
Abstract
In this paper, we present and discuss pollen data from the Early Pleistocene (1.8 to 1.6 Ma) – we use the revised timescale approved by IUGS, in which the base of the Pleistocene is defined by the GSSP of the Gelasian Stage at 2.588 (2.6) Ma (Gibbard et al. 2010) – and from the Mid-Pleistocene Transition (0.9 to 0.6 Ma) at Melka Kunture (Upper Awash , Ethiopia). At 2000 m asl in the Ethiopian highlands, these deposits yield many rich and successive archaeological sites, notably documenting the late Oldowan, the emergence of the Acheulean and the middle Acheulean. The stratigraphic position of the fifteen pollen samples is checked by 40Ar/39Ar dating and by geological investigation. Furthermore, they are now correlated to archaeological layers whose excavated lithic industries have been reinterpreted. Our study shows that mountain forest trees belonging to the present-day Afromontane complex were already established in Ethiopia at ~1.8 Ma and that the knappers of the Oldowan and early Acheulean could cope with mountain climatic conditions that had a large diurnal temperature range. Moreover, the new interpretation of pollen results emphasizes changes that occurred in the vegetation cover at 200- or 300-thousand-year snapshot intervals, one during the Early Pleistocene and another one later on, during the Mid-Pleistocene Transition . These changes concerned plant species and their respective abundance and appear to have been related to rainfall and temperature variability . The proportion of forest trees increased during wet episodes, whereas the influence of Afroalpine grassland indicators increased during cool and dry episodes. Variations in Early Pleistocene pollen data from Melka Kunture at ~1.8–1.6 Ma are consistent with isotopic evidence of precession variability as recorded at Olduvai and Turkana archaeological sites at ~2–1.8 Ma. For the Mid-Pleistocene Transition , variations in pollen data seem to match the climatic variability of isotopic and long pollen records from the Mediterranean region, notably upon the onset of dominant 100 ka-long glacial/interglacial cycles.
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others

Keywords
- Melka Kunture
- Early Pleistocene
- Mid-Pleistocene Transition
- Pollen
- Mountain forest history
- Hominin adaptation
5.1 Introduction
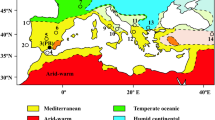
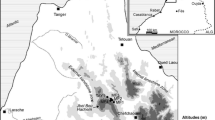
Melka Kunture is known for its extensive archaeological and paleontological record, spanning over most of the Early, Middle, and Upper Pleistocene (Chavaillon and Piperno 2004). Located in Ethiopia 50 km southwest of Addis Ababa, at 2000 m asl, in the Upper Awash valley and on the border of the Northern Ethiopian plateau, this cluster of archaeological sites is one of the few known mountain habitats of early hominins (Fig. 5.1). It is situated at a slightly lower altitude than the Gadeb site in the Bale Mountains (Southern Ethiopia), unfortunately inaccessible since it was flooded after the construction of a dam (Clark and Kurashina 1979; de la Torre 2011). The Melka Kunture sites yield evidence of Oldowan assemblages at ~1.7 Ma (Gallotti and Mussi 2015) and record the development of the Acheulean starting from its emergence at ~1.6 Ma (Gallotti 2013; Gallotti and Mussi 2018). Its location in the Ethiopian highlands is unlike that of other Acheulean sites in East Africa: those in the Ethiopian Rift , at 500 m asl (de Heinzelin et al. 2002); around Lake Turkana in Kenya (Isaac and Isaac 1997; Roche et al. 2003; de la Torre 2004; Lepre et al. 2011); and around Lake Natron as Peninj in Tanzania (de la Torre et al. 2008). Other Acheulean sites such as Olduvai in Tanzania (Leakey 1971) and Konso in southwestern Ethiopia (Beyene et al. 2013) are situated at medium altitude, i.e. at 1500 m asl.

Location map of Melka Kunture archaeological site in the Upper Awash valley, on the border of the Rift Escarpment, Ethiopia
Past vegetation is one of the environmental features that may help us understand the emergence of technological changes in lithic productions (Gallotti et al. 2010) and the long-lasting occupation of the Ethiopian highlands. Isotopic analysis and Carbon δ C13 measurements on soil carbonates or organic matter provide information on the proportion of C3 versus C4 plants in the vegetal cover, a ratio that can be interpreted as an estimate of tree cover density in past vegetation (Cerling et al. 2011; Magill et al. 2013). However, at high altitudes the C3 signal may also reflect a certain proportion of C3 grass. Moreover isotopic analyses do not yield knowledge regarding the botanical identification of the plant themselves, a critical information that is necessary for assessing vegetation type, hominin diet (Teaford 2000; Goren-Inbar et al. 2007, 2014) or habitat. Pollen grains are distinctive of the plants that produce them and provide such botanical precision. In this paper, we present and discuss fossil pollen data linked to the discovery of two hominin fossils: a massive humerus of Homo sp. found at ~1.6 Ma in level Gombore IB with early Acheulean lithic productions (Chavaillon et al. 1977; Di Vincenzo et al. 2015; Gallotti and Mussi 2018) and cranial fragments of Homo cf. heidelbergensis associated with the middle Acheulean of Gombore II -1, which is slightly younger than 0.875 Ma (Chavaillon et al. 1974; Chavaillon and Coppens 1975, 1986; Profico et al. 2016). Within this time interval, fossil pollen data were obtained from 15 distinct stratigraphic layers, that are partly associated with Oldowan and Acheulean lithic implements. They are centered around two critical periods: 1.8–1.6 Ma and 0.9–0.6 Ma, and there is additional information at ~1.4 Ma.
The sites are named after the gully or valley where they are located, for example Garba (or Gombore or Kella), followed by a Roman numeral that refers to the order in which they were discovered (e.g. Gombore II was discovered after Gombore I). At Gombore II, two distinct archaeological horizons mentioned in this paper are named Gombore II-1 and Gombore II-2, respectively. KII and KIII refer to geological sections in the Kella valley, as reported in Taieb (1974).
5.2 General Background
Melka Kunture (8°42 N; 38°36 E) is located in the tropical zone, but at high altitude, a geographical environment which modulates climatic conditions. At 2000 m asl, the maximum temperature averages 20° and the minimum one 8°C. Large diurnal temperature variations and nighttime frost may occur. The mean annual rainfall is about 860 mm at Boneja, the nearest meteorological station, 20 km north of Melka Kunture. There is great annual variability in the 1974–2010 record, with a minimum value of 388 mm, and a peak of 1419 mm. Rainfall distribution is usually bimodal, with a short rainy season in spring and long summer rains from June to September. Monsoon rainfall is primarily controlled by the seasonal migration of the Inter Tropical Convergence Zone (ITCZ) which lies north of Ethiopia at that time. The dry season is from October to February when the ITCZ lies south of Ethiopia. The northeasterly winds that blow in winter carry little moisture from the Red Sea and are dry when they reach central Ethiopia. The short rainy season is from March to May, when the ITCZ moves from south to north.
5.2.1 Vegetation
In the upper Awash valley of central Ethiopia, the archaeological area is surrounded by pastoral and intensely cultivated land that leaves very small patches of natural vegetation. In this region, agriculture developed thousands of years ago (Terwilliger et al. 2011). After a pastoral Oromo migration, in the second half of the sixteenth century, cereal cultivation was replaced by pastoralism and abandoned areas turned into bushland (Zerihun and Backéus 1991). However, the “potential vegetation” is mapped by botanical studies as a unit of the “Dry evergreen Afromontane forest and grassland complex ” (DAF) (Friis et al. 2010). DAF vegetation, widespread on the Ethiopian plateaux between 1800 and 3000 meters, includes mountain forests, woodlands, wooded grasslands and grasslands. It is a complex unit where the various vegetation types are characterized by heterogeneous distribution of trees or shrubs and herbaceous plants, but include the same botanical components. The Afromontane vegetation type is clearly distinguished from that of lowland savanna or woodland by the botanical attribution of characteristic plants, notably trees. It is defined by the presence of tall tree species such as Juniperus procera (Cupressaceae ), Podocarpus falcatus (Podocarpaceae ), Olea europea (Oleaceae), Croton macrostachys (Euphorbiaceae) associated with smaller trees such as Celtis africana (Ulmaceae) and with shrubs among which Myrsine africana (Myrsinaceae) frequently occurs. Mountain woodland , wooded grassland , and grassland are DAF subtypes and include many species of Acacia (Fabaceae, subfamily Mimosoideae) together with many Maytenus obscura (Celastraceae) and several species of endemic Echinops (Asteraceae). At its lower altitudinal limit, a drier type of vegetation marks the transition between Afromontane vegetation and the lowland vegetation described as Acacia-Commiphora bushland . Scattered Juniperus procera may occur, sometimes associated with Dracaena , Barbeya oleoides (Barbeyaceae), Tarchonanthus camphoratus (Asteraceae), or Dodonaea angustifolia (new name for Dodonaea viscosa) (Sapindaceae) and with various species of Euclea (Ebenaceae ). The vegetation zone located at 1600 m asl, like the one around the lakes in the Rift, is occupied by Combretum-Terminalia (Combretaceae) woodland (CTW), and by wooded grassland (Friis et al. 2010). Above the Afromontane forest, at ~3000 m asl, the vegetation consists of Hagenia abyssinica - Hypericum revolutum woodland , overlapping in part with an Ericaceous bushland dominated by Erica arborea ; Asteraceae and Artemisia abound in the Afroalpine grassland up to 4000 m asl (Friis 1992; Friis and Sebsebe 2001). This altitudinal distribution of modern vegetation is observed along the slopes of the Wochacha volcano, some 20 km away from Melka Kunture (Fig. 5.1). The succession of woodland , mixed forest and juniper-dominated forest, followed by remnants of the Afroalpine bushland on the top, reflects variations in ecological conditions along the altitudinal gradient. Between 2200 and 3200 m the mean annual temperature decreases by about 5 °C to 6 °C, whereas rainfall increases up to 3000 m (upper cloud limit) and then decreases above the cloud forest in the Afroalpine vegetation. Pollen analysis of present-day surface soil samples, collected under different types of vegetation still preserved on the slopes of the Wochacha and Zuqualla volcanoes, clearly indicates several pollen markers of altitudinal vegetation zones (Fig. 5.2). Grass pollen dominates in the lowermost woodland . Dodonaea shrubs and Acacia indicate the transition zone, while the forest is indicated by abundant Olea and Juniperus pollen . Ericaceae and Plantago are good markers of the Afroalpine vegetation.

Distribution of selected pollen markers in pollen analysis of soil samples collected in modern vegetation zones encountered along an altitudinal gradient on slopes of the Wochacha and Zuqualla volcanoes. New name for Dodonaea viscosa is Dodonaea angustifolia
5.2.2 Geology, Chronology, and Archaeology
The Melka Kunture sedimentary basin lies on the shoulder of the northern Ethiopian plateau, near the western margin of the active Wonji fault belt that marks the axis of the Ethiopian Rift (Mohr 1971). Outcrops of Pleistocene deposits exposed on both banks of the Awash River hardly amount to a total combined thickness of 40 meters. Silt, sand, clay, and coarse layers inter-bedded with tephra falls and tuffaceous clay were deposited by the Awash River and its tributaries. Fluvial sedimentation proceeded mainly in an alluvial braided context influenced by the deposition of volcanic ashes and lavas. The fluvial-volcanic succession accumulated on top of the “basal clays”, deposited when a shallow body of water had developed in the basin. The “basal clays” were initially measured by geophysical echosounding and found a few meters thick near the mouth of the Garba gully (Taieb 1974). A normal paleomagnetic signal places their deposition at the end of the Olduvai subchron, with an upper limit at 1.778 Ma (Tamrat et al. 2014). Recent fieldwork enabled additional observation of the basal clays in the Kella tributary valley. Evidence of erosion at the top of the clays was mentioned in Jean Chavaillon’s archives and observed again recently, when water level was low, along the Kella tributary close to its confluence in the Awash River. The earliest volcanic deposit (tuff sample 27-17) capping the basalt clays was dated at 1.874 ± 0.012 Ma (Morgan et al. 2012). Accordingly, we consider that the extant, eroded basal clays are related to the lower part of the Olduvai paleomagnetic event. The earliest dated tuff provides at ~1.9 Ma the lower limit for the archaeological and paleontological record of Melka Kunture.
Soon after the area’s archaeological potential was recognized (Bailloud 1965), exploration surveys and extensive excavations unearthed abundant Oldowan and Acheulean artifacts, as well as later lithic productions (Chavaillon 1971, 1979), whereas geological studies (Taieb 1974) were undertaken together with pollen sampling (Bonnefille 1972). Discontinuities in the sedimentary record, some indicated by erosion surfaces, as well as the lack of easily recognizable marker beds, made it difficult at first to establish stratigrafic correlations between exposures outcropping in different gullies, partly covered by abundant bushy vegetation. Initially, the succession of Oldowan, Acheulean, Middle Stone Age, and Late Stone Age lithic productions helped place the local stratigraphy within the Pleistocene through typological comparison with dated industries from Olduvai (Chavaillon 1973, 1980; Chavaillon and Piperno 2004). An estimated time range from at least 1.5 Ma to the last ten thousand years was supported by early paleomagnetic measurements (Westphal et al. 1979; Cressier 1980) until 40Ar/39Ar dating provided a more precise chronological framework (Morgan et al. 2012). With the help of new fieldwork and recent archaeological investigation (Gallotti et al. 2014; Mussi et al. 2014, 2016), formerly collected pollen samples are now securely placed in this new chronology.
Initial sampling for pollen was done in the sedimentary sequences along three gullies: Garba (G) and Gombore (Gomb) which are tributaries of the Awash River on the right bank, and mostly dry all year long, and Kella (K) on the left bank (Fig. 5.3). A few more samples were collected directly on excavated archaeological layers. Accordingly, they are numbered from top to bottom as it is usual in archaeological practice. The excavated archaeological localities are Gombore I (level C and overlying level B) and Garba IV (levels E and F and overlying level D). The pollen data discussed in this paper are constrained by 40Ar/39Ar dating and tephrochronology in a time interval starting at ~1.8 Ma and ending before 0.709 ± 0.013 Ma (Morgan et al. 2012), except for one sample which is more recent. Although initially collected at more or less regular depth interval throughout the sedimentary sequences, in light of current fieldwork the stratigraphic positions of the pollen samples appear to be grouped in two time periods. In the Early Pleistocene sequence, the pollen data correspond to the time of the late Oldowan and emerging Acheulean (1.8–1.6 Ma); during the Mid-Pleistocene Transition (0.9–0.6 Ma), they document past vegetation of the middle to final Acheulean .

Melka Kunture, location map of the litho-stratigraphic sections and archaeological localities mentioned in the text
5.3 Pollen Data from the Early Pleistocene, ~1.8–1.6 Ma
Past vegetation in the 1.8–1.6 Ma period is documented by eight pollen samples collected in deposits at the base of stratigraphic successions in the Kella, Garba, and Gombore gullies. Intercorrelation between the three sections relies on current research that focuses on volcanic material.
5.3.1 Chronostratigraphic Placement of the Pollen Samples
Four samples come from two sections, Kella II and Kella III, which are exposed 500 m apart from each other in the Kella valley (Fig. 5.3). Three of them were collected in a three-meters sedimentary sequence outcropping on a cliff at Kella III. From bottom to top, sample K263 was extracted from clayey sands with obsidian gravels, K265 from sands and K269 from cross-bedded sands (Fig. 5.4). The Kella III sequence is overlaid, 2 m above K269, by a tuff which, based on stratigraphic correlation of the outcrops along the Kella valley, corresponds to tuff layer MK27-18, dated 1.666 ± 0.009 Ma, which lies a few hundred meters downstream (Morgan et al. 2012). At Kella II, also called Butte Kella, pollen sample K242 comes from the infill of a bone extracted from a sand layer capping this same dated volcanic ash (Bonnefille 1968; Taieb 1974). Therefore K242 is slightly younger than 1.666 Ma whereas K263, K265, and K269 are older (Table 5.1). The stratigraphic position of these samples is well constrained by the 1.666 ± 0.009 Ma dated tuff intercalated between samples K269 and K242.

Stratigraphic position and correlation between the Early Pleistocene (1.8–1.6 Ma) pollen samples collected at Kella , with respect to dated volcanic deposits. *The dated sample was collected 80 m downstream of Kella III
Further chronological constraint is provided by Early Pleistocene deposits that are likewise exposed above the basal clays in the Garba and Gombore gullies near the Awash River, two km downstream (Fig. 5.3).
Two pollen samples from the excavated archaeological horizons Gombore IC and Gombore IB are separated from each other by an interval of few dozen centimeters (Chavaillon and Berthelet 2004). Archaeological layer Gombore IC lies 10–15 cm below a tuff consisting of ryolithic ashfall material that showed an elementary composition “quasi-identical” to the “Grazia Tuff” (Raynal and Kieffer 2004, p. 152), dated at <1.719 ± 0.199 Ma at the Garba IV archaeological site (sample 27-23 in Morgan et al. 2012) (Fig. 5.5). Layer Gombore IB, 20 cm above the local ryolithic tuff, is considered to be ~1.6 Ma (Gallotti and Mussi 2018).

Stratigraphic position and correlation between the Early Pleistocene (1.8–1.6 Ma) pollen samples collected at Gombore and Garba, with respect to dated volcanic deposits
In the Garba gully, the lowest and earliest Oldowan horizons, i.e., Garba IVE and Garba IVF, are older than the <1.719 ± 0.199 Ma “Grazia tuff ” (Piperno et al. 2009; Morgan et al. 2012; Gallotti and Mussi 2015) (Fig. 5.5). Pollen samples were not collected either from layer Garba IVF, or from layer Garba IVE. This is unfortunate since the latter layer yielded a hominin child mandible, tentatively attributed to Homo erectus s.l. (Condemi 2004; Zilberman et al. 2004). We have no palynological samples from archaeological horizon Garba IVD, which caps the <1.719 ± 0.199 Ma tuff and contains implements previously attributed to the Oldowan (Chavaillon and Piperno 2004) and now to the earliest Acheulean of Melka Kunture (Gallotti 2013). However two pollen samples had been collected (Bonnefille 1972) from an exposed section later described by Kieffer et al. (2002, Fig. 21, p. 89). Pollen sample G393 extracted from a manganese clay layer with embedded obsidian artifacts corresponds well to archaeological horizon Garba IVD. Sample G133 from a fine cross-bedded sand with bones 80 cm above this horizon is equivalent to subunits 3.4 or 3.5 in Kieffer et al. (2002, Fig. 21, p. 89). This sample is much older than tuff 27-19 dated 1.429 ± 0.029 Ma and located 4 m higher up in the stratigraphy (Morgan et al. 2012).
The <1.719 ± 0.199 Ma “Grazia tuff ” at Garba IV and the undated ash layer from Gombore I , which are related to the same volcanic activity, are used as a marker bed for correlating the deposits in the two sections. The ~1.66 Ma tuff at Kella III, in turn, is correlated to this marker bed by geostratigraphy. All the samples detailed above were collected in close stratigraphic proximity to each other and are considered to date from ~1.7 Ma. Although the time span cannot be estimated exactly, no more than about two hundred thousands years separate the oldest K263 from the youngest G133 (Table 5.1).
5.3.2 Pollen Results from the Early Pleistocene, ~1.8–1.6 Ma
The eight samples listed in Table 5.1 provided abundant pollen that document past vegetation around Melka Kunture during the Early Pleistocene . A total of 58 pollen taxa have been identified; 32 of them are trees and 8 to shrubs and climbers. The others are from herbs, aquatic plants, and ferns (Table 5.2). The great number of tree species indicates a highly diversified woody vegetation, although the high proportion of grass pollen (Poaceae) points to a local abundance of herbaceous plants (Fig. 5.6).

Percentage distribution for selected pollen taxa identified in samples of the Early Pleistocene (1.8–1.6 Ma), with reference to the corresponding archaeological levels and the proposed interpretation of past vegetation. Relative pollen % are calculated versus the total pollen sum. Pollen not represented in this graph are found in Table 5.2. Abbreviations: G for Garba, Gomb for Gombore, K for Kella . New name for Dodonaea viscosa is Dodonaea angustifolia
The oldest K263 pollen spectrum (n = 255) shows an unusual distribution of pollen among Asteraceae, Resedaceae and Fabaceae (including Rhynchosia) and Acacia, mostly insect pollinated plant taxa. No proportion of Acacia as high as 40% is known from any of the many soil samples analyzed from different vegetation types in tropical Africa (Bonnefille 2007). However, the association of Acacia tortilis and Fabaceae among trees, together with Caylusea, an herb of the Resedaceae family, characterizes the vegetation along wadies on the Tibesti slopes in the Sahara (White 1983). Particular events such as instantaneous ash flooding of the Kella River may have concentrated pollen from plants growing along streambed before the ashfall.
The next K265 sample (n = 406) contains a few more pollen taxa than those found in the previous sample. The abundance of Asteraceae pollen together with the occurrence of Resedaceae and the increase in Fabaceae are noteworthy. The abundant Asteraceae pollen includes Tarchonanthus, a tall shrub widespread in Africa (White 1983) and abundant in dry Juniperus-Olea woodlands or forests, which are common in southern Ethiopian highlands close to Somalia (Friis 1992). This interpretation would be consistent with the finding of a single grain of Olea and of Piliostigma , whereas there is a much smaller amount of Acacia pollen, and a much lower proportion of Poaceae.
Pollen assemblage K269 (n = 212) shows abundant and diversified tree pollen taxa, while Asteraceae pollen is less abundant. Among the tree and shrub taxa, the occurrence of Combretaceae, Euclea , Dodonaea angustifolia, Podocarpus, Olea africana, and Myrsine africana attests to a more diversified type of evergreen bushland with some forest trees. The abundance of a pollen type attributed to Ziziphus (Rhamnaceae) indicates a woody riparian vegetation, confirmed by the abundance of Achyranthes aspera , a shade-loving herb, frequent on river banks. The percentages of grasses increase. A few additional pollen grains extracted from another uppermost sample confirm the occurrence of Celtis, Syzygium, Trilepisium (Table 5.2) associated with Ricinus communis and indicate enriched wooded riverine vegetation.
Pollen sample K242 (n = 331) includes a large number of pollen taxa. Among them there are various tree taxa, the majority of which can be attributed to the mountain dry forest or evergreen bushland , whereas a few of them belong to a riparian forest (Bonnefille 1968). However, the high percentage of grass pollen and the occurrence of herbs attributed to Plantago , Brassicaceae (former Cruciferae) or Apiaceae (former Umbelliferae) attest to the opening-up of the tree cover, interpreted as mountain grassland. At the headwaters of the Atebella, a left tributary of the Awash River, the Wochacha volcano, already formed 4 Ma ago (Chernet et al. 1998), rises to 3400 m asl (Fig. 5.1). At about 20 km from Melka Kunture, its slopes show a succession of vegetation types at different altitudinal zones. The various vegetation provided pollen input in sediments deposited at Melka Kunture. There were also plants such as Arecaceae (former Palmae) (15%) and Encephalartos (10%) that belongs to the botanical family Zamiaceae of the Cycadae order, among the Gymnosperms. Encephalartos pollen is distinguished by its large size and elongated shape (30 to 40 μm), smooth exine, and invaginated sulcus (Fig. 5.7). The pollen does not disperse very well, like in the case of palms. This fossil pollen was compared to modern pollen of several known African secies and tentatively attributed to Encephalartos hildebrandtii (Bonnefille 1975). The genus Encephalartos is native to Africa and several of its species are found in South Africa, Congo, Tanzania, Kenya, Sudan, etc. Commonly known as “bread palm” because a bread-like food can be prepared from its pith, E. hildebrandtii grows in the Usambara mountains in Tanzania. Associated with Brachylaena, it was an important component of dry woodlands near the coast, north of Lamu, in the Lunghi forest of Kenya (Pichi-Sermolli 1957). A few isolated Encephalartos tree have been described on remote hills, for instance at Ololokwe, in central Kenya. Therefore, the fossil Encephalartos pollen recorded at Melka Kunture may have come from a relic of Pliocene vegetation whose geographic distribution once extended to Ethiopia, north of its present-day range. In sample K242, a larger proportion of Encephalartos is accompanied by pollen from palm trees, associated with Olea, Hagenia abyssinica , Pistacia, Psychotria , Polyscias, Loganiaceae, Myrsine africana , and Dodonaea angustifolia; all of them are components of drier types of mountain forest and bushland . We conclude that 1.7 Ma ago a diversified type of dry forest, including Syzygium guineense , Podocarpus and Myrica salicifolia , Combretaceae and the dry Sinopteridaceae ferns, grew not far from Melka Kunture. However, the percentage of grass pollen (>50%) remains high, indicating that extensive areas were still occupied by mountain woodland or dry forest, with various herbaceous plants such as Plantago , Dyschoriste , Apiaceae, Brassicae (Cruciferae) in large clearings near Melka Kunture.

Modern and fossil pollen . 1–5: Encephalartos bubalinus; 6: optical section of Phoenix reclinata (Arecaceae) for comparison showing columella in the exine; 7–13: Encephalartos hildebrandtii, showing an invaginated sulcus; 12 and 13 SEM photographs: 12 (×2000) and 13 (×5000) showing granulated sulcus membrane: 14–23: fossil pollen grains extracted from Melka Kunture Pleistocene sample K242, and related to Encephalartos hildebrandtii. Encephalartos pollen is distinguished from that of Cycas by a thicker exine membrane and granulations visible on the proximal side. The asiatic genus Gingko shows an undulated border of the sulcus
The number of pollen grains (n = 140) extracted from archaeological layer Gombore IC is smaller than in previous samples, hence a smaller number of plant taxa have been identified. Among trees, abundant Podocarpus is accompanied by small amounts of pollen from Juniperus and from some shrubs, including Dodonaea angustifolia and the climber Clematis . The herbaceous component is dominated by grasses (61%), with Plantago and Apiaceae of open grassland and the aquatic Typha . Most of these pollen are known to disperse widely, a phenomenon that leads to pollen deposition on sub-aerial surfaces rather than in fluvial deposits. The vegetation of the area was an open grassland. Few forest trees grew not far away from the site.
Pollen sample Gombore IB (n = 456) yielded more substantial results. It contains a greater amount of Juniperus (>20%) associated with Podocarpus, Olea, Polyscias, Hypericum, and Myrica; all of these plants are mostly encountered in highland forest at altitudes far above 2800 m. The percentage of arboreal pollen is as high as 27%, indicating that the dry conifer forest was close to Melka Kunture at that time, because Juniperus does not disperse its pollen very far. The proximity of bushland or forest was later confirmed when a wood fragment identified as Caesalpinioxylon was recovered from the excavated surface of Gombore IB (Chavaillon and Koeninger 1970). A juniper-dominated forest whose composition was similar to that of the modern Managasha forest expanded from the northern slopes of the Wochacha volcano toward Melka Kunture. The large proportion of grass pollen (Poaceae 63%) indicates that this forest was broken up by large open spaces in the vegetation, probably near the river. A great variety of grass species characterizes mountain grasslands . We are unable to distinguish among them because the pollen morphology is uniform throughout the various genera of the Poaceae family. However pollen from different botanical tribes is grouped in size classes. The fossil pollen is more abundantly distributed in two of the size classes. Pollen from Eragrostideae and Chlorideae grasses is in the 25–35 µm class, whereas Andropogoneae pollen is larger (35–40 μm; Bonnefille and Riollet 1980a). The size distribution of grass pollen measured in the Gombore IB sample ranges from 15 to 55 μm. The pattern of grass pollen size distribution in this fossil sample is similar to the pattern in a modern soil sample collected in the evergreen juniper open woodland at 2600 m asl (Fig. 5.8). This similarity confirms that the early Acheulean site was indeed surrounded by this type of vegetation.

Measurements of size of grass pollen grains: A in a high-elevation modern sample; B in the fossil sample from the early Acheulean horizon Gombore IB
In the Garba gully, the two pollen samples collected near archaeological layer Garba IVD and near the ~1.7 Ma “Grazia tuff ” are not rich in pollen . Counts on the order of one hundred grains provide a rough estimate of tree cover density, but not enough details on the diversity of tree species. Pollen assemblage G393 (n = 111) is dominated by grass pollen (86%) indicating a high-elevation grassland in which Plantago and various species of Asteraceae were present together. Remnants of mountain forest are attested to by a single pollen grain of Podocarpus accompanied by Myrsine africana , a shrub that most likely grew nearby, since its pollen does not disperse over long distances, unlike that of Podocarpus. Pollen sample G133 (n = 113) shows a high percentage of grass pollen (76%), and only four tree species. The total arboreal percentage is 12%, indicating the proximity of wooded riparian vegetation characterized by the presence of the highland riparian Myrica salicifolia associated with Celtis and Trema , and attesting to the deciduous character of the vegetation in seasonal climate conditions.
In conclusion, during this 200 ka-long period, the vegetation surrounding Melka Kunture underwent significant changes. They are reflected in the pollen composition by variations in the total abundance of tree taxa and changes in the predominance of different species. At ~1.7 Ma, we note the appearance of the modern Afromontane character of all the trees that belong to present-day mountain vegetation that grows from 2000 to 2800 m asl.
5.3.3 Pollen Data of the Early Pleistocene, ~1.4 Ma
One pollen sample, G101 (n = 111), comes from a sandy silt layer overlying a consolidated Mn Fer crust in the Garba gully, ~3 m above Garba IVD level. Its stratigraphic position is below volcanic ash 27-19 dated <1.429 ± 0.029 Ma, validated as a minimum age (Morgan et al. 2012) (Fig. 5.5). Another volcanic ash (ash 27-16), higher up in the stratigraphy, is dated to 1.037 ± 0.088 Ma. The pollen spectrum is dominated by grasses (77%) and includes Plantago , an herbaceous plant that produces abundant pollen in mountain grasslands . The presence of Achyranthes, a shade-loving erect herb abundant on river banks, together with more spores from Pteridophytae (3 distinct types), points to local humidity conditions that encouraged the growth of ferns. The surrounding vegetation can be interpreted as mountain woodland and wooded grassland . A few Podocarpus, Euclea , Dodonaea angustifolia, Syzygium, and Acacia trees were also present
5.4 Pollen Data of the Early/Mid-Pleistocene, ~0.9–0.6 Ma
Six pollen samples document past vegetation between ~0.9 and ~0.6 Ma. They were collected from exposed sediments along the Garba (G) and Gombore (Gomb) gullies, where several archaeological levels have been excavated.
5.4.1 Chronological Placement of the Pollen Samples
Four samples were collected in good stratigraphic succession from the westernmost part of the Garba gully, in a two-meter section atop a gray blue volcanic ash layer (“cinerite 106” in Bonnefille 1972; Taieb 1974). We correlate this cinerite with the tuff dated 0.869 ± 0.02 Ma in the Garba gully (sample 27-13 in Morgan et al. 2012) (Fig. 5.9). Sample G372 was extracted from the medullar cavity contents of a bone fragment found in the sand-and-gravel bed that contains some Acheulean stone tools, and collected at the lower contact with the dated tuff. Pollen sample G136, extracted from the medullar infill of another long bone fragment, came from the cross-bedded yellow sand lying 100 cm above the tuff, hence it is significantly younger than 0.869 Ma. Pollen sample G371 was taken from the sand-and-gravel layer outcropping 150 cm above the tuff, hence it too is younger than G136. The last pollen sample of this group, G110, comes from the 4-m-thick tuffaceous clay that forms an abrupt cliff 200 cm above the dated tuff.

Stratigraphic position of the Early/Mid-Pleistocene (0.9–0.6 Ma) pollen samples collected at Garba, with respect to a dated volcanic deposit (redrawn after Bonnefille 1972, Fig. 37)
Two other samples come from the Gombore gully, situated a hundred meters west of the Garba gully (Fig. 5.3). This section has two main archaeological horizons, bracketing the Early/Middle Pleistocene boundary (Fig. 5.10). Pollen sample Gomb173 was collected at the base of a silty–sandy deposit, 2.5 m above Gombore II -1, a middle Acheulean archaeological site (Gallotti et al. 2010) overlying a tuff dated at 0.875 ± 0.010 Ma (sample 27-08, Morgan et al. 2012). We correlate this tuff with the one dated at 0.869 ± 0.02 Ma in the Garba gully, a result coincident within 1σ. The second main archaeological horizon, nearly 1 m above sample Gomb173, is Gombore II -2, the so-called “Hippo Butchery Site” (Mussi et al. 2016), which is capped by a volcanic ash dated 0.709 ± 0.013 Ma (sample 27-09, Morgan et al. 2012). Pollen sample Gomb173, which lies between Gombore II -1 and Gombore II -2, and below the Matuyama-Brunhes boundary , as identified by Tamrat et al. (2014), is slightly younger than 0.8 Ma.

Stratigraphic position of the Early/Mid-Pleistocene (0.9–0.6 Ma) pollen samples collected at Gombore, with respect to dated volcanic deposits. The log is the outcome of the excavations carried out at Gombore since 2012, which do not include Gombore III. Gombore III has been relocated above the uppermost part of the sequence illustrated in the figure
Pollen sample Gomb349 was extracted from the infill sediment of a long bone fragment collected in a clayish tuffaceous deposit above Gombore II -2 (Mussi et al. 2016). Its estimated age is close to ~0.6 Ma.
The six pollen samples span more than 200 ka (Figs. 5.9 and 5.10).
5.4.2 Pollen Results, ~0.9–0.6 Ma
The results of pollen counts obtained from the six pollen samples included in the 0.9–0.6 Ma period are shown in Table 5.3. Among the 71 recognized pollen taxa, 39 (55%) belong to tall trees and 9 (12%) to shrubs or climbers, a proportion indicating highly diversified arboreal and forb components in the past vegetation. Identification of the pollen indicates that all the taxa belong to the Dry Afromontane forest, i.e. they are currently encountered in the vegetation of the Ethiopian highlands (Fig. 5.11).

Distribution of percentages for selected pollen taxa identified in samples dated ~0.9–0.6 Ma corresponding to the MPT period, related to archaeological evidence and to the proposed interpretation of past vegetation. Relative pollen % calculated as in Fig. 5.5. Pollen not represented in this graph are indicated in Table 5.3
The rich pollen spectrum G372 (n = 1084) is dominated by grasses (93%). Plantago is associated with Artemisia , a small shrub abundant today in the Afroalpine grassland above the forest limit at ~3000 m asl. A few forest tree taxa are nevertheless present: Myrica salicifolia and Salix from riparian vegetation encountered today at higher altitudes, above 2500 m asl. At ~0.9 Ma, the vegetation surrounding Melka Kunture can be reconstructed as an Afroalpine grassland with scattered forest trees.
G136 is an abundant pollen spectrum with a total pollen count close to 1000 pollen grains and a great number of identified pollen taxa. There is still a high percentage of grasses (84%), although it is smaller than in sample G372. The tree taxa are more numerous and more diversified. The shrubs, including Dodonaea angustifolia, Rumex , Jasminum , Rhus , and Myrsine africana indicate a dense evergreen bushland . The increasing proportion of Podocarpus, and the occurrence of Juniperus, Olea, and Rutaceae, indicate that the dry mountain forest was more diverse and closer to Melka Kunture. Acacia associated to Faidherbia albida (syn. Acacia albida), a tall Legume tree, may have been present locally (Bonnefille 1975). The landscape was more wooded and richer in species including Euphorbia and Rubiaceae. Typha (cattail) and Phragmites (reed) occupied locally humid flatlands surrounding quiet freshwater ponds, possibly pointing to wetter climate conditions.
The pollen results from sample G371 (n = 421) show an increase in the total arboreal pollen (22%) in which Juniperus becomes more abundant (6.5%). The number of tree pollen taxa is considerable, including Ulmaceae associated with Apocynaceae, Pygeum and Celastraceae whose pollen does not disperse over long distances. There is less pollen attributed to Acacia. Pollen taxa from herbaceous plants include Labiatae and Cruciferae together with Ricinus communis . Sedges decrease. Taken as a whole the pollen results indicate that a different type of diversified forest/woodland with dominant Juniperus had developed near Melka Kunture.
Pollen spectrum G110 (n = 718) contains a great number of pollen taxa although the total of arboreal pollen has decreased (15%). Juniperus is not present. The beautiful tree Hagenia abyssinica , well known in Ethiopia as Kosso, is found together with Anthospermum , a common herb of high-elevation grasslands. The notable proportion of Asteraceae (4.6%) is consistent with an Afroalpine vegetation that now occupies areas above the forest line. Pollen from Legume trees such as Caesalpiniaceae (2%) and Acacia are associated with Combretum, trees or climbers normally abundant in deciduous vegetation growing in climates characerized by seasonal rainfall. The pollen found in this deposit attests to types of vegetation that grow in drier and colder climates. Highland forest and local deciduous woodland existed somewhere whereas freshwater swamp vegetation including Typha and sedges was locally present.
Gomb173 provided a rich pollen spectrum (n = 573). Pollen from trees and shrubs accounts for 10% of the total. Almost 20 shrubs or trees taxa have been identified, thus attesting to high diversity among forest trees. Juniperus pollen is present but not abundant. We note the occurrence of Maesa, a common tree in the Shewa forests and of Ericaceae, the mountain heath. More abundant are Ebenaceae , Croton, Pterolobium , together with Acacia and Carissa edulis , associated with the shrubby Heteromorpha of the Apiaceae (Umbelliferae) family. These finding point to a dense and diversified woody bushland or shrubland in the area. Asteraceae pollen representing several species accounts for 67% of the total count and dominates the herbaceous pollen (75%), whereas grass pollen amounts to only 15%. This finding is consistent with an attribution to the evergreen bushland that is normally found above the forest belt and points to a climate that has become significantly drier and colder than it was before.
Gomb349 pollen spectrum (n = 565) at ~0.6 Ma, belongs to a late phase of the Mid-Pleistocene Transition (MPT). It contains a rich and diversified proportion of 28 tree and shrub taxa. Most of the trees are the same as in sample Gomb173; the presence of Olea points to warmer climatic conditions. With Sapium, among the Sapotaceae, Fagaropsis among the Rutaceae, Diospyros among the Ebenaceae and Osyris (Santalaceae), a tall herb, the pollen composition indicates a diversified forest with a dense canopy of trees and shrubs (17.5%).
During the Mid-Pleistocene Transition , at Melka Kunture past vegetation remains permanently in the Dry evergreen Afromontane Domain already established during the Early Pleistocene. All the plant taxa are of mountain vegetation. However, the six pollen assemblages documenting two-three hundred thousand years from ~0.9 to ~0.6 Ma show important changes in the floristic composition of past vegetation, notably in the relative abundance of various forest trees pointing to the influence of warmer or wetter conditions, while some others indicate slightly cooler conditions.
5.5 Discussion
With the exception of Encephalartos in samples K265 and K242 (Fig. 5.7), all the fossil pollen taxa refer to plants that are present today in the flora of Ethiopia (Friis 1992). A number of modern pollen taxa such as Astragalus, Bridelia , Cassia , Commelinaceae , Crotalaria , Diospyros , Drognetia , Galiniera coffeoïdes , Gardenia , Kohautia aspera , Malvaceae , Pterocarpus , Rubia cordifolia and Sterculia , all of which were identified in a mud sample collected in 1968 from the Awash River (Bonnefille 1969), are not found in the fossil pollen assemblages (Tables 5.2 and 5.3). However, since most of these plant taxa are herbaceous plants or are represented in the assemblages by only one grain, their absence from the fossil record is not statistically significant and cannot be regarded as indicating that during the Pleistocene plant diversity was lower than it is today.
5.5.1 Afromontane Character of the Past Vegetation at Melka Kunture since 1.8 Ma
In the overall fossil pollen counts (n = 2024 for 1.8–1.6 Ma; n = 4343 for 0.9–0.6 Ma) 114 different fossil pollen taxa were identified, including 70 that were not found in a modern sample from the Awash River (n = 2865; Bonnefille 1969). The fact that modern and fossil pollen assemblages do not overlap has two implications. First it confirms that the fossil assemblages were not contaminated by any modern pollen input, as can happen when rivers overflood. On the other hand, it indicates that vegetation was more diverse in the past that it is today. Indeed, the impoverishment of present-day vegetation is expected at Melka Kunture as the result of long-lasting human impact. Forests were already rare during the sixteenth century, when travelers wrote the first detailed descriptions of the landscape (Zerihun 1999). Nevertheless, except for Encephalartos, all fossil taxa are related to genera or species encountered somewhere in the modern vegetation of the northern Ethiopian highlands.
Of the 114 fossil pollen taxa identified in this study, 69 (60%) are attributed to trees, shrubs or climbers. This high number attests to a very diverse arboreal (woody) component of the past vegetation. Twenty-two fossil pollen taxa (18%) belong to herbaceous plants, whereas 23 others (22%) cannot be attributed to a well-defined plant form. Trees and shrubs taxa that belong to the montane floristic domain identified as the Dry evergreen Afromontane forest and grassland complex (DAF) dominate the pollen assemblages (~80%) (Tables 5.2 and 5.3). We conclude that highland vegetation as it is known today was already established in the Awash basin at ~1.8 Ma. Accordingly, we suggest that all hominins locally associated with the Oldowan and Acheulean had already adapted to mountain climate conditions.
5.5.2 Environmental Changes
In all samples, the percentages of grass pollen dominate because grasses produce large amounts of pollen. However, the relative abundance of each of the pollen taxa listed in Figs. 5.6 and 5.11 is subject to significant fluctuations. At Melka Kunture, Middle Pleistocene vegetation differs sharply from that of the Early Pleistocene. The Early Pleistocene has a variable but lower proportion of grass pollen , a higher proportion of arboreal pollen and greater diversity of trees and shrubs. Relict palms and cycadales are associated with « warm » taxa such as Acacia, Tamarix , Resedaceae . During the Mid-Pleistocene Transition , the pattern is different. More extensive grasslands and less tree cover are associated with a few « colder » Afroalpine grassland taxa such as Artemisia , Anthospermum , Erica arborea and Plantago .
In general, species diversity increases with total pollen counts and with the number of analyzed samples. The total pollen counts (n = 2024) for the Early Pleistocene (EP) is smaller than that of the six samples from the MPT (n = 4343). Therefore the greater number of identified pollen taxa recorded for the MPT (71 versus 58 for the EP) might not reflect greater plant diversity. However, in each period large variations are observed among the samples. Stratigraphic constraints suggest that such changes occurred at intervals on the order of a few tens of thousands of years.
5.5.3 Early Pleistocene Wet–Dry Variability (1.8–1.6 Ma)
A well-marked vegetation change is observed between the ~1.8 Ma Kella results on the one hand, and the ~1.6 Ma samples related to Garba IVD and Gombore IB archaeological sites on the other. Bushy woodland , possibly existing under warmer and drier conditions, differs sharply from the extended grasslands that dominated the landscape in the more recent samples from Garba IVD, at the time of the emergence of the Acheulean. Mountain forest trees and shrubs became more abundant at Gombore I ; a real Juniper woodland /forest grew near the Gombore IB archaeological site, close in age to Garba IVD (Gallotti and Mussi 2018), although locally open vegetation left room for an herbaceous cover and some periaquatic plants. Juniperus percentages did not reach the same values that they did under a forest cover such as prevails today in the Managesha forest, on the Wochacha volcano (Fig. 5.2). Our interpretation is that a real forest existed in the vicinity, but that locally at the site open space was covered by herbaceous vegetation. Conversely, during and after the early Acheulean occupation at Garba IVD (Gallotti 2013), mountain grassland expanded greatly. The presence of Plantago , Anthospermum , and Apiaceae indicate affinities with high-elevation grasslands, now found above the forest zone. This context explains the presence of wind-transported pollen of Podocarpus, frequently observed in the Afroalpine zone (Umer et al. 2007). In this cool grassland herbivores were present as shown by the fossil fauna, which also includes the highland rodent Tachyoryctes (Geraads et al. in press). The expansion of mountain grassland implies a significant lowering of vegetation belts explained by a colder and drier climate.
At Melka Kunture, environmental conditions differed sharply from those of the savanna existing elsewhere in the Rift Valley , in the warmer climate inferred at archaeological sites located at lower altitude. Nevertheless, vegetation changes documented in the Ethiopian northern highlands are contemporary to vegetation changes evidenced by previous pollen studies at Olduvai from ~1.8 Ma to ~1.6 Ma, during Bed I/lower Bed II times (Bonnefille and Riollet 1980a; Bonnefille et al. 1982). Olduvai is located at lower elevation in a warmer wooded grassland and woodland near a permanent salt lake (Barboni 2014). At this site, past vegetation changes also preceded and followed the emergence of the Acheulean, dated at 1.664 ± 0.019 Ma at the FLKW site (Diez-Martin et al. 2015). At both the Ethiopian and the Tanzanian sites, the availability of freshwater, the density of trees in mountain wooded grassland and the proximity of mountain forests provided various plant resources to hominins producing at first the Oldowan, and later the early Acheulean. However, at Melka Kunture, sedges, Typha or ferns were rare whereas at Olduvai the grassland surrounding a permanent salt lake and the periodically flooded adjacent marshes are thought to have produced abundant freshwater food (Magill et al. 2013) and a C4 diet exclusive to Paranthropus boisei (Ungar and Sponheimer 2011). The past vegetation of Melka Kunture did not provide abundant aquatic food, but probably did offer more resources from nearby forests, such as fruit trees.
5.5.4 Mid-Pleistocene Transition Variability
Pollen assemblages between ~0.9 and 0.6 Ma are more homogenous, with a proportion of grass pollen exceeding 70%, except in one sample characterized by overrepresentation of Asteraceae. At that time, mountain shrub grasslands were permanently established with forests and Afroalpine vegetation in the background. The expansion of cold mountain grassland at 0.9 Ma is followed by the downward expansion of the mountain juniper forest related to more humid conditions. The return of cooler conditions and the re-expansion of grasslands occur at a time close to the Bruhnes/Matuyama boundary. At ~0.6 Ma, the establishment of a rich and diversified humid forest closer to the site attests to a warmer and probably more humid climate.
5.5.5 Past Vegetation Changes linked to Global Orbital Forcing
In the tropics, variability in past vegetation is most likely explained by changes in precipitation, rainfall or available moisture. Between ~1.8–1.6 Ma, wet–dry climate variability most probably triggered the environmental changes observed at Melka Kunture. Rainfall fluctuated in response to orbital changes combined to astronomical forcing linked to precession cycles. Precessional global climate forcing dominates the tropical climate system during the Early Pleistocene (deMenocal 2004). Precession monsoon cycles have been evidenced at Olduvai, on the basis of sedimentological and isotopic studies that show several wet and dry cycles (Hay and Kyser 2001; Sikes and Ashley 2007). At Melka Kunture, the necessary high-resolution sampling at regular intervals cannot be done with the same dating accuracy. However, changes documented seven times within a ~200 ka time-span would attest to cyclicity on the order of 20 ka. This is consistent with precession change cycles evidenced by isotopic studies at Turkana, during the 2–1.85 Ma interval (Joordens et al. 2011) and at Olduvai, from 1.877 to 1.803 Ma (Deino 2012).
Following the onset and development of the continental arctic icesheet, the ocean circulation changed suddenly at 1.6 Ma (Lisiecki and Raymo 2005), and precession cyclicity was replaced by 41 ka obliquity cyclicity.
At 0.9 Ma, when the icesheet had reached a greater height and volume, the tropical climate changed to a system characterized by less influence of precession cycles but greater influence of high-amplitude glacial/interglacial successions. During the Middle Pleistocene Transition (MPT), pollen data from Melka Kunture show the response of past mountain vegetation to longer and more marked climate cycles. At the onset of the MPT, drier and cooler climate conditions indicated at Melka Kunture by pollen studies appear at the same time as greater icesheet thickness in the northern hemisphere, and as temperature decrease in an Antarctica ice core at ~750 ka (Jouzel et al. 2007). During the MPT interval, three glacial stages, MIS 22 (0.9 Ma), MIS 20 (0.8 Ma) and MIS 18 (before 0.7 Ma) have been identified in the oceanic record (Lisiecki and Raymo 2005). These glacial stages are linked to lower solar insolation at 65°N, lower amplitude in precession cycles and lower pCO2 in the atmosphere (Almogi-Labin 2011). The global climate was greatly modified, as recorded in long terrestrial sedimentary sequences such as those in the eastern Mediterranean region (Almogi-Labin 2011). In palynology , past vegetation changes are traditionally based on the fluctuations of total arboreal pollen (AP) versus non-arboreal pollen (NAP). In the Levant and Greece at the time of MIS 22, the terrestrial pollen record of lacustrine sequences shows a minimum of AP, and the extinction of a few relict Tertiary species; whereas during MIS 16 there was a steppe environment, after which a forest of modern composition with Quercus and Carpinus dominates (Tzedakis et al. 2006). The extreme glacial conditions of both MIS 22 and 16 appear to have played a major role in the disappearance of a large number of Tertiary relict species at that time. During the MPT, the absence of sapropel in Mediterranean cores reflects the weakening of the tropical monsoon in East Africa, resulting in less rainfall; the increased dryness encouraged the development of grasslands at the expense of forest. The marine pollen record offshore from the mouth of the Congo River indicates that the equatorial region experienced cyclically predominantly warm and dry conditions during interglacials, but cool and humid conditions during glacial periods (Dupont et al. 2001). In South America, during glacial periods the long terrestrial pollen record spanning MIS 22 to 19 highlights the lowering and oscillations of the upper forest line (Hooghiemstra et al. 1993).
Pollen data from long cores is not available in Africa. The Melka Kunture pollen data are the first terrestrial data for the MPT period. Colder and drier conditions documented in the Ethiopian highlands correspond to glacial stages 22, 20 and 18; whereas warmer more humid conditions, more favorable to forest development, correspond to interglacial stage 21, 19 and 17. Past vegetation changes are consistent with global glacial/interglacial cycles. We discuss elsewhere weather in the highlands the hominins were able, or not, to withstand both interglacial and glacial climates (Mussi et al. 2016).
5.6 Conclusion
The study presented here, which is based on a new assessment of the chronostratigraphy at Melka Kunture, and a new interpretation of the pollen data, emphasizes ecological variability and ecosystems changes that followed global climatic changes during the Early and Middle Pleistocene in the Ethiopian highlands. During a critical period, close in time to the Acheulean emergence (~1.7 Ma), the pattern of fluctuations related to the precessional monsoon cycle had already appeared in the Turkana basin and at Olduvai. Although the stratigraphy of Melka Kunture does not allow much precision in the high resolution, the pollen record presented here is consistent with such precession climate variability . Homo erectus s.l. adapted to mountain climate conditions characterized by large diurnal temperature ranges. Later, during the MPT transition, hominins of a species close to that of Homo heidelbergensis – the knappers of the middle Acheulean lithic industries – experienced the impact of changing environmental conditions during glacial/interglacial cycles.
5.7 Note About Palynological Processing
Sediment samples used for pollen analysis were collected on cleaned outcrops or deep trenches dug out from exposures. The initial stratigraphic position of each pollen sample (Bonnefille 1972) can be found on the same lithostratigraphic sections as geological studies (Taieb 1974). A few additional samples collected in the Simbiro gully, and at Melka Gila (Djilla) did not yield pollen . One pollen assemblage from the Tuka gully (Bonnefille 1972) is not included here because the sample cannot be securely placed in the updated chronostratigraphy.
Pollen extraction from the sediment (up to 100 g) is performed according to the classical chemical treatment, attacking the sediment with HCl and HF acids to remove the mineral fraction, followed by hot-water rinses and by treatment with diluted KOH to remove humic acids. The residue is then stained with diluted safranine and kept in glycerin. It is mounted on slides in glycerin allowing to turn pollen grains and facilitating the observation under the microscope of the pollen wall structure. The identification of pollen was performed by comparing them with a 7,000-specimen reference collection of modern pollen stored at Cerege, Aix-en-Provence. Descriptions of pollen from most common trees and plants are available (Bonnefille 1969, 1971a, b; Riollet and Bonnefille 1976; Bonnefille and Riollet 1980b), with photo in the African Pollen Database. 15% of the processed samples yielded fossil pollen grains (Bonnefille 1972). The pollen assemblages from bones infills show species distribution similar to the one in a modern sample from the Awash River. Taphonomic preservation of bone remains (Fiore and Tagliacozzo 2004) on the banks of a meandering Awash River may explain this exceptional pollen preservation. We consider pollen assemblages extracted from bone cavities to be reliable.
References
Almogi-Labin, A. (2011). The paleoclimate of the Eastern Mediterranean during the transition from early to mid Pleistocene (900 to 700 ka) based on marine and non-marine records: An integrated overview. Journal of Human Evolution, 60, 428–436.
Bailloud, G. (1965). Les gisements paléolithiques de Melka-Kontouré. Cahier n°1, Institut Ethiopien d’Archéologie.
Barboni, D. (2014). Vegetation of Northern Tanzania during the Plio-Pleistocene: A synthesis of the paleobotanical evidences from Laetoli, Olduvai, and Peninj hominin sites. Quaternary International, 322–323, 264–276.
Beyene, Y., Katoh, S., WoldeGabriel, G., Hart, W. K., Uto, K., Sudo, M., et al. (2013). The characteristics and chronology of the earliest Acheulean at Konso, Ethiopia. Proceedings of the National Academy of Sciences, 110, 1584–1591.
Bonnefille, R. (1968). Contribution de l’étude de la flore d’un niveau pléistocène de la haute vallée de l’Awash (Ethiopie). Comptes Rendus de l’Académie des Sciences Paris, D, 266, 1229–1232.
Bonnefille, R. (1969). Analyse d’un sediment recent : vases actuelles de la rivière Awash (Ethiopie). Pollen et Spores, 11, 7–16.
Bonnefille, R. (1971a). Atlas des pollens d’Ethiopie. Principales espèces des forêts de montagne. Pollen et Spores, 13, 15–72.
Bonnefille, R. (1971b). Atlas des pollens d’Ethiopie. Pollens actuels de la basse vallée de l’Omo, récoltes botaniques 1968. Adansonia, 2(11), 463–518.
Bonnefille, R. (1972). Association polliniques actuelles et quaternaires en Ethiopie (vallées de l’Awash et de l’Omo). Ph.D. Dissertation, Université de Paris.
Bonnefille, R. (1975). Présence de pollens fossiles d’Acacia albida Del. et de Cycadales dans les échantillons pléistocènes de Melka Kunturé (Ethiopie). Boissiera, 24, 33–42.
Bonnefille, R. (2007). Rain forest responses to past climatic changes in tropical Africa. In J. Flenley & M. Bush (Eds.), Tropical Rain Forest Responses to Climatic Change (pp. 118–170). New York: Springer.
Bonnefille, R., & Riollet, G. (1980a). Palynologie, végétation et climats de Bed I et Bed II à Olduvai, Tanzanie. In R. E. Leakey & B. A. Ogot (Eds.), Proceedings of the 8th Panafrican Congress of Prehistory and Quaternary Studies, Nairobi, September 1977 (pp. 123–127). Nairobi: TILLMIAP.
Bonnefille, R., & Riollet, G. (1980b). Pollens des savanes d’Afrique orientale. Paris: CNRS.
Bonnefille, R., Lobreau, D., & Riollet, G. (1982). Pollen fossile de Ximenia (Olacaceae) dans le Pléistocène inférieur d’Olduvai en Tanzanie: implications paléoécologiques. Journal of Biogeography, 9, 469–486.
Cerling, T. E., Wynn, J. G., Andanje, S. A., Bird, M. I., Korir, D. K., Levin, N. E., et al. (2011). Woody cover and hominin environments in the past 6 million years. Nature, 476, 51–56.
Chavaillon, J. (1971). Les habitats préhistoriques en Ethiopie : Melka Kunture. Sciences (Paris), 73, 44–52.
Chavaillon, J. (1973). Chronologie des niveaux paléolithiques de Melka-Kunturé. Comptes Rendus de l’Académie des Sciences Paris, D, 276, 1533–1536.
Chavaillon, J. (1979). Stratigraphie du site archéologique de Melka-Kunturé (Ethiopie). Bulletin de la Société Géologique Française, 7° série, XXI, 3, 227–232.
Chavaillon, J. (1980). Chronologie archéologique de Melka-Kunturé (Ethiopie). In R. E. Leakey & B. A. Ogot (Eds.), Proceedings of the 8th Panafrican Congress of Prehistory and Quaternary Studies, Nairobi, September 1977 (pp. 200–201). Nairobi: TILLMIAP.
Chavaillon, J., & Berthelet, A. (2004). The archaeological sites of Melka Kunture. In J. Chavaillon & M. Piperno (Eds.), Studies on the Early Paleolithic Site of Melka Kunture, Ethiopia (pp. 25–80). Florence: Origines.
Chavaillon, J., & Coppens, Y. (1975). Découverte d’Hominidé dans un site acheuléen de Melka-Kunturé. Bulletin et Mémoires de la Société d’Anthropologie, Paris, 2, 125–128.
Chavaillon, J., & Coppens, Y. (1986). Nouvelle découverte d’Homo erectus à Melka-Kunturé. Comptes Rendus de l’Académie des Sciences Paris, II, 1(303), 99–104.
Chavaillon, J., & Koeniguer, J.-C. (1970). Découverte d’un fragment de liane dans le sol oldowayen de Melka Konturé (Ethiopie). Comptes Rendus de l’Académie des Sciences Paris, D, 270, 1969–1972.
Chavaillon, J., & Piperno, M. (Eds.). (2004). Studies on the Early Paleolithic site of Melka Kunture, Ethiopia. Florence: Origines.
Chavaillon, J., Brahimi, C., & Coppens, Y. (1974). Première découverte d’Hominidé dans l’un des sites acheuléens de Melka-Kunturé (Ethiopie). Comptes Rendus de l’Académie des Sciences Paris, D, 278, 3299–3302.
Chavaillon, J., Chavaillon, N., Coppens, Y., & Senut, B. (1977). Présence d’Hominidé dans le site oldowayen de Gomboré I à Melka-Kunturé. Comptes Rendus de l’Académie des Sciences Paris, D, 285, 961–963.
Chernet, T., Hart, W. K., James, L., Aronson, J. L., Robert, C., & Walter, R. C. (1998). New age constraints on the timing of volcanism and tectonism in the northern Main Ethiopian Rift—southern Afar transition zone (Ethiopia). Journal of Volcanology and Geothermal Research, 80, 267–280.
Clark, J. D., & Kurashina, H. (1979). An Analysis of Earlier Stone Age Bifaces from Gadeb (Locality 8E), Northern Bale Highlands, Ethiopia. The South African Archaeological Bulletin, 34, 93–109.
Condemi, S. (2004). The Garba IV E mandible. In J. Chavaillon & M. Piperno (Eds.), Studies on the Early Paleolithic Site of Melka Kunture, Ethiopia (pp. 687–701). Florence: Origines.
Cressier, P. (1980). Magnétostratigraphie du gisement pléistocène de Melka-Kunturé (Ethiopie). Datation des niveaux oldowayens et acheuléens. Ph.D. Dissertation, Université Louis Pasteur, Strasbourg.
de Heinzelin, J., Clark, J. D., Schick, K., & Gilbert, W. H. E. (2002). The Acheulean and the Plio-Pleistocene Deposits of the Middle Awash Valley, Ethiopia. Tervuren: Musée Royal de l’Afrique Centrale.
de la Torre, I. (2004). Omo revisited. Current Anthropology, 45, 439–465.
de la Torre, I. (2011). The Early Stone Age lithic assemblages of Gadeb (Ethiopia) and the developed Oldowan/early Acheulean in East Africa. Journal of Human Evolution, 60, 768–812.
de la Torre, I., Mora, R., & Martínez-Moreno, J. (2008). The early Acheulean in Peninj (Lake Natron, Tanzania). Journal of Anthropological Archaeology, 27, 244–264.
Deino, A. L. (2012). 40Ar/39Ar dating of Bed I, Olduvai Gorge, Tanzania, and the chronology of early Pleistocene climate change. Journal of Human Evolution, 63, 251–273.
deMenocal, P. (2004). African climate change and faunal evolution during the Pliocene–Pleistocene. Earth and Planetary Science Letters, 220, 3–24.
Di Vincenzo, F., Rodriguez, L., Carretero, J. M., Collina, C., Geraads, D., Piperno, M., et al. (2015). The massive fossil humerus from the Oldowan horizon of Gombore I, Melka Kunture (Ethiopia, >1.39 Ma). Quaternary Science Reviews, 122, 207–221.
Diez-Martín, F., Sánchez Yustos, P., Uribelarrea, D., Baquedano, E., Mark, D. F., Mabulla, A., et al. (2015). The Origin of the Acheulean: The 1.7 Million-Year-Old Site of FLK West, Olduvai Gorge (Tanzania). Nature Scientific Reports, 5, 17839.
Dupont, L. M., Donner, B., Schneider, R., & Wefer, G. (2001). Mid-Pleistocene environmental change in tropical Africa began as early as 1.05 Ma. Geology, 29, 195–198.
Fiore, I., & Tagliacozzo, A. (2004). Taphonomic analysis of the bone remains from the Oldowan site of Garba IV. In J. Chavaillon & M. Piperno (Eds.), Studies on the Early Paleolithic site of Melka Kunture, Ethiopia (pp. 639–682). Florence: Origines.
Friis, I. (1992). Forest and forest trees of Northeast Tropical Africa: Their natural habitats and distribution pattern in Ethiopia, Djibouti and Somalia. Kew Bulletin Additional Series, 15, 1–396.
Friis, I., & Sebsebe, D. (2001). Vegetation maps of Ethiopia and Eritrea. A review of the existing maps and the need for a new map for the Flora of Ethiopia and Eritrea. Biologiske Skrifter, 54, 399–439.
Friis, I., Demissew, S., & Van Breugel, P. (2010). Atlas of the Potential Vegetation of Ethiopia. Biologiske Skrifter, 58, 1–307.
Gallotti, R. (2013). An older origin for the Acheulean at Melka Kunture (Upper Awash, Ethiopia). Techno-economic behaviours at Garba IVD. Journal of Human Evolution, 65, 594–620.
Gallotti, R., & Mussi, M. (2015). The Unknown Oldowan: ~1.7-Million-Year-Old Standardized Obsidian Small Tools from Garba IV, Melka Kunture, Ethiopia. PLoS ONE, 10, e0145101.
Gallotti, R., & Mussi, M. (2018). Before, During, and After the Early Acheulean at Melka Kunture (Upper Awash, Ethiopia): A Techno-economic Comparative Analysis. In R. Gallotti & M. Mussi (Eds.), The Emergence of the Acheulean in East Africa and Beyond. Contributions in Honor of Jean Chavaillon. Cham: Springer.
Gallotti, R., Collina, C., Raynal, J.-P., Kieffer, G., Geraads, D., & Piperno, M. (2010). The Early Middle Pleistocene site of Gombore II (Melka Kunture, Upper Awash, Ethiopia) and the issue of Acheulean Bifacial Shaping Strategies. African Archaeological Review, 27, 291–322.
Gallotti, R., Raynal, J.-P., Geraads, D., & Mussi, M. (2014). Garba XIII (Melka Kunture, Upper Awash, Ethiopia): A new Acheulean site of the late Lower Pleistocene. Quaternary International, 343, 17–27.
Geraads, D., Gallotti, R., Raynal, J. P., Bonnefille, R., & Mussi, M. (in press). Melka Kunture, Ethiopia: Lower Pleistocene faunas of the Ethiopian highlands. In S. C. Reynolds & R. Bobe (Eds.), African Paleoecology and Human Evolution. Cambridge: Cambridge University Press.
Gibbard, P. L., Head, M. J., Walker, M. J. C., & the Subcommission on Quaternary Stratigraphy (2010). Formal ratification of the Quaternary System/Period and the Pleistocene Series/Epoch with a base at 2.58 Ma. Journal of Quaternary Sciences, 25, 96–102.
Goren-Inbar, N., Melamed, Y., Zohar, I., Akhilesh, K., & Pappu, S. (2014). Beneath Still Waters—Multistage Aquatic Exploitation of Euryale ferox (Salisb.) during the Acheulian. In R. Fernandes & J. Meadows (Eds.), Human Exploitation of Aquatic Landscapes, special issue. Internet Archaeology. https://doi.org/10.11141/ia.37.1.
Goren-Inbar, N., Sharon, G., Melamed, Y., & Kislev, M. (2007). Nuts, nut cracking, and pitted stones at Gesher Benot Ya‘aqov, Israel. Proceedings of the National Academy of Sciences, 99, 2455–2460.
Hay, R. L., & Kyser, T. K. (2001). Chemical sedimentology and paleoenvironmental history of Lake Olduvai, a Pleistocene lake in northern Tanzania. Geological Society of America Bulletin, 113, 1505–1521.
Hooghiemstra, H., Melice, J. L., Berger, A., & Shackleton, N. J. (1993). Frequency spectra and paleoclimatic variability of the high-resolution 30–145 ka Funza I pollen record (eastern Cordillera Colombia). Quaternary Science Reviews, 12, 141–156.
Isaac, G. Ll., & Isaac, B. (Eds.). (1997). Koobi Fora Research Project. Plio-Pleistocene Archaeology (Vol. 5). Oxford: Clarendon Press.
Joordens, J. C. A., Vonhof, H. B., Feibel, C. S., Lourens, L. J., Dupont-Nivet, G., van der Lubbe, J. H. J. L., et al. (2011). An astronomically-tuned climate framework for hominins in the Turkana Basin. Earth and Planetary Science Letters, 307, 1–8.
Jouzel, J., Masson-Delmotte, V., Cattani, O., Dreyfus, G., Falourd, S., Hoffmann, G., et al. (2007). Orbital and millennial Antarctic climate variability over the past 800,000 years. Science, 317, 793–797.
Kieffer, G., Raynal, J.-P., & Bardin, G., 2002. Cadre structural et volcanologiques des sites du Paléolithique ancien de Melka Kunture (Awash, Ethiopie): Premiers résultats. In J.-P. Raynal, C. Albore-Livadie & M. Piperno (Eds.), Hommes et Volcans. De l’éruption à l’objet, Les Dossiers de l’Archéologie n° 2 (pp. 77–92). Liège: Archéo-Logis/CDERAD.
Leakey, M. D. (1971). Olduvai Gorge. Excavations in Bed I and II, 1960–1963 (Vol. 3). Cambridge: Cambridge University Press.
Lepre, C. J., Roche, H., Kent, D. V., Harmand, S., Quinn, R. L., Brugal, J.-P., et al. (2011). An earlier origin for the Acheulian. Nature, 477, 82–85.
Lisiecki, L. E., & Raymo, M. E. (2005). A Pliocene-Pleistocene stack of 57 globally distributed benthic δ18O records. Paleoceanography, 20, PA1003.
Magill, C. R., Ashley, G. M., & Freeman, K. H. (2013). Ecosystem variability and early human habitats in eastern Africa. Proceedings of the National Academy of Sciences, 110, 1167–1174.
Mohr, P. A. (1971). Ethiopian rift and plateaus: Some volcanic petrochemical differences. Journal of Geophysical Research, 76, 1967–1984.
Morgan, L. E., Renne, P. R., Kieffer, G., Piperno, M., Gallotti, R., & Raynal, J.-P. (2012). A chronological framework for a long and persistent archaeological record: Melka Kunture, Ethiopia. Journal of Human Evolution, 62, 104–115.
Mussi, M., Altamura, F., Bonnefille, R., De Rita, D., & Melis, R. T. (2016). The environment of the Ethiopian highlands at the Mid Pleistocene transition: Fauna, flora and hominins in the 850–700 ka sequence of Gombore II (Melka Kunture). Quaternary Science Reviews, 149, 259–268.
Mussi, M., Altamura, F., Macchiarelli, R., Melis, R. T., & Spinapolice, E. (2014). Garba III (Melka Kunture, Ethiopia): A MSA site with archaic Homo sapiens remains revisited. Quaternary International, 343, 28–39.
Pichi-Sermolli, R. E. G. (1957). Une carta geobotanica dell’Africa orientale (Eritrea Etiopia, Somalia). Webbia, 13, 15–132.
Piperno, M., Collina, C., Gallotti, R., Raynal, J.-P., Kieffer, G., Le Bourdonnec, F.-X., et al. (2009). Obsidian exploitation and utilization during the Oldowan at Melka Kunture (Ethiopia). In E. Hovers & D. R. Braun (Eds.), Interdisciplinary Approaches to the Oldowan (pp. 111–128). Dordrecht: Springer.
Profico, A., Di Vincenzo, F., Gagliardi, L., Piperno, M., & Manzi, G. (2016). Filling the gap. Human cranial remains from Gombore II (Melka Kunture, Ethiopia; ca. 850 ka) and the origin of Homo heidelbergensis. Journal of Anthropological Sciences, 94, 1–24.
Raynal, J.-P., & Kieffer, G. (2004). Lithology, dynamism and volcanic successions at Melka Kunture (Upper Awash, Ethiopia). In J. Chavaillon & M. Piperno (Eds.), Studies on the Early Paleolithic Site of Melka Kunture, Ethiopia (pp. 111–135). Florence: Origines.
Roche, H., Brugal, J.-P., Delagnes, A., Feibel, C., Harmand, S., Kibunjia, M., et al. (2003). Les sites archéologiques plio-pléistocènes de la formation de Nachukui, Ouest-Turkana, Kenya : bilan synthétique 1997–2001. Comptes Rendus Palevol, 2, 663–673.
Riollet, G., & Bonnefille, R. (1976). Pollen des Amaranthacées du Lac Rodolphe (Afrique orientale), déterminations générique et spécifique. Pollen et spores, 18/1, 67–92, 6 pl.
Sikes, N. E., & Ashley, G. M. (2007). Stable isotopic signatures of pedogenic carbonates as indicators of paleoecology in the Plio-Pleistocene (upper Bed I) western margin of Olduvai Basin, Tanzania. Journal of Human Evolution, 53, 574–594.
Taieb, M. (1974). Évolution quaternaire du bassin de l’Awash. Ph.D. Dissertation, Université de Paris.
Tamrat, E., Thouveny, N., Taieb, M., & Brugal, J. P. (2014). Magnetostratigraphic study of the Melka Kunture archaeological site (Ethiopia) and its chronological implications. Quaternary International, 343, 5–16.
Teaford, M. F., & Ungar, P. S. (2000). Diet and the evolution of the earliest human ancestors. Proceedings of the National Academy of Sciences, 97, 13506–13511.
Terwilliger, V., Eshetu, Z., Huang, Y., Alexandre, M., Umer, M., & Gebru, T. (2011). Local variation in climate and land use during the time of the major kingdoms of the Tigray Plateau in Ethiopia and Eritrea. Catena, 85, 130–143.
Tzedakis, P. C., Hooghiemstra, H., & Pälike, H. (2006). The last 1.35 million years at Tenaghi Philippon: Revised chronostratigraphy and long-term vegetation trends. Quaternary Science Reviews, 25, 3416–3430.
Umer, M., Lamb, H. F., Bonnefille, R., Lézine, A.-M., Tiercelin, J.-J., Gibert, E., et al. (2007). Late Pleistocene and Holocene vegetation history of the Bale Mountains, Ethiopia. Quaternary Science Reviews, 26, 2229–2246.
Ungar, P. S., & Sponheimer, M. (2011). The Diets of Early Hominins. Science, 334, 190–193.
Wesphal, M., Chavaillon, J., & Jaeger, J.-J. (1979). Magnétostratigraphie des dépôts pléistocènes de Melka-Kunturé (Ethiopie), premières données. Bulletin de la Société Géologique Française, 7ème série, XXI(3), 237–241.
White, F. (1983). The Vegetation of Africa. Paris: Unesco.
Zerihun, W. (1999). Vegetation Map of Ethiopia. Addis Ababa: Addis Ababa University.
Zerihun, W., & Backéus, I. (1991). The shrubland vegetation in western Shewa, Ethiopia, and its possible recovery. Journal of Vegetation Science, 2, 173–180.
Zilberman, U., Smith, P., Piperno, N., & Condemi, S. (2004). Evidence of amelogenesis imperfecta in an early African Homo erectus. Journal of Human Evolution, 46, 647–653.
Acknowledgements
The recent research at Melka Kunture was supported by grants from Sapienza Università di Roma (“Grandi scavi archeologici”) and from the Italian Foreign Ministry awarded to MM. We thank the Authority for Research & Conservation of Cultural Heritage of the Ministry of Culture & Tourism, the National Museum of Addis Ababa, and the Oromia Culture and Tourism Bureau for permits for fieldwork and access to the lithic collections. RB did the paleobotanical analysis and reexamined the environmental interpretation, RTM established the stratigraphic sequence and MM designed, encouraged and coordinated the project. The late Nicole Chavaillon and her daughters generously entrusted Jean Chavaillon’s archives to MM. We thank Guillaume Jouve (CEREGE) et Morteza Djamali (IMBE) for their help in drawing the pollen diagrams. Rosalia Gallotti participated in the discussion with the three authors. Flavio Altamura provided important information related to the stratigraphic position of samples Gomb173 and Gomb349.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2018 Springer International Publishing AG, part of Springer Nature
About this chapter

Cite this chapter
Bonnefille, R., Melis, R.T., Mussi, M. (2018). Variability in the Mountain Environment at Melka Kunture Archaeological Site, Ethiopia, During the Early Pleistocene (~1.7 Ma) and the Mid-Pleistocene Transition (0.9–0.6 Ma). In: Gallotti, R., Mussi, M. (eds) The Emergence of the Acheulean in East Africa and Beyond. Vertebrate Paleobiology and Paleoanthropology. Springer, Cham. https://doi.org/10.1007/978-3-319-75985-2_5
Download citation
DOI: https://doi.org/10.1007/978-3-319-75985-2_5
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-75983-8
Online ISBN: 978-3-319-75985-2
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)