
Abstract
This chapter describes the ecology of foliar endophytes of the Acadian Forest that dominates Canada’s Maritime Provinces extending into Eastern Quebec and Maine. Recent evidence has illuminated the ‘foraging ascomycete’ life habit of fungi that can be endophytic in conifer needles. These fungi can occupy several eco-niches other than the needles including as saprophytes in aquatic or terrestrial environments or as endophytes of understory species. Structurally diverse secondary antifungal and antiinsectan metabolites appear to mediate the exchange between plant and fungus. The plant provides nutrients and shelter, the fungus increases plant fitness by contributing to tolerance to herbivorous insects or needle pathogens. This work is enabled by the advent of affordable sequencing capability, a dedication to fieldwork and alpha taxonomy, and directed investigations of the metabolites produced by these interesting fungi.
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others


Keywords
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
1 Introduction
Endophytes are an artificial assemblage of phylogenetically diverse fungi that can asymptomatically infect and inhabit healthy plant tissue. Interest in fungal endophytes of plants was propelled by the discovery of the clavicipitaceous (Clavicipitaceae, Hypocreales) endophytes of cool-season grasses, which produce toxic alkaloids associated with livestock toxicoses and resistance to insect pests (Clay 1988; Siegel et al. 1990). These fungi systemically infect their hosts and may increase growth and seed production and confer tolerance to the host from a variety of abiotic and biotic stresses such as drought, heat, waterlogging, heavy metal toxicity, herbivory and fungal pathogens (Kuldau and Bacon 2008; Monnet et al. 2001; Rice et al. 1990; Song et al. 2015; Xia et al. 2015). Because of the economic importance of the impact on livestock and their potential use as inoculants of turfgrass against insects, clavicipitaceous endophytes received much attention and consequently shaped the endophyte research paradigm.
However, the clavicipitaceous endophyte model is not applicable to foliar endophytes of woody plants (Rodriguez et al. 2009). Instead of a phylogenetically-coherent group of taxa which display host-specific, systemic, and frequently vertically-transmitted life strategies, foliar endophytes of woody plants are phylogenetically very diverse, frequently exhibit broad host preferences, can be euryoecious, and are horizontally-transmitted. A significant knowledge gap in the general biology and taxonomy of woody plant endophytes exists. This is exacerbated by the immense phylogenetic diversity and lack of research investigating endophyte life histories.
Collectively, endophytes interact with their hosts along the endosymbiont-pathogen continuum, depending on a balance of antagonisms involving the immediate phenotype status of the fungus and host (Schulz and Boyle 2005). In trees, these host-endophyte interactions are often studied in the context of mutualism, for example investigating possible benefits bestowed upon the host by endophyte infection, such as decreased herbivory, disease resistance, drought tolerance, and plant growth promotion (e.g.: Arnold et al. 2003; Hanada et al. 2010; Khan et al. 2016; Rocha et al. 2011). Many of these mutualistic associations are explained by the production of endophytic natural products in planta. Endophytes are widely considered as an unexplored treasure trove of new biologically active compounds (Aly et al. 2010; Strobel and Daisy 2003). During the portion of their lifecycle associated with a host, there is constant metabolic interaction between the endophyte and plant. The endophyte-plant metabolic equilibrium must balance interactions between the fungal species, host, epiphytes, and plant pathogens. This type of constant interaction fosters an environment selective for endophyte genotypes capable of producing extensive secondary metabolite mixtures fulfilling an ecological function. The lack of knowledge regarding the taxonomy of foliar endophytes of woody plants provides a rare opportunity to prospect for new chemistries in understudied taxa.
Foliar endophyte communities of the Acadian forest and their natural product diversity have been the subject of extensive investigation over the past three decades. Last reviewed by Sumarah and Miller (2009) and Miller (2011), we update here the progress made studying the natural products, ecology and taxonomy of some abundant, phylogenetically diverse conifer endophytes. Consideration is given to the reintroduction of selected endophyte genotypes into seedlings to increase forest resiliency during reforestation efforts as part of integrated forest pest management strategies.
2 Challenges Detecting and Identifying Endophytes
Culture-dependent endophyte studies involve rigorous sterilization of plant tissue surfaces, for example by serial passage through ethanol and sodium hypochlorite, to kill non-endophytic fungi (e.g. epiphytes or viable propagules on the leaf surface) and other organisms. The surface-sterilized plant tissue is then cut into segments, placed on suitable agar media and incubated. Mycelia emerging from plant tissue segments are excised and subcultured to generate axenic cultures. Capturing more biodiversity from plant tissue can involve the addition of other methods such as enzymatic digestion of plant tissue, use of non-standard media, or higher throughput protocols (Greenfield et al. 2015; Prior et al. 2014). Culture-based studies generate viable strains, which enable further work such as multilocus phylogenetic analyses, omics studies, secondary metabolite profiling, inoculation experiments, and the ability to describe novel species represented by acceptable holotypes and ex-type living strains.
Traditional endophyte studies rely upon morphological characters to distinguish, group, and identify endophyte strains. Identification to genus and species is feasible when fungi produce taxonomically-informative morphological characters such as conidiomata or ascomata in vitro. Inducing sporulation in sterile cultures can be a laborious process with varying success. This requires prolonged incubation under various culture conditions, with treatments including different artificial and natural media, light regimes, incubation temperatures, gradual desiccation, mechanical stimulation, flotation in still or aerated water, and the addition of natural substrates such as sterilized plant leaves (Knapp et al. 2015; Sokolski et al. 2006a; Tanney et al. 2016a). Recalcitrant sterile cultures are generally unidentifiable because hyphae typically lack morphological characters allowing for precise taxonomic designation. Hence unidentified endophytes are often designated simply by morphotype descriptions (e.g.: “sterile white” and “sterile pigmented” in Johnson and Whitney 1989). The absence of identified strains, especially for strains not accessioned in culture collections, can result in the inability to make meaningful conclusions or comparisons between past and modern studies. Identification relying solely on morphology and other phenotypic characters can be problematic due to cryptic speciation, convergent evolution, and hybridization (Raja et al. 2017).
Previous attempts to identify particular sterile endophytes included, for example, the use of a specific absorbed immunoserum with on-section immunogold labeling to identify Lophodermium piceae hyphae in symptomless Picea abies needles by immunoelectron microscopy (Suske and Acker 1989). However, the most important development in the categorization and identification of sterile endophyte strains was the use of molecular DNA tools for fingerprinting and barcoding (Haemmerli et al. 1992; Leuchtmann and Clay 1993; Wilson et al. 1994). Most contemporary endophyte studies use the nuclear ribosomal internal transcribed spacer (ITS) barcode to identify or group endophyte strains. ITS, the official fungal barcode, is readily amplified using standard primers, well-represented in public DNA sequence databases, such as GenBank, and provides good species resolution, although insufficient interspecific variation is known among some taxa, requiring the further use of secondary barcodes (Badotti et al. 2017; Samson et al. 2014; Schoch et al. 2012; Stielow et al. 2015). The more conserved large-subunit rRNA (LSU) gene is frequently used in conjunction with ITS. Similar to ITS, LSU is also efficiently amplified using standard primers (e.g. LR0R and LR5; Vilgalys and Hester 1990) and is well-represented by abundant reference sequences (Stielow et al. 2015). LSU provides good generic or higher rank taxonomic classification and may be readily aligned across distantly related taxa, which is useful for estimating phylogenies of diverse communities, placing new fungal lineages, and analyzing basal lineages (Liu et al. 2012; Porter and Golding 2012). A good description of a typical workflow for identifying cultures using molecular tools and morphological characters is presented by Raja et al. (2017).
Next generation sequencing (NGS) technology now enables the amplification of endophyte fungal DNA directly from plant tissue, bypassing the labor and selective biases associated with culture-dependent methods and capturing a large number of reads at a relatively low cost. However, NGS studies may also have shortfalls such as amplification biases and failure to discriminate between DNA from living or dead cells (Tedersoo and Lindahl 2016; van Dijk et al. 2014).
Despite these advances, an analogous problem that plagued pre-molecular culture-based studies still confronts both culture-dependent and culture-independent studies relying on DNA sequence-based identification: the precise identification of endophyte strains or sequences lacking taxonomically-informative data. When a fungus belongs to a species, genus, or even family that is unrepresented by accurately identified reference sequences, for example in an accessible DNA sequence repository such as NCBI GenBank (https://www.ncbi.nlm.nih.gov/genbank/) or UNITE (Abarenkov et al. 2010), its identification using DNA sequences may be irresolvable. The inability to identify endophyte sequences yields results similar to that of traditional studies, however instead of “sterile white” and “sterile pigmented” there are “fungal endophyte sp.”, “Ascomycota sp.”, and “Leotiomycetes sp.”. Accordingly, many endophyte studies report endophyte taxa that are often unidentified to species or genus (Arnold and Lutzoni 2007; Huang et al. 2016; U’Ren et al. 2012).
However, unidentified endophyte DNA sequences accessioned in GenBank and other sequence repositories can be a valuable resource. Comparison of unidentified sequences connected with biogeographic data can provide insight into endophyte ecology despite the absence of a binomial name. These sequences can also be clustered into molecular operational taxonomic units (MOTU) and presented as putative species hypotheses (Abarenkov et al. 2010). The ability to cluster MOTUs into species hypotheses with associated aggregate data has led some mycologists to advocate for a sequence-based taxonomic system allowing for the description of fungi based solely on ENAS data (De Beer et al. 2016; Hawksworth et al. 2016; Hibbett and Taylor 2013; Hibbett et al. 2016). This movement is partly justified by the growing number of unidentified sequences that certainly represent a portion of the currently unknown fungal biodiversity. Names provide consistent identifiers that facilitate the communication of taxonomic concepts; for example, the name Aspergillus fumigatus provides a wealth of associated information compared with a unique digital identifier from GenBank. Taxonomic instability can also present issues in consistent identification of sequences over time as the quality of sequences databases changes, e.g. correction of misidentifications and identification of previously unidentified sequences (Arnold and Lutzoni 2007).
Although promising, the premature adoption of a taxonomic system accepting ENAS-based species descriptions is likely to create a legacy fraught with doubtful or redundant names. For example, named-but-unsequenced species will be re-described solely because of their absence from sequence databases. MOTU-delineation thresholds used in automated bioinformatics pipelines are variable across studies and dependent on the taxa being investigated. Sequence similarity thresholds used for MOTU delineation (e.g. 95–97%) attempt to compensate for intraspecific sequence variation and possible sequencing errors (Jumpponen and Jones 2009; Lindner et al. 2013; Vrålstad 2011). The result can be the inflation or underestimation of species diversity based on ITS MOTUs, especially when generic similarity thresholds values are applied to a phylogenetically diverse range of taxa with different expressions of ITS variation. In a culture-based study using multilocus phylogenetic analyses, Gazis et al. (2011) provided a demonstration of the shortfalls of species delineation using only ITS sequences for species complexes and advocated the use of more than one gene for species delineation or, if not feasible, the use of a higher ITS similarity threshold (99–100%) for a more accurate diversity estimate. Conversely, intra-isolate and even intrasporal variation of ITS and LSU can be high in Glomeromycota species, thus more stringent delineation thresholds can result in an overestimation of species diversity (Thiéry et al. 2012, 2016). The inability of ITS to discriminate species complexes or distinct species sharing similar ITS sequences is established for some well-studied genera such as Diaporthe and Penicillium, leading to the use of secondary markers (Udayanga et al. 2014; Visagie et al. 2014). A prudent approach mindful of the taxa being studied, rather than an automated taxonomic pipeline, should be adopted when describing novel taxa, including endophytes, from ENAS.
Endophyte species identification is therefore not constrained by the ability to generate strains or sequences, but by: (1) the lack of reference sequences; (2) the current approach to this taxonomic problem; and (3) a dearth of taxonomists. The growing dependence on sequence-based identification of fungal specimens and cultures and the shift towards biodiversity studies relying solely on ENAS highlights the need for accurately-identified reference sequences connected with voucher specimens and, when feasible, strains accessioned in public culture collections. If sterile endophyte strains or endophyte sequences cannot be identified by ITS barcodes and other gene sequences because of a lack of reference data, then the clear solution is to expand upon the available reference sequences—and knowing where to look.
3 Beyond the Leaf: The Foraging Ascomycete Hypothesis
Xylariaecous (Xylariaceae, Xylariales) endophytes are detected in varying abundance in most endophyte studies involving woody plants, regardless of geographic location or host (e.g. Davis et al. 2003; Dreyfuss and Petrini 1984; Okane et al. 2008, 2013; Petrini and Petrini 1985; Petrini et al. 1995). This ubiquity and apparent host-neutrality has led to much interest and speculation into the role of endophytism in fungal ecology. In this context, Carroll (1999) posited the foraging ascomycete hypothesis, suggesting that for some saprotrophs, endophytism is a secondary life-history strategy facilitating persistence and dispersal in the absence of substrates suitable for reproduction (i.e., a primary host). This latent saprotroph strategy may involve a fungus persisting as a quiescent thallus for some time until the host tissue senesces or dies, at which point the expansive colonization of the fungus into the host substrate ensues. The fungus may cycle through foliage as an endophyte for some time until it comes into contact with a suitable primary host and sexual reproduction or another facet of its life history is permitted. A fungus capable of endophytically infecting host foliage is afforded a refuge buffering it from UV radiation, drought, starvation, competition, mycophagy, and other stresses, while being provided with a source of nutrition and a vehicle for persistence and dispersal. Picea trees, for example, retain needles for several years to over a decade depending on environmental conditions (Hom and Oechel 1983; Okane et al. 2008; Osono et al. 2013; Reich et al. 1996), presenting a refuge amenable for long-term persistence by endophytes. Thomas et al. (2016) demonstrated the direct transmission of endophytes from leaves to woody substrates by means of physical contact. Thus the shedding of foliage may act as a dispersal mechanism for saprotrophic endophytes capable of growing on substrates present in the forest floor. Thomas et al. (2016) also isolated Xylariaceae endophytes from 38 tree species in a tropical cloud forest and connected all five Xylaria endophytes with saprotrophic (stromatal) life stages. Hypoxylon pulicicidum, a pantropical endophyte producing antiinsectan nodulisporic acids, was detected in several independent studies and eventually described following the collection of stromata on decaying wood (Bills et al. 2012). U’Ren et al. (2016) and Okane et al. (2008) also provided evidence connecting saprotrophic and endophytic Xylariaceae species.
Undoubtedly, life histories of foliar endophytes of conifer trees can be entirely restricted within the host foliage, for example Lophodermium piceae (Rhytismataceae, Rhytismatales). Lophodermium piceae is a ubiquitous Picea endophyte that can persist as an endophyte throughout the life of the needle, producing conidiomata (spermatia) and ascomata following needle senescence (Barklund 1987; Livsey and Barklund 1992). Ascospore discharge coincides with vernal shoot and needle flush, when the ascospores are forcibly ejected, adhere to needles by means of a gelatinous or mucoid sheath, and consequently germinate and infect the newly emerging needles (Osorio and Stephan 1991). The fungus then persists as an endophyte until needle senescence either occurs naturally or accelerated by stress.
Converging evidence indicates the foraging ascomycete hypothesis is also applicable to phylogenetically-diverse endophytes outside Xylariales, for example diverse families within Helotiales (Leotiomycetes). Conifer endophytes may produce reproductive structures on specific non-foliar tissues of the same host species or on other hosts such as hardwood trees. For example, Phialocephala scopiformis (Mollisiaceae, Helotiales) is a common foliar endophyte of Picea, also occurring as branch endophyte (Kowalski and Kehr 1995). Tanney et al. (2016a) discovered that the apothecial sexual state of P. scopiformis commonly occurred on decaying Picea wood and fallen branches, and suggested the fungus might systemically infect hosts. Conversely, Phialocephala piceae, also a foliar endophyte of Picea, instead produces its sexual state on decaying hardwood, indicating a broader host range and complex life history (Grünig et al. 2009; Menkis et al. 2004; Tanney et al. 2016a). Tanney et al. (2016a) described the first collection of Phialocephala piceae apothecia in nature, occurring erumpent from decaying Acer saccharum (sugar maple) branches, and hypothesized its presence as a cambium endophyte in the hardwood host. More sampling is required to determine the endophyte host range of P. piceae; i.e.: is it restricted to Picea or capable of infecting diverse hosts?
Some fungi are able to infect a broad range of hosts as endophytes but reproduce on a narrow range of alternate hosts with some narrow preference observed, such as the griseofulvin-producing Xylaria sp. discussed below (Fig. 1), Nemania (Xylariaceae, Xylariales), Pezicula, and wood-decaying basidiomycetes such as Peniophora and Trametes (Chen et al. 2015; Lee et al. 2016a; Martin et al. 2015; Yuan et al. 2011). Many studies have observed the presence of sporulating endophytes outside of their plant host (e.g. Chaverri and Gazis 2011; Koukol et al. 2012; Yuan and Verkley 2015; Okane et al. 2008; Tanney et al. 2016a; Thomas et al. 2016; U’Ren et al. 2016), indicating many endophytes are also capable of functioning as saprotrophs.

Hypothetical life history of a Xylaria sp. exhibiting an endophytic-saprotrophic life cycle with host-switching. The fungus is capable of endophytically infecting a broad range of overstory and understory plants, persisting and cycling within and among host canopies indefinitely. Endophytic infections are transmitted to primary substrates (e.g.: decaying Acer saccharum wood) when host leaves senesce and abscise by means of conidia disseminated by insects, rainsplash, and air currents, or by direct contact of leaves with the primary substrate. Ascomata and conidiomata are produced on the primary substrate and, via airborne ascospores or disseminated conidia, enable saprotrophic colonization of new primary substrates or endophytic infection of host foliage
Emerging evidence of alternating saprotrophic-endophytic life histories of woody plant endophytes supports the idea that, in woody plants, endophytism should also be considered a facultative life history strategy that may occupy all or only part of the fungus’ life, rather than endophytism being strictly an obligate mode of trophism. The foraging ascomycete hypothesis might explain why endophytism is common throughout Dikarya and may also provide a strategy to identify unknown endophytes, one that involves considering the broader ecology of endophytes. The strategy encouraged by Tanney (2016) is straightforward: culturable endophytes that are unidentifiable based on current sequence data or an absence of taxonomically-informative characters are identified by connecting them to identifiable specimens using DNA barcodes (Fig. 2). Specimens of named-but-unsequenced or undescribed species containing taxonomically-informative characters are inferred to represent saprotrophic reproductive states of unidentified endophytes based on hypotheses derived from phylogenetic or taxonomic evidence (Fig. 3). The resulting DNA barcodes generated from identifiable herbarium specimens, ex-type cultures, new field collections, and sporulating cultures are used to connect unidentified endophytes with names. This chapter will provide some examples of both this approach, as inspired by Carroll’s (1999) foraging ascomycete theory, and the synergy of combining chemistry and taxonomy to more effectively target endophytes that produce biologically active natural products.

General approaches to identifying endophytes, using Phialocephala scopiformis as an example. (1) Healthy, asymptomatic host leaves are surface sterilized, dissected, and incubated on an amenable media. Emerging mycelia is excised to yield an axenic endophyte culture. Sporulation is induced in vitro, enabling identification based on taxonomically-informative characters (e.g.: apothecia). Reference sequences (e.g. ITS and LSU) are generated for this species and accessioned in GenBank, allowing for its accurate identification in future studies. (2) The resulting axenic endophyte culture is sterile but rapidly identified by comparison or phylogenetic analysis with available reference sequences. (3) An identifiable, named-but-unsequenced field specimen, putatively representing the reproductive structure of an endophyte, is collected based on phylogenetically-informed inference. Sequences are generated directly from the fruiting body or from an axenic culture generated from ascospores, conidia, etc. Named sequences are accessioned into GenBank and can be used to identify this species when detected in future studies, for example from ENAS. (4) Fungal endophyte DNA is extracted and sequenced directly from host tissue. Phialocephala scopiformis is identified from the host by the availability of an authenticated reference sequence

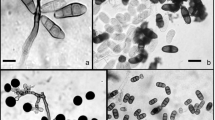
Examples of diverse Ascomycota fruiting bodies connected with endophytic life histories. A. Penicillium section Thysanophora (Trichocomaceae, Eurotiales) conidiophores erumpent through Picea rubens needle stomata. B. Rhizosphaera sp. (Dothideaceae, Dothidiales) conidiomata erumpent through needle stomata, with stomatal wax plugs attached to conidiomata. C. Hyphodiscus sp. (Hyaloscyphaceae, Helotiales) apothecia occurring on living Picea rubens twigs. D. Xylaria sp. (Xylariaceae, Xylariales) stromata on decaying, partially buried Acer saccharum branch. E. Phialocephala sp. (Mollisiaceae, Helotiales) apothecia on decaying hardwood. F. Micraspis acicola (Tympanidaceae, Helotiales) conidioma on dead needle still attached to living twig
4 Acadian Forest: A Case Study
The Acadian forest is the dominant forest region in the Canadian Maritime Provinces, into Eastern Quebec as well as Maine (USA) and including elements of the neighbouring Eastern Deciduous Forest and Boreal Forest Regions. Late-successional forest types are composed of the defining species Picea rubens (red spruce) and other long-lived, shade tolerant species including Acer saccharum (sugar maple), Betula alleghaniensis (yellow birch), Fagus grandifolia (American beech), Pinus strobus (Eastern white pine), and Tsuga canadensis (Eastern hemlock) (Mosseler et al. 2003a). Natural climax forest dynamics are generally driven by small gap-producing events, such as insect-, disease-, or wind-caused mortality, with stand-replacing disturbances being rare (Lorimer 1977; Wein and Moore 1977). However, European settlement brought intensive deforestation activity and large-scale fire disturbances (Loo and Ives 2003). Additional disruptive human activities include persistent selective harvesting for Picea as well as conversion of mixed-wood forests to Abies balsamea (balsam fir)-dominated conifer forests after spruce budworm epidemics and fires (Blais 1983). In response to the spruce budworm epidemic peaking in 1975 forest managers planted white and red spruce. Over the past 70 years, the percentage of softwood in the Acadian forest has not materially changed (Amos-Binks et al. 2010; Etheridge et al. 2005, 2006).
The Eastern spruce budworm (Choristoneura fumiferana) is one of the most destructive forest pests in Eastern Canada, capable of causing substantial decline in vigour and increased mortality in Abies balsamea and Picea species. The 1970s epidemic in Eastern North America resulted in substantial ecological disturbance and economic loss. Budworm outbreaks cause a significant loss of timber volume, an increase in forest fires, and substantial CO2 emissions (Dymond et al. 2010; Fleming et al. 2002). Budworm outbreaks have occurred at periodic intervals of about 35 years (Royama 1984). Forty years after the peak of the last epidemic, in Quebec, almost 3.4 million hectares were moderately to severely defoliated by the budworm in 2014. This is roughly equal to the total number of hectares defoliated in the province from 1987–2007 (National Forestry Database 2017). Chang et al. (2012) estimate that from 2012–2041, uncontrolled moderate budworm outbreaks could cost the New Brunswick economy CDN$3.3 billion and severe budworm outbreaks $4.7 billion.
The driving factors initiating budworm outbreak cycles have not been fully elucidated, although epidemiological studies contradict the hypothesis of one epicenter spreading across the region and suggest that populations across the landscape cycle in unison (Royama 1984; Royama et al. 2005). Royama (1984) considered this basic cycle was determined by various density-dependent mortality factors during the third to sixth larval instars: parasitism, predation, food shortage, weather, and the “fifth agent”. This last undefined variable was hypothesized to be a complex of cryptic causes to explain residual mortality. Royama deduced that population oscillation was ultimately driven by parasitism and the fifth agent.
Motivated by Royama’s intriguing “fifth agent” and the importance of the survival of feeding larvae in driving regional outbreaks, research was initiated under the hypothesis that this unknown variable consisted of toxigenic endophytes (Miller 1986, 2011). This has become a long-term collaborative research effort resulting in the discovery of antiinsectan metabolites produced by Picea endophytes, which may remediate the negative effects of Eastern spruce budworm through the inoculation and planting of endophyte-enhanced seedlings (Miller 2011). This research program is also screening Pinus endophytes for antifungal secondary metabolites to provide tolerance against white pine blister rust (Chronartium ribicola), an invasive disease which continues to decimate white pine in North America (Et-Touil et al. 1999; McMullin et al. 2015; Sumarah et al. 2015; Richardson et al. 2014, 2015).
Seedlings produced in greenhouses are typically void of endophytes (Miller et al. 2009). A primary objective of this program is to identify horizontally-transmitted toxigenic conifer endophyte genotypes to reintroduce stand diversity during reforestation efforts. An ongoing challenge remains developing knowledge regarding the biology and ecology of conifer endophyte collected from the Acadian forest (Tanney 2016). From a natural product discovery perspective, this type of information aids with the dereplication of isolates enabling new chemistries to be efficiently discovered. Akin to the challenges identifying endophytes described above, the natural product literature is full of errors with reports of metabolites produced by unidentified isolates, without deposited voucher specimens, lacking molecular data, with dubious identifications. Credible fungal natural product discovery is reliant upon proper taxonomic identification (Raja et al. 2017). Our approach of combining natural products chemistry with molecular methods, classical taxonomy, new field collections, and herbarium specimens is being implemented to provide more insight into the taxonomy and ecology of conifer endophytes. Examples of these endophytes producing ecologically important natural products collected from the Acadian forest are presented below, including relevant taxonomic, biological, and ecological discussions.
4.1 Toxigenic Endophytes of Picea
The identification of Picea needles harbouring endophytes capable of producing toxins affecting the growth of C. fumiferana pioneered subsequent surveys searching for diverse toxin-producing endophytes. Despite recent efforts to identify endophytic species of interest, some endophyte strains remain unidentified, although with improved taxonomic resolution, or represent undescribed species as part of ongoing taxonomic work. This was the case for two Picea rubens endophytes with culture filtrate extracts toxic to spruce budworm larvae in dietary bioassays. From undescribed Phaeomoniella sp. (Phaeomoniellaceae, Phaeomoniellales) CBS 121944, the sesquiterpenes 3-epiisopetasol and 15-hydroxy-3-epiisopetasol were isolated as the major constituents of the antiinsectan extract (Fig. 4; Sumarah et al. 2010). These metabolites are structurally similar to eremophilane sesquiterpenes including PR-toxin, phomenone, and eremofortines produced by several Penicillium species (McMullin et al. 2014; Nielsen et al. 2006). Three additional polyketide metabolites were characterized from Picea rubens endophyte strain CBS 121942, which cannot be confidently placed in any family or order based on rDNA sequences (Fig. 4; Sumarah et al. 2010).

Sesquiterpenes and polyketides metabolites characterized from the undescribed Picea rubens endophytes Phaeomoniella sp. CBS 121944 and unidentified strain CBS 121942
4.1.1 Dwayaangam colodena
Dwayaangam colodena is a common endophyte of Picea mariana and P. rubens in Eastern Canada, which motivated the investigation of its bioactive secondary metabolites (Sumarah et al. 2010). From a collection of P. rubens endophytes (ca. 2000 isolates) from New Brunswick, Nova Scotia, and Maine, 150 isolates were selected based on morphological characteristics and ITS sequence for further study (Sumarah et al. 2010). Sixteen of the selected isolates that grew well in vitro generated culture filtrate extracts toxic to C. fumiferana larvae when incorporated into a synthetic diet. The alkenoic acids cordyanhydrides A and B, and two structurally related precursors, were characterized from D. colodena DAOM 239833 (Fig. 5; Sumarah et al. 2010). These unusual natural products were initially isolated from the insect pathogenic fungus Cordyceps pseudomilitaris (Isaka et al. 2000). From a recent investigation of conifer endophytes of the Acadian forest, the same metabolites were detected from the antifungal extract of a D. colodena strain isolated from healthy P. mariana needles (Green 2016).

Maleic anhydrides produced by Dwayaangam colodena strains collected as endophytes of Picea mariana and P. rubens
Dwayaangam colodena (incertae sedis, Helotiales) was discovered as both a Picea endophyte and an aquatic hyphomycete in boreal and mixed-wood forest streams in Eastern Canada (Sokolski et al. 2006a, b). The distinct stauroform conidia of D. colodena were collected from surface foam in running forest streams and also induced in vitro by placing cultures in a microcosm simulating river foam (aerated sterile distilled water). These observations evoke a complex life history involving endophytism and an aquatic saptrophic and dispersal phase. Some root endophytes of riparian hosts are also aquatic hyphomycetes, a conceivable adaptation because riparian plants may extend part of their roots into streams and river (Selosse et al. 2008; Sati and Belwal 2005; Sridhar and Bärlocher 1992). Hypothetically, D. colodena-infected needles might naturally senesce, abscise, and enter nearby streams. The fungus would then initiate a more aggressive saprotrophic phase, colonizing the needle and producing water-dispersed conidia when exposed to aerated water. Sporulation might even be promoted during spring freshet events when water flow and aeration is increased, coinciding with bud break and needle extension of Picea hosts. The infection process of host foliage by D. colodena is unknown, but may be established by airborne ascospores from an as yet undescribed or unconnected sexual state.
Dwayaangam colodena conidia were also reported from rainwater collected from stands of Picea abies, Pinus sylvestris, and Vaccinium myrtillus in Europe, suggesting a wider geographic and host range and the production of conidia within the canopy (Révay and Gönczöl 2010; Sumarah et al. 2010). Conidia of so-called “terrestrial aquatic hyphomycetes” are frequently reported from arboreal environments, possibly arising from microhabitats such as bark fissures, lodged leaves and debris, phytohelma, or saprotrophic, parasitic, and epiphytic fungi sporulating ephemerally in surface condensation on leaves (Ando 1992; Chauvet et al. 2016; Révay and Gönczöl 2011; Stone et al. 1996). Thus the “terrestrial aquatic hyphomycete” conidia morphology might facilitate dispersal not only in aquatic habitats, but also through the canopy via throughfall and stemflow, along water films on plant surfaces, and by increasing trapping efficiency (Bandoni and Koske 1974; Gönczöl and Révay 2004; Sridhar and Karamchand 2009; Webster 1959).
4.1.2 Mollisiaceae
Mollisiaceae is a large, poorly sampled family comprising at least 1000 names from Belonopsis, Mollisia, Phialocephala, and Tapesia alone. Dark septate endophyte (DSE) Phialocephala species ubiquitous in roots are well-studied and recent evidence shows the prevalence of endophytism in Mollisia, a genus typically considered to be comprised of saprotrophic species (Tanney 2016). The Picea endophyte Phialocephala scopiformis DAOMC 229536 (Mollisiaceae, Helotiales) is the most extensively studied endophyte from the Acadian forest; its anthraquinones rugulosin, skyrin, and emodin represent some of the first insect toxins characterized from conifer endophytes (Fig. 6; Calhoun et al. 1992). Incorporation of P. scopiformis culture filtrate extract or its dominant toxin rugulosin into a synthetic diet inhibited the growth of spruce budworm (C. fumiferana) larvae, hemlock looper (Lambdina fiscellaria), and spruce budmoth (Zeiraphera canadensis) in the low µg/g range (Sumarah et al. 2008a). Greenhouse and nursery studies with three- and four-year-old inoculated seedlings were performed to determine the effect this toxigenic endophyte had on spruce budworm growth. When needle concentrations of rugulosin exceeded the low observed effect level of 0.5 µg g−1 for C. fumiferana, a dose response was observed (Miller et al. 2008). These experiments showed the reduction in growth was associated with effective endophyte toxin concentrations and provided incentive to further explore the biology of P. scopiformis and other Mollisiaceae species.

Anthraquinones rugulosin, skyrin and emodin synthesized by the Picea endophyte Phialocephala scopiformis
Phialocephala scopiformis, originally described as an endophyte from the periderm of living Picea abies branches, is a common foliar endophyte of Picea in Eastern Canada (Kowalski and Kehr 1995; Tanney et al. 2016a). As part of an ongoing investigation to identify unknown endophyte cultures by connecting them with sporulating field specimens, Tanney et al. (2016a) discovered the sexual state of P. scopiformis from fallen decaying Picea branches and wood. Its ascospores arise from typical mollisioid apothecia, which are grayish blue, 0.3–1 mm diam, and occur scattered to caespitose on the host substrate. The asexual state of P. scopiformis consists of dematiaceous penicillate conidiophores, which give rise to dimorphic conidia aggregating in slimy heads. Sumarah et al. (2005) successfully infected foliage of Picea glauca seedlings with Phialocephala scopiformis by wound inoculation of unlignified stems with a macerated mycelial suspension, suggesting P. scopiformis can cause systemic infections. Thus, it is also conceivable that colonization of cambium or inner bark by P. scopiformis (Barklund and Kowalski 1996) might arise from needle infections, and vice versa. Foliar infections are probably initiated by airborne ascospores and possibly by conidia vectored by insects or rainsplash, although the infection process remains to be studied in detail.
The culture filtrate extract of an unidentified Phialocephala sp. DAOMC 229535 endophyte isolated from Picea glauca produced the antibiotic macrolide vermiculine, a known sesquiterpene, and seven previously undocumented gamma-lactones (Fig. 7; Findlay et al. 2003). The sesquiterpene and four of the gamma-lactones at a concentration of 400 µg g−1 of synthetic diet were toxic to C. fumiferana larvae, whereas vermiculine was toxic to immortalized spruce budworm cells (Findlay et al. 2003). Choristoneura fumiferana mortality associated with exposure to this Phialocephala species’ natural products prompted taxonomic investigation and inoculation studies analogous to those of P. scopiformis. Based on multigene phylogenetic analyses, Phialocephala sp. DAOMC 229535 is likely an undescribed species sister to Phialocephala s.s. (Tanney, unpublished data). Similar to Phialocephala scopiformis and P. piceae, we hypothesize that Phialocephala sp. produces mollisioid apothecia from decaying wood, probably Picea and/or unrelated hardwood species (Tanney et al. 2016a). Efforts to collect corresponding apothecia to facilitate identification, description, and isolation of new strains from ascospores are ongoing.

Structures of the macrolide vermiculine, sesquiterpene and example γ-lactone produced by unidentified Phialocephala sp. DAOMC 229535
4.1.3 Mycosphaerellaceae sp.
An endophyte of Picea mariana and P. rubens that produces mixtures of ramulosin derivatives and isocoumarins toxic to spruce budworm has been the subject of investigation for some time. An isolate originally designated as Conoplea elegantula was reported from P. mariana. Its crude extract and several purified pentaketides were toxic to spruce budworm larvae and cells (Findlay et al. 1995). The crude extracts of two undescribed P. rubens strains (CBS 121943 and DAOMC 239830) were later shown to be toxic to spruce budworm in dietary feeding assays (Sumarah and Miller 2009). Based on ITS sequences, a subsequently studied strain from P. mariana (DAOMC 250863) was 100% identical to the P. rubens endophytes CBS 121943 and DAOMC 239830. However, the isolate investigated by Findlay et al. (1995) was not deposited, preventing a direct comparison (McMullin et al. 2017a).
McMullin et al. (2017a) reported that all conspecific strains were sterile, despite attempts to induce sporulation over a two-year period involving several media and growth conditions. Eight strains isolated as Picea mariana and P. glauca endophytes were included in a phylogenetic analysis using the ITS barcode, which placed them as a sister species to Nothophaeocryptopus gaeumannii. Nothophaeocryptopus gaeumannii (Mycosphaerellaceae, Capnodiales) is a common foliar endophyte and biotrophic pathogen of Douglas-fir (Pseudotsuga menziesii), and can cause substantial losses in productivity as the causal agent of Swiss needle cast disease. The disease is marked by chlorosis and low needle retention caused by reduced gas exchange due to the occlusion of stomata by erumpent ascomata (pseudothecia) (Manter et al. 2000). The endophytic Mycosphaerellaceae species from spruce and Nothophaeocryptopus gaeumannii are most closely related to Pallidocercospora and Pseudocercospora species reported as endophytes or associated with leaf spots from diverse hosts such as Acacia mangium, Calophyllum longifolium, Eucalyptus species, and Ventilago denticulata (McMullin et al. 2017a; Winton et al. 2007). Despite being relatively common, comprising 11% of total Picea mariana endophytes in Quebec (Stefani and Bérubé 2006), this species is probably undescribed. Sampling Picea needles in Eastern Canada for mycosphaerella-like pseudothecia or conidiomata will undoubtedly result in the connection of this unknown endophyte with identifiable field specimens.
4.1.4 Diaporthe maritima
Diaporthe maritima is a common Picea mariana and P. rubens endophyte described from the Acadian forest (Tanney et al. 2016b). Field collections connecting D. maritima with conidiomata erupting through twig surface and petioles on dead or dying twigs suggest it also occurs as an endophyte in healthy twigs and could potentially be a weakly opportunistic pathogen. Diaporthe maritima is a member of the Diaporthe eres species complex (DESC), which comprises at least 15 species that are morphologically highly variable and occur on diverse hosts (Gao et al. 2016; Udayanga et al. 2014; Yang et al. 2017). Species identifications within the DESC are difficult due to variability and overlap of morphology and ambiguous clades with short branches and moderate statistical support. However, recent epitypification efforts and the use of more informative genes provides more robust species concepts (Udayanga et al. 2014; Gao et al. 2016). ITS can be insufficient for differentiating closely related Diaporthe species and may result in discordant phylogenies, necessitating the use of additional secondary markers (e.g. EF1-α, Apn2, and HIS) (Udayanga et al. 2014). Consequently, endophyte surveys using only ITS or LSU will likely underestimate Diaporthe diversity.
Species within the genus Diaporthe represent multi-host endophytes and/or tree-associated fungi that may be observed on unrelated trees from different geographical locations (Pirttilä and Frank 2011; Rehner and Uecker 1994; Webber and Gibbs 1984). Phomopsolides A–C as well as a stable pyrone were characterized from two D. maritima strains (DAOMC 628553 and DAOMC 250334) isolated from healthy P. rubens needles (Fig. 8; Tanney et al. 2016b). This dihydropyrone class of compound was originally reported from Diaporthe eres (as Phomopsis oblonga) strains associated with the outer bark of healthy Ulmus trees (Grove 1985). DESC strains are known to invade the phloem tissue of Ulmus trees infected with Ophiostoma ulmi, the causal agent of Dutch elm disease (Webber 1981). Phomopsolides possess anti-feedant and anti-boring activity against adult female large elm bark beetles (Scolytus scolytus), an insect vector for O. ulmi (Claydon et al. 1985). Because of these biological activities, DESC species producing these metabolites were hypothesized to naturally mitigate Dutch elm disease by disrupting the breeding of the pathogen’s insect vector (Webber 1981; Webber and Gibbs 1984).

Dihydropyrone phomopsolides and stable pyrone structures characterized from the Picea endophyte Diaporthe maritima
Endophytic Diaporthe species are a large source of natural product diversity, with several of their natural products directly implicated in host tree-pest chemical ecologies. From these studies, phomopsolide B appears to be the most common metabolite reported (Claydon et al. 1985; Grove 1985; Horn et al. 1996; Tanney et al. 2016b). For example, a large proportion of North American and European Silax endophytes from the genus Diaporthe produced phomopsolide B (Horn et al. 1996). Besides phomopsolides, Ulmus-derived Diaporthe species synthesize pyrones, orsellinic acids, aromatics, and isocoumarins including (R)-5-methylmellein, tyrosol, and mellein. While these latter secondary metabolites are commonly produced by phylogenetically diverse fungi, interestingly, they are synthesized by P. glauca Rhytismataceae endophytes from the Acadian forest (Sumarah et al. 2008b; McMullin et al. 2017a). When incorporated into the diet of C. fumiferana larvae, the culture filtrate extracts of these P. glauca endophytes significantly reduced both their weight and head capsule size (Sumarah et al. 2008b). Together with the phomopsolides, (R)-5-methylmellein, tyrosol, and mellein all displayed in vitro antiinsectan activities, where phomopsolide A was the most active, towards the bark beetle S. scolytus (Claydon et al. 1985). Of particular interest is the identification of the same biologically active natural products synthesized by different tree-associated fungi of different geographic origin having potentially very important ecological roles.
4.1.5 Rhytismataceae
Rhytismataceae (Rhytismatales) contains 55 genera and ca. 730 species of endophytic, parasitic, and saprotrophic plant associates, many of which are exclusively associated with conifers (Kirk et al. 2008; Lantz et al. 2011). The general morphology of Rhytismataceae consists of an ascoma immersed in host tissue with a pigmented shield-like clypeus. This is closed in dry conditions and open in humid conditions to reveal a hymenium consisting of paraphyses that are simple or branched, filiform, free or embedded in a gelatinous epithecium, often connected by hyphal bridges near the bases, with cylindrical to clavate or circinate apices, and asci with undifferentiated thin-walled inamyloid apices (Fig. 9). The ascus forcibly ejects ascospores, which are typically fusiform to filiform and surrounded in gelatinous sheaths (Darker 1967; Lantz et al. 2011; Sherwood 1980). Many Rhytismataceae genera are known from their ascomata produced from needles (e.g. Bifusella, Elytroderma, Isthmiella, Lirula, Lophodermiella, and Lophodermium), branches and bark (e.g.: Coccomyces, Colpoma, Therrya, and Tryblidiopsis).

Apothecia of some Rhytismataceae species detected as conifer endophytes. A–B. Coccomyces strobi on dead self-pruned Pinus strobus branches. C. Coccomyces irretitus on living Picea rubens bark. D. Colpoma crispum on fallen Picea mariana branches. E. Tryblidiopsis sp. On dead self-pruned Picea glauca branches. F. Lophodermium nitens on fallen Pinus strobus needles. G–H. Lophodermium piceae ascomata and conidiomata on fallen Picea abies needles
Rhytismataceae species can have both narrow host and substrate preferences, such as Lophodermium nitens and L. piceae, while evidence suggests some species may have narrow host preferences but broader, specialized substrate preferences. For example, Tryblidiopsis pinastri is specific to Picea and occurs as both a cambium and foliar endophyte, but produces ascomata and conidiomata only on dead, usually self-pruned, branches and never foliage (Barklund and Kowalski 1996; Livsey and Minter 1994; Tanney 2016). Tryblidiopsis pinastri is probably a latent pioneer saprotroph associated with self-pruning of branches, existing as a cambium endophyte for prolonged periods prior to the death of its harboring branch, after which it rapidly colonizes the substrate and eventually produces ascomata and conidiomata (Stone et al. 1996). The detection of T. pinastri as a foliar endophyte does not necessarily indicate its ability to persist in foliage, rather foliar infections might be transient and a dead end (i.e.: accidental endophytes), a probability resulting from high numbers of adhesive ascospores contacting foliage. However, the infection biology of T. pinastri is unknown, and it is conceivable that twig and branch colonization originates from both wounds and expansion from needle infections.
An unidentified Rhytismataceae endophyte (DAOMC 251461) collected from healthy Picea mariana needles produced several metabolite classes including dihydropyrones, phthalides, isocoumarins, and amino acid-derived compounds (Fig. 10; McMullin et al. 2017a). The two dihydropyrones, rhytismatone A and B, were reported as new metabolites, structurally similar to the potently antifungal compound alternaric acid. A phylogenetic analysis of Rhytismataceae endophytes collected from the Acadian forest determined that Rhytismataceae sp. DAOMC 251461 was distinct from three endophyte strains previously isolated from Picea glauca (CBS 120379–CBS 120381) with culture filtrate extracts toxic to spruce budworm in dietary bioassays (McMullin et al. 2017a; Sumarah et al. 2008b). These P. glauca strains produced mixtures of polyketide structures; however, similarities to the metabolites of Rhytismataceae sp. DAOMC 251461 were observed. The amino acid-derived compound tyrosol was produced by all four conifer endophytes and both Rhytismataceae sp. DAOMC 251461 and Tryblidiopsis sp. CBS 120380 synthesised similar isocoumarins and phthalides (McMullin et al. 2017a; Sumarah et al. 2008b).

Structures of dihydropyrones, phthalides, isocoumarins, and tyrosol produced by Picea mariana Rhytismataceae endophyte DAOMC 251461
Rhytismataceae species comprise an important family of conifer associates commonly detected as endophytes. However, Rhytismataceae taxonomy is largely neglected by contemporary mycologists. Reference sequences are lacking, hindering sequence-based identification. Identification of unknown Rhytismataceae endophytes is dependent on generating reference sequences for named-but-unsequenced species from identifiable field specimens (ascomata) and actively describing novel species when encountered. Rhytismataceae sp. DAOMC 251461 is unidentifiable based on currently available reference sequences. However, an ITS phylogeny places it in a polytomous clade including genera consisting largely of branch-associated fungi, such as Coccomyces, Colpoma, and Tryblidiopsis, as well as Lophodermium species associated with foliage (McMullin et al. 2017a). Increased field sampling of conifer-associated Rhytismataceae ascomata will eventually connect this unknown species with identifiable material. For example, the Picea glauca endophyte CBS 120380 is an undescribed Tryblidiopsis species occurring on self-pruned Picea branches in Eastern Canada (McMullin et al. 2017a). Foliar endophytism should therefore be investigated in bark or branch-inhabiting Rhytismataceae with similar life histories as Tryblidipsis, such as Coccomyces, Colpoma, and Therrya species. For example, Therrya pinastri (misidentified as Tryblidiopsis pinastri) was isolated as an endophyte from Pinus halepensis needles and is possibly a branch endophyte (Botella and Diez 2011; Solheim et al. 2013). The characterization of new and ecologically important metabolites should encourage more attention on this understudied family (McMullin et al. 2015, 2017a; Reignoux et al. 2014; Sokolski et al. 2004; Sumarah et al. 2008b, 2011, 2015; Tanney and Seifert 2017; Wang et al. 2014).
4.1.6 Pezicula sporulosa
Three chlorinated cyclopentenones and (R)-mellein were characterized from the culture filtrate extract of an endophytic strain of Pezicula sporulosa (Dermateaceae, Helotiales) collected from Picea rubens in a mixed-wood stand dominated by Betula alleghaniensis and Acer rubrum (red maple) (Fig. 11; McMullin et al. 2017a). The characterization of the antifungal metabolite (+)-crytosporiopsin from this endophytic P. sporulosa strain was not unexpected. This compound was originally reported from a Pezicula species (as Cryptosporiopsis) collected from decaying yellow birch wood (Stillwell et al. 1969; Strunz et al. 1969). A survey of endophytic Pezicula species from asymptomatic deciduous and coniferous trees in Northern Germany revealed that all eighty-five strains produced (R)-mellein, whereas only six strains produced (+)-crytosporiopsin, suggesting it is taxonomically significant (Schulz et al. 1995). From this survey, other Pezicula species-specific biologically active compounds included 4-epi-ethiosolide, 2-methyoxy-4-hydroxy-6-methyoxymethyl-benzalehyde, and (-)-mycorrhizin (Schulz et al. 1995). Interestingly, strains of Lachnum papyraceum associated with wood from Europe and North America produced mycorrhizin secondary metabolites with antiinsectan and nematicidal activities together with other chemical classes (Anke et al. 1995; McMullin et al. 2017a). (R)-mellein is also produced by a strain of an endophytic Tryblidiopsis species (CBS 120381) isolated from Picea glauca (Sumarah et al. 2008b). Other Pezicula species collected as endophytes of Fagus sylvatica, Pinus sylvestris, and Tripterygium wilfordii are a source of cyclic lipopeptides with potent antimicrobial activities (Noble et al. 1991; Strobel et al. 1999). Future efforts should focus on the identification of conifer endophytes producing cyclic peptides, because members of this chemical class including the enniatins are toxic to spruce budworm (Strongman et al. 1988; Sy-Cordero et al. 2012).

Structures of the chlorinated cyclopentenones cryptosporiopsin, 5-hydroxycryptosporiopsin and cryptosporiopsinol and (R)-mellein produced by Pezicula sporulosa
Pezicula sporulosa is reported from a diverse range of hosts including conifers such as Abies, Cupressus, Larix, and Pseudotsuga, and hardwoods such as Fagus, Pterocarya, Quercus, and Tilia (Farr and Rossman 2016; Chen et al. 2015). In general, Pezicula species are usually detected as saprotrophs and sometimes weak pathogens on recently dead branches or stems, where they may produce ascomata (apothecia) and a sporodochial asexual state (= Cryptosporiopsis). Pezicula species are also commonly isolated as branch endophytes, which, combined with the habit of ascomata and conidiomata erupting through the bark, suggests they share a similar ecology with Tryblidiopsis pinastri and may be involved in the self-pruning of branches (Butin and Kowalski 1990; Kehr 1992; Kowalski and Kehr 1992). Endophytic Pezicula strains are known from roots, shoots, and leaves, and some studies described species occurring as both foliar and branch endophytes within the same host species (Barengo et al. 2000; Chen et al. 2015; Sieber 2002; Sieber et al. 1991; Sigler et al. 2005; Verkley 1999). For example, Pezicula neosporulosa, a common needle and branch endophyte described from Abies species in Europe and China, produces ascomata on necrotic bark of the same hosts (Yuan and Verkley 2015). Taken together, these observations indicate a general pattern for Pezicula species: the ability to endophytically infect a potentially broad range of hosts while exhibiting more narrow substrate preferences for the production of reproductive structures. Thus, endophytism in Pezicula might represent a secondary life history sensu the foraging ascomycete hypothesis.
4.2 Toxigenic Endophytes of Pinus strobus
Eastern white pine (Pinus strobus) is an economically important conifer for lumber and finished wood products with several potential value-added commercial opportunities. During the North American colonial period, pine was extensively harvested for ship masts and much of the Acadian forest was cleared for agriculture to support new inhabitants (Miller 2011). Natural populations of Pinus species are severely susceptible to the disease white pine blister rust (WPBR), caused by the obligate biotrophic parasite Cronartium ribicola (Fig. 12). This invasive species, native to Asia, was independently introduced to the west coast and east coast of North America during the early 1900s (Allen and Humble 2002). Since then, it has virtually eliminated P. monticola (Western white pine) as a viable commercial species in British Columbia and as of 2010, ~70% of P. strobus (Eastern white pine) populations in Quebec were afflicted with the disease (Natural Resources Canada 2012). Cronartium ribicola infects North American five-needle pines during all growth stages, with younger trees being the most susceptible, and is one of most important forest diseases. White pine blister rust is more pervasive in cooler, humid environments, making Northern latitudes particularly susceptible to the disease. The rust propagates by alternating between Pinus and Ribes (currant/gooseberry) species. During the spring, C. ribicola spores infect Ribes species, which subsequently sporulate in the autumn to infect Pinus tree needles. Left untreated, the disease spreads sequentially from the needles, branches, and trunk, potentially killing the tree within a few years (Et-Touil et al. 1999). Silviculture disease management strategies have included pruning diseased branches and eradicating alternative hosts, or the use of fungicides (Maloy 1997). Despite this, C. ribicola remains a threat to Eastern Pinus species that are extensively utilized for reforestation purposes due to their rapid growth. Recognition and planting of Pinus trees tolerant to the disease remains an important mechanism of maintaining healthy stands.

Pinus strobus branch infected with the forest disease white pine blister rust
As with other conifers, Pinus species harbour a diverse community of foliar endophytes (e.g. Oono et al. 2015; Prihatini et al. 2016; Qadri et al. 2014). Echoing our previous experiences with Picea endophytes, foliar endophytes of Eastern white pine producing antifungal natural products were targeted in an effort to identify genotypes capable of mitigating the effects of C. ribicola. A collection comprising more than 600 P. strobus endophyte isolates was assembled and isolations are still ongoing. Incorporating the taxonomy of the isolates facilitated natural product dereplication efforts to identify new chemistries from undescribed or unexplored species. From these collections, approximately 175 isolates were screened for antifungal activity using a modified Oxford assay, where approximately 25% showed antifungal activity. This approach resulted in identifying a greater percentage of biologically active extracts compared to previous collections screened where approximately 5% showed appreciable activity (Sumarah et al. 2015). An initial phylogenetic analysis of these isolates revealed that a large proportion of these strains were Lophodermium nitens (Rhytismataceae, Rhytismatales). This abundant Pinus endophyte, together with two other species, are discussed below in the context of their taxonomy, biologically active compounds, and ecological roles within the Acadian forest.
4.2.1 Lophodermium nitens
Lophodermium nitens is a ubiquitous endophyte of Pinus strobus in Eastern Canada, where its conspicuous ascomata are commonly encountered on fallen P. strobus needles (Fig. 9f). Ascomata and conidiomata develop on fallen senesced secondary needles and mature in the spring and early summer following overwintering. Infection of new foliage is initiated by ascospores, which are released coinciding with bud break and needle extension. Lophodermium nitens endophyte infections may be prolonged but are not necessarily quiescent, rather they show signs of continual slow growth intercellularly between the epidermis and hypodermis, and later exhibit increasingly aggressive intracellular hyphal proliferation when needle senescence begins (Deckert et al. 2001). Endophytic Lophodermium nitens strains can be rapidly identified by comparing ITS sequences with available reference sequences derived from ascospore isolates.
From the culture filtrate extracts of Lophodermium nitens strains collected as white pine endophytes, the antifungal macrolides pyrenophorol, pyrenophorin, and dihydropyrenophorin were characterized together with phenolic bisabolane-type sesquiterpenoids (Fig. 13; Sumarah et al. 2011, 2015). Many of the sesquiterpenoids including sydonol were produced as natural racemic mixtures (McMullin et al. 2015). From in vitro assays, the macrolide chemical class was identified as the more antifungal constituent of these Lophodermium extracts where pyrenophorol and pyrenophorin both inhibited the growth of the biotrophic pathogen Microbotryum violaceum at 5 µM (McMullin et al. 2015; Sumarah et al. 2011). The same concentration of pyrenophorol was shown to significantly reduce the dry cell mass of Cronartium ribicola, the causal agent of white pine blister rust (Sumarah et al. 2015). These early data indicated needle endophytes of P. strobus produce potently antifungal metabolites that may inhibit the growth of forest diseases.

Structures of the macrolides pyrenophorol, pyrenophorin and dihydropyrenophorin, and phenolic bisabolane-type sesquiterpenoids produced by Lophodermium nitens
4.2.2 Hendersonia pinicola
When assessing the Pinus strobus endophyte collection for in vitro antifungal activity, extracts of two strains (DAOMC 242779 and DAOMC 242780) representing an unidentified species provided the largest response. From their culture filtrate extracts, four dihydrobenzofurans and two xanthenes were characterized (Fig. 14; Richardson et al. 2015). The xanthenes and two chlorinated dihydrobenzofurans were new structures; the remaining dihydrobenzofurans were new natural products, previously identified as synthetic intermediates. Individually tested, each metabolite displayed in vitro bioactivity towards M. violaceum and the Gram-positive bacterium Bacillus subtilis; however, not as potent as the crude extract.

Dihydrobenzofurans and xanthenes produced by the Pinus strobus endophyte Hendersonia pinicola
Based on ITS and LSU phylogenetic analyses, Richardson et al. (2015) suggested these toxigenic strains represented a new genus and species within a novel lineage of Massarinaceae (Pleosporales). However, shortly after the publication of Richardson et al. (2015), Broders et al. (2015) isolated strains of Hendersonia pinicola from Pinus strobus needles, which share 100% similar ITS sequences with the unknown Massarinaceae sp. strains (Tanney, unpublished data). This exemplifies the importance of collecting and barcoding named-but-unsequenced forest fungi, as the reference sequences generated by Broders et al. (2015) facilitated the rapid identification of an unknown species previously unidentifiable to genus, despite repeated attempts to induce taxonomically-informative characters in vitro.
Hendersonia pinicola is a well-known secondary colonizer of pine needles, often found in association with Lophodermella concolor (= Hypodermella concolor), which causes a needle cast disease on two-needle pines. Darker (1967) wrote that these secondary fungi might be extremely important from an economic viewpoint by acting as natural biological control agents: “By robbing the primary fungus of its food supply they prevent the latter from fruiting and thus act as natural biological control agents. As an example, during the height of its development in 1961, a collection (DAOM 88009) of Hendersonia pinicola Wehm., secondary after Hypodermella concolor, was so abundant that the black masses of spores hung in festoons on the needles of Pinus banksiana and blackened the green leaves of vegetation below. During the following year no evidence of either the primary or the secondary species could be found at the site of the earlier infection near North Onslow, Pontiac County, Quebec. Apparently the H. concolor infestation in this small area was completely wiped out.” Additional evidence suggests the potential biocontrol of species of Lophodermella through the reduction of inoculum availability by invading secondary Hendersonia species (Czabator et al. 1971; Jalkanen and Laakso 1986; Mitchell et al. 1976; Staley and Bynum 1972). The production of antifungal mixtures by H. pinicola could explain some of these earlier observations. Hendersonia is a large, neglected, and probably polyphyletic genus (Broders et al. 2015). However, earlier observations suggesting biocontrol potential and work by Richardson et al. (2015) should motivate future taxonomic work and natural product investigations.
4.2.3 Xylaria sp.
Recent studies have shown the ecologies of conifer endophytes from the Acadian forest are more complicated than anticipated. A currently undescribed Xylaria species was collected as a common endophyte of Pinus strobus and selected for further investigation, as Xylariaceae have been a rich source of diverse metabolites (Stadler 2011). The potently antifungal compound griseofulvin was isolated as a major constituent of its antifungal culture filtrate extract, together with dechlorogriseofulvin, a dihydrobenzofuran, and piliformic acid (Fig. 15; Richardson et al. 2014). The identification of piliformic acid (2-hexylidene-3-methylsuccinic acid) is chemotaxonomically important as it is produced characteristically by several temperate Xylaria species (Whalley and Edwards 1995; Richardson et al. 2014). Griseofulvin production has been reported from a narrower group of Xylariaceae species (Lee et al. 2016b; Sica et al. 2016; Park et al. 2005; Whalley and Edwards 1995). This antifungal metabolite is known to inhibit the growth of several plant pathogens, including rusts, in the low µM range, and is not phytotoxic (Richardson et al. 2014). A corresponding study of fruit-bearing plants in Eastern Canada identified griseofulvin-producing Xylaria strains from lowbush blueberry stems (Vaccinium angustifolium). Phylogenetic analysis revealed these were the same undescribed Xylaria species and several strains isolated from both hosts produced griseofulvin in culture (Richardson et al. 2014). These observations have prompted seedling inoculation studies with this Xylaria sp. (described below) as well as further taxonomic and metabolomic investigations (Ibrahim et al., in preparation).

Griseofulvin, dechlorogriseofulvin, piliformic acid and the dihydrobenzofuran produced by endophytic strains of the discussed undescribed Xylaria species
The phylogeny presented by Richardson et al. (2014) shows this Xylaria sp. was detected in other studies, including as an endophyte of the liverwort Trichocolea tomentella (Davis et al. 2003). Xylaria sp. is now known as an endophyte from Picea glauca, P. mariana, P. rubens, and has a diverse host range including other conifers and even mosses, bryophytes, and lichens (Ibrahim et al., in preparation). A hypothetical life history for Xylaria sp. illustrated in Fig. 1. is likely applicable to many endophytic Xylaria species.
5 Endophyte Inoculation Studies
The ecological roles of conifer endophytes within the Acadian forest have been the subject of considerable study over the past three decades. As described above, phylogenetically-diverse endophytes from the Acadian forest represent a prolific source of natural products with antifungal, antibiotic, and antiinsectan activities. Studies focused on the ecology and impact of foliar endophytes on spruce budworm or other forest pest and pathogen populations are critically dependent on methods to detect both the fungi and principal metabolites in planta. The development of polyclonal antibodies specific to endophyte mycelial proteins previously represented a suitable approach to monitor the infection and persistence of selected toxigenic strains in seedlings. Together with HPLC-UV analysis for rugulosin in needles, these analytical methods enabled earlier nursery and field studies with Phialocephala scopiformis (Sumarah et al. 2008a; Miller et al. 2008, 2009). While being adequately sensitive and specific, antibody-based approaches are very labour-intensive and require large sample amounts. A shift from ELISA to molecular diagnostic methods, such as qPCR, was made feasible by more reliable reference sequences and knowledge advancements in the taxonomy and phylogenetics of conifer endophytes, a prerequisite to developing more effective and reproducible molecular diagnostic methods.
The Picea foliar endophyte Phialocephala scopiformis was extensively studied over the past few decades because it synthesizes rugulosin, a toxin that negatively affects the growth of C. fumiferana. Previous efforts studying the colonization and persistence of P. scopiformis in Picea glauca trees relied upon ELISA methods together with HPLC-UV to monitor rugulosin concentrations (Miller et al. 2009; Sumarah et al. 2005, 2008a). Using ELISA, Miller et al. (2009) showed that after three years, P. scopiformis was detected in 40% of the 250 P. glauca seedlings planted under the emerging canopies of 50 P. glauca trees previously inoculated with P. scopiformis. Armed with greater taxonomic insight for the endophyte strains producing insect toxins or antifungal compounds, qPCR methods were developed to monitor their persistence within conifers (Frasz et al. 2014; McMullin et al. 2017b). Using a P. scopiformis-specific qPCR probe targeting the ITS region, Frasz et al. (2014) tested 291 P. glauca trees eleven years after inoculation. They reported 56.7% of needle samples collected throughout the crown were colonized by P. scopiformis. Needles samples were also tested for rugulosin, where the mean (1.2 µg g−l) and median (0.93 µg g−l) rugulosin concentrations were approximately twice the in vitro minimum lowest observed effect level (0.5 µg g–l). These results combined with those of Miller et al. (2009) demonstrated that P. scopiformis persists following inoculation, is horizontally-transmitted throughout the crown, and is capable of infecting proximal naturally regenerating Picea seedlings.
A follow-up study aimed to monitor three additional endophyte genotypes discussed above from inoculated Picea glauca and Pinus strobus seedlings by qPCR (McMullin et al. 2017b). Tandem mass spectrometric methods specific to each endophytes’ principal toxin were also implemented to replace HPLC-UV analysis used previously to quantify rugulosin from needles. Seedlings were inoculated with a single endophyte species and their toxins were quantified to determine the antifungal and antiinsectan metabolites concentrations within host foliage. From inoculated Pinus strobus seedlings, endophytes and corresponding metabolites considered were Xylaria sp. DAOMC 242774 and L. nitens CBS 127939, which produce the antifungal compounds griseofulvin and pyrenophorol, respectively. From inoculated Picea glauca seedlings, the undescribed vermiculine-producing Phialocephala sp. DAOMC 229535 was evaluated together with the rugulosin-producing Phialocephala scopiformis DAOMC 229536. In all cases, there was a high concordance between the qPCR and LC-MS/MS results. All samples positive for the respective toxins were positive for the producing endophyte with the accompanying qPCR assay. The undescribed Phialocephala sp. was detected from 83% (25/30) of samples where the mean vermiculine concentration in needles was 0.5 µg g−1 from 82% (22/27) of samples (McMullin et al. 2017b). All samples (n = 25) tested for P. scopiformis DAOMC 229536 were positive for both the endophyte and its toxin. Akin to previous studies, the mean rugulosin concentration was in the range expected to affect the growth of spruce budworm larvae (> 1.0 ppm; see Frasz et al. 2014; Sumarah et al. 2005, 2008a).
Griseofulvin was detected from 78% (25/32) of Pinus strobus seedlings inoculated with Xylaria sp. DAOMC 242774, a high griseofulvin-producing strain in culture. Unlike the other endophyte metabolites quantified, mean and median griseofulvin concentrations were divided between two distinct groups. The mean griseofulvin concentration from the high toxin group equated to approximately 5 µM of the antifungal compound in needles (McMullin et al. 2017b). This concentration is in the range where griseofulvin is known to be toxic to several plant disease causal agents, including rusts (Napier et al. 1956). The mean concentration of the macrolide pyrenophorol from white pine seedlings inoculated with Lophodermium nitens CBS 127939 was also in the low µM range (McMullin et al. 2017b). A 5 µM dose of pyrenophorol reduced the dry cell mass of a Cronartium ribicola strain, collected from afflicted P. strobus seedlings (Sumarah et al. 2015). As stated above, the macrolides pyrenophorol and pyrenophorin both inhibited the growth of the biotrophic pathogen M. violaceum at the same concentration (McMullin et al. 2015; Sumarah et al. 2011). In all cases, forest pests and pathogens would be exposed to a greater toxin concentration as a proportion of the biologically available water would be bound by needle structural components. These data further support the hypothesis that species-specific strains synthesize natural products that can increase host tolerance to pests and pathogens. These studies have been enabled by advances in modern analytical instrumentation and investments into the informative taxonomy of conifer endophytes from the Acadian forest.
6 Conclusions
Forests are critical to global ecological and economic integrity, yet relatively little is known concerning the composite forest mycobiota. Endophytes are omnipresent in forest ecosystems, where they represent a wealth of undescribed biodiversity and novel natural products, and likely play an important role modulating forest health. Improving our understanding of endophyte diversity and taxonomy is critical to developing a more holistic approach to forest health, including the development of integrative forest pest management tools. For example, finding higher rugulosin producers requires access to a large number of Phialocephala scopiformis strains. Previously, isolating P. scopiformis endophytes was a more-or-less chance occurrence; however, now countless single-ascospore P. scopiformis strains can be generated by collecting apothecia commonly occurring on decaying fallen Picea rubens branches. Other practical benefits of investing in taxonomic research of endophytes, and forest mycobiota in general, include improved surveillance and monitoring for invasive pathogens and more informed forest management practices.
The evidence marshalled in this chapter should provide strong support for field collecting efforts and the mining of invaluable resources such as herbaria and culture collections. Classical taxonomic knowledge is needed now more than ever and the synergistic marriage of both classical taxonomy and modern molecular methods leads to significant advances in mycology. Mounting evidence connecting endophytic and saprotrophic life histories cannot be ignored. Endophyte research programs should include efforts to connect unidentified endophytes with field specimens, which will increase the resolution of subsequent studies and provide invaluable reference sequences for studies using NGS. In general, funding agencies should be more supportive of “traditional” methods and fieldwork, because our lack of taxonomic knowledge cannot yet be circumvented by adopting new technology (see Truong et al. 2017). Further, the identification of novel or known bioactive secondary metabolites from conifer endophytes of the Acadian Forest Region is possible only because of culturing endeavors, and screening is becoming incrementally more efficient because of taxonomy-guided chemistry. Future culture- or inoculation-based work should include connecting genomic data with phenotypic data to identify genetic signatures of endophytism and the mechanisms driving endophyte-saprotroph life histories and host-switching strategies.
This multi-disciplinary investigation of endophytes has already resulted in the discovery of novel species, new natural products, and assists the ongoing development of integrated forest pest management tools to combat forest pests and pathogens. Endemic endophytes are an overlooked source of forest biodiversity in the Northern hemisphere and may provide tools that protect managed forests from destructive native and alien species. One consequence of evidence garnered from endophyte research is that studies investigating resistance among forest tree populations must also consider the possible role of endophytes, and not just the host genotype, in expressed phenotypic variation.
Just as old growth Picea rubens forests are reservoirs of tree genetic diversity and possible superior seed sources (Mosseler et al. 2003b), these same forests also contain a great biodiversity of horizontally-transmitted endophytes and represent important sources of potentially commercially viable endophyte strains. Forest endophyte diversity is reduced by disturbances from insect pests and pathogens, fire, and commercial logging (Miller 2011). The high incidence of horizontally-transmitted endophytes detected from inoculated nursery seedlings suggests this approach provides a mechanism to reintroduce natural endophyte populations during reforestation efforts. Additionally, this knowledge also provides a pragmatic incentive to protect old growth forests and ascribes additional value to the retention of trees and coarse woody debris, which may serve as sources of endophyte inoculum for regenerating forests or for future discoveries.
References
Abarenkov K, Henrik Nilsson R, Larsson KH, Alexander IJ, Eberhardt U, Erland S, Høiland K, Kjøller R, Larsson E, Pennanen T (2010) The UNITE database for molecular identification of fungi–recent updates and future perspectives. New Phytol 186:281–285
Allen E, Humble L (2002) Nonindigenous species introductions: a threat to Canada’s forests and forest economy. Can J Plant Path 24:103–110
Aly AH, Debbab A, Kjer J, Proksch P (2010) Fungal endophytes from higher plants: a prolific source of phytochemicals and other bioactive natural products. Fungal Divers 41:1–16
Amos-Binks LJ, MacLean DA et al (2010) Temporal changes in species composition of mixed wood stands in northwest New Brunswick: 1946–2008. Can J For Res 40:1–12
Ando K (1992) A study of terrestrial aquatic hyphomycetes. Trans Mycol Soc Jpn 33:415–425
Anke H, Stadler M, Mayer A, Sterner O (1995) Secondary metabolites with nematicidal and antimicrobial activity from nematophagous fungi and Ascomycetes. Can J Bot 73:932–939
Arnold AE, Lutzoni F (2007) Diversity and host range of foliar fungal endophytes: are tropical leaves biodiversity hotspots? Ecology 88:541–549
Arnold AE, Mejía LC, Kyllo D, Rojas EI, Maynard Z, Robbins N, Herre EA (2003) Fungal endophytes limit pathogen damage in a tropical tree. Proc Natl Acad Sci 100:15649–15654
Badotti F, de Oliveira FS, Garcia CF, Vaz ABM, Fonseca PLC, Nahum LA, Oliveira G, Góes-Neto A (2017) Effectiveness of ITS and sub-regions as DNA barcode markers for the identification of Basidiomycota (Fungi). BMC Microbiol 17:42
Bandoni RJ, Koske RE (1974) Monolayers and microbial dispersal. Science 183:1079–1081
Barengo N, Sieber TN, Holdenrieder O (2000) Diversity of endophytic mycobiota in leaves and twigs of pubescent birch (Betula pubescens). Sydowia 52:305–320
Barklund P (1987) Occurrence and pathogenicity of Lophodermium piceae appearing as an endophyte in needles of Picea abies. Trans Br Mycol Soc 89:307–313
Barklund P, Kowalski T (1996) Endophytic fungi in branches of Norway spruce with particular reference to Tryblidiopsis pinastri. Can J Bot 74:673–678
Bills GF, González-Menéndez V, Martín J, Platas G, Fournier J, Peršoh D, Stadler M (2012) Hypoxylon pulicicidum sp. nov. (Ascomycota, Xylariales), a pantropical insecticide-producing endophyte. PLoS ONE 7:e46687
Blais J (1983) Trends in the frequency, extent, and severity of spruce budworm outbreaks in eastern Canada. Can J For Res 13:539–547
Botella L, Diez JJ (2011) Phylogenic diversity of fungal endophytes in Spanish stands of Pinus halepensis. Fungal Divers 47:9–18
Broders K, Munck I, Wyka S, Iriarte G, Beaudoin E (2015) Characterization of fungal pathogens associated with white pine needle damage (WPND) in Northeastern North America. Forests 6:4088–4104
Butin VH, Kowalski T (1990) Die natürliche Astreinigung und ihre biologischen Voraussetzungen V. Die Pilzflor.von Fichte, Kiefer und Lärche. Eur J For Pathol 20:44–54
Calhoun LA, Findlay JA, Miller JD, Whitney NJ (1992) Metabolites toxic to spruce budworm from balsam fir needle endophytes. Mycol Res 96:281–286
Carroll GC (1999) The foraging ascomycete. In: 16th International Botanical Congress, St. Louis, MO, USA
Chang W-Y, Lantz VA, Hennigar CR, MacLean DA (2012) Economic impacts of forest pests: a case study of spruce budworm outbreaks and control in New Brunswick, Canada. Can J For Res 42:490–505
Chauvet E, Cornut J, Sridhar KR, Selosse M-A, Bärlocher F (2016) Beyond the water column: aquatic hyphomycetes outside their preferred habitat. Fungal Ecol 19:112–127
Chaverri P, Gazis RO (2011) Linking ex planta fungi with their endophytic stages: Perisporiopsis, a common leaf litter and soil fungus, is a frequent endophyte of Hevea spp. and other plants. Fungal Ecol 4:94–102
Chen C, Verkley GJ, Sun G, Groenewald JZ, Crous PW (2015) Redefining common endophytes and plant pathogens in Neofabraea, Pezicula, and related genera. Fungal Biol 120:1291–1322
Clay K (1988) Fungal endophytes of grasses: a defensive mutualism between plants and fungi. Ecology 69:10–16
Claydon N, Grove JF, Pople M (1985) Elm bark beetle boring and feeding deterrents from Phomopsis oblonga. Phytochemistry 24:937–943
Czabator F, Staley J, Snow G (1971) Extensive southern pine needle blight during 1970–1971, and associated fungi. Plant Dis Rep 55:764–766
Darker GD (1967) A revision of the genera of the Hypodermataceae. Can J Bot 45:1399–1444
Davis EC, Franklin JB, Shaw AJ, Vilgalys R (2003) Endophytic Xylaria (Xylariaceae) among liverworts and angiosperms: phylogenetics, distribution, and symbiosis. Am J Bot 90:1661–1667
de Beer Z, Marincowitz S, Duong T, Kim J, Rodrigues A, Wingfield M (2016) Hawksworthiomyces gen. nov. (Ophiostomatales) Hawksworthiomyces gen. nov. (Ophiostomatales), illustrates the urgency for a decision on how to name novel taxa known only from environmental nucleic acid sequences (ENAS). Fungal Biol 120:1323–1340
Deckert RJ, Melville LH, Peterson RL (2001) Structural features of a Lophodermium endophyte during the cryptic life-cycle phase in the foliage of Pinus strobus. Mycol Res 105:991–997
Dreyfuss M, Petrini O (1984) Further investigations on the occurrence and distribution of endophytic fungi in tropical plants. Bot Helv 94:33–40
Dymond CC, Neilson ET, Stinson G, Porter K, MacLean DA, Gray DR, Campagna M, Kurz WA (2010) Future spruce budworm outbreak may create a carbon source in eastern Canadian forests. Ecosystems 13:917–931
Etheridge DA, MacLean DA, Wagner RG et al (2005) Changes in landscape composition and stand structure from 1945–2002 on an industrial forest in New Brunswick, Canada. Can J For Res 35:1965–1977
Etheridge DA, MacLean DA, Wagner RG, Wilson JS (2006) Effects of intensive forest management on stand and landscape characteristics in northern New Brunswick, Canada (1945–2027). Landscape Ecol 21:509–524
Et-Touil K, Bernier L, Beaulieu J, Bérubé J, Hopkin A, Hamelin R (1999) Genetic structure of Cronartium ribicola populations in eastern Canada. Phytopathology 89:915–919
Farr D, Rossman A (2016). Fungal Databases, Systematic Mycology and Microbiology Laboratory, ARS, USDA. Retrieved 25 Sept 2017, from http://nt.ars-grin.gov/fungaldatabases/
Findlay JA, Buthelezi S, Lavoie R, Peña-Rodriguez L, Miller JD (1995) Bioactive isocoumarins and related metabolites from conifer endophytes. J Nat Prod 58:1759–1766
Findlay JA, Li G, Miller JD, Womiloju TO (2003) Insect toxins from spruce endophytes. Can J Chem 81:284–292
Fleming RA, Candau J-N, McAlpine RS (2002) Landscape-scale analysis of interactions between insect defoliation and forest fire in central Canada. Clim Change 55:251–272
Frasz SL, Walker AK, Nsiama TK, Adams GW, Miller JD (2014) Distribution of the foliar fungal endophyte Phialocephala scopiformis and its toxin in the crown of a mature white spruce tree as revealed by chemical and qPCR analyses. Can J For Res 44:1138–1143
Gao Y, Liu F, Cai L (2016) Unravelling Diaporthe species associated with Camellia. Syst Biodivers 14:102–117
Gazis R, Rehner S, Chaverri P (2011) Species delimitation in fungal endophyte diversity studies and its implications in ecological and biogeographic inferences. Mol Ecol 20:3001–3013
Gönczöl J, Révay Á (2004) Fungal spores in rainwater: stemflow, throughfall and gutter conidial assemblages. Fungal Divers 16:67–86
Green BD (2016). Characterization of secondary metabolites produced by foliar endophytes of red and black spruce. MSc. Thesis. Department of Chemistry, Carleton University Ottawa
Greenfield M, Pareja R, Ortiz V, Gómez-Jiménez MI, Vega FE, Parsa S (2015) A novel method to scale up fungal endophyte isolations. Biocontrol Sci Technol 25:1208–1212
Grove JF (1985) Metabolic products of Phomopsis oblonga. Part 2. Phomopsolide A and B, tiglic esters of two 6-substituted 5, 6-dihydro-5-hydroxypyran-2-ones. J Chem Soc, Perkin Trans 1:865–869
Grünig CR, Queloz V, Duò A, Sieber TN (2009) Phylogeny of Phaeomollisia piceae gen. sp. nov.: a dark, septate, conifer-needle endophyte and its relationships to Phialocephala and Acephala. Mycol Res 113:207–221
Haemmerli UA, Brändle UE, Petrini O, McDermott JM (1992) Differentiation of isolates of Discula umbrinella (Teleomorph: Apiognomonia errabunda) from beech, chestnut, and oak using randomly amplified polymorphic DNA markers. MPMI 5:479–483
Hanada RE, Pomella AWV, Costa HS, Bezerra JL, Loguercio LL, Pereira JO (2010) Endophytic fungal diversity in Theobroma cacao (cacao) and T. grandiflorum (cupuaçu) trees and their potential for growth promotion and biocontrol of black-pod disease. Fungal Biol 114:901–910
Hawksworth DL, Hibbett DS, Kirk PM, Lücking R (2016) (308–310) Proposals to permit DNA sequence data to serve as types of names of fungi. Taxon 65:899–900
Hibbett DS, Taylor JW (2013) Fungal systematics: is a new age of enlightenment at hand? Nat Rev Microbiol 11:129–133
Hibbett D, Abarenkov K, Kõljalg U, Öpik M, Chai B, Cole J, Wang Q, Crous P, Robert V, Helgason T (2016) Sequence-based classification and identification of Fungi. Mycologia 108:1049–1068
Hom J, Oechel W (1983) The photosynthetic capacity, nutrient content, and nutrient use efficiency of different needle age-classes of black spruce (Picea mariana) found in interior Alaska. Can J For Res 13:834–839
Horn WS, Simmonds MS, Schwartz RE, Blaney WM (1996) Variation in production of phomodiol and phomopsolide B by Phomopsis spp. Mycologia: 588–595
Huang Y-L, Devan MN, U’Ren JM, Furr SH, Arnold AE (2016) Pervasive effects of wildfire on foliar endophyte communities in montane forest trees. Microb Ecol 71:452–468
Ibrahim A, Fei F, Tanney JB, McDowell T, Ejim L, Seifert KA, Sørensen D, Capretta A, Sumarah MW (2018) Metabome-guided discovery of cyclic non-ribosomal peptides from Xylaria elissi—a new griseofulvin-producing endophyte from Vaccinium angustifolium (in preparation)
Isaka M, Tanticharoen M, Thebtaranonth Y (2000) Cordyanhydrides A and B. Two unique anhydrides from the insect pathogenic fungus Cordyceps pseudomilitaris BCC 1620. Tetrahedron Lett 41:1657–1660
Jalkanen R, Laakso R (1986) Hendersonia acicola in an epidemic caused by Lophodermella sulcigena with special reference to biological control. Karstenia 26:49–56
Johnson JA, Whitney NJ (1989) An investigation of needle endophyte colonization patterns with respect to height and compass direction in a single crown of balsam fir (Abies balsamea). Can J Botany 67:723–725
Jumpponen A, Jones K (2009) Massively parallel 454 sequencing indicates hyperdiverse fungal communities in temperate Quercus macrocarpa phyllosphere. New Phytol 184:438–448
Kehr R (1992) Pezicula canker of Quercus rubra L., caused by Pezicula cinnamomea (DC.) Sacc. II. Morphology and biology of the causal agent. For Pathol 22:29–40
Khan Z, Rho H, Firrincieli A, Hung SH, Luna V, Masciarelli O, Kim S-H, Doty SL (2016) Growth enhancement and drought tolerance of hybrid poplar upon inoculation with endophyte consortia. Curr Plant Biol 6:38–47
Kirk P, Cannon P, Minter D, Stalpers J (2008) Dictionary of the Fungi, 10th edn. CAB International, University Press, Cambridge
Knapp DG, Kovács GM, Zajta E, Groenewald JZ, Crous P (2015) Dark septate endophytic pleosporalean genera from semiarid areas. Persoonia 35:87–100
Koukol O, Kolařík M, Kolářová Z, Baldrian P (2012) Diversity of foliar endophytes in wind-fallen Picea abies trees. Fungal Divers 54:69–77
Kowalski T, Kehr R (1992) Endophytic fungal colonization of branch bases in several forest tree species. Sydowia 44:137–168
Kowalski T, Kehr R (1995) Two new species of Phialocephala occurring on Picea and Alnus. Can J Bot 73:26–32
Kuldau G, Bacon C (2008) Clavicipitaceous endophytes: their ability to enhance resistance of grasses to multiple stresses. Biol Control 46:57–71
Lantz H, Johnston P, Park D, Minter DW (2011) Molecular phylogeny reveals a core clade of Rhytismatales. Mycologia 103:57–74
Lee B-H, Eo J-K, Eom A-H (2016a) Notes on five endophytic fungal species isolated from needle leaves of conifers in Korea. Korean J Mycol 44:51–56
Lee HB, Mun HY, Nguyen TTT, Kim J-C, Stone JK (2016b) Abieticola koreana gen. et sp. nov., a griseofulvin-producing endophytic xylariaceous ascomycete from Korea. Mycotaxon 131:749–764
Leuchtmann A, Clay K (1993) Nonreciprocal compatibility between Epichloë typhina and four host grasses. Mycologia 85:157–163
Lindner DL, Carlsen T, Henrik Nilsson R, Davey M, Schumacher T, Kauserud H (2013) Employing 454 amplicon pyrosequencing to reveal intragenomic divergence in the internal transcribed spacer rDNA region in fungi. Ecol Evolut 3:1751–1764
Liu K-L, Porras-Alfaro A, Kuske CR, Eichorst SA, Xie G (2012) Accurate, rapid taxonomic classification of fungal large-subunit rRNA genes. Appl Environ Microbiol 78:1523–1533
Livsey S, Barklund P (1992) Lophodermium piceae and Rhizosphaera kalkhoffii in fallen needles of Norway spruce (Picea abies). Eur J For Path 22:204–216
Livsey S, Minter D (1994) The taxonomy and biology of Tryblidiopsis pinastri. Can J Bot 72:549–557
Loo J, Ives N (2003) The Acadian forest: historical condition and human impacts. Forest Chronicle 79:462–474
Lorimer CG (1977) The presettlement forest and natural disturbance cycle of northeastern Maine. Ecology 58:139–148
Maloy OC (1997) White pine blister rust control in North America: a case history. Annu Rev Phytopathol 35:87–109
Manter DK, Bond BJ, Kavanagh KL, Rosso PH, Filip GM (2000) Pseudothecia of Swiss needle cast fungus, Phaeocryptopus gaeumannii, physically block stomata of Douglas fir, reducing CO2 assimilation. New Phytol 148:481–491
Martin R, Gazis R, Skaltsas D, Chaverri P, Hibbett D (2015) Unexpected diversity of basidiomycetous endophytes in sapwood and leaves of Hevea. Mycologia 107:284–297
McMullin DR, Nsiama TK, Miller JD (2014) Secondary metabolites from Penicillium corylophilum isolated from damp buildings. Mycologia 106:621–628
McMullin DR, Green BD, Miller JD (2015) Antifungal sesquiterpenoids and macrolides from an endophytic Lophodermium species of Pinus strobus. Phytochem Lett 14:148–152
McMullin DR, Green BD, Prince NC, Tanney JB, Miller JD (2017a) Natural products of Picea endophytes from the Acadian forest. J Nat Prod 80:1475–1483
McMullin DR, Nguyen HDT, Daly GJ, Menard BS, Miller JD (2017b) Detection of foliar endophytes and their metabolites in Picea and Pinus seedling needles. Fungal Ecol 31:1–8
Menkis A, Allmer J, Vasiliauskas R, Lygis V, Stenlid J, Finlay R (2004) Ecology and molecular characterization of dark septate fungi from roots, living stems, coarse and fine woody debris. Mycol Res 108:965–973
Miller JD (1986) Toxins of endophytic and epiphytic fungi of conifer needles. In: Fokkema NJ, Van Huevel J (eds) Microbiology of the phyllosphere. Cambridge University Press, Cambridge, pp 223–231
Miller JD (2011) Foliar endophytes of spruce species found in the Acadian forest: basis and potential for improving the tolerance of the forest to spruce budworm. In: Endophytes of Forest Trees. Springer, Berlin, pp 237–249
Miller JD, Sumarah MW, Adams GW (2008) Effect of a rugulosin-producing endophyte in Picea glauca on Choristoneura fumiferana. J Chem Ecol 34:362–368
Miller JD, Cherid H, Sumarah MW, Adams GW (2009) Horizontal transmission of the Picea glauca foliar endophyte Phialocephala scopiformis CBS 120377. Fungal Ecol 2:98–101
Mitchell C, Williamson B, Millar C (1976) Hendersonia acicola on pine needles infected by Lophodermella sulcigena. For Pathol 6:92–102
Monnet F, Vaillant N, Hitmi A, Coudret A, Sallanon H (2001) Endophytic Neotyphodium lolii induced tolerance to Zn stress in Lolium perenne. Physiol Plant 113:557–563
Mosseler A, Lynds J, Major J (2003a) Old-growth forests of the Acadian Forest Region. Environ Rev 11:S47–S77
Mosseler A, Major J, Rajora O (2003b) Old-growth red spruce forests as reservoirs of genetic diversity and reproductive fitness. Theor Appl Genet 106:931–937
Napier EJ, Turner DI, Rhodes A (1956) The in vitro action of griseofulvin against pathogenic fungi of plants. Ann Bot 20:461–466
National Forestry Database (2017) Forest insects: spruce budworm, 1975–2015. Accessed: April 27, 2018. http://nfdp.ccfm.org/data/graphs/graph_41_a_e.php
Natural Resources Canada (2012) White pine blister rust fact sheet. Retrieved 25 Sept 2017, from http://www.tidcfnrcangcca/diseases/factsheet/24
Nielsen KF, Sumarah MW, Frisvad JC, Miller JD (2006) Production of metabolites from the Penicillium roqueforti complex. J Agric Food Chem 54:3756–3763
Noble HM, Langley D, Sidebottom P, Lane S, Fisher P (1991) An echinocandin from an endophytic Cryptosporiopsis sp. and Pezicula sp. in Pinus sylvestris and Fagus sylvatica. Mycol Res 95:1439–1440
Okane I, Srikitikulchai P, Toyama K, Læssøe T, Sivichai S, Hywel-Jones N, Nakagiri A, Potacharoen W, K-i Suzuki (2008) Study of endophytic Xylariaceae in Thailand: diversity and taxonomy inferred from rDNA sequence analyses with saprobes forming fruit bodies in the field. Mycoscience 49:359–372
Oono R, Lefèvre E, Simha A, Lutzoni F (2015) A comparison of the community diversity of foliar fungal endophytes between seedling and adult loblolly pines (Pinus taeda). Fungal Biol 119:917–928
Osono T, Tateno O, Masuya H (2013) Diversity and ubiquity of xylariaceous endophytes in live and dead leaves of temperate forest trees. Mycoscience 54:54–61
Osorio M, Stephan BR (1991) Life cycle of Lophodermium piceae in Norway spruce needles. Eur J For Path 21:152–163
Park JH, Choi G, Lee HB, Kim K, Jung HS, Lee S-W, Jang KS, Cho KY, Kim J-C (2005) Griseofulvin from Xylaria sp. strain F0010, an endophytic fungus of Abies holophylla and its antifungal activity against plant pathogenic fungi. J Microbiol Biotechnol 15:112–117
Petrini L, Petrini O (1985) Xylariaceous fungi as endophytes. Sydowia 38:216–234
Petrini O, Petrini L, Rodrigues K (1995) Xylariaceous endophytes: an exercise in biodiversity. Fitopatol Bras 20:531–539
Pirttilä AM, Frank C (2011) Endophytes of forest trees: biology and applications. Springer, Berlin
Porter TM, Golding GB (2012) Factors that affect large subunit ribosomal DNA amplicon sequencing studies of fungal communities: classification method, primer choice, and error. PLoS ONE 7(4):e35749
Prihatini I, Glen M, Wardlaw TJ, Mohammed CL (2016) Diversity and identification of fungi associated with needles of Pinus radiata in Tasmania. South Forest: J For Sci 78:19–34
Prior R, Görges K, Yurkov A, Begerow D (2014) New isolation method for endophytes based on enzyme digestion. Mycol Prog 13:849–856
Qadri M, Rajput R, Abdin MZ, Vishwakarma RA, Riyaz-Ul-Hassan S (2014) Diversity, molecular phylogeny, and bioactive potential of fungal endophytes associated with the Himalayan blue pine (Pinus wallichiana). Microb Ecol 67:877–887
Raja HA, Miller AN, Pearce CJ, Oberlies NH (2017) Fungal identification using molecular tools: a primer for the natural products research community. J Nat Prod 80:756–770
Rehner SA, Uecker FA (1994) Nuclear ribosomal internal transcribed spacer phylogeny and host diversity in the coelomycete Phomopsis. Can J Bot 72:1666–1674
Reich PB, Oleksyn J, Modrzynski J, Tjoelker MG (1996) Evidence that longer needle retention of spruce and pine populations at high elevations and high latitudes is largely a phenotypic response. Tree Physiol 16:643–647
Reignoux SN, Green S, Ennos RA (2014) Molecular identification and relative abundance of cryptic Lophodermium species in natural populations of Scots pine, Pinus sylvestris L. Fungal Biol 118:835–845
Révay Á, Gönczöl J (2010) Rainborne hyphomycete conidia from evergreen trees. Nova Hedwig 91:151–163
Révay Á, Gönczöl J (2011) Canopy fungi (“terrestrial aquatic hyphomycetes”) from twigs of living evergreen and deciduous trees in Hungary. Nova Hedwig 92:303–316
Rice J, Pinkerton B, Stringer W, Undersander D (1990) Seed production in tall fescue as affected by fungal endophyte. Crop Sci 30:1303–1305
Richardson SN, Walker AK, Nsiama TK, McFarlane J, Sumarah MW, Ibrahim A, Miller JD (2014) Griseofulvin-producing Xylaria endophytes of Pinus strobus and Vaccinium angustifolium: evidence for a conifer-understory species endophyte ecology. Fungal Ecol 11:107–113
Richardson SN, Nsiama TK, Walker AK, McMullin DR, Miller JD (2015) Antimicrobial dihydrobenzofurans and xanthenes from a foliar endophyte of Pinus strobus. Phytochemistry 117:436–443
Rocha AC, Garcia D, Uetanabaro AP, Carneiro RT, Araújo IS, Mattos CR, Góes-Neto A (2011) Foliar endophytic fungi from Hevea brasiliensis and their antagonism on Microcyclus ulei. Fungal Divers 47:75–84
Rodriguez R, White J Jr, Arnold A, Redman R (2009) Fungal endophytes: diversity and functional roles. New Phytol 182:314–330
Royama TO (1984) Population dynamics of the spruce budworm Choristoneura fumiferana. Ecological Mon 54:429–462
Royama TO, MacKinnon WE, Kettela EG, Carter NE, Hartling LK (2005) Analysis of spruce budworm outbreak cycles in New Brunswick, Canada, since 1952. Ecology 86:1212–1224
Samson RA, Visagie CM, Houbraken J, Hong S-B, Hubka V, Klaassen CH, Perrone G, Seifert KA, Susca A, Tanney JB (2014) Phylogeny, identification and nomenclature of the genus Aspergillus. Stud Mycol 78:141–173
Sati S, Belwal M (2005) Aquatic hyphomycetes as endophytes of riparian plant roots. Mycologia 97:45–49
Schoch CL, Seifert KA, Huhndorf S, Robert V, Spouge JL, Levesque CA, Chen W, Bolchacova E, Voigt K, Crous PW (2012) Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc Natl Acad Sci 109:6241–6246
Schulz B, Boyle C (2005) The endophytic continuum. Mycol Res 109:661–686
Schulz B, Sucker J, Aust H, Krohn K, Ludewig K, Jones P, Döring D (1995) Biologically active secondary metabolites of endophytic Pezicula species. Mycol Res 99:1007–1015
Selosse MA, Vohník M, Chauvet E (2008) Out of the rivers: are some aquatic hyphomycetes plant endophytes? New Phytol 178:3–7
Sherwood MA (1980) Taxonomic studies in the Phacidiales: the genus Coccomyces (Rhytismataceae). Occas Pap Farlow Herbarium Cryptogamic Bot 15:1–120
Sica VP, Rees ER, Tchegnon E, Bardsley RH, Raja HA, Oberlies NH (2016) Spatial and temporal profiling of griseofulvin production in Xylaria cubensis using mass spectrometry mapping. Front Microbiol 7:544
Sieber TN (2002) Fungal root endophytes. Plant roots: the hidden half. Marcel Dekker, New York, pp 887–917
Sieber T, Sieber-Canavesi F, Dorworth C (1991) Endophytic fungi of red alder (Alnus rubra) leaves and twigs in British Columbia. Can J Bot 69:407–411
Siegel M, Latch G, Bush L, Fannin F, Rowan D, Tapper B, Bacon C, Johnson M (1990) Fungal endophyte-infected grasses: alkaloid accumulation and aphid response. J Chem Ecol 16:3301–3315
Sigler L, Allan T, Lim SR, Berch S, Berbee M (2005) Two new Cryptosporiopsis species from roots of ericaceous hosts in western North America. Stud Mycol 53:53–62
Sokolski S, Piché Y, Bérubé JA (2004) Lophodermium macci sp. nov., a new species on senesced foliage of five-needle pines. Mycologia 96:1261–1267
Sokolski S, Piche Y, Chauvet É, Bérubé JA (2006a) A fungal endophyte of black spruce (Picea mariana) needles is also an aquatic hyphomycete. Mol Ecol 15:1955–1962
Sokolski S, Piché Y, Laitung B, Bérubé J (2006b) Streams in Quebec boreal and mixed-wood forests reveal a new aquatic hyphomycete species, Dwayaangam colodena sp. nov. Mycologia 98:628–636
Solheim H, Torp TB, Hietala AM (2013) Characterization of the ascomycetes Therrya fuckelii and T. pini fruiting on Scots pine branches in Nordic countries. Mycol Prog 12:37–44
Song M, Li X, Saikkonen K, Li C, Nan Z (2015) An asexual Epichloë endophyte enhances waterlogging tolerance of Hordeum brevisubulatum. Fungal Ecol 13:44–52
Sridhar K, Bärlocher F (1992) Endophytic aquatic hyphomycetes of roots of spruce, birch and maple. Mycol Res 96:305–308
Sridhar K, Karamchand K (2009) Diversity of water-borne fungi in stemflow and throughfall of tree canopies in India. Sydowia 61:327–344
Stadler M (2011) Importance of secondary metabolites in the Xylariaceae as parameters for assessment of their taxonomy, phylogeny, and functional biodiversity. Curr Res Environ Appl Mycol 1:75–133
Staley JM, Bynum HH (1972) A new Lophodermella on Pinus ponderosa and P. attenuata. Mycologia 64:722–726
Stefani F, Bérubé J (2006) Biodiversity of foliar fungal endophytes in white spruce (Picea glauca) from southern Québec. Botany 84:777–790
Stielow J, Lévesque C, Seifert K, Meyer W, Irinyi L, Smits D, Renfurm R, Verkley G, Groenewald M, Chaduli D (2015) One fungus, which genes? Development and assessment of universal primers for potential secondary fungal DNA barcodes. Persoonia 35:242–263
Stillwell M, Wood F, Strunz G (1969) A broad-spectrum antibiotic produced by a species of Cryptosporiopsis. Can J Microbiol 15:501–507
Stone JK, Sherwood MA, Carroll GC (1996) Canopy microfungi: function and diversity. Northwest Sci 70:37–45
Strobel G, Daisy B (2003) Bioprospecting for microbial endophytes and their natural products. Microbiol Mol Biol Rev 67:491–502
Strobel GA, Miller RV, Martinez-Miller C, Condron MM, Teplow DB, Hess W (1999) Cryptocandin, a potent antimycotic from the endophytic fungus Cryptosporiopsis cf. quercina. Microbiology 145:1919–1926
Strongman DB, Strunz GM, Giguère P, Yu C-M, Calhoun L (1988) Enniatins from Fusarium avenaceum isolated from balsam fir foliage and their toxicity to spruce budworm larvae, Choristoneura fumiferana (Clem.) (Lepidoptera: Tortricidae). J Chem Ecol 14:753–764
Strunz G, Court A, Komlossy J, Stillwell M (1969) Cryptosporiopsin, an amended structure. Can J Chem 47:3700–3701
Sumarah MW, Miller JD, Adams GW (2005) Measurement of a rugulosin-producing endophyte in white spruce seedlings. Mycologia 97:770–776
Sumarah MW, Adams GW, Berghout J, Slack GJ, Wilson AM, Miller JD (2008a) Spread and persistence of a rugulosin-producing endophyte in Picea glauca seedlings. Mycol Res 112:731–736
Sumarah MW, Puniani E, Blackwell BA, Miller JD (2008b) Characterization of polyketide metabolites from foliar endophytes of Picea glauca. J Nat Prod 71:1393–1398
Sumarah MW, Miller JD (2009) Anti-insect secondary metabolites from fungal endophytes of conifer trees. Nat Prod Commun 4:1497–1504
Sumarah MW, Kesting JR, Sørensen D, Miller JD (2011) Antifungal metabolites from fungal endophytes of Pinus strobus. Phytochemistry 72:1833–1837
Sumarah MW, Puniani E, Sørensen D, Blackwell BA, Miller JD (2010) Secondary metabolites from anti-insect extracts of endophytic fungi isolated from Picea rubens. Phytochemistry 71:760–765
Sumarah MW, Walker AK, Seifert KA, Todorov A, Miller JD (2015) Screening of fungal endophytes isolated from eastern white pine needles. The formation, structure and activity of phytochemicals. Springer, Berlin, pp 195–206
Suske J, Acker G (1989) Identification of endophytic hyphae of Lophodermium piceae in tissues of green, symptomless Norway spruce needles by immunoelectron microscopy. Can J Bot 67:1768–1774
Sy-Cordero AA, Pearce CJ, Oberlies NH (2012) Revisiting the enniatins: a review of their isolation, biosynthesis, structure determination, and biological activities. J Antibiot 65:541
Tanney JB (2016) A taxonomic and phylogenetic investigation of conifer endophytes of Eastern Canada. PhD Thesis. Department of Biology. Carleton University, Ottawa
Tanney JB, Douglas B, Seifert KA (2016a) Sexual and asexual states of some endophytic Phialocephala species of Picea. Mycologia: 15–136
Tanney JB, McMullin DR, Green BD, Miller JD, Seifert KA (2016b) Production of antifungal and antiinsectan metabolites by the Picea endophyte Diaporthe maritima sp. nov. Fungal Biol 120:1448–1457
Tanney JB, Seifert KA (2017) Lophodermium resinosum sp. nov. from red pine (Pinus resinosa) in Eastern Canada. Botany 95:773–784
Tedersoo L, Lindahl B (2016) Fungal identification biases in microbiome projects. Environ Microbiol Rep 8:774–779
Thiéry O, Moora M, Vasar M, Zobel M, Öpik M (2012) Inter-and intrasporal nuclear ribosomal gene sequence variation within one isolate of arbuscular mycorrhizal fungus, Diversispora sp. Symbiosis 58:135–147
Thiéry O, Vasar M, Jairus T, Davison J, Roux C, Kivistik PA, Metspalu A, Milani L, Saks Ü, Moora M (2016) Sequence variation in nuclear ribosomal small subunit, internal transcribed spacer and large subunit regions of Rhizophagus irregularis and Gigaspora margarita is high and isolate-dependent. Mol Ecol 25:2816–2832
Thomas DC, Vandegrift R, Ludden A, Carroll GC, Roy BA (2016) Spatial ecology of the fungal genus Xylaria in a tropical cloud forest. Biotropica 48:381–393
Truong C, Mujic AB, Healy R, Kuhar F, Furci G, Torres D, Moretto A (2017) How to know the fungi: combining field inventories and DNA-barcoding to document fungal diversity. New Phytol 214:913–919
U’Ren JM, Lutzoni F, Miadlikowska J, Laetsch AD, Arnold AE (2012) Host and geographic structure of endophytic and endolichenic fungi at a continental scale. Am J Bot 99:898–914
U’Ren JM, Miadlikowska J, Zimmerman NB, Lutzoni F, Stajich JE, Arnold AE (2016) Contributions of North American endophytes to the phylogeny, ecology, and taxonomy of Xylariaceae (Sordariomycetes, Ascomycota). Mol Phylogen Evol 98:210–232
Udayanga D, Castlebury LA, Rossman AY, Chukeatirote E, Hyde KD (2014) Insights into the genus Diaporthe: phylogenetic species delimitation in the D. eres species complex. Fungal Divers 67:203–229
van Dijk EL, Jaszczyszyn Y, Thermes C (2014) Library preparation methods for next-generation sequencing: tone down the bias. Exp Cell Res 322:12–20
Verkley G (1999) A monograph of the genus Pezicula and its anamorphs. Stud Mycol 44:5–171
Vilgalys R, Hester M (1990) Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J Bacteriol 172:4238–4246
Visagie CM, Houbraken J, Frisvad JC, Hong S-B, Klaassen CHW, Perrone G, Seifert KA, Varga J, Yaguchi J, Samson RA (2014) Identification and nomenclature of the genus Penicillium. Stud Mycol 78:343–371
Vrålstad T (2011) ITS, OTUs and beyond—fungal hyperdiversity calls for supplementary solutions. Mol Ecol 20:2873–2875
Wang S, Cannon P, Li Z-J, Hou C-L (2014) Multigene phylogenetic analysis detects cryptic species of Tryblidiopsis in China. Mycologia 106:95–104
Webber J (1981) A natural biological control of Dutch elm disease. Nature 292:449–451
Webber J, Gibbs JN (1984) Colonization of elm bark by Phomopsis oblonga. Trans Br Mycol Soc 82:348–352
Webster J (1959) Experiments with spores of aquatic hyphomycetes. I. Sedimentation and impaction on smooth surfaces. Ann Bot 23:595–611
Wein RW, Moore JM (1977) Fire history and rotations in the New Brunswick Acadian Forest. Can J For Res 7:285–294
Whalley AJS, Edwards RL (1995) Secondary metabolites and systematic arrangement within the Xylariaceae. Can J Bot 73(S1):802–810
Wilson RW, Wheatcroft RGC, Miller JD, Whitney NJ (1994) Genetic diversity among natural populations of endophytic Lophodermium pinastri from Pinus resinosa. Mycol Res 98:740–744
Winton LM, Stone JK, Hansen EM, Shoemaker R (2007) The systematic position of Phaeocryptopus gaeumannii. Mycologia 99:240–252
Xia C, Zhang X, Christensen MJ, Nan Z, Li C (2015) Epichloë endophyte affects the ability of powdery mildew (Blumeria graminis) to colonise drunken horse grass (Achnatherum inebrians). Fungal Ecol 16:26–33
Yang Q, Fan X, Du Z, Tian C (2017) Diaporthe juglandicola sp. nov. (Diaporthales, Ascomycetes), evidenced by morphological characters and phylogenetic analysis. Mycosphere 8:817–826
Yuan Z-L, Rao L-B, Chen Y-C, Zhang C-L, Wu Y-G (2011) From pattern to process: species and functional diversity in fungal endophytes of Abies beshanzuensis. Fungal Biol 115:197–213
Yuan Z, Verkley GJ (2015) Pezicula neosporulosa sp. nov. (Helotiales, Ascomycota), an endophytic fungus associated with Abies spp. in China and Europe. Mycoscience 56:205–213
Acknowledgements
We thank many undergraduate and graduate students and post-doctoral fellows who have contributed to this research over the past three decades. We are grateful for the support and advice of Greg Adams, Shona Millican and Andrew McCartney from JD Irving Limited, Dr. Keith Seifert from Agriculture Agi-Food Canada (Ottawa), and Prof. David Malloch. We thank the Natural Sciences and Engineering Research Council of Canada, the Province of Ontario through the Mitacs program and JD Irving Limited for supporting this research. Jason Karakehian kindly provided the image of Colpoma crispum (Fig. 9d).
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2018 Springer International Publishing AG, part of Springer Nature
About this chapter

Cite this chapter
Tanney, J.B., McMullin, D.R., Miller, J.D. (2018). Toxigenic Foliar Endophytes from the Acadian Forest. In: Pirttilä, A., Frank, A. (eds) Endophytes of Forest Trees. Forestry Sciences, vol 86. Springer, Cham. https://doi.org/10.1007/978-3-319-89833-9_15
Download citation
DOI: https://doi.org/10.1007/978-3-319-89833-9_15
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-89832-2
Online ISBN: 978-3-319-89833-9
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)