
Abstract
Survival in microhabitats that experience extreme fluctuations in water availability and temperature requires extreme adaptations. Antonie van Leeuwenhoek was the first who describe the phenomenon of the resurrection of a desiccated rotifer in 1702. As with some rotifers and other small organisms, tardigrades enter a desiccated state known as anhydrobiosis to withstand such environmental conditions. This allows them to cope with the temporal variation of available water and to extend their lifespan in an anhydrobiotic state by up to 20 years without biological aging, according to the Sleeping Beauty hypothesis. Heat shock proteins serve as molecular chaperones to preserve or restore protein integrity, and tardigrade-specific intrinsically disordered proteins (TDPs) as well as metabolite help prevent the formation of damaging cellular compartments aggregates during water stress.
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others


10.1 Life Without Water
The first to describe the phenomenon of the resurrection of a desiccated animal was the Dutch naturalist Antonie van Leeuwenhoek. He discovered that when dry and apparently lifeless dust from a roof gutter was rehydrated with clean water in a small glass, many “animalcules” (small organisms) became active within an hour (van Leeuwenhoek 1702). He noticed: “I confess, I never thought that there could be any living creature in a substance so dried as this was.” These animals were probably of the bdelloid rotifer species Philodina roseola (Tunnacliffe and Lapinski 2003). In the past 300 years, various names were used for this phenomenon until David Keilin, who is most known for his research and rediscovery of cytochrome in the 1920s, published a benchmark review and defined the term cryptobiosis. He called it “the state of an organism when it shows no visible signs of life and when its metabolic activity becomes hardly measurable, or comes reversibly to a standstill” (Keilin 1959). Of course, the difference between a “hardly measurable” metabolism and one that is at a “reversible standstill” is of considerable significance. The latter is difficult to interpret, because it ultimately means neither alive nor dead. Due to the fact that, for example, anhydrobiotic organism contains no or only a little water means that also no functional enzymes are available, and therefore no metabolism is possible. Despite this circumstance, the anhydrobiotes are able to survive desiccation and to continue to live normally after rehydration. Cryptobiosis results from desiccation (anhydrobiosis), low temperature (cryobiosis), lack of oxygen (anoxybiosis), high or low salt concentration (osmobiosis), or combinations of these (Keilin 1959). However, there are always similarities between these kinds of cryptobiotic states, for example, between the state of anhydrobiosis and cryobiosis. In both states, the amount of free water in the cells is reduced by desiccation or ice formation. Nevertheless, various studies suggest that the two conditions are not equivalent and that different mechanisms of survival have been developed (Crowe et al. 1992). The ability to enter cryptobiosis is quite common in nature. It occurs in many invertebrate taxa like sponges, crustaceans, rotifers, nematodes, and tardigrades (Crowe and Clegg 1973, 1978; Lapinski and Tunnacliffe 2003; Womersley 1987; Wright 2001). But also many prokaryotes such as bacteria and blue-green algae (Potts 2001), plant seeds (Alpert 2000; Chandler and Bartels 1999; Ingram and Bartels 1996; Priestley 1986; Vertucci and Farrant 1995), and also tissues of some higher plants have developed this ability. Particularly noteworthy here is the resurrection plant Selaginella lepidophylla, also known as the “Rose of Jericho” (Scott 2000; Tomos 1992).
10.2 Tardigrades and Their Longevity
10.2.1 Longevity in Anhydrobiosis
To date, few comparative studies have been carried out to obtain information on how long anhydrobiotes can remain in anhydrobiosis without losing their vitality after rehydration. Most studies were performed in the first half of the twentieth century (Baumann 1927; Fielding 1951; Franceschi 1948; Goodey 1923; Lee 1961; Rahm 1923; Steiner and Albin 1946). More information is available about parasitic nematodes of plants. For example, Steiner and Albin (1946) reported on two nematode species that successfully survived 28 and 30 years, respectively, in anhydrobiosis. Fielding (1951) also reported a nematode species that survived dehydrated for 20–28 years. Goodey (1923) and Lee (1961) in turn found a maximum time span of 9 and 10 years, respectively. There is much less information about tardigrades (Fig. 10.1a, b) and rotifers. One of the first who did extensive experiments was Baumann (1927). One species of the genus Macrobiotus was successfully rehydrated after 7 years (Baumann 1927). Richtersius oberhaeuseri survived up to 1604 days and Echiniscus sp. survived up to 1085 days in anhydrobiosis (Franceschi 1948). Franceschi (1948) even reported a tardigrade that came from a 120-year-old moss and showed briefly movement after rehydration before it died. Meanwhile, it is doubted whether the observed animal was alive. Presumably, passive rehydration caused the animal’s supposed movements (Jönsson and Bertolani 2001). The longest reliable documented survival time in anhydrobiosis is known from the heterotardigrade Echiniscus testudo with 20 years (Jørgensen et al. 2007).

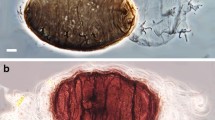
(a) SEM picture of an active tardigrade of the species Paramacrobiotus richtersi and (b) anhydrobiotic tun state
10.2.2 Sleeping Beauty Hypothesis
Very little is known about the effects of anhydrobiosis on life cycle and longevity achieved thereby. So far, little information is available, mainly about nematodes and rotifers (Ricci 2001; Ricci and Caprioli 2005; Ricci and Covino 2005; Ricci and Pagani 1997; Ricci et al. 1987; Wharton 2003; Wharton and Aalders 1999). Ricci and Pagani (1997) postulated three hypotheses of what effects anhydrobiosis could have on the life of organisms. The first hypothesis assumes that the time an animal spends in anhydrobiosis is ignored and biological aging is halted. This hypothesis is also referred to as the “Sleeping Beauty” model. The second hypothesis is that the internal clock and thus biological aging is slowed down. The third model describes unrestricted biological aging, even during anhydrobiosis. Studies on the rotifer species Macrotrachela quadricornifera and Adineta ricciae clearly demonstrated that the “Sleeping Beauty” model is correct and that the life cycle is not affected by occasional anhydrobiosis (Ricci and Caprioli 1998; Ricci et al. 1987). Even with nematodes, the internal clock seems to be stopped because, despite a short lifetime, they can be rehydrated again after months or years and can continue to live normally (Ricci and Covino 2005; Ricci et al. 1987). Since tardigrades, like nematodes and rotifers, can spend long periods in anhydrobiosis (Baumann 1927; Bertolani et al. 2004; Guidetti and Jönsson 2002), the “Sleeping Beauty” model has been investigated with the species Milnesium tardigradum (Hengherr et al. 2008). They were exposed to alternate dry periods of 7 days each compared to an active control group. The animals of the control group reached an age of 82.7 ± 2.7 days. The age of the animals exposed to the periodic drought periods, minus the times spent in anhydrobiosis, was 79.7 ± 5.7 days. The total time period including the dry periods was 133.2 ± 11.7 days. The result shows that the “Sleeping Beauty” model is valid for the tardigrades, and the time spent in anhydrobiosis is not counted as biological age (Hengherr et al. 2008). Nevertheless, there seems to be an upper limit to persist in anhydrobiosis. One possible explanation is that the anhydrobionts die during anhydrobiosis due to cellular damage from chemical aging (Schöneich 1999), since no repair mechanisms can become active in the desiccated state (Clegg 1967; Crowe and Madin 1975; Örstan 1998; Wright 2001).
10.2.3 Desiccation Tolerance in Embryonic Stages
In recent decades, a variety of experiments have shown that adult tardigrades can spend long periods, from months to several years, in the anhydrobiotic state. Bertolani et al. reported the successful hatching of four juveniles of the species Richtersius oberhaeuseri from 13 eggs, which were rehydrated after 9 years (Bertolani et al. 2004). This was the first study to show that not only adults have the ability to enter anhydrobiosis but also embryos. In the species Ramazzottius oberhaeuseri and Milnesium tardigradum, successful hatching after 1604 days of anhydrobiosis has been demonstrated (Rebecchi et al. 2006). The anhydrobiosis of embryos is considered to be an adaptation strategy, since Paramacrobiotus richtersi is also able to undergo a time-shifted hatching depending on the environmental conditions (Altiero et al. 2006). For rotifers of the species Adineta vaga (Örstan 1995) and other species (Ricci 1998), desiccation tolerance, depending on the stage of development, has already been demonstrated. The first comparative study in tardigrades has been done with the parthenogenetic tardigrade species Milnesium tardigradum which lives in microhabitats that dry out regularly (Ramazzotti and Maucci 1983; Wright 1991). It was shown that both factors, the humidity and the state of development (several stages from the blastula stage to the hatching juveniles), are decisive for desiccation tolerance (Schill and Fritz 2008). The less the embryos were developed, the lower was the survival rate for all humidities between 10 and 81%. When the developing embryos were dehydrated during the first 3 days of their development at low humidities, the survival rates were particularly low. The older they are, the better is the survival rate, especially for those which are close to hatching and which were dried slowly at high humidities (Schill and Fritz 2008). Tardigrades are thus successfully adapted to microhabitats in all stages of life, which are subject to regular dehydration and rehydration processes (Fig. 10.2).

A slow desiccation of a tardigrade results in biochemical and biophysical adaptations and a high survival rate after rehydration
10.3 Tardigrades and Stress Proteins
Heat shock reactions were first observed in 1962 as a puffing pattern in Drosophila larvae correlated with a temperature increase and later shown to produce heat shock proteins (Hsp) (Tissières et al. 1974). Although called heat shock proteins, their induction is not only due to temperature effects but to a whole series of other stressors. They play a critical role in intracellular protection against protein-denaturing factors, acting as molecular chaperones, helping to fold newly synthesized proteins, and preventing stress-induced denaturation or aggregation (proteotoxicity). Furthermore, they are involved in the renaturation and transmembrane transport of proteins. Their molecular and cellular physiological functions have been studied extensively in various fields of biology and have been the subject of numerous review articles (e.g., Feder and Hofmann 1999; Gething and Sambrook 1992; Morimoto 1993; Parsell and Lindquist 1993). Their classification is based on their respective molecular weights. The best known are the very conserved families Hp60, Hsp70, and Hsp90. Among them Hsp70 is one of the best studied families; their induction mechanism is by means of HSF (heat shock factor) and HSE (heat shock element). There is a whole range of heterogeneous low molecular weight Hsps with a molecular weight below 30 kDa (low molecular weight (LMW), which are also referred to as small Hsps).
10.3.1 Small Stress Proteins
Large amounts of two small stress proteins, artemin (Arrigo and Müller 2002; De Graaf et al. 1990) and p26 (Clegg et al. 1994, 1995), were found in stress-resistant, encysted embryos of the brine shrimp species Artemia franciscana. Both proteins have been extensively studied in the context of anhydrobiosis and osmobiosis (Chen et al. 2003, 2007; Clegg et al. 1995, 1999; Liang et al. 1997a; Liang and MacRae 1999; Warner et al. 2004; Willsie and Clegg 2002). p26 belongs to the small heat shock/α-crystallin protein family, which has molecular chaperone activity in vitro (Liang et al. 1997a, b) and probably also in vivo (Liang and MacRae 1999). During a long-term study over several years, Artemia franciscana embryos showed no evidence of misfolded proteins or protein aggregation (Clegg 1997; Clegg et al. 1999). The amount of artemin and p26 was between 10 and 15% of the non-yolk protein in these embryos. However, the proteins have been found only in the early stages during development, not in the adult animals (Clegg et al. 1999). There is strong evidence that p26 in particular plays an important role as a molecular chaperone and is responsible for the high tolerance of embryos (Clegg 2007). Other studies indicate that artemin might even play a role as a molecular chaperone for RNA (Warner et al. 2004). Several cDNA libraries and thus expressed sequence tags (EST) of Milnesium tardigradum were created, and thus a whole range of different stress protein families were identified (Reuner et al. 2009). The in silico analysis of two existing sequences showed that Milnesium tardigradum possesses proteins that contain a small Hsp/α-crystallin domain. Based on their amino acid sequence, the molecular weights were determined and the proteins designated as MtHsp19.5 and MtHsp17.2. Small Hsps form large complexes of several hundred kilodaltons during heat stress, which are able to stabilize the structure of other proteins. Expression analyses in Milnesium tardigradum showed that Mthsp17.2 is significantly upregulated by heat stress. In contrast to p26 in Artemia, however, no increased expression of Mthsp19.5 and Mthp17.2 was detectable in anhydrobiotic tardigrades. A cDNA library of Milnesium tardigradum also contained the complete, coding sequence for Hsp10 (chaperonin). Hsp10, like Hsp60, is a chaperone found in the mitochondria and cytoplasm. Hsp60 shows chaperone activity, while Hsp10 serves as a functional regulator (Reuner et al. 2009).
10.3.2 Hsp60 and Hsp70
The first tardigrade partial heat shock protein gene sequences with a molecular weight of 70 kDa were found in the species Milnesium tardigradum (Schill et al. 2004). All sequences showed a different expression pattern but were inducible by heat shock. Additional Hsp70 isoforms and one Hsp90 sequence were found later in a cDNA library (Reuner et al. 2009). Only one of three Hsp70 isoforms was significantly expressed during dehydration and is still found in the anhydrobiotic state. Maybe the RNA will be translated only after rehydration, so that the chaperones can fold new proteins or renature others. Accumulation of RNAs associated with anhydrobiosis is known from prokaryotic and eukaryotic cells (Albertson et al. 1990). For example, a high amount of mRNAs was detected in the cryptobiotic stages of the ciliates Colpoda inflata (Benítez and Gutiérrez 1997) and Sterkiella histriomuscorum (Tourancheau et al. 1999). Likewise, various fungi that form surviving spores are known to store mRNAs (Camonis et al. 1982). Byers et al. (Byers et al. 1991; Martinez-Guitarte et al. 2007) and Gutierrez et al. (Martinez-Guitarte et al. 2007) could also detect Hsp70 mRNA in anhydrobiotic stages of Colpoda inflata and Colpoda nova. However, two yeast studies with Saccharomyces cerevisiae and Debaryomyces hansenii showed that Hsp70 does not protect the yeast cells from dehydration stress during desiccation (Guzhova et al. 2008). This may also be the case for tardigrades in anhydrobiosis.
Unlike Hsp70, the cytoplasmic Hsp90 is not generally involved in the folding of new proteins (Nathan et al. 1997). Hsp90 differs from Hsp70, that most of the known substrates are signal transduction proteins (e.g., steroid receptors and signal kinases) (Picard et al. 1990; Xu and Lindquist 1993). Therefore, it plays an important role in the network of cellular signaling pathways and is part of a broader chaperone mechanism (Bose et al. 1996; Freeman and Morimoto 1996; Schumacher et al. 1996). The complexity of Hsp90 also makes it much harder to investigate its capabilities compared to Hsp70. In tardigrades, it is only significantly expressed in anhydrobiosis. Which function it has shortly before complete dehydration, or whether it is translated only after renewed rehydration, requires further studies.
10.4 Metabolites for Cell Protection
Several mechanisms probably play a role in the protection of living cells in the anhydrobiotic state. These include the maintenance of the biologically relevant structures of macromolecules through the accumulation of disaccharides and other metabolites (Buitink and Leprince 2004; Crowe et al. 1987; Crowe 2002). Drying and freezing of cells usually lead to a massive damage of cell proteins and membranes, which normally ends in cell death. However, there are a number of organisms that have the extraordinary ability to survive complete dehydration like tardigrades (Crowe et al. 1992). The removal of intracellular water causes drastic changes in inter- and intramolecular interactions. During desiccation, intracellular proteins and membranes compensate for the loss of hydrogen bonds with water through hydrogen bonding with other molecules. This leads to forced interactions between molecules that would normally not react in the presence of water. Protein–protein interactions induced by water loss can result in irreversible conformational changes and result in the loss of enzyme activity (Carpenter et al. 1987; Hanafusa 1969; Prestrelski et al. 1993). In membranes, water loss can lead to a phase transition from the biologically active liquid crystalline to the gel phase (Crowe et al. 1992, 1997). In addition, water loss can result in the fusion of cell organelles. In cells that exhibit dehydration tolerance, molecular interactions during drying are controlled by replacing lost hydrogen bonds with reversible molecular interactions. This means that biomolecules and cellular structures can be protected from degradation by dehydration and rehydrate to their previous natural conformation. In dehydration-sensitive cells, on the other hand, the lack of such control means that the biomolecules and cellular structures within these cells cannot regain their original functional conformation after the stressor has disappeared.
Many anhydrobiotic organisms, like some nematodes and yeast cells, accumulate trehalose (Crowe et al. 1997; Womersley 1987), while sucrose is accumulated in the tissues of higher plants, especially in seeds and pollen (Hoekstra 1986; Koster and Leopold 1988). However, there are also exceptions such as the plant Myrothamnus flabellifolia which concurrently enriches sucrose and trehalose (Bianchi et al. 1993; Drennan et al. 1993). As a reason for the stabilizing effect of dissacharides, the “water replacement” (or “water substitute”) and “vitrification” (or “glassy state”) hypotheses are discussed (Crowe 1971; Webb et al. 1965). These are not alternative, but rather complementary explanations. Both are necessary but do not seem to be sufficient to fully protect the membranes during drying and rehydration.
10.4.1 Water Replacement Hypothesis
In the early 1960s, S. J. Webb postulated the water replacement hypothesis (Webb et al. 1965). It explains the stabilizing effect of free hydroxyl groups of sugar molecules that are available to proteins during and after removal of water to form hydrogen bonds. This lowers the free enthalpy of the system, which stabilizes native proteins. When unprotected membranes dehydrate, the water molecules that help maintain the distance between the phospholipid headgroups are removed. This allows the lipid molecules to come closer together and increase the membrane phase transition temperature (Tm), resulting in a phase change. In rehydration, a renewed phase change occurs in membranes, resulting in a brief release of soluble substances in cells through the membranes (Crowe et al. 1997). By contrast, when membranes are dried in the presence of trehalose, the water replacement hypothesis postulates that sugar molecules prevent the closest approach of lipids through interaction with phospholipid head groups during dehydration (Crowe et al. 1988). When water is returned to such a system, there is no phase change and the membranes retain their barrier function (Crowe et al. 1992, 1994; Harrigan et al. 1990).
10.4.1.1 Trehalose in Anhydrobionts
Trehalose was detected in anhydrobiotic states of the freshwater sponge Trochospongilla sp. (gemmules), the bryozoan Cristatella mucedo (statoblasts), and embryos of the crustaceans Daphnia magna, Daphnia pulex, Triops longicaudatus, and Triops cancriformis (Hengherr et al. 2011 ; Hengherr and Schill 2011). In particular, anhydrobiotic embryos of the brine shrimp Artemia franciscana have been the subject of intense research over the past three decades. They increase to 15–18% trehalose on a dry weight basis as they enter the anhydrobiotic stage (Clegg 1986; Clegg and Conte 1980; Clegg and Drost-Hansen 1990). The chironomid larva Polypedilum vanderplanki also stores around 18% trehalose (Watanabe et al. 2002) and nematodes, depending on the species, 4 to 15% (Madin and Crowe 1975). The first quantitative detection of trehalose in tardigrades was performed in the species Paramacrobiotus areolatus (Crowe 1975). Later, trehalose was also detected in Amphibolus nebulosus and Richtersius coronifer (Westh and Ramlov 1988). Quantitative studies in Richtersius coronifer showed that the highest amount of trehalose is found in the anhydrobiotic state but with a rather low content of up to 2.3% trehalose based on dry weight (Westh and Ramlov 1991). In a comparative quantitative amperometric chromatography study with several tardigrade species, it was shown that in species of the genera Paramacrobiotus and Macrobiotus, trehalose is accumulated during dehydration, whereas in Milnesium tardigradum trehalose could not be detected either in the active or in the anhydrobiotic or the respective transitional stages (Hengherr et al. 2008). The highest trehalose amount was measured with 0.472 ± 0.037% dry weight in a Paramacrobiotus species (Hengherr et al. 2008). This is much less than measured by Westh and Ramlov (1991), possibly due to different measuring methods. Although small amounts of trehalose could be detected in the heterotardigrades Echiniscus testudo and Echiniscus granulatus, accumulation of trehalose did not occur during dehydration (Hengherr et al. 2008). The rotifer species Philodina roseola and Adineta vaga, which also have the ability to completely dehydrate, are known to have no trehalose and no corresponding gene for trehalose-6-phosphate synthase (Lapinski and Tunnacliffe 2003). Anhydrobiosis without incorporation of trehalose for cell stabilization seems to be possible, at least for the tardigrade species Milnesium tardigradum, as well as these rotifer species. Nevertheless, sugars appear to play an important role in the tolerance to desiccation stress in many microorganisms, invertebrates, and plants and to impart stability to dried biomolecules and membranes in vitro (Tunnacliffe et al. 2001).
10.4.2 Vitrification Hypothesis
The second important protective function of dissacharides, as well as other carbohydrates and proteins, is vitrification at low water content (Crowe et al. 1998; Crowe 2002; Sun and Leopold 1997). The “glassy state” hypothesis postulates the formation of glassy states to immobilize proteins, which greatly reduces the reaction kinetics. The glass of carbohydrates is a solid-like amorphous material which prevents diffusion-limiting destruction processes, such as membrane fusions. In order to convert proteins into a “solid” state, the glass transition temperature of the amorphous system must be lowered. In the temperature range just above the glass transition temperature, but below the eutectic point of the constituents contained, the product is in a highly viscous state. In this range, a gradual transformation of an unstable amorphous liquid into a metastable amorphous solid state occurs. Not only sugars but also proteins may be involved in cell stabilization. In model systems, proteins have had a considerable effect on the molecular properties of carbohydrate glasses (Bell and Hageman 1996; Kalichevsky et al. 1992; Wolkers et al. 1998). For example, the addition of proteins to a glass caused a much higher glass transition temperature (T g) (Slade and Levine 1991) and a mean hydrogen bonding strength of the mixture compared to a pure sugar matrix (Wolkers et al. 1998). A mixture of protein and sugar is more dense than a pure carbohydrate or protein glass. In addition, proteins increase the temperature difference between the glass transition temperature and the critical temperature at which the dynamics of the glass transition from a solid-like to a liquid state. Proteins and carbohydrates probably interact through hydrogen bonds during the dry state in the cytoplasm of anhydrobiotes. By differential scanning calorimetry (DSC) measurements and Fourier transform infrared (FTIR) analyses, the “water replacement” and “vitrification” hypothesis could be examined for the first time in the drought-tolerant African chironomid larva Polypedilum vanderplanki (Sakurai et al. 2008). It was found that the anhydrobiotic larvae were in a glassy state; at a temperature above 65 ° C, the stabilizing glass melted. The previously mentioned high concentrations of trehalose have been detected throughout the organism, and the authors believe that trehalose plays an important role in water replacement and intracellular glass formation. In order to detect possible vitrification in tardigrades, DSC measurements were used in combination with experiments on heat tolerance in several species of the genera Paramacrobiotus, Macrobiotus, Echiniscus, and Milnesium (Fig. 10.3). In the species of the genera Paramacrobiotus and Macrobiotus, a glass transition and thus a vitrified state could be detected (Hengherr et al. 2009). When temperature tolerance of these species is considered, in parallel, it can be seen that when the glass transition temperature (T g) of about 80 °C is exceeded, the survival rates of the tardigrades drop sharply. This could indicate the loss of the protective function of the glassy state or the importance of vitrification in cellular protection. Macrobiotus hufelandi survived a short exposure to temperatures between 120 and 125 °C (Doyère 1842). Later, Baumann (1927) reported that anhydrobiotic tardigrades could tolerate 100 °C for 6 hours, and Richtersius coronifer survived temperatures up to approximately 70 °C for 60 minutes without any decrease in survival (Ramløv and Westh 2001). In the species Milnesium tardigradum, Echiniscus granulatus, and Echiniscus testudo, in which no or little trehalose was detected, no vitrification has been observed (Hengherr et al. 2009). It was found that exactly these species have a much better tolerance to high temperatures. At temperatures up to 90 °C, Milnesium tardigradum, Echiniscus granulatus, and Echiniscus testudo still showed survival rates of 90.0 ± 5.4% and 54.2 ± 13.7%, respectively, and 30.0% ± 11.2% after subsequent rehydration. The all-time record is held by Milnesium tardigradum at 100 °C with a survival rate of 91.7 ± 6.9%. Only higher temperatures led to a marked decline in survival rates, at 110 °C 1 ± 2% still survived (Hengherr et al. 2009). The temperature at which glass devitrifies (T g) depends on several factors (Slade and Levine 1991). Interestingly, recent evidence suggests that various stress or late embryogenesis abundant (LEA) proteins that accumulate during dehydration may also be involved in glass formation and improve overall stability (Wolkers et al. 1999, 2001). This could at least explain why tardigrades have a glass transition and thus a vitrified state, although no or only small amounts of trehalose could be detected.

As long as the heat exposure of a vitrified tardigrade is below the glass transition temperature (T g), the protective function of the glassy state is available and the tardigrade becomes alive after rehydration
10.5 Tardigrade-Specific Intrinsically Disordered Proteins (TDPs)
10.5.1 Late Embryogenesis Abundant (LEA) Proteins
In plants, and more recently in several animals, induction of high levels of hydrophilic proteins, in particular the late embryogenesis abundant (LEA) proteins, has been associated with water stress. LEA proteins were first identified 30 years ago in plants, where they are produced during seed development (Galau et al. 1986; Grzelezak et al. 1982). However, their precise function is poorly understood. To be classified as molecular chaperones, LEA proteins must not only prevent aggregation, they must additionally form transient, non-covalent complexes (Ellis 2004). Unfavorable protein–protein interactions, however, can lead to irreversible conformational changes and, in enzymes, a loss of catalytic activity (Crowe et al. 1987). LEA proteins might simply function as “molecular shields,” forming a physical barrier between partially unfolded neighboring proteins and preventing contact between them (Tunnacliffe and Wise 2007). LEA protein expression has more recently been demonstrated in other organisms and is also linked to desiccation stress and the acquisition of desiccation tolerance. So far these organisms include microorganisms (Battista et al. 2001), nematodes (Browne et al. 2002, 2004; Goyal et al. 2003, 2005), rotifers (Tunnacliffe et al. 2005), chironomid larvae (Kikawada et al. 2006), and Collembola (Bahrndorff et al. 2008). Research on LEA proteins has recently focused on tardigrades because of the evidence that these proteins play an important role in protecting cellular proteins. For the first time, putative LEA proteins have been detected in the anhydrobiotic state of the species Macrobiotus hufelandi (McGee et al. 2004). With high-throughput, high-accuracy proteomics in combination with a newly developed tardigrade-specific protein database (Schokraie et al. 2012), more than 3000 proteins were identified in Milnesium tardigradum. This comprehensive proteome resource includes protein families such as chaperones, antioxidants, ribosomal proteins, cytoskeletal proteins, transporters, protein channels, nutrient reservoirs, and developmental proteins. Within these proteins, also those belonging to the LEA family (group 3) were identified.
10.5.2 CAHS, SAHS, and MAHS Proteins
Tanaka et al. (2015) found in Ramazzottius varieornatus two novel mitochondrial heat-soluble proteins, RvLEAM (a group 3 LEA) and MAHS (mitochondrial abundant heat soluble), as potent mitochondrial protectants. The MAHS protein is also a heat-soluble protein that might have protective roles in tardigrades. In the tardigrade species Ramazzottius varieornatus, a further five abundant heat-soluble proteins have been found, but all of them showed no sequence similarity with LEA proteins. They formed two novel protein families, the cytoplasmic abundant heat soluble (CAHS) and secretory abundant heat soluble (SAHS) protein families, according to their localization (Yamaguchi et al. 2012). Both protein families were also found in Hypsibius dujardini and Milnesium tardigradum, but not in other phyla. Therefore LEA, CAHS, SAHS, and MAHS families have become referred to as the tardigrade-specific intrinsically disordered proteins (TDPs). Boothby et al. (2017) showed that the TDP genes are constitutively expressed at high levels or induced during desiccation in the tardigrade species Hypsibius dujardini and Paramacrobiotus richtersi. All TDPs form noncrystalline amorphous solids upon desiccation, and this vitrified state mirrors their protective capabilities.
10.6 DNA Damage During Anhydrobiosis
One reason for the decline in survival with increasing time in anhydrobiosis could be oxidative damage caused by reactive oxygen species (ROS) (Womersley 1987). Since enzymes do not work without water and therefore no metabolism can be detected, even energy-dependent repair systems cannot be activated. This causes an accumulation of DNA damage until it reaches a lethal limit, eventually leading to the death of the whole organism (Lindahl 1993). Deoxyribonucleic acids are a preferred biological target of ROS (Gros et al. 2002). For example, they may spontaneously result from the chemical degradation of various substances in the metabolism and have a deleterious effect on proteins, membranes, and DNA (Mattimore and Battista 1996). The phenomenon of induced double-strand breaks by dehydration is already known in bacteria (Billi 2009). Blasius et al. (2008), as well as Mattimore and Battista (1996), believe in effective repair of DNA as one of the most important survival mechanisms (Blasius et al. 2008; Mattimore and Battista 1996). To visualize and detect DNA damage, e.g., single- and double-strand breaks, incomplete excision repair, alkali-labile sites, and cross-linking, storage cells of anhydrobiotic tardigrades of the species Milnesium tardigradum were examined with single-cell gel electrophoresis (comet assay) (Neumann et al. 2009). The animals which spent 2 days in anhydrobiosis showed little DNA damage (2.09 ± 1.98% DNA) in the comet tail, compared with the DNA damage in the comet tail of active animals (0.44 ± 0.74%). With increasing time in anhydrobiosis, DNA damage increased. After 6 weeks of anhydrobiosis, 13.63 ± 6.41% DNA was found in the comet tail and 23.66 ± 7.56% DNA after 10 months (Neumann et al. 2009). The amount of DNA damage correlated with the duration of anhydrobiosis, since storage cells from animals that spend only 2 days in anhydrobiosis have relatively little damage and cells that are longer in anhydrobiosis show significantly greater damage. However, it is assumed that the DNA damage does not occur during the dehydration process but rather during anhydrobiosis. They can accumulate DNA damage over time. Of course, a repair process can only become activated after rehydration. The exact causes of the damage are not yet sufficiently known, but a direct or indirect involvement of reactive oxygen species and/or free radicals is likely.
In the last decade, many data from the tardigrade species Echiniscus testudo, Milnesium tardigradum, Hypsibius dujardini, Paramacrobiotus richtersi, and Ramazzottius varieornatus have been generated with state-of-the-art methods in genomics, transcriptomics, proteomics, and bioinformatics (e.g., Arakawa 2018; Arakawa et al. 2016; Beisser et al. 2012; Bemm et al. 2016; Boothby et al. 2015, 2017; Borner et al. 2014; Förster et al. 2009, 2011a, b; Kondo et al. 2015; Mali et al. 2008; Schokraie et al. 2010, 2012; Tanaka et al. 2015; Yamaguchi et al. 2012; Yoshida et al. 2017). This is an excellent basis for further studies to understand the mechanisms of desiccation tolerance in tardigrades.
References
Albertson NH, Nyström T, Kjelleberg S (1990) Functional mRNA half-lives in the marine Vibrio sp. S14 during starvation and recovery. J Gen Microbiol 136:2195–2199
Alpert P (2000) The discovery, scope, and puzzle of desiccation tolerance in plants. Plant Ecol 151:5–17
Altiero T, Rebecchi L, Bertolani R (2006) Phenotypic variations in the life history of two clones of Macrobiotus richtersi (Eutardigrada, Macrobiotidae). Hydrobiologia 558:33–40
Arakawa K (2018) The complete mitochondrial genome of Echiniscus testudo (Heterotardigrada: Echiniscidae). Mitochondrial DNA Part B 3:810–811
Arakawa K, Yoshida Y, Tomita M (2016) Genome sequencing of a single tardigrade Hypsibius dujardini individual. Sci Data 3:16006
Arrigo A-P, Müller WEG (2002) Small stress proteins. Springer, Berlin, I–XV, 1–270 pp
Bahrndorff S, Tunnacliffe A, Wise MJ, McGee B, Holmstrup M, Loeschcke V (2008) Bioinformatics and protein expression analyses implicate LEA proteins in the drought response of Collembola. J Insect Physiol 55:210–217
Battista JR, Park MJ, McLemore AE (2001) Inactivation of two homologues of proteins presumed to be involved in the desiccation tolerance of plants sensitizes Deinococcus radiodurans R1 to desiccation. Cryobiology 43:133–139
Baumann H (1927) Anabiosis of tardigrades. Zool Anz 72:175–179
Beisser D, Grohme M, Kopka J, Frohme M, Schill RO, Hengherr S, Dandekar T, Klau GW, Dittrich M, Müller T (2012) Integrated pathway modules using time-course metabolic profiles and EST data from Milnesium tardigradum. BMC Syst Biol 6:72
Bell LN, Hageman MJ (1996) Glass transition explanation for the effect of polyhydroxy compounds on protein denaturation in dehydrated solids. J Food Sci Technol 61:372–378
Bemm F, Weiss CL, Schultz J, Förster F (2016) Genome of a tardigrade: horizontal gene transfer or bacterial contamination? Proc Natl Acad Sci U S A 113:E3054–E3056
Benítez L, Gutiérrez JC (1997) Encystment – specific mRNA is accumulated in the resting cysts of the ciliate Colpoda inflata. Biochem Mol Biol Int 41:1137–1141
Bertolani R, Guidetti R, Jönsson KI, Altiero T, Boschini D, Rebecchi L (2004) Experiences with dormancy in tardigrades. J Limnol 63:16–25
Bianchi G, Gamba A, Limiroli R, Pzzi N, Ester R, Salamini F, Bartels D (1993) The unusual sugar composition in leaves of the resurrection plant Myrothamnus flabellifolia. Physiol Plant 87:223–226
Billi D (2009) Subcellular integrities in Chroococcidiopsis sp. CCMEE 029 survivors after prolonged desiccation revealed by molecular probes and genome stability assays. Extremophiles 13:49–57
Blasius M, Hubscher U, Sommer S (2008) Deinococcus radiodurans: what belongs to the survival kit? Crit Rev Biochem Mol Biol 43:221–238
Boothby TC, Tapia H, Brozena AH, Piszkiewicz S, Smith AE, Giovannini I, Rebecchi L, Pielak GJ, Koshland D, Goldstein B (2017) Tardigrades use intrinsically disordered proteins to survive desiccation. Mol Cell 65(975–984):e975
Boothby TC, Tenlen JR, Smith FW, Wang JR, Patanella KA, Nishimura EO, Tintori SC, Li Q, Jones CD, Yandell M, Messina DN, Glasscock J, Goldstein B (2015) Evidence for extensive horizontal gene transfer from the draft genome of a tardigrade. Proc Natl Acad Sci U S A 112:15976–15981
Borner J, Rehm P, Schill RO, Ebersberger I, Burmester T (2014) A transcriptome approach to ecdysozoan phylogeny. Mol Phylogenet Evol 80:79–87
Bose S, Weikl T, Bugl H, Buchner J (1996) Chaperone function of Hsp90-associated proteins. Science 274:1715–1717
Browne J, Tunnacliffe A, Burnell A (2002) Anhydrobiosis – plant desiccation gene found in a nematode. Nature 416:38
Browne JA, Dolan KM, Tyson T, Goyal K, Tunnacliffe A, Burnell AM (2004) Dehydration-specific induction of hydrophilic protein genes in the anhydrobiotic nematode Aphelenchus avenae. Eukaryot Cell 3:966–975
Buitink J, Leprince O (2004) Glass formation in plant anhydrobiotes: survival in the dry state. Cryobiology 48:215–228
Byers TJ, Kim BG, King LE, Hugo ER (1991) Molecular aspects of the cell cycle and encystment of Acanthamoeba. Rev Infect Dis 5:373–384
Camonis J, Julien J, Ayala J, Jaequet M (1982) Polyadenylated RNA population present in dormant spores of Dictyostelium discoideum. Cell Differ (1):55–61
Carpenter JF, Crowe LM, Crowe JH (1987) Stabilization of phosphofructokinase with sugars during freeze-drying characterization of enhanced protection in the presence of divalent cations. Biochim Biophys Acta 923:109–115
Chandler J, Bartels D (1999) Plant desiccation. In: Lerner HR (ed) Plant responses to environmental stresses: from phytohormones to genome reorganization. Marcel Dekker, New York, pp 575–590
Chen T, Amons R, Clegg JS, Warner AH, MacRae TH (2003) Molecular characterization of artemin and ferritin from Artemia franciscana. Eur J Biochem 270:137–145
Chen T, Villeneuve TS, Garant KA, Amons R, MacRae TH (2007) Functional characterization of artemin, a ferritin homolog synthesized in Artemia embryos during encystment and diapause. FEBS J 274:1093–1101
Clegg JS (1967) Metabolic studies of crytobiosis in encysted embryos of Artemia salina. Comp Biochem Physiol 20:801–809
Clegg JS (1986) The physical properties and metabolic status of Artemia cysts at low water contents: the water replacement hypothesis. In: Leopold AC (ed) Membranes, metabolism and dry organisms. Cornell University Press, New York, pp 169–187
Clegg JS (1997) Embryos of Artemia franciscana survive four years of continuous anoxia: the case for complete metabolic rate depression. J Exp Biol 200:467–475
Clegg JS (2007) Protein stability in Artemia embryos during prolonged anoxia. Biol Bull 212:74–81
Clegg JS, Conte F (1980) A review of the cellular and developmental biology of Artemia. In: Persoone GP, Sorgeloos P, Roels O, Jaspers E (eds) The brine shrimp, Artemia. Universa Press, Wetteren, pp 11–54
Clegg JS, Drost-Hansen W (1990) On the biochemistry and cell physiology of water. In: Hochachka PW, Mommsen TP (eds) Biochemistry and molecular biology of fishes. Elsevier, Amsterdam, pp 1–23
Clegg JS, Jackson SA, Liang P, MacRae TH (1995) Nuclear-cytoplasmic translocations of protein p26 during aerobic-anoxic transitions in embryos of Artemia franciscana. Exp Cell Res 219:1–7
Clegg JS, Jackson SA, Warner AH (1994) Extensive intracellular translocations of a major protein accompany anoxia in embryos of Artemia franciscana. Exp Cell Res 212:77–83
Clegg JS, Willsie JK, Jackson SA (1999) Adaptive significance of a small heat shock/alpha-crystallin protein (p26) in encysted embryos of the brine shrimp, Artemia franciscana. Am Zool 39:836–847
Crowe JH (1971) Anhydrobiosis: an unsolved problem. Am Nat 105:563–573
Crowe JH (1975) The physiology of cryptobiosis in tardigrades. Memorie dell’Istituto Italiano di Idrobiologica 32(Suppl):37–59
Crowe LM (2002) Lessons from nature: the role of sugars in anhydrobiosis. Comp Biochem Physiol A Mol Integr Physiol 131:505–513
Crowe JH, Clegg JS (1973) Anhydrobiosis. Dowden, Hutchinson and Ross, Stroudsburg, p 477
Crowe JH, Clegg JS (1978) Dry biological systems. Academic Press, New York
Crowe JH, Madin KAC (1975) Anhydrobiosis in nematodes: evaporative water loss and survival. J Exp Zool 193:323–333
Crowe JH, Crowe LM, Carpenter JF, Wistrom CA (1987) Stabilization of dry phospholipid bilayers and proteins by sugars. Biochem J 242:1–10
Crowe JH, Crowe LM, Carpenter JF, Rudolph AS, Wistrom CA, Spargo BJ, Anchordoguy TJ (1988) Interactions of sugars with membranes. Biochim Biophys Acta 947:367–384
Crowe JH, Hoekstra FA, Crowe LM (1992) Anhydrobiosis. Annu Rev Physiol 54:579–599
Crowe JH, Leslie SB, Crowe LM (1994) Is vitrification sufficient to preserve liposomes during freeze-drying? Cryobiology 31:355–366
Crowe JH, Crowe LM, Petrelski S, Hoekstra FA, Araujo PD, Panek AD (1997) Anhydrobiosis: cellular adaptation to extreme dehydration. In: Dantzler WH (ed) Handbook of physiology. Oxford University Press, New York
Crowe JH, Carpenter JF, Crowe LM (1998) The role of vitrification in anhydrobiosis. Annu Rev Physiol 60:73–103
De Graaf J, Amons R, Möller W (1990) The primary structure of artemin from artemia cysts. Eur J Biochem 193:737–750
Doyère PLN (1842) Memoires sur les Tardigrades. Sur le facilité que possedent les Tardigrades, les rotiferes, les anguillules des toits et quelques autres animalcules, de revenir à la vie après été completement desséchées. Ann Sci Nat Zool Biol Anim 2e:5–35
Drennan P, Smith M, Goldsworthly D, van Staden J (1993) The occurence of trehalose in the leaves of the desiccation-tolerant angiosperm Myrothamnus flabellifolius. J Plant Physiol 142:493–496
Ellis RJ (2004) From chloroplasts to chaperones: how one thing led to another. Photosynth Res 80:333–343
Feder ME, Hofmann GE (1999) Heat-shock proteins, molecular chaperones, and the stress response: evolutionary and ecological physiology. Annu Rev Physiol 61:243–282
Fielding MJ (1951) Observations on the length of dormancy in certain plant infecting nematodes. Proc Helminthol Soc Wash 18:110–112
Förster F, Beisser D, Frohme M, Schill RO, Dandekar T (2011a) Tardigrade bioinformatics: molecular adaptations, DNA j-family and dynamical modeling. J Zool Syst Evol Res 49:120–126
Förster F, Beisser D, Grohme M, Liang C, Mali B, Siegl AM, Engelmann JC, Shkumatov AV, Schokraie E, Müller T, Schnölzer M, Schill RO, Frohme M, Dandekar T (2011b) Transcriptome analysis in tardigrade species reveals specific molecular pathways for stress adaptations. Bioinf Biol Insights 6:69–96
Förster F, Liang C, Shkumatov A, Beisser D, Engelmann JC, Schnölzer M, Frohme M, Müller T, Schill RO, Dandekar T (2009) Tardigrade workbench: comparing stress-related proteins, sequence-similar and functional protein clusters as well as RNA elements in tardigrades. BMC Genomics 10:469–479
Franceschi T (1948) Anabiosi nei tardigradi. Boll Mus Ist Biot Univ Genova:47–49
Freeman BC, Morimoto RI (1996) The human cytosolic molecular chaperones hsp90, hsp70 (hsc70) and hdj-1 have distinct roles in recognition of a non-native protein and protein refolding. EMBO J 15:2969–2979
Galau GA, Hughes DW, Dure L III (1986) Abscisic-acid induction of cloned cotton gossypium-hirsutum late embryogenesis-abundant lea messenger rna species. Plant Mol Biol 7:155–170
Gething M-J, Sambrook J (1992) Protein folding in the cell. Nature 355:33–45
Goodey T (1923) Quiescence and reviviscence in nematodes, with special reference to Tylenchus tritici and Tylenchus dipsaci. J Helminthol 1:47–52
Goyal K, Pinelli C, Maslen SL, Rastogi RK, Stephens E, Tunnacliffe A (2005) Dehydration-regulated processing of late embryogenesis abundant protein in a desiccation-tolerant nematode. FEBS Lett 579:4093–4098
Goyal K, Tisi L, Basran A, Browne J, Burnell A, Zurdo J, Tunnacliffe A (2003) Transition from natively unfolded to folded state induced by desiccation in an anhydrobiotic nematode protein. J Biol Chem 278:12977–12984
Gros L, Saparbaev MK, Laval J (2002) Enzymology of the repair of free radicals-induced DNA damage. Oncogene 21:8905–8925
Grzelezak ZF, Sattalo MH, Hanley-Bowdoin LK, Kennedy TD, Lane BG (1982) Synthesis and turnover of proteins and mRNA in germinating wheat embryos. Can J Biochem Physiol:389–397
Guidetti R, Jönsson KI (2002) Long-term anhydrobiotic survival in semi-terrestrial micrometazonas. J Zool (Lond) 257:181–187
Guzhova I, Krallish I, Khroustalyova G, Margulis B, Rapoport A (2008) Dehydration of yeast: changes in the intracellular content of Hsp70 family proteins. Process Biochem 43:1138–1141
Hanafusa H (1969) Rapid transformation of cells by rous sarcoma virus. Proc Natl Acad Sci 63:318–325
Harrigan PR, Madden TD, Cullis PR (1990) Protection of liposomes during dehydration or freezing. Chem Phys Lipids 52:139–149
Hengherr S, Heyer AG, Brümmer F, Schill RO (2008) Trehalose as protecting agent in aquatic invertebrates during dormancy induced by desiccation. Comp Biochem Physiol A Comp Physiol 151:S34
Hengherr S, Heyer AG, Brümmer F, Schill RO (2011) Trehalose and vitreous states: desiccation tolerance in dormant stages of the crustaceans Triops and Daphnia. Physiol Biochem Zool 84:147–153
Hengherr S, Schill RO (2011) Dormant stages in freshwater bryozoans – an adaptation to transcend environmental constraints. J Insect Physiol 57:595–601
Hengherr S, Worland MR, Reuner A, Brümmer F, Schill RO (2009) High-temperature tolerance in anhydrobiotic tardigrades is limited by glass transition. Physiol Biochem Zool 82:749–755
Hoekstra FA (1986) Water content in relation to stress in pollen. In: Leopold CA (ed) Membranes, metabolism and dry organisms. Cornell University Press, New York
Ingram J, Bartels D (1996) The molecular basis of dehydration tolerance in plants. Annu Rev Plant Physiol Plant Mol Biol 47:377–403
Jönsson KI, Bertolani R (2001) Facts and fiction about long-term survival in tardigrades. J Zool (Lond) 255:121–123
Jørgensen A, Møbjerg N, Kristensen RM (2007) A molecular study of the tardigrade Echiniscus testudo (Echiniscidae) reveals low DNA sequence diversity over a large geographical area. J Limnol 66:77–83
Kalichevsky MT, Jaroszkiewicz EM, Ablett S, Blanshard JMV, Lillford PJ (1992) The glass transition of amylopectin measured by DSC, DMTA and NMR. Carbohydr Polym 18:77–88
Keilin D (1959) The leeuwenhoek lecture. The problem of anabiosis or latent life: history and current concept. Proc R Soc Lond B Biol Sci 150:149–191
Kikawada T, Nakahara Y, Kanamori Y, Iwata K-i, Watanabe M, McGee B, Tunnacliffe A, Okuda T (2006) Dehydration-induced expression of LEA proteins in an anhydrobiotic chironomid. Biochem Biophys Res Commun 348:56–61
Kondo K, Kubo T, Kunieda T (2015) Suggested involvement of PP1/PP2A activity and de novo gene expression in anhydrobiotic survival in a tardigrade, Hypsibius dujardini, by chemical genetic approach. PLoS One 10:e0144803
Koster KL, Leopold AC (1988) Sugars and desiccation tolerance in seeds. Plant Physiol (Rockv) 88:829–832
Lapinski J, Tunnacliffe A (2003) Anhydrobiosis without trehalose in bdelloid rotifers. FEBS Lett 553:387–390
Lee DL (1961) Two new species of cryptobiotic (anabiotic) freshwater nematodes, Actinolaimus hintoni sp. nov. and Dorylaimus keilini sp. nov. (Dorylaimidae). Parasitology 51:237–240
Liang P, Amons R, Clegg JS, MacRae TH (1997a) Molecular characterization of a small heat shock/alpha-crystallin protein in encysted Artemia embryos. J Biol Chem 272:19051–19058
Liang P, Amons R, MacRae TH, Clegg JS (1997b) Purification, structure and in vitro molecular-chaperone activity of Artemia p26, a small heatshock a-crystallin protein. Eur J Biochem 243:225–232
Liang P, MacRae TH (1999) The synthesis of a small heat shock/alpha-crystallin protein in Artemia and its relationship to stress tolerance during development. Dev Biol 207:445–456
Lindahl T (1993) Instability and decay of the primary structure of DNA. Nature 362:709–715
Madin KAC, Crowe JH (1975) Anhydrobiosis in nematodes: carbohydrate and lipid metabolism during dehydration. J Exp Zool 193:335–342
Mali B, Grohme M, Wełnicz W, Dandekar T, Schnölzer M, Reuter D, Schill RO, Frohme M (2008) Genomic analyses of cryptobiotic tardigrades. In: Wissenschaftliche Beiträge 2008. Wildau
Martinez-Guitarte JL, Planello R, Morcillo G (2007) Characterization and expression during development and under environmental stress of the genes encoding ribosomal proteins L11 and L13 in Chironomus riparius. Comp Biochem Physiol B Biochem Mol Biol 147:590–596
Mattimore V, Battista JR (1996) Radioresistance of Deinococcus radiodurans: functions necessary to survive ionizing radiation are also necessary to survive prolonged desiccation. J Bacteriol 178:633–637
McGee B, Schill RO, Tunnacliffe A (2004) Hydrophilic proteins in invertebrate anhydrobiosis. Integr Comp Biol 44:679–679
Morimoto RI (1993) Cells in stress: transcriptional activation of heat shock genes. Science 259:1409–1410
Nathan DF, Vos MH, Lindquist S (1997) In vivo functions of the Saccharomyces cerevisiae Hsp90 chaperone. Proc Natl Acad Sci U S A 94:12949–12956
Neumann S, Reuner A, Brümmer F, Schill RO (2009) DNA damage in storage cells of anhydrobiotic tardigrades. Comp Biochem Physiol A Mol Integr Physiol 153:425–429
Örstan A (1995) Desiccation survival of the eggs of the rotifer Adineta vaga (Davis, 1873). Hydrobiologia 313/314:373–375
Örstan A (1998) Factors affecting long-term survival of dry bdelloid rotifers: a preliminary study. Hydrobiologia 387/388:327–331
Parsell DA, Lindquist S (1993) The function of heat-shock proteins in stress tolerance: degradation and reactivation of damaged proteins. Annu Rev Genet 27:437–496
Picard D, Khursheed B, Garabedian MJ, Fortin MG, Lindquist S, Yamamoto KR (1990) Reduced levels of hsp90 compromise steroid receptor action in vivo. Nature 348:166–168
Potts M (2001) Desiccation tolerance: a simple process? Trends Microbiol 9:553–559
Prestrelski SJ, Tedeschi N, Arakawa T, Carpenter JF (1993) Dehydration-induced conformational transitions in proteins and their inhibition by stabilizers. Biophys J 65:661–671
Priestley DA (1986) Seed aging. Cornell University Press, New York
Rahm PG (1923) Biologische und physiologische Beiträge zur Kenntnis der Moosfauna. Zeitschrift für allgemeine Physiologie 20:1–34
Ramazzotti G, Maucci W (1983) Il phylum Tardigrada. Memorie dell’Istituto Italiano di ldrobiologia. Istituto Italiano di idrobiologia, Verbana Pallanza, Italien, 1014 pp
Ramløv H, Westh P (2001) Cryptobiosis in the eutardigrade Adorybiotus (Richtersius) coronifer: tolerance to alcohols, temperature and de novo protein synthesis. Zool Anz 240:517–523
Rebecchi L, Guidetti R, Borsari S, Altiero T, Bertolani R (2006) Dynamics of long-term anhydrobiotic survival of lichen-dwelling tardigrades. Hydrobiologia 558:23–30
Reuner A, Hengherr S, Mali B, Förster F, Arndt D, Reinhardt R, Dandekar T, Frohme M, Brümmer F, Schill RO (2009) Stress-response in tardigrades: differential gene expression of molecular chaperones. Cell Stress Chaperones 15:423–430
Ricci C (1998) Anhydrobiotic capabilities of bdelloid rotifers. Hydrobiologia 387/388:321–326
Ricci C (2001) Dormancy patterns in rotifers. Hydrobiologia 446/447:1–11
Ricci C, Pagani M (1997) Desiccation of Panagrolaimus rigidus (nematoda): survival, reproduction and the influence on the internal clock. Hydrobiologia 347:1–13
Ricci C, Caprioli M (1998) Stress during dormancy: effect on recovery rates and life-history traits of anhydrobiotic animals. Aquat Ecol 32:353–359
Ricci C, Caprioli M (2005) Anhydrobiosis in bdelloid species, populations and individuals. Integr Comp Biol 45:759–763
Ricci C, Covino C (2005) Anhydrobiosis of Adineta ricciae: costs and benefits. Hydrobiologia 546:307–314
Ricci C, Vaghi L, Manzini ML (1987) Desiccation of rotifers (Macrotrachela quadricornifera): survival and reproduction. Ecology 68:1488–1494
Sakurai M, Furuki T, Akao K-i, Tanaka D, Nakahara Y, Kikawada T, Watanabe M, Okuda T (2008) Vitrification is essential for anhydrobiosis in an african chironomid, Polypedilum vanderplanki. Proc Natl Acad Sci 105:5093–5098
Schill RO, Fritz GB (2008) Desiccation tolerance in embryonic stages of the tardigrade. J Zool (Lond) 276:103–107
Schill RO, Steinbrück GHB, Köhler H-R (2004) Stress gene (hsp70) sequences and quantitative expression in Milnesium tardigradum (Tardigrada) during active and cryptobiotic stages. J Exp Biol 207:1607–1613
Schokraie E, Hotz-Wagenblatt A, Warnken U, Mali B, Frohme M, Forster F, Dandekar T, Hengherr S, Schill RO, Schnölzer M (2010) Proteomic analysis of tardigrades: towards a better understanding of molecular mechanisms by anhydrobiotic organisms. PLoS One 5:e9502.9501–e9502.9537
Schokraie E, Warnken U, Hotz-Wagenblatt A, Grohme M, Hengherr S, Förster F, Schill RO, Frohme M, Dandekar T, Schnölzer M (2012) Comparative proteome analysis of Milnesium tardigradum in early embryonic state versus adults in active and anhydrobiotic state. PLoS One 7:e45682
Schöneich C (1999) Reactive oxygen species and biological aging: amechanistical approach. Exp Gerontol 34:19–34
Schumacher RJ, Hansen WJ, Freeman BC, Alnemri E, Litwack G, Toft DO (1996) Cooperative action of Hsp70, Hsp90, and DnaJ proteins in protein renaturation. Biochemistry 35:14889–14898
Scott P (2000) Resurrection plants and the secrets of eternal leaf. Ann Bot (Lond) 85:159–166
Slade L, Levine H (1991) Beyond water activity: recent advances based on an alternative approach to the assessment of food quality and safety. Crit Rev Food Sci Nutr 30:115–360
Steiner G, Albin FE (1946) Resuscitation of the nematode Tylenchus polyhypnus n. sp. after almost 39 years’ dormancy. J Wash Acad Sci 36:97–99
Sun WQ, Leopold AC (1997) Cytoplasmic vitrification and survival of anhydrobiotic organisms. Comp Biochem Physiol A Physiol 117:327–333
Tanaka S, Tanaka J, Miwa Y, Horikawa DD, Katayama T, Arakawa K, Toyoda A, Kubo T, Kunieda T (2015) Novel mitochondria-targeted heat-soluble proteins identified in the anhydrobiotic Tardigrade improve osmotic tolerance of human cells. PLoS One 10:e0118272
Tissières A, Mitchell HK, Tracy UM (1974) Protein synthesis in the salivary glands of Drosophila melanogaster. J Mol Biol 84:389–398
Tomos D (1992) Life without water. Curr Biol 2:594–596
Tourancheau AB, Morin L, Yang T, Perasso R (1999) Messenger RNA in dormant cells of sterkiella histriomuscorum (oxytrichidae): identification of putative regulatory gene transcripts. Protist 150:137–147
Tunnacliffe A, de Castro AG, Manzanera M (2001) Anhydrobiotic engineering of bacterial and mammalian cells: is intracellular trehalose sufficient? Cryobiology 43:124–132
Tunnacliffe A, Lapinski J (2003) Resurrecting van Leeuwenhoek’s rotifers: a reappraisal of the role of disaccharides in anhydrobiosis. Philos Trans R Soc Lond Ser B Biol Sci 358:1755–1771
Tunnacliffe A, Lapinski J, McGee B (2005) A putative LEA protein, but no trehalose, is present in anhydrobiotic bdelloid rotifers. Hydrobiologia 546:315–321
Tunnacliffe A, Wise MJ (2007) The continuing conundrum of the LEA proteins. Naturwissenschaften 94:791–812
van Leeuwenhoek A (1702) On certain animalcules found in the sediments in gutters of the roofs of houses. Letter 144. In: The selected works of Anton van Leeuwenhoek. London, pp 207–213
Vertucci CW, Farrant JM (1995) Acquisition and loss of desiccation. Marcel Dekker, New York
Warner AH, Brunet RT, MacRae TH, Clegg JS (2004) Artemin is an RNA-binding protein with high thermal stability and potential RNA chaperone activity. Arch Biochem Biophys 424:189–200
Watanabe M, Kikawada T, Minagawa N, Yukuhiro F, Okuda T (2002) Mechanism allowing an insect to survive complete dehydration and extreme temperatures. J Exp Biol 205:2799–2802
Webb SJ, Dumasia MD, Bhorjee JS (1965) Bound water, inositol, and the biosynthesis of temperate and virulent bacteriophages by air-dried Escherichia coli. Can J Microbiol 11:141–150
Westh P, Ramlov H (1988) Cryptobiosis in arctic tardigrades with special attention to the appearance of trehalose. In: Petersen GH (eds) Bericht über die Grönland-Exkursion des Instituts für Polarökologie. Universität für Polarökologie, Kiel, pp 227–245
Westh P, Ramlov H (1991) Trehalose accumulation in the tardigrade Adorybiotus coronifer during anhydrobiosis. J Exp Zool 258:303–311
Wharton DA (2003) The environmental physiology of antarctic terrestrial nematodes: a review. J Comp Physiol B Biochem Syst Environ Physiol 173:621–628
Wharton DA, Aalders O (1999) Desiccation stress and recovery in the anhydrobiotic nematode Ditylenchus dipsaci (Nematoda : Anguinidae). Eur J Entomol 96:199–203
Willsie JK, Clegg JS (2002) Small heat shock protein p26 associates with nuclear lamins and HSP70 in nuclei and nuclear matrix fractions from stressed cells. J Cell Biochem 84:601–614
Wolkers WF, McCready S, Brandt WF, Lindsey GG, Hoekstra FA (2001) Isolation and characterization of a D-7 LEA protein from pollen that stabilizes glasses in vitro. Biochim Biophys Acta Protein Struct Mol Enzymol 1544:196–206
Wolkers WF, Oldenhof H, Alberda M, Hoekstra FA (1998) A Fourier transform infrared microspectroscopy study of sugar glasses: application to anhydrobiotic higher plant cells. Biochim Biophys Acta 1379:83–96
Wolkers WF, Tetteroo FA, Alberda M, Hoekstra FA (1999) Changed properties of the cytoplasmic matrix associated with desiccation tolerance of dried carrot somatic embryos. An in situ fourier transform infrared spectroscopic study. Plant Physiol 120:153–164
Womersley CZ (1987) A re-evaluation of strategies employed by nematode anhydrobiotes in relation to their natural environment. In: Veech JA, Dickson DW (eds) Vistas on nematology. Society of Nematologists, Hyattsville, pp 165–173
Wright JC (1991) The significance of four xeric parameters in the ecology of terrestrial tardigrada. J Zool (Lond) 224:59–77
Wright JC (2001) Cryptobiosis 300 years on from van Leuwenhoek: what have we learned about tardigrades? Zool Anz 240:563–582
Xu Y, Lindquist S (1993) Heat-shock protein hsp90 governs the activity of pp60v-src kinase. Proc Natl Acad Sci U S A 90:7074–7078
Yamaguchi A, Tanaka S, Yamaguchi S, Kuwahara H, Takamura C (2012) Two novel heat-soluble protein families abundantly expressed in an anhydrobiotic tardigrade. PLoS One 7:e44209
Yoshida Y, Koutsovoulos G, Laetsch DR, Stevens L, Kumar S, Horikawa DD, Ishino K, Komine S, Kunieda T, Tomita M, Blaxter M, Arakawa K (2017) Comparative genomics of the tardigrades Hypsibius dujardini and Ramazzottius varieornatus. PLoS Biol 15:e2002266
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2018 Springer Nature Switzerland AG
About this chapter

Cite this chapter
Schill, R.O., Hengherr, S. (2018). Environmental Adaptations: Desiccation Tolerance. In: Schill, R. (eds) Water Bears: The Biology of Tardigrades. Zoological Monographs, vol 2. Springer, Cham. https://doi.org/10.1007/978-3-319-95702-9_10
Download citation
DOI: https://doi.org/10.1007/978-3-319-95702-9_10
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-95701-2
Online ISBN: 978-3-319-95702-9
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)