
Abstract
Prominent among the many factors now affecting the sustainability of forest ecosystems are anthropogenically-generated carbon dioxide (CO2) and ozone (O3). CO2 is the substrate for photosynthesis and thus can accelerate tree growth, whereas O3 is a highly reactive oxygen species and interferes with basic physiological functions. This review summarizes the impacts of CO2 and O3 on tree chemical composition and highlights the consequences thereof for trophic interactions and ecosystem dynamics. CO2 and O3 influence phytochemical composition by altering substrate availability and biochemical/physiological processes such as photosynthesis and defense signaling pathways. Growth of trees under enriched CO2 generally leads to an increase in the C/N ratio, due to a decline in foliar nitrogen and concomitant increases in carbohydrates and phenolics. Terpenoid levels generally are not affected by atmospheric CO2 concentration. O3 triggers up-regulation of antioxidant defense pathways, leading to the production of simple phenolics and flavonoids (more so in angiosperms than gymnosperms). Tannins levels generally are unaffected, while terpenoids exhibit variable responses. In combination, CO2 and O3 exert both additive and interactive effects on tree chemical composition. CO2-and O3-mediated changes in plant chemistry influence host selection, individual performance (development, growth, reproduction), and population densities of herbivores (primarily phytophagous insects) and soil invertebrates. These changes can effect shifts in the amount and temporal pattern of forest canopy damage and organic substrate deposition. Decomposition rates of leaf litter produced under elevated CO2 and O3 may or may not be altered, and can respond to both the independent and interactive effects of the pollutants. Overall, however, CO2 and O3 effects on decomposition will be influenced more by their impacts on the quantity, rather than quality, of litter produced. A prominent theme to emerge from this and related reviews is that the effects of elevated CO2 and O3 on plant chemistry and ecological interactions are highly context- and species-specific, thus frustrating attempts to identify general, global patterns. Many of the interactions that govern above- and below-ground community and ecosystem processes are chemically mediated, ultimately influencing terrestrial carbon sequestration and feeding back to influence atmospheric composition. Thus, the discipline of chemical ecology is fundamentally important for elucidating the impacts of humans on the health and sustainability of forest ecosystems. Future research should seek to increase the diversity of natural products, species, and biomes studied; incorporate long-term, multi-factor experiments; and employ a comprehensive “genes to ecosystems” perspective that couples genetic/genomic tools with the approaches of evolutionary and ecosystem ecology.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Forest ecosystems have played roles of unparalleled importance in the evolution and ecology of life on Earth. Currently, forests cover ∼30% of the earth’s land surface, store ∼45% of terrestrial carbon, and contribute ∼50% of terrestrial net primary production (Bonan 2008). They provide vital ecological, economic and social services, including regulation of biogeochemical cycles, production of food and fiber, preservation of biodiversity, governing of climate systems, and buffering against natural and anthropogenic disasters (Hassan et al. 2005; Millenium Ecosystem Assessment 2005).
Earth’s forests are now subject to a veritable Pandora’s Box of environmental evils, the magnitude and diversity of which have never occurred previously in human history. Worldwide, climate and land use changes, invasive species, pest outbreaks, and pollution are exacting an increasing toll on the health, biological diversity, and sustainability of forests (Laurence and Peres 2006; Hari and Kulmala 2008; Kurz et al. 2008; Raffa et al. 2008; van Mantgem et al. 2009). Against this backdrop of massive and seemingly intractably complex socio-environmental ills, the question arises: “of what relevance is chemical ecology?”
In a word, plenty. Naturally-produced chemicals, particularly phytochemicals, govern many, if not most, of the interactions that characterize community and ecosystem function. They respond to global environmental change; they perpetuate, via interaction networks, the consequences of global change; and they feed back to influence future global change. Consider, for example, the carbon cycle (Fig. 1). Enriched atmospheric CO2 influences plant physiology, with consequences for plant production and chemical composition. Plant chemical composition in turn influences trophic interactions and decomposition, which ultimately feed back to affect atmospheric CO2 concentrations (Peñuelas and Estiarte 1998; Beedlow et al. 2004; Lindroth and Dearing 2005). Or, consider the isoprenoids and related biogenic volatile organic carbons (BVOCs) released in large quantities by some species of trees (e.g., eucalypts, poplars, and conifers). These compounds are precursors for the formation of tropospheric ozone (O3), which in turn negatively impacts plant production (Fig. 1) and human health (Fowler et al. 1999; Lerdau and Gray 2003; Monson 2003; Laothawornkitkul et al. 2009). An understanding of the roles of natural products as responders to, and effectors of, global change will improve our ability to predict ecosystem responses to future change, and inform management decisions with respect to mitigation and adaptation strategies.

Carbon cycling and storage in forest ecosystems. Solid boxes represent major pools; dashed boxes represent major plant physiological processes (adapted from Lindroth and Dearing 2005)
Analogous in some respects to intestinal villi, forest trees provide structurally expansive and heterogeneous surfaces through which the exchange of chemicals between the atmosphere and biosphere is effected. The same properties enhance exposure of forest trees to gaseous pollutants. Two such gases, CO2 and O3, are considered the most important and ubiquitous, affecting forests worldwide (Saxe et al. 1998; Fowler et al. 1999; Felzer et al. 2004; Karnosky et al. 2005, 2007). Independent of their function as greenhouse gases, CO2 and O3 are directly and indirectly affecting forest ecosystems now, with increasingly pronounced impacts predicted for the future (Fig. 2). (Note that although both CO2 and O3 occur naturally, they are appropriately considered “pollutants” because of their anthropogenic production and effects on living systems.)

Complex of interacting factors that feed forward and feed back to influence food webs, community and ecosystem structure and function, and atmospheric composition
The purpose of this review is to assess the impacts of enriched atmospheric CO2 and O3, independently and interactively, on the chemical ecology of forest ecosystems. Several recent reviews provide an assessment of the effects of CO2 and O3 on plant chemistry; that information will be summarized and updated here. I will then explore the consequences of CO2- and O3-mediated changes in tree chemistry for trophic interactions (plant—animal—natural enemy) and ecosystem dynamics (rates of plant damage, organic substrate deposition and decomposition) (Fig. 1). I will not detail the links between ozone and plant volatiles, as those have been covered in several very recent reviews, including elsewhere in this issue (Laothawornkitkul et al. 2009; Pinto et al. 2010; Yuan et al. 2009). Finally, although CO2 and O3 are both greenhouse gases, the effects of climate change per se on ecological interactions (e.g., Bale et al. 2002; Parmesan 2006; Berggren et al. 2009; Dukes et al. 2009; Pelini et al. 2009) are beyond the scope of this review.
Temporal Trends in Concentrations of CO2 and O3
Concentrations of atmospheric CO2 now approach 386 ppm, higher than at any time in the last 26 million years (Pearson and Palmer 2000) and 40% higher than at the dawn of the industrial revolution. Emissions scenarios predict a range of substantially elevated concentrations by the end of this century, from a low of 550 ppm to a high of 900 ppm (Karl et al. 2009). Over the last several decades, combustion of fossil fuels, coupled with a smaller contribution from manufacture of cement, have accounted for approximately 80% of human-caused emissions. The remaining 20% has resulted from changes in land use, primarily deforestation (Denman et al. 2007; Karl et al. 2009).
Since the evolution and spread of large, vascular plants, about 380 MYA, forests have played prominent, regulating roles with respect to atmospheric CO2 concentrations. They were largely responsible for the massive drawdown of atmospheric CO2 (peaking at upwards of 5,000 ppm) in the late Devonian and Carboniferous Periods. Forests exerted two major effects on the global carbon cycle, both of which reduced atmospheric CO2 concentrations (Berner 2005). First, they produced massive amounts of lignified tissue that was refractory to decomposition. This organic material, upon sedimentary burial, led to the formation of enormous coal deposits. Second, large and deep root systems greatly accelerated nutrient uptake from rock substrates, enhancing the weathering of Ca-Mg silicate minerals. Ca and Mg cations, in the presence of carbonic acid (derived from atmospheric CO2 and water), formed Ca-Mg carbonates, which precipitated in ocean sediments to form limestone and dolomite.
Tropospheric ozone is recognized as the most damaging and widespread pollutant affecting agricultural and forest ecosystems in North America and Europe (Chameides et al. 1994; Ollinger et al. 1997; Skärby et al. 1998; Fuhrer and Booker 2003; Ashmore 2005; Karnosky et al. 2007; Wittig et al. 2009). Concentrations of O3 have increased an average of 38% (range of 20–50%) since the pre-industrial age (Denman et al. 2007). Although O3 is a regional pollutant, large areas of the earth already are impacted, background levels are steadily increasing, and 60% of the world’s forests are expected to be affected detrimentally by the end of this century (Fowler et al. 1999; Vingarzan 2004; Denman et al. 2007).
O3 is a secondary pollutant, produced via the catalytic oxidation of hydrocarbons (e.g., VOCs) by OH radicals, in the presence of nitrogen oxides and sunlight (Wennberg and Dabdub 2008). Because O3 is highly reactive, it persists for short periods (hours to several weeks) in the atmosphere. Rapid degradation, coupled with substantial spatial and temporal variation in production, contribute to highly variable distribution of this pollutant. Regional VOC and NOx emission control policies are expected to reduce peak O3 levels in North American and Europe, whereas in Asia and other regions, emissions of these precursors are increasing (Ashmore 2005). Moreover, background levels of O3 (not attributable to local origin) are increasing steadily worldwide (Vingarzan 2004).
Plant-derived, biogenic VOCs can be important contributors to the formation of O3 (Sharkey and Lerdau 1999; Lerdau and Gray 2003; Laothawornkitkul et al. 2009; Yuan et al. 2009). When BVOCs such as isoprene, monoterpenes or methylbutenol are oxidized in the presence of NOx, substantial amounts of O3 can be produced. Indeed, large inputs of BVOCs may handicap efforts to reduce ozone pollution through controlling emissions of anthropogenic VOCs. Moreover, ecological interactions may amplify ozone production. A modeling study by Litvak et al. (1999) indicated that even modest (10%) damage by defoliating insects in coniferous forests is sufficient to increase local concentrations of O3.
In contrast, in the presence of low NOx concentrations, BVOCs are oxidized by O3, leading to a decrease in atmospheric O3 and a concomitant increase in secondary organic aerosols (Laothawornkitkul et al. 2009). Thus, forest ecosystems can serve as effective quenchers of anthropogenic O3, but in the process contribute to atmospheric aerosol loading that influences climate. Kurpius and Goldstein (2003) documented that for a Pinus ponderosa forest during summer, 51% of atmospheric O3 is broken down via gas phase chemistry (reactions with BVOCs), 30% is lost to stomatal uptake, and 19% is lost to surface deposition.
Effects of CO2 and O3 on Plant Physiology and Growth
Carbon Dioxide
As the substrate for photosynthesis, and ultimate source of all carbon fixed via primary production, atmospheric CO2 strongly influences plant physiology and growth. Photosynthesis is catalyzed by Rubisco (ribulose-1,5-bisphosphate carboxylase oxygenase), which fixes carbon to form carbohydrates. In C3 plants, including most forest tree species, enriched CO2 atmospheres increase the intracellular CO2:O2 ratio, thus reducing photorespiration (oxygenation) and increasing photosynthesis (carboxylation). Carbohydrates then are transported throughout the plant and subsequently converted to various carbon-containing compounds required for plant metabolism, structure, storage and defense (Fig. 3).

Biosynthetic pathways for the production of the major classes of plant secondary compounds (adapted from Taiz and Zeiger 2002). Dashed lines indicate a control (signal transduction) pathway. PAR = photosynthetically active radiation
The effects of elevated atmospheric CO2 on tree physiology and growth have been covered in numerous reviews (e.g., McGuire et al. 1995; Saxe et al. 1998; Long et al. 2004; Ainsworth and Long 2005; Norby et al. 2005; Körner 2006; Leakey et al. 2009). In brief, N use efficiency improves as the N allocated to Rubisco (the largest single pool of N in leaves) can be re-allocated to other metabolic processes. Stomatal conductance generally declines, decreasing transpiration and improving water use efficiency. Enhanced photosynthesis, coupled with improved nitrogen and water use efficiencies, often accelerates plant growth, i.e., the “fertilizer effect” of enriched CO2 (Beedlow et al. 2004). The growth stimulation afforded by enriched CO2 is especially strong in young forest stands, characterized by abundant soil resources and open canopies. In contrast, studies in mature stands, where tree growth is tightly coupled to soil nutrients and canopies are closed, indicate smaller to zero growth enhancement (Asshoff et al. 2006; Körner 2006).
A key, unresolved issue in global change biology is to what extent improved tree growth can be sustained into the future, affording greater carbon sequestration and thereby mitigating climate change. To date, open-air FACE studies generally indicate that accelerated rates of photosynthesis and growth are sustained over the short to medium term and across a broad range of productivity (Long et al. 2004; Norby et al. 2005; Huang et al. 2007; but see Körner et al. 2005 for a contrary example with mature trees). Still, the duration of these studies is short compared with the lifespan of trees. The Progressive Nitrogen Limitation (PNL) hypothesis (Luo et al. 2004) posits that CO2-enhanced growth and carbon sequestration will diminish over time: as storage of N in plant biomass and soil organic matter increases, available soil N becomes limiting to growth. If not nitrogen, then a host of other factors (climate change, fire, insects, ozone, etc.) is predicted to constrain the fertilization effect of atmospheric CO2 enrichment (Beedlow et al. 2004; Huang et al. 2007). For example, Cole et al. (2009) reported that atmospheric CO2 enrichment over the last 50 years has increased growth rates of natural stands of Populus tremuloides by an astonishing 53%, but enhancement is diminished in periods of low precipitation. Finally, elevated atmospheric CO2 may, via climate change impacts, ultimately lead to reduced carbon sequestration at the landscape level. In Canada, for example, extensive areas of forest recently have transitioned from CO2 sinks to CO2 sources, due to insect outbreaks and fires (Kurz et al. 2008).
Ozone
Reactions of tropospheric O3 with plant chemistry can occur in gaseous, solid, or liquid phases (Fuhrer and Booker 2003). Gaseous reactions with plant VOCs are described by Pinto et al. (2010). Solid phase reactions may occur, for example, with plant cuticular components. The most important reactions for plants, however, occur in the liquid phase, after ozone has entered through the stomata and diffused into the aqueous media of interstellar spaces (Fuhrer and Booker 2003).
O3 elicits a cascade of damaging physiological consequences in plants, described in detail in numerous recent reviews (e.g., Andersen 2003; Fuhrer and Booker 2003; Ashmore 2005; Kangasjärvi et al. 2005; Wittig et al. 2007, 2009). Because it is so highly reactive, O3 is minimally distributed following dissolution into intercellular fluids. It reacts with lipid and protein components of cell walls and plasma membranes, leading to the formation of aldehydes, peroxides and assorted reactive oxygen species (ROS). These products can then cause cell death, or activate various transduction pathways for defense responses, such as stomatal closure, production of anti-oxidants such as ascorbate and phenolics, and programmed cell death (a component of the hypersensitive response) (Fuhrer and Booker 2003; Valkama et al. 2007; Fig. 3). Reduced stomatal conductance, compromised photosynthesis, consumption of carbohydrates in localized tissue repair, and inhibition of phloem transport at the leaf level combine to decrease the supply of carbohydrates throughout the plant (Andersen 2003; Wittig et al. 2007, 2009). As a consequence, senescence is accelerated and whole-plant growth is inhibited.
A recent, comprehensive meta-analysis by Wittig et al. (2009) evaluated the effects of O3 on the growth of trees representative of northern temperate and boreal forests. They found that, relative to preindustrial levels, current levels of tropospheric O3 account for an average 7% reduction in tree biomass growth. Moreover, as O3 levels continue to rise, further decreases of 11% and 17% are predicted for 2050 and 2100, respectively.
Understanding of the interactive effects of CO2 and O3 on tree growth is critically important to our ability to predict the efficacy of forests to serve as global carbon sinks in the future (Beedlow et al. 2004). A modeling study by Sitch et al. (2007) indicated that tropospheric O3 may reduce terrestrial carbon sinks, and thereby effect indirect radiative forcing of climate change. This conclusion was subsequently corroborated by the work of Wittig et al. (2009), which indicated increasingly pronounced reductions in tree growth under elevated concentrations of O3 in the future. To date, only one experiment has addressed the long-term (10+ years) effects of independent and combined CO2 and O3 treatments on forest trees. Karnosky et al. (2005) reported that at Aspen FACE (Wisconsin, U.S.A.), O3 reduced the growth-enhancement effect of CO2, although responses differed among tree species.
Effects of CO2 and O3 on Phytochemistry
CO2 and O3 directly and indirectly influence carbon assimilation and nutrient (e.g., N) acquisition, and thereby the pools and fluxes of key precursors for the production of secondary metabolites (Fig. 3). They also may influence the signal transduction pathways that govern gene expression and subsequent synthesis of secondary compounds. Not surprisingly then, growth under CO2- or O3-enriched atmospheres typically alters the chemical composition of forest trees.
Numerous reviews have addressed the effects of CO2 and O3 on plant chemistry (Kangasjärvi et al. 1994; Peñuelas et al. 1997; Bezemer and Jones 1998; Cotrufo et al. 1998a; Koricheva et al. 1998a; Peñuelas and Estiarte 1998; Norby et al. 2001; Zvereva and Kozlov 2006; Stiling and Cornelissen 2007; Valkama et al. 2007; Bidart-Bouzat and Imeh-Nathaniel 2008; Wittig et al. 2009). Unless noted otherwise, the general trends for forest trees, summarized below, are drawn from that comprehensive set of reviews. Specific studies will be mentioned to illustrate particular patterns or highlight recent findings.
To date, research on the effects of CO2 and O3 on forest trees has been restricted to a small set of primarily temperate species. Most commonly studied are species of Acer, Betula, Fagus, Pinus, Populus, and Quercus. Although these are prominent, if not “foundation,” genera, a major challenge to the global change research community is to broaden the taxonomic and geographic representation of trees, especially tropical species.
Carbon Dioxide
Enriched atmospheric CO2 produces generally consistent effects on concentrations of primary metabolites in trees (Cotrufo et al. 1998a; Koricheva et al. 1998a; Saxe et al. 1998; Stiling and Cornelissen 2007). Foliar concentrations of nitrogen typically decline (on average, 14–17% relative to ambient CO2), due to reductions in Rubisco concentrations and dilution by accumulating carbohydrates. Levels of simple sugars may or may not increase, whereas levels of starch generally increase substantially.
Studies of the effects of CO2 on secondary metabolites in trees have been restricted to “carbon-based” compounds, particularly phenolics and terpenoids. Consequently, this research often has been framed in the context of source-sink balance hypotheses [e.g., carbon-nutrient balance hypothesis (Bryant et al. 1983), growth-differentiation balance hypothesis (Herms and Mattson 1992)]. These hypotheses predict that elevated CO2 will increase plant concentrations of carbon-based secondary compounds, especially if soil nutrient availability is limiting to growth (Peñuelas and Estiarte 1998).
In general, results have not been as readily predictable as the hypotheses would suggest (reviewed in Koricheva et al. 1998a; Peñuelas and Estiarte 1998; Zvereva and Kozlov 2006; Stiling and Cornelissen 2007; Bidart-Bouzat and Imeh-Nathaniel 2008). Moreover, the shikimic acid pathway, leading to production of phenolics, is more strongly influenced by CO2 levels than are the mevalonic acid and methylerythritol phosphate pathways, leading to production of terpenoids (Fig. 3). Enriched atmospheric CO2 typically, but not invariably, increases tannin concentrations. Common, but less consistent, are increases in simple phenolics, including phenolic acids and glycosides (e.g., salicylates). Growth under elevated CO2 also tends to increase lignin concentrations in tree leaves (Coûteaux et al. 1999; Norby et al. 2001), although more recent results, from long-term FACE studies, have shown no such effects (Finzi et al. 2001; Parsons et al. 2008; Liu et al. 2009). Overall, effects of CO2 on terpenoid concentrations are minimal; although a few studies have revealed increases, most have found no change or decreases in concentrations.
The C/N ratio is a widely recognized, general index of overall tissue quality, relevant to both herbivory and decomposition. Enriched CO2 atmospheres almost invariably increase C/N ratios in tree foliage (Coûteaux et al. 1999), due to a combination of decreased N concentrations and increased carbohydrate and phenolic concentrations.
Aside from such general indices, however, empirical studies have frustrated attempts to develop general, predictive models of the impacts of CO2 on plant secondary chemistry. In short, a host of interacting factors have, over evolutionary time, combined to determine the chemical profiles of trees, and the effects of any one environmental factor can be masked by the direct or interactive effects of others.
One of the major confounding factors is taxon-specificity. Phytochemical responses of trees to CO2 enrichment differ at the level of class (e.g., gymnosperm vs. angiosperm), species, and genotype (Julkunen-Tiitto et al. 1993; Lindroth et al. 2001, 2002; Zvereva and Kozlov 2006; Stiling and Cornelissen 2007; Bidart-Bouzat and Imeh-Nathaniel 2008). These differences represent taxonomic variation in evolutionary strategies that optimize solutions for the conflicting demands of carbon allocation toward growth, reproduction, and defense. For example, Cseke et al. (2009) recently compared leaf transcription profiles, physiology, and biochemistry between CO2-responsive and unresponsive clones of Populus tremuloides grown at Aspen FACE. They found that the CO2-responsive clone partitions carbon into pathways associated with “active” defense and stress responses, carbohydrate synthesis, and subsequent growth, whereas the unresponsive clone partitions carbon into pathways associated with cell wall compounds such as lignin, derived from the shikimic acid pathway.
Interactions with abiotic environmental factors also influence chemical responses of trees to CO2. The availability of key resources (e.g., nutrients, water, light) can strongly shape plant chemical responses to CO2, but, again, the magnitude and even direction of responses differ among chemical constituents and tree species (e.g., Lavola and Julkunen-Tiitto 1994; Kinney et al. 1997; McDonald et al. 1999; Booker and Maier 2001; Coley et al. 2002; Koike et al. 2006). Of particular importance in the context of global environmental change are potential interactions between atmospheric CO2 and temperature. To that end, Zvereva and Kozlov (2006) conducted a meta-analysis of studies that assessed the simultaneous elevation of CO2 and temperature on plant quality. Again, results were compound- and taxon-specific. Thus, responses to enriched CO2 may be: 1) independent of temperature (e.g., foliar nitrogen and phenolics in Angiosperms); 2) offset by elevated temperature (e.g., carbohydrates; foliar terpenes in gymnosperms); or 3) apparent only with elevated temperature (e.g., foliar nitrogen in gymnosperms, phenolics, and terpenoids in woody tissues). More recently, Veteli et al. (2007) reported for three tree species (Betula and Salix) that carbon allocation to phenolics was increased under elevated CO2, decreased under elevated temperature, and unchanged under a combination of elevated CO2 and temperature.
Interactions with biotic agents also may influence chemical responses of trees to CO2, although much less research has been conducted with biotic than with abiotic factors. Herbivory frequently elicits induced chemical changes in trees, particularly in levels of phenolics (Nykänen and Koricheva 2004). Thus, enriched CO2 atmospheres have been predicted to enhance induced chemical responses. Evidence to date, however, suggests that CO2 does not modify induced chemical responses to defoliation in trees, although only a few studies have been conducted (Lindroth and Kinney 1998; Roth et al. 1998; Agrell et al. 1999; Rossi et al. 2004; Hall et al. 2005; Huttunen et al. 2008).
A final factor complicating attempts to draw general conclusions about the effects of CO2 on tree chemistry is ontogeny (genetically-determined developmental changes). Many species of trees exhibit strong ontogenetic shifts in chemical composition (e.g., Bryant and Julkunen-Tiitto 1995; Donaldson et al. 2006), which reflect shifting physiological demands and defense strategies as trees mature (Boege and Marquis 2005). How ontogeny may interact with CO2 to affect chemical profiles in trees is unknown. Such interactions may be one reason, however, for why chemical responses of plants in long-term FACE studies appear to be less pronounced than in short-term pot studies (Ainsworth and Long 2005; Lindroth, unpublished data).
Ozone
Elevated levels of tropospheric O3 can modify plant chemical composition via several mechanisms. By disrupting photosynthesis, O3 inhibits production of carbohydrates and the flow of precursors into pathways of primary and secondary metabolism (Fig. 3). Ozone also functions as a general abiotic elicitor of defense signaling pathways, particularly of phenolic compounds (Kangasjärvi et al. 1994; Heath 2008; Betz et al. 2009a). The two mechanisms may, of course, interact, as reductions in carbohydrate precursors for the shikimic acid pathway may constrain induced defensive responses. Moreover, these mechanisms may play out differently over different time scales, resulting in varying responses to acute versus chronic ozone stress.
The effects of O3 on levels of primary metabolites vary among compounds and tree species. Levels of nutrients (e.g., N, P, K, Ca) may increase, decrease, or not change in response to O3 fumigation. Levels of simple sugars generally are unaffected, whereas levels of starch typically decline in angiosperms, but not gymnosperms.
As is true for CO2, studies of the effects of O3 on secondary compounds in trees have been restricted almost entirely to phenolics and terpenoids. In general, elevated O3 leads to increases in concentrations of phenolic acids and flavonoids in angiosperms but not in gymnosperms, and no overall change in concentrations of tannins. Foliar lignin concentrations also are generally unresponsive to O3 fumigation (Boerner and Rebbeck 1995; Booker et al. 1996; Liu et al. 2005; Oksanen et al. 2005). Effects of O3 on terpenoid levels have been studied in only a few tree species, and results are species-specific: increases in Pinus sylvestris and Populus species, and no change in Picea abies (Valkama et al. 2007; Blande et al. 2007). Moreover, the magnitude of response varies among types of terpenes, with diterpenes responding more strongly to O3 than mono- or sesquiterpenes.
That O3 elicits increases in the levels of some carbon-based secondary metabolites without corresponding increases in carbohydrates runs counter to the predictions of source-sink balance hypotheses (e.g., Bryant et al. 1983), and may be explained by the fact that ozone triggers up-regulation of antioxidant defense systems linked to the shikimic acid pathway (Valkama et al. 2007). Ozone causes oxidative damage, coupled to the proliferation of oxygen radicals. Gene transcription and activity of numerous enzymes in the shikimic acid pathway are elevated in response to O3 exposure, leading to the production of antioxidants such as flavonoids and other simple phenolics, and shifts in the monomeric composition of lignin (Kangasjärvi et al. 1994; Heath 2008; Betz et al. 2009a, b). Indeed, ozone-induced phenolics have been considered potential bioindicators of O3 stress in plants under natural conditions (Sager et al. 2005; Bidart-Bouzat and Imeh-Nathaniel 2008).
Ozone exposure also influences the amount and chemical composition (e.g., alkyl esters and fatty acids vs. alkanes and alkanols) of cuticular waxes in tree leaves (Karnosky et al. 2002; Kontunen-Soppela et al. 2007; Percy et al. 2009). These changes in turn determine the physical structure of the cuticular surface, with repercussions for leaf surface properties such as wettability.
In contrast to a large body of research with CO2, little work has addressed how O3 pollution interacts with other environmental factors to shape plant chemical profiles. Lindroth et al. (1993a) evaluated the effects of elevated O3 and light intensity on several foliar constituents of hybrid poplar and sugar maple (Acer saccharum), and found that O3 and light interacted to affect only concentrations of N in maple. Given that forest ecosystems are exposed to multiple, simultaneous stress factors (e.g., climate change, insect outbreaks) in addition to increasing O3 damage, and that some of those stressors induce the same chemical defense pathway as does O3, such research is sorely needed.
Interactions Between CO2 and O3
The environmental factor that has been investigated most thoroughly in combination with O3 pollution is CO2. Enriched CO2 may reduce ozone damage to trees both by decreasing stomatal conductance (and consequent oxidative stress) and by increasing carbohydrate pools for the synthesis of antioxidant compounds. The recent meta-analysis by Valkama et al. (2007) evaluated the effects of O3, and O3 combined with CO2, on foliar chemistry of 22 species of trees. They found that enriched CO2 can offset O3-induced chemical changes, but their analysis did not differentiate between simple additive, versus interactive, effects of CO2 and O3.
Indeed, CO2 and O3 exert both independent (additive) and interactive effects on tree chemical profiles. Elevated O3 concentrations exacerbated the CO2-mediated reductions in N concentrations in Betula papyrifera (Kopper et al. 2001) and Populus tremuloides (Holton et al. 2003), but offset reductions in N in Betula pendula (significant CO2 × O3 interactions). Peltonen et al. (2005) assessed the effects of CO2 and O3 on over thirty simple phenolics in B. pendula. They identified interactive effects in approximately three-quarters, and independent effects in one-quarter, of the cases. The interactive effects revealed that O3-mediated induction of phenolics disappeared under elevated CO2. Tannin levels also are influenced by both additive and interactive effects of CO2 and O3. Condensed tannin concentrations responded to the additive, independent effects of CO2 and O3 in Fagus crenata and various species of Betula (Kopper et al. 2001; Peltonen et al. 2005; Karonen et al. 2006), but to the interactive effects of CO2 and O3 in Populus tremuloides (Holton et al. 2003). Similarly, the effects of CO2 and O3 on monoterpenes and sesquiterpenes in Pinus sylvestris sometimes were additive and sometimes interactive (Sallas et al. 2001). In short, whether CO2 and O3 function in an additive or interactive manner depends on the particular chemical constituent and tree species studied.
Effects of CO2 and O3 on Trophic Interactions
Plant chemical composition is a strong ecological and evolutionary driver of trophic interactions, particularly between plants and herbivores. Given that green plants and phytophagous insects comprise nearly half of Earth’s recognized species (Strong et al. 1984), it is not surprising that these taxa occupy the vast majority of all studies on plant-herbivore interactions. Several recent reviews have addressed the effects of CO2 and O3 on plant-insect interactions (Zvereva and Kozlov 2006; Stiling and Cornelissen 2007; Valkama et al. 2007; Bidart-Bouzat and Imeh-Nathaniel 2008). Composite results will be summarized below and supplemented with additional, relevant studies of tree-feeding herbivores.
Carbon Dioxide
Little evidence exists to suggest that elevated CO2, upwards of 1,000 ppm predicted for this century, will directly affect insects (Coviella and Trumble 1999). Work by Stange (1997) showed that atmospheric CO2 concentrations influence host location by Cactoblastis cactorum, but no similar work has been done with forest insects. Rather, the major effect of enriched CO2 on herbivores will be indirect, mediated via changes in plant quality (Fig. 2). Overall, elevated CO2 reduces the quality of tree foliage as food, due to decreases in nitrogen (an index of protein) and minerals, and increases in carbohydrates and phenolics. These changes typically, but not always, alter the preference and reduce the performance of herbivores.
CO2 concentration influences selection of plants by insects and mammals. Elevated CO2 reduced leafminer oviposition on Populus tremuloides (Kopper and Lindroth 2003a), but increased birch aphid oviposition on one of two clones of B. pendula (Peltonen et al. 2006). The few feeding studies conducted to date suggest that preferences of insects for ambient versus elevated CO2 foliage are not easily predicted, as shifts in both directions, and no change, have been reported (Traw et al. 1996; Kuokkanen et al. 2003; Agrell et al. 2005; Knepp et al. 2007). Elevated CO2 decreased the palatability of winter-dormant Betula species to hares and rabbits (Mattson et al. 2004), but not to voles (Kuokkanen et al. 2004). Of greater relevance in the universally CO2-enriched world of the future, however, is whether enrichment alters relative preferences of herbivores for various host species. Minimal evidence to date suggests that such is the case. For example, Agrell et al. (2005) demonstrated that forest tent caterpillars switch relative preferences between Populus tremuloides and Betula papyrifera, as well as between P. tremuloides genotypes, grown in high CO2 environments.
Elevated CO2 also influences the individual performance of herbivores, although the effects are highly species-specific for both trees and insects. Among the more uniform (though still variable) of responses is food consumption. In general, insects increase both consumption rates, and total consumption, on CO2-enriched foliage, ostensibly as a means to compensate for reduced N concentrations. Compensatory feeding responses may be constrained, however, by the simultaneous consumption of higher concentrations of secondary metabolites such as phenolics. Approximate digestibility of food typically is not significantly affected, but conversion efficiencies of digested food into biomass generally decline. Little is known about how CO2 environment affects insect detoxication capacities; a study with gypsy moths revealed enhanced oxidase, reductase, and esterase activities in larvae fed CO2-enriched P. tremuloides, but not Acer saccharum (Lindroth et al. 1993b). Development times usually are prolonged, while growth rates and final pupal and adult weights decline or are unaffected, but rarely improve. Effects on reproduction have rarely been investigated. Lindroth et al. (1997) found no effect of CO2 on the number and mass of eggs produced by gypsy moths, whereas Awmack et al. (2004) reported deceased fecundity in aphids on B. papyrifera under elevated CO2.
Species-specificity in terms of insect herbivore responses to enriched CO2 may be linked to differences among feeding guilds (e.g., chewers, miners, sap-feeders, seed-feeders). Bezemer and Jones (1998) identified such guild effects for plants and phytophagous insects in general. Too few studies have been conducted with tree-feeding insects to evaluate differences among guilds. However, the minimal, if not positive effects of CO2 on tree-feeding aphids (Docherty et al. 1997; Awmack et al. 2004), in contrast to effects on chewing insects, suggest that guild-specific responses to CO2 may exist for forest insects.
Just as interactions with various environmental factors influence the effects of CO2 on plant chemistry, so may they modulate effects on herbivores. For example, the effects of elevated CO2 on herbivore performance can be ameliorated or exacerbated by the availability of light, water, and soil nutrients (e.g., Kinney et al. 1997; Lawler et al. 1997; Roth et al. 1997; Hättenschwiler and Schafellner 1999; Agrell et al. 2000). Potential interactions with climate warming are of particular interest, as warm temperatures generally affect insects in a manner opposite that of CO2, i.e., accelerated development and enhanced growth and reproductive performance (Bale et al. 2002). Indeed, Zvereva and Kozlov (2006) conclude from their recent meta-analysis that temperature increases are likely to mitigate predicted negative effects of enriched CO2 on insects, and emphasize the need for future research to address the two factors in parallel.
In addition to the bottom-up effects of altered plant chemistry, elevated CO2 may influence the top-down effects of natural enemies (predators, parasitoids, and pathogens) on insect herbivores (Fig. 2). Enriched CO2 could affect natural enemies by altering the behavioral or physiological defenses of prey, modifying the quality of prey (e.g., insect size, concentrations of plant-derived toxins), or by altering the efficacy of host location and disease transmission. Few studies have addressed these possibilities, however, especially with forest systems. Elevated atmospheric CO2 dramatically diminished the pheromone-mediated, predator escape behavior of aphids on Populus tremuloides (Mondor et al. 2004). In contrast, two studies have found the bottom-up effects of CO2 on parasitoid performance (survivorship, development, and growth) to be minimal to nil (Roth and Lindroth 1995; Holton et al. 2003). The virulence of pathogens such as nuclearpolyhedrosis virus (NPV) is linked to host plant chemistry [e.g., tannins (Foster et al. 1992)], so CO2-enriched atmospheres may influence the dynamics of disease outbreaks. CO2-induced increases in condensed tannins in P. tremuloides did not, however, alter the susceptibility of gypsy moths to NPV (Lindroth et al. 1997). At this time insufficient information is available to predict whether the bottom-up effects of CO2 on natural enemies will in general be buffered, amplified, or, more likely, context- and species-specific.
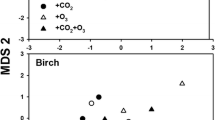
The impacts of elevated CO2 concentrations on forest insects at the levels of populations and communities will be determined by a combination of bottom-up, top-down, and abiotic factors (Fig. 2). Historically, inferences about population-level responses have been drawn from studies of individual insects, but such conclusions can be misleading (Awmack et al. 2004). To date, however, few studies have addressed the impacts of elevated CO2 at higher levels of organization. Several studies with tree-feeding aphids have shown no population-level effects (Docherty et al. 1997; Awmack et al. 2004), while another revealed positive effects of enriched CO2 on aphid population size (Percy et al. 2002). In contrast, Stiling et al. (1999, 2002, 2003, 2009) found reduced densities (number per leaf) of herbivores, especially leafminers and leaftiers, in several woody species of a scrub-oak community exposed to elevated CO2. Reduced herbivore densities were attributed to higher rates of both plant- and parasitoid-induced mortality, and were offset in several Quercus species by increased foliar production under high CO2 (Stiling et al. 2009). Monitoring of aspen blotch leafminers (Phyllonorycter tremuloidiella) at Aspen-FACE indicated that elevated CO2 roughly doubled plant-mediated mortality, substantially reduced predator-mediated mortality, and minimally influenced parasitoid-mediated mortality (Fig. 4).

Effects of CO2 and O3, in isolation and combination, on plant-, predator-, and parasitoid-induced mortality of aspen blotch leafminers (Phyllonorycter tremuloidiella). Data reveal major sources of mortality, prior to adult emergence, for miners on Populus tremuloides. Numerals below bars indicate aspen genotypes. Observations recorded in July 2001 at the Aspen FACE research site (Awmack and Lindroth, unpublished data)
Even fewer studies have addressed the effects of enriched CO2 on speciose insect communities. Following 4 years of pan-trapping at Aspen FACE, Hillstrom and Lindroth (2008) reported that elevated CO2 reduced abundance of phloem-feeding herbivores and increased abundance of chewing herbivores, although results were only marginally significant. Enriched CO2 increased numbers of figitid and ichneumonid parasitoids, but not of braconid or chalcid parasitoids. Insect community composition differed between ambient and elevated CO2 plots in 3 out of 4 years. In the most comprehensive study to date for forest canopy insects, Hillstrom and Lindroth (unpublished data) visually monitored insects on Populus tremuloides and Betula papyrifera over 3 years at Aspen FACE, cataloging over 36,000 insects from nearly 300 species. Overall, enriched CO2 tended to increase the abundance of phloem-feeders and decrease the abundance of chewers and gallers on P. tremuloides, although effects were variable among species and over time. Few consistent effects on abundance were found, however, for insects on B. papyrifera. Elevated CO2 did not affect species diversity of canopy insects, except for insects on P. tremuloides in 1 year. Elevated CO2 also did not influence insect community composition, except for insects on B. papyrifera in 1 year.
Ozone
Increasing evidence suggests that tropospheric ozone pollution may affect directly insects. Gate et al. (1995) found that under elevated O3, searching efficiency of a hymenopteran parasitoid, as well as the proportion of hosts parasitized, declined. O3 likely interacts with the complex of BVOCs that function as “infochemicals” to govern key interactions between insects and both their hosts (plant or insect) and natural enemies (predators and parasitoids) (Laothawornkitkul et al. 2009; Yuan et al. 2009; Pinto et al. 2010, this volume). Chemical degradation or transformation of BVOCs by O3 is likely to increasingly disrupt olfactory cues in atmospheres of the future.
Selection of plants as oviposition or food hosts is influenced by O3 concentrations. O3-treated foliage reduced leaf beetle oviposition onto Populus deltoides (Jones and Coleman 1988) and leafminer oviposition onto Populus tremuloides (Kopper and Lindroth 2003a). Few feeding studies have compared the preferences of insects for fumigated vs. nonfumigated foliage, and those indicate a range of responses. Leaf beetles preferred O3-treated P. deltoides (Jones and Coleman 1988) and common leaf weevils preferred O3-treated hybrid aspen (Freiwald et al. 2008), whereas forest tent caterpillars preferred untreated P. tremuloides (Agrell et al. 2005). Forest tent caterpillars greatly increased their preference for Betula papyrifera, relative to P. tremuloides, under elevated O3, and also shifted their preferences among P. tremuloides genotypes (Agrell et al. 2005).
O3 fumigation changes the individual performance of phytophagous insects, although, as for CO2, effects are highly species-specific. The meta-analysis by Valkama et al. (2007) summarized results for 22 species of trees and ten species of insects. Food consumption rates typically are unaffected, as are survival and reproduction rates. Development times usually are reduced, and pupal masses increased, under elevated O3. For example, Kopper and Lindroth (2003b) reported that for forest tent caterpillars reared on high-O3 Populus tremuloides, development times decreased an average of 14% and pupal weights increased an average of 31% compared with insects on control trees. Relative growth rates of chewing, but not sucking, insects generally increase under elevated O3. On average, insect fecundity is not affected by O3, although Awmack et al. (2004) reported decreased fecundity for aphids on Betula papyrifera.
Under natural conditions, interactions between trees and herbivores likely are influenced by multiple, interacting stressors, including O3 (Koricheva et al. 1998b). With the exception of interactions with CO2 (addressed below), however, exceedingly little is known about how O3 may interact with other environmental factors to influence trophic interactions. Lindroth et al. (1993a) reported that gypsy moth larval performance was influenced by the independent, but not interactive, effects of O3 and light intensity. With the likelihood of increased pest damage to forests under future scenarios of global environmental change, assessments of the combined effects of ozone and other stressors should be a high priority.
A growing body of literature reveals that O3 pollution likely will affect interactions between tree-feeding insects and their natural enemies. As indicated above and described elsewhere in detail (Yuan et al. 2009; Pinto et al. 2010), O3 may directly affect the behavior and fitness of predators and parasitoids. Alternatively, elevated O3 may alter the behavioral or physiological defenses of prey, or their quality as food. For example, Mondor et al. (2004) reported that at Aspen FACE, high O3 environments markedly enhanced predator escape behaviors of aphids on Populus tremuloides. Also at Aspen FACE, O3 fumigation reduced survivorship of the parasitoid Compsilura concinnata (Holton et al. 2003). Decreased fitness was consistent with improved performance of its forest tent caterpillar host, which in turn was linked to decreased levels of phenolic glycosides in P. tremuloides.
Despite accumulating evidence for impacts of O3 at the level of individual insect performance, little work has addressed impacts at the level of populations. Awmack et al. (2004) found that O3 fumigation at Aspen FACE increased population densities of aphids on Betula papyrifera when protected from natural enemies, but not when unprotected. Also at Aspen FACE, elevated O3 levels tended to increase plant-mediated mortality, reduce predator-mediated mortality. and increase parasitoid-mediated mortality of aspen blotch leafminers (Phyllonorycter tremuloidiella), although magnitudes of effects varied among insects on different aspen genotypes (Fig. 4).
At the level of insect communities, pan-trapping at Aspen FACE revealed positive effects of elevated O3 on abundance of some phloem-feeding insects, and strong suppressive effects on a diversity of parasitoids (Hillstrom and Lindroth 2008). O3 influenced the community composition of insects captured by traps in 2 out of 4 years. In our more recent visual censuses of canopy insects at Aspen FACE, we found that O3 fumigation increased abundances of several species of chewing insects, but decreased numbers of several phloem-feeders, on Populus tremuloides (Hillstrom and Lindroth, unpublished data). O3 did not, however, consistently affect abundances of insects on Betula papyrifera. O3 also had generally little impact on the species diversity and community composition of canopy insects on several tree species, and across multiple years, at Aspen FACE.
Interactions Between CO2 and O3
Valkama et al. (2007) sought to differentiate the effects of O3 alone, versus O3 in combination with CO2, on the performance of tree-feeding insects. Their meta-analysis revealed that many of the positive effects of O3 on insect performance, especially for leaf chewers, are negated with simultaneous exposure to enriched CO2. Their review did not, however, differentiate between simple additive, versus truly interactive, effects of CO2 and O3 on insects.
Indeed, relatively few studies have been designed to detect potential interactive effects of CO2 and O3 on forest insect performance, and those have shown mixed results. Several studies (Awmack et al. 2004; Peltonen et al. 2006) have assessed the independent and combined effects of CO2 and O3 on feeding, growth, and reproductive performance of aphids, and revealed no significant interactions. Similarly, no significant interactions were observed with respect to CO2 and O3 effects on leafminers on Populus tremuloides (Kopper and Lindroth 2003a), whitemarked tussock moths feeding on Betula papyrifera (Kopper et al. 2001), or forest tent caterpillar feeding preferences for P. tremuloides and B. papyrifera (Agrell et al. 2005). In contrast, CO2 and O3 interacted to influence development and growth of forest tent caterpillars on P. tremuloides: O3-mediated enhancements in performance were greater at ambient than elevated CO2. In short, the limited evidence to date suggests that the combined effects of CO2 and O3 on insect herbivores will be primarily additive, although the standard caveat of context- and species-specificity applies.
Effects of CO2 and O3 on Ecosystem Dynamics
At the level of forest ecosystems, plant chemistry plays a host of important roles, the most prominent of which arguably is regulation of the transfer of fixed carbon to herbivores and decomposers—the dominant pathways of material flow (Fig. 1). Accumulating evidence suggests that elevated concentrations of CO2 and O3 will modify the complex of physiological and ecological interactions that determine rates of plant damage, organic substrate deposition, and nutrient cycling. In short, the effects of CO2 and O3 on plant chemistry will in turn influence the sequestration and cycling of carbon and nutrients in forest ecosystems.
Forest Canopy Damage
Phytophagous insects generally remove 2–15% of primary production in temperate deciduous forests (Cebrian 1999; Cyr and Pace 1993; Cebrian and Lartigue 2004), although during outbreaks, nearly 100% can be consumed (e.g., Donaldson and Lindroth 2008). Canopy damage rates are a function of the host preferences, individual consumption rates, population densities, and community composition of herbivorous insects, all of which can be influenced by levels of atmospheric CO2 and O3.
The effects of CO2 on loss of primary production to herbivores have been explored in only a few forest systems. In a Florida scrub oak community, enriched CO2 decreased the frequency of damage to Quercus myrtifolia (Stiling et al. 2002). Later work in the same system documented a decline in the frequency of damage by 4 of 6 insect feeding groups, across three Quercus species, under elevated CO2 (Hall et al. 2005). Working with four understory tree species at the FACTS-1 FACE site (loblolly pine—hardwood forest), Hamilton et al. (2004) reported declines of 10–46% in foliar damage rates in high CO2 plots. These declines were driven largely, however, by the response of one species (Ulmus alata); damage rates in other species were not statistically significant from those of control trees. Knepp et al. (2005) followed up that work by censusing foliar damage rates in twelve species of potted saplings at FACTS-1 FACE. Enriched CO2 reduced damage rates across all species in 2001, but not in 2002–2003. In the most comprehensive study to date, Couture, Meehan, and Lindroth (unpublished data) evaluated damage rates on leaves collected from Populus tremuloides and Betula papyrifera over 3 years at Aspen FACE. Elevated CO2 concentrations did not markedly affect herbivory rates in 1 year, but increased damage rates by 2–3-fold in 2 years of the study. Overall, these studies indicate that canopy damage rates are likely to change under CO2 concentrations of the future. But, consistent with effects on individual insects, the magnitude and direction of change will be both species-specific and temporally variable.
To date, no published study has evaluated the effects of elevated tropospheric O3 on community-wide, canopy damage in forest ecosystems. Our work at Aspen FACE (Couture, Meehan and Lindroth, unpublished data) revealed that high levels of O3 reduced damage to both Populus tremuloides and Betula papyrifera in 2 of 3 years.
Calculations of losses to primary production based on leaf areas damaged by herbivores may significantly underestimate true reductions in production. Aldea et al. (2006) showed that in a variety of deciduous tree species, chewing damage caused modest and localized suppression of photosynthetic efficiency in adjacent, undamaged leaf tissue, whereas galling damage caused large and extensive suppression. The authors suggest that the spatial propagation of reduced photosynthesis may be influenced by enriched CO2, but did not observe such effects in their study. More recently, however, P. Nabity and M. Hillstrom (pers. comm.) found that elevated CO2 decreased the propagation of reduced photosynthesis caused by insects feeding on Populus tremuloides and Betula papyrifera at Aspen FACE.
Organic Substrate Deposition
Herbivorous insects mediate the transfer of organic materials from the forest canopy to the forest floor by depositing frass, greenfall, and insect biomass, and altering the timing and amounts of leachate througfall and leaf litterfall. These processes have received little attention in the context of global change research.
We recently evaluated the effects of fumigation treatments at Aspen FACE on the quality of frass produced by whitemarked tussock moth larvae feeding on Populus tremuloides, and the effects of both frass and greenfall on soil microbial respiration and nitrogen leaching (Hillstrom, Meehan, Kelly and Lindroth, unpublished data). Insect frass had slightly decreased N levels in high O3 environments, and increased tannin levels in high CO2 environments. However, neither frass nor greenfall quality affected microbial respiration (CO2 efflux) or nitrogen (NO −3 ) leaching in soil microcosms. In contrast, the quantity of frass and greenfall added to microcosms markedly affected both respiration and N leaching, under all atmospheric conditions. Thus, insects may affect carbon and nitrogen mineralization in forests of the future more through their effects on the quantity (and timing), than quality, of substrate deposition.
Indeed, atmospheric CO2 and O3 concentrations appear to influence rates of insect-mediated organic substrate deposition in forests. At Aspen FACE, rates of combined frass and greenfall deposition were 35% higher in elevated vs. ambient CO2 stands, and 13% lower in elevated vs. ambient O3 stands, over three field seasons (Meehan, Couture, Bennett and Lindroth, unpublished data).
Finally, insect feeding and atmospheric composition both may alter the timing of litterfall in forest ecosystems. In general, feeding damage accelerates leaf abscission, whereas enriched CO2 may either accelerate (Stiling et al. 2002) or decelerate (Karnosky et al. 2003) abscission rates. How herbivory and atmospheric chemistry may interact to influence the magnitude and timing of leaf abscission is unknown.
Soil Invertebrates
Soil invertebrates play important roles in the nutrient cycling dynamics of forest ecosystems, from the comminution and microbial inoculation of plant litter to the vertical transport of organic matter through soil horizons. These processes are governed both by the quality and quantity of litterfall, which in turn are influenced by atmospheric composition (Fig. 1).
Several studies have evaluated the effects of leaf litter from elevated CO2 plots on feeding preferences of isopods. Using short-term (3 day) feeding choice studies, Cotrufo et al. (1998b) found that isopods (Oniscus asellus) consumed 16% less high-CO2 litter than ambient-CO2 litter from Fraxinus excelsior. Subsequently, Cotrufo et al. (2005a) fed isopods (Porcellio sp.) for 2 weeks on litter from six tree species grown under ambient and elevated CO2, and found no difference in palatability for five species, and increased preference for high-CO2 litter for one species. Using no-choice feeding studies and two species of isopods, Hättenschwiler et al. (1999) found increased consumption of high-CO2 beech litter, but no effect of CO2 on consumption of spruce litter. Hättenschwiler and Bretscher (2001) reported that growth of trees under elevated CO2 altered the relative preference of O. asellus for different litter species—increasing preference for one, decreasing preference for another, and not affecting yet another. Thus, as is the case for insects feeding on green leaves, the consequences of CO2 for detritus-feeding invertebrates are species-specific.
Additional studies have assessed the effects of leaf litter from elevated CO2 or O3 environments on fitness indices and population growth of soil invertebrates. Individual growth of juvenile earthworms (Lumbricus terrestris) was reduced when fed high-CO2 or high-O3 birch (Betula pendula) litter (Kasurinen et al. 2007), and high-CO2 (but not high-O3) aspen (P. tremuloides) litter (Meehan, Crossley and Lindroth, unpublished data). Growth and mortality rates of isopods, however, generally have been shown not to be affected by CO2 or O3 treatments (Hättenschwiler and Bretscher 2001; Kasurinen et al. 2007). Population growth of collembola (Sinella curviseta) decreased when reared on high-CO2 aspen litter, but increased when reared on high-O3 aspen litter (Meehan, Crossley and Lindroth, unpublished data).
Several studies have evaluated the effects of CO2 and O3 on soil invertebrate communities under field conditions. Hansen et al. (2001) investigated the effects of elevated CO2 concentrations on soil microarthropods at the FACTS-1 FACE site. Over the first 19 months of CO2 treatment, total microathropod abundance declined by 34% in high-CO2 plots, relative to ambient plots. The decline was driven by changes in abundance of oribatid mites, and was attributed to factors other then changes in litter quality. Haimi et al. (2005) censused populations of enchytraeids, mites and collembola in soil samples surrounding chambered Pinus sylvestris trees. Elevated CO2 concentrations contributed to low numbers of acaridid mites, but did not significantly affect other soil fauna. Loranger et al. (2004) assessed responses of soil invertebrates to CO2 and O3 enrichment at Aspen FACE. They found that numbers of collembola and Oribatid mites, but not various other soil animals, declined in elevated CO2 plots. Mites, but not other fauna, also decreased in high O3 plots.
In summary, CO2- and O3-mediated changes in litter quality are likely to influence the performance of soil invertebrates of forests in the future. Some evidence exists to suggest that reduced litter quality will negatively affect the fitness of soil animals. In turn, their roles in nutrient cycling and soil carbon sequestration are likely to be altered. Overall, however, as noted by Couteaûx and Bolger (2000), no general patterns of response have yet emerged, likely due to the species- and context-specific nature of these interactions.
Litter Decomposition
Rates of litter decomposition and attendant nutrient mineralization are governed primarily by climate, litter chemistry, and soil organisms. Climate (especially temperature and moisture) is the most important factor on a global scale, whereas chemical composition is most influential within a geographic region (Aerts 1997). All of these regulating factors are influenced directly, or indirectly, by CO2 and O3 (Fig. 1).
Litter quality is determined by carbon availability (which ranges from labile sugars to recalcitrant lignin), mineral content, and chemical modifiers such as tannins (Swift et al. 1979; Anderson 1991). Tannins and related phenolic constituents alter decomposition by complexing with other litter components (e.g., amino acids) and/or by inhibiting the degradative activity of enzymes and soil fauna (Horner et al. 1988; Anderson 1991; Coûteaux et al. 1995). For similar litter types in common environments, nitrogen, lignin, and tannin concentrations appear to act as “a hierarchical series of controls” (Anderson 1991). In high quality, readily decomposable litters, tannins may be rate retardants, whereas in low quality, slowly decomposable litters, lignin concentration, or lignin: N ratio typically determine decomposition rates (Anderson 1991). Any factor that markedly affects the nitrogen or C-based secondary metabolite concentration of litter also will affect decomposition, and consequently, nutrient availability (Chapin 1991).
The “litter quality” hypothesis, introduced by Strain and Bazzaz (1983), proposed that CO2 enrichment will decrease plant nitrogen (relative to carbon) concentrations, and thereby reduce litter decomposition and soil N availability. Progressive soil nitrogen limitation is anticipated to eventually feed back to reduce plant production (Luo et al. 2004; Johnson 2006). Thus, CO2-mediated changes in the chemical quality of litter have been of key interest with respect to understanding the cycling and storage of carbon in forest ecosystems. Indeed, litter from trees grown under elevated CO2 characteristically exhibits reduced N concentrations and increased lignin concentrations and C:N ratios (Ceulemans et al. 1999; Norby et al. 2001).
To date, however, studies of the effects of elevated CO2 on litter decomposition (measured as mass loss or microbial respiration) have not demonstrated a consistent direction of response. A comprehensive meta-analysis by Norby et al. (2001), including data for 33 woody species, showed that data do not support the “litter quality” hypothesis of reduced decomposition rates for litter produced under elevated CO2. Decomposition studies published since that review have been increasingly realistic, employing more long-term and field-based approaches with open-top chambers or FACE sites. Still, a consistent pattern has yet to emerge. Several studies have documented reduced decomposition of leaf litter produced under elevated CO2. For example, litter from three Populus species grown under elevated CO2 at POPFACE exhibited reduced decomposition rates (Cotrufo et al. 2005b), as did litter from Betula papyrifera and Populus tremuloides from young (sapling) forest stands grown under elevated CO2 at Aspen FACE (Parsons et al. 2004, 2008). In contrast, CO2 growth environment did not alter subsequent litter decomposition rates for Quercus myrtifolia in a scrub oak community (Hall et al. 2006), for five tree species at the FACTS-1 FACE site (Finzi et al. 2001), or for B papyrifera and P. tremuloides in older, closed-canopy stands at Aspen FACE (Liu et al. 2009). Diverse responses among experiments have been attributed to a number of factors, including soil fertility, decomposition environment, CO2 exposure mechanism, study duration, stand development, and, genotype- and species-specific effects (Norby et al. 2001; Kasurinen et al. 2006; Hall et al. 2006; Liu et al. 2009).
Relatively little research has investigated the effects of atmospheric O3 on litter quality and decomposition. Growth of three species of deciduous tree seedlings (Boerner and Rebbeck 1995; Scherzer et al. 1998) and two species of Pinus seedlings and saplings (Scherzer et al. 1998; Kainulainen et al. 2003) under elevated O3 did not influence subsequent decomposition rates of leaf litter. Findlay et al. (1996), however, found that cottonwood leaves produced under high O3 levels exhibited reduced decomposition rates (in aquatic microcosms), which were linked to increased phenolic concentrations. Similarly, Kasurinen et al. (2006) observed reduced decomposition in Betula pendula leaves produced under high O3 concentrations. Parsons et al. (2008) found that early in stand development at Aspen FACE, growth under elevated O3 reduced 2-year decomposition rates of Populus tremuloides litter, independent of CO2 treatment. In contrast, elevated O3 accelerated decomposition of Betula papyrifera litter from ambient CO2 treatments, but decreased decomposition of litter from enriched CO2 treatments (a significant O3 × CO2 interaction). Later work at Aspen FACE, however, showed that elevated O3 decreased the decomposition of P. tremuloides and B. papyrifera litter from closed-canopy stands during only the first year of a 2.6-year study (Liu et al. 2009). As is the case for atmospheric CO2, the effects of elevated O3 on litter decomposition appear to be species-, age-, and context-specific.
Although too early for definitive conclusions, the developing scientific consensus is that atmospheric CO2 and O3 will affect litter decomposition and subsequent nutrient cycling more through their effects on the quantity, than quality, of foliage produced, and by shifting the species composition of forests (Finzi and Schlesinger 2002; Cotrufo et al. 2005b; Liu et al. 2007, 2009; Hillstrom, Meehan, Kelly and Lindroth, unpublished data). For example, soil carbon sequestration at Aspen FACE appears to be more closely related to CO2- and O3-mediated changes in input quantity and species composition than to qualitative changes in litter chemical composition (Loya et al. 2003; Liu et al. 2007, 2009).
Conclusions and Recommendations for Future Research
Human-induced changes in the levels of atmospheric CO2 and O3 clearly influence the quantity and quality of primary production. In turn, changes in plant chemical composition cascade through above- and below-ground ecological interactions (e.g., herbivory, decomposition) to alter ecosystem structure and function, ultimately feeding back to affect terrestrial carbon sequestration and atmospheric composition. Many of these interactions are governed by processes that are fundamentally chemical in nature (double entendre intended). Thus, chemical ecologists have much to offer in terms of elucidating the mechanisms that integrate biological organization—from genomes to ecosystems—in a globally changing world.
An admittedly frustrating theme to emerge from this and other recent reviews (e.g., Stiling and Cornelissen 2007; Bidart-Bouzat and Imeh-Nathaniel 2008; Tylianakis et al. 2008) of global environmental change is that the effects of any single change factor on any particular ecological interaction are highly variable. Responses tend to be species- and context-specific, and thus appear idiosyncratic. Global change science must advance considerably further before predictions about impacts on natural systems can be made with accuracy and precision. The work of chemical ecologists should be central to such efforts: it will provide mechanistic insight into ecosystem function and provide data critical to the accurate parameterization of global change and risk assessment models.
Below I provide a non-exhaustive list of some of the more pressing needs for the contributions of chemical ecologists toward furthering our understanding of the effects of CO2 and O3 on forest ecosystems. Funding of large, long-term, multi-investigator projects is essential to the success of these efforts.
-
1.
Increase the diversity of natural products studied. Our understanding of the effects of global change on the phytochemistry of forest ecosystems is based on a few classes of chemical compounds, primarily phenolics and terpenoids. Almost nothing is known about effects on other classes of secondary metabolites important to at least some tree species, e.g., alkaloids, cyanogenic glycosides, nonprotein amino acids, coumarins, and iridoid glycosides.
-
2.
Broaden the representation of species and biomes investigated. To date, research has focused on a narrow range of deciduous and coniferous tree species and biomes, and mostly lepidopteran herbivores. Both tropical and boreal forests are critically important to the global carbon cycle, and poorly represented in global change research. Future studies should emphasize foundation tree species (Ellison et al. 2005) and functional groups (e.g., insect borers, miners) and taxa (e.g., mammals) known to be important to the health and sustainability of forest ecosystems. The coniferous forests and irruptive bark beetles of western North America are a prime example (Raffa et al. 2008). In a similar vein, future research should address non-exploitative ecological interactions, such as mutualistic associations of plants with pollinators and mycorrhizae.
-
3.
Emphasize simultaneous investigation of multiple global change drivers. As documented in this review and others (e.g., Tylianakis et al. 2008), the effects of any particular global change factor on ecological interactions are context-dependent. Multiple drivers should be investigated simultaneously to more realistically approximate future environments and to provide a diversity of conditions under which to test the effects of individual drivers (Tylianakis et al. 2008). Altered temperature and precipitation are arguably the most important additional factors to evaluate in the context of CO2 and O3 effects on forests.
-
4.
Conduct long-term experiments. The majority of studies published to date have been executed over short time frames (up to several years), with only a few approaching 10 years—considerably less than the lifespan of forest trees. Some ecosystem processes (e.g., soil nutrient depletion and carbon storage) require assessment over the course of a decade or longer, logistical and funding constraints nothwithstanding. Long-term experiments also could allow for studies of evolutionary responses of insects to CO2- and O3-mediated changes in tree chemistry, which are conspicuously absent in the published literature and important to understanding the effects of global change on forest ecosystems.
-
5.
Employ “genes to ecosystems” research perspectives. “Genes to ecosystems” research has emerged as a new disciplinary frontier in genetics and ecology (Whitham et al. 2006). It combines genetic/genomic tools with the approaches of evolutionary ecology to understand the functional significance of genes in ecological context and the dynamics of ecosystems in evolutionary context. The discipline of chemical ecology can play a key role in elucidating the “metabolomes” that link gene expression to ecosystem processes in the context of global environmental change.
References
Aerts, R. 1997. Climate, leaf litter chemistry and leaf litter decomposition in terrestrial ecosystems: a triangular relationship. Oikos 79:439–449.
Agrell, J., Mcdonald, E. P., and Lindroth, R. L. 1999. Responses to defoliation in deciduous trees: effects of CO2 and light. Ecol. Bull. 47:84–95.
Agrell, J., Mcdonald, E. P., and Lindroth, R. L. 2000. Effects of CO2 and light on tree phytochemistry and insect performance. Oikos 88:259–272.
Agrell, J., Kopper, B., Mcdonald, E. P., and Lindroth, R. L. 2005. CO2 and O3 effects on host plant preferences of the forest tent caterpillar (Malacosoma disstria). Glob. Chang Biol. 11:588–599.
Ainsworth, E. A., and Long, S. P. 2005. What have we learned from 15 years of free-air CO2 enrichment (FACE)? A meta-analytic review of the responses of photosynthesis, canopy properties and plant production to rising CO2. New Phytol. 165:351–371.
Aldea, M., Hamilton, J. G., Resti, J. P., Zangerl, A. R., Berenbaum, M. R., Frank, T. D., and Delucia, E. H. 2006. Comparison of photosynthetic damage from arthropod herbivory and pathogen infection in understory hardwood saplings. Oecologia 149:221–232.
Andersen, C. P. 2003. Source-sink balance and carbon allocation below ground in plants exposed to ozone. New Phytol. 157:213–228.
Anderson, J. M. 1991. The effects of climate change on decomposition processes in grassland and coniferous forests. Ecol. Appl. 1:326–347.
Ashmore, M. R. 2005. Assessing the future global impacts of ozone on vegetation. Plant Cell Environ. 28:949–964.
Asshoff, R., Zotz, G., and Körner, C. 2006. Growth and phenology of mature temperate forest trees in elevated CO2. Glob. Chang. Biol. 12: 848–861.
Awmack, C. S., Harrington, R., and Lindroth, R. L. 2004. Aphid individual performance may not predict population responses to elevated CO2 or O3. Glob. Chang. Biol. 10:1414–1423.
Bale, J. S., Masters, G. J., Hodkinson, I. D., Awmack, C., Bezemer, T. M., Brown, V. K., Butterfield, J., Buse, A., Coulson, J. C., Farrar, J., Good, J. E. G., Harrington, R., Hartley, S., Jones, T. H., Lindroth, R. L., Press, M. C., Symrnioudis, I., Watt, A. D., and Whittaker, J. B. 2002. Herbivory in global climate change research: direct effects of rising temperature on insect herbivores. Global Change Biol. 8:1–16.
Beedlow, P. A., Tingey, D. T., Phillips, D. L., Hogsett, W. E., and Olszyk, D. M. 2004. Rising atmospheric CO2 and carbon sequestration in forests. Front. Ecol. Environ. 2:315–322.
Berggren, A., Björkman, C., Bylund, H., and Ayres, M. P. 2009. The distribution and abundance of animal populations in a climate of uncertainty. Oikos 118:1121–1126.
Berner, R. A. 2005. The rise of trees and how they changed the Paleozoic atmosphere CO2, climate, and geology, pp. 1–7, in Ehleringer, J. R., Cerling, T. E., Dearing, M. D., (eds.). A History of Atmospheric CO2 and Its Effects on Plants, Animals, and Ecosystems. Springer, New York.
Betz, G. A., Gerstner, E., Stich, S., Winkler, B., Welzl, G., Kremmer, E., Langebartels, C., Heller, W., Sandermann, H., and Ernst, D. 2009a. Ozone affects shikimate pathway genes and secondary metabolites in saplings of European beech (Fagus sylvatica L.) grown under greenhouse conditions. Trees—Struct. Funct. 23:539–553.
Betz, G. A., Knappe, C., Lapierre, C., Olbrich, M., Welzl, G., Langebartels, C., Heller, W., Sandermann, H., and Ernst, D. 2009b. Ozone affects shikimate pathway transcripts and monomeric lignin composition in European beech (Fagus sylvatica L.). Eur. J. Forest Res. 128:109–116.
Bezemer, T. M., and Jones, T. H. 1998. Plant-insect herbivore interactions in elevated atmospheric CO2: quantitative analyses and guild effects. Oikos 82:212–222.
Bidart-Bouzat, M. G., and Imeh-Nathaniel, A. 2008. Global change effects on plant chemical defenses against insect herbivores. Journal of Integrative Plant Biology 50:1339–1354.
Blande, J. D., Tiiva, P., Oksanen, E., and Holopainen, J. K. 2007. Emission of herbivore-induced volatile terpenoids from two hybrid aspen (Populus tremula x tremuloides) clones under ambient and elevated ozone concentrations in the field. Glob. Chang. Biol. 13:2538–2550.
Boege, K., and Marquis, R. J. 2005. Facing herbivory as you grow up: the ontogeny of resistance in plants. Trends Ecol. Evol. 20:441–448.
Boerner, R. E. J., and Rebbeck, J. 1995. Decomposition and nitrogen release from leaves of three hardwood species grown under elevated O3 and/or CO2. Plant Soil 170:149–157.
Bonan, G. B. 2008. Forests and climate change: forcings, feedbacks, and the climate benefits of forests. Science 320:1444–1449.
Booker, F. L., and Maier, C. A. 2001. Atmospheric carbon dioxide, irrigation, and fertilization effects on phenolic and nitrogen concentrations in loblolly pine (Pinus taeda) needles. Tree Physiol. 21:609–616.
Booker, F. L., Anttonen, S., and Heagle, A. S. 1996. Catechin, proanthocyanidin and lignin contents of loblolly pine (Pinus taeda) needles after chronic exposure to ozone. New Phytol. 132:483–492.
Bryant, J. P., and Julkunen-Tiitto, R. 1995. Ontogenic development of chemical defense by seedling resin birch: energy cost of defense production. J. Chem. Ecol. 21:883–896.
Bryant, J. P., Chapin, F. S. III, and Klein, D. R. 1983. Carbon/nutrient balance of boreal plants in relation to vertebrate herbivory. Oikos 40:357–368.
Cebrian, J. 1999. Patterns in the fate of production in plant communities. Am. Nat. 154:449–468.
Cebrian, J., and Lartigue, J. 2004. Patterns of herbivory and decomposition in aquatic and terrestrial ecosystems. Ecol. Monogr. 74:237–259.
Ceulemans, R., Janssens, I. A., and Jach, M. E. 1999. Effects of CO2 enrichment on trees and forests: lessons to be learned in view of future ecosystem studies. Ann. Bot. 84:577–590.
Chameides, W. L., Kasibhatla, P. S., Yienger, J., and Levy II, H. 1994. Growth of continental-scale metro-agro-plexes, regional ozone pollution, and world food production. Science 264:74–77.
Chapin, F. S. III. 1991. Effects of multiple environmental stresses on nutrient availability and use, pp. 67–88, in H. A. Mooney, W. E. Winner, and E. J. Pell (eds.). Response of Plants to Multiple Stresses. Academic Press, Inc., New York, NY.
Cole, C. T., Anderson, J. E., Lindroth, R. L., and Waller, D. M. 2009. Rising concentrations of atmospheric CO2 have increased growth in natural stands of quaking aspen (Populus tremuloides). Glob. Chang. Biol. doi:10.1111/j.1365-2486.2009.02103.x.
Coley, P. D., Massa, M., Lovelock, C. E., and Winter, K. 2002. Effects of elevated CO2 on foliar chemistry of saplings of nine species of tropical tree. Oecologia 133:62–69.
Cotrufo, M. F., Ineson, P., and Scott, A. 1998a. Elevated CO2 reduces the nitrogen concentration of plant tissues. Glob. Chang. Biol. 4:43–54.
Cotrufo, M. F., Briones, M. J., and Ineson, P. 1998b. Elevated CO2 affects field decomposition rate and palatability of tree leaf litter: importance of changes in substrate quality. Soil Biol. Biochem. 36:1565–1571.
Cotrufo, M. F., Drake, B., and Ehleringer, J. R. 2005a. Palatability trials on hardwood leaf litter grown under elevated CO2: a stable carbon isotope study. Soil Biol. Biochem. 37:1105–1112.
Cotrufo, M. F., De Angelis, P., and Polle, A. 2005b. Leaf litter production and decomposition in a poplar short-rotation coppice exposed to free air CO2 enrichment (POPFACE). Glob. Chang. Biol. 11:971–982.
Couteaûx, M.-M., and Bolger, T. 2000. Interactions between atmospheric CO2 enrichment and soil fauna. Plant Soil 224:123–134.
Coûteaux, M.-M., Bottner, P., and Berg, B. 1995. Litter decomposition, climate and litter quality. Trends Ecol. Evol. 10:63–66.
Coûteaux, M. M., Kurz, C., Bottner, P., and Raschi, A. 1999. Influence of increased atmospheric CO2 concentration on quality of plant material and litter decomposition. Tree Physiol. 19:301–311.
Coviella, C., and Trumble, J. T. 1999. Effects of elevated atmospheric CO2 on insect-plant interactions. Conserv. Biol. 13:700–712.
Cseke, L. J., Tsai, C. J., Rogers, A., Nelsen, M. P., White, H. L., Karnosky, D. F., and Podila, G. K. 2009. Transcriptomic comparison in the leaves of two aspen genotypes having similar carbon assimilation rates but different partitioning patterns under elevated [CO2]. New Phytol. 182:891–911.
Cyr, H., and Pace, M. L. 1993. Magnitude and patterns of herbivory in aquatic and terrestrial ecosystems. Nature 361:148–150.
Denman, K. L., Brasseur, G., Chidthaisong, A., Ciais, P., Cox, P. M., Dickinson, R. E., Hauglustaine, D., Heinze, C., Holland, E., Jacob, D., Lohmann, U., Ramachandran, S., da Silva Dias, P. L., Wofsy S. C., and Zhang, X. 2007. Couplings between changes in the climate system and biogeochemistry, in S. Solomon, D. Qin, M. Manning, Z. Chen, M. Marquis, K. B. Averyt, M. Tignor and H. L. Miller (eds.). Climate change 2007: the physical science basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press, New York.
Docherty, M., Wade, F. A., Hurst, D. K., Whittaker, J. B., and Lea, P. J. 1997. Responses of tree sap-feeding herbivores to elevated CO2. Glob. Chang. Biol. 3:51–59.
Donaldson, J. R., and Lindroth, R. L. 2008. Effects of variable phytochemistry and budbreak phenology on defoliation of aspen during a forest tent caterpillar outbreak. Agric. For. Entomol. 10:399–410.
Donaldson, J. R., Stevens, M. T., Barnhill, H. R., and Lindroth, R. L. 2006. Age-related shifts in leaf chemistry of clonal aspen (Populus tremuloides). J. Chem. Ecol. 32:1415–1429.
Dukes, J. S., Pontius, J., Orwig, D., Garnas, J. R., Rodgers, V. L., Brazee, N., Cooke, B., Theoharides, K. A., Stange, E. E., Harrington, R., Ehrenfeld, J., Gurevitch, J., Lerdau, M., Stinson, K., Wick, R., and Ayres, M. 2009. Responses of insect pests, pathogens, and invasive plant species to climate change in the forests of northeastern North America: what can we predict? Can. J. For. Res. 39:231–248.
Ellison, A. M., Bank, M. S., Clinton, B. D., Colburn, E. A., Elliott, K., Ford, C. R., Foster, D. R., Kloeppel, B. D., Knoepp, J. D., Lovett, G. M., Mohan, J., Orwig, D. A., Rodenhouse, N. L., Sobczak, W. V., Stinson, K. A., Stone, J. K., Swan, C. M., Thompson, J., Von Holle, B., and Webster. J. R. 2005. Loss of foundation species: consequences for the structure and dynamics of forested ecosystems. Front. Ecol. Environ. 3:479–486.
Felzer, B. D., Kicklighter, D., Melillo, J., Wang, C., ZHUANG, Q., and PRINN, R. 2004. Effects of ozone on net primary production and carbon sequestration in the conterminous United States using a biogeochemistry model. Tellus 56B:230–248.
Findlay, S., Carreiro, M., Krischik, V., and Jones, C. G. 1996. Effects of damage to living plants on leaf litter quality. Ecol. Appl. 6:269–275.
Finzi, A. C., and Schlesinger, A. H. 2002. Species control variation in litter decomposition in a pine forest exposed to elevated CO2. Glob. Chang. Biol. 8:1217–1229.
Finzi, A. C., Allen, A. S., Delucia, E. H., Ellsworth, D. S., and Schlesinger, W. H. 2001. Forest litter production, chemistry, and decomposition following 2 years of free-air CO2 enrichment. Ecology 82:470–484.
Foster, M. A., Schultz, J. C., and Hunter, M. D. 1992. Modelling gypsy moth-virus-leaf chemistry interactions: implications of plant quality for pest and pathogen dynamics. J. Anim. Ecol. 61:509–520.
Fowler, D., Cape, J. N., Coyle, M., Flechard, C., Kuylenstierna, J., Hicks, K., Derwent, D., Johnson, C., and Stevenson, D. 1999. The global exposure of forests to air pollutants. Water Air Soil Pollut. 116:5–32.
Freiwald, V., Haikio, E., Julkunen-Tiitto, R., Holopainen, J. K., and Oksanen, E. 2008. Elevated ozone modifies the feeding behaviour of the common leaf weevil on hybrid aspen through shifts in developmental, chemical, and structural properties of leaves. Ent. Exp. Appl. 128:66–72.
Fuhrer, J., and Booker, F. 2003. Ecological issues related to ozone: agricultural issues. Environ. Int. 29:141–154.
Gate, I. M., Mcneill, S., and Ashmore, M. R. 1995. Effects of air pollution on the searching behavior of an insect parasitoid. Water Air Soil Pollut. 85:1425–1430.
Haimi, J., Laamanen, J., Penttinen, R., Raty, M., Koponen, S., Kellomaki, S., and Niemela, P. 2005. Impacts of elevated CO2 and temperature on the soil fauna of boreal forests. Appl. Soil Ecol. 30:104–112.
Hall, M. C., Stiling, P., Hungate, B. A., Drake, B. G., and Hunter, M. D. 2005. Effects of elevated CO2 and herbivore damage on litter quality in a scrub oak ecosystem. J. Chem. Ecol. 31:2343–2356.
Hall, M. C., Stiling, P., Moon, D. C., Drake, B. G., and Hunter, M. D. 2006. Elevated CO2 increases the long-term decomposition rate of Quercus myrtifolia leaf litter. Glob. Chang. Biol. 12:568–577.
Hamilton, J. G., Zangerl, A. R., Berenbaum, M. R., Pippen, J., Aldea, M., and Delucia, E. H. 2004. Insect herbivory in an intact forest understory under experimental CO2 enrichment. Oecologia 138:566–573.
Hansen, R. A., Williams, R. S., Degenhardt, D. C., and Lincoln, D. E. 2001. Non-litter effects of elevated CO2 on forest floor microarthropod abundances. Plant Soil 236:139–144.
Hari, P., and Kulmala, L. (eds.). 2008. Boreal Forest and Climate Change. Advances in Global Change Research, Vol. 34. Springer, New York.
Hassan, R., Scholes, R., and Ash, N. (eds.). 2005. Ecosystems and Human Well-Being: Current State and Trends, Vol. 1. Island, Washington
Hättenschwiler, S., and Bretscher, D. 2001. Isopod effects on decomposition of litter produced under elevated CO2, N deposition and different soil types. Glob. Chang. Biol. 7:565–579.
Hättenschwiler, S., and Schafellner, C. 1999. Opposing effects of elevated CO2 and N deposition on Lymantria monacha larvae feeding on spruce trees. Oecologia 118:210–217.
Hättenschwiler, S., Bühler, S., and Körner, C. 1999. Quality, decomposition and isopod consumption of tree litter produced under elevated CO2. Oikos 85:271–281.
Heath, R. L. 2008. Modification of the biochemical pathways of plants induced by ozone: what are the varied routes to change? Environ. Pollut. 155:453–463.
Herms, D. A., and Mattson, W. J. 1992. The dilemma of plants: to grow or defend. Q. Rev. Biol. 67:283–335.
Hillstrom, M.L., and Lindroth, R. L. 2008. Elevated atmospheric carbon dioxide and ozone alter forest insect abundance and community composition. Insect Conserv. Diversity 1:233–241.
Holton, M. K., Lindroth, R. L., and Nordheim, E. V. 2003. Foliar quality influences tree-herbivore-parasitoid interactions: effects of elevated CO2, O3, and plant genotype. Oecologia 137:233–244.
Horner, J. D., Gosz, J. R., and Cates, R. G. 1988. The role of carbon-based plant secondary metabolites in decomposition in terrestrial ecosystems. Am. Nat. 132:869–883.
Huang, J. G., Bergeron, Y., Denneler, B., Berninger, F., and Tardif, J. 2007. Response of forest trees to increased atmospheric CO2. Crit. Rev. Plant Sci. 26:265–283.
Huttunen, L., Niemelä, P., Julkunen-Tiitto, R., Heiska, S., Tegelberg, R., Rousi, M., and Kellomaki, S. 2008. Does defoliation induce chemical and morphological defenses in the leaves of silver birch seedlings under changing climate? Chemoecology 18:85–98.
Johnson, D. W. 2006. Progressive N limitation in forests: review and implications for long-term responses to elevated CO2. Ecology 87:64–75.
Jones, C. G., and Coleman, J. S. 1988. Plant stress and insect behavior: cottonwood, ozone and the feeding and oviposition preference of a beetle. Oecologia 76:51–56.
Julkunen-Tiitto, R., Tahvanainen, J., and Silvola, J. 1993. Increased CO2 and nutrient status changes affect phytomass and the production of plant defensive secondary chemicals in Salix myrsinifolia (Salisb.). Oecologia 95:495–498.
Kainulainen, P., Holopainen, T., and Holopainen, J. K. 2003. Decomposition of secondary compounds from needle litter of scots pine grown under elevated CO2 and O3. Glob. Chang. Biol. 9:295–304.
Kangasjärvi, J., Talvinen, J., Utriainen, M., and Karjalainen, R. 1994. Plant defence systems induced by ozone. Plant Cell Environ. 17:783–794.
Kangasjärvi, J., Jaspers, P., and Kollist, H. 2005. Signalling and cell death in ozone-exposed plants. Plant Cell Environ. 28:1021–1036.
Karl, T. R., Melillo, J. M., Peterson, T. C. (eds.). 2009. Global Climate Change Impacts in the United States. Cambridge University Press, New York.
Karnosky, D. F., Percy, K. E., Xiang, B. X., Callan, B., Noormets, A., Mankovska, B., Hopkin, A., Sober, J., Jones, W., Dickson, R. E., and Isebrands, J. G. 2002. Interacting elevated CO2 and tropospheric O3 predisposes aspen (Populus tremuloides Michx.) to infection by rust (Melampsora medusae f. sp. tremuloidae). Glob. Chang. Biol. 8:329-338.
Karnosky, D. F., Zak, D. R., Pregitzer, K. S., Awmack, C. S., Bockheim, J. G., Dickson, R. E., Hendrey, G. R., Host, G. E., King, J. S., Kopper, B. J., Kruger, E. L., Kubiske, M. E., Lindroth, R. L., Mattson, W. J., Mcdonald, E. P., Noormets, A., Oksanen, E., Parsons, W. F. J., Percy, K. E., Podila, G. K., Riemenschneider, D. E., Sharma, P., Thakur, R., Sober, A., Sober, J. et al. 2003. Tropospheric O3 moderates responses of temperate hardwood forests to elevated CO2: a synthesis of molecular to ecosystem results from the Aspen FACE project. Funct. Ecol. 17:289-304.
Karnosky, D. F., Pregitzer, K. S., Zak, D. R., Kubiske, M. E., Hendrey, G. R., Weinstein, D., Nosal, M., and Percy, K. E. 2005. Scaling ozone responses of forest trees to the ecosystem level in a changing climate. Plant Cell Environ. 28:965-981.
Karnosky, D. F., Skelly, J. M., Percy, K. E., and Chappelka, A. H. 2007. Perspectives regarding 50 years of research on effects of tropospheric ozone air pollution on US forests. Environ. Pollut. 147:489–506.
Karonen, M., Ossipov, V., Ossipova, S., Kapari, L., Loponen, J., Matsumura, H., Kohno, Y., Mikami, C., Sakai, Y., Izuta, T., and Pihlaja, K. 2006. Effects of elevated carbon dioxide and ozone on foliar proanthocyanidins in Betula platyphylla, Betula ermanii, and Fagus crenata seedlings. J. Chem. Ecol. 32:1445–1458.
Kasurinen, A., Riikonen, J., Oksanen, E., Vapaavuori, E., and Holopainen, T. 2006. Chemical composition and decomposition of silver birch leaf litter produced under elevated CO2 and O3. Plant Soil 282:261–280.
Kasurinen, A., Peltonen, P. A., Julkunen-Tiitto, R., Vapaavuori, E., Nuutinen, V., Holopainen, T., and Holopainen, J. K. 2007. Effects of elevated CO2 and O3 on leaf litter phenolics and subsequent performance of litter-feeding soil macrofauna. Plant Soil 292:25–43.
Kinney, K. K., Lindroth, R. L., Jung, S. M., and Nordheim, E. V. 1997. Effects of CO2 and NO −3 availability on deciduous trees: phytochemistry and insect performance. Ecology 78:215–230.
Knepp, R. G., Hamilton, J. G., Mohan, J. E., Zangerl, A. R., Berenbaum, M. R., and Delucia, E. H. 2005. Elevated CO2 reduces leaf damage by insect herbivores in a forest community. New Phytol. 167:207–218.
Knepp, R. G., Hamilton, J. G., Zangerl, A. R., Berenbaum, M. R., and Delucia, E. H. 2007. Foliage of oaks grown under elevated CO2 reduces performance of Antheraea polyphemus (Lepidoptera: Saturniidae). Environ. Entomol. 36:609–617.
Koike, T., Tobita, H., Shibata, T., Matsuki, S., Konno, K., Kitao, M., Yamashita, N., and Maruyama, Y. 2006. Defense characteristics of seral deciduous broad-leaved tree seedlings grown under differing levels of CO2 and nitrogen. Popul. Ecol. 48:23-29.
Kontunen-Soppela, S., Ossipov, V., Ossipova, S., and Oksanen, E. 2007. Shift in birch leaf metabolome and carbon allocation during long-term open-field ozone exposure. Glob. Chang. Biol. 13:1053–1067.
Kopper, B. J., and Lindroth, R. L. 2003a. Responses of trembling aspen (Populus tremuloides) phytochemistry and aspen blotch leafminer (Phyllonorycter tremuloidiella) performance to elevated level of CO2 and O3. Agric. For. Entomol. 5:17–26.
Kopper, B. J., and Lindroth, R. L. 2003b. Effects of elevated carbon dioxide and ozone on the phytochemistry of aspen and performance of an herbivore. Oecologia 134:95–103.
Kopper, B. J., Lindroth, R. L., and Nordheim, E. V. 2001. CO2 and O3 effects on paper birch (Betulaceae: Betula papyrifera) phytochemistry and whitemarked tussock moth (Lymantriidae: Orgyia leucostigma) performance. Environ. Entomol. 30:1119–1126.
Koricheva, J., Larsson, S., Haukioja, E., and Keinänen, M. 1998a. Regulation of woody plant secondary metabolism by resource availability: hypothesis testing by means of meta-analysis. Oikos 83:212–226.
Koricheva, J., Larsson, S., and Haukioja, E. 1998b. Insect performance on experimentally stressed woody plants: a meta-analysis. Annu. Rev. Entomol. 43:195–216.
Körner, C. 2006. Plant CO2 responses: an issue of definition, time and resource supply. New Phytol. 172:393-411.
Körner, C., Asshoff, R., Bignucolo, O., Hattenschwiler, S., Keel, S. G., Pelaez-Riedl, S., Pepin, S., Siegwolf, R. T. W., and Zotz, G. 2005. Carbon flux and growth in mature deciduous forest trees exposed to elevated CO2. Science 309:1360–1362.
Kuokkanen, K., Yan, S. C., and Niemela, P. 2003. Effects of elevated CO2 and temperature on the leaf chemistry of birch Betula pendula (Roth) and the feeding behaviour of the weevil Phyllobius maculicornis. Agric. For. Entomol. 5:209–217.
Kuokkanen, K., Niemela, P., Matala, J., Julkunen-Tiitto, R., Heinonen, J., Rousi, M., Henttonen, H., Tahvanainen, J., and Kellomaki, S. 2004. The effects of elevated CO2 and temperature on the resistance of winter-dormant birch seedlings (Betula pendula) to hares and voles. Glob. Chang. Biol. 10:1504–1512.
Kurpius, M. R., and Goldstein, A. H. 2003. Gas-phase chemistry dominates O3 loss to a forest, implying a source of aerosols and hydroxyl radicals to the atmosphere. Geophys. Res. Lett. 30:1371.
Kurz, W. A., Stinson, G., Rampley, G. J., Dymond, C. C., and Neilson, E. T. 2008. Risk of natural disturbances makes future contribution of Canada’s forests to the global carbon cycle highly uncertain. Proc. Natl. Acad. Sci. USA 105:1551–1555.
Laothawornkitkul, J., Taylor, J. E., Paul, N. D., and Hewitt, C. N. 2009. Biogenic volatile organic compounds in the Earth system. New Phytol. 183:27–51.
Laurence, W. F., and Peres, C. A. 2006. Emerging Threats to Tropical Forests. University of Chicago Press, Chicago.
Lavola, A., and Julkunen-Tiitto, R. 1994. The effect of elevated carbon dioxide and fertilization on primary and secondary metabolites in birch, Betula pendula (Roth). Oecologia 99:315–321.
Lawler, I. R., Foley, W. J., Woodrow, I. E., and Cork, S. J. 1997. The effects of elevated CO2 on the nutritional quality of Eucalyptus foliage and its interaction with soil nutrient and light availability. Oecologia 109:59–68.
Leakey, A. D. B., Ainsworth, E. A., Bernacchi, C. J., Rogers, A., Long, S. P., and Ort, D. R. 2009. Elevated CO2 effects on plant carbon, nitrogen, and water relations: six important lessons from FACE. J. Exp. Bot. 60:2859–2876.
Lerdau, M., and Gray, D. 2003. Ecology and evolution of light-dependent and light-independent phytogenic volatile organic carbon. New Phytol. 157:199–211.
Lindroth, R. L., and Dearing, M. D. 2005. Herbivory in a world of elevated CO2, pp. 468–486, in J. R. Ehleringer, T. E. Cerling, and M. D. Dearing (eds.). A History of Atmospheric CO2 and Its Effect on Plants, Animals, and Ecosystems. Springer Science, Inc, New York.
Lindroth, R. L., and Kinney, K. K. 1998. Consequences of enriched atmospheric CO2 and defoliation for foliar chemistry and gypsy moth performance. J. Chem. Ecol. 24:1677–1695.
Lindroth, R. L., Reich, P. B., Tjoelker, M. G., Volin, J. C., and Oleksyn, J. 1993a. Light environment alters response to ozone stress in Acer saccharum Marsh. and hybrid Populus L. seedlings. III. Consequences for gypsy moth performance. New Phytol. 124:647–651.
Lindroth, R. L., Jung, S. M., and Feuker, A. M. 1993b. Detoxication activity in the gypsy moth: effects of host CO2 and NO −3 availability. J. Chem. Ecol. 19:357–367.
Lindroth, R. L., Roth, S., Kruger, E. L., Volin, J. C., and Koss, P. A. 1997. CO2-mediated changes in aspen chemistry: effects on gypsy moth performance and susceptibility to virus. Glob. Chang. Biol. 3:279–289.
Lindroth, R. L., Roth, S., and Nordheim, E. V. 2001. Genotypic variation in response of quaking aspen (Populus tremuloides) to atmospheric CO2 enrichment. Oecologia 126:371–379.
Lindroth, R. L., Wood, S. A., and Kopper, B. J. 2002. Response of quaking aspen genotypes to enriched CO2: foliar chemistry and tussock moth performance. Agric. For. Entomol. 4:315–323.
Litvak, M. E., Madronich, S., and Monson, R. K. 1999. Herbivore-induced monoterpene emissions from coniferous forests: potential impact on local tropospheric chemistry. Ecol. Appl. 9:1147–1159.
Liu, L. L., King, J. S., and Giardina, C. P. 2005. Effects of elevated concentrations of atmospheric CO2 and tropospheric O3 on leaf litter production and chemistry in trembling aspen and paper birch communities. Tree Physiol. 25:1511–1522.
Liu, L., King, J. S., and Giardina, C. P. 2007. Effects of elevated atmospheric CO2 and tropospheric O3 on nutrient dynamics: decomposition of leaf litter in trembling aspen and paper birch communities. Plant Soil 299:65–82.
Liu, L. L., King, J. S., Giardina, C. P., and Booker, F. L. 2009. The influence of chemistry, production and community composition on leaf litter decomposition under elevated atmospheric CO2 and tropospheric O3 in a northern hardwood ecosystem. Ecosystems 12:401–416.
Long, S. P., Ainsworth, E. A., Rogers, A., and Ort, D. R. 2004. Rising atmospheric carbon dioxide: plants FACE the future. Annu. Rev. Plant Biol. 55:591–628.
Loranger, G. I., Pregitzer, K. S., and King, J. S. 2004. Elevated CO2 and O3t concentrations differentially affect selected groups of the fauna in temperate forest soils. Soil Biol. Biochem. 36:1521–1524.
Loya, W. M., Pregitzer, K. S., Karberg, N. J., King, J. S., and Giardina, C. P. 2003. Reduction of soil carbon formation by tropospheric ozone under increased carbon dioxide levels. Nature 425:705–707.
Luo, Y., Su, B., Currie, W. S., Dukes, J. S., Finzi, A., Hartwig, U., Hungate, B., Mcmurtrie, R. E., Oren, R., Parton, W. J., Pataki, D. E., Shaw, M. R., Zak, D. R., and Field, C. B. 2004. Progressive nitrogen limitation of ecosystem responses to rising atmospheric carbon dioxide. Bioscience 54:731–739.
Mattson, W. J., Kuokkanen, K., Niemelä, P., Julkunen-Tiitto, R., Kellomäki, S., and Tahvanainen, J. 2004. Elevated CO2 alters birch resistance to Lagomorpha herbivores. Glob. Chang. Biol. 10:1402–1413.
Mcdonald, E. P., Agrell, J., and Lindroth, R. L. 1999. CO2 and light effects on deciduous trees: growth, foliar chemistry, and insect performance. Oecologia 119:389–399.
Mcguire, A. D., Melillo, J. M., and Joyce, L. A. 1995. The role of nitrogen in the response of forest net primary production to elevated atmospheric carbon dioxide. Annu. Rev. Ecol. Syst. 26:473–503.
Millenium Ecosystem Assessment. 2005. Ecosystems and Human Well-Being: Biodiversity Synthesis. World Resources Institute, Washington
Mondor, E. B., Tremblay, M. N., Awmack, C. S., and Lindorth, R. L. 2004. Divergent pheromone-mediated insect behaviour under global atmospheric change. Glob. Chang. Biol. 10:1820–1824.
Monson, R. K. 2003. The many faces of plant carbon relations: forging an ecophysiological identity in the age of human influence. New Phytol. 157:167–173.
Norby, R. J., Cotrufo, M. F., Ineson, P., O’neill, E. G., and Canadell, J. G. 2001. Elevated CO2, litter chemistry, and decomposition: a synthesis. Oecologia 127:153–165.
Norby, R. J., Delucia, E. H., Gielen, B., Calfapietra, C., Giardina, C. P., King, J. S., Ledford, J., Mccarthy, H. R., Moore, D. J. P., Ceulemans, R., De Angelis, P., Finzi, A. C., Karnosky, D. F., Kubiske, M. E., Lukac, M., Pregitzer, K. S., Scarascia-Mugnozza, G. E., Schlesinger, W. H., and Oren, R. 2005. Forest response to elevated CO2 is conserved across a broad range of productivity. Proc. Natl. Acad. Sci. USA 102:18052–18056.
Nykänen, H., and Koricheva, J. 2004. Damage-induced changes in woody plants and their effects on insect herbivore performance: a meta-analysis. Oikos 104:247–268.
Oksanen, E., Riikonen, J., Kaakinen, S., Holopainen, T., and Vapaavuori, E. 2005. Structural characteristics and chemical composition of birch (Betula pendula) leaves are modified by increasing CO2 and ozone. Glob. Chang. Biol. 11:732–748.
Ollinger, S. V., Aber, J. D., and Reich, P. B. 1997. Simulating ozone effects on forest productivity: interactions among leaf-, canopy-, and stand-level processes. Ecol. Appl. 7:1237–1251.
Parmesan, C. 2006. Ecological and evolutionary responses to recent climate change. Annu. Rev. Ecol. Syst. 37:637–669.
Parsons, W. F. J., Lindroth, R. L., and Bockheim, J. G. 2004. Decomposition of Betula papyrifera leaf litter under the independent and interactive effects of elevated CO2 and O3. Glob. Chang. Biol. 10:1666–1677.
Parsons, W. F. J., Bockheim, J. G., and Lindroth, R. L. 2008. Independent, interactive, and species-specific responses of leaf litter decomposition to elevated CO2 and O3 in a northern hardwood forest. Ecosystems 11:505–519.
Pearson, P. N., and Palmer, M. R. 2000. Atmospheric carbon dioxide concentrations over the past 60 million years. Nature 406:695–699.
Pelini, S. L., Prior, K. M., Parker, D. J., Dzurisin, J. D. K., Lindroth, R. L., and Hellmann, J. J. 2009. Climate change and temporal and spatial mismatches in insect communites, pp. 215–231, in Letcher, T. M., (ed.) Climate Change: Observed Impacts on Planet Earth. Elsevier, Oxford.
Peltonen, P. A., Vapaavuori, E., and Julkunen-Tiitto, R. 2005. Accumulation of phenolic compounds in birch leaves is changed by elevated carbon dioxide and ozone. Global Change Biol. 11:1305–1324.
Peltonen, P. A., Julkunen-Tiitto, R., Vapaavuori, E., and Holopainen, J. K. 2006. Effects of elevated carbon dioxide and ozone on aphid oviposition preference and birch bud exudate phenolics. Glob. Chang. Biol. 12:1670–1679.
Peñuelas, J., and Estiarte, M. 1998. Can elevated CO2 affect secondary metabolism and ecosystem function? Trends Ecol. Evol. 13:20–24.
Peñuelas, J., Estiarte, M., and Llusià, J. 1997. Carbon-based secondary compounds at elevated CO2. Photosynthetica 33:313–316.
Percy, K. E., Awmack, C. S., Lindroth, R. L., Kubiske, M. E., Kopper, B. J., Isebrands, J. G., Pregitzer, K. S., Hendrey, G. R., Dickson, R. E., Zak, D. R., Oksanen, E., Sober, J., Harrington, R., and Karnosky, D. F. 2002. Altered performance of forest pests under atmospheres enriched by CO2 and O3. Nature 420:403–407.
Percy, K. E., Sirkku, M., Karl-Heinz, H., Heerdt, C., Werner, H., Henderson, G. W., Rainer M. 2009. Effect of 3 years’ free-air exposure to elevated ozone on mature Norway spruce (Picea abies (L.) Karst.) needle epicuticular wax physicochemical characteristics. Environ. Pollut. 157:1657–1665.
Pinto, D. M., Blande, J. D., Souza, S. R., Nerg, A.-M., Holopainen, J. K. 2010. Plant volatile organic compounds (VOCs) in ozone (O3) polluted atmospheres: the ecological effects. J. Chem. Ecol. — this issue.
Raffa, K. F., Aukema, B. H., Bentz, B. J., Carroll, A. L., Hicke, J. A., Turner, M. G., and Romme, W. H. 2008. Cross-scale drivers of natural disturbances prone to anthropogenic amplification: the dynamics of bark beetle eruptions. Bioscience 58:501–517
Rossi, A. M., Stiling, P., Moon, D. C., Cattell, M. V., and Drake, B. G. 2004. Induced defensive response of myrtle oak to foliar insect herbivory in ambient and elevated CO2. J. Chem. Ecol. 30:1143–1152.
Roth, S. K., and Lindroth, R. L. 1995. Elevated atmospheric CO2: effects on phytochemistry, insect performance and insect-parasitoid interactions. Glob. Chang. Biol. 1:173–182.
Roth, S., Mcdonald, E. P., and Lindroth, R. L. 1997. Atmospheric CO2 and soil water availability: consequences for tree-insect interactions. Can. J. For. Res. 27:1281–1290.
Roth, S., Lindroth, R. L., Volin, J. C., and Kruger, E. L. 1998. Enriched atmospheric CO2 and defoliation: effects on tree chemistry and insect performance. Glob. Chang. Biol. 4:419–430.
Sager, E. P., Hutchinson, T. C., Croley, T. R. 2005. Foliar phenolics in sugar maple (Acer saccharum) as a potential indicator of tropospheric ozone pollution. Environ. Monit. Assess. 105:419–430.
Sallas, L., Kainulainen, P., Utriainen, J., Holopainen, T., and Holopainen, J. K. 2001. The influence of elevated O3 and CO2 concentrations on secondary metabolites of Scots pine (Pinus sylvestris L.) seedlings. Glob. Chang. Biol. 7:303–311.
Saxe, H., Ellsworth, D. S., and Heath, J. 1998. Tree and forest functioning in an enriched CO2 atmosphere. New Phytol. 139:395–436.
Scherzer, A. J., Rebbeck, J., and Boerner, R. E. J. 1998. Foliar nitrogen dynamics and decomposition of yellow-poplar and eastern white pine during four seasons of exposure to elevated ozone and carbon dioxide. For. Ecol. Manag. 109:355–366.
Sharkey, T. D., and Lerdau, M. T. 1999. Atmospheric chemistry and hydrocarbon emissions from plants. Ecol. Appl. 9:1107–1108.
Sitch, S., Cox, P. M., Collins, W. J., and Huntingford, C. 2007. Indirect radiative forcing of climate change through ozone effects on the land-carbon sink. Nature 448:791–794.
Skärby, L., Ro-Poulsen, H., Wellburn, F. A. M., and Sheppard, L. J. 1998. Impacts of ozone on forests: a European perspective. New Phytol. 139:109–122.
Stange, G. 1997. Effects of changes in atmospheric carbon dioxide on the location of hosts by the moth, Cactoblastis cactorum. Oecologia 110:539–545.
Stiling, P., and Cornelissen, T. 2007. How does elevated carbon dioxide (CO2) affect plant-herbivore interactions? A field experiment and meta-analysis of CO2-mediated changes on plant chemistry and herbivore performance. Glob. Chang. Biol. 13:1823–1842.
Stiling, P., Rossi, A. M., Hungate, B., Dijkstra, P., Hinkle, C. R., Knott, W. M. III, and Drake, B. 1999. Decreased leaf-miner abundance in elevated CO2: reduced leaf quality and increased parasitoid attack. Ecol. Appl. 9:240–244.
Stiling, P., Cattell, M., Moon, D. C., Rossi, A., Hungate, B. A., Hymus, G., and Drake, B. 2002. Elevated atmospheric CO2 lowers herbivore abundance, but increases leaf abscission rates. Glob. Chang. Biol. 8:658–667.
Stiling, P., Moon, D. C., Hunter, M. D., Colson, J., Rossi, A. M., Hymus, G. J., and Drake, B. G. 2003. Elevated CO2 lowers relative and absolute herbivore density across all species of a scrub-oak forest. Oecologia 134:82–87.
Stiling, P., Moon, D., Rossi, A., Hungate B. A., and Drake, B. 2009. Seeing the forest for the trees: long-term exposure to elevated CO2 increases some herbivore densities. Glob. Chang. Biol. 15:1895–1902.
STRAIN, B. R., and BAZZAZ, F. A. 1983. Terrestrial plant communities, pp. 177–222, in E. R. Lemon (ed.). CO2 and Plants. The Response of Plants to Rising Levels of Atmospheric Carbon Dioxide. Westview Press, Inc., Boulder, CO.
Strong, D. R., Lawton, J. H., and Southwood, R. 1984. Insects on Plants. Harvard University Press, Cambridge.
Swift, M. J., Heal, O. W., and Anderson, J. M. 1979. Decomposition in Terrestrial Ecosystems. Blackwell Publications, Oxford, England.
Taiz L, Zeiger E (2002) Plant Physiology, 3rd ed. Sinauer Associates, Sunderland.
Traw, M. B., Lindroth, R. L., and Bazzaz, F. A. 1996. Decline in gypsy moth (Lymantria dispar) performance in an elevated CO2 atmosphere depends upon host plant species. Oecologia 108:113–120.
Tylianakis, J. M., Didham, R. K., Bascompte, J., and Wardle, D. A. 2008. Global change and species interactions in terrestrial ecosystems. Ecol. Lett. 11:1351–1363.
Valkama, E., Koricheva, J., and Oksanen, E. 2007. Effects of elevated O3, alone and in combination with elevated CO2, on tree leaf chemistry and insect herbivore performance: a meta-analysis. Glob. Chang. Biol. 13:184–201.
Van Mantgem, P. J., Stephenson, N. L., Byrne, J. C., Daniels, L. D., Franklin, J. F., Fule, P. Z., Harmon, M. E., Larson, A. J., Smith, J. M., Taylor, A. H., and Veblen, T. T. 2009. Widespread increase of tree mortality rates in the western United States. Science 323:521–524.
Veteli, T. O., Mattson, W. J., Niemelä, P., Julkunen-Tiitto, R., Kellomäki, S., Kuokkanen, K., and Lavola, A. 2007. Do elevated temperature and CO2 generally have counteracting effects on phenolic phytochemistry of boreal trees? J. Chem. Ecol. 33: 287–296.
Vingarzan, R. 2004. A review of surface ozone background levels and trends. Atmos. Environ. 38:3431–3442.
Wennberg, P. O., and Dabdub, D. 2008. Rethinking ozone production. Science 319:1624–1625.
Whitham, T. G., Bailey, J. K., Schweitzer, J. A., Shuster, S. M., Bangert, R. K., Leroy, C. J., Lonsdorf, E., Allan, G. J., Difazio, S. P., Potts, B. M., Fischer, D. G., Gehring, C. A., Lindroth, R. L., Marks, J., Hart, S. C., Wimp, G. M., and WOOLEY, S. C. 2006. A framework for community and ecosystem genetics: from genes to ecosystems. Nat. Rev., Genet. 7:510–523.
Wittig, V. E., Ainsworth, E. A., and Long, S. P. 2007. To what extent do current and projected increases in surface ozone affect photosynthesis and stomatal conductance of trees? A meta-analytic review of the last three decades of experiments. Plant Cell Environ. 30:1150–1162.
Wittig, V. E., Ainsworth, E. A., Naidu, S. L., Karnosky, D. F., and Long, S. P. 2009. Quantifying the impact of current and future tropospheric ozone on tree biomass, growth, physiology and biochemistry: a quantitative meta-analysis. Glob. Chang. Biol. 15:396–424.
Yuan, J. S., Himanen, S. J., Holopainen, J. K., Chen, F., and Stewart, C. N. 2009. Smelling global climate change: mitigation of function for plant volatile organic compounds. Trends Ecol. Evol. 24:323–331.
Zvereva, E. L., and Kozlov, M. V. 2006. Consequences of simultaneous elevation of carbon dioxide and temperature for plant-herbivore interactions: a metaanalysis. Glob. Chang. Biol. 12:27–41.
Acknowledgments
Two decades of global change science by my research group have been generously supported by grants from the National Science Foundation, U.S. Department of Agriculture (National Research Initiatives) and the U.S. Department of Energy (Office of Science; currently, grant DE-FG02-06ER64232). Comments from two reviewers, including Stephan Hättenschwiler, improved the manuscript. I am deeply appreciative of the contributions of numerous undergraduate, graduate, postdoctoral and technical associates who have contributed to nearly 20 years of global change research in the Lindroth Lab, and grateful to the Source of grace that, like rain, falls on us all.
Author information
Authors and Affiliations
Corresponding author
Additional information
Silverstein-Simeone Award Lecture, 2009
Rights and permissions
About this article
Cite this article
Lindroth, R.L. Impacts of Elevated Atmospheric CO2 and O3 on Forests: Phytochemistry, Trophic Interactions, and Ecosystem Dynamics. J Chem Ecol 36, 2–21 (2010). https://doi.org/10.1007/s10886-009-9731-4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10886-009-9731-4