
Abstract
Tsunami may strongly impact beach ecosystems. To assess its magnitude five beaches along the Sendai Bay, Japan, were studied 2 months after the 11th March 2011 Tohoku-oki tsunami with focus on their recovery and meiofauna assemblages within few weeks after the event. The beaches recovered and new meiofauna assemblages established, which were strongly correlated to sediment grain size. The new data and review of previous works suggest that for beach ecosystems tsunami plays a role of ecosystem disturbance, not a catastrophe.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
9.1 Introduction
Tsunami is a series of waves characterised by small amplitude and very long wavelengths. They are caused by displacement of a large volume of water, usually attributed to earthquake, landslide or volcanic eruptions. As tsunami approaches the coast and the waters become shallow the wave amplitude significantly increases and seawater may inundate the coastal zone up to several tens of meters above sea level. Large tsunami waves (e.g. 2004 Indian Ocean tsunami, 11th March 2011 Tohoku-oki tsunami) caused death of thousands of humans, generate massive damages of houses and infrastructure, erode, transport and deposit huge amounts of sediments, and affect the coastal zone environment. The environmental impact of tsunami includes for instance translation of large amount of saltwater on land, deposition of thick layer of salty sediment covering plants and former soils and damage of natural habitats due to erosion.
Coastal ecosystems are affected in various ways by tsunami (e.g. Szczuciński et al. 2006). Apart from widely studied impacts on coral reefs (e.g. Baird et al. 2005; Brown 2005) and mangrove forests (e.g. Dam Roy and Krishnan 2005; Cochard et al. 2008; Yanagisawa et al. 2009), the most evident effect is an alteration of distribution and abundance of living benthic organisms. Biological investigations of tsunami impact on benthic communities have focused mainly on the larger macrofauna organism, which can readily be counted and identified (Kendall et al. 2009; Whanpetch et al. 2010; Lomovasky et al. 2011). Smaller organisms (meiofauna) received less attention, despite their significant roles in ecosystem functions, such as nutrient cycling and energy flow (for details see Giere 2009). Sandy beaches provide the habitat with the most diverse meiofauna communities. At the same time, beaches are one of the most tsunami affected parts of the coastal zone.
Reports from the previous tsunamis documented almost complete beach erosion (MacInnes et al. 2009; Paris et al. 2009), but also relatively fast beach recovery, completed its reversion back to the equilibrium stage within 1 year after the tsunami disturbance (Choowong et al. 2009). Both processes obviously have an effect on beach dwellers. So far the studies of possible impacts on meiofauna of episodic events such as tsunami are limited to few studies after 2004 Indian Ocean tsunami (Altaff et al. 2005; Kotwicki and Szczuciński 2006) suggesting quick recovery of meiofauna organisms.
Here we report the ecological status of meiofaunal assemblages at five sandy beaches shortly after the 2011 Tohoku-oki tsunami, with the aim of evaluating the natural meiofauna resilience and recovery after tsunami damage. To our knowledge this is the first report investigating the impact of the Tohoku-oki tsunami on beach faunal biota. The available records on tsunami impact on beach meiofauna are discussed in terms of its ecological significance.
9.2 Study Area and 11th March 2011 Tsunami
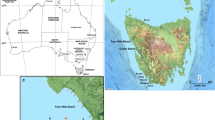
The devastating tsunami generated by the Mw 9.0 earthquake (Simons et al. 2011) struck northeastern Japan on 11th March 2011. The survey area includes five sandy beaches located at the Sendai Plain, directly exposed to tsunami waves impact (Figs. 9.1 and 9.2). Tsunami inundation of coastal plain in the investigated area extended up to 5 km inland. Flow heights (tsunami elevation above a mean sea-level datum) were over 10 m near the coastal dune ridges, and decreased landward to less than 3 m close to the inundation limit (Mori et al. 2011; The 2011 Tohoku Earthquake Tsunami Joint Survey Group 2011).

Study area, location of the studied transects (at each transect meiofauna and sediment were sampled at three positions- at low tide, mean sea level and high tide position). Simplified bathymetry after Saito (1989). Inset map shows location of the study area in Japan

Satellite images (source Google Earth) showing pre- and post-tsunami beach changes with indication of sampling transects
Changes in daily tidal range are app. 1 m (Goto et al. 2012). Offshore of the Sendai coast the seabed is covered by sand or silt with no bedrock exposed (Miyagi Prefecture 2000).
9.2.1 Beach Erosion and Recovery
Although more than 10 m high tsunami inundated the Sendai beaches, there was only minor beach erosion (Goto et al. 2011). The recovery of Arahama beach was studied by Goto et al. (2012) based on the analyses of the field and DEM (digital elevation model) data measured in April, June and July. Moreover, the topographic profile before (2005–2006) and after (March 19–23, 2011) the tsunami using 5 m DEM data were extracted. These data clarified that the berm was fully eroded and the beach profile was smoothed by the tsunami. However, new berm, which is nearly the same scale to that before the tsunami, was quickly recovered until the early June. In fact, the beach width was not markedly affected by tsunami erosion with difference of only 10 m in width before (app. 130 m) and after (app. 120 m) the tsunami (Goto et al. 2012).
Although the beach was one of the major sediment sources for tsunami deposits left on land (e.g. Szczuciński et al. 2012) the observations from the Sendai Bay coast by Goto et al. (2012), Richmond et al. (2012), Tappin et al. (2012) and Udo et al. (2012), point that most of the eroded beaches in the investigated area were rebuilt with sand within 1 month after the tsunami.
9.3 Material and Methods
9.3.1 Sampling and Samples Processing
The five studied beaches (Table 9.1) were sampled approximately 50 days after the 11th March Tohoku tsunami event (4th–10th May 2011). At each beach’s transect meiofauna and sediments were sampled in three positions – at low tide, mean sea level and high tide position using a meiocore sampler (inner diameter 3.6 cm). The upper 5 cm of sediment was taken and, for meiofauna analysis, immediately fixed in 70 % alcohol. In laboratory, a standard decantation technique was used to extract the animals from the sediment. Samples were washed over a 500 μm mesh, then sieved on a 32 μm mesh to retain the meiofauna fraction and stained with Bengal Rose. All metazoan meiobenthic organisms were classified at higher taxon level following Higgins and Thiel (1988) and counted under a stereo-microscope. Meiofaunal abundance was calculated per 10 cm2.
Sediment samples were subjected to grain size analysis with optical diffractometry method on laser diffraction based Mastersizer 2000 Particle Analyzer. All the results are presented in phi scale (larger value means finer sediments). Conversion of metric scale into phi values is based on:
where D is the size in mm. The grain size statistics (mean, sorting, skewness and kurtosis) were calculated using logarithmic method of moments with Gradistat software (Blott and Pye 2001).
The multivariate community data on major taxa level (square-root transformed, Bray-Curtis similarity was used to calculate resemblance) was analysed by means of non-parametric permutational ANOVA (PERMANOVA) to assess differences between investigated beaches and sampling position within the beach. In the case of a significant result in the PERMANOVA main test, pairwise tests for significant term were performed. Due to a restricted number of possible permutations in pairwise tests, p-values were obtained from Monte Carlo sampling (Anderson and Robinson 2003). Principal coordinates analysis (PCO) was used to visualise the PERMANOVA results. Total densities per 10 cm2, richness (S), and diversity (Shannon diversity index-H’loge) were calculated for each investigated beach. For analysis of selected biological variables separate univariate PERMANOVA tests were performed. To analyse and model the relationship between meiofauna assemblages and the environmental variables (grain size statistics: mean, sorting, skewness, kurtosis) DISTLM (distance-based linear model) routine was used (log transformed, Euclidean distance was used to calculate resemblance). All the multivariate analyses and calculations of S and H′ were performed using the PRIMER v6 with PERMANOVA+ add-on software package (Anderson et al. 2008).
9.4 Results
9.4.1 Beach Sediments
The studied surface beach sediments were composed mostly of medium-grained sand (Table 9.1). The mean grain size ranged from 0.67Φ at Natori N to 1.84Φ at Shichigahama site (Fig. 9.3).

Images of investigated beaches with indication of sampling position (left panel), LW low tide, MW mean water, HW high tide. Total meiofauna density, mean grain size and Shannon-Wiener index (right panel)
Coarse sand was encountered only in Natori S and Natori N beaches. In general the finer sediments were found on beaches protected by artificial structures (pier in case of Shichigahama, breakwaters at Arahama beach). Moderately well sorted sand constituted most of the beach sediments, apart from the Arahama site where sediments were composed of well sorted sand. The grain size distribution was mostly symmetrical, with some exception of fine and very fine skewed sediments encountered at Arahama and Shichigahama sites, and mesokurtic. Only Shichigahama beach sediments exhibited different grain distribution being characterised by very leptokurtic grained sediments (Table 9.1).
9.4.2 Meiofauna Analysis
The meiofauna in the study area was composed of 12 higher taxa, three represented by larval stages – copepods nauplii, polychaetes and insects (Table 9.2). Between one to eight higher meiofaunal taxa were recorded at single sampling point and significant differences in number of taxa were observed between the investigated beaches (Table 9.3A). Five taxa accounted for 96 % of meiofaunal communities: nematodes, copepods nauplii, gastrotrichs, turbellarians and ostracods. Nematodes were by far the most abundant group (~73 % of the total individuals), presented the highest density (mean abundance 307 indiv./10 cm2) and as the only taxon occurred in all the samples. Turbellarians and gastrotrichs were also very common taxa, as they were present in 80 % of the analysed samples. Nematodes dominated at almost all sampling stations. Only at Natori N (LW, MW) and Arahama beach (LW) percentage of this taxon was lower (Fig. 9.3), and meiofauna community was dominated by polychaetes, copepods nauplii and turbellarians, respectively. Other taxa such as water mites (Acari) and Bivalvia occurred only occasionally and/or in low numbers per sampling station, e.g. Harpacticoida. Total meiofaunal abundances varied significantly between the investigated beaches, being the highest at the Shichiganama (Table 9.3A, Fig. 9.3). Densities at other beaches were much lower, with great differences between sampling position (Fig. 9.3). At Arahama, Natori N and Shichiganama the highest densities were recorded at mean water level, while at Natori S and Natori C this position was characterised by lower abundance.
The variability of the Shannon-Wiener diversity index (H’) at the studied beaches was high and statistically significant (Table 9.3A). Consistent spatial meiofaunal diversity pattern was observed, with decreasing value of H’ from low to high water level. The only exception was Shichiganama, where generally H’ was very low and almost no differences between sampling points were observed (Fig. 9.3).
Results of multivariate data showed that investigated beaches harbour distinct meiofaunal assemblages (Table 9.3B). The most different from other beaches was Shichiganama. Pair-wise comparisons showed that also Arahama and Natori N were significantly different from each other.
When analysing meiofaunal structure in a PCO (Fig. 9.4), the first two components explained almost 68 % of the total variability. The graph showed a clear separation between Shichigahama, Arahama and other beaches. Observed differences are mainly connected with presence of nematodes and their gradual increase in abundance from Natori beaches to Shichigahama, while Natori beaches are characterised by increasing relationship with gastrotrichs, copepods nauplii and harpacticoids.

Principal coordinates ordination plot for meiofauna higher taxa showing all taxa which were significantly correlated with the PCO coordinates (r > 0.7). Vectors length corresponds to the correlation values
Four environmental variables were included in the DISTLM procedure. The best fitted model (illustrated in Fig. 9.5) that explains the meiofaunal structure at the investigated beaches includes all of the variables, together explaining 41 % of total variation. However, only mean grain size, which is the main contributor in explaining observed variability in meiofauna assemblages (19.8 %) was verified as statistically significant.

Distance based redundancy plot illustrating the DISTLM model based on the higher meiofauna taxa assemblage data and fitted environmental variables with their vector (strength and direction of effect of the variable on the ordination plot)
9.5 Discussion and Conclusions
Many studies have shown meiofauna to be useful in assessing environmental disturbance, with the general conclusions that the resilience of meiofauna is relatively strong; even though, the response depends on the intrinsic resilience of the particular community, intensity of disturbance, or such stochastic factors as seasonal or trophic conditions in the disturbed area. Regardless of nature of the disturbance (anthropogenic, natural, chronic, episodic), such events generally result in a reduction in meiofauna density and/or diversity (e.g. Schratzberger and Warwick 1999; Lee et al. 2001). Nevertheless, some studies indicate that significant meiofauna population can occur even within 1 week of disturbance (deposited dredging material, Schratzberger et al. 2006). Similar observation was made by Altaff et al (2005) at an Indian beach soon after tsunami in December 2004, which flooded the coastline with a tsunami flow depth of up to 6 m. Five days after tsunami meiofauna density achieved typical values for that beach.
These reports and own observations may indicate that almost 2 months period, from tsunami event to sampling time, was long enough to colonize the newly built sandy beaches, and observed densities resulted from fast growth rates, typical for meiofaunal species life cycles. According to Heip et al. (1985) some of common nematode species, dominant taxon in most habitats, have annual number of generations even up to 17. However, since no data from pre-tsunami period are available, it is difficult to predict if established meiofauna assemblages at the investigated beaches are stable.
Probably, the composition of the post-disturbed meiofauna community will never be exactly the same as it was before, although the general structural traits may be predictable (Altaff et al. 2005; Grzelak et al. 2009). This suggestion seems even more reliable particularly when we look at the beach recovery process, which has been very rapid in comparison to previous studies (Goto et al. 2012), such as the 2004 Indian Ocean tsunami. For instance at Arahama transect the beach had almost completely reverted back to the pre-tsunami conditions within 3 months after the event, while beach recovery on the western coast of Thailand was estimated on 60 % within 6 months after tsunami 2004 event (Goto et al. 2012; Choowong et al. 2009, respectively). Moreover, the sediment characteristics of investigated beaches indicate relatively stable conditions.
In Thailand, beach sediments shortly after the 2004 Indian Ocean tsunami (app. 50 days after) were generally poorly sorted and very coarse skewed (Kotwicki and Szczuciński 2006). Comparable sediment characteristics to present result (better sorted and grain size distribution much closer to symmetrical) were achieved in Thailand within 1 year. Sediment properties and composition of sand particles, which form interstices, are undoubtedly one of the most important factors for the abundance, composition and colonization processes of meiofauna. Those variables determine spatial and structural conditions, as well as indirectly determine the physical and chemical regime of the sediment. Thus, little variability of the sediment, indicated by its grain size statistics, pointed at the existence of relatively favourable conditions for meiofauna organisms’ development and re-entry to beach ecosystem.
Observed differences between investigated beaches in meiofauna densities and composition may result from natural spatial variation and differences in complex of environmental conditions, for instance wave regime and sediment characteristic. As was shown by present study significant impact on meiofauna assemblages at investigated beaches has grain size (Fig. 9.5). Although it explained only 20 % of total variation in meiofauna assemblages, what means that several varied factors influence those communities (e.g. quantities and qualities of food available, predation) and should be taken into consideration during similar future investigations, grain size remains an important foundation for the living conditions and ecological status of meiobenthos.
High variability of meiofauna abundance was comparable to observation made by Kotwicki and Szczucinski (2006) at three Thai beaches impacted by tsunami in 2004. Generally speaking, the spatial distribution of meiofauna is extremely patchy and unpredictably variable (Giere 2009). Nevertheless, densities recorded in present study are of the same range as densities reported for other marine subtropical and tropical regions (Kotwicki et al. 2005; Giere 2009 and references therein). The same is true in terms of taxonomical composition.
The results on meiofauna assemblages in newly (after the tsunami) rebuild parts of the studied beaches suggest that recolonisation process occurs very fast. It is not surprising, since experimental studies have shown that the first stage of the recolonisation proceeds rapidly, as long as the ecological niches remain wide and can be occupied by the range of opportunistic generalists (e.g. De Troch et al. 2005).
Nematodes were dominant group at all investigated beaches. This taxon is characterised by the greatest species richness in the benthic zone, what resulting in utilization of all trophic resources. Set of morphological features together with their frequent occurrence make nematodes possible to occupy wide range of ecological niches with different trophic requirements and sediment properties. Moreover, nematodes appear to be among the most abundant rafting organisms (Thiel and Gutow 2005), what makes them good colonizers. For this reason nematodes are usually observed in high densities, even after severe disturbance events and are considered to be the most valuable tools for pollution studies (e.g. Sherman and Coull 1980; Coull and Chandler 1992; Lee et al. 2001; Altaff et al. 2005; Kotwicki and Szczuciński 2006; Szymelfenig et al 2006; Hua et al. 2010).
Harpacticoids are usually second the most abundant taxa in meiobenthic samples. Representatives of this group are considered as the classical ‘emergers’ among the meiobenthos. It means that they have high ability to intensive dispersal, redistribution and colonization of new or disturbed habitats. As was shown by experimental studies dispersal rates of harpacticoids might be even 65-fold higher than those of nematodes (Commito and Tita 2002). That is why, their very low abundance or even absence at Shichigahama beach might suggest lack of donor communities in the proximity of investigated beaches, which is important factor in colonization potential and success (Derycke et al. 2007).
Meiofauna is usually considered as less sensitive to disturbance and less persistently affected than macrofauna (Kennedy and Jacoby 1999; Giere 2009). In particular, responds rapidly to environmental changes, both of natural and anthropogenic origin. Meiofauna can resist disturbance such as tsunami waves because their small body size may allow them to be re-suspended and short life cycles allow them to withstand high mortality. Thus, it appears that the initial respond of meiofauna assemblages to tsunami event is similar, regardless of the climatic zone, and benthic meiofaunal communities are highly developed, even shortly after disturbance.
Although the studies of environmental impact of tsunami on beach environment are still very limited it seems that although the beach is usually subjected to strong erosion, the new beach sediments are rapidly colonized by meiofauna organisms both in tropical and temperate climates. Consequently, in terms of beach ecosystems tsunami events shall be considered as ecological disturbance not a catastrophe.
References
Altaff K, Sugumaran J, Naveed MS (2005) Impact of tsunami on meiofauna of Marina beach, Chennai, India. Curr Sci 89(1):34–38
Anderson MJ, Robinson J (2003) Generalized discriminant analysis based on distances. Aust NZ J Stat 45:301–318
Anderson MJ, Gorley RN, Clarke KR (2008) PERMANOVA+ for PRIMER: guide to software and statistical methods. PRIMER-E, Plymouth
Baird AH, Campbell SJ, Anggoro AW, Ardiwijaya RL, Fadli N, Herdiana Y, Kartawijaya T, Mahyiddin D, Mukminin A, Pardede ST, Pratchett MS, Rudi E, Siregar AM (2005) Acehnese reefs in the wake of the Asian tsunami. Curr Biol 15(21):1926–1930
Blott SJ, Pye K (2001) Gradistat: a grain size distribution and statistics package for the analysis of unconsolidated sediments. Earth Surf Proc Land 26:1237–1248
Brown BE (2005) The fate of coral reefs in the Andaman Sea, eastern Indian Ocean following the Sumatran earthquake and tsunami, 26 December 2004. Geogr J 171:372–374
Choowong M, Phantuwongraj S, Charoentitirat T, Chutakositkanon V, Yumuang S, Charusiri P (2009) Beach recovery after 2004 Indian Ocean tsunami from Phang-Nga, Thailand. Geomorphology 104:134–142
Cochard R, Ranamukhaarachchi SL, Shivakoti GP, Shipin OV, Edwards PJ, Seeland KT (2008) The 2004 tsunami in Aceh and Southern Thailand: a review on coastal ecosystems, wave hazards and vulnerability. Perspect Plant Ecol, Evol Syst 10(1):3–40
Commito JA, Tita G (2002) Differential dispersal rates in an intertidal meiofauna assemblage. J Exp Mar Biol Ecol 268:237–256
Coull BC, Chandler GT (1992) Pollution and meiofauna: field, laboratory, and mesocosm studies. Oceanogr Mar Biol Annu Rev 30:191–271
Dam Roy S, Krishnan P (2005) Mangrove stands of Andamans vis-à-vis tsunami. Curr Sci 89(11):1800–1804
De Troch M, Vandepitte L, Reas M, Suarez-Morales E, Vincx M (2005) A field colonization experiment with meiofauna and seagrass mimics: effect of time, distance and leaf surface area. Mar Biol 148:73–86
Derycke S, Van Vynckt R, Vanaverbeke J, Vincx M, Moens T (2007) Colonization patters of nematoda on decomposing algae in the estuarine environment: community assembly and genetic structure of the dominant species Pellioditis marina. Limnol Oceanogr 52:992–1001
Giere O (2009) Meiobenthology. The microscopic motile fauna of aquatic sediments. Springer, Berlin
Goto K, Chagué-Goff C, Fujino S, Goff J, Jaffe B, Nishimura Y, Richmond B, Sugawara D, Szczuciński W, Tappin DR, Witter R, Yulianto E (2011) New insights into tsunami hazard from the 2011 Tohoku-oki event. Mar Geol 290:46–50
Goto K, Sugawara D, Abe T, Haraguchi T, Fujino S (2012) Liquefaction as an important source of the 2011 Tohoku-oki tsunami deposits at Sendai Plain, Japan. Geology 40(10):887–890
Grzelak K, Kotwicki L, Szczuciński W (2009) Monitoring of sandy beach meiofaunal assemblages and sediments after the 2004 tsunami in Thailand. Pol J Environ Stud 18(1):43–51
Heip C, Vincx M, Vranken G (1985) The ecology of marine nematodes. Oceanogr Mar Biol Ann Rev 23:399–489
Higgins RP, Thiel H (1988) Introduction to the study of meiofauna. Smithsonian Institution Press, London
Hua E, Zhang ZN, Yu ZS, Zhang Y (2010) Preliminary study on the immediate response of the nematode community to Typhoon Soudelor. Deep-Sea Res II 57:1064–1070
Kendall MA, Aryuthaka C, Chimonides J, Daungnamon D, Hills J, Jittanoon C, Komwachirapitak P, Kongkaew V, Mittermeyr A, Monthum Y, Nimsantijaroen S, Paterson GLJ, Foster-Smith R, Foster-Smith J, Thongsin N (2009) Post-tsunami recovery of shallow water biota and habitats on Thailand’s Andaman coast. Pol J Environ Stud 18(1):69–76
Kennedy AD, Jacoby CA (1999) Biological indicators of marine environmental health: meiofauna- a neglected benthic component? Environ Monit Assess 54:47–68
Kotwicki L, Szczuciński W (2006) Meiofauna assemblages and sediment characteristic of sandy beaches on the west coast of Thailand after the 2004 tsunami event. Phuket Mar Biol Centre Res Bul 67:39–47
Kotwicki L, Szymelfenig M, De Troch M, Urban-Malinga B, Weslawski JM (2005) Latitudinal biodiversity patterns of meiofauna from sandy litoral beaches. Biodivers Conserv 14:461–474
Lee H, Vanhove S, Peck LS, Vincx M (2001) Recolonisation of meiofauna after catastrophic iceberg scouring in shallow Antarctic sediments. Polar Biol 24:918–925
Lomovasky BJ, Firstater FN, Salaraz AG, Mendo J, Iribarne OO (2011) Macro benthic community assemblage before and after the 2007 tsunami and earthquake at Paracas Bay, Peru. J Sea Res 65:205–212
MacInnes BT, Bourgeois J, Pinegina TK, Kravchunovskaya EA (2009) Tsunami geomorphology: erosion and deposition from the 15 November 2006 Kuril Island tsunami. Geology 37:995–998
Miyagi Prefecture (2000) Report of the committee for the technical survey at Sendai coast. 73 pp
Mori N, Takahashi T, Yasuda T, Yanagisawa H (2011) Survey of 2011 Tohoku earthquake tsunami inundation and run-up. Geophys Res Lett 38, L00G14. doi:10.1029/2011GL049210
Paris R, Wassmer P, Sartohadi J, Lavigne F, Barthomeuf B, Desgages E, Grancher D, Baumert P, Vautier F, Brunstein D, Gomez C (2009) Tsunamis as geomorphic crises: lessons from the December 26, 2004 tsunami in Lhok Nga, West Banda Aceh (Sumatra, Indonesia). Geomorphology 104:59–72
Richmond B, Szczuciński W, Chagué-Goff C, Goto K, Sugawara D, Witter R, Tappin DR, Jaffe B, Fujino S, Nishimura Y, Goff J (2012) Erosion, deposition and landscape change on the Sendai coastal plain, Japan, resulting from the March 11, 2011 Tohoku-oki tsunami. Sed Geol 282:27–39
Saito Y (1989) Modern storm deposits in the inner shelf and their recurrence intervals, Sendai Bay, northeast Japan. In: Taira A, Masuda F (eds) Sedimentary facies in the active plate margin. Terra Scientific Publishing Company, Tokyo, pp 331–344
Schratzberger M, Warwick RM (1999) Differential effects of various types of disturbances on the structure of nematode assemblages: an experimental approach. Mar Ecol Prog Ser 181:227–236
Schratzberger M, Bolam S, Whomersley P, Warr K (2006) Differential response of nematode colonist communities to the intertidal placement of dredged material. J Exp Mar Biol Ecol 334:244–255
Sherman K, Coull BC (1980) The response of meiofauna to sediment disturbance. J Exp Mar Biol Ecol 46:59–71
Simons M, Minson SE, Sladen A, Ortega F, Jiang J, Owen SE, Meng L, Ampuero JP, Wei S, Chu R, Helmberger DV, Kanamori H, Hetland E, Moore AW, Webb FH (2011) The 2011 magnitude 9.0 Tohoku-oki earthquake: mosaicking the megathrust from seconds to centuries. Science 332:1421–1425
Szczuciński W, Chaimanee N, Niedzielski P, Rachlewicz G, Saisuttichai D, Tepsuwan T, Lorenc S, Siepak J (2006) Environmental and geological impacts of the 26 December 2004 tsunami in coastal zone of Thailand – overview of short and long-term effects. Pol J Environ Stud 15(5):793–810
Szczuciński W, Kokociński M, Rzeszewski M, Chagué-Goff C, Cachão M, Goto K, Sugawara D (2012) Sediment sources and sedimentation processes of 2011 Tohoku-oki tsunami deposits on the Sendai Plain, Japan – insights from diatoms, nannoliths and grain size distribution. Sed Geol 282:40–56
Szymelfenig M, Kotwicki L, Graca B (2006) Benthic re-colonization in post-dredging pits in the Puck Bay (Southern Baltic Sea). Estuar Coast Shelf Sci 68:489–498
Tappin DR, Evans HM, Jordan CJ, Richmond B, Sugawara D, Goto K (2012) Coastal changes in the Sendai area from the impact of the 2011 Tōhoku-oki tsunami: interpretations of time series satellite images, helicopter-borne video footage and field observations. Sed Geol 282:151–174
The 2011 Tohoku Earthquake Tsunami Joint Survey Group (2011) Nationwide field survey of the 2011 off the Pacific coast of Tohoku earthquake tsunami. J Japan Soc Civil Eng, Ser B2 (Coast Eng) 67:63–66
Thiel M, Gutow L (2005) The ecology of rafting in the marine environment. II. The rafting organisms and community. Oceanogr Mar Biol Annu Rev 43:279–418
Udo K, Sugawara D, Tanaka H, Imai K, Mano A (2012) Impact of the 2011 Tohoku earthquake and tsunami on beach morphology along the northern Sendai coast. Coast Eng J 54, art. no. 1250009
Whanpetch N, Nakaoka M, Muaki H, Suzuki T, Nojima S, Kawai T, Aryuthaka C (2010) Temporal changes in benthic communities of seagrass beds impacted by a tsunami in the Andaman Sea, Thailand. Estuar Coast Shelf Sci 87:246–252
Yanagisawa H, Koshimura S, Goto K, Miyagi T, Imamura F, Ruangrassamee A, Tanavud C (2009) The reduction effects of mangrove forest on a tsunami based on field surveys at Pakarang Cape, Thailand and numerical analysis. Estuar Coast Shelf Sci 81(1):27–37
Acknowledgements
The fieldwork was supported by Adam Mickiewicz University in Poznań, Poland and Tohoku University in Sendai, Japan. The help of International Tsunami Survey Team members (C. Chagué-Goff, S. Fujino, J. Goff, K. Goto, B. Jaffe, Y. Nishimura, B. Richmond, D. Tappin, R. Witter and E. Yulianto) during the fieldwork and of Michał Rzeszewski during the lab measurements is greatly acknowledged.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2014 Springer Science+Business Media Dordrecht
About this chapter
Cite this chapter
Grzelak, K., Szczuciński, W., Kotwicki, L., Sugawara, D. (2014). Ecological Status of Sandy Beaches After Tsunami Events: Insights from Meiofauna Investigations After the 2011 Tohoku-oki Tsunami, Sendai Bay, Japan. In: Kontar, Y., Santiago-Fandiño, V., Takahashi, T. (eds) Tsunami Events and Lessons Learned. Advances in Natural and Technological Hazards Research, vol 35. Springer, Dordrecht. https://doi.org/10.1007/978-94-007-7269-4_9
Download citation
DOI: https://doi.org/10.1007/978-94-007-7269-4_9
Published:
Publisher Name: Springer, Dordrecht
Print ISBN: 978-94-007-7268-7
Online ISBN: 978-94-007-7269-4
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)