
Abstract
This review summarises the distribution of dissolved nutrients (nitrate, phosphate and silicate) over the period of modern measurements (~30 years) and aspects of the biogeochemical processes which control their distribution in the Eastern Mediterranean Sea (EMS). The levels of nitrate, phosphate and silicic acid (~6 μ mol N kg−1, 0.25 μ mol P kg−1 and 11 μ mol Si kg−1 in deep water) in the EMS are much lower than all other parts of the ocean due to the unusual anti-estuarine circulation which exports nutrient replete intermediate water at the Straits of Sicily and to the relatively young age of the deep water (τ ≈ 120 years). Dissolved oxygen decrease and nutrients increase eastwards, in the direction of the deep-water circulation with nitrate increasing by ~0.5 μ mol kg−1 from the Western Ionian to the Eastern Levantine, phosphate by 0.05 μ mol kg−1 and silicic acid by ~4.5 μ mol kg−1 representing the amount (rate) of organic matter and biogenic silica (BSi) breakdown over the residence time of Deep Water in the basin. The East Mediterranean Transient interrupted this simple pattern causing lower nutrients in the deep water and upwelled nutrients into the upper layers. The high nitrate/phosphate found in the deep water (25–28:1) is due to a combination of high Nitrate:Phosphate waters advected from the surface of the Adriatic during deep water formation during the P-limited winter bloom and P recycling more efficiently than N from the descending particulate organic matter (POM). High N:P ratios in POM and dissolved organic matter (DOM) show that the entire EMS is P starved. There are seasonal changes in nutrient limitation with conventional P limitation during the winter phytoplankton bloom which becomes N&P co-limitation in early summer and can, under some circumstances, become even N limited in mid-summer in the nutrient starved upper waters. N2 fixation rates are constantly low in the EMS compared to somewhat higher levels in the western MS. Nutrient budgets have been used to explain that the reason for the unusual N:P ratio in the basin is high N:P ratio in the external inputs combined with low denitrification rates caused by the ultra-oligotrophic status of the basin. However the external nutrient (both riverine and atmospheric) inputs have increased dramatically between 1950 and 2000. Estimates of the pristine flux of nutrients by riverine sources to the EMS are 2.0 × 109 moles N/y and 0.095 × 109 moles P/y while the atmospheric flux has increased by 85 % for NOx and 65 % for NH3 between 1910 and the end of the twentieth century. It is suggested that climate change may result in dramatic threshold changes in trophic status of the EMS if surface circulation rates decrease as has been predicted.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
Introduction
The Eastern Mediterranean Sea (EMS) is an unusual part of the global ocean. It is ultra-oligotrophic with primary productivity approximately half that of the Sargasso Sea (Krom et al. 2003). The nutrient concentrations in the deep water are amongst the lowest measured anywhere in the oceans despite the fact that the EMS is almost land-locked and has considerable nutrient inputs (both atmospheric and riverine), much of it anthropogenic, particularly from its northern shores. Its low nutrient concentration is due to a combination of the relatively young age of the water masses and to the unusual anti-estuarine circulation in which nutrient depleted waters flow in through the Straits of Sicily while nutrient enriched intermediate waters flow out. The nitrate: phosphate (N:P) ratio in the deep water is highly unusual being almost twice the Redfield ratio (Krom et al. 1991) while the N: P ratios of both dissolved and particulate organic matter is also far in excess of 16:1 making the whole system P starved. Recent work has explained this unusual N: P ratio as being due to high N: P ratios in the external supply combined with very low denitrification rates due to the ultra-oligotrophic nature of the basin (Krom et al. 2010). Together these unusual properties make the EMS an ideal natural laboratory to examine the nature of the controls on nutrient cycling and hence important processes related to carbon uptake in the ocean.
In this chapter we review the distribution of inorganic nutrients in the water column of the EMS over the past 30 years, the period over which modern nutrient measurements have been made. This time period includes the changes in the physical circulation caused by the Eastern Mediterranean Transient (EMT) event. The vertical distribution of dissolved and particulate organic matter is presented together with changes in chlorophyll-a and Deep Chlorophyll Maximum (DCM) across the basin. In examining the biogeochemical and nutrient cycling processes in the EMS, we describe why the deep waters of the EMS have such an unusual N:P ratio even though the uptake by phytoplankton in surface waters is approximately Redfieldian. Although primary productivity in the EMS is P limited during the main winter phytoplankton bloom, later in the year the nature of the nutrient limitation changes for different parts of the ecosystem including the phytoplankton becoming N and P co-limited. We also review the most recent measurements of N2 fixation rates across the basin which display very low rates throughout the year. Thus, in the EMS new N sources from current N2 fixation in the photic layers are negligible and contribute only a small percentage to new and primary production in the basin.
The review continues by examining the use of nutrient budgets to define biogeochemical processes within the basin. We show how the input terms in those budgets have changed with time and discuss the implications as regards the proportion of anthropogenic nutrients in the system. We finish by describing how changes in the physical circulation have caused significant modifications in biogeochemical processes in the past and speculate, how environmental and climate change might cause major changes in the future and what further studies are needed to understand the workings of this semi-enclosed sea.
General and Mesoscale Circulation and Changes in the Circulation
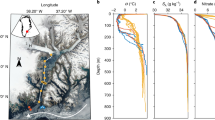
The general circulation of the Mediterranean Sea is anti-estuarine. Surface water of Atlantic origin (AW), low in nutrients, flows through Gibraltar and the Sicily Straits and spreads to the Ionian and Levantine, being modified along the way . This Modified Atlantic water (MAW) circulation in the EMS displays a number of mesoscale features with large variability in extent, shape and time scales (Malanotte-Rizzoli et al. 1999; POEM-Group 1992; Robinson et al. 1991). Levantine Intermediate Water (LIW), formed in the Levantine Basin, with higher nutrient concentrations, flows at intermediate depths (200–500 m) westwards from the Levantine Basin, through the Cretan Passage with the major flow directed towards the Sicily Straits, and a minor part introduced to the Adriatic Sea. The deep waters in the EMS are isolated from the Western Basin by the shallow sill depth of the Sicily Straits. Until the early 1990s the deep layer was occupied exclusively by dense water formed in the Southern Adriatic, known as Adriatic Deep Water (ADW) (Fig. 4.1). A combination of meteorological and hydrological factors led to the establishment of the Aegean as a new source of deep water in the area at the beginning of the 1990s, changing the circulation in the basin (Roether et al. 2007 and references therein). The event was termed the Eastern Mediterranean Transient (EMT). The EMT formed a new water mass, the Cretan Sea Outflow water (CSOW) (Klein et al. 1999). This new, younger, dense water was warmer and more saline than the older ADW and could be recognised by an inversion in the TS diagrams. The CSOW was also characterized by higher dissolved oxygen and lower nutrient concentrations than the ADW (Roether et al. 1996). The main outflow of CSOW (ca. 75 %) was delivered between mid 1992 and late 1994. It propagated preferentially westwards and more slowly towards the east. In 1991 there were strong signals of the CSOW at stations south of Crete located only to the north of the Eastern Mediterranean Ridge (EMRidge). By 1995 all the stations in the Levantine showed strong TS inversions while by 1999, the EMT was well established in the whole EMS (Roether et al. 1996; 2007; Klein et al. 2003; Kress et al. 2003) (Fig. 4.2). A snapshot of the evolution of the CSOW in the Levantine Basin taken in September 2008 (Fig. 4.3) showed the continuing change and erosion of the deep water structure. From 27 °E eastwards, potential temperature and salinity increased from the original Adriatic Deep water (ADWo) towards the bottom due to the CSOW, while close to Crete, a new deep water was present (ADWn) which was slightly less saline and colder compared to the CSOW (Manca et al. 2002, 2006; Rubino and Hainbucher 2007).

Vertical sections of salinity, potential temperature, dissolved oxygen, silicic acid, nitrate and phosphate along the west–east cross section of the Eastern Mediterranean in 1987. Stations and data points are indicated by filled dots (see inset map) (Adapted from Schlitzer et al. 1991; Klein et al. 1999)

Vertical sections of salinity, dissolved oxygen, silicic acid, nitrate and phosphate along the west–east cross section of the Eastern Mediterranean in 1999. Stations and data points are indicated by filled dots (see inset map) (Adapted from Kress et al. 2003)

Vertical sections of salinity, dissolved oxygen, silicic acid, nitrate and phosphate along the west–east cross section of the Levantine Basin in 2008. Stations and data points are indicated by filled dots (see inset map) (Adapted from Kress et al. 2011)
Description of the Present Distribution of Nutrients and Dissolved Oxygen and Changes Over the Past 25 years
Description of Typical Vertical Profiles of Nutrients and Dissolved Oxygen
The upper layer (0–150 m) across the EMS is quite homogeneous in dissolved oxygen and nutrient concentrations, influenced only by the strong mesoscale field (Figs. 4.1, 4.2, and 4.3). The surface waters are generally saturated with dissolved oxygen; indeed in summer a slight oversaturation often appears in the MAW (Kress and Herut 2001). The nutrient concentrations are very low, with phosphate in the nanomolar level throughout the year, while nitrate is at the nanomolar level in the summer and slightly higher in winter (~0.5–1 μmole kg−1) and silicic acid close to 1 μmole kg−1 (e.g. Krom et al. 2005; Kress et al. 2003; Pujo-Pay et al. 2011)
Typical composite depth profiles of dissolved oxygen, nitrate and silicic acid in the Levantine and central Ionian Basin after the EMT event are presented in Fig. 4.4 (Kress et al. 2003). Dissolved oxygen concentrations are high at the surface, decrease to minimal values at the 500–1,500 m layer and increase towards the bottom (Levantine) or down to 2,500 m, the concentrations remaining then essentially constant (Ionian). The nitrate depth profile is a mirror image of the dissolved oxygen profile: low concentrations at the surface, an increase to maximal values at the 500–1,500 m layer and a decrease towards the bottom or down to 2,500 m. The silicic acid depth profile is similar to that of nitrate, but the maximal layer is located deeper, due to the different remineralization processes, biological for nitrate and chemical dissolution for silicic acid.

Composite depth profiles of dissolved oxygen, nitrate and silicic acid in the Levantine (left panel, filled circles) and the central Ionian basin (right panel, open circles) in April–May 1999, after the EMT event (Adapted from Kress et al. 2003)
Cross Basin Distribution and How They Have Changed with Time
In comparison to the physical evolution, the changes in the distributions of dissolved oxygen and dissolved nutrients across the Eastern Mediterranean in the mid and deep waters (i.e. >300 m) have been less described in the literature (Schlitzer et al. 1991; Roether et al. 1996; Klein et al. 1999; 2003; Kress et al. 2011; 2003). In 1987, prior to the EMT (Fig. 4.1), the cross-basin distribution of dissolved oxygen and nutrients in the deep water was essentially uniform from 1,000 m and below, except for the Ionian that exhibited a mid-depth layer at ca. 1,200 m with minimum oxygen and maximum nutrients concentrations (MinOx/MaxNut) (with 178, 9.3, 5.0 and 0.16 μmole kg−1 for oxygen, silicic acid, nitrate and phosphate, respectively in the Mid Ionian) due to the intrusion of newly added ADW at depth toward the base of the western continental slope (Schlitzer et al. 1991). Dissolved oxygen decreased and nutrients increased eastwards, in agreement with the direction of the deep-water circulation. No mid depth MinOx/MaxNut layer was present in the Levantine Basin, and the waters were essentially homogeneous below 1,200 m depth with 170 μmole kg−1 dissolved oxygen and 10.5, 4.9 and 0.21 μmole kg−1 silicic acid, nitrate and phosphate, respectively. The nitrate increased by ~0.5 μmole kg−1 from the Western Ionian to the Eastern Levantine, while the phosphate increased by only 0.05 μmole kg−1 . Silicic acid increased by ~4.5 μ mol kg−1 while dissolved oxygen decreased by 20 μmole kg−1 . These nutrient changes represent the amount (rate) of organic matter and biogenic silica (BSi) breakdown at depth in the system over the residence time of ADW in the basin.
This simple pattern changed during the 1990s as a result of the EMT event. In 1991, the CSOW was confined to the vicinity of the Cretan Arc, but by 1995, ca. 6 years after the start of the EMT event its influence was present in most of the EMS. The deep water was no longer uniform and a pronounced MinOx/MaxNut layer was detected at the 500–1,500 m depth range depending on the basin sampled (Klein et al. 1999; Lascaratos et al. 1999). This layer corresponded to the older ADWo that was pushed up by the younger CSOW. The MinOx/MaxNut layer continued to change as the EMT evolved (Klein et al. 2003; Kress et al. 2003).
Nine years from its onset, in 1999, the CSOW was found in the whole basin and clearly discernible in the dissolved oxygen and nutrient vertical distributions (Fig. 4.2) (Klein et al. 2003; Kress et al. 2003). The MinOx/MaxNut layer, the old ADW, was found at the 300–1,500 m depth interval, thickening and deepening eastwards. The Minox concentrations were similar in the Levantine and Western Ionian (175 μmole kg−1) but located at different depth layers, 600–1,500 m and 300–800 m, respectively (Fig. 4.4). The MinOx at the Cretan Passage was slightly higher (175–180 μmole kg−1) indicating the presence of younger water. The maximal concentrations of phosphate and nitrate (> 0.175 and > 5.5 μmol kg−1, respectively) were similar in the Levantine and the Western Ionian with lower concentrations at the Cretan Passage and Eastern Ionian. The maximal concentration of silicic acid increased eastwards, from ca. 8.5 to >9.5 μmol kg−1 due to increasing age of the water mass. Maximal values were centered at 1,200–1,400 m, deeper than the maxima for nitrate and phosphate due to the different remineralisation pathways (slower chemical dissolution for silicic acid as opposed to biological remineralisation for N and P). The CSOW was very noticeable in the Levantine, the concentrations of nutrients decreasing from the maxima in the ADW to minima of 4.7, 0.17 and 8.2 μmol kg−1 for nitrate, phosphate and silicic acid, respectively in the CSOW. Before the EMT event, nitrate and phosphate concentrations were constant below 1,200 m while silicic acid concentrations continued to increase with depth (Kress and Herut 2001). In the Eastern Ionian and Western Crete (19–25.5 oE), below 2,000 m, the concentrations of nutrients were lower (<4.4, <0.15 and < 7.8 μmol kg−1 for nitrate, phosphate and silicic acid, respectively) and dissolved oxygen higher than those found in the Levantine and in the westernmost Ionian, indicating a source of younger water.
By 2008, ca. 12 years after the maximum CSOW outflow, the effect of the EMT could still be identified in the Levantine Basin (Fig. 4.3). The MinOx layer was centred at ca. 900 m, more emphasized in the eastern part of the transect (<175 μmol kg−1 up to 27 oE) and eroded towards the west (minimal concentrations of 180–175 μmol kg−1). The concentrations increased towards the bottom, in particular westwards of 27 oE, indicating mixing and erosion of the CSOW in the east and penetration of younger waters in the west. The MaxNO3/PO4 was located at 400–1,500 m depth interval with maximal concentrations of > 5 and > 0.2 μmol kg−1, respectively, the upper isoline (i.e. lines of constant concentration) shoaling eastwards up to 400 m depth. The concentrations decreased towards the bottom, except for phosphate at 25.5–26.5 oE. The MaxSi was positioned at the 700–1,750 m depth interval, centered at 1,200 m, similar to the position of the MinSal/T . This layer shoaled eastwards with the upper 9 μmol kg−1 isoline reaching 700 m depth. There were indications of deep intrusion of seawater with lower silicic acid concentration in the vicinity of Crete.
Dissolved and Particulate Organic Carbon and Nutrients
The concentration of dissolved organic carbon (DOC) measured in the SE. Levantine Basin (Krom et al. 2005) is similar to that measured in the Ionian Sea (Seritti et al. 2003) and across the entire EMS in both the surface layers and deep waters (Santinelli et al. 2010; Pujo-Pay et al. 2011) (Table 4.1). These DOC concentrations are only slightly lower than values measured in the subtropical gyres of the N.Atlantic and the N.Pacific (Hansell 2002) which are similar to the concentrations measured in the N.W. Mediterranean (Raimbault et al. 1999a). Likewise the dissolved organic nitrogen (DON) values are relatively constant across the EMS and similar to values in other oligotrophic regions of the world’s ocean (Table 4.2). It is somewhat surprising that the DOC and DON contents are as high as those determined in other sub-tropical areas since the annual primary production rates in the EMS are much lower (Psarra et al. 2000; Siokou-Frangou et al. 2010) . A likely contributory factor to this is that while DOC and DON produced during respiration and grazing of phytoplankton is probably low in the EMS, their consumption by heterotrophic bacteria is likely also to be low because the bacteria are strongly P limited (Thingstad et al. 2005). Sufficient data are not available yet to determine whether there is a measurable temporal trend in DOC and/or DON in surface waters as shown in the Sargasso Sea (BATS station, http://bats.bios.edu/) and as predicted by a I-D nutrient flux-reservoir model by Powley et al. (2012)
By contrast, the dissolved organic phosphorus (DOP) concentration in surface waters of the EMS (40–60 nmol kg−1) is considerably lower than values obtained in both the Sargasso Sea (20–500 nmol kg−1) and the N.Pacific Subtropical Gyre (150–270 nmol kg−1; Table 4.3) . Despite this, DOP (dissolved organic phosphorus) is the largest reservoir of phosphorus in the surface water column of the EMS compared to a maximum of a few nanomoles of DIP and < 18 nmol kg−1 of particulate P (Table 4.4). This residual DOP was considered biologically unreactive during the CYCLOPS experiment (Thingstad et al. 2005; Zohary et al. 2005) because there was only limited primary production even though N was present as ammonium (~80 nmol kg−1).
However, it is becoming increasingly evident that inorganic and organic P species other than the traditional ortho-phosphate can be utilized by phytoplankton and bacteria to sustain productivity, in particular in oligotrophic seas (Mather et al. 2008; Ranhofer et al. 2009; Diaz et al. 2008; Dyhrman et al. 2006). A measure of the degree to which DOP is utilized by micro-organisms is the activity of the alkaline phosphatase (APA), an enzyme that can hydrolyze P-esters to phosphate (Thingstad and Mantoura 2005; Ranhofer et al. 2009). The measured activity of APA in the Levantine Basin in May 2002 was 2.2 nM P h−1 (Thingstad and Mantoura 2005), similar to the APA activity measured at BATS, in summer and fall (Lomas et al. 2010) but more than 10 times the values observed in the North and South Pacific Gyres (Duhamel et al. 2010) which, though oligotrophic, are not particularly P limited. Further studies are needed to understand the nature and bioavailability of DOP in the EMS in comparison with other less P starved low productivity waters.
Biogeochemical and Nutrient Cycling Processes
Processes Related to the Unusually High N:P Ratio in the EMS
Nitrate: Phosphate Ratio in the Deep Water
One of the first unusual features to be recognised in the EMS was the high nitrate to phosphate ratio in deep waters of the Eastern Mediterranean, much higher than the Redfield ratio of 16 (Redfield et al. 1963). High N: P molar ratios in the deep waters were measured by Krom et al. (1991) – N: P = 28 ± 3:1; Civitarese et al. (1998) – N: P = 25–27:1; Yilmaz and Tugrul (1998) – N: P = 27–28.5:1; Krasakopoulou et al. (1999) – N: P = 24–36:1; and Kress and Herut (2001) – N: P = 25 ± 3:1, Kress et al. (2003) – 26–31:1, among others. These values are higher than those in the Western Mediterranean (23:1) and almost double the values found in the Eastern North Atlantic (16:1) and almost everywhere else in the global ocean.
Various hypotheses had been put forward to explain the unusually high ratio including high rates of nitrogen fixation, adsorption of phosphate onto Saharan dust, preferential remineralisation of P over N, nitrate enriched waters forming the ADW. The high N:P ratio in the EMS has now been explained as being due to high N:P ratio in the external sources of nutrients (atmospheric and riverine) (Table 4.5) combined with low rates of denitrification in the EMS (Krom et al. 2004, 2010). This is caused by the ultraoligotrophic status of the basin which results in few areas having enough residual OM to cause the sediment to become anoxic and hence to enable microbial denitrification to take place.
The Eastern Mediterranean as a P Starved System
Recent data has shown that it is not only the major dissolved inorganic nutrients which have a ratio of > 16:1 but all nutrient species (particulate and organic) have N: P ratios far in excess of Redfield (Table 4.4). The DOC:DON:DOP ratios in the photic zone is 106: 8–9: 0.06–0.07, which is depleted in N and highly depleted in P relative to the Redfield ratio (Krom et al. 2005; Pujo-Pay et al. 2011) . The POC:PON:POP ratio of particulate matter measured within the photic zone, a combination of live and dead cells and faecal pellets, is 106:10.3–11.3:0.28–0.47, much greater than Redfield’s, indicating nutrient depletion in particulate matter as well. High carbon to nutrient ratios are not uncommon in ultra-oligotrophic systems where carbon fixation by primary producers continues after N and P become totally depleted (e.g. Berman-Frank and Dubinsky 1999) and rapid recycling occurs. However the very high N:P ratios observed in the photic zone of the EMS combined with higher release of N than P below the photic zone was interpreted by Krom et al. (2005) as evidence of the rapid and very efficient recycling of P within the photic zone compared to that of N as seen in other areas such as the N. Pacific (Loh and Bauer 2000). They concluded that DOP is preferentially mineralised compared to DOC and DON resulting in increased C:P and N:P ratios with depth. In the EMS, by contrast, there is a much smaller decrease in DOP with depth than for either DON or DOC . This implies that in the EMS, DOP is recycled very efficiently in the photic zone and at lower depths the DOP is too refractory to break down much.
Seasonal Changes in Primary Production and Nutrient Limitation
The high N:P ratio was the basis of the hypothesis that the productivity of the Eastern Mediterranean was simply limited by the availability of phosphorus. However, recent results have shown that the nutrient limitation in the Eastern Mediterranean is more complex than it at first appeared and that there are seasonal differences in the ultimate nutrient limiting specific phytoplankton groups as well as different members of the surface microbial community (Thingstad et al. 2005; Zohary et al. 2005; Tanaka et al. 2011).
The annual major phytoplankton bloom in the EMS starts as soon as deep winter mixing occurs in early winter (e.g. December – Fig. 4.5) and nutrients are supplied into the photic zone. The bloom continues and increases in intensity throughout the winter months as the depth of mixing increases adding more nutrients until early spring, typically March in the SE. Levantine when the seasonal thermocline begins to form (Hecht et al. 1988). This unusual pattern is probably controlled by the typical weather pattern in the region which consists of a few days of cold often wet weather which causes the upper layers of the water column to mix, followed by several days of relatively warmer clear weather which causes a short term water column stability and allows the phytoplankton to bloom thereby exploiting the nutrients mixed into the surface waters.

Monthly SeaWiFS “climatological” images of chlorophyll over the seasonal cycle. The images are based on input data from Sept 1997 to August 2004 and chl-a was computed using the Bricaud et al. (2002) algorithm for the Mediterranean (Groom et al. 2005). The images were provided by Steve Groom (NERC Earth Observation Data Acquisition and Analysis Service, Plymouth)
This winter phytoplankton bloom is conventionally P limited. Deep winter mixing advects water with an N:P ratio >16:1 to the photic zone (e.g. Krom et al. 1992) containing nutrient concentrations typical of LIW (Levantine intermediate water) that is on the order of 0.03 μmolP kg−1 (Kress and Herut 2001) and between 1.3 and 2.0 μmolN kg−1 (Schlitzer et al. 1991) i.e. with an N:P ratio of 40–60:1. The bloom ceases as soon as the waters in the photic zone run out of the phosphate (Fig. 4.6). Characteristically this occurs when there is 0.3–1 μmolesN kg−1 of nitrate remaining in the surface waters (Krom et al. 1992; Kress and Herut 2001). This residual nitrate has been shown to be isotopically heavy (16–40 ‰) compared with 3–12 ‰ at depth (Emeis et al. 2010; Struck et al. 2001). This pattern of heavier N isotopes is characteristic of water in which the phytoplankton runs out of the limiting factor, in this case phosphate, before all the nitrate has been taken up by biological productivity. Additional biological evidence for P-limitation of the surface waters of the Eastern Mediterranean has been obtained from a series of observations on phytoplankton activity and specific microcosm experiments (Vukadin and Stojanovski 1976; Becacos-Kontos 1977; Pojed and Kveder 1977; Berland et al. 1980; Bonin et al. 1989). Zohary and Robarts (1998) also showed that in winter, bacteria from various sites in the EMS are also P-Limited.

Nutrient distribution during simple P-limited phytoplankton bloom in winter (February 1989) in the warm core Cyprus eddy showing that all the phosphate has been consumed leaving residual nitrate in the water column (Modified from Krom et al. 1992)
Starting generally in March/April a seasonal thermocline develops firstly in the SE. Levantine which then expands to the north and west (Fig. 4.5). As this happens, the primary productivity and plankton biomass is reduced in the uppermost layers and becomes concentrated at the top of the nutricline in a deep chlorophyll maximum (DCM). The DCM, which is characteristic of the summer in the EMS (April to November) is approximately the same depth (100–120 m) across the basin with decreasing concentrations of chlorophyll (a proxy to total phytoplankton biomass) and concurrent decrease in nutrients (Fig. 4.7). However this pattern is not simple because of the complex mesoscale circulation in the EMS.

Vertical section of in situ chl-a fluorescence (arbitrary units) across the Eastern Mediterranean sampled in April-May 1999. Stations and data points are indicated by filled dots (Adapted from Kress et al. 2011)
By late spring and early summer (May-June), the phytoplankton production has become N & P co-limited (Tanaka et al. 2011) although heterotrophic bacterial growth is still P limited (Thingstad et al. 2005). At that time, the seasonal thermocline is present, a deep chlorophyll maximum has developed, and the surface waters have become depleted in nitrate as well as phosphate (e.g. Kress and Herut 2001; Krom et al. 2005). This change in phytoplankton nutrient limitation in May was observed and identified by microcosm experiments carried out in the framework of the CYCLOPS P addition Lagrangian study (Krom et al. 2005; Zohary et al. 2005). In the microcosm experiment (Fig. 4.8) surface waters preconditioned with phosphate added to a patch of water (IN) and water from outside the patch (control) were incubated with 1.6 μmol kg−1 of ammonium. The results showed the classic behaviour of an N & P co-limited system with the only large increase in Chl-a concentrations in the treatment where ammonium was added to seawater preconditioned with phosphate. This change in nutrient limitation was explained by Thingstad et al. (2005). At the end of winter mixing, the surface water has residual nitrate and no free phosphate (see Fig. 4.6). After stratification develops, the phytoplankton in the water column above the nutricline receives very limited amounts of new nutrients and most of the production is sustained by recycled nutrients produced by respiration, grazing and biological oxidation of organic matter (PON – particulate organic nitrogen -, and POP – particulate organic phosphorus). The recycled nutrients include ammonium, nitrate, phosphate, that are directly available for phytoplankton uptake, and DON and DOP. While some of the DON compounds can be made available to phytoplankton, most of it cannot. Therefore the N available for phytoplankton growth decreases with recycling. The situation for P is different. When the particulate P is recycled there is no significant pool of non-bioavailable P formed i.e. if significant amounts of DOP are formed then it is either directly bioavailable to phytoplankton or after the action of enzymes such as APA. The surface waters have high activity of APA which is excreted by both bacteria and microphytoplankton to enable them to access part of the DOP pool (e.g. Thingstad and Mantoura 2005; Tanaka et al. 2011). As a result, more of the recycled P is available for phytoplankton uptake than N, eventually removing all of the ‘excess’ dissolved N (mainly nitrate) from the ‘winter excess’ and causing productivity to become N & P co-limited. Heterotrophic bacteria that are able to access the DON pool were P limited during the same time (Thingstad et al. 2005).

A shipboard microcosm experiment in which the effect of ammonium addition on phytoplankton growth as determined by changes in Chlorophyll-a concentration was followed using IN patch (phosphate enriched waters) and OUT patch (control waters). For more details see Zohary et al. (2005)
In the summer (June/July), similar nutrient limitation experiments were performed in the framework of the BOUM cruise (BOUM: Biogeochemistry from the Oligotrophic to the Ultra-oligotrophic Mediterranean) using phytoplankton collected from 20 m i.e. phytoplankton from the highly nutrient depleted waters well above the DCM (Tanaka et al. 2011). These results showed that although the organisms and system were P starved with DON:DOP values of 114–190:1 and PON:POP values of 24–28:1, there was only an increase in growth of osmotrophs when N + P or N only were added i.e. the system was no longer P limited. They suggested that as the stratified period progresses the pool of bioavailable N and P becomes very small in the surface layers and the most limiting nutrient for osmotrophic growth shifts sporadically among N, P and N&P (c.f. Hecky and Kilham 1988)
Is the Nutrient Uptake in the Surface Waters Redfieldian?
While the data is somewhat equivocal it seems that nutrient uptake in the EMS is often close to Redfieldian (i.e. 16:1). In a series of microcosm experiments carried out as part of the CYCLOPS programme, Kress et al. (2005) added ammonium and phosphate to EMS surface waters and obtained a nutrient uptake ratio of 18 ± 3.1. Zohary et al. (2005) subsequently carried out similar microcosm experiments in the core of the Cyprus Eddy and found an uptake ratio of 25:1. However these experiments were carried out by adding ammonium to surface water collected within artificially phosphate-enriched patch from the CYCLOPS addition experiment. Prior exposure to excess dissolved phosphate may have preconditioned the phytoplankton to take up nutrients in an N:P ratio exceeding 16:1. The nutrient budget calculations of Krom et al. (1992) yielded an N:P ratio for new production of phytoplankton in the core Cyprus eddy of 20:1. In another indirect estimate of the N:P ratio of nutrient uptake, assuming the nutrient supply to the photic zone contains concentrations typical of LIW (Kress and Herut 2001; Schlitzer et al. 1991) and that the phytoplankton uptake occurs in a Redfieldian ratio of 16;1, then the residual nitrate which would be left in the surface water would be 0.8–1 μmol kg−1, which agrees with the typical range of observed values in winter in the EMS (Kress and Herut 2001; Thingstad and Mantoura 2005). Thus phytoplankton growth in the surface water is approximately Redfieldian, certainly much lower than the 28:1 molar ratio measured in the Deep Water. However it has been suggested that P is more efficiently recycled from POM than N in surface (see section “Seasonal changes in primary production and nutrient limitation ”) and intermediate water. This results in the POM which reaches Deep Water having a N:P ratio higher than 16:1. Indeed the measurements made of POM in deep water by Krom et al. (2005) found N:P of 26:1 though Pujo-Pay et al. (2011) determined values close to 16:1 (Table 4.4). When this POM is broken down it releases more N than P.
In addition, there is an unusual (high) nutrient ratio of preformed nutrients in the EMS (Civitarese et al. 1998). It is known that ADW is formed in the S.Adriatic in winter simultaneously with the P limited major phytoplankton bloom i.e. when phosphate is entirely depleted and residual nitrate remains (see section “Seasonal changes in primary production and nutrient limitation”). The measured nitrate:phosphate ratio in the middle of the S.Adriatic in winter is ~33:1 (Zavatarelli et al. 1998) and this is the water advected into the deep Ionian Basin. The results of the 1-D flux reservoir model (Powley et al., 2012) confirm this explanation.
Nitrogen Fixation
Biological Dinitrogen (N2) fixation can contribute new N sources to N starved systems and significantly induce further nutrient uptake by subsequent blooms of primary producers. Early geochemical and isotopic models suggested that N2 fixation rates were high in the Mediterranean Sea (Bethoux and Copin-Montegut 1986, Bonin et al. 1989, Bethoux et al. 1992, Gruber and Sarmiento 1997, Pantoja et al. 2002, Sachs and Repeta 1999). Yet, actual measured rates utilizing the 15 N-uptake method (Montoya et al. 1996) are now accumulating from throughout the Mediterranean Basin bridging spatial and temporal scales (Garcia et al. 2006, Rees et al. 2006, Sandroni et al. 2007, Bar Zeev et al. 2008, Marty et al. 2008, Ibello et al. 2010, Yogev et al. 2011, Bonnet et al. 2011, Rahav et al. 2013a, b). The published N2 fixation rates from the Western Basin are typically higher both volumetrically and areally than those measured from the Eastern Basin during all measured seasons (Berman-Frank and Rahav 2012, Rahav et al. 2013a, b). Volumetric rates ranged from non-detected to 0.3 nmol N L−1 day−1 in the Eastern Basin, compared to non-detected to 17 nmol N L−1 day−1 in the Western Basin (Fig 4.9a). Areal integrated rates changed spatially and temporally with rates in the Western Basin ranging from > 3 to 50 μmol N m−2 day−1 (Sandroni et al. 2007, Ibello et al. 2010), compared with usually less than 10 μmol N m−2 day−1 for the Eastern Basin (Berman-Frank and Rahav 2012; Yogev et al. 2011; including a coastal Turkish station with rates of 4 μmol N m−2 day−1 (Ibello et al. 2010) and 0–0.4 μmol N m−2 day−1 during summertime in an anticyclonic eddy in the Ionian Sea (Bonnet et al. 2011)). Interestingly, low rates of two studies from the EMS are accompanied by the observations that ~35 % of all samples taken (accounting for spatial and temporal samplings) were below the detection limit (Ibello et al. 2010, Yogev et al. 2011). The contribution of N2 fixation to primary productivity in the EMS is extremely low with a maximum of 5 % (Fig. 4.9b) compared to the WMS, where N2 fixation comprised up to 16.5 % of primary productivity (Fig. 4.9b).

Summary depth distribution of measured volumetric rates of N2 fixation measured across the Mediterranean Sea (a) and the percentage contribution of the fixed N to primary productivity (b). Data were compiled from Yogev et al. 2011, Rahav et al. (2013a, b) (EMS), Ibello et al. (2010), Bonnet et al. (2011) (EMS + WMS), Rees et al. (2006), Garcia et al. (2006), Sandroni et al. (2007), Marty et al. (2008) (WMS)
The low measured rates of N2 fixation correspond with a taxonomically diverse yet limited abundance of diazotrophic organisms found throughout the EMS including unicellular representatives, heterotrophic diazotrophs, filamentous, and symbiotic associations (Man-Aharonovich et al. 2007; Bar-Zeev et al. 2008; Yogev et al. 2011; Bonnet et al. 2011). Of the few conspicuous diazotrophs in the Eastern Mediterranean Sea, the diatom-cyanobacterial associations (Rhizosolenia-Richelia, Hemiaulus-Richelia) were consistently present (10–55 heterocysts L−1 maximal abundance) in two routinely monitored stations off the Israeli coast (Bar Zeev et al. 2008). Despite the microscopic and molecular evidence no blooms of Hemiaulus-R. intracellularis were observed in over 8 years of monitoring and sampling in the EMS and in the above stations (personal observation; Bar Zeev et al. 2008).
Conspicuously rare is the appearance of the globally important filamentous non-heterocystous Trichodesmium in the Mediterranean Sea. In the EMS two species of Trichodesmium (T. erythraeum and T. thiebautii) have been sporadically observed in extremely low abundance (<1 trichome per 1,000 L) and only one bloom (from the Aegean Sea) has been reported (Spatharis et al. 2012). The rarity of Trichodesmium in the Mediterranean is surprising since this organism is routinely observed in both coastal and open waters of the adjacent Red Sea (Stihl et al. 2001) and in the Atlantic Ocean (Capone et al. 1997). Limitation by bioavailable inorganic P may control Trichodesmium populations in the Mediterranean although Trichodesmium can utilize other forms of P such as phosphonates (Dyhrman et al. 2006).
The limited contributions of N2 fixation in the Mediterranean Sea and particularly in the EMS have been routinely attributed to P limitation (Bonnet et al. 2011, Ridame et al. 2011) as dinitrogen fixers require P for the energetically expensive process of N2 fixation (16 ATP per mole of N2 fixed) (Postgate 1998). However, this explanation is not always consistent with experimental results. While some P additions increased N2 fixation rates in an anticyclonic eddy of the EMS (BOUM Station. C) during the stratified summer (Ridame et al. 2011), at other times and stations P enrichments did not enhance N2 fixation (e.g. BOUM, Station B (Ridame et al. 2011); pelagic EMS (Yogev et al. 2011); and cyclonic and anticyclonic eddies (Rahav et al. 2013a). Furthermore, dust enrichment in microcosm experiments (high N and P in dust) did not show consistent trends with regards to N2 fixation in the EMS. A significant increase was observed at the anticyclonic Station C (Ridame et al. 2011), while N2 fixation was not elevated at the Ionian Sea (Station B, Ridame et al. 2011), and at either the Cyprus Eddy or the Rhodes Gyre during July 2009 (Rahav et al. 2013a). Moreover, the published studies do not report any significant correlations between N2 fixation rates and any of the physio-chemical parameters such as temperature, nutrient availability, or biological competition. The limited samplings, methodological underestimation of 15 N uptake (Mohr et al. 2010), or the generally low N2 fixation rates measured in the EMS that were near or below detection limits at times, constrain our current understanding of what parameters limit N2 fixation in the EMS.
Physics Induced Bio-chemical Processes
The EMT event provided the opportunity to investigate the effect of physical changes in circulation on chemical and biological depth distribution and processes. As mentioned in section “Cross basin distribution and how they have changed with time ”, the younger CSOW was introduced into the deep waters of the EMS and caused the older nutrient rich ADW to be uplifted to mid depths. In the Eastern Levantine Basin, from 2002 to 2010, this uplift increased the nitrate and phosphate concentrations at the base of the nutricline by 0.5 and 0.04 μmole kg−1, respectively (400–500 m). Concurrently, the DCM depth range widened and became shallower, with a slight increase in chl-a concentrations, most evident at the continental slope. The concentrations at the surface waters increased as well since March 2010 in the open sea and earlier (Sep 2008) in the surface waters at the continental slope. There was no concurrent trend of increase in nitrate and phosphate at the DCM, but silicic acid concentrations decreased at the upper layers. Therefore, it is reasonable to assume that the increase in concentration at the base of the nutricline increased the supply of nutrients to the ultra-oligotrophic photic zone, where they were rapidly utilized by the phytoplankton. It is possible that the increased supply of nitrate and phosphate from below changed the phytoplankton community structure in the upper layers, increasing the relative contribution of diatoms and hence depleting silicic acid. However, there are no data on the phytoplankton community from this time series that can directly support this hypothesis.
Basin-wide oceanographic transects conducted in 1991 (Yacobi et al. 1995), 2001 (Kress et al. 2011), and 2008 did not show a clear trend in Chl-a concentrations, nor did satellite imagery analysis up to 2002 (D’Ortenzio et al. 2003). In October 1991, Chl-a ranged from 0.01 to 0.25 μg l−1 (depth integrated chlorophyll ranged from 17 to 35 mg m−2 with higher values in the Western Levantine). In October 2001, concentrations ranged from 0.01 to 0.28 μg l−1 (integrated values of 16.5–23.1 mg m−2) while in September 2008, from 0.01 to 0.33 μg l−1 (depth integrated 10–27.1 mg m−2). It is evident that the occasional basin-wide snapshots are not enough to indicate trends that may be cyclic and short lived. High frequency surveys are needed if we are to identify long term trends in this important parameter. However it is clear that despite large changes in external nutrient supply (see section “Changes in the nutrient supply from 1950 to 2000 ”), there have been no equivalent large changes in chlorophyll content or any other regional measurement of phytoplankton productivity.
Use of Nutrient Budgets to Define Biogeochemical Processes
Oceanic, Terrestrial and Atmospheric Nutrient Inputs
The Mediterranean in general and the Eastern Basin in particular represent almost closed systems in which total nutrient budgets have been used successfully to explain and quantify the relative importance of various biogeochemical processes within the system (e.g. Ribera d’Alcalà et al. 2003; Krom et al. 2004; Béthoux et al. 2002). The most recent total nutrient budget of the Eastern Basin first described by Krom et al. (2004, 2010) has been updated here (Table 4.5) to include more recent atmospheric inputs to the basin. The model output is balanced (~10 % error) for total N with 200 ×109 molesN/year imported to the basin and 179 ×109 molesN/year exported mainly through flux of nutrients with intermediate water through the Straits of Sicily. The major conclusion drawn from this budget was that N2 fixation is not significant in the EMS contrary to previous suggestions (Bethoux et al. 1986, 2002; Pantoja et al. 2002; Rees et al. 2006 and others) . This conclusion was confirmed subsequently by in-situ measurements (Bar-Zeev et al. 2008; Ibello et al. 2010; Yogev et al. 2011; Bonnet et al. 2011; Rahav et al. 2013a, b) (see section “Nitrogen fixation ”)
There are currently two problems with this budget : the phosphate budget does not balance and the external input of nutrients does not change with time. Considerably more P is exported from the basin (5.4 × 109 molesP/year) than is input (2.3 × 109 molesP/year). The export values are rather well constrained since they were calculated from measured values for dissolved phosphate at the Straits of Sicily and total P deposition rates in the sediment. By contrast the values for the input of bioavailable P is far less known. It is possible that both atmospheric P and riverine P fluxes do not take into account chemical species which become bioavailable within the basin. Thus the atmospheric input measured by Kocak et al. (2010) is only the leachable P and does not include any non-leachable P such as iron-bound P and apatite which are known to be present in Saharan dust (Nenes et al. 2011) and which might be bioavailable under the P starved conditions of the EMS water column. Likewise, the riverine input was calculated from dissolved inorganic P alone and does not include any particulate P or organic P which are brought down the rivers and might subsequently be converted to bioavailable P in the adjacent water column and sediment. DOP comprises an additional possible source of bioavailable P not included in the total budget. DOP is somewhat higher in surface waters (i.e. incoming waters) than in the outflowing LIW. However there are no data available for DOP fluxes through the Straits of Sicily nor is there any information of the relative bioavailability of surface vs intermediate water DOP. In a flux-reservoir model designed to quantitatively model the biogeochemical cycling in the EMS, van Cappellen et al. (2012) increased the P inputs by adding DOP which come close to closing the P budget.
Changes in External Nutrient Fluxes into the EMS with Time
Changes in the Nutrient Supply from 1950 to 2000
The other problem with the total nutrient budgets calculated thus far is that they do not take into account the changing fluxes of important parameters with time. It is known that the external supply of nutrients (N & P) from both rivers and atmospheric inputs to the EMS has increased significantly over the past century, particularly since 1960 as a result of anthropogenic inputs.
Thus the total N and P inputs from riverine sources to the EMS have been calculated by Ludwig et al. (2009) from 1963 to 1998 (Fig. 4.10). These data show a systematic increase in total N to the basin with the flux increasing by a factor of 5. The changes in riverine P are more complex. There was an increase in P through 1965 followed by a sharp decrease caused by the effect of the completion of the Aswan dam stopping the flow of dissolved and particle borne P into the S.E. Levantine Basin (Ludwig et al. 2009). Over the next 15 years, up to 1980, P increased as a result of anthropogenic discharges into rivers from sewage and other sources. However from the late 1980s to the present day, P fluxes into the basin have decreased as a result of abatement procedures designed to reduce the effect of P pollution within the rivers.

Figure showing the relative changes in external nutrient input to the EMS from the beginning of the twentieth century to 2000. The riverine data is from Ludwig et al. (2009) extrapolated back to 1920 assuming the major rivers behave in a similar manner to the Rhine (Zobrist and Stumm 1981). The Atmospheric flux uses data from an Alpine ice core modified by modelling from Preunkert et al. (2003) and Fagereli et al. (2007). In order to calculate a total atmospheric input it was assumed that 2/3 of the N flux to EMS is NOX and 1/3 is NH4 based on measurements made in Crete (Kocak et al. 2010)
The changes in atmospheric N (NOx and NH4) have been estimated using measured data from alpine ice cores modelled to calculate atmospheric fluxes for NOx (Preunkert et al. 2003) and for NH4 (Fagerli et al. 2007). Pro-rating these data for the EMS using measured atmospheric flux data from Crete (Kocak et al. 2010) and Israel (Herut et al. 1999; Carbo et al. 2005), it has been possible to determine the changing flux of N since 1950. The changing flux of P is more difficult to estimate since no long term record has been developed. It is complicated by the fact that there is a large ‘natural’ source of P to the EMS in the form of Saharan dust which contains an unknown proportion of bioavailable P. Recent data have shown that the fraction of bioavailable P in this dust has been modified by the effect of acidic atmospheric processes caused mainly by anthropogenic inputs from Southern Europe (Nenes et al. 2011). Further work is needed to understand the nature and magnitude of these atmospheric processes and their effect on the supply of this key nutrient to the EMS.
What Was the Level of Nutrient Supply Prior to Anthropogenic Inputs?
There are no data available to directly determine the anthropogenic increases in nutrients in rivers flowing into the EMS prior to 1950. However calculations to determine the nutrient content of the ‘pristine’ Rhine suggests that there was significant nutrient pollution of the river already by 1950 and it is likely that there would be similar changes in rivers such as the Nile and Po flowing into the EMS. Powley et al. (2012) have used these values and estimated that the riverine inputs to the EMS were N = 2.0 × 109 MolesN/year and P = 0.095 × 109 MolesP/year (i.e. ~5 % and 10 % of the 2,000 values). The relative increase in external supply is shown in Fig. 4.10. The input flux of atmospheric supply for both N species has increased by 85 % for NOx and by 65 % for NH4 between 1910 and 20 and the end of the century.
Possible Factors Which Will Influence Future Changes in Nutrient Distribution
Future Circulation Changes That Might Affect Nutrient Distribution
The most dramatic change in circulation in the recent past has been the EMT event which has caused changes in nutrient distribution in the EMS. It is still not determined whether this is part of an on-going cyclic pattern of changes in the EMS which may be natural or as a result of anthropogenically caused environmental and/or climate change. At present the most striking temporal change observed in physical parameters has been the increase in salinity (from 39.3 to 39.6) at the surface of the Eastern Levantine that started by mid-2005, with no concurrent changes in temperature. Salinity has continued to increase within the upper layers (down to 300 m) until the end of 2010. Historical data has shown a cyclic pattern in salinity: lower salinities at the beginning of the 1980s and 2000s and higher salinities at the beginning of the 1990s (onset of the EMT) and since 2006. This salinity increase, identified at the easternmost part of the basin, should be followed and analyzed not only in the local context but basinwide, for it is assumed that a similar increase in salinity in the area may have conditioned the EMT event in the 1990s. Cyclic pattern changes have also been observed in the upper layer circulation in the Ionian Basin that changed from cyclonic to anti-cyclonic circulation and back (Civitarese et al. 2010; Gacic et al. 2010). Both these changes in circulation resulted in observed changes in nutrient distribution and it is likely that future changes will also modify nutrients and hence primary productivity.
Is the EMS Particularly Vulnerable to Climate and Environmental Change?
In a more general way, it is known that the nutrient status of the EMS has changed dramatically over the past 10 k years as a result of natural climate change. The evidence for this is found in sapropels deposited in the deeper waters (>500 m) of the EMS basin. The present ultra-oligotrophic conditions in the EMS result are characterised by calcareous marl sediments with very low concentrations of organic C (0.1–0.2 %) . However interbedded with these sediments are a series of organic C rich sapropel layers. The most recent of these sapropel layers (S-1) began at ~9.8 ky 14C BP and finished at ~5.7 ky 14C BP (de Lange et al. 2008). During the S-1 period there was both higher primary productivity and restricted circulation within the deeper water which resulted in sub-oxic to anoxic conditions. While the precise controls which gave rise to this dramatic change in trophic status in the EMS is still being debated, it is generally agreed that the climate during that period was more humid and river flow into the basin (e.g. the river Nile) was higher. These changes in climate resulted in both reduced water exchange through the Straits of Sicily and more restricted formation of deep water. The changes in circulation predicted as a result of modern climate change (e.g. Somot et al. 2006) may also have profound effects on the export rate of nutrients in the EMS and thus have the potential to result in drastic changes in the trophic status of the basin
References
Bar Zeev E, Yogev T, Man-Aharonovich D, Kress N, Herut B, Beja O, Berman-Frank I (2008) Seasonal dynamics of the endosymbiotic, nitrogen-fixing cyanobacterium Richelia intracellularis in the eastern Mediterranean sea. ISME J 2:911–923
Becacos-Kontos T (1977) Primary production and environmental factors in an oligotrophic biome in the Aegean sea. Mar Biol 42:93–98
Berland BR, Bonin DJ, Maestrini SY (1980) Azote ou phosphore? Considérations sur le “paradoxe nutritionnel” de la Mer Méditerranée. Oceanol Acta 3:135–141
Berman-Frank I, Dubinsky Z (1999) Balanced growth of photosynthesizing aquatic organisms: myth and reality. Bioscience 49:29–37
Berman-Frank I, Rahav E (2012) Ch. 8: Nitrogen fixation as a source for new production in the Mediterranean Sea: a review. In: Stambler NE (ed) Life in the Mediterranean Sea: a look at habitat changes. Nova Science Publishers, New York, pp 199–226
Bethoux JP, Copin-Montegut G (1986) Biological fixation of atmospheric nitrogen in the Mediterranean Sea. Limnol Oceanogr 31:1353–1358
Bethoux JP, Morin P, Madec C, Gentili B (1992) Phosphorus and nitrogen behavior in the Mediterranean Sea. Deep-Sea Res 39:1641–1654
Béthoux JP, Morin P, Ruiz-Pino DP (2002) Temporal trends in nutrient ratios: chemical evidence of Mediterranean ecosystem changes driven by human activity. Deep-Sea Res II 49:2007–2016
Bonin DJ, Bonin MC, Berman T (1989) Mise en evidence experimentale des facteurs nutritifs limitants de la production du micro-nanoplancton et de l’ultraplancton dans une eau cotiere de la Mediterranee orientale (Haifa Israel). Aquat Sci 51:129–159
Bonnet S, Grosso O, Moutin T (2011) Planktonic dinitrogen fixation along a longitudinal gradient across the Mediterranean Sea during the stratified period (BOUM cruise). Biogeosciences 8:2257–2267
Bricaud A, Bosc E, Antoine D (2002) Algal biomass and sea surface temperature in the Mediterranean Basin: intercomparison of data from various satellite sensors, and implications for primary production estimates. Remote Sens Environ 81:163–178
Capone DG, Zehr JP, Paerl HW, Bergman B, Carpenter EJ (1997) Trichodesmium, a globally significant marine cyanobacterium. Science 276:1221–1229
Carbo P, Krom MD, Homoky WB, Benning LG, Herut B (2005) Impact of atmospheric deposition on N and P geochemistry in the southeastern Levantine basin. Deep-Sea Res II 52:3041–3053
Cavender-Bares KK, Karl DM, Chisholm SW (2001) Nutrient gradients in the Western North Atlantic Ocean: relationship to microbial community structure and comparison to patterns in the Pacific Ocean. Deep-Sea Res I 48:2373–2395
Civitarese G, Gacic M, Vetrano A, Boldrin A, Bregant D, Rabitti S, Souvermezoglou E (1998) Biogeochemical fluxes through the Strait of Otranto (Eastern Mediterranean). Cont Shelf Res 18:773–789
Civitarese G, Gacic M, Lipizer M, Borzelli GLE (2010) On the impact of the Bimodal Oscillating System (BiOS) on the biogeochemistry and biology of the Adriatic and Ionian seas (Eastern Mediterranean). Biogeosciences 7:3987–3997
D’Ortenzio F, Ragni M, Marullo S, Ribera d’Alcalà M (2003) Did biological activity in the Ionian sea change after the eastern Mediterranean Transient? Results from the analysis of remote sensing observations. J Geophys Res-Oceans 108:8113
de Lange GJ, Thomson J, Reitz A, Slomp CP, Principato MS, Erba E, Corselli C (2008) Synchronous basin-wide formation and redox-controlled preservation of a Mediterranean sapropel. Nat Geosci 1:606–610
Diaz J, Ingall E, Benitez-Nelson C, Paterson D, de Jonge MD, McNulty I, Brandes JA (2008) Marine polyphosphate: a key player in geologic phosphorus sequestration. Science 320:652–655
Duhamel S, Dyhrman ST, Karl DM (2010) Alkaline phosphatase activity and regulation in the North Pacific Subtropical Gyre. Limnol Oceanogr 55:1414–1425
Dyhrman ST, Chappell PD, Haley ST, Moffett JW, Orchard ED, Waterbury JB, Webb EA (2006) Phosphonate utilization by the globally important marine diazotroph Trichodesmium. Nature 439:68–71
Emeis KC, Mara P, Schlarbaum T, Möbius J, Dähnke K, Struck U, Mihalopoulos N, Krom M (2010) External N inputs and internal N cycling traced by isotope ratios of nitrate, dissolved reduced nitrogen, and particulate nitrogen in the eastern Mediterranean sea. J Geophys Res 115, G04041
Fagerli H, Legrand M, Preunkert S, Vestreng V, Simpson D, Cerqueira M (2007) Modeling historical long-term trends of sulfate, ammonium, and elemental carbon over Europe: a comparison with ice core records in the Alps. J Geophys Res 112:D23S13
Gacic M, Borzelli GLE, Civitarese G, Cardin V, Yari S (2010) Can internal processes sustain reversals of the ocean upper circulation? The Ionian sea example. Geophys Res Lett 37, L09608
Garcia N, Raimbault P, Gouze E, Sandroni V (2006) Nitrogen fixation and primary production in western Mediterranean. C R Biol 329:742–750
Groom S, Herut B, Brenner S, Zodiatis G, Psarra S, Kress N, Krom MD, Law CS, Drakopoulos P (2005) Satellite-derived spatial and temporal biological variability in the Cyprus Eddy. Deep-Sea Res II 52:2990–3010
Gruber N, Sarmiento JL (1997) Global patterns of marine nitrogen fixation and denitrification. Global Biogeochm Cycle 11:235–266
Hansell DA (2002) DOC in the global ocean carbon cycle. In: Hansell DA, Carlson CA (eds) Biogeochemistry of marine dissolved organic matter. Academic, San Diego
Hansell DA, Carlson CA (2001) Biogeochemistry of total organic carbon and nitrogen in the Sargasso sea: control by convective overturn. Deep-Sea Res II 48:1649–1667
Hecht A, Pinardi N, Robinson AR (1988) Currents, water masses, eddies and jets in the Mediterranean Levantine Basin. J Phys Oceanogr 18:1320–1353
Hecky RE, Kilham P (1988) Nutrient limitation of phytoplankton in freshwater and marine environments: a review of recent evidence on the effects of enrichment. Limnol Oceanogr 33:796–822
Herut B, Zohary T, Robarts RD, Kress N (1999) Adsorption of dissolved phosphate onto loess particles in surface and deep eastern Mediterranean water. Mar Chem 64:253–265
Ibello V, Cantoni C, Cozzi S, Civitarese G (2010) First basin-wide experimental results on N2 fixation in the open Mediterranean Sea. Geophys Res Lett 37, L03608, doi:10.1029/2009GL041635
Klein B, Roether W, Manca BB, Bregant D, Beitzel V, Kovacevic V, Luchetta A (1999) The large deep water transient in the Eastern Mediterranean. Deep-Sea Res I 46:371–414
Klein B, Roether W, Kress N, Manca BB, d’Alcala MR, Souvermezoglou E, Theocharis A, Civitarese G, Luchetta A (2003) Accelerated oxygen consumption in eastern Mediterranean deep waters following the recent changes in thermohaline circulation. J Geophys Res Oceans 108:8107
Kocak M, Kubilay N, Tugrul S, Mihalopoulos N (2010) Long-term atmospheric nutrient inputs to the eastern Mediterranean: sources, solubility and comparison with riverine inputs. Biogeosciences 7:4037–4050
Koike I, Tupas L (1993) Total dissolved nitrogen in the northern North Pacific assessed by a high-temperature combustion method. Mar Chem 41:209–214
Krasakopoulou E, Souvermezoglou E, Pavlidou A, Kontoyannis H (1999) Oxygen and nutrient fluxes through the Straits of the Cretan Arc (March 1994–January 1995). Prog Oceanogr 44:601–624
Kress N, Herut B (2001) Spatial and seasonal evolution of dissolved oxygen and nutrients in the southern Levantine Basin (eastern Mediterranean sea): chemical characterization of the water masses and inferences on the N : P ratios. Deep-Sea Res I 48:2347–2372
Kress N, Manca BB, Klein B, Deponte D (2003) Continuing influence of the changed thermohaline circulation in the eastern Mediterranean on the distribution of dissolved oxygen and nutrients: physical and chemical characterization of the water masses. J Geophys Res Oceans 108:8109
Kress N, Herut B, Gertman I (2011) Nutrient distribution in the Eastern Mediterranean before and after the transient event. In: Stambler N (ed) Life in the Mediterranean Sea: a look at habitat changes. Nova, New York
Kress N, Thingstad TF, Pitta P, Psarra S, Tanaka T, Zohary T, Groom S, Herut B, Mantoura RFC, Polychronaki T, Rassoulzadegan F, Spyres G (2005) Effect of P and N addition to oligotrophic Eastern Mediterranean waters influenced by near-shore waters: a microcosm experiment. Deep-Sea Res II 52:3054–3073
Krom MD, Kress N, Brenner S, Gordon LI (1991) Phosphorus limitation of primary productivity in the eastern Mediterranean sea. Limnol Oceanogr 36:424–432
Krom MD, Brenner S, Kress N, Neori A, Gordon LI (1992) Nutrient dynamics and new production in a warm-core eddy from the eastern Mediterranean sea. Deep-Sea Res A 39:467–480
Krom MD, Groom S, Zohary T (2003) The Eastern Mediterranean. In: Black KD, Shimmield GB (eds) The biogeochemistry of marine systems. Blackwell Publishing, Oxford
Krom MD, Herut B, Mantoura RFC (2004) Nutrient budget for the eastern Mediterranean: implications for phosphorus limitation. Limnol Oceanogr 49:1582–1592
Krom MD, Woodward EMS, Herut B, Kress N, Carbo P, Mantoura RFC, Spyres G, Thingstad TF, Wassmann P, Wexels-Riser C, Kitidis V, Law CS, Zodiatis G (2005) Nutrient cycling in the south east Levantine basin of the eastern Mediterranean: results from a phosphorus starved system. Deep-Sea Res II 52:2879–2896
Krom MD, Emeis KC, Van Cappellen P (2010) Why is the eastern Mediterranean phosphorus limited? Prog Oceanogr 85:236–244
Lascaratos A, Roether W, Nittis K, Klein B (1999) Recent changes in deep water formation and spreading in the eastern Mediterranean sea: a review. Prog Oceanogr 44:5–36
Lomas MW, Burke AL, Lomas DA, Bell DW, Shen C, Dyhrman ST, Ammerman JW (2010) Sargasso sea phosphorus biogeochemistry: an important role for dissolved organic phosphorus (DOP). Biogeosciences 7:695–710
Loh AN, Bauer JE (2000) Distribution, partitioning and fluxes of dissolved and particulate organic C, N and P in the eastern North Pacific and Southern Oceans. Deep-Sea Res 47:2287–2316
Ludwig W, Dumont E, Meybeck M, Heussner S (2009) River discharges of water and nutrients to the Mediterranean and Black sea: major drivers for ecosystem changes during past and future decades? Prog Oceanogr 80:199–217
Man-Aharonovich D, Kress N, Bar Zeev E, Berman-Frank I, Beja O (2007) Molecular ecology of nifH genes and transcripts in the eastern Mediterranean Sea. Environ Microbiol 9:2354–2363.
Malanotte-Rizzoli P, Manca BB, d’Alcala MR, Theocharis A, Brenner S, Budillon G, Ozsoy E (1999) The eastern Mediterranean in the 80s and in the 90s: the big transition in the intermediate and deep circulations. Dynam Atmos Oceans 29:365–395
Manca BB, Kovacevic V, Gacic M, Viezzoli D (2002) Dense water formation in the southern Adriatic sea and spreading into the Ionian sea in the period 1997–1999. J Marine Syst 33–34:133–154
Manca BB, Ibello V, Pacciaroni M, Scarazzato P, Giorgetti A (2006) Ventilation of deep waters in the Adriatic and Ionian seas following changes in thermohaline circulation of the eastern Mediterranean. Climate Res 31:239–256
Marty JC, Garcia N, Rairnbault P (2008) Phytoplankton dynamics and primary production under late summer conditions in the NW Mediterranean Sea. Deep-Sea Res I 55:1131–1149
Mather RL, Reynolds SE, Wolff GA, Williams RG, Torres-Valdes S, Woodward EMS, Landolfi A, Pan X, Sanders R, Achterberg EP (2008) Phosphorus cycling in the north and south Atlantic ocean subtropical gyres. Nat Geosci 1:439–443
Montoya JP, Voss M, Kahler P, Capone DG (1996) A simple, high-precision, high-sensitivity tracer assay for N2 fixation. Appl Environ Microbiol 62:986–993
Nenes A, Krom MD, Mihalopoulos N, Van Cappellen P, Shi Z, Bougiatioti A, Zarmpas P, Herut B (2011) Atmospheric acidification of mineral aerosols: a source of bioavailable phosphorus for the oceans. Atmos Chem Phys 11:6265–6272
Pantoja S, Repeta DJ, Sachs JP, Sigman DM (2002) Stable isotope constraints on the nitrogen cycle of the Mediterranean Sea water column. Deep-Sea Res I 49:1609–1621
POEM-Group (1992) General circulation of the Eastern Mediterranean. Earth Sci Rev 32:285–309
Pojed I, Kveder S (1977) Investigation of nutrient limitation of phytoplankton production in northern Adriatic. Thalass Jugosl 13:13–27
Postgate J (1998) The origins of the unit of nitrogen fixation at the University of Sussex. Notes Rec R Soc 52:355–362
Preunkert S, Wagenbach D, Legrand M (2003) A seasonally resolved alpine ice core record of nitrate: comparison with anthropogenic inventories and estimation of preindustrial emissions of NO in Europe. J Geophys Res 108:4681
Psarra S, Tselepides A, Ignatiades L (2000) Primary productivity in the oligotrophic Cretan sea (NE Mediterranean): seasonal and interannual variability. Prog Oceanogr 46:187–204
Pujo-Pay M, Conan P, Oriol L, Cornet-Barthaux V, Falco C, Ghiglione JF, Goyet C, Moutin T, Prieur L (2011) Integrated survey of elemental stoichiometry (C, N, P) from the Western to Eastern Mediterranean Sea. Biogeosciences 8:883–899
Rahav E, Herut B, Stambler N, Bar-Zeev E, Mulholland MR, Berman-Frank I (2013a) Uncoupling between dinitrogen fixation and primary productivity in the Eastern Mediterranean Sea. JGR Biogeosci 18:195–202
Rahav E, Herut B, Levi A, Mulholland MR, Berman-Frank I (2013b) Springtime contribution of dinitrogen fixation to primary production across the Mediterranean. Ocean Sci 9:1–10
Raimbault P, Diaz F, Pouvesle W, Boudjellal B (1999a) Simultaneous determination of particulate organic carbon, nitrogen and phosphorus collected on filters, using a semi-automatic wet-oxidation method. Marine Ecol-Progr Ser 180:289–295
Raimbault P, Slawyk G, Boudjellal B, Coatanoan C, Conan P, Coste B, Garcia N, Moutin T, Pujo-Pay M (1999b) Carbon and nitrogen uptake and export in the equatorial Pacific at 150°W: evidence of an efficient regenerated production cycle. J Geophys Res 104:3341–3356
Ranhofer M, Lawrenz E, Pinckney J, Benitez-Nelson C, Richardson T (2009) Cell-specific alkaline phosphatase expression by phytoplankton from Winyah Bay, South Carolina, USA. Estuar Coasts 32:943–957
Redfield AC, Ketchum BH, Richards FA (1963) The influence of organisms on the composition of sea-water. In: Hill MN (ed) The sea. Interscience, New York
Rees AP, Law CS, Woodward EMS (2006) High rates of nitrogen fixation during an in-situ phosphate release experiment in the Eastern Mediterranean Sea. Geophys Res Lett 33, L10607, doi:10.1029/2006GL025791
Ribera d’Alcalà M, Civitarese G, Conversano F, Lavezza R (2003) Nutrient ratios and fluxes hint at overlooked processes in the Mediterranean sea. J Geophys Res 108:8106
Ridame C, Le Moal M, Guieu C, Ternon E, Biegala IC, L’Helguen S, Pujo-Pay M (2011) Nutrient control of N2 fixation in the oligotrophic Mediterranean Sea and the impact of Saharan dust events. Biogeosciences 8:2773–2783
Robinson AR, Golnaraghi M, Leslie WG, Artegiani A, Hecht A, Lazzoni E, Michelato A, Sansone E, Theocharis A, Ünlüata Ü (1991) The eastern Mediterranean general circulation: features, structure and variability. Dynam Atmos Oceans 15:215–240
Roether W, Manca B, Klein B, Bregant D, Georgopoulos D, Beitzel V, Kovacevic V, Luchetta A (1996) Recent changes in eastern Mediterranean deep water. Science 271:333–335
Roether W, Klein B, Manca BB, Theocharis A, Kioroglou S (2007) Transient eastern Mediterranean deep waters in response to the massive dense-water output of the Aegean sea in the 1990s. Prog Oceanogr 74:540–571
Rubino A, Hainbucher D (2007) A large abrupt change in the abyssal water masses of the eastern Mediterranean. Geophys Res Lett 34, L23607
Sachs JP, Repeta DJ (1999) Oligotrophy and nitrogen fixation during eastern Mediterranean sapropel events. Science 286:2485–2488
Sandroni V, Raimbault P, Migon C, Garcia N, Gouze E (2007) Dry atmospheric deposition and diazotrophy as sources of new nitrogen to northwestern Mediterranean oligotrophic surface waters. Deep-Sea Res I 54:1859–1870
Santinelli C, Nannicini L, Seritti A (2010) DOC dynamics in the meso and bathypelagic layers of the Mediterranean sea. Deep-Sea Res II 57:1446–1459
Schlitzer R, Roether W, Oster H, Junghans HG, Hausmann M, Johannsen H, Michelato A (1991) Chlorofluoromethane and oxygen in the eastern Mediterranean. Deep-Sea Res A 38:1531–1551
Seritti A, Manca BB, Santinelli C, Murru E, Boldrin A, Nannicini L (2003) Relationships between dissolved organic carbon (DOC) and water mass structures in the Ionian sea (winter 1999). J Geophys Res 108:8112
Siokou-Frangou I, Christaki U, Mazzocchi MG, Montresor M, Ribera d’Alcalá M, Vaqué D, Zingone A (2010) Plankton in the open Mediterranean sea: a review. Biogeosciences 7:1543–1586
Smith SV, Kimmerer WJ, Walsh TW (1986) Vertical flux and biogeochemical turnover regulate nutrient limitation of net organic production in the north Pacific Gyre. Limnol Oceanogr 31:161–167
Somot S, Sevault F, Deque M (2006) Transient climate change scenario simulation of the Mediterranean Sea for the twenty-first century using a high-resolution ocean circulation model. Climate Dyn 27:851–879
Spatharis S, Skliris N, Meziti A, Kormas KA (2012) First record of a Trichodesmium erythraeum bloom in the Mediterranean Sea. Can J Fish Aquat Sci 69:1444–1455
Stihl A, Sommer U, Post AF (2001) Alkaline phosphatase activities among populations of the colony-forming diazotrophic cyanobacterium Trichodesmium spp. (cyanobacteria) in the Red Sea. J Phycol 37:310–317
Struck U, Emeis K-C, Voß M, Krom MD, Rau GH (2001) Biological productivity during sapropel S5 formation in the eastern Mediterranean sea: evidence from stable isotopes of nitrogen and carbon. Geochim Cosmochim Acta 65:3249–3266
Tanaka T, Thingstad TF, Christaki U, Colombet J, Cornet-Barthaux V, Courties C, Grattepanche JD, Lagaria A, Nedoma J, Oriol L, Psarra S, Pujo-Pay M, Van Wambeke F (2011) Lack of P-limitation of phytoplankton and heterotrophic prokaryotes in surface waters of three anticyclonic eddies in the stratified Mediterranean sea. Biogeosciences 8:525–538
Thingstad TF, Mantoura RFC (2005) Titrating excess nitrogen content of phosphorous-deficient eastern Mediterranean surface water using alkaline phosphatase activity as a bio-indicator. Limnol Oceanogr Methods 3:94–100
Thingstad TF, Krom MD, Mantoura RFC, Flaten GAF, Groom S, Herut B, Kress N, Law CS, Pasternak A, Pitta P, Psarra S, Rassoulzadegan F, Tanaka T, Tselepides A, Wassmann P, Woodward EMS, Riser CW, Zodiatis G, Zohary T (2005) Nature of phosphorus limitation in the ultraoligotrophic eastern Mediterranean. Science 309:1068–1071
Vukadin I, Stojanovski L (1976) C:N:Si:P ratio in the waters of the middle and south Adriatic. Commun Int ExplSci Mer Medit Rapp PV Reun 23:41–43
Williams PM, Carlucci AF, Olson R (1980) Deep profile of some biologically important properties in the Central North Pacific Gyre. Oceanol Acta 3:471–476
Wu J, Sunda W, Boyle EA, Karl DM (2000) Phosphate depletion in the western north Atlantic ocean. Science 289:759–762
Yacobi YZ, Zohary T, Kress N, Hecht A, Robarts RD, Waiser M, Wood AM (1995) Chlorophyll distribution throughout the southeastern Mediterranean in relation to the physical structure of the water mass. J Marine Syst 6:179–190
Yilmaz A, Tugrul S (1998) The effect of cold- and warm-core eddies on the distribution and stoichiometry of dissolved nutrients in the Northeastern Mediterranean. J Marine Syst 16:253–268
Yogev T, Rahav E, Bar-Zeev E, Man-Aharonovich D, Stambler N, Kress N, Béjà O, Mulholland MR, Herut B, Berman-Frank I (2011) Is dinitrogen fixation significant in the Levantine Basin, east Mediterranean sea? Environ Microbiol 13:854–871
Zavatarelli M, Raicich F, Artegiani A, Bregant D, Russo A (1998) Climatological biogeochemical characteristics of the Adriatic sea. J Marine Syst 18:227–263
Zobrist J, Stumm W (1981) Chemical dynamics of the Rhine catchment area in Switzerland, extrapolation to the ‘pristine’ Rhine river input to the ocean. In: Martin JM, Burton JD, Eisma D (eds) River inputs to ocean systems. United Nations, New York
Zohary T, Robarts RD (1998) Experimental study of microbial P limitation in the eastern Mediterranean. Limnol Oceanogr 43:387–395
Zohary T, Herut B, Krom MD, Fauzi C, Mantoura R, Pitta P, Psarra S, Rassoulzadegan F, Stambler N, Tanaka T, Frede Thingstad T, Malcolm S, Woodward E (2005) P-limited bacteria but N and P co-limited phytoplankton in the eastern Mediterranean – a microcosm experiment. Deep-Sea Res II 52:3011–3023
Acknowledgements
The ideas which are contained in this review have developed over many years as a result of conversations with many colleagues. While many of those interactions ended up as papers which have been referenced here, others were more informal. Our first scientific interaction with the Eastern Mediterranean was inspired by Hava Hornung, Tom Berman, Arthur Hecht and the POEM group. Since then MK has particularly appreciated the interactions with Frede Thingstad, Fauzi Mantoura, Barak Herut and the other members of the CYCLOPS team, and with Kay Emeis, Philippe van Cappellen and Helen Powley who together have developed a quantitative description of processes in the EMS.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2014 Springer Science+Business Media Dordrecht
About this chapter
Cite this chapter
Krom, M., Kress, N., Berman-Frank, I., Rahav, E. (2014). Past, Present and Future Patterns in the Nutrient Chemistry of the Eastern Mediterranean. In: Goffredo, S., Dubinsky, Z. (eds) The Mediterranean Sea. Springer, Dordrecht. https://doi.org/10.1007/978-94-007-6704-1_4
Download citation
DOI: https://doi.org/10.1007/978-94-007-6704-1_4
Published:
Publisher Name: Springer, Dordrecht
Print ISBN: 978-94-007-6703-4
Online ISBN: 978-94-007-6704-1
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)