
Abstract
The effects of climate change on insect outbreaks, wildfire, invasive species, and pathogens in forest ecosystems will greatly exceed the effects of warmer temperature on gradual changes in forest processes. Increased frequency and extent of these disturbances will lead to rapid changes in vegetation age and structure, plant species composition, productivity, carbon storage, and water yield. Insect outbreaks are the most pervasive forest disturbance in the United States, and rapid spread of bark beetles in the western United States has been attributed to a recent increase in temperature. Wildfire area burned has increased in recent decades, although frequency and severity have not changed, and is expected to greatly increase by 2050 (at least twice as much area burned annually in the West). More frequent occurrence of fire and insects will create landscapes in which regeneration of vegetation will occur in a warmer environment, possibly with new species assemblages, younger age classes, and altered forest structure. Increased fire and insects may in turn cause more erosion and landslides. Invasive plant species are already a component of all forest ecosystems, and a warmer climate will likely facilitate the spread of current and new invasives, particularly annuals that compete effectively in an environment with higher temperature and frequent disturbance. The interaction of multiple disturbances and stressors, or stress complexes, has the potential to alter the structure and function of forest ecosystems, especially when considered in the context of human land-use change. Occurring across large landscapes over time, these stress complexes will have mostly negative effects on ecosystem services, requiring costly responses to mitigate them and active management of forest ecosystems to enhance resilience.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
1 Introduction
Disturbances such as wildfire, insect outbreaks, pathogens, invasive species, drought, and storms are part of the ecological history of most forest ecosystems, influencing vegetation age and structure, plant species composition, productivity, carbon (C) storage, water yield, nutrient retention, and wildlife habitat. Climate influences the timing, frequency, and magnitude of disturbances (Dale et al. 2001). As the climate continues to change, we expect increased disturbance through more frequent extreme weather events, including severe drought, wind storms, and ice storms. Indirect effects may amplify these changes, with conditions that favor wildfire, insects, pathogens, and invasive species.
If frequency and severity of disturbances increase in the future, they will almost certainly have a bigger impact on forest ecosystems than gradual changes in other forest processes in response to higher temperature (see Chap. 3). This will lead to rapid changes in forest structure and function. It will also create landscapes in which regeneration of vegetation will occur in a warmer environment, possibly with new competitive relationships among species. In this way, the indirect effects of climate change in forest ecosystems may be more important than direct effects.
2 Wildfire
Climate and fuels are the two most important factors controlling patterns of wildfire within forest ecosystems. Climate controls the frequency of weather conditions that promote fire, whereas the amount and arrangement of fuels influence fire intensity and spread. Climate influences fuels on longer time scales by shaping species composition and productivity (Marlon et al. 2008; Power et al. 2008), and large-scale climatic patterns such as the El Niño Southern Oscillation, Pacific Decadal Oscillation, Atlantic Multidecadal Oscillation, and Arctic Oscillation are important drivers of forest productivity and susceptibility to disturbance (Duffy et al. 2005; Collins et al. 2006; Fauria and Johnson 2006; Kitzberger et al. 2007).
Current and past land use, including timber harvest, forest clearing, fire suppression, and fire exclusion through grazing (Swetnam and Betancourt 1998; Allen et al. 2002) have affected the amount and structure of fuels in the United States. For example, in montane forests in the Southwest (Allen et al. 2002) and other dry forests in the interior West, removal of fine fuels by grazing and fire suppression have increased the number of trees and amount of fuels; these forest conditions have increased fire size and intensified fire behavior. In colder and wetter forests in the western United States, such as subalpine forests in Yellowstone National Park and forests in the maritime Northwest, grazing and fire suppression have not altered fire regimes as extensively. Forests in the northeastern United States (Foster et al. 2002) and the upper Midwest developed after widespread timber harvest, land clearing, and forest re-growth after land abandonment. Compared to other regions of the United States, forests in the Northeast and upper Midwest burn less often and with smaller fires. Forests in the southeastern United States are often managed for timber, and prescribed fire is generally more prevalent than uncontrolled ignitions (National Interagency Coordination Center 2011). Prescribed fire is applied every 2–4 years in some fire-dependent ecosystems in the Southeast (Mitchell et al. 2006). Fire suppression and deer herbivory in the central hardwoods section of the eastern United States have pushed the composition toward more mesic and fire intolerant species (e.g., from oak dominated to maple dominated) (Nowacki and Abrams 2008).
Weather remains the best predictor of how much area will burn, despite changes in land use and the resulting effects on fuels. Correlations between weather and either area burned by fire or number of large fires are similar for both pre-settlement fires and fires of the last few decades. These syntheses of fire-weather relationships for both pre-settlement and modern records exist in several subregions of the West (Northwest: Heyerdahl et al. 2002, 2008a; Hessl et al. 2004. Southwest: Swetnam and Betancourt 1998; Grissino-Mayer and Swetnam 2000. Northern Rocky Mountains: Heyerdahl et al. 2008b. Westwide: Westerling et al. 2003, 2006; Littell et al. 2009) and the East (Hutchinson et al. 2008). Pre-settlement fire-weather relationships are derived from trees scarred by fires or age classes of trees established after fire and independently reconstructed climate; modern fire-weather comparisons are derived from observed fire events and observed weather in seasons leading up to the fire. Drought and increased temperature promote large fires, but effects differ by forest and region (Westerling et al. 2003; Littell et al. 2009). Weather can also influence fire through higher precipitation, increasing understory vegetation growth, which later becomes fuel (Swetnam and Betancourt 1998; Littell et al. 2009). Increased temperature and altered precipitation also affect fuel moisture and the length of time during which wildfires can burn during a given year.
The potential effects of climate change on forest fire area have been assessed using statistical models that project area burned from climatic variables, and by using global climate models to project future climatic variables (Westwide: McKenzie et al. 2004; Spracklen et al. 2009; Littell et al. 2010. Pacific Northwest: Littell et al. 2010; Yellowstone region: Westerling et al. 2011). Estimated future changes in annual area burned in the West ranges from declines of 80 % to increases greater than 500 %, depending on the region, timeframe, methods, and climate model/emission scenario (Bachelet et al. 2001). Future fire potential is expected to increase in summer and autumn from low to moderate in eastern regions of the South, and from moderate to high levels in western regions of the South (Liu et al. 2010).
The risk posed by future fire activity in a changing climate can be assessed by its likely effects on human and ecological systems. At the wildland-urban interface (WUI), higher population and forest density have created forest conditions that are likely to experience more area burned and possibly greater fire severity than in the historical record. Fire risk is likely to increase in a warmer climate because of the longer duration of the fire season, and the greater availability of fuels if temperature increases and precipitation does not sufficiently increase to offset summer water balance deficit. Where fuels management is common, forest fuel reduction and restoration to pre-settlement tree density and surface fire regimes help mitigate fire hazard under current and future climatic conditions. Finally, future fire risk may depend on whether extreme fire weather conditions will change in step with monthly to seasonal climate changes. Even if fire weather and ignitions do not change, it is likely that risk driven only by seasonal climate changes will increase; particularly in the WUI and managed forests, where fire has been historically rare or fully suppressed. The current increase in annual area burned may be partially related to increased fuels in frequent-fire forest types, in addition to more frequent weather conditions conducive to fire.
The effects of climate change intersecting with increased fuel loads in frequent-fire forests will be an exceptional challenge for resource managers on both public and private lands. As noted above, active management is highly effective in reducing fuel quantity and continuity, thus reducing fire intensity and mortality in the forest overstory (see Chap. 9). Prescribed fire is applied routinely and extensively in pine forests in the Southeast, but funding for fuel treatment in Western forests is sufficient to treat only a small portion of the landscape that currently has elevated fuels. Fire suppression is currently a large proportion of federal agency budgets (approximately 50 % for the U.S. Forest Service). If area burned does in fact increase by 100 % or more in future decades, this will pose a major budgetary and policy issue, and create challenges in managing landscapes increasingly occupied by younger forests.
3 Insects and Pathogens
3.1 General Concepts
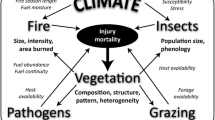
Biotic disturbances are natural features of forests that play key roles in ecosystem processes (Adams et al. 2010; Boon 2012; Hicke et al. 2012a). Epidemics by forest insects and pathogens affect more area and result in greater economic costs than other forest disturbances in the United States (Dale et al. 2001). By causing local to widespread tree mortality or reductions in forest productivity, insect and pathogen outbreaks have broad ecological and socioeconomic effects (Tkacz et al. 2010; Pfeifer et al. 2011). The first National Climate Assessment (Melillo et al. 2001) projected increased disturbance in forests, especially from insects, and especially from bark beetles, because of their high physiological sensitivity to climate, short generation times, high mobility, and explosive reproductive potential. These projections have been upheld, and current observations suggest that disturbances are occurring more rapidly than imagined a decade ago. Understanding how these disturbances are influenced by climate change is therefore critical for quantifying and projecting effects (Fig. 4.1).

General pathways by which atmospheric changes associated with increasing greenhouse gases can influence forest disturbance from insects and pathogens. CO 2 carbon dioxide, CH 4 methane
The powerful general effect of temperature on insects and pathogens is well known (Gillooly et al. 2002). Clear examples exist of climatic effects on insects, yet the most important insects and pathogens in American forests remain poorly studied with respect to how interactions with climate will affect forests. Climatic warming can influence biotic disturbances of forests through effects on (1) the physiology of insects and pathogens that cause changes in their abundance and distribution, (2) tree defenses and tolerance, and (3) interactions of insects and pathogens with enemies, competitors, and mutualists (Fig. 4.1). Higher temperature has reduced winter mortality of insects, increased their range northward (Trân et al. 2007; Paradis et al. 2008; Safranyik et al. 2010), and increased their development rate during the growing season (Gillooly et al. 2002; Bentz et al. 2010). Temperature increases can also alter phenology, such as bringing leaf maturation into synchrony with insect feeding (Jepsen et al. 2011) or changing the life cycle synchrony of bark beetles, which depend on mass attack to overwhelm tree defenses (Powell et al. 2000; Friedenberg et al. 2007; Bentz et al. 2010).
A broader set of atmospheric drivers affects tree defenses against, and tolerance to, herbivores and pathogens (Lindroth 2010; Sturrock et al. 2011). Deficiencies of water or mineral nutrients can both increase and decrease tree defenses, depending on the severity of the deficiency, biochemical pathways, and the type of defense (Lombardero et al. 2000; Breshears et al. 2005; Worrall et al. 2008). In addition, tree mortality from severe drought may facilitate an increase in bark beetles, which then become sufficiently abundant to successfully attack healthy trees (Greenwood and Weisberg 2008; Raffa et al. 2008). Information on the effects of climate on tree-pathogen interactions is sparse, despite a theoretical expectation for temperature and moisture to have significant effects (Grulke 2011; Rohrs-Richey et al. 2011; Sturrock et al. 2011).
Outbreak dynamics of forest insects respond to interactions between herbivores and their enemies (Dwyer et al. 2004), and these interactions should be sensitive to temperature (Berggren et al. 2009; Klapwijk et al. 2012), but empirical studies are rare (Siegert et al. 2009). Similarly, for the many forest insects that involve mutualisms with fungi, it is logical that outbreak dynamics will be sensitive to climatic effects on the mutualism (Lombardero et al. 2000; Hofstetter et al. 2007; Six and Bentz 2007; Evans et al. 2011).
3.2 Climate and Biotic Disturbances
3.2.1 Bark Beetles
Multiple species of indigenous bark beetles affect millions of hectares of coniferous forests in North America. Major species include mountain pine beetle (Dendroctonus ponderosae Hopkins), the most important disturbance agent of pines in the western United States (Box 4.1; see Chap. 1); southern pine beetle (D. frontalis Zimmermann) in pine forests of the southeastern United States (Box 4.2), and spruce beetle (D. rufipennis Kirby). In the early 2000s, severe drought, coupled with several species of bark beetles, killed trees of several conifer species in the Southwest (Ganey and Vojta 2011), most notably pinyon pine (Pinus edulis Engelm.) attacked by pinyon ips (Ips confusus LeConte) across 1.2 million ha (Breshears et al. 2005).

3.2.3 Defoliating Insects
Defoliating insects are a continentally important biotic disturbance in American forests. For example, western spruce budworm (Choristoneura occidentalisFreeman) is currently important in the West (USDA FS 2010), and Eastern boreal forests have been affected by many cycles of spruce budworm (Archips fumiferana Clemens) outbreaks (Candau and Fleming 2005). Other important defoliators include tussock moths, tent caterpillars, gypsy moths, and jack pine budworm (A. pinus Freeman). Most defoliating insects are indigenous to American forests, and many have cyclical outbreak dynamics involving predators, parasitoids, and pathogens (Dwyer et al. 2004).
Climatic effects on these predator–prey interactions remain largely unstudied (Klapwijk et al. 2012). In general, it is less clear (compared to what is known about bark beetles) how climatic patterns influence the frequency, extent, and geographic distribution of defoliators in American forests. There is limited evidence in some forest systems of climatic effects on winter populations (Thomson et al. 1984; Kemp et al. 1985; Williams and Liebhold 1995a; but see Reynolds et al. 2007; Thomson and Benton 2007), drought stress of host trees (Williams and Liebhold 1995b; Campbell et al. 2006), and phenological synchronization of larval emergence and bud break (Thomson et al. 1984). Considerable uncertainty remains about future responses of defoliators to climate change (Dukes et al. 2009; Rodenhouse et al. 2009). Hemlock woolly adelgid (Adelges tsugae Annand), a non-native, stem-feeding insect, has been spreading in the eastern United States (Box 4.3).
3.2.5 Plant Pathogens
We identified 21 plant pathogens that are notable agents of disturbance in U.S. forests and may respond to climate change. Climatic effects on these pathogens are generally not well studied, but we expect that some of these pathogens will be affected directly by climatic influences on sporulation and infection, indirectly by predisposing trees to infection, or both (Sturrock et al. 2011). For pathogens that involve associations with insects, climatic effects on the animal associates may also be important.
A few cases of climate-pathogen interactions have been documented. For example, Swiss needle cast (Phaeocryptopus gaeumannii T. Rohde), a native foliar pathogen in the Northwest, is influenced by winter warming and spring precipitation. Climatic projections suggest an increase in Swiss needle cast distribution and severity (Stone et al. 2008). The susceptibility of alder to a cankering pathogen is related to the phenology of the plant, the pathogen, and water availability (Grulke 2011; Rohrs-Richey et al. 2011). Outbreaks of some virulent invasive pathogens can also be enhanced by climate (e.g., sudden oak death; Sturrock et al. 2011), whereas others are not very sensitive to climate (Garnas et al. 2011b).
The potential effects of climate change on root pathogens are difficult to project (Ayres and Lombardero 2000), but it will be important to understand this relationship because endemic root diseases are widespread and often have a major influence on forest dynamics and management. One would expect root diseases to be affected by both the distribution of host species and the effects of a changing climate on susceptibility of host species and prevalence of fungal pathogens. If a warmer climate increases physiological stress in a particular tree species, then it may be less resistant to some root diseases, potentially causing lower tree vigor, higher mortality in mature trees and seedlings, and lower C storage. Although some initial modeling of future changes in root pathogens has been attempted (Armillaria spp.; Klopfenstein et al. 2009), geographic specificity for host-pathogen relationships is highly uncertain based on current knowledge.
3.2.6 Non-native and Emerging Insects and Pathogens
Invasive, non-native insects and pathogens are becoming an increasingly important component of forest disturbance (Lovett et al. 2006; Seppälä et al. 2009), and warming and precipitation shifts associated with climate change can affect forest vulnerability (Paradis et al. 2008; Sturrock et al. 2011). For example, the geographic range and incidence of dothistroma needle blight (Dothistroma septosporum [Dorog.] M. Morelet and D. pini Hulbary), which reduces growth of many conifers by causing premature needle defoliation, may shift with changing precipitation patterns (Woods et al. 2010).
At present, the primary cause of biological invasions is global commerce. However, increasing temperatures are generally expanding the geographic zones where potential invasive species could survive and reproduce if they arrive, for example, at ports of entry on the Eastern seaboard and in the Great Lakes waterway. The potential for global, climate-driven increases in invasion risks has prompted international organizations to discuss changes in trade restrictions to manage associated phytosanitation risks (Standards and Trade Development Facility 2009).
Outbreaks of lesser known forest insects have recently occurred in U.S. forests. Aspen leaf miner (Phyllocnistis populiella Chambers), which reduces longevity of aspen leaves, has damaged 2.5 million ha of quaking aspen (Populus tremuloides Michx.) in Alaska since 1996 (Wagner et al. 2008). Large areas of willows were damaged during two eruptive outbreaks of the willow leafblotch miner (Micurapteryx salicifoliella Chambers) in the 1990s in two major river drainages in Alaska (Furniss et al. 2001); outbreaks of this leaf miner had not been previously reported. Substantial defoliation by Janet’s looper (Nepytia janetae Rindge) of stressed trees in Southwestern spruce-fir forests was preceded by uncharacteristically warm winters; defoliation by Janet’s looper encouraged attack by opportunistic bark beetles. These examples demonstrate that previously rare native insects that displayed new eruptive behavior and caused notable forest disturbances.
3.3 Effects and Interactions with Other Disturbances
Through their effects on tree growth and mortality, insects and pathogens have broad effects on ecosystem processes (see Chap. 3). Insects and pathogens, by virtue of their host preferences, can alter tree species composition within stands, remove most host trees from some landscapes (Lovett et al. 2006), and modify forest types (e.g., from conifers to hardwoods) (Veblen et al. 1991; Orwig et al. 2002; Collins 2011). Forests typically shift toward younger, smaller trees after biotic disturbances (Ylioja et al. 2005; Garnas et al. 2011a; Tchakerian and Couslon 2011), which can affect wildlife habitat and biodiversity by quickly modifying multiple trophic levels (Chan-McLeod 2006; Drever et al. 2009). Both positive and negative effects occur depending on species, time since disturbance, surviving vegetation, ecosystem type, and spatial extent of outbreak.
Fire and biotic disturbances interact in several ways. Fires lead to younger stands that may be less susceptible to attack, and dead trees provide a food resource for some insects and pathogens (Parker et al. 2006). Insect-killed trees influence fuels and therefore fire behavior, although the effect depends on a number of factors, including the number of attacked trees within a stand and time since outbreak (e.g., Ayres and Lombardero 2000; Hicke et al. 2012b; Jenkins et al. 2008; Simard et al. 2011), and fire-induced increases in tree defenses can mitigate bark beetle risks (Lombardero and Ayres 2011).
Extreme soil water deficits (drought) that reduce tree growth might also reduce tree defenses to insects and pathogens (Bentz et al. 2009; Sturrock et al. 2011), although previous studies suggest there may be either no effect (Gaylord et al. 2007; McNulty et al. 1997) or the opposite effect (Lombardero et al. 2000). Drought also facilitates population increases of western bark beetles. Some aggressive species such as mountain pine beetle are able to maintain epidemics after return to normal conditions, whereas others such as pinyon ips decline with alleviation of drought stress (Raffa et al. 2008).
Insects and pathogens clearly affect the economic value of forests that are intended for harvest for wood products, and direct economic effects occur for tree removal and replacement, such as the $10 billion spent after emerald ash borer (Agrilus planipennis Fairmaire) infestations (Kovacs et al. 2010, 2011). A more complete valuation of socioeconomic effects is challenging because it is difficult to quantify all ecosystem services, especially those with non-market values (Holmes et al. 2010). Regions with dead and dying trees have reduced aesthetic value (Sheppard and Picard 2006) and may have reduced housing prices (Holmes et al. 2010; Price et al. 2010).
4 Invasive Plants
4.1 Introduction
Invasive plants are recent introductions of non-native, exotic, or nonindigenous species that are (or have the potential to become) successfully established or naturalized, and that spread into new localized natural habitats or ecoregions with the potential to cause economic or environmental harm (Lodge et al. 2006). This definition of “invasive” (1) does not consider native species that have recently expanded their range, such as juniper (Juniperus spp.) in the western United States (Miller and Wigand 1994; Miller et al. 2005), (2) involves defined temporal and spatial scales, and (3) considers social values related to economic and environmental effects.
An estimated 5,000 nonnative plant species exist in U.S. natural ecosystems (Pimentel et al. 2005) (Table 4.1). The effects of invasive plants include reduced native biodiversity, altered species composition, loss of habitat for dependent species (e.g., wildlife), changes in biogeochemical cycling, changes in ecosystem water use, and altered disturbance regimes. Billions of dollars are spent every year to mitigate invasive plants or control their effects (Pimentel et al. 2005). Negative environmental effects are scale-dependent (Powell et al. 2011), with some subtle beneficial properties (Sage et al. 2009) on ecosystem function (Myers et al. 2000; Zavaleta et al. 2001). For example, some consider species in the genus Tamarix to be among the most aggressively invasive and detrimental plants in the United States (Stein and Flack 1996), but others point out benefits, including sediment stabilization and the creation of vertebrate habitat in riparian areas that can no longer support native vegetation (Cohn 2005).
The spatial extent of many invasive plants at any point in time has been difficult to determine, limiting assessment of overall consequences. One assessment (Duncan et al. 2004) for the western United States indicates that 16 invasive plants account for most current invasive plant problems. Centaurea species are particularly widespread and persistent in the West. Cogongrass (Imperata cylindrica [L.] Raeusch.), which has invaded extensive forested areas of the Southeast, is considered to be one of the most problematic invasive plants in the world (Box 4.4). Mountain ecosystems tend to have fewer invasive plant species than other regions because of a short growing season, limited settlement history, relatively low frequency of seed sources, and prevalence of closed-canopy conifer forests that limit light in the understory and acidify the soil (Parks et al. 2005).

4.2 Interactions Between Climate Change and Plant Invasion
Plant invasions can be influenced by warmer temperatures, earlier springs and earlier snowmelt, reduced snowpack, changes in fire regimes, elevated nitrogen (N) deposition, and elevated CO2 concentrations. The responses of invasive plants to climate change should be considered separately from those of native species, because invasive plants (1) have characteristics that may differ from native species, (2) can be highly adaptive (Sexton et al. 2002), (3) have life history characteristics that facilitate rapid population expansion, and (4) often require different management approaches than for native species (Hellmann et al. 2008). Successful invasion of areas dominated by native plants depends on environment, disturbance, resource availability, biotic resistance, and propagule pressure (Davis et al. 2000; D’Antonio et al. 2001; Levine et al. 2004; Eschtruth and Battles 2009; Pauchard et al. 2009). Climate change may influence all of these drivers of invasion, with high variability across space and time.
4.2.1 Temperature, Precipitation, and CO2
Climate change will alter the abiotic conditions under which plant species can establish, survive, reproduce, and spread. These effects are expected to increase plant stress and decrease survival in the drier, warmer, and lower elevation portions of species ranges (Allen and Breshears 1998). Abiotic factors probably constrain the range of many invasive plants and limit their successful establishment (Alpert et al. 2000; Pauchard et al. 2009). With climate change, however, new habitat, once too cold or wet, may become available, enabling plants to survive outside their historical ranges and expand beyond their current ranges.
Many native plants are projected to move northward or upward in elevation with climate change. Examples of invasive plants projected to follow this pattern are rare, but information on species tolerances provides insight on potential responses. For example, the northern limit of Japanese barberry (Berberis thunbergii DC.), an invasive shrub in the eastern United States, is determined by low temperature tolerance, the southern limit by cold stratification requirements for germination, and the western limit by drought tolerance (Silander and Klepeis 1999). The widespread invasive tree of heaven (Ailanthus altissima [P. Mill.] Swingle) is limited by cold and prolonged snow cover to lower mountain slopes, but it may be able to colonize during several successive years of mild climate, conditions that may become more common under climate change (Miller 1990). Soil water availability and regional changes in climatic water balance may be important for plant invasions, particularly at lower elevations (Chambers et al. 2007; Crimmins et al. 2011). Species growth, productivity, and reproduction may also change as climatic conditions change. For example, invasive plants may be better able to adjust to rapid changes in abiotic conditions by tracking seasonal temperature trends and shifting their phenologies (e.g., earlier spring warming) (Willis et al. 2010).
Increased productivity in response to elevated CO2 has been documented under controlled conditions for several invasive plant species, including cheatgrass (Bromus tectorum L.) (Box 4.4), Canada thistle (Cirsium arvense [L.] Scop.), spotted knapweed (Centaurea melitensis L.), yellow star-thistle (C. solstitialis L.), and kudzu (Pueraria montana [Lour.] Merr.) (Dukes et al. 2011; Ziska and Dukes 2011; Ziska and George 2004). Response to CO2 enrichment is less predictable when plants are grown in the field (Dukes and Mooney 1999; Ziska and Dukes 2011), where response may be limited by nutrients and water availability. Carbon dioxide enrichment can also increase water use efficiency, which can partially ameliorate conditions associated with decreased water availability, particularly for C3 plants (Eamus 1991). This phenomenon may be partially responsible for global patterns of encroachment of C3 plants in grasslands dominated by C4 plants or mixed species (Bond and Midgley 2000).
4.2.2 Disturbance and Resource Availability
Disturbances such as fire, landslides, volcanic activity, logging, and road building open forest canopies, reduce competition, and expose mineral soil, increasing light and nutrient availability. Invasive plants are generally well adapted to use increased resources. Fluctuating resource availability, coinciding with available propagules, facilitates regeneration and establishment of invasive species associated with forest development after disturbance (Halpern 1989; Davis et al. 2000; Parks et al. 2005). Opportunities for invasions may also be created by forest thinning, fuel treatments, and biofuel harvesting (Bailey et al. 1998; Silveri et al. 2001; Nelson et al. 2008). However, the spatial extent of invasions may be limited (Nelson et al. 2008), especially for shade intolerant species in closed-canopy Western forests.
The reintroduction of fire is a high priority for restoration and management of fire-adapted forests such as ponderosa pine (Pinus ponderosa Douglas ex P. Lawson & C. Lawson), longleaf pine (P. palustris Mill.), and loblolly pine (P. taeda L.). Invasive plants, especially annual grasses (Box 4.4), can spread rapidly after fire, particularly in high-severity burns (D’Antonio 2000; Kerns et al. 2006; Keeley and McGinnis 2007). Forest sites treated with prescribed fire, which are often near the wildland-urban interface and roads, are also well positioned for invasive plant introduction and spread (Keeley et al. 2003).
The success of plant invasions is regulated by competition from resident plants (Levine 2000; Seabloom et al. 2003), and land managers can alter post-disturbance (logging, fire) invasive establishment by seeding to increase native plant competition. Although native plant competition can be overwhelmed by invasive plant seed abundance (D’Antonio et al. 2001; Lonsdale 1999), resistance related to soil properties is more likely to withstand seed abundance. Native plant competition with invasive plants can also be affected by the effects of predation, herbivory, and pathogens associated with native species. Native plant competition may change as temperature and ambient CO2 increase; numerous studies have documented that weedy plants are more productive in an elevated CO2 environment (Ziska and George 2004).
Propagule pressure, which includes seed size, numbers, and temporal and spatial patterns, is perhaps the most important driver of successful invasions in forest ecosystems (Tilman 1997; Colautti et al. 2006; Eschtruth and Battles 2009; Simberloff 2009). For invasive plants, propagule pressure is largely controlled by factors other than climate. For example, the most critical factors projecting plant invasion in eastern hemlock (Tsuga canadensis [L.] Carrière) forests in the eastern United States are overstory canopy disturbance and propagule pressure (Eschtruth and Battles 2009). However, little is known about how biotic and abiotic resistance factors interact with propagule supply to influence exotic plant invasion (D’Antonio et al. 2001; Lonsdale 1999).
Atmospheric CO2 may influence seed production through enhanced flowering under elevated CO2, increasing the probability that a smaller seed can establish a viable population (Simberloff 2009). Of greater concern is how climate change may alter human activities that transfer seeds. For example, climate change could alter tourism and commerce, enhance survival of seeds during transport (Hellmann et al. 2008), and shift recreation to higher elevations. Changes in atmospheric circulation patterns could also alter wind-dispersed species, allowing new species to arrive in areas that previously had few seeds.
Climate change will affect invasive plants in forests because of the potential for increased ecological disturbance, effects of warming on species distributions, enhanced competitiveness of invasive plants owing to elevated CO2, and increased stress to native species and ecosystems (Dukes and Mooney 1999; Breshears et al. 2005; Pauchard et al. 2009; Ziska and Dukes 2011). Warming will increase the risk of invasion in temperate mountainous regions because cold temperature has tended to limit the establishment of invasive plants.
Empirical models suggest that a warmer climate could result in both range expansion and contraction for common invasive plants (Sasek and Strain 1990; Pattison and Mack 2008; Bradley et al. 2009; Kerns et al. 2009), although these types of species distribution models do not account for species ecophysiology and biotic interactions. Process-based models may ultimately prove more robust for prediction, although model parameters are quantified from experimental data or the research literature, which themselves have uncertainties.
For management responses to plant invasions to be cost effective and successful, assertive action is needed in the early phase of invasion. A potentially useful approach is a climate change-based modification of the Early Detection and Rapid Response System (National Invasive Species Council 2001). For example, risk assessment could be done over broader geographic areas than has been performed in the past (Hellman et al. 2008). Unfortunately, some biocontrol methods may no longer be effective in a warmer climate (Hellmann et al. 2008), and some herbicides are less effective on plants grown in elevated CO2 (Ziska and Teasdale 2000). The successful control of invasive plants over large forest landscapes will depend on knowledge about resistance of native species to invasion and our ability to limit propagule pressure.
5 Erosion, Landslides, and Precipitation Variability
Based on analysis of recent climate records and the projections of climate change simulations, hydroclimatic extremes will become more prominent with a warming climate (O’Gorman and Schneider 2009; Trenberth et al. 2009), with potential increases in flood frequency, droughts and low flow conditions, saturation events, landslide occurrence, and erosion. Ecosystems are expected to differ in their response to changes in precipitation intensity and inter-storm length because of differences in geomorphic conditions, climate, species assemblages, and susceptibility to drought. For erosion, these differences may be predictable with a general mass balance framework, but other processes are poorly understood, such as the effect of drought on tree mortality, vegetation resistance to insects and pathogens, and subsequent feedbacks to erosion processes. The indirect effects of disturbances (e.g., fire, insect infestations, pathogens) to shifts in water balance will complicate the response of erosion. Changing species composition will also potentially affect forest ecosystem water balance (see Chap. 3).
5.1 Erosion and Landslides
Changes in precipitation intensity, and in the magnitude and frequency of precipitation events that saturate soil and cause runoff, will interact with mass wasting and erosion. Potential annual increases and decreases in precipitation will directly contribute to the amount of water available to drive mass wasting at seasonal and event scales. Increases in extremes of precipitation intensity (Easterling et al. 2000; Karl and Knight 1998), rain-on-snow during mid-winter melt (Hamlet and Lettenmaier 2007; Wenger et al. 2011), and transport of moisture in atmospheric rivers (Ralph et al. 2006; Dettinger 2011) can increase pore water pressure on hillslopes, thus increasing the risk of landslides, erosion, and gully formation for individual storms. Seasonal to annual changes in precipitation will contribute to soil moisture and groundwater levels, which can amplify or mitigate individual events.
Direct effects of some climatic changes on sediment yield and mass wasting may be overshadowed by longer term, indirect effects through vegetation response (Istanbulluoglu and Bras 2006; Collins and Bras 2008; Goode et al. 2011). Although decreasing precipitation in some places might suggest reduced risks of erosion or landslides, this change may have indirect effects on mortality and thinning of vegetation and fire risk, which could in turn increase erosion and landslides through lower root reinforcement of soil and higher exposure of soil to precipitation. For example, paleoclimatic and paleoecological evidence links periods of drought and severe fire to severe erosion events (Briffa 2000; Meyer and Pierce 2003; Whitlock et al. 2003; Pierce et al. 2004; Marlon et al. 2006). At shorter time scales, years of widespread fire are linked to severely dry and warm years (e.g., McKenzie et al. 2004; Morgan et al. 2008; Littell et al. 2009). As we shift toward a drier and warmer climate in the western United States, more areas are likely to burn annually (e.g., Littell et al. 2009; Spracklen et al. 2009), with resulting postfire debris flows (Meyer and Pierce 2003; Luce 2005; Shakesby and Doerr 2006; Moody and Martin 2009; Cannon et al. 2010). Breshears et al. (2005) documented drought-induced canopy mortality of ponderosa pine, followed by erosional loss of topsoil and nutrients, with subsequent species replacement by pinyon pine and juniper. These types of state transitions may indicate the type of complex feedbacks that will lead to permanent shifts in dominant vegetation, rather than to recovery following disturbance.
Adjustment of canopy density and root distributions to longer inter-storm periods may increase the efficiency of use of rain or snowmelt (Hwang et al. 2009; Brooks et al. 2011). The response of both annual runoff and runoff from extreme events may be amplified or mitigated by forest canopy adjustment to temperature, moisture, N, and atmospheric CO2. Increased precipitation intensity and amount, combined with lower root biomass from a drier climate, can yield more unstable slopes. Shifts in species dominance can also cause changes in root depth and cohesion (Hales et al. 2009). The spatial pattern of unstable slope conditions that can lead to landslides is influenced by interactions among the lateral redistribution of soil water in large events, the resulting pattern of high pore pressures with topographic slope, and root cohesive strength (Band et al. 2011).
5.2 Drought and Water Supply
Projections of drought extent over the next 75 years show that the proportion of global land mass experiencing drought will double from 15 to 30 % (Burke et al. 2006), and on most land masses, dry season precipitation is expected to decline by 15 % (Solomon et al. 2009). Projections for the largest declines in the United States are in the Southwest, strongly affecting water supply (Barnett and Pierce 2008; Rajagopalan et al. 2009). As noted above, lower precipitation will probably increase both forest mortality (Allen et al. 2010; Holden et al. 2011a) and fire risk (Westerling et al. 2011); however, forest mortality may not substantially mitigate runoff reductions associated with decreased precipitation (Adams et al. 2012). Historical observations of interannual variability in precipitation in the western United States have shown substantial increases in variability in the last 50 years (Luce and Holden 2009; Pagano and Garen 2005), even in areas not projected to show precipitation declines. Short-term severe droughts have consequences for vegetation (Holden et al. 2011b; van Mantgem et al. 2009) and water supply.
Although there has been interest in using forest harvest to augment water supplies, most increases in water yield after harvest occur in wet years (Brown et al. 2005; Ford et al. 2011), which may be less frequent in the future. In addition, water yield increases in snow environments occur earlier in the year, exacerbating flow timing issues caused by climate change (Troendle et al. 2010). Finally, in warmer and moister locations, increases in water yields can be replaced by decreases as young vegetation reestablishes within a few years (Brown et al. 2005; Ford et al. 2011).
6 Disturbance Interactions
6.1 Disturbances and Thresholds
Understanding interactions among disturbance regimes is a significant challenge for projecting the effects of climate change on forest ecosystems (Bigler et al. 2005; Busby et al. 2008) (Box 4.5). For example, how will massive outbreaks of bark beetles, which kill trees by feeding on cambial tissues, increase the potential for large severe wildfires in a warming climate (Box 4.1)? Interactions between processes can amplify or mute the overall effects of changes in complex forest ecosystems. The predominance of negative and positive feedbacks within and between processes will determine the stability or instability of the system.
Box 4.5: Response of Western Mountain Ecosystems to Climatic Variability and Change: The Western Mountain Initiative
The Western Mountain Initiative (WMI) uses paleoecological studies, contemporary studies, and modeling to understand responses to climatic variability and change in mountainous landscapes in the 11 large conterminous Western states (http://westernmountains.org). Initiated in 1991, the WMI consists of ten scientific laboratories in two federal agencies and four universities.
Research has documented how climatic variability and change affect long-term patterns of snow, glaciers, and water geochemistry; forest productivity, vigor, and demography; and changing patterns of treeline dynamics and forest disturbances. Empirical and simulation modeling indicates that major changes in hydrologic function and ecological disturbance will occur in a warming climate. WMI data show that extreme disturbances have rapidly altered the structure and function of forest ecosystems over the past decade (Peterson et al. 2012).
WMI research on disturbance interactions and their effects on ecosystem processes indicates that synergistic interactions between disturbances produce larger effects than would occur from an individual disturbance, especially when combined with chronic stressors such as air pollution, periodic drought, and reduced snowpack. For example, bark beetle outbreaks have been linked to increased likelihood of stand-replacing fire and changes in fire behavior, with the nature of the effect depending on the time since outbreak. Combined with increasing climatic stress on tree populations and growth, disturbance interactions can alter forest structure and function faster than could be expected from species redistribution or disturbance alone. Simultaneous climatically driven shifts in the locations of species optima, ecosystem productivity, disturbance regimes, and interactions between them can reset forest succession over large areas and short timeframes. (Figure is a conceptual model of relative time scales for disturbance versus climate change alone to alter ecosystems. The focus is on fire, but the same logic applies to insect outbreaks. Adapted from McKenzie et al. (2004))

Disturbance interactions may rapidly bring ecosystems to thresholds (Groffman et al. 2006). For example, Allen and Breshears (1998) and Breshears et al. (2005) documented rapid dieback of pinyon pine across the arid Southwest. Mature trees were pushed over a threshold by a combination of “global-change type drought” (Breshears et al. 2005) and an opportunistic bark beetle invasion. Regeneration of pinyon pine will determine whether this mortality represents a threshold for the ecosystem. Characteristic patterns of patchiness or continuity may indicate thresholds that have been approached or crossed (Scheffer et al. 2009). For example, the invasion of sagebrush (Artemisia tridentata Nutt.) steppe by cheatgrass (Fischer et al. 1996) and of the Sonoran Desert by buffelgrass (Cenchrus ciliaris L.) (Esque et al. 2007) provide fuel continuity and the potential for much more extensive wildfires than non-invaded areas with patchy fuels.
A notable threshold response to multiple stressors is the reproductive cycle of mountain pine beetle (Logan and Powell 2001) (see Sect. 4.3), whose outbreaks have killed mature trees across millions of hectares of pine in western North America. Within particular ranges of winter temperatures and growing-season degree days, the reproductive cycle is synchronized to the seasonal cycle, permitting maximum survival and epidemic population size. This “adaptive seasonality,” combined with drought-caused and age-related vulnerability of the host species, may promote an abrupt increase in mortality of lodgepole pine (Hicke et al. 2006).
Conceptually, thresholds are fairly well understood. Modeling of thresholds has by necessity taken place in simplified (often virtual) ecosystems, and a major challenge remains to apply such sophistication to real-world systems outside the specific examples chosen by modelers to test their hypotheses. A larger challenge will always be the unpredictability of the occurrence of contingent, interacting events that push systems across thresholds.
6.2 Stress Complexes: From Conceptual to Quantitative Models
In the context of the effects of climate change on ecosystems, sensitivity to disturbance interactions is extended to environmental drivers not usually identified as disturbances. For example, extreme temperatures, drought, and air pollution put forest ecosystems under stress, which may increase their vulnerability to “true” disturbances such as fire, insect outbreaks, and pathogens. Following McKenzie et al. (2009), we refer to interacting stresses as stress complexes and present three examples from the Sierra Nevada, Alaska, and the Southeast.
A striking feature of mixed conifer forests in the southern Sierra Nevada and southern California is ambient air pollution, particularly elevated ozone, which affects plant vigor by reducing net photosynthesis and therefore growth (Peterson et al. 1991) and is often concentrated at middle and upper elevations (Brace and Peterson 1998). Air pollution exacerbates drought stress from warmer temperatures, which amplifies biotic stresses such as insects and pathogens (Ferrell 1996). The stress complex for California forests is represented in Fig. 4.2; interacting disturbances form the core of drivers of ecosystem change, modified by climate, management, and air pollution.

Conceptual model of stress complexes in mixed conifer forests of the southern Sierra Nevada and southern California. The effects of insects and fire disturbance regimes (red box) and of fire exclusion are exacerbated by higher temperature. Stand-replacing fires and drought-induced mortality both contribute to species changes and invasive species (Modified from McKenzie et al. 2009)
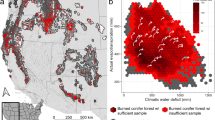
Alaska has experienced massive fires in the last decade, including the five largest fires in the United States. Over 2.5 million ha burned in the interior in 2004. Concurrently (1990s), massive outbreaks of the spruce bark beetle occurred on and near the Kenai Peninsula in south-central Alaska (Berg et al. 2006) (Fig. 4.3). Although periodic outbreaks have occurred throughout the historical record, both in south-central Alaska and the southwestern Yukon, these most recent outbreaks may be unprecedented in both extent and percentage mortality (over 90 % in many places) (Berg et al. 2006). Both wildfire and bark beetle outbreaks are associated with warmer temperatures in recent decades (Duffy et al. 2005; Werner et al. 2006). At the same time, major hydrological changes are underway from the cumulative effects of warming. Permafrost degradation is widespread in central Alaska, shifting ecosystems from birch forests to wetland types such as bogs and fens (Jorgenson et al. 2001). If broad-scale water balances become increasingly negative, peatlands may begin to support upland forest species (Klein et al. 2005). The stress complex for Alaska is represented conceptually in Fig. 4.4; upland and lowland ecosystems may follow parallel but contrasting paths toward new structure and species composition.

Mortality of white spruce from bark beetle attack on the Kenai Peninsula, Alaska (Photo by W.M. Ciesla, Forest Health Management International, Bugwood.org)

Conceptual model of stress complexes in the interior and coastal forests of Alaska. Rapid increases in the severity of disturbance regimes (insects and fire) are triggered by a warmer climate. Stand-replacing fires, massive mortality from insects, and permafrost degradation contribute to species changes and conversion to deciduous life forms (Modified from McKenzie et al. 2009)
Much of the forested landscape in the southeastern United States is adapted to frequent fire, and prescribed fire is a mainstay of ecosystem management. Fire-adapted inland forests overlap geographically with coastal areas affected by hurricanes and potentially by sea-level rise (Ross et al. 2009), such that interactions between wildfires and hurricanes are synergistic (Fig. 4.5). For example, dry-season (prescribed) fires may have actually been more severe than wet-season (lightning) fires in some areas, causing structural damage via cambium kill and subsequent increased vulnerability to hurricane damage (Platt et al. 2002). The stress complex for the Southeast is represented conceptually in Fig. 4.6.

Interactions between wildfire and hurricanes are synergistic in the southern United States. Figure depicts a longleaf pine/saw palmetto flatwoods stand on the Atlantic coastal plain, 2.5 years after a hurricane and with a previous history of prescribed fire (Courtesy of the Fire and Environmental Research Applications team, U.S. Forest Service, Digital Photo Series)

Conceptual model of stress complexes in the interior and coastal forests of the Southeast. Increases in the severity of hurricanes are triggered by global warming as sea level rises. Warmer and drier climate in uplands leads to longer periods with flammable fuels. Changes in fire and hydrologic regimes, and responses to them, lead to species change and altered C dynamics
6.3 Uncertainties
Current knowledge about multiple stressors is mainly qualitative, despite case studies in various ecosystems that have measured the effects of interactions and even followed them over time (Hicke et al. 2012b). In the three examples above, the directional effects of warming-induced stressors may be clear (e.g., in California, species composition shifts to those associated with frequent fire). However, the magnitudes of these effects are not, nor are the potentially irreversible crossings of ecological thresholds. Given the complexity and diversity of potential interacting stressors in U.S. forests, a fruitful way to advance quantitative knowledge may be with explicit simulations with models of “intermediate complexity” to ascertain the sensitivity of ecosystems to uncertainties associated with key parameters (e.g., the thickness of the arrows in Figs. 4.2, 4.4 and 4.6). As the climate continues to warm, new empirical data will incrementally help to quantify disturbance and stressor interactions, providing greater certainty about the nature of stress complexes in forest ecosystems.
References
Adams, H. D., Macalady, A. K., Breshears, D. D., et al. (2010). Climate-induced tree mortality: Earth system consequences. EOS, Transactions of the American Geophysical Union, 91, 153.
Adams, H. D., Luce, C. H., Breshears, D. D., et al. (2012). Ecohydrological consequences of drought- and infestation-triggered tree die-off: Insights and hypotheses. Ecohydrology, 5, 145–149.
Allen, C. D., & Breshears, D. D. (1998). Drought-induced shift of a forest-woodland ecotone: Rapid landscape response to climate variation. Proceedings of the National Academy of Sciences, USA, 95, 14839–14842.
Allen, C. D., Savage, M., Falk, D. A., et al. (2002). Ecological restoration of Southwestern ponderosa pine ecosystems: A broad perspective. Ecological Applications, 12, 1418–1433.
Allen, C. D., Macalady, A. K., Chenchouni, H., et al. (2010). A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. Forest Ecology and Management, 259, 660–684.
Alpert, P., Bone, E., & Holzapfel, C. (2000). Invasiveness, invasibility and the role of environmental stress in the spread of non-native plants. Perspectives in Plant Ecology, Evolution and Systematics, 3, 52–66.
Ayres, M. P., & Lombardero, M. J. (2000). Assessing the consequences of global change for forest disturbance from herbivores and pathogens. The Science of the Total Environment, 262, 263–286.
Bachelet, D., Neilson, R. P., Lenihan, J. M., & Drapek, R. J. (2001). Climate change effects on vegetation distribution and carbon budget in the United States. Ecosystems, 4, 164–185.
Bailey, J. D., Mayrohn, C., Doescher, P. S., et al. (1998). Understory vegetation in old and young Douglas-fir forests of western Oregon. Forest Ecology and Management, 112, 289–302.
Band, L. E., Hwang, T., Hales, T. C., et al. (2011). Ecosystem processes at the watershed scale: Mapping and modeling ecohydrological controls of landslides. Geomorphology, 137, 159–167.
Barnett, T. P., & Pierce, D. W. (2008). When will Lake Mead go dry? Water Resources Research, 44, W03201.
Bentz, B., Logan, J., MacMahon, J., et al. (2009). Bark beetle outbreaks in western North America: Causes and consequences (44pp). Salt Lake City: University of Utah Press.
Bentz, B. J., Régnière, J., Fettig, C. J., et al. (2010). Climate change and bark beetles of the Western United States and Canada: Direct and indirect effects. BioScience, 60, 602–613.
Berg, E. E., Henry, J. D., Fastie, C. L., et al. (2006). Spruce beetle outbreaks on the Kenai Peninsula, Alaska, and Kluane National Park and Reserve. Yukon Territory: Relationship to summer temperatures and regional differences in disturbance regimes. Forest Ecology and Management, 227, 219–232.
Berggren, A., Böjrkman, C., Bylund, H., & Ayres, M. P. (2009). The distribution and abundance of animal populations in a climate of uncertainty. Oikos, 118, 1121–1126.
Bigler, C., Kulakowski, D., & Veblen, T. T. (2005). Multiple disturbance interactions and drought influence fire severity in Rocky Mountain subalpine forests. Ecology, 86, 3018–3029.
Bond, W. J., & Midgley, G. F. (2000). A proposed CO2-controlled mechanism of woody plant invasion in grasslands and savannas. Global Change Biology, 6, 865–869.
Boon, S. (2012). Snow accumulation following forest disturbance. Ecohydrology, 5, 279–285.
Brace, S., & Peterson, D. L. (1998). Spatial patterns of tropospheric ozone in the Mount Rainier region of the Cascade Mountains, U.S.A. Atmospheric Environment, 32, 3629–3637.
Bradley, B. A., Oppenheimer, M., & Wilcove, D. S. (2009). Climate change and plant invasions: Restoration opportunities ahead? Global Change Biology, 15, 1511–1521.
Breshears, D. D., Cobb, N. S., Rich, P. M., et al. (2005). Regional vegetation die-off in response to global-change-type drought. Proceedings of the National Academy of Sciences, USA, 102, 15144–15148.
Briffa, K. R. (2000). Annual climate variability in the Holocene: Interpreting the message of ancient trees. Quaternary Science Reviews, 19, 87–105.
Brooks, P. D., Troch, P. A., Durcik, M., et al. (2011). Quantifying regional scale ecosystem response to changes in precipitation: Not all rain is created equal. Water Resources Research, 47, W00J08.
Brown, A. E., Zhang, L., McMahon, T. A., et al. (2005). A review of paired catchment studies for determining changes in water yield resulting from alterations in vegetation. Journal of Hydrology, 310, 28–61.
Burke, E. J., Brown, S. J., & Christidis, N. (2006). Modeling the recent evolution of global drought and projections for the twenty-first century with the Hadley Centre climate model©. Journal of Hydrometeorology, 7, 1113–1125.
Busby, P. E., Motzkin, G., & Foster, D. R. (2008). Multiple and interacting disturbances lead to Fagus grandifolia dominance in coastal New England. Journal of the Torrey Botanical Society, 135, 346–359.
Campbell, R., Smith, D. J., & Arsenault, A. (2006). Multicentury history of western spruce budworm outbreaks in interior Douglas-fir forests near Kamloops, British Columbia. Canadian Journal of Forest Research, 36, 1758–1769.
Candau, J.-N., & Fleming, R. A. (2005). Landscape-scale spatial distribution of spruce budworm defoliation in relation to bioclimatic conditions. Canadian Journal of Forest Research, 35, 2218–2232.
Cannon, S. H., Gartner, J. E., Rupert, M. G., et al. (2010). Predicting the probability and volume of postwildfire debris flows in the intermountain western United States. Geological Society of America Bulletin, 122, 127–144.
Chambers, J. C., Roundy, B. A., Blank, R. R., et al. (2007). What makes Great Basin sagebrush ecosystems invasible by Bromus tectorum? Ecological Monographs, 77, 117–145.
Chan-McLeod, A. C. A. (2006). A review and synthesis of the effects of unsalvaged mountain-pine-beetle-attacked stands on wildlife and implications for forest management. BC Journal of Ecosystems and Management, 7, 119–132.
Cohn, J. P. (2005). Tiff over tamarisk: Can a nuisance be nice, too? Bioscience, 55, 648–654.
Colautti, R. I., Grigorovich, I. A., & MacIsaac, H. J. (2006). Propagule pressure: A null model for biological invasions. Biological Invasions, 8, 1023–1037.
Collins, D. B. G., & Bras, R. L. (2008). Climate control of sediment yield in dry lands following climate and land cover change. Water Resources Research, 44, W10405.
Collins, B. M., Omi, P. N., & Chapman, P. L. (2006). Regional relationships between climate and wildfire-burned area in the Interior West, USA. Canadian Journal of Forest Research, 36, 699–709.
Collins, B. J., Rhoades, C. C., Hubbard, R. M., & Battaglia, M. A. (2011). Tree regeneration and future stand development after bark beetle infestation and harvesting in Colorado lodgepole pine stands. Forest Ecology and Management, 261, 2168–2175.
Crimmins, S. M., Dobrowski, S. Z., Greenberg, J. A., et al. (2011). Changes in climate water balance drive downhill shifts in plant species optimum elevations. Science, 331, 324–327.
Cudmore, T. J., Bjorkland, N., Carroll, A. L., & Lindgren, B. S. (2010). Climate change and range expansion of an aggressive bark beetle: Evidence of higher beetle reproduction in naïve host tree populations. Journal of Applied Ecology, 47, 1036–1043.
D’Antonio, C. M. (2000). Chapter 4: Fire, plant invasions, and global changes. In H. A. Mooney & R. J. Hobbs (Eds.), Invasive species in a changing world (pp. 65–94). Washington, DC: Island Press.
D’Antonio, C., Levine, J., & Thomsen, M. (2001). Ecosystem resistance to invasion and the role of propagule supply: A California perspective. Journal of Mediterranean Ecology, 2, 233–245.
Dale, V. H., Joyce, L. A., McNulty, S., et al. (2001). Climate change and forest disturbances. BioScience, 51, 723–734.
Davis, M. A., Grime, J. P., & Thompson, K. (2000). Fluctuating resources in plant communities: A general theory of invasibility. Journal of Ecology, 88, 528–534.
Dettinger, M. (2011). Climate change, atmospheric rivers, and floods in California—A multimodel analysis of storm frequency and magnitude changes. Journal of the American Water Resources Association, 47, 514–523.
Drever, M. C., Goheen, J. R., & Martin, K. (2009). Species-energy theory, pulsed resources, and regulation of avian richness during a mountain pine beetle outbreak. Ecology, 90, 1095–1105.
Duffy, P. A., Walsh, J. E., Graham, J. M., et al. (2005). Impacts of large-scale atmospheric-ocean variability on Alaskan fire season severity. Ecological Applications, 15, 1317–1330.
Dukes, J. S., & Mooney, H. A. (1999). Does global change increase the success of biological invaders? Trends in Ecology & Evolution, 14, 135–139.
Dukes, J. S., Pontius, J., Orwig, D., et al. (2009). Responses of insect pests, pathogens, and invasive plant species to climate change in the forests of northeastern North America: What can we predict? Canadian Journal of Forest Research, 39, 231–248.
Dukes, J. S., Chiariello, N. R., Loarie, S. R., & Field, C. B. (2011). Strong response of an invasive plant species (Centaurea solstitialis L.) to global environmental changes. Ecological Applications, 21, 1887–1894.
Duncan, C. A., Jachetta, J. J., Brown, M. L., et al. (2004). Assessing the economic, environmental, and societal losses from invasive plants on rangeland and wildlands. Weed Technology, 18, 1411–1416.
Dwyer, G., Dushoff, J., & Yee, S. H. (2004). The combined effects of pathogens and predators on insect outbreaks. Nature, 430, 341–345.
Eamus, D. (1991). The interaction of rising CO2 and temperatures with water use efficiency. Plant, Cell & Environment, 14, 843–852.
Easterling, D. R., Evans, J. L., Groisman, P. Y., et al. (2000). Observed variability and trends in extreme climate events: A brief review. Bulletin of the American Meteorological Society, 81, 417–425.
Eschtruth, A. K., & Battles, J. J. (2009). Assessing the relative importance of disturbance, herbivory, diversity, and propagule pressure in exotic plant invasion. Ecological Monographs, 79, 265–280.
Esque, T. C., Schwalbe, C. R., Lissow, J. A., et al. (2007). Buffelgrass fuel loads in Saguaro National Park, Arizona, increase fire danger and threaten native species. Park Science, 24, 33–37.
Evangelista, P. H., Kumer, S., Stohlgren, T. J., & Young, N. E. (2011). Assessing forest vulnerability and the potential distribution of pine beetles under current and future climate scenarios in the Interior West of the US. Forest Ecology and Management, 262, 307–316.
Evans, L. M., Hofstetter, R. W., Ayres, M. P., & Klepzig, K. D. (2011). Temperature alters the relative abundance and population growth rates of species within the Dendroctonus frontalis (Coleoptera: Curculionidae) community. Environmental Entomology, 40, 824–834.
Fauria, M. M., & Johnson, E. A. (2006). Large-scale climatic patterns control large lightning fire occurrence in Canada and Alaska forest regions. Journal of Geophysical Research, 111, G04008.
Ferrell, G. T. (1996). Chapter 45: The influence of insect pests and pathogens on Sierra forests. In Sierra Nevada Ecosystem Project: Final report to Congress, Vol. II, Assessments and scientific basis for management options (pp. 1177–1192). Davis: University of California, Centers for Water and Wildland Resources.
Fischer, R. A., Reese, K. P., & Connelly, J. W. (1996). An investigation on fire effects within xeric sage grouse brood habitat. Journal of Range Management, 49, 194–198.
Ford, C. R., Laseter, S. H., Swank, W. T., & Vose, J. M. (2011). Can forest management be used to sustain water-based ecosystem services in the face of climate change? Ecological Applications, 21, 2049–2067.
Foster, D. R., Clayden, S., Orwig, D. A., et al. (2002). Oak, chestnut and fire: Climatic and cultural controls of long-term forest dynamics in New England, USA. Journal of Biogeography, 29, 1359–1379.
Friedenberg, N. A., Powell, J. A., & Ayres, M. P. (2007). Synchrony’s double edge: Transient dynamics and the Allee effect in stage structured populations. Ecology Letters, 10, 564–573.
Furniss, M. M., Holsten, E. H., Foote, M. J., & Bertram, M. (2001). Biology of a willow leafblotch miner, Micrurapteryx salicifoliella, (Lepidoptera: Gracillariidae) in Alaska. Environmental Entomology, 30, 736–741.
Ganey, J. L., & Vojta, S. C. (2011). Tree mortality in drought-stressed mixed-conifer and ponderosa pine forests, Arizona, USA. Forest Ecology and Management, 261, 162–168.
Garnas, J. R., Ayres, M. P., Liebhold, A. M., & Evans, C. (2011a). Subcontinental impacts of an invasive tree disease on forest structure and dynamics. Journal of Ecology, 99, 532–541.
Garnas, J. R., Houston, D. R., Ayres, M. P., & Evans, C. (2011b). Disease ontogeny overshadows effects of climate and species interactions on population dynamics in a nonnative forest disease complex. Ecography, 35, 412–421.
Gaylord, M. L., Kolb, T. E., Wallin, K. F., & Wagner, M. R. (2007). Seasonal dynamics of tree growth, physiology, and resin defenses in a northern Arizona ponderosa pine forest. Canadian Journal of Forest Research, 37, 1173–1183.
Gillooly, J. F., Charnov, E. L., West, G. B., et al. (2002). Effects of size and temperature on developmental time. Nature, 417, 70–73.
Goode, J. R., Luce, C. H., & Buffington, J. M. (2011). Enhanced sediment delivery in a changing climate in semi-arid mountain basins: Implications for water resource management and aquatic habitat in the northern Rocky Mountains. Geomorphology, 139/140, 1–15.
Greenwood, D. L., & Weisberg, P. J. (2008). Density-dependent tree mortality in pinyon-juniper woodlands. Forest Ecology and Management, 255, 2129–2137.
Grissino-Mayer, H. D., & Swetnam, T. W. (2000). Century-scale climate forcing of fire regimes in the American Southwest. The Holocene, 10, 213–220.
Groffman, P. M., Baron, J. S., Blett, T., et al. (2006). Ecological thresholds: The key to successful environmental management or an important concept with no practical application? Ecosystems, 9, 1–13.
Grulke, N. E. (2011). The nexus of host and pathogen phenology: Understanding the disease triangle with climate change. New Phytologist, 189, 8–11.
Hales, T. C., Ford, C. R., Hwang, T., et al. (2009). Topographic and ecologic controls on root reinforcement. Journal of Geophysical Research, 114, F03013.
Halpern, C. B. (1989). Early successional patterns of forest species: Interactions of life history traits and disturbance. Ecology, 70, 704–720.
Hamlet, A. F., & Lettenmaier, D. P. (2007). Effects of 20th century warming and climate variability on flood risk in the western U.S. Water Resources Research, 43, W06427.
Hellmann, J. J., Byers, J. E., Bierwagen, B. G., & Dukes, J. S. (2008). Five potential consequences of climate change for invasive species. Special section. Conservation Biology, 22, 534–543.
Hessl, A. E., McKenzie, D., & Schellhaas, R. (2004). Drought and Pacific Decadal Oscillation linked to fire occurrence in the inland Pacific Northwest. Ecological Applications, 14, 425–442.
Heyerdahl, E. K., Brubaker, L. B., & Agee, J. K. (2002). Annual and decadal climate forcing of historical regimes in the interior Pacific Northwest, USA. The Holocene, 12, 597–604.
Heyerdahl, E. K., McKenzie, D., Daniels, L. D., et al. (2008a). Climate drivers of regionally synchronous fires in the inland Northwest (1651–1900). International Journal of Wildland Fire, 17, 40–49.
Heyerdahl, E. K., Morgan, P., & Riser, J. P. (2008b). Multi-season climate synchronized historical fires in dry forests (1650–1900), Northern Rockies, U.S.A. Ecology, 89, 705–716.
Hicke, J. A., Logan, J. A., Powell, J., & Ojima, D. S. (2006). Changes in temperature influence suitability for modeled mountain pine beetle (Dendroctonus ponderosae) outbreaks in the Western United States. Journal of Geophysical Research, 11, G02019.
Hicke, J. A., Allen, C. D., Desai, A. R., et al. (2012a). Effects of biotic disturbances on forest carbon cycling in the United States and Canada. Global Change Biology, 18, 7–34.
Hicke, J. A., Johnson, M. C., Hayes, J. L., & Preisler, H. K. (2012b). Effects of bark beetle-caused tree mortality on wildfire. Forest Ecology and Management, 271, 81–90.
Hofstetter, R. W., Dempsey, T. D., Klepzig, K. D., & Ayres, M. P. (2007). Temperature-dependent effects on mutualistic, antagonistic, and commensalistic interactions among insects, fungi and mites. Community Ecology, 8, 47–56.
Holden, Z. A., Abatzoglou, J. T., Luce, C. H., & Baggett, L. S. (2011a). Empirical downscaling of daily minimum air temperature at very fine resolutions in complex terrain. Agricultural and Forest Meteorology, 151, 1066–1073.
Holden, Z. A., Luce, C. H., Crimmins, M. A., & Morgan, P. (2011b). Wildfire extent and severity correlated with annual streamflow distribution and timing in the Pacific Northwest, USA (1984–2005). Ecohydrology. doi:10.1002/eco.257.
Holmes, T. P., Liebhold, A. M., Kovacs, K. F., & Von Holle, B. (2010). A spatial-dynamic value transfer model of economic losses from a biological invasion. Ecological Economics, 70, 86–95.
Hutchinson, T. F., Long, R. P., Ford, R. D., & Sutherland, E. K. (2008). Fire history and the establishment of oaks and maples in second-growth forests. Canadian Journal of Forest Research, 38, 1184–1198.
Hwang, T., Band, L., & Hales, T. C. (2009). Ecosystem processes at the watershed scale: Extending optimality theory from plot to catchment. Water Resources Research, 45, W11425.
Istanbulluoglu, E., & Bras, R. L. (2006). On the dynamics of soil moisture, vegetation, and erosion: Implications of climate variability and change. Water Resources Research, 42, W06418.
Jenkins, M. J., Hebertson, E., Page, W., & Jorgensen, C. A. (2008). Bark beetles, fuels, fires and implications for forest management in the Intermountain West. Forest Ecology and Management, 254, 16–34.
Jepsen, J. U., Kapari, L., & Hagen, S. B. (2011). Rapid northwards expansion of a forest insect pest attributed to spring phenology matching with sub-Arctic birch. Global Change Biology, 17, 2071–2083.
Jorgenson, M. T., Racine, C. H., Walters, J. C., & Osterkamp, T. E. (2001). Permafrost degradation and ecological changes associated with a warming climate in central Alaska. Climatic Change, 48, 551–571.
Karl, T. R., & Knight, R. W. (1998). Secular trends of precipitation amount, frequency, and intensity in the United States. Bulletin of the American Meteorological Society, 79, 231–241.
Keeley, J. E., & McGinnis, T. W. (2007). Impact of prescribed fire and other factors on cheatgrass persistence in a Sierra Nevada ponderosa pine forest. International Journal of Wildland Fire, 16, 96–106.
Keeley, J. E., Lubin, D., & Fotheringham, C. J. (2003). Fire and grazing impacts on plant diversity and alien plant invasions in the southern Sierra Nevada. Ecological Applications, 13, 1355–1374.
Kemp, W. P., Everson, D. O., & Wellington, W. G. (1985). Regional climatic patterns and western spruce budworm outbreaks (Tech. Bull. 1693, 31pp). Washington, DC: U.S. Department of Agriculture, Forest Service, Canada/United States Spruce Budworms Program.
Kerns, B. K., Thies, W. G., & Niwa, C. G. (2006). Season and severity of prescribed burn in ponderosa pine forests: Implications for understory native and exotic plants. Ecoscience, 13, 44–55.
Kerns, B. K., Naylor, B. J., Buonopane, M., et al. (2009). Modeling tamarisk (Tamarix spp.) habitat and climate change effects in the Northwestern United States. Invasive Plant Science and Management, 2, 200–215.
Kitzberger, T., Brown, P. M., Heyerdahl, E. K., et al. (2007). Contingent Pacific-Atlantic Ocean influence on multicentury wildfire synchrony over western North America. Proceedings of the National Academy of Sciences, USA, 104, 543–548.
Klapwijk, M. J., Ayres, M. P., Battisti, A., & Larsson, S. (2012). Assessing the impact of climate change on outbreak potential. In P. Barbosa, D. L. Letourneau, & A. A. Agrawal (Eds.), Insect outbreaks revisited (pp. 429–450). New York: Wiley-Blackwell.
Klein, E., Berg, E. E., & Dial, R. (2005). Wetland drying and succession across the Kenai Peninsula Lowlands, south-central Alaska. Canadian Journal of Forest Research, 35, 1931–1941.
Klopfenstein, N. B., Kim, M.-S., Hanna, J. W., et al. (2009). Approaches to predicting potential impacts of climate change on forest disease: An example with Amillaria root disease (Res. Pap. RMRS-RP-76, 10pp). Fort Collins: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station.
Kovacs, K. F., Haight, R. G., McCollough, D. G., et al. (2010). Cost of potential emerald ash borer damage in U.S. communities, 2009–2019. Ecological Economics, 69, 569–578.
Kovacs, K. F., Mercader, R. J., Haight, R. G., et al. (2011). The influence of satellite populations of emerald ash borer on projected economic costs in U.S. communities, 2010–2020. Journal of Environmental Management, 92, 2170–2181.
Levine, J. M. (2000). Species diversity and biological invasions: Relating local process to community pattern. Science, 288, 852–854.
Levine, J. M., Adler, P. B., & Yelenik, S. G. (2004). A meta-analysis of biotic resistance to exotic plant invasions. Review. Ecology Letters, 7, 975–989.
Lindroth, R. L. (2010). Impacts of elevated atmospheric CO2 and O3 on forests: Phytochemistry, trophic interactions, and ecosystem dynamics. Journal of Chemical Ecology, 36, 2–21.
Littell, J. S., McKenzie, D., Peterson, D. L., & Westerling, A. L. (2009). Climate and wildfire area burned in western U.S. ecoprovinces, 1916–2003. Ecological Applications, 19, 1003–1021.
Littell, J. S., Oneil, E. E., McKenzie, D., et al. (2010). Forest ecosystems, disturbance, and climatic change in Washington State, USA. Climatic Change, 102, 129–158.
Liu, Y., Stanturf, J., & Goodrick, S. (2010). Trends in global wildfire potential in a changing climate. Forest Ecology and Management, 259, 685–697.
Lodge, D. M., Williams, L., MacIsaac, H. J., et al. (2006). Biological invasions: Recommendations for U.S. policy and management. Ecological Applications, 16, 2035–2054.
Logan, J. A., & Powell, J. A. (2001). Ghost forests, global warming, and the mountain pine beetle (Coleoptera: Scolytidae). American Entomologist, 47, 160–173.
Lombardero, M. J., & Ayres, M. P. (2011). Factors influencing bark beetle outbreaks after forest fires on the Iberian Peninsula. Environmental Entomology, 40, 1007–1018.
Lombardero, M. J., Ayres, M. P., Ayres, B. D., & Reeve, J. D. (2000). Cold tolerance of four species of bark beetle (Coleoptera: Scolytidae) in North America. Environmental Entomology, 29, 421–432.
Lonsdale, W. M. (1999). Global patterns of plant invasions and the concept of invasibility. Ecology, 80, 1522–1536.
Lovett, G. M., Canham, C. D., Arthur, M. A., et al. (2006). Forest ecosystem responses to exotic pests and pathogens in eastern North America. BioScience, 56, 395–405.
Luce, C. H. (2005). Land use and land cover effects on runoff processes: Fire. In M. G. Anderson (Ed.), Encyclopedia of hydrological sciences (pp. 1831–1838). Hoboken: Wiley.
Luce, C. H., & Holden, Z. A. (2009). Declining annual streamflow distributions in the Pacific Northwest United States, 1948–2006. Geophysical Research Letters, 36, L16401.
Marlon, J., Bartlein, P. J., & Whitlock, C. (2006). Fire-fuel-climate linkages in the northwestern USA during the Holocene. The Holocene, 16, 1059–1071.
Marlon, J. R., Bartlein, P. J., Carcaillet, C., et al. (2008). Climate and human influences on global biomass burning over the past two millennia. Nature Geoscience, 1, 697–702.
McKenzie, D., Gedalof, Z., Peterson, D. L., & Mote, P. (2004). Climatic change, wildfire and conservation. Conservation Biology, 18, 890–902.
McKenzie, D., Peterson, D. L., & Littell, J. J. (2009). Global warming and stress complexes in forests of western North America. In A. Bytnerowicz, M. J. Arbaugh, A. R. Riebau, & C. Andersen (Eds.), Wildland fires and air pollution (pp. 319–337). Amsterdam/London: Elsevier. Developments in Environmental Science 8. Chapter 15.
McNulty, S. G., Lorio, P. L., Ayres, M. P., & Reeve, J. D. (1997). Predictions of southern pine beetle populations under historic and projected climate using a forest ecosystem model. In R. A. Mickler & S. Fox (Eds.), The productivity and sustainability of southern forest ecosystems in a changing environment (pp. 617–634). New York: Springer.
Melillo, J. M., Janetos, A. C., & Karl, T. R. (2001). Climate change impacts on the United States: The potential consequences of climate variability and change. Cambridge: Cambridge University Press.
Meyer, G. A., & Pierce, J. L. (2003). Climatic controls on fire-induced sediment pulses in Yellowstone National Park and central Idaho: a long-term perspective. Forest Ecology and Management, 178, 89–104.
Miller, J. H. (1990). Ailanthus altissima (Mill.) Swingle ailanthus. In R. M. Burns, & B. H. Honkala (Tech. Coords.), Silvics of North America: Vol. 2. Hardwoods. Agriculture handbook 654 (pp. 101–104). Washington, DC: U.S. Department of Agriculture, Forest Service.
Miller, R. F., & Wigand, P. E. (1994). Holocene changes in semiarid pinyon-juniper woodlands. Bioscience, 44, 465–474.
Miller, R. F., Bates, J. D., Svejcar, T. J., et al. (2005). Biology, ecology, and management of western juniper (Juniperus occidentalis) (Tech. Bull. 152, 82pp). Corvallis: Oregon State University, Agricultural Experiment Station.
Mitchell, R. J., Hiers, J. K., O’Brien, J. J., et al. (2006). Silviculture that sustains: The nexus between silviculture, frequent prescribed fire, and conservation of biodiversity in longleaf pine forests of the southeastern United States. Canadian Journal of Forest Research, 36, 2724–2736.
Moody, J. A., & Martin, D. A. (2009). Synthesis of sediment yields after wildland fire in different rainfall regimes in the Western United States. International Journal of Wildland Fire, 18, 96–115.
Morgan, P., Heyerdahl, E. K., & Gibson, C. E. (2008). Multi-season climate synchronized widespread forest fires throughout the 20th century, Northern Rockies, USA. Ecology, 89, 717–728.
Moser, J. C., Fitzgibbon, B. A., & Klepzig, K. D. (2005). The Mexican pine beetle, Dendroctonus mexicanus: First record in the United States and co-occurrence with the southern pine beetle— Dendroctonus frontalis (Coleoptera: Scolytidae or Curculionidae: Scolytidae). Entomological News, 116, 235–243.
Myers, J. H., Simberloff, D., Kuris, A. M., & Carey, J. R. (2000). Eradication revisited: Dealing with exotic species. Trends in Ecology & Evolution, 15, 316–320.
National Interagency Coordination Center. (2011). GACC predictive services intelligence. http://www.predictiveservices.nifc.gov/intelligence/intelligence.htm. Accessed 25 Jan 2012.
National Invasive Species Council. (2001). Meeting the invasive species challenge: National invasive species management plan (p. 80). Washington, DC: U.S. Department of Agriculture.
Nelson, C. R., Halpern, C. B., & Agee, J. K. (2008). Thinning and burning results in low-level invasion by nonnative plants but neutral effects on natives. Ecological Applications, 18, 762–770.
Nowacki, G. J., & Abrams, M. D. (2008). The demise of fire and “mesophication” of forests in the eastern United States. Bioscience, 58, 112–128.
O’Gorman, P. A., & Schneider, T. (2009). The physical basis for increases in precipitation extremes in simulations of 21st-century climate change. Proceedings of the National Academy of Sciences, USA, 106, 14773–14777.
Orwig, D. A., Foster, D. R., & Mausel, D. L. (2002). Landscape patterns of hemlock decline in New England due to the introduced hemlock woolly adelgid. Journal of Biogeography, 29, 1475–1487.
Pagano, T., & Garen, D. (2005). A recent increase in Western U.S. streamflow variability and persistence. Journal of Hydrometeorology, 6, 173–179.
Paradis, A., Elkinton, J., Hayhoe, K., & Buonaccorsi, J. (2008). Role of winter temperature and climate change on the survival and future range expansion of the hemlock woolly adelgid (Adelges tsugae) in eastern North America. Mitigation and Adaptation Strategies for Global Change, 13, 541–554.
Parker, T. J., Clancy, K. M., & Mathiasen, R. L. (2006). Interactions among fire, insects and pathogens in coniferous forests of the interior western United States and Canada. Agricultural and Forest Entomology, 8, 167–189.
Parks, C. G., Radosevich, S. R., Endress, B. A., et al. (2005). Natural and land-use history of the Northwest mountain ecoregions (USA) in relation to patterns of plant invasions. Perspectives in Plant Ecology, Evolution and Systematics, 7, 137–158.
Pattison, R. R., & Mack, R. N. (2008). Potential distribution of the invasive tree Triadica sebifera (Euphorbiaceae) in the United States: Evaluating CLIMEX predictions with field trials. Global Change Biology, 14, 813–826.
Pauchard, A., Kueffer, C., Dietz, H., et al. (2009). Ain’t no mountain high enough: Plant invasions reaching new elevations. Frontiers in Ecology and the Environment, 7, 479–486.
Peterson, D. L., Arbaugh, M. J., & Robinson, L. J. (1991). Growth trends of ozone-stressed ponderosa pine (Pinus ponderosa) in the Sierra Nevada of California, USA. The Holocene, 1, 50–61.
Peterson, D. L., Allen, C. D., Baron, J. S., et al. (2012). Response of Western mountain ecosystems to climatic variability and change: A collaborative research approach. In J. Bellant & E. Beever (Eds.), Ecological consequences of climate change: Mechanisms, conservation, and management (pp. 163–190). New York: Taylor & Francis.
Pfeifer, E. M., Hicke, J. A., & Meddens, A. J. H. (2011). Observations and modeling of aboveground tree carbon stocks and fluxes following a bark beetle outbreak in the Western United States. Global Change Biology, 17, 339–350.
Pierce, J. L., Meyer, G. A., & Jull, A. J. T. (2004). Fire-induced erosion and millennial-scale climate change in northern ponderosa pine forests. Nature, 432, 87–90.
Pimentel, D., Zuniga, R., & Morrison, D. (2005). Update on the environmental and economic costs associated with alien-invasive species in the United States. Ecological Economics, 52, 273–288.
Platt, W. J., Beckage, B., Doren, R. F., & Slater, H. H. (2002). Interactions of large-scale disturbances: Prior fire regimes and hurricane mortality of savanna pines. Ecology, 83, 1566–1572.
Powell, J. A., Jenkins, J. L., Logan, J. A., & Bentz, B. J. (2000). Seasonal temperature alone can synchronize life cycles. Bulletin of Mathematical Biology, 62, 977–998.
Powell, K. I., Chase, J. M., & Knight, T. M. (2011). Synthesis of plant invasion effects on biodiversity across spatial scales. American Journal of Botany, 98, 539–548.
Power, M. J., Marlon, J., Ortiz, N., et al. (2008). Changes in fire regimes since the Last Glacial Maximum: An assessment based on a global synthesis and analysis of charcoal data. Climate Dynamics, 30, 887–907.
Price, J. I., McCollum, D. W., & Berrens, R. P. (2010). Insect infestation and residential property values: A hedonic analysis of the mountain pine beetle epidemic. Forest Policy and Economics, 12, 415–422.
Raffa, K. F., Aukema, B. H., Bentz, B. J., et al. (2008). Cross-scale drivers of natural disturbances prone to anthropogenic amplification: The dynamics of bark beetle eruptions. BioScience, 58, 501–517.
Rajagopalan, B., Nowak, K., Prairie, J., et al. (2009). Water supply risk on the Colorado River: Can management mitigate? Water Resources Research, 45, W08201.
Ralph, F. M., Neiman, P. J., Wick, G. A., et al. (2006). Flooding on California’s Russian River: Role of atmospheric rivers. Geophysical Research Letters, 33, L13801.
Reynolds, L. V., Ayres, M. P., Siccama, T. G., & Holmes, R. T. (2007). Climatic effects on caterpillar fluctuations in northern hardwood forests. Canadian Journal of Forest Research, 37, 481–491.
Rodenhouse, N. L., Christenson, L. M., Parry, D., & Green, L. E. (2009). Climate change effects on native fauna of northeastern forests. Canadian Journal of Forest Research, 39, 249–263.
Rohrs-Richey, J. K., Mulder, C. P. H., Winton, L. M., & Stanosz, G. (2011). Physiological performance of an Alaskan shrub (Alnus fruticosa) in response to disease (Valsa melanodiscus) and water stress. New Phytologist, 189, 295–307.
Ross, M. S., Obrien, J. J., Ford, R. G., et al. (2009). Disturbance and the rising tide: The challenge of biodiversity management on low-island ecosystems. Frontiers in Ecology and the Environment, 7, 471–478.
Safranyik, L., Carroll, A. L., Régnière, D. W., et al. (2010). Potential for range expansion of mountain pine beetle into the boreal forest of North America. Canadian Entomologist, 142, 415–442.
Sage, R. F., Coiner, H. A., Way, D. A., et al. (2009). Kudzu [Pueraria montana (Lour.) Merr. var lobata]: A new source of carbohydrate for bioethanol production. Biomass and Bioenergy, 33, 57–61.
Salinas-Moreno, Y., Ager, A., Vargas, C. F., et al. (2010). Determining the vulnerability of Mexican pine forests to bark beetles of the genus Dendroctonus Erichson (Coleoptera: Curculionidae: Scolytinae). Forest Ecology and Management, 260, 52–61.
Sasek, T. W., & Strain, B. R. (1990). Implications of atmospheric CO2 enrichment and climatic change for the geographical distribution of two introduced vines in the U.S.A. Climatic Change, 16, 31–51.
Scheffer, M., Bascompte, J., Brock, W. A., et al. (2009). Early-warning signals for critical transitions. Nature, 461, 53–59.
Seabloom, E. W., Harpole, W. S., Reichman, O. J., & Tilman, D. (2003). Invasion, competitive dominance, and resource use by exotic and native California grassland species. Proceedings of the National Academy of Sciences, USA, 100, 13384–13389.
Seppälä, R., Buck, A., Katila, P. (Eds.). (2009). Adaptation of forests and people to climate change: A global assessment report (224pp). Helsinki: International Union of Forest Research Organizations.
Sexton, J. P., McKay, J. K., & Sala, A. (2002). Plasticity and genetic diversity may allow saltcedar to invade cold climates in North America. Ecological Applications, 12, 1652–1660.
Shakesby, R. A., & Doerr, S. H. (2006). Wildfire as a hydrological and geomorphological agent. Earth-Science Reviews, 74, 269–307.
Sheppard, S., & Picard, P. (2006). Visual-quality impacts of forest pest activity at the landscape level: A synthesis of published knowledge and research needs. Landscape and Urban Planning, 77, 321–342.
Sherriff, R. L., Berg, E. E., & Miller, A. E. (2011). Climate variability and spruce beetle (Dendroctonus rufipennis) outbreaks in south-central and southwest Alaska. Ecology, 92, 1459–1470.
Siegert, N. W., McCullough, D. G., Venette, R. C., et al. (2009). Assessing the climatic potential for epizootics of the gypsy moth fungal pathogen Entomophaga maimaiga in the north central United States. Canadian Journal of Forest Research, 39, 1958–1970.
Silander, J. A., Jr., & Klepeis, D. M. (1999). The invasion ecology of Japanese barberry (Berberis thunbergii) in the New England landscape. Biological Invasions, 1, 189–201.
Silveri, A., Dunwiddie, P. W., & Michaels, H. J. (2001). Logging and edaphic factors in the invasion of an Asian woody vine in a mesic North American forest. Biological Invasions, 3, 379–389.
Simard, M., Romme, W. H., Griffin, J. M., & Turner, M. G. (2011). Do mountain pine beetle outbreaks change the probability of active crown fire in lodgepole pine forests? Ecological Monographs, 81, 3–24.
Simberloff, D. (2009). The role of propagule pressure in biological invasions. Annual Review of Ecology, Evolution, and Systematics, 40, 81–102.
Six, D. L., & Bentz, B. J. (2007). Temperature determines symbiont abundance in a multipartite bark beetle-fungus ectosymbiosis. Microbial Ecology, 54, 112–118.
Solomon, S., Plattner, G.-K., Knutti, R., & Friedlingstein, P. (2009). Irreversible climate change due to carbon dioxide emissions. Proceedings of the National Academy of Sciences, USA, 106, 1704–1709.
Spracklen, D. V., Mickley, L. J., Logan, J. A., et al. (2009). Impacts of climate change from 2000 to 2050 on wildfire activity and carbonaceous aerosol concentrations in the Western United States. Journal of Geophysical Research, 114, D20301.
Standards and Trade Development Facility. (2009). Climate change and agriculture trade: Risks and responses. http://www.standardsfacility.org/Climate_change.htm. Accessed 24 Jan 2012.
Stein, B. A., & Flack, S. R. (Eds.). (1996). America’s least wanted: Alien species invasions of U.S. ecosystems (31pp). Arlington: The Nature Conservancy.
Stone, J. K., Coop, L. B., & Manter, D. K. (2008). Predicting effects of climate change on Swiss needle cast disease severity in Pacific Northwest forests. Canadian Journal of Plant Pathology, 30, 169–176.
Sturrock, R. N., Frankel, S. J., Brown, A. V., et al. (2011). Climate change and forest diseases. Plant Pathology, 60, 133–149.
Swetnam, T. W., & Betancourt, J. L. (1998). Mesoscale disturbance and ecological response to decadal climatic variability in the American Southwest. Journal of Climate, 11, 3128–3147.
Tchakerian, M. D., & Couslon, R. N. (2011). Ecological impacts of southern pine beetle. In R. N. Coulson, & K. D. Klepzig (Eds.), Southern pine beetle II (Gen. Tech. Rep. SRS-140, pp. 223–234). Asheville: U.S. Department of Agriculture, Forest Service, Southern Research Station.
Thomson, A. J., & Benton, R. (2007). A 90-year sea warming trend explains outbreak patterns of western spruce budworm on Vancouver Island. The Forestry Chronicle, 83, 867–869.
Thomson, A. J., Shepherd, R. F., Harris, J. W. E., & Silversides, R. H. (1984). Relating weather to outbreaks of western spruce budworm, Choristoneura occidentalis (Lepidoptera: Tortricidae), in British Columbia. The Canadian Entomologist, 116, 375–381.
Tilman, D. (1997). Community invasibility, recruitment limitation, and grassland biodiversity. Ecology, 78, 81–92.
Tkacz, B., Brown, H., Daniels, A., et al. (2010). National roadmap for responding to climate change (FS-957b). Washington, DC: U.S. Department of Agriculture, Forest Service.
Trân, J. K., Ylioja, T., Billings, R. F., et al. (2007). Impact of minimum winter temperatures on the population dynamics of Dendroctonus frontalis. Ecological Applications, 17, 882–899.
Trenberth, K. E., Fasullo, J. T., & Kiehl, J. (2009). Earth’s global energy budget. Bulletin of the American Meteorological Society, 90, 311–323.
Troendle, C. A., MacDonald, L. H., Luce, C. H., & Larsen, I. J. (2010). Fuel management and water yield. In W. J. Elliot, I. S Miller, & L. Audin (Eds.), Cumulative watershed effects of fuel management in the Western United States (Gen. Tech. Rep. RMRS-GTR-231, pp. 124–148). Fort Collins: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station. Chapter 7.
U.S. Department of Agriculture, Forest Service (USDA FS). (2010). Major forest insect and disease conditions in the United States: 2009 update (FS-952, 28pp). Washington, DC: U.S. Forest Service.
van Mantgem, P. J., Stephenson, N. L., Byrne, J. C., et al. (2009). Widespread increase of tree mortality rates in the Western United States. Science, 323, 521–524.
Veblen, T. T., Hadley, K. S., Reid, M. S., & Rebertus, A. J. (1991). The response of subalpine forests to spruce beetle outbreak in Colorado. Ecology, 72, 213–231.
Wagner, D. L., Defoliart, L., Doak, P., & Schneiderheinze, J. (2008). Impact of epidermal leaf mining by the aspen leaf miner (Phyllocnistis populiella) on the growth, physiology, and leaf longevity of quaking aspen. Oecologia, 157, 259–267.
Wenger, S. J., Isaak, D. J., Luce, C. H., et al. (2011). Flow regime, temperature, and biotic interactions drive differential declines of trout species under climate change. Proceedings of the National Academy of Sciences, USA, 108, 14175–14180.
Werner, R. A., Holsten, E. H., Matsouka, S. M., & Burnside, R. E. (2006). Spruce beetles and forest ecosystems in south-central Alaska: A review of 30 years of research. Forest Ecology and Management, 227, 195–206.
Westerling, A. L., Gershunov, A., Brown, T. J., et al. (2003). Climate and wildfire in the Western United States. Bulletin of the American Meteorological Society, 84, 595–604.
Westerling, A. L., Hidalgo, H. G., Cayan, D. R., & Swetnam, T. W. (2006). Warming and earlier spring increase western U.S. forest wildfire activity. Science, 313, 940–943.
Westerling, A. L., Turner, M. G., Smithwick, E. A. H., et al. (2011). Continued warming could transform Greater Yellowstone fire regimes by mid-21st century. Proceedings of the National Academy of Sciences, USA, 108, 13165–13170.
Whitlock, C., Shafer, S. L., & Marlon, J. (2003). The role of climate and vegetation change in shaping past and future fire regimes in the northwestern US and the implications for ecosystem management. Forest Ecology and Management, 178, 5–21.
Williams, D. W., & Liebhold, A. M. (1995a). Forest defoliators and climatic change: Potential changes in spatial distribution of outbreaks of western spruce budworm (Lepidoptera: Tortricidae) and gypsy moth (Lepidoptera: Lymantriidae). Environmental Entomology, 24, 1–9.
Williams, D. W., & Liebhold, A. M. (1995b). Herbivorous insects and global change: Potential changes in the spatial distribution of forest defoliator outbreaks. Journal of Biogeography, 22, 665–671.
Willis, C. G., Ruhfel, B. R., Promack, R. B., et al. (2010). Favorable climate change response explains non-native species’ success in Thoreau’s woods. PLoS One, 5, e8878.
Woods, A. J., Heppner, D., Kope, H. H., et al. (2010). Forest health and climate change: A British Columbia perspective. The Forestry Chronicle, 86, 412–422.
Worrall, J. J., Egeland, L., & Eager, T. (2008). Rapid mortality of Populus tremuloides in southwestern Colorado, USA. Forest Ecology and Management, 255, 686–696.
Ylioja, T., Slone, D. H., & Ayres, M. P. (2005). Mismatch between herbivore behavior and demographics contributes to scale-dependence of host susceptibility in two pine species. Forest Science, 51, 522–531.
Zavaleta, E. S., Hobbs, R. J., & Mooney, H. A. (2001). Viewing invasive species removal in a whole-ecosystem context. Trends in Ecology & Evolution, 16, 454–459.
Ziska, L. H., & Dukes, J. S. (2011). Weed biology and climate change (248pp). Ames: Wiley-Blackwell.
Ziska, L. H., & George, K. (2004). Rising carbon dioxide and invasive, noxious plants: Potential threats and consequences. World Resource Review, 16, 427–447.
Ziska, L. H., & Teasdale, J. R. (2000). Sustained growth and increased tolerance to glyphospsate observed in a C3 perennial weed quackgrass (Elytrigia repens), grown at elevated carbon dioxide. Australian Journal of Plant Physiology, 27, 159–166.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2014 Springer Science+Business Media Dordrecht (outside the USA)
About this chapter
Cite this chapter
Ayres, M.P. et al. (2014). Disturbance Regimes and Stressors. In: Peterson, D., Vose, J., Patel-Weynand, T. (eds) Climate Change and United States Forests. Advances in Global Change Research, vol 57. Springer, Dordrecht. https://doi.org/10.1007/978-94-007-7515-2_4
Download citation
DOI: https://doi.org/10.1007/978-94-007-7515-2_4
Published:
Publisher Name: Springer, Dordrecht
Print ISBN: 978-94-007-7514-5
Online ISBN: 978-94-007-7515-2
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)