
Abstract
Unlike immotile cilia, which protrude from most cells in our body, motile cilia are restricted to sperm cells and epithelial cells lining the airways, the oviduct, the paranasal sinuses, and the brain ventricles. The best-known function of these cilia is that their coordinated beating generates extracellular flow that clears mucus from the airways, moves ova from the oviducts toward the uterus, and propels cerebrospinal fluid (CSF) through the cerebral ventricles. The vertebrate brain forms around a ventricular cavity in which the CSF, secreted by the choroid plexus in each ventricle, flows continuously. The CSF, which contains many growth factors and morphogens, is present from the first stages of brain development and plays crucial roles throughout life. Ependymal cells are specialized glial cells that extend multiple motile cilia into the cerebral ventricles. These cells appear at early postnatal stages and line all cerebral ventricles in adult mammals. In this chapter, we will review current knowledge on ependymal motile cilia development, organization, functions, and their associated pathologies.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
7.1 Introduction
Motile cilia are microtubule-based structures protruding from the apical surface of epithelial cells in different organs such as the airways, the female reproductive tract and the brain. Each cilium is arranged in the classic 9 + 2 configuration (9 pairs of microtubules in the periphery and 2 pairs in the center) and coordinately beat to produce large-scale fluid flows crucial for the development and physiology of these organs. Indeed, genetic diseases in which ciliary beating is affected often leads to loss of ciliary flow linked to bronchiestasis and chronic sinusitis, ectopic pregnancies, hydrocephalus and defects in neurogenesis. In the brain, multiciliated ependymal cells line all ventricular walls in which CSF flows continuously thanks to the coordinated beating of their motile cilia. We discuss the main current knowledge on development and functions of brain motile cilia and ependyma cells.
7.2 The Brain Ventricular System
7.2.1 The Brain Ventricles
The vertebrate brain forms around a ventricular system composed of four interconnected cavities filled with cerebrospinal fluid (CSF): The two lateral ventricles located in the cerebrum are linked to the third ventricle located in the diencephalon via the interventricular foramen of Monro. The cerebral aqueduct links the third and fourth ventricle, which lies between the cerebellum and pons. The fourth ventricle joins the spinal cord canal and the subarachnoid space that envelops the brain via the left lateral aperture (foramen of Lushka) and the median aperture (foramen of Magendie and the spinal cord canal) (Fig. 7.1)

Drawing of a sagittal view of mammalian cerebral ventricular system. CSF is secreted (blue arrows) by the choroid plexuses (red) in each ventricle. The bulk CSF flows unidirectionally (yellow arrows) and into the suarachnoid space via the apertures (green arrows)
7.2.2 The Cerebrospinal Fluid (CSF)
The CSF is constantly produced by the choroid plexus in each ventricle, by filtering blood through choroidal capillaries and epithelial cells. The bulk CSF flows unidirectionally, from the lateral to the third and fourth ventricles and then out into the subarachnoid space surrounding the brain where it is absorbed into the hematopoietic circulatory and lymphatic systems. The driving force of the CSF bulk flow is actively supported by arterial pulsations and pressure gradients produced by secretion and absorption (Bering 1955; Wagshul et al. 2006). Inside each ventricle and depending on their size and shape (i.e. in large mammals like humans or sheep compared to rodents), the CSF may flow locally against the bulk flow, creating local turbulence. Along the ventricular walls, the laminar flow of CSF is mainly directed by the coordinated beating of ependymal cilia in the direction of the bulk flow (Nguyen et al. 2001; Fig. 7.1). The total volume of the CSF in humans is around 140 ml and constitutes 18% of the volume of an adult human brain: about 30 ml is inside the ventricles, the remainder surrounds the brain (Segal 2000). The CP produces around 500 ml of CSF per day in a young adult, suggesting that human CSF is completely replaced about three times every 24 h. In the young adult rat, which has about 200 μl CSF, it takes about 2 h to replace the CSF (Veening and Barendregt 2010). In a healthy normal individual, the CSF contains more than 2,600 different peptides and proteins (Schutzer et al. 2010). The CSF is vital to the health of the brain; it protects it from physical damage, facilitates the removal of metabolites and provides nutrients (Segal 2001). It also contributes to brain development, homeostasis, adult neurogenesis and repair by providing physical and chemical cues (Miyan et al. 2003).
7.2.3 Cells in Contact with the Cerebral Ventricles
7.2.3.1 Cells Extending Motile Cilia
-
Multiciliated ependymal cells (E1 cells) are the most abundant epithelial cells lining the lateral, third and fourth ventricles, and are CD24+, S100β+, FoxJ1+, Sox2+ and CD133+ but nestin− and GFAP−. These cells are cuboidal with a light cytoplasm and spherical uninvaginated nuclei containing dispersed chromatin. Abundant mitochondria are localized near the basal bodies (Doetsch et al. 1997; Mirzadeh et al. 2008). The lateral processes of adjacent ependymal cells are heavily interdigitated and contain apical adherens and tight junctional complexes and gap junctions. Ependymal cilia are motile and measure 8–15 μm in length (average size: 11.5 μm), protruding in the cerebral ventricles (Mirzadeh et al. 2008, 2010; Hirota et al. 2010; Fig. 7.2). The surface exposed to the ventricular cavity is variable in size (from 100 to 600 μm2, average size: 265 μm2) and contains a patch of 32–73 (mean: 49) basal bodies separated by 250 nm. In contrast to other epithelia containing motile cilia, in which basal bodies cover the entire apical surface, the basal body patch occupies only 4–35% of the total apical surface of the ependymal cells and is positioned downstream with respect to the direction of ciliary beating. In the adult, multiciliated ependymal cells do not divide (Capela and Temple 2002; Spassky et al. 2005) and the best known function of ependymal cilia is to create a constant laminar flow of CSF through the cerebral ventricles. For efficient fluid flow, the ependymal cilia must beat in the same direction and in a coordinated fashion. This coordinated or “metachronic” beating of adjacent cilia, seen as waves propagating over the surface of ciliated cells, is possibly mediated by tight coupling of the cells through gap junctions (Scott et al. 1974; Fig. 7.3). The close spatial relationships among cilia and the hydrodynamic interactions generated by their beating are believed, however, to be the most important factors for ciliary coordination on ependymal surfaces (Sanderson and Sleigh 1981; Salathe 2007; Guirao and Joanny 2007; Guirao et al. 2010; Fig. 7.3). The beating of ependymal cilia is highly asymmetrical, similar to that of motile cilia in other organs, such as airway epithelia and the oviduct. The beat begins with the formation of a large curve at the base of the cilium that sweeps the cilium forward in an effective stroke. The cilium then returns to its initial position through the propagation of the curve during the recovery stroke (Lechtreck et al. 2008). The waveform of the ependymal ciliary beat is nearly planar, creating a constant laminar flow of fluid: the effective stroke significantly displaces the fluid in the direction of the beat, whereas the recovery stroke has a minimal effect on fluid displacement. The frequency of ciliary beats is around 30 Hz under basal conditions, but may be modulated by external signals; for example, it may increase to 50 Hz after application of serotonin (5-HT, which causes a prolonged increase of intracellular Ca2+ due to the opening of Ca2+ release-activated Ca2+ channels on the plasma membrane; Nguyen et al. 2001) or adenosine receptor agonists (due to A2B receptor activation, Genzen et al. 2009). It can be decreased by application of ATP through intracellular Ca2+-independent, purinergic receptors and an intracellular cAMP-mediated pathway (Nguyen et al. 2001).
Fig. 7.2 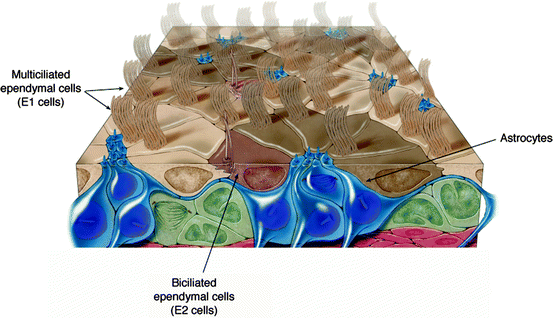
Drawing of cells in contact with the lateral ventricles in the mouse adult brain (Modified from Mirzadeh et al. (2008))
Fig. 7.3 
Drawing of ependymal ciliary beating coordination leading to metachronal waves propagating over the surface of the cells and efficient fluid flow in the cerebral ventricles
-
Bi-ciliated ependymal cells: are sparsely distributed throughout the lateral wall of the lateral ventricles (5% of cells contacting the lateral ventricle; called E2 cells; Fig. 7.2) and are the most common cells observed in the adult mouse spinal cord (Ecc cells). At the ultrastructural level, E2 and Ecc have deeply interdigitated cell membranes with long lateral extensions, light cytoplasm, small dictyosomes (abundant cisternae of rough endoplasmic reticulum and polyribosomes) and spherical uninvaginated nuclei containing dispersed chromatin. They have abundant mitochondria concentrated around the nucleus and two complex basal bodies anchored at the apical surface with 9 + 2 motile cilia. They are stained with antibodies against CD24, vimentin, Dlx2, S100β and GFAP (Doetsch et al. 1997; Danilov et al. 2009). They form specialized intercellular tight and adherens junctions with E1 cells. It has been proposed that E2 cells may serve as mechanical or chemical sensors of CSF flow or composition (Bruni 1998; Mirzadeh et al. 2008). It is not known whether these cells are mitotically active. In the mouse spinal cord, Ecc cells are Vimentin+, CD24+, FoxJ1+, Sox2+ and CD133+ but nestin− and GFAP−. These cells resemble E2 cells in the lateral ventricles, but their basal bodies are different: Each basal body is associated with complex pericentriolar electron-dense particles organized into multiple (up to six) radial spikes. Ecc cells are mitotically active, but the timing of their greatest proliferation together with the distribution of pairs of BrdU-labeled cells at different time points after injection suggest that proliferation of these cells contributes to the extension of the central canal during spinal cord growth (Alfaro-Cervello et al. 2012).


7.2.3.2 Other Cells
-
Tanycytes are bipolar cells with microvilli and primary cilia (9 + 0 structure) that mostly contact the third ventricle, although a few have been observed in the lateral walls of the lateral ventricles (Doetsch et al. 1997) and in discrete locations around the central canal of the spinal cord. Tanycytes share some features with radial glia and astrocytes: they stain strongly with antibodies against GFAP, nestin, Glast, vimentin Sox2, S100β and the functional receptors for the neurotransmitters γ-aminobutyrate (GABA) and glutamate (Rodriguez et al. 2005; Hamilton et al. 2009). In the third ventricle, tanycytes connects the CSF to the hypothalamus portal system and several studies provide molecular features suggesting that they link the CSF to neuroendocrine events (Rodriguez et al. 2005).
-
Astrocytes are multipolar cells localized in the subventricular zone of the lateral ventricles. These cells represent one-third of all lateral ventricle-contacting cells and were demonstrated to be the adult neural stem cells (Doetsch et al. 1999). These cells were also observed along the central canal of the adult mouse spinal cord (Martens et al. 2002), the most dorsal of which were identified as neural stem cells (Sabourin et al. 2009). In the lateral ventricle, their apical process contains a primary cilium and form rosette-like clusters surrounded by multiciliated ependymal cells, although the functional significance of this architecture is still unknown (Mirzadeh et al. 2008; Fig. 7.2).
-
An extensive plexus of axons originating mainly from serotoninergic neurons in the dorsal raphe nucleus in the reticular formation have been found on the ventricular surface of multiciliated ependymal cells, in direct contact with the cerebral ventricles. These neurons have been shown to use serotonin and glutamate as co-transmitters. Interestingly, their axons often terminate in expansions near the base of ependymal cilia and tufts of cilia are often encircled by these axons. It has been proposed that glutamate provided by supra-ependymal axons may supplement metabolic pathways in multiciliated ependymal cells to fuel the high energy demands of ciliary beating (Harandi et al. 1986; Robinson et al. 1996). However, the roles of these supra-ependymal axons and their possible interactions with ependymal cells and CSF still need to be understood.
7.3 Development of Multiciliated Ependymal Cells
7.3.1 Description of the Main Stages of Ependymal Cells Development
During embryonic development, bipolar radial glial cells express the radial glial markers RC2 and GLAST and extend both a pial and ventricular process with a 9 + 0 cilium that project into the ventricular lumen (Tramontin et al. 2003). A subpopulation of radial glial cells is specified for the ependymal lineage around E15 in mice (Rakic and Sidman 1968 ; Spassky et al. 2005). Around birth, the nuclei of ependymal progenitors invaginate, and electron-dense aggregates and multiple deuterosomes appear in the cytoplasm. Deuterosomes are spherical structures of unknown composition, which act as nucleation centers for ciliary basal bodies in multiciliated cells (Dirksen 1971). These cells express both the radial glial cell marker GLAST and the ependymal cell markers S100β and CD24. Future basal bodies then detach from deuterosomes and migrate toward the apical surface where they dock to the cell membrane and start to extend 9 + 2 cilia. Basal bodies are first broadly distributed in the apical membrane and gradually become asymmetrically localized within the apical membrane, accumulating at the anterior region of each ependymal cell (Hirota et al. 2010). This aspect of ependymal cells planar polarity is called “translational polarity” and is unique to multiciliated cells in the brain; cells with motile cilia in the trachea and oviduct contain basal bodies covering their entire apical surface. Concomitant to the establishment of translational polarity, cilia increase in length and start beating, producing fluid flow that orients the basal bodies in the same direction. This is referred to “rotational polarity”. The orientation of each basal body is determined by the positioning of its accessory structure called the basal foot, which points in the direction of the effective stroke (Mitchell et al. 2007). The cilia first beat in independent directions and progressively orient in a common and fixed direction (Guirao et al. 2010). At this mature stage, the features characteristic of immature cells (deuterosomes and aggregates) have disappeared and the planar polarized beating of the cilia directs the flow of CSF through the cerebral ventricles, which is crucial for brain development and function (Ibanez-Tallon et al. 2004; Sawamoto et al. 2006).
7.3.2 Transcription Factors Involved in Ependymal Cell Development
Some transcription factors required for ependymal cells development (Vax1, Sox2, Six3) have been identified, although the molecular mechanisms by which radial glial cells become ependymal cells still remain to be elucidated (Soria et al. 2004; Ferri et al. 2004; Lavado and Oliver 2011), the transcription factors FoxJ1 (also known as hepatocyte nuclear factor-3 and forkhead homolog 4) and RFX (regulatory factor X homologs) appear to be implicated. In FoxJ1 mutant mice, axonemal dyneins and kinesins are downregulated, causing defective apical migration of basal bodies and defects in the genesis of motile cilia (Brody et al. 2000; Stubbs et al. 2008; Jacquet et al. 2009; Yu et al. 2008). It was recently proposed that FoxJ1 contributes to ependymal cell differentiation by controlling ankyrin G (Ank3) expression in ependymal cells. Ank3 is a large adaptor molecule that binds to E- and N-cadherin at cell membranes (Paez-Gonzalez et al. 2011). RFX3 belongs to the regulatory factor X (RFX) family of transcription factors, which are involved in the assembly and function of cilia in nematodes, drosophila and mice (Swoboda et al. 2000; Dubruille et al. 2002; Bonnafe et al. 2004). In mice, an RFX3 deficiency leads to hydrocephalus, partly due to defects in the differentiation of the subcommissural organ and choroid plexuses (Baas et al. 2006). Interestingly, RFX3 binds to FoxJ1 promoter and to the promoters of the genes encoding two axonemal dyneins involved in ciliary motility, suggesting that RFX3 regulates both ciliary assembly and motility (Zein El et al. 2009).
7.3.3 Mechanisms Regulating Ependymal Cell Polarity
-
Translational polarity: Although the functional significance of the ependymal translational polarity in the adult is still unknown, the mechanisms involved in its regulation are starting to be deciphered. Specific ablation of radial glial cells primary cilia using mouse genetics leads to random distribution of the patch of basal bodies at the apical surface of ependymal cells, suggesting that the primary cilium provides important positional information for correct basal body accumulation in ependymal cells (Mirzadeh et al. 2010). It was also shown that during mouse brain development, non-muscle myosin II (NMII) is involved in the rostral migration of basal bodies, independently of the planar cell polarity pathway (Hirota et al. 2010; Fig. 7.4).
Fig. 7.4 
Development of ependymal cells in wild-type and mutant animals. Radial glial cells extending a primary cilium transform into multiciliated ependymal cells. Motile cilia orient their beating in the direction of the flow: patch of basal bodies migrate to the anterior side of the cell and basal feet (red) point in the direction of the flow. Pink indicate the microtubule lattices and orange indicate the localisation of Vangl proteins at the apical and posterior side of the cells. In PCP mutants or in cilia defective ependymal cells or in OFD2 mutant trachea cells, motile cilia beat in random directions and leads to rotational polarity defects. In cilia defective radial glial cells, patch of basal bodies are randomly distributed along the apical surface of ependymal cells and leads to translational polarity defects
-
Rotational polarity: The oriented and coordinated ependymal ciliary beating is crucial for CSF flow and adult neurogenesis, and depends on the rotational polarity of each basal body in the cell. During development, ependymal cilia grow and start beating in random orientations and then progressively align in each cell and along the tissue axis. Using mutant mice, siRNA, or dominant-negative constructs, it was recently shown that the core planar cell polarity genes Dishevelled2, Van Gogh-like2 (Vangl2), cadherin, EGF, laminin, seven-pass receptors2 and 3 (Celsr2 and Celsr3) are required for alignment of ependymal basal feet during development (Guirao et al. 2010; Hirota et al. 2010; Tissir et al. 2010; Fig. 7.4). Most interestingly, it was shown that hydrodynamic forces among beating cilia help determine the direction of ependymal ciliary beating during maturation. Since Vangl2 is localized along cilia at these stages, it was hypothesized that Vangl2 may be part of a mechanosensor complex able to convert hydrodynamic forces into intracellular mechanisms for global basal foot alignment (Guirao et al. 2010). How PCP proteins interact is unknown, but perturbation of the cellular distribution of Vangl2 in cells in which Celsr2 and 3 are invalidated suggests that these genes may cooperate in the regulation of rotational polarity (Tissir et al. 2010). Most interestingly, it was recently shown that targeted elimination of exons 6 and 7 of Odf2, encoding a basal body and centrosome-associated protein Odf2/cenexin, leads to basal bodies lacking basal feet, disruption of the polarized organization of the apical microtubule lattice and uncoordinated beating of motile cilia in the trachea without affecting planar cell polarity. Altogether, these results suggest a model in which maturing motile cilia may orient their beating in response to a coupling between hydrodynamic forces and planar cell polarity, thanks to a downstream planar cell polarity-based basal foot-dependent organization of the apical microtubule lattice (Kunimoto et al. 2012).

7.4 Roles of Ependymal Motile Cilia
7.4.1 Contribution to the CSF Flow
Multiciliated ependymal cells lining the cerebral ventricles extend around 50 long motile cilia beating in a coordinated fashion, indicating that cilia beat as part of a metachronal wave (wave generated by the synchronized beating along the longitudinal ciliary rows). Although the bulk transport of CSF along the brain ventricles is mostly achieved by the changing blood pressures of the brain vessels during systole and diastole (Bradley et al. 1986), coordinated beating of ependymal cilia may also contribute to CSF flow along the ventricular walls (Sawamoto et al. 2006; Guirao et al. 2010). Depending on the size and shape of each brain ventricle (especially in large mammals), and given that ependymal cilia are around 12 μm long, ependymal ciliary beating may contribute to CSF flow at least in the narrowest regions of the ventricular system, i.e. the cerebral aqueduct. As proof of principle, it is noteworthy that in mutant mouse models in which cilia formation or cilia motility is affected (e.g. Hydin, stumpy protein, Tg737orpk, Mdnah5 mutant mice), mice develop hydrocephalus in postnatal stages due to aqueduct closure and massive dilatation of the third and lateral ventricles (Ibanez-Tallon et al. 2004; Banizs et al. 2005; Lechtreck et al. 2008; Town et al. 2008). It is thus clear that ependymal ciliary beating is crucial for CSF laminar flow along all brain ventricular walls, especially in the narrowest regions of the ventricular system (the cerebral aqueduct) where it prevents aqueduct stenosis and ventricle enlargement (Ibanez-Tallon et al. 2003, 2004).
7.4.2 Contribution to CSF-Brain Exchanges
In human airway epithelia, motile cilia beat to propel harmful inhaled material out of the lung. It was recently demonstrated that motile cilia in the airway are able to sense noxious substances entering the airways and rapidly increase ciliary beat frequency (CBF) as a defense mechanism (Shah et al. 2009). Similarly, an increase in ependymal ciliary beating in brain ventricles might minimize the layer of unstirred CSF covering the ependyma and optimize the dispersion or spreading of neural messengers in the CSF to other regions of the CNS or move debris in the direction of CSF bulk flow. Conversely, reducing CBF may allow ciliated ependymal cells to retain or localize certain messengers in the CSF, generating short-range effects on the exchange of neural messengers at the CSF-brain interface (Cathcart and Worthington 1964; Roth et al. 1985). These possibilities suggest that motile cilia in the brain, as in the airways, might have sensory roles leading to their active participation in brain-CSF exchanges. It was observed, for example, that ependymal cells express morphogens and growth factors (Noggin, Fibroblast Growth Factor 2, Vascular Endothelial Growth Factor, Hepatocyte Growth Factor, Insulin Growth Factor Binding Protein, etc.…) (Lim et al. 2000; Hayamizu et al. 2001; Arai et al. 1998; Calvo et al. 2011), suggesting that ependymal cells may contribute to autocrine or paracrine trophic support, perhaps through exocytosis from motile cilia. Interestingly, PDGF-α and EGF receptors are localized along ependymal cilia although these cells are post-mitotic; their roles in ependymal cilia are still unknown (del Bigio 2010; Danilov et al. 2009). It was recently shown that ependymal cells along the central canal of the spinal cord extend two motile cilia (Alfaro-Cervello et al. 2012), although it is unknown whether ciliary beating in the spinal cord is coordinated and whether this might contribute to exchanges between CSF and neural cells.
7.4.3 Contribution to Neuronal Migration in the Adult
In the adult mammalian brain, new neurons are generated in the subventricular zone (SVZ) of the lateral ventricles (see Chap. 4 on adult neurogenesis). These progenitors migrate long distances toward the olfactory bulb where they differentiate into interneurons. The choroid plexus is a source of chemorepulsive factors, including members of the Slit family, which influence SVZ cell migration (Hu 1999; Nguyen-Ba-Charvet et al. 2004). Infusion of a recombinant Slit2-alkaline phosphatase fusion protein into the adult lateral ventricles revealed that the alkaline phosphatase signal is mainly found on the surface of the ependymal layer and in a gradient along the dorsal SVZ, where neuroblasts form longitudinal arrays of chains and migrate predominantly in the rostral direction. Most importantly, this gradient does not form when Slit2-AP is injected in Tg737orpk adult mutant mice, in which cilia and CSF are abnormal, suggesting that ependymal ciliary beating is crucial for CSF flow in adult mouse brain and for the formation of the Slit2 gradient in the SVZ in vivo (Sawamoto et al. 2006).
7.5 Associated Pathologies
In humans, the main pathology associated with defective ependymal cilia is hydrocephalus. Hydrocephalus is a progressive pathological condition characterized by the excessive accumulation of CSF, which can be caused by impaired CSF flow, excess CSF production or a lack of CSF reabsorption (Bruni et al. 1985). Current treatments involve surgical insertion of a ventricular shunt to facilitate drainage of excess CSF. Among ciliopathies, primary ciliary dyskinesia is the main pathology involving hydrocephalus. It arises from ultrastructural defects that perturb the motility of all motile cilia and cause a variety of symptoms that include fertility problems (ectopic pregnancy, sperm immotility) sinusitis, otitis, bronchiectasis, hydrocephalus and situs inversus (Zariwala et al. 2007).
In mice, all mutant in which cilia formation or motility is affected develop hydrocephalus (Ibanez-Tallon et al. 2004; Banizs et al. 2005; Lechtreck et al. 2008; Town et al. 2008). The lack of ependymal flow causes a secondary closure of the aqueduct and subsequent formation of triventricular hydrocephalus during early postnatal brain development. In humans, ependymal ciliary dysmotility is not sufficient to cause hydrocephalus but increases the risk of aqueduct closure; there is a 1:40 incidence of hydrocephalus caused by aqueduct stenosis in primary ciliary dyskinesia patients compared to 3:10,000 in the general population (Ibanez-Tallon et al. 2003). Defects in ependymal motile cilia are responsible for only some forms of hydrocephalus. For example, excess CSF production by the choroid plexus also leads to hydrocephalus but is primarily due to chemosensory defects of primary cilia in the choroid plexus cells (Banizs et al. 2005; Narita et al. 2010).
References
Alfaro-Cervello C, Soriano-Navarro M, Mirzadeh Z et al (2012) Biciliated ependymal cell proliferation contributes to spinal cord growth. J Comp Neurol. doi:10.1002/cne.23104 (ahead of publication)
Arai Y, Deguchi K, Takashima S (1998) Vascular endothelial growth factor in brains with periventricular leukomalacia. Pediatr Neurol 19:45–49
Baas D, Meiniel A, Benadiba C et al (2006) A deficiency in RFX3 causes hydrocephalus associated with abnormal differentiation of ependymal cells. Eur J Neurosci 24:1020–1030. doi:10.1111/j.1460-9568.2006.05002.x
Banizs B, Pike MM, Millican CL et al (2005) Dysfunctional cilia lead to altered ependyma and choroid plexus function, and result in the formation of hydrocephalus. Development 132:5329–5339
Bering EA Jr (1955) Choroid plexus and arterial pulsation of cerebrospinal fluid; demonstration of the choroid plexuses as a cerebrospinal fluid pump. AMA Arch Neurol Psychiatry 73(2):165–172
Bonnafe E, Touka M, AitLounis A et al (2004) The transcription factor RFX3 directs nodal cilium development and left-right asymmetry specification. Mol Cell Biol 24:4417–4427. doi:10.1128/MCB.24.10.4417-4427.2004
Bradley WJ, Kortman K, Burgoyne B (1986) Flowing cerebrospinal fluid in normal and hydrocephalic states: appearance on MR images. Radiology 159(3):611–616
Brody SL, Yan XH, Wuerffel MK et al (2000) Ciliogenesis and left-right axis defects in forkhead factor HFH-4-null mice. Am J Respir Cell Mol Biol 23:45–51
Bruni JE (1998) Ependymal development, proliferation, and functions: a review. Microsc Res Tech 41:2–13. doi:10.1002/(SICI)1097-0029(19980401)41:1<2::AID-JEMT2>3.0.CO;2-Z
Bruni JE, Del Bigio MR, Clattenburg R (1985) Ependyma: normal and pathological. A review of the literature. Brain Res 356(1):1–19
Calvo CF, Fontaine RH, Soueid J et al (2011) Vascular endothelial growth factor receptor 3 directly regulates murine neurogenesis. Genes Dev 25:831–844. doi:10.1101/gad.615311
Capela A, Temple S (2002) LeX/ssea-1 is expressed by adult mouse CNS stem cells, identifying them as nonependymal. Neuron 35:865–875
Cathcart RS 3rd, Worthington WC Jr (1964) Ciliary movement in the rat cerebral ventricles: clearing action and directions of currents. J Neuropathol Exp Neurol 23:609–618
Danilov AI, Gomes-Leal W, Ahlenius H et al (2009) Ultrastructural and antigenic properties of neural stem cells and their progeny in adult rat subventricular zone. Glia 57:136–152. doi:10.1002/glia.20741
Del Bigio MR (2010) Ependymal cells: biology and pathology. Acta Neuropathol 119:55–73. doi:10.1007/s00401-009-0624-y
Dirksen ER (1971) Centriole morphogenesis in developing ciliated epithelium of the mouse oviduct. J Cell Biol 51:286–302
Doetsch F, Garcia-Verdugo JM, Alvarez-Buylla A (1997) Cellular composition and three-dimensional organization of the subventricular germinal zone in the adult mammalian brain. J Neurosci 17:5046–5061
Doetsch F, Caille I, Lim DA et al (1999) Subventricular zone astrocytes are neural stem cells in the adult mammalian brain. Cell 97:703–716
Dubruille R, Laurençon A, Vandaele C et al (2002) Drosophila regulatory factor X is necessary for ciliated sensory neuron differentiation. Development 129:5487–5498. doi:10.1242/dev.00148
Ferri AL, Cavallaro M, Braida D et al (2004) Sox2 deficiency causes neurodegeneration and impaired neurogenesis in the adult mouse brain. Development 131:3805–3819. doi:10.1242/dev.01204
Genzen JR, Yang D, Ravid K, Bordey A (2009) Activation of adenosine A2B receptors enhances ciliary beat frequency in mouse lateral ventricle ependymal cells. Cerebrospinal Fluid Res 6:15. doi:10.1186/1743-8454-6-15
Guirao B, Joanny JF (2007) Spontaneous creation of macroscopic flow and metachronal waves in an array of cilia. Biophys J 92:1900–1917. doi:10.1529/biophysj.106.084897
Guirao B, Meunier A, Mortaud S et al (2010) Coupling between hydrodynamic forces and planar cell polarity orients mammalian motile cilia. Nat Cell Biol 12:341–350. doi:10.1038/ncb2040
Hamilton LK, Truong MKV, Bednarczyk MR et al (2009) Cellular organization of the central canal ependymal zone, a niche of latent neural stem cells in the adult mammalian spinal cord. Neuroscience 164:1044–1056. doi:10.1016/j.neuroscience.2009.09.006
Harandi M, Didier M, Aguera M et al (1986) GABA and serotonin (5-HT) pattern in the supraependymal fibers of the rat epithalamus: combined radioautographic and immunocytochemical studies. Effect of 5-HT content on [3] GABA accumulation. Brain Res 370(2):241–249
Hayamizu TF, Chan PT, Johanson CE (2001) FGF-2 immunoreactivity in adult rat ependyma and choroid plexus: responses to global forebrain ischemia and intraventricular FGF-2. Neurol Res 23:353–358
Hirota Y, Meunier A, Huang S et al (2010) Planar polarity of multiciliated ependymal cells involves the anterior migration of basal bodies regulated by non-muscle myosin II. Development 137:3037–3046. doi:10.1242/dev.050120
Hu H (1999) Chemorepulsion of neuronal migration by Slit2 in the developing mammalian forebrain. Neuron 23:703–711
Ibanez-Tallon I, Heintz N, Omran H (2003) To beat or not to beat: roles of cilia in development and disease. Hum Mol Genet 12(Spec No 1):R27–R35
Ibanez-Tallon I, Pagenstecher A, Fliegauf M et al (2004) Dysfunction of axonemal dynein heavy chain Mdnah5 inhibits ependymal flow and reveals a novel mechanism for hydrocephalus formation. Hum Mol Genet 13:2133–2141
Jacquet BV, Salinas-Mondragon R, Liang H et al (2009) FoxJ1-dependent gene expression is required for differentiation of radial glia into ependymal cells and a subset of astrocytes in the postnatal brain. Development 136:4021–4031. doi:10.1242/dev.041129
Kunimoto K, Yamazaki Y, Nishida T et al (2012) Coordinated ciliary beating requires Odf2-mediated polarization of basal bodies via basal feet. Cell 148:189–200. doi:10.1016/j.cell.2011.10.052
Lavado A, Oliver G (2011) Six3 is required for ependymal cell maturation. Development 138:5291–5300. doi:10.1242/dev.067470
Lechtreck KF, Delmotte P, Robinson ML et al (2008) Mutations in Hydin impair ciliary motility in mice. J Cell Biol 180:633–643. doi:10.1083/jcb.200710162
Lim DA, Tramontin AD, Trevejo JM et al (2000) Noggin antagonizes BMP signaling to create a niche for adult neurogenesis. Neuron 28:713–726
Martens DJ, Seaberg RM, van der Kooy D (2002) In vivo infusions of exogenous growth factors into the fourth ventricle of the adult mouse brain increase the proliferation of neural progenitors around the fourth ventricle and the central canal of the spinal cord. Eur J Neurosci 16:1045–1057
Mirzadeh Z, Merkle FT, Soriano-Navarro M et al (2008) Neural stem cells confer unique pinwheel architecture to the ventricular surface in neurogenic regions of the adult brain. Cell Stem Cell 3:265–278. doi:10.1016/j.stem.2008.07.004
Mirzadeh Z, Han YG, Soriano-Navarro M et al (2010) Cilia organize ependymal planar polarity. J Neurosci 30:2600–2610. doi:10.1523/JNEUROSCI.3744-09.2010
Mitchell B, Jacobs R, Li J et al (2007) A positive feedback mechanism governs the polarity and motion of motile cilia. Nature 447:97–101
Miyan JA, Nabiyouni M, Zendah M (2003) Development of the brain: a vital role for cerebrospinal fluid. Can J Physiol Pharmacol 81:317–328. doi:10.1139/y03-027
Narita K, Kawate T, Kakinuma N, Takeda S (2010) Multiple primary cilia modulate the fluid transcytosis in choroid plexus epithelium. Traffic 11:287–301. doi:10.1111/j.1600-0854.2009.01016.x
Nguyen T, Chin WC, O’Brien JA et al (2001) Intracellular pathways regulating ciliary beating of rat brain ependymal cells. J Physiol 531:131–140
Nguyen-Ba-Charvet KT (2004) Multiple roles for slits in the control of cell migration in the rostral migratory stream. J Neurosci 24:1497–1506. doi:10.1523/JNEUROSCI.4729-03.2004
Paez-Gonzalez P et al (2011) Ank3-dependent SVZ niche assembly is required for the continued production of new neurons. Neuron 71:61–75. doi:10.1016/j.neuron.2011.05.029
Rakic P, Sidman RL (1968) Subcommissural organ and adjacent ependyma: autoradiographic study of their origin in the mouse brain. Am J Anat 122:317–335. doi:10.1002/aja.1001220210
Robinson SR, Noone DF, O’Dowd BS (1996) Ependymocytes and supra-ependymal axons in rat brain contain glutamate. Glia 17:345–348. doi:10.1002/(SICI)1098-1136(199608)17:4<345::AID-GLIA9>3.0.CO;2-Y
Rodríguez EM, Blázquez JL, Pastor FE et al (2005) Hypothalamic tanycytes: a key component of brain-endocrine interaction. Int Rev Cytol 247:89–164. doi:10.1016/S0074-7696(05)47003-5
Roth Y, Kimhi Y, Edery H, Aharonson E (1985) Ciliary motility in brain ventricular system and trachea of hamsters. Brain Res 330(2):291–297
Sabourin J-C, Ackema KB, Ohayon D et al (2009) A mesenchymal-like ZEB1 + niche harbors dorsal radial glial fibrillary acidic protein-positive stem cells in the spinal cord. Stem Cells 27:2722–2733. doi:10.1002/stem.226
Salathe M (2007) Regulation of mammalian ciliary beating. Annu Rev Physiol 69:401–422. doi:10.1146/annurev.physiol.69.040705.141253
Sanderson MJ, Sleigh MA (1981) Ciliary activity of cultured rabbit tracheal epithelium: beat pattern and metachrony. J Cell Sci 47:331–347
Sawamoto K, Wichterle H, Gonzalez-Perez O et al (2006) New neurons follow the flow of cerebrospinal fluid in the adult brain. Science 311:629–632
Schutzer SE, Liu T, Natelson BH et al (2010) Establishing the proteome of normal human cerebrospinal fluid. PLoS One 5:e10980. doi:10.1371/journal.pone.0010980.g003
Scott DE, Kozlowski GP, Sheridan MN (1974) Scanning electron microscopy in the ultrastructural analysis of the mammalian cerebral ventricular system. Int Rev Cytol 37:349–388
Segal MB (2000) The choroid plexuses and the barriers between the blood and the cerebrospinal fluid. Cell Mol Neurobiol 20:183–196. doi:10.1002/1097-0029(20010101)52:1<38::AID-JEMT6>3.0.CO;2-J
Segal MB (2001) Transport of nutrients across the choroid plexus. Microsc Res Tech 52(1):38–48
Shah AS, Ben-Shahar Y, Moninger TO et al (2009) Motile cilia of human airway epithelia are chemosensory. Science 325:1131–1134. doi:10.1126/science.1173869
Soria JM, Taglialatela P, Gil-Perotin S et al (2004) Defective postnatal neurogenesis and disorganization of the rostral migratory stream in absence of the Vax1 homeobox gene. J Neurosci 24:11171–11181. doi:10.1523/JNEUROSCI.3248-04.2004
Spassky N, Merkle FT, Flames N et al (2005) Adult ependymal cells are postmitotic and are derived from radial glial cells during embryogenesis. J Neurosci 25:10–18
Stubbs JL, Oishi I, Izpisua Belmonte JC, Kintner C (2008) The forkhead protein Foxj1 specifies node-like cilia in Xenopus and zebrafish embryos. Nat Genet 40:1454–1460. doi:10.1038/ng.267
Swoboda P, Adler HT, Thomas JH (2000) The RFX-type transcription factor DAF-19 regulates sensory neuron cilium formation in C. elegans. Mol Cell 5:411–421
Tissir F, Qu Y, Montcouquiol M et al (2010) Lack of cadherins Celsr2 and Celsr3 impairs ependymal ciliogenesis, leading to fatal hydrocephalus. Nat Neurosci 13:700–707. doi:10.1038/nn.2555
Town T, Breunig JJ, Sarkisian MR et al (2008) The stumpy gene is required for mammalian ciliogenesis. Proc Natl Acad Sci USA 105:2853–2858. doi:10.1073/pnas.0712385105
Tramontin AD, Garcia-Verdugo JM, Lim DA, Alvarez-Buylla A (2003) Postnatal development of radial glia and the ventricular zone (VZ): a continuum of the neural stem cell compartment. Cereb Cortex 13:580–587
Veening JG, Barendregt HP (2010) The regulation of brain states by neuroactive substances distributed via the cerebrospinal fluid; a review. Cerebrospinal Fluid Res 7:1. doi:10.1186/1743-8454-7-1
Wagshul ME, Chen JJ, Egnor MR et al (2006) Amplitude and phase of cerebrospinal fluid pulsations: experimental studies and review of the literature. J Neurosurg 104(5):810–819
Yu X, Ng CP, Habacher H, Roy S (2008) Foxj1 transcription factors are master regulators of the motile ciliogenic program. Nat Genet 40:1445–1453. doi:10.1038/ng.263
Zariwala MA, Knowles MR, Omran H (2007) Genetic defects in ciliary structure and function. Annu Rev Physiol 69:423–450. doi:10.1146/annurev.physiol.69.040705.141301
Zein El L, Ait-Lounis A, Morle L et al (2009) RFX3 governs growth and beating efficiency of motile cilia in mouse and controls the expression of genes involved in human ciliopathies. J Cell Sci 122:3180–3189. doi:10.1242/jcs.048348
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2013 Springer Science+Business Media Dordrecht
About this chapter
Cite this chapter
Spassky, N. (2013). Motile Cilia and Brain Function: Ependymal Motile Cilia Development, Organization, Function and Their Associated Pathologies. In: Tucker, K., Caspary, T. (eds) Cilia and Nervous System Development and Function. Springer, Dordrecht. https://doi.org/10.1007/978-94-007-5808-7_7
Download citation
DOI: https://doi.org/10.1007/978-94-007-5808-7_7
Published:
Publisher Name: Springer, Dordrecht
Print ISBN: 978-94-007-5807-0
Online ISBN: 978-94-007-5808-7
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)