
Abstract
Numerous non-indigenous species have established populations in New Zealand lake ecosystems, including some relatively recently, and many others are likely unrecognised. Control or eradication as part of lake restoration programmes is more achievable for vertebrate than invertebrate species once established. Focus for management of smaller species must, therefore, be placed on preventing initial establishment rather than post-establishment control. Eradication of the Australian marron (Cherax tenuimanus) from New Zealand is a rare example of eradication success for an invertebrate. Even for vertebrates, while control is possible with ongoing effort, complete eradication is typically difficult, even within a single waterbody. This has been achieved using piscicides and drainage in some parts of New Zealand, although examples exist overseas of successful eradication of fish through concerted integrated management. Netting, trapping, electro-fishing and use of cages, baits and one-way barriers have been used for control purposes in New Zealand, but the ecological outcomes have largely gone unmonitored. Emerging and future technologies that may assist in the management of non-indigenous fish include the use of pheromones to enhance capture rates, the introduction of taxon-specific pathogens and the genetic modification of fish to produce single-sex (male-only) progeny. Environmental DNA (eDNA) techniques show promise for early detection of vertebrate invaders. Warming of lakes due to climate change may increase the number of species able to establish populations and also the potential for serious parasites to be carried by some invaders.
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others


Keywords
9.1 Introduction
Invasions by non-indigenous species are a multistep process, involving entrainment in a vector, transportation to a new site, introduction, followed by establishment and spread (Williamson 1996; Feature Box 9.1). Management of non-indigenous aquatic species can occur at any stage along this process and may involve prevention of establishment, control of numbers once established, or eradication of species (Feature Box 9.2). Such management is essential for the conservation of native flora and fauna, and numerous methods are employed in New Zealand for control or eradication of terrestrial pests. Control of non-indigenous species is also important for achieving biodiversity and water quality outcomes in lake restoration, but methods are much less advanced for freshwater compared to terrestrial ecosystems.
Invasions are commonly self-perpetuating with reproduction allowing the problem to potentially grow and spread through time, unlike water quality issues which will typically abate once the source activities cease. In the same way that the water quality of a lake cannot be restored long-term without considering the catchment, management of invasions cannot be considered without taking into account species not yet introduced to a site. Thus, in-lake control or eradication of non-indigenous animals makes little sense without reducing the probabilities of invasion or re-invasion. Eradication of smaller species in lakes, particularly invertebrates that produce resistant diapausing stages, is typically impossible once establishment has occurred, making restoration to a predetermined biological state out of the question (Galil 2002; Duggan et al. 2006). As such, we take a more holistic view of ‘restoration’, not just focussing on in-lake restoration, but also on prevention—as the old adage goes, ‘an ounce of prevention is worth a pound of cure’ (Leung et al. 2002).
In this chapter, we review invasions of animals into New Zealand lakes, beginning with inventories of non-indigenous species, including their distributions and known effects. We limit ourselves where possible to free-living animal species that have been accidentally or deliberately released by humans into standing freshwaters of New Zealand’s main islands. Thus, we do not consider species living in saline coastal lakes, species found to date only in flowing waters, introduced parasites of freshwater species, offshore or sub-Antarctic islands, or species seemingly self-introduced since human arrival. We also limit ourselves to species that spend a significant amount of their lives within freshwater (i.e. non-native waterfowl, which primarily live on the water, are not considered). Translocations of native species are also not considered, although they may compromise lake restoration outcomes through effects on food webs; for example, the native common smelt (Retropinna retropinna) has been introduced into many North Island lakes (see Chap. 10) to provide food for introduced trout. We review management techniques for non-indigenous species, including pre-border, border and post-border controls, and in-lake restoration approaches involving invasive species management.
Box 9.1 Ballast Water Management in the Laurentian Great Lakes
Hugh J. MacIsaac grid.267455.7 0000 0004 1936 9596 Great Lakes Institute for Environmental Research The University of Windsor Windsor ON CanadaBallast water has been a principal vector for introduction of non-indigenous species (NIS) since its advent approximately one century ago. Ballast water is rather unique among vectors transmitting NIS in that it may harbour an enormous diversity of species, each present at different abundance. In the Great Lakes, ballast water accounts for at least 55% of established NIS reported since the modern Seaway opened in 1959. Species introduced originate primarily from Europe, with a strong contingent from the Black-Caspian Sea region. This contingent includes highly invasive molluscs (zebra and quagga mussels), fishes (round goby) and crustaceans (fishhook waterfleas), all of which entered the lakes in the last 30 years. Benthic invertebrate biomass in regions of the Great Lakes may be heavily dominated by these species (e.g. >99% dreissenid mussels in Lake Erie).
Global trade has resulted in new opportunities for NIS to colonise areas that were previously inaccessible. This pattern may result from hub-and-spoke or stepping stone invasions patterns. In addition, invasions by NIS are unlike traditional forms of pollution which, when countered by an appropriate management action, may abate in severity. By contrast, self-replicating NIS may continue to plague affected systems even after vector management has been implemented. It is for this reason that preventing further invasions is essential to reducing spread of even more NIS. Regulatory ballast water management was first implemented in the Great Lakes in 1993 for vessels arriving from overseas with filled ballast tanks. This policy was then extended to loaded vessels carrying only ballast residuals, beginning in 2006. Currently, all vessels arriving from foreign destinations must ensure that ballast water or residuals have been exchanged or flushed on the open ocean, thereby minimising opportunities for freshwater NIS to remain in tanks upon entry to the system. The policy appears to be successful in that no new ballast-mediated NIS have been reported in the system since 2006. This dramatic reduction in apparent invasion rate seemingly occurred as a result of a reduction in propagule pressure for freshwater invaders carried rather than the number of high risk species carried in the tanks. In this regard, the current ballast water issue for the Great Lakes resembles that in New Zealand, where vessels do not enter freshwater and, thus, are incapable of discharging NIS into vulnerable ecosystems.
Box 9.2 Carp Eradication: Tasmania
Chris Wisniewski grid.493010.f 0000 0004 5929 3979 Inland Fisheries Service New Norfolk Tasmania AustraliaAfter Cyprinus carpio (European carp) were found in lakes Crescent and Sorell in the central highlands of Tasmania, the Inland Fisheries Service (IFS) established the Carp Management Program (CMP). The CMP has successfully eradicated carp from Lake Crescent, using a range of techniques that continue to be used in Lake Sorell today. In January 1995, an angler found a fish being eaten by a sea eagle on a bank at Lake Crescent. He handed the remains of the fish to the IFS which confirmed it was carp. The Service then surveyed the lake using backpack electrofishing and confirmed the presence of carp. Soon after, carp were also found in Lake Sorell. Both lakes were closed to the public using provisions of the Inland Fisheries Act 1959. As part of the CMP, outflows from both lakes were modified to enable screening of water to 1 mm. Twenty years of surveys show that the screens have been successful as no carp have been found downstream from the lakes. The CMP uses persistent physical effort through a range of techniques including biotelemetry, trapping, netting, electrofishing, spot poisoning, spawning sabotage and pheromone attractants. 7797 carp were removed from Lake Crescent. The last wild carp, a mature female, was caught in December 2007. This is a significant result given the 12 years of effort including the development of the approach that integrates such a broad range of techniques. Lake Crescent was declared carp free in 2009 (see Fig. 9.1). In 2009, estimates indicated that carp numbers in Lake Sorell had been reduced to below 50. However, efforts were setback when the carp were able to spawn. The CMP’s current priority is to prevent any further spawning while continuing to fish out the 2009 cohort. About 14 km of purpose built barrier netting is being used to block the extensive wetlands that are preferred spawning sites. Over 40,000 carp have been removed from Lake Sorell. A capture/mark/recapture population study that started in 2011 indicates that around 7000 maturing carp remain. These fish grow slowly in Tasmanian conditions with some males beginning to mature in their fourth year with the first of the females at least a year behind. The CMP continues to refine old methods and develop new in its integrated approach to carp eradication.

Monthly carp captures from Lake Crescent
9.2 Inventories and Distributions
Numerous aquatic species have established non-indigenous populations in New Zealand lakes, while many others are currently entrained in vectors such as aquaria (e.g. numerous fish species) or freshwater aquaculture facilities (e.g. Malaysian freshwater prawns, Macrobrachium rosenbergii, and grass carp, Ctenopharyngodon idella). Some species have been introduced to natural waters for specific purposes (e.g. the grass carp for aquatic weed control) but fail to establish self-sustaining populations because they are unable to reproduce in the recipient environments. The following section provides an overview of non-indigenous aquatic invertebrate and vertebrate species that have established populations in standing freshwaters in New Zealand.
9.2.1 Invertebrates
At least 23 species of non-indigenous invertebrate have established populations in New Zealand freshwater lakes (Table 9.1), and it is likely a number of others are currently unrecognised. The vast majority of these non-indigenous invertebrates have been introduced unintentionally, with many entering incidentally through the aquarium trade. The exceptions are Lymnaea stagnalis introduced as trout food (Pullan et al. 1972) and marron (Cherax tenuimanus) introduced for aquaculture purposes. In general, the more recent species recorded as established are smaller in size than those introduced earlier, most likely reflecting more stringent border controls.
At least eight zooplankton species have established non-indigenous populations in New Zealand lakes, four of which have been discovered since 2000 (Table 9.1). All are likely to have been unintentionally introduced with flora or fauna intentionally imported or in association with contaminated equipment (e.g. fishing gear, dam building equipment). Three are Daphnia species, adding to our two currently recognised native Daphnia species (Burns et al. 2017). The North American Daphnia galeata has become the most common and widespread Daphnia species in the North Island since its first record in 2000, and it is now also spreading through the South Island (early records of Daphnia dentifera reported by Duggan et al. (2006) have since been confirmed as D. galeata). Another North American species, Daphnia ‘pulex’, was first observed in South Island lakes in 2005 and is now spreading rapidly throughout that island (Duggan et al. 2012). More recently, this species has also been found to be common in Auckland ponds (Branford and Duggan 2017). Daphnia obtusa has been recorded near Dunedin (Chapman et al. 2011), but has not been observed for a number of years, either through lack of sampling or because it has been extirpated. If present, the identity of this species requires re-evaluation, as the taxonomy of this and closely related species has long been confused and is only being clarified through recent genetic advances (Benzie 2005). However, D. obtusa has also been recognised in Australia from a dam impoundment constructed in 1964, indicating it is non-indigenous there also (Benzie and Hodges 1996). Further, a record of Daphnia lumholtzi, which is native to Africa, Australia and Asia, in Lake Rotoroa, South Island (Jolly 1955), is doubtful.
The effects of non-native Daphnia species in New Zealand are unclear. However, Daphnia, including the native species, were uncommon inhabitants of New Zealand lakes prior to 1990, but they are now extremely widespread and abundant and, as noted earlier, populations are dominated by non-indigenous species. Balvert et al. (2009) noted a large reduction in rotifer densities, likely resulting from the superior competitive abilities of Daphnia galeata following its invasion in a North Island lake. In addition, due to their high efficiency as filter-feeders relative to other zooplankton, a large increase in water clarity followed, at least in the short-term (Balvert et al. 2009). Being larger than most native zooplankton, the new Daphnia species may alter energy flows to higher consumer levels through, for example, greater susceptibilities to predation than smaller species.
Five non-indigenous calanoid copepod species have been recorded, primarily associated with constructed waters (e.g. dams, retired quarries, ornamental ponds; Banks and Duggan 2009). Due to distinct morphologies and geographical distributions within the group, species can be readily identified and their origin determined. The Australian species Boeckella symmetrica and Boeckella minuta have been known for many years from the North Island, confined to water supply and hydroelectricity reservoirs, a retired quarry and an ornamental pond. The North American species Skistodiaptomus pallidus and the Japanese Sinodiaptomus valkanovi were both found in 2000, but likely existed in New Zealand for longer, being found initially in constructed urban ponds not typically sampled by New Zealand freshwater biologists (Duggan et al. 2006). All of these calanoid copepods were, until recently, thought to be confined to constructed waters.
The populations of S. valkanovi can be traced genetically to the north of Honshu, Japan (Makino et al. 2010), but as with other calanoid copepod species the localities provide little clue as to how this species came to be in New Zealand. This species, first described from a botanic garden in Bulgaria, was recorded in New Zealand from the Auckland Wintergardens glasshouse, suggesting that it may have been moved in association with aquatic macrophytes. Skistodiaptomus pallidus was also first recorded in a botanic garden (Auckland Regional Botanic Garden ponds) and was found soon after among Daphnia carinata cultures being sold as live food in aquarium stores (Duggan et al. 2006), highlighting another potential vector of zooplankton invasion. Skistodiaptomus pallidus has recently spread into natural lakes, and this has been linked to movement of grass carp for the control of macrophytes (Duggan et al. 2014; Branford and Duggan 2017). Although all recent records have been from lakes where grass carp have been released, some of the earliest records are not, indicating this species may have been introduced by other means and is being spread by different vectors. Finally, the Australian species Calamoecia ampulla was recorded once at an unknown South Island location. Overall, little is known of the ecological effects of these calanoid copepod species, although in the longitudinal study of Duggan et al. (2014) from Lake Kereta, Auckland region, S. pallidus was observed to reach high abundances following establishment and suppressed the abundances of the native calanoid Calamoecia lucasi.
The majority (nine) of the remaining non-indigenous invertebrate species are molluscs, and all but one of these are gastropod species (Table 9.1). Most will have arrived in New Zealand in association with aquatic plants and many have spread widely in lakes. The snail species all feed on benthic algae and as such may compete with other grazers in New Zealand. For example, Physa acuta is thought to have displaced the native Glyptophysa variabilis in much of its range (Winterbourn 1973; Collier 1993). Also, aquatic snails are intermediate hosts for a number of diseases. Lymnaea stagnalis is an intermediate host of the fluke Echinostoma, the cause of echinostomiasis in humans, while Planorbarius corneus can carry the gapeworm nematode (Syngamus trachea) which affects birds, and P. acuta is likely an intermediate host for swimmer’s itch (Champion et al. 2012). In addition to species listed from standing waters in Table 9.1, Melanoides tuberculata is established in a geothermal stream at Golden Springs (Duggan 2002), and an individual apple snail Pomacea diffusa has been recorded from the Waikato River (Collier et al. 2011); neither species would be likely to form self-sustaining populations in lakes, except under circumstances where water is heated through geothermal activity or by industrial cooling-water discharges.
The freshwater jellyfish Craspedacusta sowerbii, native to the Yangtze Valley in China, was first recorded in New Zealand in the 1950s. Observations to date have primarily been of the medusa (jellyfish) stage, which periodically blooms in some lakes. Boothroyd et al. (2002) examined the feeding of the medusa and determined that although zooplanktivorous, they are unlikely to have a major impact on the zooplankton community except possibly a temporally limited impact during bloom periods. A recent survey of Craspedacusta polyps showed the species to be in a high proportion of North Island lakes, in places where medusa have never been observed (Duggan and Eastwood 2012); nothing is known of the feeding effects of this life stage in New Zealand.
At least two annelid species are considered potentially non-indigenous in standing waters in New Zealand, although they occur in a broad range of habitats internationally; the leech Barbronia weberi (Mason 1976—from man-made fish ponds) and the oligochaete Lumbriculus variegatus (Brinkhurst 1971; Talbot and Ward 1987—from lakes). When first observed in New Zealand, Mason (1976) noted B. weberi was found only in artificial environments such as man-made fish ponds or aquaria, giving clues to its vector for introduction. Where introduced elsewhere, it is also noted that it has a close association with common aquarium plants (e.g. Genoni and Fazzone 2008). Branchiura sowerbyi has been found in flowing waters in New Zealand, but elsewhere is commonly found in stagnant ponds (e.g. Aston 1968; Georgieva et al. 2012); as such, it likely inhabits New Zealand ponds where few benthic macroinvertebrate studies have been conducted. Another non-indigenous leech, Helobdella europaea (Collier et al. 2014), and the non-indigenous oligochaetes, Eiseniella tetraedra (Marshall and Winterbourn 1978) and Stylodrilus heringianus (Marshall 1978), are also known only from New Zealand flowing waters; a number of Naididae species are possibly also non-indigenous in New Zealand (Glasby et al. 2007). All these species currently occupying lotic habitats in New Zealand also have the potential to invade standing waters.
Finally, several non-indigenous insects are considered established or vagrant in New Zealand. The mosquito Culex quinquefasciatus has been recorded widely in New Zealand from small artificial standing waters (e.g. tyres, gully traps) and is a known vector elsewhere for the parasitic roundworm Wuchereria bancrofti (the major cause of lymphatic filariasis), Plasmodium (avian malaria), myxomatosis, and other diseases (Holder 1999). Other insect species, including the damselfly Ischnura aurora; the dragonflies Hemianax papuensis, Tramea loewii and Hemicordulia australiae (Rowe 1987); waterboatman Agraptocorixa hirtifrons (Young 2010); the whirligig beetle Gyrinus convexiusculus (Wise 1989); and the lacewing Sisyra rufistigma (Wise 1998), are likely self-introduced from Australia and are thus not included in Table 9.1.
A number of other invertebrate species are likely to have invaded freshwaters, but have not yet been discovered or recognised. For example, Duggan (2010) found two species of harpacticoid copepod, Nitokra pietschmanni and Elaphoidella sewelli, amongst the bottom sediments of home aquaria. Although these are both likely to survive in New Zealand lakes, neither has yet been recorded in the wild, although perhaps due only to a lack of expertise and surveys for these taxa. The ostracod Eucypris virens, otherwise considered native to the Holarctic, is considered an invader in Australia (Koenders et al. 2012) and has been recorded in New Zealand from temporary waterbodies and farm ponds around Auckland (Barclay 1968). Genetic analysis of this species, and other small invertebrates shared between New Zealand and elsewhere, is required to better determine their status as native or non-indigenous.
9.2.2 Fish
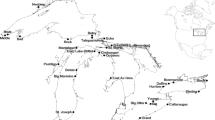
Of the 63 freshwater fish species known to occur in New Zealand (i.e. outside of captivity), 21 are introduced and 19 are (or were) considered established; 17 of these species spend a significant period in freshwater lakes (Table 9.2). All of these non-indigenous fish have been deliberately introduced, either legally, particularly by acclimatisation societies as sport fish, or illegally with a range of intentions. Several of these fish have the potential to compromise lake restoration goals when present in high numbers, for example, by exacerbating other pressures and adversely affecting water quality, habitat and/or native biodiversity. Mechanisms for these effects include (1) bioturbation that can decrease water clarity and, along with excretion, transfer bioavailable nutrients to the water column, (2) degradation of habitat through mobilisation of sediment and direct consumption of aquatic plants and (3) food-web effects through top-down or bottom-up control of other biota (Collier et al. 2015). The larvae of all invasive fish species feed on zooplankton and the high fecundity of coarse fish in particular means there is considerable potential for their larvae to have cascading effects on algal grazing by zooplankton and, therefore, water quality.
Salmonids, mainly brown trout (Salmo trutta) and rainbow trout (Oncorhynchus mykiss), occur throughout the country, but are less common in warmer parts of northern North Island. There are four species of trout and two species of salmon present in New Zealand, but only brown and rainbow trout are widespread in lakes; feral stocks of Salmo salar are considered close to extinction. Trout typically spawn in lake tributaries but can also use exposed shorelines where conditions are suitable, although generally spawning habitat is limited and stocking occurs for recreational fishing purposes (Rowe and Graynoth 2000). Adult trout are opportunistic feeders in lakes, including on native fish and large invertebrates. The other trout species and salmon have restricted distributions and are rarely abundant, although tiger trout (brown trout-brook trout hybrids) are stocked into some Rotorua lakes (D.K. Rowe, personal communication).
Common carp (Cyprinus carpio) are widespread in northern New Zealand, but do not occur in the South Island following eradication of an isolated incursion. Carp can be considered as ‘nutrient pumps’, consuming organic sediments and excreting bioavailable nutrients into the water column, contributing to elevated levels of chlorophyll a and cyanobacteria (Weber and Brown 2015). Carp rarely consume macrophytes directly but dislodge roots from fine sediment during feeding, and the increased turbidity they create attenuates light for macrophyte growth (Weber and Brown 2009). Carp feeding also undermines banks and degrades benthic habitats, partly accounting for an observed negative relationship between carp biomass and benthic invertebrate taxa richness (Vilizzi et al. 2014). Carp move into interconnected shallow lakes, as well as inundated floodplains and wetlands, in spring and summer where they congregate in spawning aggregations in shallow, weedy areas (Tempero et al. 2006; Hicks and Ling 2015).
Goldfish (Carassius auratus) occur throughout both the North and South Islands but are much more numerous in the north. Goldfish can withstand temperatures from freezing to over 35 °C, very low oxygen levels and have been associated with adverse impacts on water quality through resuspension of sediment and nutrients during feeding, in a similar way to carp. It has been shown that growth of cyanobacteria is stimulated by passage through goldfish intestines (Kolmakov and Gladyshev 2003), suggesting high numbers may contribute to the development of algal blooms in enriched waterbodies (Morgan and Beatty 2007). Their main impact on native fish is likely to be through competition for food and other resources. Spawning occurs in spring and summer among macrophytes in shallow margins of lakes (Hicks et al. 2010).
The catfish brown bullhead (Ameiurus nebulosus) is widespread throughout the middle of the North Island with scattered records from the South Island, and it has been associated with impacts on benthic and littoral food-webs in lakes. In Lake Taupo, for example, catfish from weedy habitats were found to feed predominantly on gastropods, cadddisflies, cladocerans and chironomids (Barnes and Hicks 2003), and elsewhere they have also been reported to feed on eggs of native fish and extensively on crayfish (Rowe and Graynoth 2000; Clearwater et al. 2014). The wide-ranging diet of catfish makes them likely to both compete with other benthivorous native fish such as eels and they can include native fish in their diets (Collier et al. 2018). They can survive long periods out of water if the skin is kept moist and are, therefore, extremely resilient to adverse conditions, difficult to eradicate and easy to spread inadvertently. As well as modifying invertebrate communities, their benthic feeding has the potential to affect ecosystem processes and nutrient status through stirring up bottom sediments. Thus, increased rates of nutrient cycling caused by catfish may contribute to higher productivity in lakes (e.g. Hicks et al. 2001).
Gambusia (Gambusia affinis) was introduced into New Zealand for mosquito control but is ineffective at this. The species occurs mainly in upper half of the North Island (north of Taupo), with populations also in Taranaki, Hawkes Bay, Manawatu and Wellington, but they are absent from the South Island except at a few sites in and around Nelson where eradication is underway (Grainger 2015). Gambusia proliferates in shallow margins, mainly around aquatic plants in summer and autumn months. Rapid growth and turnover makes them capable of colonising new habitats rapidly. Each female is capable of producing up to 130 live young and 2–3 broods between November and April each year, although this varies with size and up to 9 broods over 6–7 months have been reported (Pyke 2005). High numbers of gambusia have been associated with negative effects upon a range of fish, invertebrate and amphibian species worldwide, through direct predation or competition (e.g. Lloyd et al. 1986). In New Zealand, there are reports of gambusia feeding upon juveniles and fry of threatened black mudfish (Neochanna diversus) which can inhabit lake margins, aggressively attacking dwarf inanga (Galaxias gracilis) in a dune lake, forcing small indigenous fish species into deeper waters, and competing with native fish (e.g. bully, smelt) for space and food in shallow lake margins when they occur at high densities (e.g. Barrier and Hicks 1994; Rowe 1998; Ling 2004; Rowe et al. 2007). Gambusia is also responsible for the extinction of Galaxias gracilis in Lake Kai Iwi, Northland (Rowe 2003; Pingram 2005).
Rudd (Scardinius erythrophthalmus) have been introduced to many lakes in the North Island where they are a concern for native macrophyte populations because of the preference by adults for feeding on native aquatic plants, giving introduced macrophytes a competitive advantage (Wells 1999). Their feeding may also suppress the regeneration of macrophytes in turbid lakes and thereby prevent re-establishment of native Nitella and Potamogeton species in restoration efforts because they preferentially graze on growing tips or young plants (Lake et al. 2002). High densities of rudd may contribute to reduced water clarity in shallow lakes through increased wind disturbance following macrophyte loss and a reduction in large zooplankton from larval feeding. Because they only digest about 30% of plant material and damage plants while feeding, rudd release phosphorus to the water (Ravera and Jamet 1991), potentially promoting the succession of submerged macrophytes to algal dominance and eutrophication in lakes. Diet of small rudd (56–65 mm fork length) overlaps significantly with common smelt and shoaling galaxiids [e.g. dwarf inanga and koaro (Galaxias brevipinnis)], which feed at mid-water and from the surface, suggesting the possibility of competition for food resources with some native fish (Cadwallader 1977; Lake et al. 2002). Large fish can also prey on native bullies and populations increase rapidly as a result of multiple spawnings, making them difficult to control (Rowe and Graynoth 2000). Rudd can also have sociocultural impacts on trout angling as they can take lures more readily than trout leading to angler disinterest (see Rowe and Champion 1994).
Adult tench (Tinca tinca) inhabit shallow lakes, reservoirs, ponds and wetlands in the northern and central parts of the North Island and in scattered locations around the South Island. They are benthivorous and feed mainly at night, consuming benthic invertebrates and detritus, although some fish can feed solely on zooplankton (Rowe and Graynoth 2000). Tench thrive in both clear and turbid waters, so the high suspended solids levels occurring in turbid lowland New Zealand lakes are unlikely to affect them. Generally, tench seem unlikely to have direct effects on other fish which they do not eat, but have been implicated in reduced densities of some benthic invertebrates (Rowe 2004). Tench are also known to be selective planktivores and so may exert top-down effects (i.e. a reduction in zooplankton) on some lake ecosystems, thereby increasing phytoplankton and reducing water clarity (Rowe and Champion 1994). Tench prefer temperatures 20–21 °C and are rarely found in waters over 25 °C.
Perch (Perca fluviatilis) occur throughout most of the west coast of the North Island and the east coast of the South Island of New Zealand. In lakes and ponds, they are found mostly around margins or shallow water close to large beds of macrophytes and/or emergent plants such as rushes. In lakes, larval perch are generally pelagic zooplankton feeders that form shoals in shallow, open waters and along littoral zones. Adults are aggressive piscivores, and they have been implicated in the decline of native fish and crayfish populations in ponds, lakes and small South Island tarns and a northern New Zealand dune lake (Rowe and Smith 2001; Closs et al. 2003; Ludgate and Closs 2003). Stunted populations of perch can contribute to reduced water clarity through consumption of zooplankton and consequent reduction in algal grazing (Rowe 2007). However, reduction in adult perch numbers can lead to the proliferation of zooplanktivorous fish, and overseas perch stocking has been utilised to suppress these with the aim of improving water quality. Perch spawn during late winter–spring and eggs die when water temperatures rapidly increase above 12 °C; consequently, temperature is a key factor limiting perch distribution and growth in freshwaters.
Many lakes have multiple species of invasive fish and, in combination with native species, form novel communities with potentially complex and unknown interactions that may have additive, synergistic or even antagonistic effects on lake ecology and water quality (Feature Box 9.3). For example, Rowe (2007) reported that the number of fish introduced into small North Island lakes affected the relationship between water clarity and lake depth, and he concluded that control of just one species may not result in an improvement in water clarity because of the interacting effects of multi-species assemblages on lake trophic processes. Thus, where invasive species control forms part of a restoration action plan in lowland lakes, a knowledge of community-level interactions and trophic linkages is required to predict ecological and water quality outcomes.
9.2.3 Other Vertebrates
Of the three species of frog introduced from Australia, two bell frogs spend part of their lives in standing waters. Litoria aurea is restricted to the top half of the North Island, whereas Litoria raniformis is widespread throughout the country. Both were introduced for mosquito control. A major cause of concern regarding these bell frogs is that they serve as vectors of amphibian diseases, such as Chytridiomycosis, which was responsible for a dramatic decline in populations of the native frog Leiopelma archeyi on the Coromandel Peninsula in 2001 (Bishop 2008). Litoria tadpoles largely feed on algae and dead insects (Pyke and White 2001). Red-eared slider terrapins (Trachemys scripta elegans) and snake-neck turtles (Chelodina longicollis) have been recorded periodically in New Zealand lakes, but are not known to have established self-sustaining populations due to temperature requirements for breeding. Both species are long-lived and feed on a range of aquatic plants, small invertebrates and fish (e.g. Dreslik 1999).
Box 9.3 Managing Invasive Fish in the Laurentian Great Lakes; ‘Darwin’s Dreampond’ or Nightmare?
Randall M. Claramunt Tracy L. Galarowicz Matthew E. Herbert Andrew J. Tucker W. Lindsay Chadderton grid.448352.c Michigan Department of Natural Resources Lansing MI USA grid.253856.f 0000 0001 2113 4110 Central Michigan University Mount Pleasant MI USA grid.422375.5 0000 0004 0591 6771 The Nature Conservancy Arlington VA USAThe Laurentian Great Lakes are likely the most heavily invaded freshwater ecosystem globally, with food webs dominated by non-native species that have altered ecosystem services and functions with huge consequential reductions in native biodiversity (Pagnucco et al. 2015; Ricciardi 2006). Yet, some have claimed that continual manipulation of the fishery has maintained a productive equilibrium, in contrast to devastation wrought by introductions in other great lakes ecosystems (e.g. Nile Perch in Lake Victoria, sensu Goldschmidt 1996). Management of these binational North American waters has certainly resulted in invasive species control success stories, notably the ongoing suppression of Sea Lamprey (Petromyzon marinus). A combination of traps, barriers, systematic monitoring and larval lampricide treatment has successfully sustained a 90% reduction in sea lamprey abundance (Christie and Goddard 2003). Similarly, stocking of Pacific salmon has significantly reduced populations of Alewife (Alosa pseudoharengus) and Rainbow Smelt (Osmerus mordax) (Bunnell et al. 2013). This strategy has allowed Great Lakes fisheries managers to develop an economically important salmon fishery, while simultaneously controlling these two invasive prey fish, whose high abundance adversely affected native fisheries and in the case of the former regularly fouled beaches and water intakes during major die-off events.
However, over the last 15 years, the fish forage base has become increasingly unpredictable following invasive quagga mussel (Dreissena bugensis) proliferation, which has resulted in concentrations of nutrients in nearshore and benthic zones (Cuhel and Aguilar 2013) and increasing risk of fishery collapse. The recent failure of the Chinook Salmon (Oncorhynchus tshawytscha) fishery in Lake Huron (O’Keefe and Miller 2011) following a crash of the alewife population is case-in-point to this daunting management challenge. This nutrient disruption and the subsequent loss in offshore alewife and salmon production is observed across the Great Lakes in varying degrees, but has opened the door for the recovery of native fish such as Cisco (Coregonus artedi), Lake Trout (Salvelinus namaycush), Lake Whitefish (C. clupeaformis) and Walleye (Sander vitreus). Unfortunately, gains in native fish recovery linked with Alewife and Rainbow Smelt declines now appear to be hampered by a new suite of invasive species that consume native fish eggs and cause recruitment failures (Claramunt et al. 2005). The round goby (Neogobius melanostomus) and rusty crayfish (Orconectes rusticus) have now invaded the Great Lakes coastal zone. These benthic egg predators (see trout-egg-baited traps in Fig. 9.2) are reaching abundance levels three to five times that of the native species which previously occupied coastal habitats (Bronte et al. 2003; Jonas et al. 2005). Round goby have become the most abundant and conspicuous near shore benthic predator in the Great Lakes, and to complicate matters further is also an increasingly important part of the prey base (Crane et al. 2015).

Baited underwater images taken on shallow lake trout spawning reef habitat in Grand Traverse Bay, Lake Michigan, showing round goby and rusty crayfish congregating around a bait bag containing lake trout eggs. Note also live and dead dressenid mussels in interstitial spaces of cobble habitat in the right image. Images by Krista Robinson, Central Michigan University
The tension between coping with the complex and evolving synergistic interactions between native and invasive species, responding to the next looming invasion crisis (e.g. Asian carp), coupled with effects of other anthropogenic stressors (Pagnucco et al. 2015; Allan et al. 2013) continues to be a monumental challenge for Great Lakes protection and management. For Darwin’s dreampond, the Laurentian Great Lakes is an important testing ground globally, for both the concepts of species adaptation in a rapidly changing environment as well as development of tools and approaches to prevent or manage species invasions (Bailey et al. 2011; Jerde et al. 2011; Kolar and Lodge 2002; Christie and Goddard 2003). Darwin’s nightmare, however, may very well be that the species most unable to adapt to the rapid environmental changes in the Great Lakes, to which they are most critically linked and dependent on, are not the fish but the humans.
9.3 Review of Control Techniques
Management and control of non-indigenous species can occur at a number of stages in the invasion process. Management can occur prior to the transportation phase (pre-border), during the transportation phase (i.e. border control or post-border) or following introduction or establishment to a new waterbody (post-border). Pre-establishment management is covered here as in-lake restoration is largely impossible for most invaders once established. Indeed, ignoring the possibility for reducing the probability of establishment of new non-indigenous species in lakes is akin to disregarding the catchment of a lake when restoring the effects of nutrient additions.
9.3.1 Pre-border Controls
A number of laws and regulations exist in New Zealand to reduce the number of species able, or allowed, to enter the country. This pre-border control is the first line of defence in reducing the establishment rate of non-indigenous species into lakes, attempting to allow entry of only a subset of species that have low probability of survival, reproduction or potential for effects. The primary pieces of legislation with respect to the importation of living things, intentionally or accidentally, are the Biosecurity Act 1993 and the Hazardous Substances and New Organisms Act 1996. The Biosecurity Act states that importers of risk goods (including live animals) must take all reasonable steps to ensure that the goods comply with applicable ‘Import Health Standards’. These standards are based on risk assessments (Hayden and Whyte 2003), and specify requirements that need to be met before importation, providing explicit directions on what measures need to be taken before goods can be imported.
Several standards are applicable to animals that might establish populations in New Zealand lakes. One standard that allows for the deliberate importation of many freshwater species is the ‘Import Health Standard for Ornamental Fish and Marine Invertebrates from all Countries’. This is essentially a ‘white list’, naming 1010 permissible freshwater aquarium fish species. Previously, this list comprised 280 names, of which 178 were genera, some of which contained multitudes of species (McDowall 2004). A subset of the 1010 permissible species (44 species) is identified as ‘high risk’, due to being susceptible to one or more diseases, and must meet additional pre-quarantine or quarantine measures before clearance.
The Hazardous Substances and New Organisms Act 1996 regulates the importation of ‘new organisms’ into New Zealand. In this Act, the definition of a new organism is one that was not present in New Zealand prior to 29 July 1998; any present before that date do not represent new organisms. If an importer wants to introduce a species, or a genetically modified organism, that is not listed on an import health standard, or was not officially present prior to 29 July 1998, an import permit must be obtained from the Environmental Protection Agency (EPA). The EPA undertakes a risk assessment of potential adverse effects of importing that organism and takes a precautionary approach. As a minimum standard, the EPA must be satisfied that, after entry to the country, the species could not: (1) form self-sustaining populations anywhere in New Zealand; (2) displace or reduce a valued species; (3) cause deterioration of natural habitats; (4) be a parasite, or be a vector or reservoir for human, plant or animal disease; or (5) have any adverse effects on human health and safety or the environment. Biosecurity inspectors are able to seize any organism that they have reason to believe may be a new organism. The Biosecurity Act 1993 also states a person must not provide false, misleading, or incomplete information about goods to be imported or undeclared goods, or take steps that are likely to hinder the detection of undeclared goods by an official.
Several standards also are aimed at preventing aquatic fauna being introduced to the country accidentally. Aquarium plants, which may be contaminated with small invertebrates, including worms, copepods and snails (Duggan 2010), must meet the standard for ‘Importation of Nursery Stock’. Aquarium plants such as Anubius spp., Hygrophila difformis (wisteria) and Echinodorus amazonicus (Amazon sword plant), for example, have had a requirement that they be inspected immediately prior to export for the presence of snails, snail eggs, worms or leeches, and be transported in an aquarium containing a dilute solution of copper sulphate. There is also an Import Health Standard for ‘Used Equipment Associated with Animals or Water’. With respect to freshwaters, used equipment associated with freshwater aquatic animals or activities may be allowed provided they are cleaned, checked and dried prior to arrival into New Zealand, and the equipment needs to be visibly clean and free from contamination with organic material including sediment, soil, weeds or other aquatic organisms. As such, this should also reduce the potential for small aquatic invertebrates to enter, live or as diapausing stages.
Although New Zealand primarily works on a ‘white-list’ system, the Freshwater Fisheries Regulations 1983 also provide a ‘black list’ for some species. The regulation states that no person shall have in their possession or under their control, or rear, raise, hatch or consign, any ‘noxious’ fish. The regulations provide a short list of ‘noxious’ taxa, including walking catfish (Clarias batrachus), pike (Esox lucius), piranha (Pygocentrus spp., Rooseveltiella spp., Serrasalmus spp.) and tilapia (Tilapia spp. or Sarotherodon sp.). Further, the Hazardous Substances and New Organisms Act 1996 specifically prohibits entry of the family Esocidae (e.g. pikes, muskellunge) or stickleback (Gasterosteus spp.). The rationale for listing these in the legislation, and not others that might potentially establish, is not provided. Nevertheless, the provision of white lists for species within Import Health Standards effectively make these listings redundant.
9.3.2 Border Control
Since New Zealand comprises a series of islands, it is in a better position to implement protection at the border to reduce the rate of invasions than many countries elsewhere. At international airports, passengers must complete a ‘Passenger Arrival Card’, which includes a question regarding any animals (or animal products) that may be carried. Further, passengers are typically questioned, and luggage is commonly checked, for organisms that are not allowed into New Zealand. Aquatic animals are occasionally intercepted at the border. For example, in 2013 a Vietnamese man was caught attempting to smuggle seven live tropical cichlids through Auckland airport, hidden in plastic bags in his pockets; he was only caught when water was seen dripping down his trouser leg.
All items of international mail are also checked for biological material. In 2008, six Siamese fighting fish (Betta splendens) sent from Thailand were intercepted at the International Mail Centre, Auckland, with the owner having provided an incorrect declaration; ironically, this species can be bought legally in New Zealand or can be legally imported with a permit. In October 2010, the secretary of the New Zealand Killifish Association was similarly caught importing fish eggs from Sweden and Denmark. An intercepted parcel to the man contained three small plastic bags containing fresh peat moss with killifish eggs. Interceptions of freshwater species have also been made on shipping vessels, such as the Asian tiger mosquito (Aedes albopictus) on a cargo ship arriving from Vanuatu in 2011. These examples highlight the importance of border control to prevent the incursion of non-indigenous species that could compromise lake restoration outcomes in the future.
9.3.3 Post-border, Pre-establishment Management
Post-border legislation is complicated, with a number of different Acts and Regulations being relevant depending on the species involved. These statutes include the Biosecurity Act 1993, the Freshwater Fisheries Regulations 1983 and the Conservation Act 1987. The Biosecurity Act 1993 is the main piece of legislation providing for post-border management or eradication of non-native species.
Post-border control can include species that have passed through the border, but have not yet been released to the wild, or the management of species after they have been introduced or have established in natural waters. Despite pre-border and border control, there is ample evidence that the border is leaky, with a number of incursions of prohibited freshwater animals detected. One species that made it through border controls, but was eradicated prior to entering natural waters, is the North American tadpole shrimp Triops longicaudatus, a species sold elsewhere as diapausing eggs and hatched as pets. In 2005, Triops, received as a present by a 6-year-old boy, was seized at Warkworth Primary School and subsequently destroyed. In 2014, the species was again found being advertised on a New Zealand online auction site; the winner voluntarily surrendered the kit to authorities. In 2007, illegally imported, genetically modified individuals of the zebra fish species Danio danio were found at four properties in Christchurch, from which they were sold and/or bred. Although this is an allowable species in New Zealand and unlikely to establish populations in the wild, being genetically modified designated these individuals as ‘new organisms’ in breach of the Hazardous Substances and New Organisms Act.
Deliberate illegal introduction of species into New Zealand has occurred for some time. In the 1960s, carp were illegally imported by a fish breeder, by incorrectly labelling them as shubunkin (multi-coloured goldfish), and these are now a major problem in many northern New Zealand lakes. Similarly, in 1967, rudd were smuggled into New Zealand in the vegetable room of the liner Rangitoto (Winters 2012). More recently, in 1999, a plastic tube arrangement was used to smuggle gudgeon eggs into New Zealand, passing through customs undetected (Winters 2012).
9.3.4 Post-establishment Control Measures: Invertebrates
Once established, different sets of laws are intended to reduce the spread of species. The Conservation Act 1987 states that ‘no person shall transfer live aquatic life or release live aquatic life into any freshwater in a new location where the species does not already exist, including fish farms’. The Biosecurity Act 1993 also provides for post-border management, allowing for the eradication or effective management of harmful organisms. Strangely, the Freshwater Fisheries Regulations 1983 states more explicitly that no person shall ‘place, liberate, or introduce into any lake, river, or stream any indigenous or exotic species of Mollusca, Crustacea, Protozoa, Insecta, or of annelid, nematode, platyhelminth worm, or oligochaete worm’. While this covers most non-indigenous invertebrate species in New Zealand, it does not include all of them, such as the freshwater jellyfish Craspedacusta sowerbii.
For invertebrates, and particularly for zooplankton, little can be done once a species becomes established other than reducing the rate of spread. By this stage, eradication will typically be impossible, and to date little effort has, therefore, been made to do so. One exception, however, has been the large freshwater crayfish, Western Australian marron, which was originally introduced to New Zealand in 1986 for aquaculture. However, in the early 1990s, a change in government policy led to the commercial marron farm being closed down and animals being destroyed, following concerns about the impact these crayfish could have on native fauna, particularly native crayfish (Paranephrops spp.). The major threat posed by marron was the possibility that the smaller female koura might prefer to mate with the larger male marron (D.K. Rowe, personal communication).
In 1993, an estimated 500,000 marron were destroyed at the Warkworth farm after the fish farmer had his permit revoked. In 2005, however, a lone marron was found on a footpath outside a disused West Auckland service station and an inspection of an adjacent property revealed more marron and also the gudgeon (Gobio gobio). Subsequent investigations revealed two ponds at South Head, Kaipara, containing more specimens of both species. About 500 marron were destroyed by draining the ponds. These marron may have originated from the farm at Warkworth; it is suspected some were removed prior to authorities taking over management of the farm. Elsewhere in the world, non-native crayfish have been eradicated from ponds using pyrethroids (Peay et al. 2006; Sandodden and Johnsen 2010). While chemicals such as pyrethroids or other insecticides are also applicable to crustacean zooplankton (e.g. Wendt-Rasch et al. 2003), the utilisation of resistant diapausing stages by that group, which lie dormant within sediments until conditions are appropriate, will render these techniques temporary at best for restoring indigenous lake zooplankton communities.
A trend among invertebrate invasions in inland waters, both in New Zealand and elsewhere, is that the first sites of establishment are commonly constructed waters (e.g. retired mines and quarries, and dammed river impoundments for hydroelectricity or water supply (e.g. Johnson et al. 2008; Banks and Duggan 2009)). These constructed waters typically have zooplankton species less specialised for pelagic conditions than natural waters, as noted by Parkes and Duggan (2012). Indeed, these authors suggested that newly constructed waterbodies should have native species introduced, so that they develop ‘biotic resistance’ more rapidly as a means to reduce establishment rates of non-indigenous species. Taylor and Duggan (2012) demonstrated this effect experimentally, whereby tanks seeded with sediments from natural lakes developed zooplankton communities more rapidly, and after 1 year more readily repelled the invasion of new species introduced to experimental tanks. In particular, establishment rates of the North American calanoid copepod S. pallidus were reduced, particularly when native calanoid copepods were present, indicating that some species may be key to reducing establishment of particular taxa. Skistodiaptomus pallidus has recently been observed in natural lakes and a link was made by Duggan et al. (2014) between its establishment and the legal releases of grass carp into lakes for aquatic macrophyte control. This link has since been supported by the finding of this copepod in grass carp aquaculture ponds (Duggan and Pullan 2017) and in further ponds that have received grass carp from these farms (Branford and Duggan 2017). The Ministry for Primary Industries is thus in the process of strengthening its requirements for the release of grass carp and silver carp from farming operations, in an attempt to reduce the rate of incidental invasions from this vector.
Management strategies designed to reduce the rate of movement of macrophytes also will aid in reducing the inadvertent movement of invertebrates. For example, signage at popular recreational lakes for boat operators to check and remove weeds from their vessels, motors and trailers is aimed at slowing the rate of invasion of these weeds and the many invertebrates, particularly snails, associated with them. Attempts have apparently not been made to eradicate non-native molluscs in New Zealand to date, likely due to their seemingly negligible current effects on the economy and human or other animal health. Elsewhere molluscicides, such as niclosamide, have been used to eradicate disease carrying snails (Clearwater et al. 2008). With warming of New Zealand due to climate change, and an associated increased risk of tropical diseases being carried by these snails, toxins may need to be considered in the future. Another alternative is complete destruction of habitats in which the species are found if distribution is restricted and the habitats do not have high native biodiversity values. This approach is not desirable or feasible for most lakes, but may be done for small constructed ponds. For example, the calanoid copepod S. valkanovi was eradicated from Bulgaria following the destruction of the pond in Sofia Botanic Gardens from which this species was first described (Duggan et al. 2006). In New Zealand, one population of the copepod Boeckella symmetrica has been eradicated following the infilling of Wiri Quarry (Branford and Duggan 2017).
9.3.5 Control Measures: Vertebrates
A range of options is available for control of non-indigenous fish as part of lake restoration. However, some species are easier to target than others and complete eradication is difficult (Collier and Grainger 2015). To achieve effective control, a combination of methods is typically required and re-invasion pathways need to be blocked. The high fecundity of most invasive fish means that control measures must be either total (i.e. eradication by piscicide) or sustained over the long term to achieve desired ecological outcomes, as spawning by only a few remaining fish can rapidly reinstate population densities. In addition, single species control may lead to ecological surprises, such as releasing other species from competitive suppression or top-down control. An understanding of community-level food-web interactions is, therefore, required to predict the outcomes of invasive fish control and its relative importance in terms of meeting lake restoration targets compared to other factors such as managing catchment land-use practices. In addition, a knowledge of species biology, behaviour and environmental cues is important so key life-cycle bottlenecks can be exploited, such as spawning aggregations and migration routes (Collier and Grainger 2015; David 2015).
Eradication of invasive fish is most cost-effectively achieved through the use of piscicides and drainage. Two types of toxin have been used in New Zealand for invasive fish control: lime [calcium hydroxide (Ca(OH)2) and rotenone (cube root powder)]. Use of lime was restricted to small ponds and involved drainage to reduce water volume before spraying a mixture of water and hydrated lime to achieve a pH of greater than 9 (West 2015). This method is no longer used now that rotenone is more readily available in New Zealand. Rotenone was first used in New Zealand to eradicate rudd, tench and grass carp in Parkinson’s Lake (near Auckland), and native fish were then restocked into the lake (see Rowe and Champion 1994). It is now applied as a cube root slurry (1.2–1.8% rotenone) using a helicopter for large ponds and lakes; to date there have been 81 rotenone operations across the country targeting gambusia, carp, rudd and tench, with three applications on lakes of 10–17 ha (West 2015). Eradication is easier on small waterbodies where outlets can be closed off, although outcomes may be compromised by inputs from springs providing clean-water refugia for fish during poisoning operations. Rotenone was successfully used to eradicate trout from Upper Karori Reservoir and its tributary streams, Wellington, resulting in a dramatic improvement in native galaxiid fish and crayfish numbers in the inlet stream and a shift in zooplankton composition (Pham et al. 2013; Duggan et al. 2015). However, using poison in high quality lakes may result in collateral damage to native species, which is an important consideration when deciding on which control methods to deploy.
Various combinations of nets and other fishing methods can be used to reduce invasive fish biomass in lakes [see Case Study—Rotopiko (Serpentine)], although netting is not effective for all species (e.g. carp). In Lake Ohinewai, the combination of multiple other large nets and traps, seining and electrofishing over several months achieved a reduction in carp biomass from 373 to c.100 kg/ha (see Case Study—Ohinewai). The nets employed included ‘pod traps’, which have one-way doors that prevent fish from escaping and automated feeders that frequently add fresh bait to the trap to improve catch rates (Hicks et al. 2015). Baiting of nets and traps has been shown to improve catch rates of carp but baits lose most of their attraction properties within an hour. Baits can be laced with toxins such as rotenone, although sodium nitrate has been shown to be just as effective (Morgan et al. 2014). However, there can be some aversion to toxins requiring the use of masking agents (Morgan et al. 2013). Boat electrofishing was also used in Lower Karori Reservoir, Wellington, to reduce top-down control of perch populations on zooplankton, leading to improvements in water quality (Hicks et al. 2015). However, once fishing ceased, perch numbers increased reinforcing the need for sustained control of invasive fish numbers to meet lake restoration targets.
Case Study 9.1 Rotopiko (Serpentine) Lakes
This lake complex is located to the south of Hamilton in the central North Island and comprises three small connected peat lakes that have high conservation value due to healthy indigenous macrophyte communities and intact native marginal terrestrial and wetland vegetation (de Winton 2014; Price and Gumbley 2015). In addition to several native fish species, including smelt which do not naturally occur there, the lakes also contain the catfish brown bullhead, goldfish, gambusia and rudd, with the latter species of most concern because of its propensity to consume aquatic plants (Lake et al. 2002). A restoration programme for the lakes has been ongoing for several years and, since 2001, has included a netting programme for pest fish targeted at rudd control. Fine mesh monofilament gill nets are deployed annually in spring prior to spawning, and submerged aquatic vegetation is monitored twice yearly (de Winton et al. 2006).
Gill netting appears to have been effective for suppressing rudd numbers, but the effect on indigenous fish populations is difficult to define due to the lack of observations before netting started. However, the indigenous fish populations of the Rotopiko (Serpentine) Lakes are relatively large compared to pest fish population (Wu et al. 2013), pointing towards a positive effect of the netting programme. Notwithstanding the decline in rudd populations, compensatory changes in catfish and goldfish populations may continue to pose a threat to macrophyte populations in these lakes, but their effects are as yet unknown. No improvement in scores of a macrophyte health index (LakeSPI; see Chap. 8) was evident following initiation of fishing in 2001, although the absence of data collected before fish removal makes drawing conclusions difficult. It is possible that planktonic algae in the lake water or self-shading by luxuriant macrophyte growths, promoted by enrichment, limits the amount of light available for macrophytes. Under this situation, a certain level fish grazing may, in fact, help to enhance macrophyte cover and diversity, suggesting that management of rudd numbers rather than eradication could be considered for restoration purposes.
Case Study 9.2 Lake Ohinewai
This is a shallow (4.5 m depth), 16.8 ha lake on the floodplain of the Waikato River, central North Island, where catchments are highly developed for agriculture and extensive drainage, and flood control measures regulate river and lake levels. Lake Ohinewai deteriorated from a stable oligotrophic (macrophyte-dominated) state to a stable eutrophic (algal-dominated) state during the early 1990s. This lake was selected for invasive fish removal with the ultimate goal of improving water quality and restoring indigenous biodiversity. The main fish species of concern is carp, but the lake also supports high numbers of goldfish, brown bullhead and gambusia, along with some rudd.
The immediate goal was to reduce carp to a level at or below 100 kg/ha with the intention of improving water quality and macrophyte cover. Mark-recapture fishing, using a combination of fyke nets, minnow traps, electrofishing, beach seining and baited traps, was successful in reducing carp biomass from 373 kg/ha to below 100 kg/ha (Tempero et al. 2015). To limit reinvasion, a one-way barrier was installed in the lake outlet allowing adult common carp to leave but not re-enter (Fig. 9.3b), and this resulted in a further 50% drop in biomass to approximately 45 kg/ha as fish exited the lake but could not return. The barrier was designed with horizontal bars to allow debris <30 mm to pass through unobstructed and was hinged at the top to allow for easy cleaning in the case of blockage. The bar spacing of the one-way gate installed in the barrier was based on the fish trap design of Thwaites et al. (2010) and included a set of weighted swinging bars at the base of the trap that would allow adult carp and eels to push through the trap when moving downstream but would not allow adult carp to return to the lake for spawning.

Fish barriers deployed downstream of lakes to prevent upstream movement of invasive fish. (a) Lake D—rods prevent fish jumping over concrete weir; (b) Lake Ohinewai carp screen with one-way finger gate below water surface (photo: A. Daniel)
Water quality (Secchi disk transparency and total suspended solids) showed early signs of improvement in Lake Ohinewai, and aquatic macrophytes are reappearing around the margins, although these are mainly introduced Ludwigia and Myriophyllum species. Thus, while regarded as a success in terms of carp control, the ecological outcomes in terms of macrophyte recovery have been compromised by the dominance of invasive plant species, although there may be longer term recovery in native species if seed banks on the lake bed are still viable. Modelling of land management and invasive fish control scenarios showed that integrated catchment management of tributary stream riparian zones and constructed wetlands would be required along with carp control to achieve desired improvements in lake health (Allan 2016).
Recent overseas developments in cage and barrier design for point source control and restriction of movement have been aimed at controlling carp or preventing silver carp (Hypophthalmic molitrix) from accessing the Great Lakes, USA. Barriers can be physical, visual, acoustic or electrical obstructions and may limit movement in one direction or both upstream and downstream directions. Zielinskia et al. (2014) found that curtains of graded and coarse bubbles reduced directional movement of carp by 75–85%, and they concluded that sound and fluid motion were more important than visual cues for restricting movement past the curtains. However, bio-acoustic fish fences can be expensive to implement (Brammeier et al. 2008), and neither these nor electric barriers (e.g. Verrill et al. 1995) have been implemented in New Zealand. Rather, vertical physical barriers ≥1 m high have proven ideal for invasive fish exclusion in this country due to the climbing ability of many migratory native fish and the inability for the current suite of invasive fish to pass such barriers. Vertical barriers can be as simple as a timber weir or a perched culvert, and if necessary weirs can include metal bars or plunge pools to limit upstream ingress by jumping (e.g. Fig. 9.3a; see also Gumbley and Daniel 2015). Native fish passes can be fitted if necessary using ramps, ropes or other devices to ensure populations of most indigenous fish are sustained in restored lakes (David et al. 2014; see also Chap. 10).
One-way barriers exploit the pushing behaviour of carp, and a finger-trap design has been successfully deployed on the outlet of Lake Ohinewai to prevent re-invasion by carp following biomass reduction (Tempero et al. 2015; see Case Study—Lake Ohinewai; Fig. 9.3b). These devices have also been effective in trapping non-native fish at the Lake Waikare separation cage (Fig. 9.4), which removed around 10 tonnes of carp, goldfish and catfish in 2 years (David 2015). Less than 20 individual non-target fish were captured during that period. Upstream netting indicated that large numbers of smaller native fish, but also invasive species (particularly small catfish and goldfish <200 mm), passed through the cage design unharmed, indicating that the fish trap is an effective and reliable tool for mass adult invasive fish harvest with minimal by-catch, but it does not exclude all invasive fish (David 2015). While barriers and exclusion screens can prevent access to target habitats such as high-value lakes (e.g. Thwaites et al. 2010), separation cages can be used to isolate carp in containment areas by capitalising on their tendency to jump out of the water when they encounter a barrier (e.g. Stuart et al. 2006).

Carp-N neutral showing the carp cage on the left and Lake Waikare in the background (photo: B. David)
9.4 Conclusions and Future Prospects
While pre-border and border controls should reduce the rate of new invasions to New Zealand lakes, they clearly still occur and will continue to do so in the future, particularly for smaller invertebrates. Once established, it can be difficult or impossible to eradicate species from the system, making restoration to a pristine state problematic. Fewer control options are available for smaller species than larger species, reinforcing the need for continued border and post-border surveillance so as not to compromise the outcomes of lake restoration efforts. Vectors have yet to be identified for a number of recent invaders, meaning that re-invasion may be difficult to prevent, rendering restoration of native biotic communities futile (i.e. the aim to achieve non-native free sites).
A large number of non-indigenous animals have invaded New Zealand lakes to date, but the current census is likely to be an underestimate, particularly for smaller species. For example, non-indigenous species of harpacticoid copepod are known from amongst the bottom sediments of home aquaria, but they are not yet recognised in the wild. Other non-indigenous species potentially present may simply not be recognised, highlighting a need for genetic screening. Further, many non-indigenous fish species intentionally carried by the aquarium trade are not yet known to be established in the wild but could pose a future threat of establishment with rising temperatures resulting from climate change. Similarly, as a number of non-indigenous invertebrate species in New Zealand are carriers of serious parasites and diseases elsewhere, the potential for some invaders to have effects on human and other animal health correspondingly increases.
Emerging and future technologies for controlling or eradicating non-indigenous fish include the use of pheromones to enhance capture rates, the introduction of taxon-specific pathogens and the genetic modification of fish to produce single-sex (male-only) progeny. As such, opportunities for the control or eradication of some species will increase through time. Additionally, the methods for detecting species are also improving. Techniques that are able to detect an incursion at an early stage, following introduction or early establishment, may greatly increase the possibilities for eradication. For example, ‘environmental DNA’ (eDNA) techniques are an emerging method to detect the presence of vertebrate species, where DNA from material sloughed from animals is detected from water samples (Jerde et al. 2011; see also Chap. 12). The usefulness of eDNA was evaluated during the eradication of brown trout in streams flowing into a Wellington reservoir, with PCR product specific to brown trout able to be amplified from samples before, but not after, eradication (Wood et al. 2013). Continued development of new detection and control technologies will be essential to keep pace with the increasing threat of invasive aquatic animals establishing in New Zealand lakes.
References
Allan M (2016) A coupled hydrodynamic-ecological model of management options for restoration of Lake Ohinewai. ERI Report 68, University of Waikato, Hamilton, New Zealand
Allan JD, McIntyre PB, Smith SDP, Halpern BS, Boyer GL, Buchsbau A, Burton GA Jr, Campbell LM, Chadderton WL, Ciborowski JJH, Doran PJ, Eder T, Infante DM, Johnson LB, Joseph CA, Marino AL, Prusevich A, Read JG, Rose JB, Rutherford ES, Sowa SP, Steinman AD (2013) Joint analysis of stressors and ecosystem services to enhance restoration effectiveness. Proc Natl Acad Sci 110:372–377
Aston RJ (1968) The effect of temperature on the life cycle, growth and fecundity of Branchiura sowerbyi (Oligochaeta: Tubificidae). J Zool 154:129–140
Bailey SA, Deneau MG, Jean L, Wiley CJ, Leung B, MacIsaac HJ (2011) Evaluating efficacy of an environmental policy to prevent biological invasions. Environ Sci Technol 45:2554–2561
Balvert SF, Duggan IC, Hogg ID (2009) Zooplankton seasonal dynamics in a recently filled mine pit lake: the effect of non-indigenous Daphnia establishment. Aquat Ecol 43:403–413
Banks CM, Duggan IC (2009) Lake construction has facilitated calanoid copepod invasions in New Zealand. Divers Distrib 15:80–87
Barclay MH (1968) Additions to the freshwater ostracod fauna of New Zealand. N Z J Mar Freshw Res 2:67–80
Barnes GE, Hicks BJ (2003) Brown bullhead catfish (Ameiurus nebulosus) in Lake Taupo. In: Managing invasive freshwater fish in New Zealand. Department of Conservation workshop, Hamilton, May 2001, pp 27–35
Barrier RFG, Hicks BJ (1994) Behavioural interactions between black mudfish (Neochanna diversus, Stokell, 1949: Galaxiidae) and mosquitofish (Gambusia affinis Baird & Girard, 1854). Ecol Freshw Fish 3:93–99
Benzie JAH (2005) The Genus Daphnia (including Daphniopsis) (Anomopoda: Daphniidae). Guides to the Identification of the Microinvertebrates of the Continental Waters of the World, vol 21. Kenobi Productions, Ghent and Backhuys, Leiden
Benzie JAH, Hodges AMA (1996) Daphnia obtusa Kurz, 1874 emend Scourfield, 1942 from Australia. Hydrobiologia 333:195–199
Bishop PJ (2008) Bell frog populations in New Zealand – good news or bad news? Aust Zool 34:408–413
Boothroyd IKG, Etheredge MK, Green JD (2002) Spatial distribution, size structure, and prey of Craspedacusta sowerbyi Lankester in a shallow New Zealand lake. Hydrobiologia 468:23–32
Brammeier J, Polls I, Mackey S (2008) Preliminary feasibility of ecological separation of the Mississippi River and the Great Lakes to prevent the transfer of aquatic invasive species. Great Lakes Fisheries Commission, Chicago
Branford SN, Duggan IC (2017) Grass carp (Ctenopharyngodon idella) translocations, including hitchhiker introductions, alter zooplankton communities in receiving ponds. Mar Freshw Res 68:2216–2227
Brinkhurst RO (1971) The aquatic Oligochaeta known from Australia, New Zealand, Tasmania, and the adjacent islands. Univ Queensland papers, Dept Zool 8:99–128
Bronte CR, Jonas J, Holey ME, Toneys ML, McKee P, Breidert B, Claramunt RM, Ebener MP, Eshenroder RL, Krueger CC, Wright G, Hess R (2003) Possible impediments to lake trout restoration in Lake Michigan. Lake Michigan Lake Trout Task Group Report. Great Lakes Fishery Commission, Ann Arbor
Bunnell DB, Madenjian CP, Desorcie TJ, Kostich MJ, Smith KR, Adams JV (2013) Status and trends of prey fish populations in Lake Michigan in 2012. Report to the Lake Michigan Committee, Mar 2013. https://pubs.er.usgs.gov/publication/70160584. Accessed 8 Sept 2018
Burns CW, Duggan IC, Banks JC, Hogg ID (2017) A new, subalpine species of Daphnia (Cladocera, Anomopoda) in the D. carinata species complex, in the South Island, New Zealand. Hydrobiologia 798:151–169
Cadwallader PL (1977) Introduction of rudd Scardinius erythrophthalmus into New Zealand – review of the ecology of rudd and the implications of its introduction into New Zealand. Ministry of Agriculture and Fisheries Technical Report No. 147, Ministry of Agriculture and Fisheries, New Zealand
Champion P, Rowe D, Smith B, Wells R, Kilroy C, de Winton M (2012) Freshwater pests of New Zealand. NIWA Report. National Institute of Water and Atmospheric Research, Hamilton, New Zealand
Chapman MA, Lewis MH, Winterbourn MJ (2011) An introduction to the freshwater Crustacea of New Zealand. Collins, Auckland
Christie GC, Goddard CI (2003) Sea Lamprey international symposium (SLIS II): advances in the integrated management of sea lamprey in the Great Lakes. J Great Lakes Res 29(Supp 1):1–14
Claramunt RM, Jonas JL, Fitzsimons JD, Marsden JE (2005) Influences of spawning habitat characteristics and interstitial predators on lake trout egg deposition and mortality. Trans Am Fish Soc 134:1048–1057
Clearwater SJ, Hickey CW, Martin ML (2008) Overview of potential piscicides and molluscicides for controlling aquatic pest species in New Zealand. Science for Conservation 283. Department of Conservation, Wellington, New Zealand
Clearwater SJ, Kusabs IA, Budd R, Bowman E (2014) Strategic evaluation of kōura populations in the upper Waikato River. NIWA Client Report HAM2014-086, National Institute of Water and Atmospheric Research, Hamilton, New Zealand
Closs GP, Ludgate V, Goldsmith RJ (2003) Controlling European perch (Perca fluviatilis): lessons from an experimental removal. In: Managing invasive freshwater fish in New Zealand. Proceedings of a workshop hosted by Department of Conservation, Hamilton, 10–12 May 2001
Collier KJ (1993) Review of the status, distribution, and conservation of freshwater invertebrates in New Zealand. N Z J Mar Freshw Res 27:339–356
Collier KJ, Grainger N (eds) (2015) New Zealand invasive fish management handbook. LERNZ (The University of Waikato) and Department of Conservation, Hamilton, New Zealand
Collier KJ, Demetras NJ, Duggan IC, Johnston TM (2011) Wild record of an apple snail in the Waikato River, Hamilton, New Zealand, and their incidence in freshwater aquaria. N Z Nat Sci 36:1–9
Collier KJ, Hamer MP, Moore SC (2014) Littoral and benthic macroinvertebrate community responses to contrasting stressors in a large New Zealand river. N Z J Mar Freshw Res 48:560–576
Collier KJ, Allan M, Rowe D (2015) Invasive fish community impacts. In: Collier KJ, Grainger N (eds) New Zealand invasive fish management handbook. LERNZ (University of Waikato) and Department of Conservation, Hamilton, New Zealand, pp 23–28
Collier KJ, Pingram MA, Francis L, Garrett-Walker J, Melchior M (2018) Trophic overlap between non-native brown bullhead (Ameiurus nebulosus) and native shortfin eel (Anguilla australis) in shallow lakes. Ecol Freshw Fish. https://doi.org/10.1111/eff.12400
Crane DP, Farrell FM, Einhouse DW, Lantry JR, Markham JL (2015) Trends in body condition of native piscivores following invasion of Lakes Erie and Ontario by round goby. Freshw Biol 60:111–124
Cuhel RL, Aguilar C (2013) Ecosystem transformations of the Laurentian Great Lake Michigan by nonindigenous biological invaders. Annu Rev Mar Sci 5:289–320
David BO (2015) Tools for drafting, counting and trapping invasive fish. In: Collier KJ, Grainger N (eds) New Zealand invasive fish management handbook. LERNZ (University of Waikato) and Department of Conservation, Hamilton, New Zealand, pp 60–66
David BO, Tonkin JD, Tapeti KWT, Hokianga HT (2014) Learning the ropes: mussel spat ropes improve fish and shrimp passage through culverts. J Appl Ecol 51:214–223
de Winton M (2014) Serpentine Lakes 2014 Macrophyte Assessment. NIWA Client Report HAM2014-054 prepared for the Department of Conservation. National Institute of Water and Atmospheric Research, Hamilton, New Zealand
de Winton MD, Taumoepeau A, Wells R (2006) Submerged vegetation in the Serpentine Lakes 2005/06: current condition and weed threat. NIWAClient Report HAM2006-045 prepared for the Department of Conservation. National Institute of Water and Atmospheric Research, Hamilton, New Zealand
Dreslik MJ (1999) Dietary notes on the red-eared slider (Trachemys scripta) and River Cooter (Pseudemys concinna) from southern Illinois. Trans Ill State Acad 92:233–241
Duggan IC (2002) First record of a wild population of the tropical snail Melanoides tuberculata in New Zealand natural waters. N Z J Mar Freshw Res 36:825–829
Duggan IC (2010) The freshwater aquarium trade as a vector for incidental invertebrate fauna. Biol Invasions 12:3757–3770
Duggan IC, Eastwood KR (2012) Detection and distribution of Craspedacusta sowerbii: observations of medusa are not enough. Aquat Invasions 7:271–275
Duggan IC, Pullan SG (2017) Do freshwater aquaculture facilities provide an invasion risk for zooplankton hitchhikers? Biol Invasions 19:307–314
Duggan IC, Green JD, Burger DF (2006) First New Zealand records of three non-indigenous zooplankton species: Skistodiaptomus pallidus, Sinodiaptomus valkanovi and Daphnia dentifera. N Z J Mar Freshw Res 40:561–569
Duggan IC, Robinson KV, Burns CW, Banks JC, Hogg ID (2012) Identifying invertebrate invasions using morphological and molecular analyses: North American Daphnia ‘pulex’ in New Zealand fresh waters. Aquat Invasions 7:585–590
Duggan IC, Neale MW, Robinson KV, Verburg P, Watson NTN (2014) Skistodiaptomus pallidus (Copepoda: Diaptomidae) establishment in New Zealand natural lakes, and its effects on zooplankton community composition. Aquat Invasions 9:195–202
Duggan IC, Wood SA, West DW (2015) Brown trout (Salmo trutta) removal by rotenone alters zooplankton and phytoplankton community composition in a shallow mesotrophic reservoir. N Z J Mar Freshw Res 49:356–365
Galil BS (2002) Control and eradication of invasive aquatic invertebrates. In: Gherardi F, Corti C, Gualtieri M (eds) Biodiversity conservation and habitat management. The Encyclopedia of life support systems. EOLSS/UNESCO, Oxford, pp 1–14
Genoni P, Fazzone A (2008) Barbronia weberi (R. Blanchard, 1897) (Hirudinea: Salifidae), an Asian leech species new to Italy. Aquat Invasions 3:77–79
Georgieva G, Varadinova E, Uzunov Y (2012) Distribution of non-indigenous tubificid worm Branchiura sowerbyi (Beddard, 1892) in Bulgaria. J BioSci Biotechnol 2012:105–113
Glasby CJ, Read GB, Lee KE, Blakemore RJ, Fraser PM, Pinder AM, Erséus C, Moser WE, Burreson EM, Govedich FR, Davies RW, Dawson EW (2007) Phylum Annelida: bristleworms, earthworms, leeches. In: Gordon DP (ed) New Zealand inventory of biodiversity, Kingdom Animalia: Radiata, Lophotrochozoa, and Deuterostomia, vol 1. Canterbury University Press, Christchurch, New Zealand, pp 312–358
Goldschmidt T (1996) Darwin’s dreampond: drama in Lake Victoria. MIT Press, Cambridge, UK
Grainger N (2015) Control of invasive fish incursions in the northern South Island. In: Collier KJ, Grainger N (eds) New Zealand invasive fish management handbook. LERNZ (University of Waikato) and Department of Conservation, Hamilton, New Zealand, pp 95–99
Gumbley J, Daniel A (2015) Role of screens and barriers for management of invasive fish. In: Collier KJ, Grainger N (eds) New Zealand invasive fish management handbook. LERNZ (University of Waikato) and Department of Conservation, Hamilton, New Zealand, pp 67–71
Hayden BJ, Whyte CF (2003) Invasive species management in New Zealand. In: Ruiz GM, Carlton JT (eds) Invasive species: vectors and management strategies. Island Press, Washington, DC, pp 270–291
Hicks BJ, Ling N (2015) Common carp as an invasive species. In: Pietsch C, Hirsch P (eds) Biology and ecology of carp. Science Publishers, Boca Raton, pp 244–281
Hicks BJ, Reynolds GB, Jamieson PM, Laboyrie JL (2001) Fish populations of Lake Ngaroto, Waikato, and fish passage at the outlet weir. CBER (Centre for Biodiversity and Ecology Research), Contract Report 14, prepared for the Waipa District Council. University of Waikato, Hamilton, New Zealand
Hicks BJ, Ling N, Wilson BJ (2010) Introduced fish. In: Collier KJ, Hamilton DP, Vant WN, Howard-Williams C (eds) Waters of the Waikato: ecology of New Zealand’s longest river. University of Waikato and Environment Waikato, Hamilton, New Zealand, pp 209–228
Hicks BJ, Daniel A, Ling N, Morgan D, Gauthier S (2015) Costs and effectiveness of different methods for capturing invasive fish. In: Collier KJ, Grainger N (eds) New Zealand invasive fish management handbook. LERNZ (University of Waikato) and Department of Conservation, Hamilton, pp 123–132
Holder P (1999) The mosquitoes of New Zealand and their animal disease significance. Surveillance 26:12–15
Jerde CL, Mahon AR, Chadderton WL, Lodge DM (2011) “Sight-unseen” detection of rare aquatic species using environmental DNA. Conserv Lett 4:150–157
Johnson PTJ, Olden JD, Vander Zanden MJ (2008) Dam invaders: impoundments facilitate biological invasions into freshwaters. Front Ecol Environ 6:357–363
Jolly VH (1955) A review of the freshwater Cladocera of New Zealand. MSc thesis, University of Otago, Dunedin, New Zealand
Jonas JL, Claramunt RM, Fitzsimons JD, Marsden JE, Ellrott BJ (2005) Estimates of egg deposition and effects of lake trout (Salvelinus namaycush) egg predators in three regions of the Great Lakes. Can J Fish Aquat Sci 62:2254–2264
Koenders A, Martens K, Halse S, Schön I (2012) Cryptic species of the Eucypris virens species complex (Ostracoda, Crustacea) from Europe have invaded Western Australia. Biol Invasions 14:2187–2201
Kolar CS, Lodge DM (2002) Ecological predictions and risk assessments for alien species. Science 298:1233–1236
Kolmakov VI, Gladyshev MI (2003) Growth and potential photosynthesis of cyanobacteria are stimulated by viable gut passage in crucian carp. Aquat Ecol 37:237–242
Lake MD, Hicks BJ, Wells RDS, Dugdale TM (2002) Consumption of submerged aquatic macrophytes by rudd (Scardinius erythrophthalmus L.) in New Zealand. Hydrobiologia 47:13–22
Leung B, Lodge DM, Finnoff D, Shogren JF, Lewis MA, Lamberti G (2002) An ounce of prevention or a pound of cure: bioeconomic risk analysis of invasive species. Proc R Soc Lond B 269:2407–2413
Ling N (2004) Gambusia in New Zealand: really bad or just misunderstood. N Z J Mar Freshw Res 38:473–480
Lloyd LN, Arthington AH, Milton DA (1986) The mosquitofish: a valuable mosquito control agent or a pest? In: Kitching RL (ed) The ecology of exotic animals and plants: some Australian case histories. Wiley, Brisbane, Australia
Ludgate BG, Closs GP (2003) Responses of fish communities to sustained removals of perch (Perca fluviatilis). Science for Conservation 210. Department of Conservation, Wellington, New Zealand
Makino W, Knox MA, Duggan IC (2010) Invasion, genetic variation and species identity of the calanoid copepod Sinodiaptomus valkanovi. Freshw Biol 55:375–386
Marshall JW (1978) The first records of Stylodrilus heringianus (Oligochaeta: Lumhriculidae) from the Southern Hemisphere. N Z J Zool 5:781–782
Marshall JW, Winterbourn MJ (1978) An ecological study of a small New Zealand stream with particular reference to the Oligochaeta. Hydrobiologia 65:199–208
Mason J (1976) Studies on the freshwater and terrestrial leeches of New Zealand. 2. Orders Gnathobdelliformes and Pharyngobdelliformes. J R Soc N Z 6:255–276
McDowall RM (2004) Shoot first, and then ask questions: a look at aquarium fish imports and invasiveness in New Zealand. N Z J Mar Freshw Res 38:503–510
Morgan DL, Beatty SJ (2007) Feral goldfish (Carassius auratus) in Western Australia: a case study from Vasse River. J R Soc West Aust 90:151–156
Morgan DK, Kumar H, Ling N, Hicks BJ (2013) The palatability of flavoured novel floating pellets made with brewer’s spent grain to captive carp. N Z J Zool 40:170–174
Morgan DK, Verbeek CJR, Rosentrater KA, Hicks BJ (2014) Toxicity effects of oral gavage of aqueous solutions of cube-root powder and sodium nitrite in common carp. Wildl Res 40:647–653
O’Keefe DM, Miller SR (2011) 2009 Michigan charter fishing study. Michigan Sea Grant Extension report MICHU-11-200. Ann Arbor, MI
Pagnucco KS, Maynard GA, Fera S, Yan ND, Nalepa TF, Ricciardi A (2015) The future of species invasions in the Great Lakes-St. Lawrence River basin. J Great Lakes Res 41(Supp 1):96–107
Parkes SM, Duggan IC (2012) Are zooplankton invasions in constructed waters facilitated by simple communities? Divers Distrib 18:1199–1210
Peay S, Hiley PD, Collen P, Martin I (2006) Biocide treatment of ponds in Scotland to eradicate signal crayfish. Bull Fr Pêche Piscic 380–381:1363–1379
Pham L, West D, Closs P (2013) Reintroduction of a native galaxiid (Galaxias fasciatus) following piscicide treatment in two streams: response and recovery of the fish population. Ecol Freshw Fish 22:361–373
Pingram MA (2005) Ecology of freshwater fish in the littoral zone of Lake Waikere, Kai Iwi Lakes, Northland, New Zealand: for the conservation of the dune lakes galaxiid (Galaxias sp.). MSc Thesis, University of Otago, Dunedin, New Zealand
Price J, Gumbley J (2015) Effects of rudd control in Rotopiko (Serpentine) lakes. In: Collier KJ, Grainger N (eds) New Zealand invasive fish management handbook. LERNZ (University of Waikato) and Department of Conservation, Hamilton, pp 100–104
Pullan NB, Climo FM, Mansfield CB (1972) Studies on the distribution and ecology of the family Lymnaeidae (Mollusca: Gastropoda) in New Zealand. J R Soc NZ 2:393–405
Pyke GH (2005) A review of the biology of Gambusia affinis and G. holbrooki. Rev Fish Biol Fish 15:339–365
Pyke GH, White AW (2001) A review of the biology of the green and golden bell frog Litoria aurea. Aust Zool 31:563–598
Ravera O, Jamet JL (1991) The diet of the rudd (Scardinius erythrophthalmus, L.) in relation to the possible consequences for the removal of this species from an eutrophic lake. Arch Hydrobiol 123:99–109
Ricciardi A (2006) Patterns of invasion of the Laurentian Great Lakes in relation to changes in vector activity. Divers Distrib 12:425–433
Rowe RJ (1987) The dragonflies of New Zealand. Auckland University Press, Auckland
Rowe DK (1998) Management trials to restore dwarf inanga show mosquitofish threat to native fish. Water Atmos 6:10–12
Rowe DK (2003) Balancing native fish diversity, exotic fish impacts and angling in New Zealand, North Island dune lakes. In: Beumer J, Grant A, Smith DC (eds) Aquatic protected areas – what works best and how do we know. Proceedings of the world congress on aquatic protected areas, Cairns, August 2002, Australian Society for Fish Biology, pp 96–102.
Rowe DK (2004) Potential effects of tench (Tinca tinca) in New Zealand freshwater ecosystems. NIWA Client Report HAM2004-005, National Institute of Water and Atmospheric Research, Hamilton, New Zealand
Rowe DK (2007) Exotic fish introductions and the decline of water clarity in small North Island, New Zealand lakes: a multi-species problem. Hydrobiologia 583:345–358
Rowe DK, Champion PD (1994) Biomanipulation of plants and fish to restore Lake Parkinson: a case study and its implications. In: Collier KJ (ed) Restoration of aquatic habitats. Selected papers from the second day of the New Zealand Limnological Society 1993 annual conference. Department of Conservation, Wellington
Rowe DK, Graynoth E (2000) Fish populations in New Zealand lakes. NIWA Client Report MfE00223 prepared for Ministry for the Environment. National Institute of Water and Atmospheric Research, Hamilton, New Zealand
Rowe DK, Smith JP (2001) The role of exotic fish in the loss of macrophytes and increased turbidity of Lake Wainamu, Auckland. Auckland Regional Council Technical Report 2008/003, Auckland, New Zealand
Rowe DK, Smith JP, Baker C (2007) Agonistic interactions between Gambusia affinis and Galaxias maculatus: implications for whitebait fisheries in New Zealand rivers. J Appl Ichthyol 23:668–674
Sandodden R, Johnsen SI (2010) Eradication of introduced signal crayfish Pasifastacus leniusculus using the pharmaceutical BETAMAX VET®. Aquat Invasions 5:75–81
Stuart IG, Williams A, McKenzie J, Holt T (2006) Managing a migratory pest species: a selective trap for common carp. N Am J Fish Manage 26:888–893
Talbot JM, Ward JC (1987) Macroinvertebrates associated with aquatic macrophytes in Lake Alexandria, New Zealand. N Z J Mar Freshw Res 21:199–213
Taylor CM, Duggan IC (2012) Can biotic resistance be utilized to reduce establishment rates of non-indigenous species in constructed waters? Biol Invasions 14:307–322
Tempero GW, Ling N, Hicks BJ, Osborne MW (2006) Age composition, growth, and reproduction of koi carp (Cyprinus carpio L.) in the lower Waikato, New Zealand. N Z J Mar Freshw Res 40:571–583
Tempero G, Daniel AJ, Ling N (2015) Removal and exclusion of koi carp from Lake Ohinewai. In: Collier KJ, Grainger N (eds) New Zealand invasive fish management handbook. LERNZ (University of Waikato) and Department of Conservation, Hamilton, pp 90–94
Thwaites LA, Smith BB, Decelis M, Fleer D, Conallin A (2010) A novel push trap element to manage carp (Cyprinus carpio L.): a laboratory trial. Mar Freshw Res 61:42–48
Verrill D, Berry C, Donovan V, Charles R (1995) Effectiveness of an electrical barrier and lake drawdown for reducing common carp and bigmouth buffalo abundances. N Am J Fish Manage 15:137–141
Vilizzi L, Thwaite LA, Smith BB, Nicol JM, Madden CP (2014) Ecological effects of common carp (Cyprinus carpio) in a semi-arid floodplain wetland. Mar Freshw Res 65:802–817
Weber MJ, Brown ML (2009) Effects of common carp on aquatic ecosystems 80 years after “Carp as a dominant”: ecological insights for fisheries management. Rev Fish Sci 17:524–537
Weber MJ, Brown ML (2015) Biomass-dependent effects of age-0 common carp on aquatic ecosystems. Hydrobiologia 742:71–80
Wells R (1999) Are rudd a threat to water plants? NIWA Water Atmos 7(4):11–13
Wendt-Rasch L, Friberg-Jensen U, Woin P, Christoffersen K (2003) Effects of the pyrethroid insecticide cypermethrin on a freshwater community studied under field conditions. II. Direct and indirect effects on the species composition. Aquat Toxicol 63:373–389
West D (2015) Piscicides and drainage for management of invasive fish. In: Collier KJ, Grainger N (eds) New Zealand invasive fish management handbook. LERNZ (University of Waikato) and Department of Conservation, Hamilton, pp 54–59
Williamson M (1996) Biological invasions. Chapman & Hall, London
Winterbourn MJ (1973) A guide to the freshwater Mollusca of New Zealand. Tuatara 20:141–158
Winters B (2012) That pommie bastard. Ocean Books, Mount Maunganui, New Zealand
Wise KAJ (1989) The family Gyrinidae (Hexapoda: Coleoptera) in New Zealand: with a description of a northern population. Rec Auckl Inst Museum 26:83–102
Wise KAJ (1998) A species of the family Sisyridae (Insecta: Neuroptera) in New Zealand. N Z Entomol 21:11–16
Wood SA, Smith KF, Banks JC, Tremblay LA, Rhodes L, Mountfort D, Cary SC, Pochon X (2013) Molecular genetic tools for environmental monitoring of New Zealand’s aquatic habitats, past, present and the future. NZ J Mar Freshw Res 47:90–119
Wu N, Daniel A, Tempero G (2013) Fish biomass and gonad development in the Rotopiko (Serpentine) lakes. Environmental Research Institute Report No. 20 prepared for Department of Conservation. University of Waikato, Hamilton, New Zealand
Young EC (2010) The water boatmen and backswimmers of New Zealand. Hemiptera: Corixidae and Notonectidae. A research guide. Witherlea Press, Nelson, New Zealand
Zielinskia DP, Vollera VR, Svendsenb JC, Hondzoa M, Mensingerc AF, Sorensen P (2014) Laboratory experiments demonstrate that bubble curtains can effectively inhibit movement of common carp. Ecol Eng 67:95–103
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2018 Springer International Publishing AG, part of Springer Nature
About this chapter

Cite this chapter
Duggan, I.C., Collier, K. (2018). Management of Non-indigenous Lacustrine Animals. In: Hamilton, D., Collier, K., Quinn, J., Howard-Williams, C. (eds) Lake Restoration Handbook. Springer, Cham. https://doi.org/10.1007/978-3-319-93043-5_9
Download citation
DOI: https://doi.org/10.1007/978-3-319-93043-5_9
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-93042-8
Online ISBN: 978-3-319-93043-5
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)