
Abstract
Alcohol dependence is a puzzling brain disorder causing enormous suffering and financial costs world-wide. One of the few common denominators of all addictive drugs is activation of the mesolimbic dopamine system resulting in increased dopamine levels in the nucleus accumbens. In order to understand the development of addiction and find new efficient treatment strategies we need to understand how addictive drugs increase dopamine following acute and chronic administration of drugs. In the search for mechanisms underlying ethanol’s ability to increase dopamine in the nucleus accumbens we have found taurine to be of major importance, although the complete picture remains to be disclosed. The aim of the present study was to explore whether chronic voluntary ethanol intake influences the ethanol-induced elevation of taurine. By means of in vivo micro-dialysis we found that voluntary intake of large amounts of ethanol for 12 weeks only had a modest influence on ethanol-induced elevations of taurine in the rat.
Access provided by CONRICYT-eBooks. Download conference paper PDF
Similar content being viewed by others


Keywords
1 Introduction
Alcohol use and abuse is continuously causing disease, suffering as well as enormous costs for societies world-wide. If we are to prevent and treat alcohol use disorders, we need to understand the mechanisms underlying the development of addiction. One of the few common denominators among addictive substances is that they increase dopamine in the nucleus accumbens (nAc) (Di Chiara and Imperato 1988). This dopamine elevation has been linked to the rewarding and reinforcing properties (Koob 1992; Wise and Rompre 1989; Spanagel 2009), which is also why this neurotransmitter often is in focus in addiction research. Thus, understanding the mechanisms underlying ethanol’s ability to increase dopamine is of high importance.
Apart from dopamine ethanol is also known to increase extracellular levels of the amino acid taurine (De Witte et al. 1994; Dahchour et al. 1996). In addition, local administration of taurine in the nAc increases dopamine via the same neuronal circuitry as ethanol (Ericson et al. 2006) involving glycine receptors in the nAc as well as nicotinic acetylcholine receptors in the ventral tegmental area (Söderpalm and Ericson 2013). Interestingly, we also found that in order for ethanol to produce an elevation of dopamine in the nAc an increase of extracellular taurine is required (Ericson et al. 2011). This would then implicate taurine as a major participant in the reinforcing properties of ethanol. Further studies are needed to validate or discard this hypothesis.
Several studies have linked chronic drug/alcohol intake to decreased endogenous levels of dopamine in the mesolimbic dopamine system (Diana et al. 1993; Volkow et al. 2007; Feltmann et al. 2016). This adaptation appears also to be a common denominator for addictive drugs but the underlying mechanism to this event is not known. Whether chronic intake of ethanol also produces altered extracellular levels of taurine is not well studied. Thus, in the present study we aimed to measure basal and ethanol-induced extracellular levels of taurine and dopamine in naïve rats and in rats with a history of high voluntary ethanol intake.
2 Methods
2.1 Voluntary Ethanol Intake
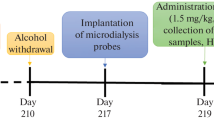
A total of 48 male Wistar rats (Taconic, Ejeby, Denmark) weighing 160–180 g at arrival were housed with regular 12 h light cycle and had ad lib access to tap water and standard rodent chow. Following 1 week of acclimatization to the animal facilities the rats were placed in single cages with continuous access to a bottle of water and a bottle of 6% (v/v) ethanol solution. After 3 weeks the rats with the highest ethanol consumption were selected (n = 8) and were administered 12% ethanol solution in the bottle in addition to the water bottle for another 8 weeks. The bottles were weighted twice a week and body weight was measured once a week (Fig. 1). All experiments were approved by the Ethics Committee for Animal Experiments, Gothenburg, Sweden.


Ethanol intake calculated as g/kg/day (a) and weight gain (b) for the eight selected ethanol high preferring rats placed in a voluntary ethanol consumption paradigm. The first 3 weeks the rats received 6% ethanol solution in addition to the regular water bottle and 12% ethanol solution for the remainder of the experiment
2.2 In Vivo Microdialysis
In vivo microdialysis was carried out in freely moving rats with or without a history of ethanol consumption (naïve). Dialysis probes were surgically implanted in the nAc as previously described (Clarke et al. 2014) using the coordinates (A/P: +1.85, M/L: −1.2 mm to bregma, V/D: −7.8 mm relative to dura; Paxinos and Watson 2007). Following 2 days of recovery the microdialysis experiment was initiated. The dialysis probe was connected to a microperfusion pump via a swivel and the probe was perfused with Ringer’s solution at a rate of 2 μL/min. Dialysate samples were collected every 15 min and analyzed for dopamine and taurine using two separate high-performance liquid chromatography systems as previously described (Lido et al. 2009). After a stable baseline had been obtained the rats received ethanol (300 mM) in the perfusion medium for 45 min or were maintained on Ringer’s solution (control animals). After the experiment the rats were sacrificed and the probe placement was verified.
2.3 Statistic Analysis
Statistical significance was determined by Student’s t-test. Each value was expressed as the mean ± SEM. Differences were considered statistically significant when the calculated P value was less than 0.05.
3 Results
3.1 Ethanol-Induced Elevation of nAc Taurine Is Modest in Ethanol-Experienced Rats
In the present study we wanted to further explore the ethanol-induced elevation of extracellular taurine in the nAc. As we previously demonstrated an ethanol-induced elevation of dopamine needs to be preceded by an increase of extracellular taurine (Ericson et al. 2011) possible adaptations following chronic ethanol intake may be of importance. Here we found that voluntary consumption of rather high amounts of ethanol (6.22 ± 0.59 g/kg/day in average calculated from the last three recordings) did not have a major impact on ethanol’s ability to increase extracellular levels of taurine (Fig. 2a).


Extracellular levels of taurine in the nAc as measured by in vivo microdialysis in naïve rats or rats with a high ethanol intake (n = 5–7). Ethanol (300 mM) was perfused via the dialysis probe as indicated by the black line in (a). Baseline levels of taurine is shown in (b), and the histogram in (c) highlights extracellular taurine levels at time-point 30. All data are expressed as mean ± SEM, * = p < 0.05 as calculated against the Ringer group; # = p < 0.05 as compared to the ethanol naïve group
Rats with a previous history of ethanol consumption responded with increased extracellular levels of taurine following acute ethanol administration (Ringer vs. ethanol exposed rats, p < 0.001). Only at time-point 30, i.e. 30 min following the initiation of ethanol perfusion, a blunted response was revealed (Fig. 2c; ethanol naïve vs. ethanol exposed, p = 0.006). When comparing baseline levels of extracellular taurine, we found that the two groups of naïve rats (Ringer and ethanol naïve groups) had significantly different extracellular levels of taurine (p = 0.004). However, when pooling the two groups with ethanol naïve rats their basal levels of taurine did not differ from rats with a previous history of voluntary ethanol consumption.
3.2 Ethanol-Induced Elevation of nAc Dopamine Is Lower in Ethanol-Experienced Rats
When analyzing dopamine, we found that the ethanol-induced elevation of dopamine was less prominent in rats chronically exposed to ethanol (Fig. 3a). Although the ethanol high preferring rats did respond with a significant elevation of dopamine following acute exposure (Ringer vs. ethanol exposed, p = 0.042) ethanol was able to produce a larger increase in the ethanol naïve rats (Fig. 3c; ethanol naïve vs. ethanol exposed, p = 0.004). When comparing basal levels of dopamine, no differences were found between the groups (Ringer vs. ethanol naive rats, p = 0.427, Ringer vs. ethanol exposed rats, p = 0.210).


Extracellular levels of dopamine in the nAc as measured by in vivo microdialysis in naïve rats or rats with a high ethanol intake (n = 6–7). Ethanol (300 mM) was perfused via the dialysis probe as indicated by the black line in (a). Baseline levels of dopamine is shown in (b), and the histogram in (c) highlights extracellular dopamine levels at time-point 30. All data are expressed as mean ± SEM, * = p < 0.05 as calculated against the Ringer group; # = p < 0.05 as compared to the ethanol naïve group
4 Discussion
In the present study using rats with almost 3 months of high voluntary ethanol consumption, we found only very modest alterations of taurine in comparison to ethanol naïve rats. Ethanol has the ability to increase extracellular taurine both in naïve rats and rats with 3 months of high ethanol intake. This is in line with a study from Lallemand and co-workers (2011) where they only found a minor reduction in ethanol’s ability to increase taurine in rats following 3 weeks of forced ethanol intake (gavage). This would then indicate that the ethanol-induced increase of extracellular taurine is a rather robust phenomenon or that a longer period of alcohol exposure is needed to induce adaptations. When dopamine was analyzed in the same samples we found a more pronounced difference in response to acute ethanol administration between naïve rats and rats that had consumed ethanol for 12 weeks. The ethanol-experienced rats displayed a significant but modest dopamine elevation. It is not possible to determine whether the diminished dopamine elevation is due to adaptations following ethanol consumption or if the animals voluntarily choose to consume large amounts of ethanol due to an unresponsive endogenous dopamine system. It has also been suggested that the time point for measuring dopamine following ethanol exposure is delicate. Immediately after withdrawal dopamine levels are low whereas after a few weeks of abstinence the dopamine levels are higher than normal (Hirth et al. 2016). Further studies addressing this question is needed.
Low levels of endogenous striatal dopamine were suggested to both be a cause for, as well as consequence of, excessive ethanol intake (Kashem et al. 2012). When monitoring the basal dopamine levels in the present study we found no differences between naïve animals and those that hade been drinking large amounts of ethanol. Since the naïve rats did not have the opportunity to demonstrate whether they were ethanol preferring rats or not, a spontaneously low dopaminergic tone could have masked any ethanol-induced adaptations in dopamine transmission. However, including more animals and utilizing another method (such as no-net-flux microdialysis) could in future studies shed light on these issues.
Analyzing basal levels of taurine and dopamine in the same samples produced an interesting result. The two groups with naïve rats differed substantially in basal levels of extracellular taurine. This would indicate that the basal extracellular levels of taurine can be very different between individuals, an interesting phenomenon that should be investigated more in depth. In the present study, although statistically significant findings, we are unable to make any conclusions regarding possible adaptations of a taurinergic tone following ethanol consumption. We will continue to investigate differences in extracellular taurine levels in relation to ethanol consumption and ethanol-induced dopamine elevation.
5 Conclusion
In the present study we found that ethanol high preferring rats, consuming ethanol for 12 weeks, displayed a modestly attenuated ethanol-induced elevation of taurine and a diminished increase of extracellular dopamine. Whether this is due to ethanol-induced adaptations or if this is an underlying factor as to why some rats choose to voluntarily consume large amounts of ethanol remains to be established.
Abbreviations
- nAc :
-
Nucleus accumbens
References
Clarke RB, Adermark L, Chau P, Soderpalm B, Ericson M (2014) Increase in nucleus accumbens dopamine levels following local ethanol administration is not mediated by acetaldehyde. Alcohol Alcohol 49:498–504
Dahchour A, Quertemont E, De Witte P (1996) Taurine increases in the nucleus accumbens microdialysate after acute ethanol administration to naive and chronically alcoholised rats. Brain Res 735:9–19
De Witte P, Dahchour A, Quertemont E (1994) Acute and chronic alcohol injections increase taurine in the nucleus accumbens. Alcohol Alcohol Suppl 2:229–233
Di Chiara G, Imperato A (1988) Drugs abused by humans preferentially increase synaptic dopamine concentrations in the mesolimbic system of freely moving rats. Proc Natl Acad Sci U S A 85:5274–5278
Diana M, Pistis M, Carboni S, Gessa GL, Rossetti ZL (1993) Profound decrement of mesolimbic dopaminergic neuronal activity during ethanol withdrawal syndrome in rats: electrophysiological and biochemical evidence. Proc Natl Acad Sci U S A 90(17):7966–7969
Ericson M, Molander A, Stomberg R, Soderpalm B (2006) Taurine elevates dopamine levels in the rat nucleus accumbens; antagonism by strychnine. Eur J Neurosci 23:3225–3229
Ericson M, Chau P, Clarke RB, Adermark L, Soderpalm B (2011) Rising taurine and ethanol concentrations in nucleus accumbens interact to produce dopamine release after ethanol administration. Addict Biol 16:377–385
Feltmann K, Fredriksson I, Wirf M, Schilström B, Steensland P (2016) The monoamine stabilizer (−)-OSU6162 counteracts downregulated dopamine output in the nucleus accumbens of long-term drinking Wistar rats. Addict Biol 21:438–449
Hirth N, Meinhardt MW, Noori HR, Salgado H, Torres-Ramirez O, Uhrig S, Broccoli L, Vengeliene V, Roßmanith M, Perreau-Lenz S, Köhr G, Sommer WH, Spanagel R, Hansson AC (2016) Convergent evidence from alcohol-dependent humans and rats for a hyperdopaminergic state in protracted abstinence. Proc Natl Acad Sci U S A 113:3024–3029
Kashem MA, Ahmed S, Sarker R, Ahmed EU, Hargreaves GA, McGregor IS (2012) Long-term daily access to alcohol alters dopamine-related synthesis and signaling proteins in the rat striatum. Neurochem Int 61:1280–1288
Koob GF (1992) Neural mechanisms of drug reinforcement. The neurobiology of drug and alcohol addiction, vol 654, PW Kalivas and HH Samson. New York. pp 171–191
Lallemand F, Ward RJ, De Witte P, Verbanck P (2011) Binge drinking +/− chronic nicotine administration alters extracellular glutamate and arginine levels in the nucleus accumbens of adult male and female Wistar rats. Alcohol Alcohol 46:373–382
Lido HH, Stomberg R, Fagerberg A, Ericson M, Soderpalm B (2009) The glycine reuptake inhibitor org 25935 interacts with basal and ethanol-induced dopamine release in rat nucleus accumbens. Alcohol Clin Exp Res 33:1151–1157
Paxinos G, Watson C (2007) The Rat brain in stereotaxic coordinates. Academic Press, Sydney
Söderpalm B, Ericson M (2013) Neurocircuitry involved in the development of alcohol addiction: the dopamine system and its access points. Curr Top Behav Neurosci 13:127–161
Spanagel R (2009) Alcoholism: a systems approach from molecular physiology to addictive behavior. Physiol Rev 89:649–705
Volkow ND, Wang GJ, Telang F, Fowler JS, Logan J, Jayne M, Ma Y, Pradhan K, Wong C (2007) Profound decreases in dopamine release in striatum in detoxified alcoholics: possible orbitofrontal involvement. J Neurosci 27:12700–12706
Wise RA, Rompre PP (1989) Brain dopamine and reward. Annu Rev Psychol 40:191–225
Acknowledgements
The authors are thankful for the technical assistance from Mrs. Rosita Stomberg and Mahmod Panahi. This work was supported by Swedish Medical Research Council (Diary numbers 2014–3888, 2014–3887, and 2015-02894) and the Alcohol Research Council of the Swedish Alcohol Retailing Monopoly.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2017 Springer Science+Business Media B.V.
About this paper
Cite this paper
Ericson, M., Ulenius, L., Adermark, L., Söderpalm, B. (2017). Minor Adaptations of Ethanol-Induced Release of Taurine Following Chronic Ethanol Intake in the Rat. In: Lee, DH., Schaffer, S.W., Park, E., Kim, H.W. (eds) Taurine 10. Advances in Experimental Medicine and Biology, vol 975. Springer, Dordrecht. https://doi.org/10.1007/978-94-024-1079-2_19
Download citation
DOI: https://doi.org/10.1007/978-94-024-1079-2_19
Publisher Name: Springer, Dordrecht
Print ISBN: 978-94-024-1077-8
Online ISBN: 978-94-024-1079-2
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)