
Abstract
It is well accepted that the ability of a biologically molecule to selectively find its target influences its potential as a successful therapeutic drug. For many molecules the molecular target is located inside sub-cellular structures. Molecules with such sub cellular targets and the inability to specifically accumulate at the location of the target can potentially be made more active by targeting strategies that improve their accumulation at the target. Pharmaceutical nanocarriers form the basis of several such targeting strategies. This chapter deals with the rational approach underlying the current uses of nanocarriers to deliver bioactive molecules to sub cellular compartments.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


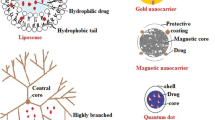
Keywords
1 Introduction
Drug therapy at the most fundamental level is based on the interaction of two molecules. An exogenous molecule administered to a patient and the molecule in the patient that the administered molecule interacts with to initiate a physiological response. In an ideal scenario, the administered molecule interacts with only one physiological molecule and produces a physiological response that improves a patient’s condition. In this context it is clear that the term target may be applied to the physiological molecule and the administered molecule is a drug. The concept of targeting has multiple definitions. From a drug discovery perspective, targeting is very often described in terms of the drug molecule’s ability to interact only with the target. This concept is more appropriately described by the use of the term selectivity and is very different from the concept of targeting from a military perspective where the term arguably first originated. Consider the firing of a bullet from a gun as an example. The object the bullet is intended to hit is the target, while targeting is associated with the act of aiming the gun so the bullet hits the target. The action that the bullet produces is destruction of the target. This action is indiscriminate in that if the bullet hits an object other than the target, that object will be destroyed as well. Using the gunshot analogy to illustrate the drug discovery perspective on targeting would involve firing bullets that only destroy the target but leave the non-targets unharmed. Most approaches to disease therapy have followed such an argument: finding such selective molecules has been relatively easy when there were significant differences between the disease causing process and normal human biochemical pathways. Not surprisingly, infectious diseases are relatively easier to treat than inherent disorders. The selectivity is however dose dependent and most drugs that are considered to be selectively toxic to invading pathogens are in fact toxic to human cells as well, but at higher doses.
The current challenges in drug therapy lie in the treatment of diseases associated with malfunctions of normal human biochemical pathways in certain tissues. More often that not, even dose dependent selectivity is hard to achieve. Therefore the concept of targeting is becoming more and more associated with selective delivery. The term ‘targeting’ should ideally imply that the molecule is in some way able to selectively accumulate at an intended site of action and that the selective accumulation is associated with its selective action. This distinction is particularly important in developing targeted therapy for a disease like cancer. Unless unique molecular targets found exclusively (or at sufficiently higher levels) in the diseased state and not in normal state are discovered, selective accumulation at the disease site is crucial to the improvement of therapy. In summary, it can be said that there appear to be two distinct approaches to targeting in the context of drug therapy. The first involves selective action on the target while the second involves selective accumulation at the target. Most if not all examples of targeting seem to end up being the combination of some degree of selective action on the target and some degree of selective accumulation at the site of the target. Improving the degree of selective accumulation has the added advantage, even for molecules with high target selective action, of reducing the required dose and hence should be a major focus of all targeting approaches.
In the context of drug molecules the properties of selective accumulation are associated with the concept of bioavailability and biodistribution that are related to the physico-chemical properties of the molecule. To overcome the limitations that a compound’s physico-chemical properties can impose on its potential pharmaceutical application, the process of large-scale screening of chemical libraries has been extended beyond identifying desired bioactivity. Screening approaches routinely incorporate selection for physico-chemical properties that are known to confer high bioavailability as well. Unfortunately, this approach often leads to many potent molecules being excluded from further development. These molecules often have a potent pharmacological action at a desired molecular target but aren’t able to find their way exclusively to that target. It is almost certain that there is a growing list of such molecules that are in essence potential drugs if only a delivery strategy can be devised to get them to their molecular target in the human body. Delivery strategies aimed at mediating the selective accumulation of a biologically active molecule fall into two broad classes. The first involves direct chemical modification of the molecule and includes the traditional approaches of designing chemical analogs of the molecule as well as more recent approaches of conjugation to macromolecules or ligands capable of directing site-specific binding of the bioactive molecule. The second approach is the use of pharmaceutical carriers in particular those approaches that involve physical entrapment of the bioactive in the carrier thus offering what might be viewed as a non-chemical approach to modify the disposition of drug molecules. All chemistry can be performed on the components of the nanocarrier system that can then be loaded with the drug to afford targeted delivery.
Generally speaking, the role of nanocarriers in drug targeting to tissues is well accepted. For example, nanocarriers are able to improve the tumor specific accumulation of anticancer drugs by virtue of the Enhanced Permeability and Retention (EPR) effect (Maeda et al. 2000; Greish 2010; Fang et al. 2003). Such targeting approaches have been applied to anticancer drugs like doxorubicin (Working et al. 1999; Northfelt et al. 1996; Gabizon et al. 1989a, b), daunorubicin (Gill et al. 1996; Cervetti et al. 2003), paclitaxel (Yang et al. 2007; Sharma et al. 1996, 1997), and cisplatin (Stathopoulos 2010) with commercial success in some cases (eg Doxil, Daunosome). Current approaches also routinely focus on improving cell specific accumulation of the nanocarriers as well. However, nanocarrier targeting at a sub-cellular level has until recently not been as widely pursued perhaps due to technological limitations or the argument that once a drug gets inside a cell it will eventually find its way to the subcellular target. There often seems to be an assumption that mediating cell cytosolic internalization is adequate to ensure the interaction of the drug molecule with its final sub-cellular target by virtue of simple diffusion of the drug molecule and random interaction with various sub-cellular structures in the cell. However, it has become increasingly evident during the last decade that such an assumption cannot always be made (Duvvuri and Krise 2005; Breunig et al. 2008; Torchilin 2006; Li and Huang 2008; Kaufmann and Krise 2007; Panyam and Labhasetwar 2003; Weissig 2003, 2005; Weissig et al. 2004, 2007). The potential role that nanocarriers play in controlling the subcellular disposition of bioactives is, therefore, certainly worth examining.
2 Potential Roles of the Nanocarrier in Controlling Sub-cellular Disposition of Bioactives
Bioactive molecules can potentially act in various sub cellular locations that can be roughly divided into general cytosolic locations, the surface of organelles and specific regions inside organelles. The nature of the molecule and its potential sites of action together influence the design of the nanocarrier delivery strategy. Cytosolic access, transport through the cytosol and organelle entry thus become the major barriers that need to be overcome to mediate the efficient delivery of the bioactive to its intended site of action.
2.1 Cytosolic Access
In the case of nanocarrier based intracellular delivery strategies, access to the cytosol is not a trivial matter. Based on current understanding it is accepted that all nanocarrier systems are subject to some form of endocytosis either receptor mediated or more commonly non specific endocytosis. Nanocarriers may be surface functionalized with endocytic targeting moieties such as transferrin, folic acid, low-density lipoprotein, cholera toxin, riboflavin, nicotinic acid and the tripeptide RGD, which lead to internalization by either clathrin-dependent receptor mediated endocytosis, caveolin-assisted endocytosis, lipid-raft assisted endocytosis or macropinocytosis (Bareford and Swaan 2007; Rajendran 2010). Strictly speaking a nanocarrier and the associated drug that is inside an endosome is not in the cytosol and therefore much research has been devoted to mediating the release of the bioactive cargo from the endosome into the cytosol (Rajendran 2010). Endosomal release may be mediated by a fusogenic mechanism where lipid-containing nanocarriers fuse with the endosomal membrane increasing the fluidity of the membrane and therefore the release of the cargo (Martin and Rice 2007). Alternatively endosomolytic agents that can disrupt the endosomal membrane by the so called proton sponge effect leading to the release of endosomal contents (Yessine and Leroux 2004) or photochemical internalization (PCI) using photosensitizer molecules that can rupture the endosomal membrane can also be utilized (Shiraishi and Nielsen 2006). Such approaches are arguably necessary in the case of agents such as siRNA since the target of the drug molecule is in the cytoplasm itself (Patil and Panyam 2009; Yuan et al. 2006; Tahara 2010; Song 2010). On the other hand retention in the endosome followed by subsequent lysosomal fusion is a desirable outcome in the case of bioactives requiring delivery into the lysosome. Recombinant human acid sphingomyelinase for example has been delivered intracellularly by nanocarriers surface functionalized with the anti-ICAM antibody that mediates endosomal uptake and subsequent lysosomal association of the enzyme. This approach has shown promise in the treatment of Nieman Pick’s disease type A and B (Muro et al. 2006). Most viruses utilize a cell penetrating peptide (CPP) that aids in the internalization of the virus in the cell. Some of these peptides can be anchored with the therapeutic drug molecule to facilitate its internalization into the cell. HIV’s Tat peptide is one such CPP, which has aided in the delivery of large proteins and DNA into the cell either when it is directly conjugated to the molecule (Fittipaldi and Giacca 2005; Schwarze et al. 1999; Fawell et al. 1994) or conjugated to a nanocarrier such as a liposome (Torchilin et al. 2001). Other CPPs, D-penetratin and Syn-B, have been used to enhance the intracellular delivery of doxorubicin in the brain (Rousselle et al. 2000).
2.2 Transport Through the Cytoplasm
The cytoplasm is a highly viscous space (Seksek et al. 1997; Goins et al. 2008), which has a high concentration of dissolved solutes (Ellis and Minton 2003; Minton 2006) and as such can present a significant barrier to the movement of many potential bioactive molecules especially those with high molecular weight. However, in the eukaryotic cells, transport of biomolecules such as proteins and lipids is governed by the actin and microtubule cytoskeletons (Vale 1987). The extent to which this transport system can be utilized for nanocarrier mediated intracellular transport remains to be explored. Interestingly it has recently been reported that PEGylation of nanoparticles improves their cytoplasmic transport presumably by allowing nanoparticles to evade non specific interactions in the highly crowded environment of the cytoplasm (Suh et al. 2007). It is also interesting to consider in this context the fact that endosomes are routinely trafficked through the cytosol in their normal progression to lysosomes. If the endosomal vesicle can somehow be rerouted to afford association with other membrane bound organelles like mitochondria or the nucleus, there may well be no need to have the biologically active molecule enter the cytosol. If for example, the nanocarrier components were to undergo a redistribution to become part of the endosome and the targeting ligand was able to redistribute to the surface of the endosomal vesicle, it might be possible then that the vesicle would have an altered sub-cellular fate that could involve transport to and association with a target compartment other than the lysosome. While there is emerging evidence to suggest that in fact cells actively traffic nanocarriers in cell membrane-derived vesicles (Ruan et al. 2007), the concept remains speculative until more is understood about the mechanisms of intracellular molecular versus vesicular transport. For now, the trend towards mediating endosomal release of internalized nanocarrier and associated bioactive is arguably based on insights gleaned from intracellular dynamics of viral particles that are in essence naturally occurring nanocarriers. Viral particles are endocytosed and then are able to mediate endosomal escape and subsequent nucleus specific delivery of their DNA. Based on the premise that to efficiently deliver DNA to the nucleus, a delivery system must penetrate through the plasma membrane and the nuclear envelope, prior to DNA release in the nucleus, a strategy that involved step-wise membrane fusion was devised. Using a multi-layered nanoparticle called a Tetra-lamellar Multi-functional Envelope-type Nano Device (T-MEND) consisting of a DNA-polycation condensed core coated with two nuclear membrane-fusogenic inner envelopes and two endosome-fusogenic outer envelopes, which are shed in stepwise fashion transgene expression in non-dividing cells was reported to be dramatically increased (Akita et al. 2009). A similar approach in designing a mitochondria specific delivery sytem has been reported as well. Liposomal carriers called MITO-Porters which carry octaarginine surface modifications to stimulate their entry into cells as intact vesicles (via macropinocytosis) were prepared with lipid compositions that were identified in various experiments to promote both fusion with the mitochondrial membrane and the release of liposomal cargo to the intra-mitochondrial compartment in living cells. Using GFP protein as a model cargo it was shown that MITOporter liposomes are able to selectively deliver their cargo to mitochondria (Yamada and Harashima 2008; Yamada et al. 2008). It is also possible that conferring upon a nanocarrier the properties of the membrane of a particular subcellular compartment might direct transport through the cytosol to the organelle of interest. This concept has been explored with a liposomal formulation of a crude mitochondrial fraction (Inoki et al. 2000). These so calle proteoliposomes were reported to colocalize with endogenous mitochondria when microinjected into pre implantation embryos.
In addition to particle composition, it has also been observed that changes in nanoparticle architecture affect sub-cellular disposition (Xu et al. 2008). Fluorescein isothiocyanate labeled layered double hydroxide (LDH) nanoparticles were prepared from Mg2Al under conditions that yielded either hexagonal sheets (5–150 nm wide and 10–20 nm thick) or nanorods (30–60 nm wide and 100–200 nm long). A comparison of the sub-cellular distribution of these two types of preparations revealed that the nanorods trafficked to the nucleus but the hexagonal sheets remained in the cytoplasm (Xu et al. 2008). An active microtubule mediated transport process is hypothesized to be responsible for the observed rapid nuclear accumulation of the nanorods (Xu et al. 2008).
2.3 Organelle Entry
As with the previous two, barriers hitchhiking existing biological processes extend to mediating organellar entry in most delivery approaches. Proteins synthesized in the cytosol bear short peptide sequences within the protein as ‘molecular zip codes’ that determine the preferential transport of a protein into a membrane bound subcellular organelle (Walter et al. 1982). These leader sequences or localization signals are typically recognized by specialized import proteins that are associated with the organelle (Allen et al. 2000; Stoffler et al. 1999). Localization sequences for the nucleus and the mitochondria are well known and have been explored for mediating the entry of a variety of molecular cargos as well as nanocarriers into these organelles. The potential of leader sequence peptides for overcoming intracellular barriers to DNA delivery was first demonstrated a decade ago (Zanta et al. 1999). To facilitate the import of exogenous DNA into the nucleus, a capped 3.3-kbp CMVLuciferase-NLS gene containing a single nuclear localization signal peptide (PKKKRKVEDPYC) was synthesized. The resulting transfection enhancement due to the nuclear leader peptide was about 10-fold to 1,000-fold irrespective of the cationic vector or the cell type used. At that time the authors hypothesized that the 3 nm wide DNA present in the cytoplasm was initially docked to and translocated through a nuclear pore by the nuclear import machinery and as DNA enters the nucleus, it was quickly condensed into a chromatin-like structure, which provided a mechanism for threading the remaining worm-like molecule through the pore (Zanta et al. 1999). In this context an interesting question arises as to whether localization signals should be directly conjugated to the bioactive molecule or the nano-carrier. Besides protein import machinery, other properties of organellar compartments like membrane lipid composition (Fernandez-Carneado et al. 2005), membrane potential (D’Souza et al. 2008) and even intra-organellar pH (Torchilin et al. 2009) have been explored for achieving selective delivery to organelles.
3 Prevalent Nanocarrier Design Approaches
It currently appears that nanocarrier design for subcellular targeting is based on the fractal symmetry between the case of drug delivery to a cell and drug delivery to a molecular target inside a sub-cellular compartment. The cell could be viewed as being a small, slightly simpler but nonetheless highly organized “body” with “organs” (organelles) and “cells” (defined structures and molecular arrangements) within these organs. Therefore principles that have been explored for organ and cell specific targeting are being applied at the subcellular level. Nanocarriers are either being modified with sub cellular targeting ligands or are being prepared from materials that have inherent subcellular accumulation characteristics. As alluded to in the previous section, much of this line of thinking is based on current understanding of viral particles. Viruses could be considered naturally occurring nanocarriers with the ability to selectively deliver their DNA cargo to a sub-cellular target (the nucleus). It is perhaps safe to say that much of what we know about the cellular interaction and sub-cellular disposition of nanocarriers has some how been associated with investigations into mimicking the DNA delivery capability of viruses using artificial nanocarriers.
3.1 Nanocarriers Modified with Sub-cellular Targeting Ligands
Most nanocarriers are believed to enter the cell by endocytic mechanisms and could therefore be considered as having a predisposition for accumulation in endosomes and potentially lysosomes as well. This predisposition of particulate systems is particularly useful as pathological conditions associated with endosomes and lysosomes could potentially benefit from therapies targeting these pathways (Bareford and Swaan 2007; Gregoriadis and Ryman 1971; Castino et al. 2003; Tate and Mathews 2006). The fate of the nanocarrier is dependent on the mechanism of vesicular internalization (Bareford and Swaan 2007). For example, nanoscale drug carrier systems taken up by clathrin-dependent receptor-mediated endocytosis (RME) are most likely to undergo lysosomal degradation, while clathrin-independent RME may lead to endosomal accumulation (Bareford and Swaan 2007). Consequently, the type of targeting moiety displayed by the nanocarrier system determines whether the carrier delivers its cargo to either endosomes or lysosomes. Several endocytic targeting moieties have been studied and include folic acid, low-density lipoprotein, cholera toxin B, mannose-6-phosphate, transferrin, riboflavin, the tripeptide RGD, ICAM-1 antibody and nicotinic acid (Bareford and Swaan 2007). Perhaps the most widely used endocytic targeting ligands for the functionalizing of nanoscale drug delivery systems are transferrins (comprehensively reviewed in (Qian et al. 2002), a family of large nonheme iron-binding glycoproteins. The efficient cellular uptake of transferrins (Tf) has been and still is being explored for the intracellular delivery of anticancer agents, and also proteins and therapeutic genes. Iron-loaded transferrin binds to a specific cell-surface receptor (TfR1) and upon endocytosis via clathrin-coated pits the transferrin-receptor complex is routed into the endosomal compartment avoiding lysosomal digestion. This is an important feature of TfR1 for drug delivery, since normally glycoproteins taken up via receptor-mediated endocytosis are destined to eventually fuse with lysosomes. Such intracellular sorting of endocytosed transferrin from other endocytosed asialoglycoprotein has been found to occur immediately after cell internalization (Stoorvogel et al. 1987). Following loss of the clathrin coat, the endosome containing the Tf-TfR1 complex then starts taking up protons which causes the quick acidification of the lysosomal lumen to a pH of around 5.5. Recently a homologue to TfR1 was cloned, called TfR2 (Trinder and Baker 2003). Of importance for anticancer drug delivery, TfR2 was found to be frequently expressed in human cancer cell lines (Calzolari et al. 2007). Encapsulation of doxorubicin into liposomes bearing transferrin on the distal end of liposomal polyethylene glycol (PEG) chains resulted in significantly increased doxorubicin uptake into glioma cells, which are known to overexpress the transferrin receptor with the extent of overexpression correlated to the severity of the tumor (Eavarone et al. 2000). Transferrin modification of Doxorubicin-loaded palmitoylated glycol chitosan (GCP) vesicles resulted in higher uptake and increased cytotoxicity as compared to GCP Doxorubicin alone (Dufes et al. 2004). Tf vesicles were taken up rapidly with a plateau after 1–2 h and Doxorubicin reached the nucleus after 60–90 min.
Low-density lipoprotein (LDL) represents another endocytic targeting ligand. Furthermore, LDL itself actually provides a highly versatile natural nanoplatform for the delivery of diagnostic and therapeutic agents to normal and neoplastic cells that over express LDL receptors (LDLR) (Glickson et al. 2008, 2009). LDL-loading of contrast or therapeutic agents has been achieved by covalent attachment to protein side chains, intercalation into the phospholipid monolayer and extraction and reconstitution of the triglyceride/cholesterol ester core (Zheng et al. 2005). Glickson and coworkers have constructed a semi-synthetic nanoparticle by coating magnetite iron oxide nanoparticles with carboxylated cholesterol and overlaying a monolayer of phospholipid to which Apo A1, Apo E or synthetic amphoteric alpha-helical polypeptides were adsorbed for targeting HDL, LDL or folate receptors, respectively (Zheng et al. 2002). These semisynthetic particles have potential utility for the in situ loading of magnetite into cells for magnetic resonance imaging (MRI) monitored cell tracking or gene therapy (Zheng et al. 2005). In addition to the surface ligand, carrier geometry also might play a role in the endocytic process (Muro et al. 2008). Disks were found to display longer half-lives in circulation and higher targeting specificity in mice, whereas spheres underwent a more rapid endocytosis. Most interestingly from the aspect of intracellular drug delivery it was also found that the size of the carrier might determine its intracellular fate. While micron-size carriers had prolonged residency in prelysosomal compartments, submicron carriers trafficked more readily to lysosomes (Muro et al. 2008).
In addition to endolysosomal targeting there is a growing body of work that suggests the feasibility of modifying nanocarriers to redirect delivery of their cargo to other sub-cellular compartments. A fusigenic viral liposome fused with the Hemagglutinating virus of Japan (HVJ) envelope protein was used to efficiently encapsulate and deliver DNA to the cytoplasm through fusion of the liposome with the plasma membrane (Dzau et al. 1996). In another study, pH sensitive liposomes composed of dioleylphosphatidylethanolamine and cholestryl hemisuccinate have been reported to be efficient carriers of N-butylnojirimycin to the endoplasmic reticulum (ER) (Costin et al. 2002); this can potentially be applied to the treatment of melanoma. Surface functionalization with ligands like cholera toxin, shiga toxin due to which the nanocarrier will be internalized into the cell by caveolae or lipid raft mediated endocytosis surpasses the lysosome and delivers the moiety to the ER or golgi complex (Le and Nabi 2003; Tarrago-Trani and Storrie 2007). Such ER liposomes may be used for the delivery of anti virals (Pollock et al. 2010). On the other hand, presence of an ER retrieval sequence on a ligand may confer to it ER targeting abilities. Acyl coenzyme-A binding protein (ACBP) is a ligand consisting of a potential ER retrieval signal, a dilysine (KK) motif near its C-terminus, which was found, by live cell imaging and indirect immunohistochemistry, to preferentially accumulate in the endoplasmic reticulum and the golgi complex (Hansen et al. 2008). Liposomes modified with mitochondriotropic ligands have been shown to improve the efficacy of an anticancer drug both in vitro and in vivo (Boddapati et al. 2008). To render liposomes mitochondria-specific, the liposomal surface was modified with triphenyl phosphonium (TPP) cations (Boddapati et al. 2005). Methyltriphenylphosphonium cations (MTPP) are rapidly taken up by mitochondria in living cells (Liberman et al. 1969) and have been extensively explored for the delivery of biological active molecules to and into mitochondria (Murphy 2008; Murphy and Smith 2007; Ross et al. 2008; Smith et al. 2003). The replacement of the methyl group in MTPP with a stearyl residue was shown to facilitate the attachment of TPP cations to the surface of liposomes (Boddapati et al. 2005).
Solid nanoparticles prepared from polymers or colloidal metals also fall under the umbrella of pharmaceutical nanocarriers. Gold nanoparticles (AuNPs) are a flexible nanoscale platform for the conjugation of a variety of targeting ligands based on the affinity of thiol and amino groups for the gold surface. Of particular interest here is the report of the conjugation of the triphenyl phosphonium mitochondriotropic ligand (Horobin et al. 2007) to the surface of AuNPs (Ju-Nam et al. 2006). Triphenyl-phosphonioalkylthiosulfate and potassium tetrachloroaurate were dissolved in dichloromethane followed by drop-wise addition into an aqueous solution of sodium borohydride to generate 5–10 nm sized AuNPs with surface-attached triphenylphosphonium residues. While data describing the intracellular localization of these potentially mitochondriotropic AuNP’s have not yet been made available, AuNPs have already been targeted to the nucleus using the adenoviral nuclear localization signal (NLS) and integrin binding domain (Tkachenko et al. 2004). Such an approach has been reported to be useful in the development of probes for cell tracking by surface enhanced raman scattering. Gold nanoparticles were surface functionalized with the SV40 NLS, which led to accumulation of the nanoparticles in the nucleus of HeLa cells (Xie et al. 2009). Quantum dots conjugated to the amino group of the mitochondrial targeting sequence, Mito-8 were reported to be colocalized in the mitochondria (Hoshino et al. 2004). Chlorotoxin conjugated iron oxide nanoparticles coated with a copolymer of chitosan-PEG-PEI were found to specifically translocate to the nucleus of cancer cells permitting a dual targeting approach to a specific cell type and an organelle within the cell type; this approach was successful in vivo, both for imaging and therapeutic purposes Kievit (2010).
Modification with a leader sequence peptide has also been applied to creating delivery systems for mitochondria. A mitochondrial leader peptide (MLP), derived from the nucleo-cytosol expressed but mitochondria localized ornithine transcarbamylase was recently used to render polyethylenimine (PEI) mitochondriotropic (Lee et al. 2007). PEI had been developed in the mid 1990s as a versatile vector for gene and oligonucleotide transfer into cells in culture and in vivo (Boussif et al. 1995; Demeneix and Behr 2005). Lee et al. (2007) conjugated the mitochondrial leader peptide to PEI via a disulfide bond and confirmed the complex formation of PEI-MLP with DNA by a gel retardation assay. In vitro delivery tests of rhodamine-labeled DNA into living cells demonstrated that PEI-MLP/DNA complexes were localized at mitochondrial sites in contrast to controls carried out with PEI-DNA complexes lacking MLP. The author’s data suggest that PEI-MLP can deliver DNA to the mitochondrial sites and may be useful for the development of direct mitochondrial gene therapy, a strategy for the cure of mitochondrial DNA diseases proposed earlier (Seibel et al. 1995; Weissig and Torchilin 2000, 2001a, b) as an alternative to allotropic expression (Ellouze et al. 2008; Oca-Cossio et al. 2003; Zullo 2001; Gray et al. 1996).
3.2 Nanocarriers Prepared from Self-assembling Molecules with Known Sub-cellular Accumulation
All the examples discussed in the previous section share a common assumption that unless a targeting ligand is incorporated into the design, the nanocarriers would remain in the endolysosomal compartment. However it is interesting to also consider the disposition of a nanocarrier made exclusively of a molecule with a predisposition for a sub-cellular compartment. A good example of a molecule that has a strong affinity for a sub-cellular compartment and that is also capable of self assembling to form a potential carrier system is the mitochondriotropic amphiphile dequalinium chloride. Vesicles prepared exclusively from dequalinium chloride (called DQAsomes) have been explored for the delivery of several bioactive molecules to mitochondria (D’Souza et al. 2003, 2005, 2008; Weissig et al. 2000, 2001). Most recently, the antitumor efficiency of DQAsomal encapsulated paclitaxel was enhanced by modifying the DQAsomal surface with folic acid (FA) (Vaidya et al. 2009). However, DQAsomes currently serve as a preliminary proof of concept and are still far from being the perfect delivery system. In order to design similar carriers for other sub-cellular compartments it is necessary to find suitable self-assembling molecules with an affinity for the intended sub-cellular compartment. To this end, recent work on the sub-cellular distribution of micelle forming agents offers some interesting insights (Bae et al. 2005; Maysinger et al. 2007; Savic et al. 2006, 2009; Xiong et al. 2008).
Imaging studies based on the use of a variety of organelle-specific dyes, gold and fluorescent polymers have provided detailed insight into the sub-cellular distribution of block copolymer micelles (Maysinger et al. 2007; Savic et al. 2003). Both imaging techniques, i.e. confocal fluorescence microscopy (to detect the fluorophore-labeled copolymers) and transmission electron microscopy (to detect the gold-labeled copolymers) demonstrate that poly(caprolactone)-b-poly(ethylene oxide) micelles (PCL-b-PEO micelles) do not enter the nucleus. With respect to the cytosolic distribution of PCL-b-PEO micelles, however, the two different imaging techniques used in these studies suggest quite a different sub-cellular disposition. TEM images show most of the gold labeled micelles to be localized in endosomes/lysosomes and a few of them were seen at or in mitochondria (Maysinger et al. 2007). Confocal fluorescence microscopic images, on the other hand, show fluorescent PCL-b-PEO micelles almost evenly distributed throughout the cytosol (Savic et al. 2003). Therefore, not surprisingly, cell staining with organelle-specific dyes and overlaying the corresponding confocal fluorescence images reveal partial colocalization of PCL-b-PEO micelles with lysosomes, with the Golgi apparatus and the Endoplasmic Reticulum, with the mitochondria and the Endoplasmic Reticulum and with mitochondria alone. Considering the nature of the micelle corona, which is entirely made up of non-functionalized polyethylene oxide, a highly hydrophilic polymer, any specific interaction with or any specific affinity for any of the cell organelles could not be expected per se. It would be very interesting to see to what extent modifying the micelle corona with organelle-specific ligands would alter the intracellular distribution of such micelles, which then potentially could become nanocontainers that distribute cargo to defined cytoplasmic organelles. However, the distinctive distribution of nonfunctionalized PCL-b-PEO micelles throughout the cytosol makes them highly suitable for multiple cytoplasmic targeting (Savic et al. 2003), which has most recently been proven to be relevant for the delivery of effector molecules of the cell signaling pathways that are activated in the cytosol (Savic et al. 2009). This study suggests that micelle-based intracellular delivery of potent, poorly water-soluble, cell-death-pathway inhibitors may represent a useful addition to established delivery of cytocidal block-copolymer micelle-incorporated bioactives (Savic et al. 2009).
4 Conclusion and Perspectives
From the examples discussed so far it would seem that nanocarrier systems could be designed to achieve true molecular level targeting inside cells. However to say that these systems will be available soon is perhaps premature given what little is currently known about the sub-cellular dynamics associated with nanocarrier trafficking. There are several unanswered questions. For example, do all nanocarriers remain intact upon cell entry and subsequent disposition? Are there differences in the disposition of vesicles in comparison to solid particles? What is the true influence of size on the intracellular disposition of various nanocarriers? Most important is however the question of the mechanism by which the nanocarrier is able to achieve selective uptake and delivery into the sub-cellular compartment. Despite several reports of altered or improved sub-cellular accumulation associated with an improvement in activity, it is still unclear how exactly this happens. It will take much active research to answer these and other questions before sub-cellular targeted nanocarrier-based therapies become commonplace.
References
Akita, H., et al., Multi-layered nanoparticles for penetrating the endosome and nuclear membrane via a step-wise membrane fusion process. Biomaterials, 2009. 30(15): p. 2940–9.
Allen, T.D., et al., The nuclear pore complex: mediator of translocation between nucleus and cytoplasm. J Cell Sci, 2000. 113 (Pt 10): p. 1651–9.
Bae, Y., et al., Preparation and biological characterization of polymeric micelle drug carriers with intracellular pH-triggered drug release property: tumor permeability, controlled subcellular drug distribution, and enhanced in vivo antitumor efficacy. Bioconjug Chem, 2005. 16(1): p. 122–30.
Bareford, L.M. and P.W. Swaan, Endocytic mechanisms for targeted drug delivery. Adv Drug Deliv Rev, 2007. 59(8): p. 748–58.
Boddapati, S.V., et al., Mitochondriotropic liposomes. J Liposome Res, 2005. 15(1–2): p. 49–58.
Boddapati, S.V., et al., Organelle-targeted nanocarriers: specific delivery of liposomal ceramide to mitochondria enhances its cytotoxicity in vitro and in vivo. Nano Lett, 2008. 8(8): p. 2559–63.
Boussif, O., et al., A versatile vector for gene and oligonucleotide transfer into cells in culture and in vivo: polyethylenimine. Proc Natl Acad Sci U S A, 1995. 92(16): p. 7297–301.
Breunig, M., S. Bauer, and A. Goepferich, Polymers and nanoparticles: intelligent tools for intracellular targeting? Eur J Pharm Biopharm, 2008. 68(1): p. 112–28.
Calzolari, A., et al., Transferrin receptor 2 is frequently expressed in human cancer cell lines. Blood Cells Mol Dis, 2007. 39(1): p. 82–91.
Castino, R., M. Demoz, and C. Isidoro, Destination ‘lysosome’: a target organelle for tumour cell killing? J Mol Recognit, 2003. 16(5): p. 337–48.
Cervetti, G., et al., Efficacy and toxicity of liposomal daunorubicin included in PVABEC regimen for aggressive NHL of the elderly. Leuk Lymphoma, 2003. 44(3): p. 465–9.
Costin, G.E., et al., pH-sensitive liposomes are efficient carriers for endoplasmic reticulum-targeted drugs in mouse melanoma cells. Biochem Biophys Res Commun, 2002. 293(3): p. 918–23.
Demeneix, B. and J.P. Behr, Polyethylenimine (PEI). Adv Genet, 2005. 53: p. 217–30.
D’Souza, G.G., et al., DQAsome-mediated delivery of plasmid DNA toward mitochondria in living cells. J Control Release, 2003. 92(1–2): p. 189–97.
D’Souza, G.G., S.V. Boddapati, and V. Weissig, Mitochondrial leader sequence--plasmid DNA conjugates delivered into mammalian cells by DQAsomes co-localize with mitochondria. Mitochondrion, 2005. 5(5): p. 352–8.
D’Souza, G.G., et al., Nanocarrier-assisted sub-cellular targeting to the site of mitochondria improves the pro-apoptotic activity of paclitaxel. J Drug Target, 2008. 16(7): p. 578–85.
Dufes, C., et al., Anticancer drug delivery with transferrin targeted polymeric chitosan vesicles. Pharm Res, 2004. 21(1): p. 101–7.
Duvvuri, M. and J.P. Krise, Intracellular drug sequestration events associated with the emergence of multidrug resistance: a mechanistic review. Front Biosci, 2005. 10: p. 1499–509.
Dzau, V.J., et al., Fusigenic viral liposome for gene therapy in cardiovascular diseases. Proc Natl Acad Sci U S A, 1996. 93(21): p. 11421–5.
Eavarone, D.A., X. Yu, and R.V. Bellamkonda, Targeted drug delivery to C6 glioma by transferrin-coupled liposomes. J Biomed Mater Res, 2000. 51(1): p. 10–4.
Ellis, R.J. and A.P. Minton, Cell biology: join the crowd. Nature, 2003. 425(6953): p. 27–8.
Ellouze, S., et al., Optimized allotopic expression of the human mitochondrial ND4 prevents blindness in a rat model of mitochondrial dysfunction. Am J Hum Genet, 2008. 83(3): p. 373–87.
Fang, J., T. Sawa, and H. Maeda, Factors and mechanism of “EPR” effect and the enhanced antitumor effects of macromolecular drugs including SMANCS. Adv Exp Med Biol, 2003. 519: p. 29–49.
Fawell, S., et al., Tat-mediated delivery of heterologous proteins into cells. Proc Natl Acad Sci U S A, 1994. 91(2): p. 664–8.
Fernandez-Carneado, J., et al., Highly efficient, nonpeptidic oligoguanidinium vectors that selectively internalize into mitochondria. J Am Chem Soc, 2005. 127(3): p. 869–74.
Fittipaldi, A. and M. Giacca, Transcellular protein transduction using the Tat protein of HIV-1. Adv Drug Deliv Rev, 2005. 57(4): p. 597–608.
Gabizon, A., et al., Systemic administration of doxorubicin-containing liposomes in cancer patients: a phase I study. Eur J Cancer Clin Oncol, 1989. 25(12): p. 1795–803.
Gabizon, A., R. Shiota, and D. Papahadjopoulos, Pharmacokinetics and tissue distribution of doxorubicin encapsulated in stable liposomes with long circulation times. J Natl Cancer Inst, 1989. 81(19): p. 1484–8.
Gill, P.S., et al., Randomized phase III trial of liposomal daunorubicin versus doxorubicin, bleomycin, and vincristine in AIDS-related Kaposi’s sarcoma. J Clin Oncol, 1996. 14(8): p. 2353–64.
Glickson, J.D., et al., Lipoprotein nanoplatform for targeted delivery of diagnostic and therapeutic agents. Mol Imaging, 2008. 7(2): p. 101–10.
Glickson, J.D., et al., Lipoprotein nanoplatform for targeted delivery of diagnostic and therapeutic agents. Adv Exp Med Biol, 2009. 645: p. 227–39.
Goins, A.B., H. Sanabria, and M.N. Waxham, Macromolecular crowding and size effects on probe microviscosity. Biophys J, 2008. 95(11): p. 5362–73.
Gray, R.E., et al., Allotopic expression of mitochondrial ATP synthase genes in nucleus of Saccharomyces cerevisiae. Methods Enzymol, 1996. 264: p. 369–89.
Gregoriadis, G. and B.E. Ryman, Liposomes as carriers of enzymes or drugs: a new approach to the treatment of storage diseases. Biochem J, 1971. 124(5): p. 58P.
Greish, K., Enhanced permeability and retention (EPR) effect for anticancer nanomedicine drug targeting. Methods Mol Biol, 2010. 624: p. 25–37.
Hansen, J.S., et al., Acyl-CoA-binding protein (ACBP) localizes to the endoplasmic reticulum and Golgi in a ligand-dependent manner in mammalian cells. Biochem J, 2008. 410(3): p. 463–72.
Horobin, R.W., S. Trapp, and V. Weissig, Mitochondriotropics: a review of their mode of action, and their applications for drug and DNA delivery to mammalian mitochondria. J Control Release, 2007. 121(3): p. 125–36.
Hoshino, A., et al., Quantum dots targeted to the assigned organelle in living cells. Microbiol Immunol, 2004. 48(12): p. 985–94.
Inoki, Y., et al., Proteoliposomes colocalized with endogenous mitochondria in mouse fertilized egg. Biochem Biophys Res Commun, 2000. 278(1): p. 183–91.
Ju-Nam, Y., et al., Phosphonioalkylthiosulfate zwitterions--new masked thiol ligands for the formation of cationic functionalised gold nanoparticles. Org Biomol Chem, 2006. 4(23): p. 4345–51.
Kaufmann, A.M. and J.P. Krise, Lysosomal sequestration of amine-containing drugs: analysis and therapeutic implications. J Pharm Sci, 2007. 96(4): p. 729–46.
Kievit, F.M., et al., Chlorotoxin labeled magnetic nanovectors for targeted gene delivery to glioma. ACS Nano, 2010. 4(8): p. 4587–94.
Le, P.U. and I.R. Nabi, Distinct caveolae-mediated endocytic pathways target the Golgi apparatus and the endoplasmic reticulum. J Cell Sci, 2003. 116(Pt 6): p. 1059–71.
Lee, M., et al., DNA delivery to the mitochondria sites using mitochondrial leader peptide conjugated polyethylenimine. J Drug Target, 2007. 15(2): p. 115–22.
Li, S.D. and L. Huang, Pharmacokinetics and biodistribution of nanoparticles. Mol Pharm, 2008. 5(4): p. 496–504.
Liberman, E.A., et al., Mechanism of coupling of oxidative phosphorylation and the membrane potential of mitochondria. Nature, 1969. 222(5198): p. 1076–8.
Maeda, H., et al., Tumor vascular permeability and the EPR effect in macromolecular therapeutics: a review. J Control Release, 2000. 65(1–2): p. 271–84.
Martin, M.E. and K.G. Rice, Peptide-guided gene delivery. AAPS J, 2007. 9(1): p. E18–29.
Maysinger, D., et al., Fate of micelles and quantum dots in cells. Eur J Pharm Biopharm, 2007. 65(3): p. 270–81.
Minton, A.P., How can biochemical reactions within cells differ from those in test tubes? J Cell Sci, 2006. 119(Pt 14): p. 2863–9.
Muro, S., E.H. Schuchman, and V.R. Muzykantov, Lysosomal enzyme delivery by ICAM-1-targeted nanocarriers bypassing glycosylation- and clathrin-dependent endocytosis. Mol Ther, 2006. 13(1): p. 135–41.
Muro, S., et al., Control of endothelial targeting and intracellular delivery of therapeutic enzymes by modulating the size and shape of ICAM-1-targeted carriers. Mol Ther, 2008. 16(8): p. 1450–8.
Murphy, M.P., Targeting lipophilic cations to mitochondria. Biochim Biophys Acta, 2008. 1777(7–8): p. 1028–31.
Murphy, M.P. and R.A. Smith, Targeting antioxidants to mitochondria by conjugation to lipophilic cations. Annu Rev Pharmacol Toxicol, 2007. 47: p. 629–56.
Northfelt, D.W., et al., Doxorubicin encapsulated in liposomes containing surface-bound polyethylene glycol: pharmacokinetics, tumor localization, and safety in patients with AIDS-related Kaposi’s sarcoma. J Clin Pharmacol, 1996. 36(1): p. 55–63.
Oca-Cossio, J., et al., Limitations of allotopic expression of mitochondrial genes in mammalian cells. Genetics, 2003. 165(2): p. 707–20.
Panyam, J. and V. Labhasetwar, Biodegradable nanoparticles for drug and gene delivery to cells and tissue. Adv Drug Deliv Rev, 2003. 55(3): p. 329–47.
Patil, Y. and J. Panyam, Polymeric nanoparticles for siRNA delivery and gene silencing. Int J Pharm, 2009. 367(1–2): p. 195–203.
Pollock, S., et al., Uptake and trafficking of liposomes to the endoplasmic reticulum. FASEB J, 2010. 24(6): p. 1866–78.
Qian, Z.M., et al., Targeted drug delivery via the transferrin receptor-mediated endocytosis pathway. Pharmacol Rev, 2002. 54(4): p. 561–87.
Rajendran, L., et al., Subcellular targeting strategies for drug design and delivery. Nat Rev Drug Discovery, 2010. 9(1): p. 29–42.
Ross, M.F., et al., Rapid and extensive uptake and activation of hydrophobic triphenylphosphonium cations within cells. Biochem J, 2008. 411(3): p. 633–45.
Rousselle, C., et al., New advances in the transport of doxorubicin through the blood-brain barrier by a peptide vector-mediated strategy. Mol Pharmacol, 2000. 57(4): p. 679–86.
Ruan, G., et al., Imaging and tracking of tat peptide-conjugated quantum dots in living cells: new insights into nanoparticle uptake, intracellular transport, and vesicle shedding. J Am Chem Soc, 2007. 129(47): p. 14759–66.
Savic, R., et al., Micellar nanocontainers distribute to defined cytoplasmic organelles. Science, 2003. 300(5619): p. 615–8.
Savic, R., et al., Assessment of the integrity of poly(caprolactone)-b-poly(ethylene oxide) micelles under biological conditions: a fluorogenic-based approach. Langmuir, 2006. 22(8): p. 3570–8.
Savic, R., et al., Block-copolymer micelles as carriers of cell signaling modulators for the inhibition of JNK in human islets of Langerhans. Biomaterials, 2009.
Schwarze, S.R., et al., In vivo protein transduction: delivery of a biologically active protein into the mouse. Science, 1999. 285(5433): p. 1569–72.
Seibel, P., et al., Transfection of mitochondria: strategy towards a gene therapy of mitochondrial DNA diseases. Nucleic Acids Res, 1995. 23(1): p. 10–7.
Seksek, O., J. Biwersi, and A.S. Verkman, Translational diffusion of macromolecule-sized solutes in cytoplasm and nucleus. J Cell Biol, 1997. 138(1): p. 131–42.
Sharma, A., U.S. Sharma, and R.M. Straubinger, Paclitaxel-liposomes for intracavitary therapy of intraperitoneal P388 leukemia. Cancer Lett, 1996. 107(2): p. 265–72.
Sharma, A., et al., Activity of paclitaxel liposome formulations against human ovarian tumor xenografts. Int J Cancer, 1997. 71(1): p. 103–7.
Shiraishi, T. and P.E. Nielsen, Enhanced delivery of cell-penetrating peptide-peptide nucleic acid conjugates by endosomal disruption. Nat Protoc, 2006. 1(2): p. 633–6.
Smith, R.A., et al., Delivery of bioactive molecules to mitochondria in vivo. Proc Natl Acad Sci U S A, 2003. 100(9): p. 5407–12.
Song, W.J., et al., Gold nanoparticles capped with polyethyleneimine for enhanced siRNA delivery. Small, 2010. 6(2): 239–46.
Stathopoulos, G.P.,Liposomal Cisplatin: a new cisplatin formulation. Anticancer Drugs, 2010. 21(8): p. 732–6.
Stoffler, D., B. Fahrenkrog, and U. Aebi, The nuclear pore complex: from molecular architecture to functional dynamics. Curr Opin Cell Biol, 1999. 11(3): p. 391–401.
Stoorvogel, W., H.J. Geuze, and G.J. Strous, Sorting of endocytosed transferrin and asialoglycoprotein occurs immediately after internalization in HepG2 cells. J Cell Biol, 1987. 104(5): p. 1261–8.
Suh, J., et al., PEGylation of nanoparticles improves their cytoplasmic transport. Int J Nanomedicine, 2007. 2(4): p. 735–41.
Tahara, K., et al., Chitosan-modified poly(D,L-lactide-co-glycolide) nanospheres for improving siRNA delivery and gene-silencing effects. Eur J Pharm Biopharm, 2010. 74(3): p. 421–6.
Tarrago-Trani, M.T. and B. Storrie, Alternate routes for drug delivery to the cell interior: pathways to the Golgi apparatus and endoplasmic reticulum. Adv Drug Deliv Rev, 2007. 59(8): p. 782–97.
Tate, B.A. and P.M. Mathews, Targeting the role of the endosome in the pathophysiology of Alzheimer’s disease: a strategy for treatment. Sci Aging Knowledge Environ, 2006. 2006(10): p. re2.
Tkachenko, A.G., et al., Cellular trajectories of peptide-modified gold particle complexes: comparison of nuclear localization signals and peptide transduction domains. Bioconjug Chem, 2004. 15(3): p. 482–90.
Torchilin, V.P., Recent approaches to intracellular delivery of drugs and DNA and organelle targeting. Annu Rev Biomed Eng, 2006. 8: p. 343–75.
Torchilin, V.P., et al., TAT peptide on the surface of liposomes affords their efficient intracellular delivery even at low temperature and in the presence of metabolic inhibitors. Proc Natl Acad Sci U S A, 2001. 98(15): p. 8786–91.
Torchilin, V.P., Nanotechnology for Intracellular Delivery and Targeting, in Nanotechnology in Drug Delivery, M.M. de Villiers, G.S. Kwon, and P. Aramwit, Editors. 2009, Springer Publications: New York. p. 313–348.
Trinder, D. and E. Baker, Transferrin receptor 2: a new molecule in iron metabolism. Int J Biochem Cell Biol, 2003. 35(3): p. 292–6.
Vaidya, B.P., R., Rai, S., Khatri, K., Goyal, A.K., Mishra, N., Vyas, S.P., Cell-selective mitochondrial targeting: A new approach for cancer therapy. Cancer Therapy, 2009. 7: p. 141–148.
Vale, R.D., Intracellular transport using microtubule-based motors. Annu Rev Cell Biol, 1987. 3: p. 347–78.
Walter, P., et al., The protein translocation machinery of the endoplasmic reticulum. Philos Trans R Soc Lond B Biol Sci, 1982. 300(1099): p. 225–8.
Weissig, V., Mitochondrial-targeted drug and DNA delivery. Crit Rev Ther Drug Carrier Syst, 2003. 20(1): p. 1–62.
Weissig, V., Targeted drug delivery to mammalian mitochondria in living cells. Expert Opin Drug Deliv, 2005. 2(1): p. 89–102.
Weissig, V. and V.P. Torchilin, Mitochondriotropic cationic vesicles: a strategy towards mitochondrial gene therapy. Curr Pharm Biotechnol, 2000. 1(4): p. 325–46.
Weissig, V. and V.P. Torchilin, Towards mitochondrial gene therapy: DQAsomes as a strategy. J Drug Target, 2001. 9(1): p. 1–13.
Weissig, V. and V.P. Torchilin, Cationic bolasomes with delocalized charge centers as mitochondria-specific DNA delivery systems. Adv Drug Deliv Rev, 2001. 49(1–2): p. 127–49.
Weissig, V., C. Lizano, and V.P. Torchilin, Selective DNA release from DQAsome/DNA complexes at mitochondria-like membranes. Drug Deliv, 2000. 7(1): p. 1–5.
Weissig, V., G.G. D’Souza, and V.P. Torchilin, DQAsome/DNA complexes release DNA upon contact with isolated mouse liver mitochondria. J Control Release, 2001. 75(3): p. 401–8.
Weissig, V., S.M. Cheng, and G.G. D’Souza, Mitochondrial pharmaceutics. Mitochondrion, 2004. 3(4): p. 229–44.
Weissig, V., et al., Mitochondria-specific nanotechnology. Nanomed, 2007. 2(3): p. 275–85.
Working, P.K., et al., Reduction of the cardiotoxicity of doxorubicin in rabbits and dogs by encapsulation in long-circulating, pegylated liposomes. J Pharmacol Exp Ther, 1999. 289(2): p. 1128–33.
Xie, W., et al., Nuclear targeted nanoprobe for single living cell detection by surface-enhanced Raman scattering. Bioconjug Chem, 2009. 20(4): p. 768–73.
Xiong, X.B., et al., Multifunctional polymeric micelles for enhanced intracellular delivery of doxorubicin to metastatic cancer cells. Pharm Res, 2008. 25(11): p. 2555–66.
Xu, Z.P., et al., Subcellular compartment targeting of layered double hydroxide nanoparticles. J Control Release, 2008. 130(1): p. 86–94.
Yamada, Y. and H. Harashima, Mitochondrial drug delivery systems for macromolecule and their therapeutic application to mitochondrial diseases. Adv Drug Deliv Rev, 2008. 60(13–14): p. 1439–62.
Yamada, Y., et al., MITO-Porter: A liposome-based carrier system for delivery of macromolecules into mitochondria via membrane fusion. Biochim Biophys Acta, 2008. 1778(2): p. 423–32.
Yang, T., et al., Enhanced solubility and stability of PEGylated liposomal paclitaxel: in vitro and in vivo evaluation. Int J Pharm, 2007. 338(1–2): p. 317–26.
Yessine, M.A. and J.C. Leroux, Membrane-destabilizing polyanions: interaction with lipid bilayers and endosomal escape of biomacromolecules. Adv Drug Deliv Rev, 2004. 56(7): p. 999–1021.
Yuan, X., et al., SiRNA drug delivery by biodegradable polymeric nanoparticles. J Nanosci Nanotechnol, 2006. 6(9–10): p. 2821–8.
Zanta, M.A., P. Belguise-Valladier, and J.P. Behr, Gene delivery: a single nuclear localization signal peptide is sufficient to carry DNA to the cell nucleus. Proc Natl Acad Sci USA, 1999. 96(1): p. 91–6.
Zheng, G., et al., Low-density lipoprotein reconstituted by pyropheophorbide cholesteryl oleate as target-specific photosensitizer. Bioconjug Chem, 2002. 13(3): p. 392–6.
Zheng, G., et al., Rerouting lipoprotein nanoparticles to selected alternate receptors for the targeted delivery of cancer diagnostic and therapeutic agents. Proc Natl Acad Sci USA, 2005. 102(49): p. 17757–62.
Zullo, S.J., Gene therapy of mitochondrial DNA mutations: a brief, biased history of allotopic expression in mammalian cells. Semin Neurol, 2001. 21(3): p. 327–35.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2011 Springer Science+Business Media B.V.
About this chapter
Cite this chapter
Solomon, M., D’Souza, G.G.M. (2011). Approaches to Achieving Sub-cellular Targeting of Bioactives Using Pharmaceutical Nanocarriers. In: Prokop, A. (eds) Intracellular Delivery. Fundamental Biomedical Technologies, vol 5. Springer, Dordrecht. https://doi.org/10.1007/978-94-007-1248-5_2
Download citation
DOI: https://doi.org/10.1007/978-94-007-1248-5_2
Published:
Publisher Name: Springer, Dordrecht
Print ISBN: 978-94-007-1247-8
Online ISBN: 978-94-007-1248-5
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)