthesis and the physiological significance of ascorbate peroxidase in photosynthesizing organisms. Biosci Biotechnol Biochem 72(5):1143–1154
Kruk J, Holländer-Czytko H, Oettmeier W, Trebst A (2005) Tocopherol as singlet oxygen scavenger in photosystem II. J Plant Physiol 162(7):749–757
Lattanzio V, Lattanzio VML, Cardinali A (2006) Role of phenolics in the resistance mechanisms of plants against fungal pathogens and insects. Phytochem: Adv Res 5:23–67
Lazzarotto F, Teixeira FK, Rosa SB, Dunand C, Fernandes CL, de Vasconcelos Fontenele AV, Silveira JAG, Verli H, Margis R, Margis-Pinheiro M (2011) Ascorbate peroxidase related (APx-R) is a new heme-containing protein functionally associated with ascorbate peroxidase but evolutionarily divergent. New Phytol 191(1):234–250
Ledford HK, Niyogi KK (2005) Singlet oxygen and photo-oxidative stress management in plants and algae. Plant Cell Environ 28(8):1037–1045
Lerfall J (2016) Carotenoids: Occurrence, properties and determination. In: Caballero B, Finglas PM, Toldrá F (eds) Encyclopedia of food and health. Elsevier, pp 663–6669
Lide DR (ed) (2006) CRC handbook of chemistry and physics, 87th edn. CRC Press Press
Lu Y, Feng Z, Bian L, Xie H, Liang J (2010) miR398 regulation in rice of the responses to abiotic and biotic stresses depends on CSD1 and CSD2 expression. Funct Plant Biol 38(1):44–53
McClung CR (1997) Regulation of catalases in Arabidopsis. Free Radic Biol Med 23(3):489–496
Menezes-Benavente L, Teixeira FK, Alvim Kamei CLA, Margis-Pinheiro M (2004) Salt stress induces altered expression of genes encoding antioxidant enzymes in seedlings of a Brazilian indica rice (Oryza sativa L.). Plant Sci 166(2):323–331
Mhamdi A, Queval G, Chaouch S, Vanderauwera S, Van Breusegem FV, Noctor G (2010) Catalase function in plants: a focus on Arabidopsis mutants as stress-mimic models. J Exp Bot 61(15):4197–4220
Miller AF (2012) Superoxide dismutases: ancient enzymes and new insights. FEBS Lett 586(5):585–595
Millar AH, Mittova V, Kiddle G, Heazlewood JL, Bartoli CG, Theodoulou FL, Foyer CH (2003) Control of ascorbate synthesis by respiration and its implications for stress responses. Plant Physiol 133(2):443–447
Mittler R (2002) Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci 7(9):405–410
Mittler R, Zilinskas BA (1991) Purification and characterization of pea cytosolic ascorbate peroxidase. Plant Physiol 97(3):962–968
Moller IM (2001) Plant mitochondria and oxidative stress: electron transport, NADPH turnover, and metabolism of reactive oxygen species. Annu Rev Plant Physiol Plant Mol Biol 52:561–591
Møller IM, Jensen PE, Hansson A (2007) Oxidative modifications to cellular components in plants. Annu Rev Plant Biol 58:459–481
Ngamwonglumlert L, Devahastin S, Chiewchan N (2017) Natural colorants: pigment stability and extraction yield enhancement via utilization of appropriate pretreatment and extraction methods. Crit Rev Food Sci Nutr 57(15):3243–3259
Noctor G (2006) Metabolic signalling in defence and stress: the central roles of soluble redox couples. Plant Cell Environ 29(3):409–425
Noctor G, Foyer CH (1998) Ascorbate and glutathione: keeping active oxygen under control. Annu Rev Plant Physiol Plant Mol Biol 49:249–279
Otles S, Cagindi O (2008) Carotenoids as natural colorants. In: Socaciu C (ed) Food colorants: chemical and functional properties. CRC Press Press, pp 51–70
Panieri E, Santoro MM (2015) ROS signaling and redox biology in endothelial cells. Cell Mol Life Sci 72(17):3281–3303
Pérez-Pérez ME, Lemaire SD, Crespo JL (2012) Reactive oxygen species and autophagy in plants and algae. Plant Physiol 160(1):156–164
Pilon M, Ravet K, Tapken W (2011) The biogenesis and physiological function of chloroplast superoxide dismutases. Biochim Biophys Acta 1807(8):989–998
Regelsberger G, Jakopitsch C, Plasser L, Schwaiger H, Furtmüller PG, Peschek GA, Zámocký M, Obinger C (2002) Occurrence and biochemistry of hydroperoxidases in oxygenic phototrophic prokaryotes (cyanobacteria). Plant Physiol Biochem 40(6–8):479–490
Rice-Evans CA, Miller NJ, Paganga G (1997) Antioxidant properties of phenolic compounds. Trends Plant Sci 2(4):152–159
Rosa SB, Caverzan A, Teixeira FK, Lazzarotto F, Silveira JAG, Ferreira-Silva SL, Abreu-Neto J, Margis R, Margis-Pinheiro M (2010) Cytosolic APx knockdown indicates an ambiguous redox responses in rice. Phytochemistry 71(5–6):548–558
Scandalios JG (2002) The rise of ROS. Trends Biochem Sci 27(9):483–486
Scandalios JG (2005) Oxidative stress: molecular perception and transduction of signals triggering antioxidant gene defenses. Braz J Med Biol Res 38(7):995–1014
Schroeter H, Boyd C, Spencer JP, Williams RJ, Cadenas E, Rice-Evans C (2002) MAPK signaling in neurodegeneration: influences of flavonoids and of nitric oxide. Neurobiol Aging 23(5):861–880
Sharma P, Dubey RS (2005) Drought induces oxidative stress and enhances the activities of antioxidant enzymes in growing rice seedlings. Plant Growth Regul 46(3):209–221
Sharma P, Jha AB, Dubey RS, Pessarakli M (2012) Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J Bot 2012:1–26
Smirnoff N (2011) Vitamin C: the metabolism and functions of ascorbic acid in plants. Advances in botanical research. In: Rebeille F, Douce R (eds) Biosynthesis of vitamin S in plants: Vitamin S B6, B8, B9, C, E, K, vol 2, 1st edn. Academic Press, pp 107–177
Srivastava V, Srivastava MK, Chibani K, Nilsson R, Rouhier N, Melzer M, Wingsle G (2009) Alternative splicing studies of the reactive oxygen species gene network in Populus reveal two isoforms of high-isoelectric-point superoxide dismutase. Plant Physiol 149(4):1848–1859
Traber MG, Stevens JF (2011) Vitamins C and E: beneficial effects from a mechanistic perspective. Free Radic Biol Med 51(5):1000–1013
Wu YS, Tang KX (2004) MAP kinase cascades responding to environmental stress in plants. Acta Bot Sin 46:127–136
Zamocky M, Furtmüller PG, Obinger C (2008) Evolution of catalases from bacteria to humans. Antioxid Redox Signal 10(9):1527–1548
Author information
Authors and Affiliations
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2022 The Author(s), under exclusive license to Springer Nature Singapore Pte Ltd.
About this chapter

Cite this chapter
Sharma, S.K., Singh, D., Pandey, H., Jatav, R.B., Singh, V., Pandey, D. (2022). An Overview of Roles of Enzymatic and Nonenzymatic Antioxidants in Plant. In: Aftab, T., Hakeem, K.R. (eds) Antioxidant Defense in Plants. Springer, Singapore. https://doi.org/10.1007/978-981-16-7981-0_1
Download citation
DOI: https://doi.org/10.1007/978-981-16-7981-0_1
Published:
Publisher Name: Springer, Singapore
Print ISBN: 978-981-16-7980-3
Online ISBN: 978-981-16-7981-0
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)