
Abstract
Tigers (Panthera tigris) are critically endangered worldwide, with the geographical range of native populations reduced to less than 7% in the last hundred years. Currently, there are only 6 subspecies residing in thirteen range countries. The main causes of their decline are habitat loss and fragmentation, prey depletion and poaching for the illegal wildlife trade. Addressing these issues has been severely constrained by inadequate spatial information on tiger distributions, movements, habitat preferences and behaviour. Focusing on the Indo-Bhutan Manas Tiger Conservation Landscape (IBMTCL) in the Indian subcontinent (India and Bhutan), this study sought to demonstrate the use of field-based camera traps for providing ground-based observations of tigers and other large cats, namely leopards (Panthera pardus) and clouded leopards (Neofelis nebulosa), and their prey. These observations were also coupled with classifications of land cover, elevation data and fire observations from Landsat TM and the Terra ASTER and MODIS respectively. The study indicated a large but variable range for individual tigers, a preference of tigers for forest cover and proximity to burned areas (attributed to greater access to prey), and a spatial separation from populations of other large cats. The study illustrates how earth observation data can provide some of the elements needed to better understand how large cats utilise the landscapes they inhabit thereby contributing to efforts aimed at long-term conservation of these endanged species across their range.
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others


Keywords
- C12
- Tiger
- Panthera tigris
- Carnivores
- Royal Manas National Park
- Manas National Park
- Camera trap
- Earth observation
- Fire
- Topography
- Land cover
Introduction
The tiger (Panthera tigris) is the largest and one of the most charismatic and endangered cat species in the world. It is a flagship icon for wildlife conservation, being at the apex of the food chain in the majority of the eco-regions in Asia (Sunquist et al. 1999; Jhala et al. 2008, 2015). The tiger has been held in mythical reverence across the world, is worshipped in certain faiths and religions and is the national animal for India, Bangladesh, Nepal, Malaysia and both North and South Korea. Until the early nineteenth century, nine subspecies (Luo et al. 2004) of tiger were found across Asia, from the Caucasus and the Caspian Sea to Siberia and Indonesia. Three of these subspecies are now extinct; the Bali Tiger (Panthera tigris balica), Javan Tiger (P.t.sondaica) and Caspian Tiger (P.t.virgata). The remaining six occur as fragmented populations in the Russian Far East, northeast China and the Asian subcontinent.
India supports about 70% of the World’s tiger population and one of the key areas of importance is the Indo-Bhutan Manas Tiger Conservation Landscape (IBMTCL), which offers a contiguous protected landscape of over 6000 sq km and constitutes the core area of Manas National Park (NP) in India and Royal Manas NP in Bhutan. Between November 2010 and February 2011, a large camera trap survey was conducted in Manas NP and the southern section of Royal Manaus NP by organisations in both India and Bhutan. This survey was undertaken to locate, identify and track the movements of individual Royal Bengal tigers (Panthera tigris tigris) and two other sympatric species, the leopard (Panthera pardus) and clouded leopard (Neofelis nebulosa). Landsat-derived classifications of land cover, historical MODIS fire records and spatial information on human development of the landscape were then used in combination with the survey results to provide a better understanding of niche separation as well as influences on the movement and range of these large cat species over the period of observation. Through this approach, the potential contribution of integrated field campaigns and earth observation to supporting management of Tiger Conservation Landscapes (TCLs) was assessed.
Background
Why Are Tigers Endangered?
Whilst once widespread, the range of tigers has been reduced to a mere 7% (less than 1,184,911 km2) of their former geographical range, and approximately 55% of the inhabited landscape is essentially non-tiger habitat (Sanderson et al. 2006). A general compilation of national estimates suggests a global population of approximately 4240 tigers remaining in the wild (Goodrich et al. 2015). The causes for their decline has largely been habitat loss and poaching and the pressures continue to escalate (Sanderson et al. 2006; Dinerstein et al. 2007). They are also vulnerable to extinction because of their low densities (relative to other mammals, including their prey species) and relatively low recruitment rates (where few animals raise offspring that survive to join the breeding population) (Kerley et al. 2003). Tigers also require large undisturbed landscapes with ample prey to raise young and maintain long term genetic and demographic viability (Seidensticker and McDougal 1993; Karanth and Sunquist 1995; Carbone et al. 1999; Jhala et al. 2008). However, such areas are contracting or becoming increasingly fragmented because of high anthropogenic pressures on natural resources. Whilst India has the world’s largest population of wild tigers, it has also suffered the highest range contraction for the species. Poaching is mainly for the supply of organs, bones and hides that are smuggled for use in traditional medicine, mainly into China and south-east Asia (Gratwicke et al. 2008; Nowell 2009).
Efforts to save the tiger started rather late. For example, Project Tiger was initiated in India in 1973 (GOI 2005). However, the strategies used were not time-tested and were also often weak on scientific rigor, partly because of a general lack of baseline information on tiger distributions (ibid). Conservation strategies for tigers had previously focused only on protecting the species in pristine wilderness areas and were based on the premise that estimates of tiger densities derived from sign-based indices (e.g., pugmark census techniques) were adequate to provide information on the survival status of the species. However, detecting tigers through traditional field survey methods is a challenging task. Tigers require sufficient numbers of large-bodied prey and therefore roam across large that may exceed 100s of kilometres. They also occur in low densities and often as lone individuals, particularly in closed forests (Gittleman and Harvey 1982; Carbone et al. 2001), are cryptic by nature and are rarely seen. As a consequence, records relating to tiger distributions and their use of habitats have been inadequate and this has severely impeded the formulation and implementation of appropriate conservation measures (Blake and Hedges 2004; Sanderson et al. 2006).
For these reasons, there has been an increasing and now urgent need to devise rapid yet rigorous population analysis and viability methods with spatial and temporal observations that are repeatable across the varied landscapes that tigers occupy (Linkie et al. 2006). It is also widely accepted that a landscape approach to tiger conservation that takes into account the distribution and condition of habitat also fares better than those that focus only on protecting the species. Although breeding populations are currently found in eight Asian range states, there are few regular empirical studies to indicate trends on a countrywide basis (Goodrich et al. 2015). Sanderson et al. (2006) put forward the concept of establishing Tiger Conservation Landscapes (TCLs) and estimating tiger populations across their entire range using multiple methods; from expert opinion to land cover (e.g., vegetation) and change assessments based on satellite sensor data. The study also highlighted that 76 TCLs held the major tiger subpopulations in the world. An additional 543 fragments were also identified, but they were considered too small to support long-term viable populations. 491 areas were designated as Tiger Survey Landscapes (TSL; total area of 1.1 million km2), where the current tiger status was unknown but the area was considered sufficiently large to support at least five tigers. The study highlighted that it would be a major challenge to survey and monitor these TCLs on a regular basis unless new options for mapping and monitoring their state, condition and dynamics were developed and implemented.
Determining the Distribution and Abundance of Tigers
Whilst the ecology of wild tigers has been studied in depth (Sanderson et al. 2006), information on their present status is limited and scattered over a number of publications, with many in the grey literature. In a review conducted in support of this study, 35 papers were identified that provided information relating specifically to assessing tiger populations and their habitat, with these based largely on primary data collection. Approximately half of these studies used capture-recapture methods with a lesser proportion focusing on sign-based indices (such as tracks, visual estimates, scat DNA analysis; 25%), and satellite tracking (7%). Only one paper (Imam et al. 2009) explicitly used remote sensing data to assess tiger habitat suitability in the landscape. The review highlighted the general paucity of information, with only a few local-scale studies, but also the varied and often inconsistent approaches for estimating population distributions and sizes. Furthermore, many population estimates were found to be based on single-studies, typically field-based surveys, with these relying primarily on pugmark census, camera traps, satellite telemetry, scent-matching using dogs, fecal genetics and large scale occupancy surveys. Most studies also relied on capture-recapture techniques conducted over a specified period (Table 1).
From the 1970s to 2005, the pugmark census technique has been widely used and has relied on sign-based abundance estimates. The census is based on intensive monitoring of tigers within areas, identifying individual tigers by visual inspection of the pugmark tracings/plaster casts, measuring pug mark variables (e.g., for toe to pad distance, pad length and width etc.) and subsequently using statistical techniques to identify individuals and numbers (Choudhury 1970; Sharma et al. 2005). Satellite telemetry allows animals with radio collars to be tracked from the ground or air. Animal locations are plotted on base maps from which their home range can be approximated. This method is particularly useful for detecting the movement of individuals and understanding tiger ecology and behaviour (Smith et al. 1998).
Camera trapping has long been used to estimate populations of faunal species (Otis et al. 1978; Balme et al. 2009; Carbone et al. 2001; Harihar et al. 2009) and is currently the most accepted technique for studying tiger populations in the Indian Subcontinent (Karanth and Sunquist 1995; Karanth et al. 2002; Karanth et al. 2006; Royle et al. 2009; Wegge and Storaas 2009). This method works on the a priori premise that individual tigers have unique biometrics, such as stripe patterns or tail to body ratios, that can be captured by placing automated camera traps in optimal field locations. The biometric information can then be analyzed within a robust capture-recapture statistical framework to arrive at estimates of numbers. The technique has been used for determining the spatial location and movements of carnivores such as tigers (Karanth et al. 2006; O’Brian et al. 2003; Linkie et al. 2006), leopards (Harihar et al. 2011), pumas (Kelly et al. 2008), jaguars (Davis et al. 2010) and other large predators. New techniques that have been used in conjunction with camera traps include the use of scent-matching dogs for Amur tigers (Kerley and Salkina 2007), fecal genetics to prepare DNA profiles of individual tigers (Samrat et al. 2009) and large-scale occupancy surveys for tigers and their prey (Jhala et al. 2008).
Use of Remote Sensing Data for Tiger Habitat Mapping
To address the mapping of tigers distributions, a handful of studies (e.g., Sanderson et al. 2006; Imam et al. 2009) have used thematic or continuous layers (e.g., representing vegetation, land use and elevation data) derived from remote sensing data as input to Species Distribution Models (SDM) (e.g., occupancy) or population models. Most have focused on using freely available pre-processed optical satellite sensor data, pixel-based image classifications and geometry-based geospatial analysis (Karanth et al. 2009; Ranganathan et al. 2008; Jhala et al. 2008). However, their generation and use has often been compromised by uncertainty and errors (positional and thematic) related to geometric correction accuracy, poor atmospheric and topographic correction (particularly in the mountainous areas that tigers commonly occupy), and inconsistent use of classification and accuracy assessment techniques. There is, however, considerable interest in the use of remote sensing data, including as input to SDMs and population models, and the increased quality, diversity, availability and accessibility of data and derived products is contributing to greater uptake. A summary of information that can be obtained from remotely sensed data and which is relevant to understanding and/or modelling tiger distributions, movement and behaviour is provided in Table 2.
The Indo-Bhutan Manas Tiger Conservation Landscape (IBMTCL) and Its Conservation Significance
The IBMTCL lies at the critical juncture of the Indo-Malayan and the Indo-Gangetic bio-geographical pathways and has been lauded for its outstanding natural beauty and unparalleled diversity as a UNESCO World Heritage site (UNESCO 2008). The IBMTCL forms an important connecting link between the Buxa, Nameri and Pakke Tiger Reserves in India and the Hukaung Valley Tiger Reserve in Myanmar. Within this region, a number of Park Areas (PAs) exist, including Jigme Singye National Park (NP), Royal Manas NP, Phibsoo Wildlife Sanctuary (WLS), Khaling WLS in Bhutan and the Manas Tiger Reserve (MTR) in India. Together, these form the single-largest TCL for tigers (Panthera tigris tigris) in the world (Dorji and Santiapillai 1989; Sanderson et al. 2006).
The IBMTCL is of historical evolutionary significance as tigers here share the connecting gene pool with the south eastern tiger population and the area represents the entry point of tigers into the Indian sub-continent (Jhala et al. 2011). Administratively, the IBMTCL is comprised of six Protected Areas (PAs; Table 3) but there are several potential landscapes which do not have PA status but nevertheless are able to support tigers (e.g., the Ripu-Chirang Forest complexes in India).
Data Collected
Mammal Surveys
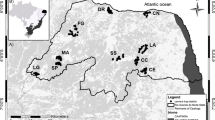
Various organizations, including NGOs such as WWF-India, Aaranyak and the Wildlife Trust of India, assisted the Manas NP Authority in India to lay camera traps between November 2010 and February 2011. This was part of a nation-wide tiger sampling exercise carried out within most of the tiger reserves in India (Jhala et al. 2011). The infrared-triggered camera-traps used within the two ranges of Manas NP (Bansbari and Bhuyanpara) were the Cuddeback (Non Typical Inc. Wisconsin), TrailMaster (Goodson and Associates, Kansas, USA) and Panthera Camera Trap V3 (Panthera, USA). A pair of camera traps was placed within individual 2 × 2 km sized cells and the total area covered by all camera traps approximated 300 km2 (in India) (Table 4). In a separate but simultaneous exercise in Royal Manas NP, camera trapping was undertaken by the Department of Agriculture, Royal Government of Bhutan (RGoB) in collaboration with the Ugyen Wangchuk Institute for Conservation and Environment (UWICE) and the Bhutan Foundation, specifically to monitor the tiger population (Fig. 1). This study focused only on the camera trapping undertaken in Manas NP.

The location of camera traps placed at IBMTCL between November 2010 and February 2011
Remote Sensing and Other Spatial Data
For the 21 and 30 January and 6 and 7 February 2010, relatively cloud free Landsat Enhanced Thematic Mapper (ETM+) data were downloaded from the Glovis (http://glovis.usgs.gov/) and Land Processes Distributed Active Archive Centre (https://lpdaac.usgs.gov/get_data/data_pool). These data were converted to surface reflectance (%) using the 6S atmospheric correction software. Radiometric normalization between scenes was also undertaken using the approach outlined by Homer et al. (1997). All images were geo-referenced by first establishing ground control points between locations identified from existing maps and the most cloud free Landsat sensor data, warping these images and then generating a mosaic covering the study area. Control points were then established subsequently between the remaining Landsat sensor data and this reference mosaic. The processing of the Landsat sensor data was undertaken using the open source software RSGISLib (www.rsgislib.org; Bunting et al. 2013).
The history of burning with the IBMTCL over the period 2000–2012 was documented by referencing the MODIS product MOD 14 (Thermal Anomalies – Fires and Biomass Burning). This product includes fire occurrence (day/night) and location, the logical criteria used for the fire selection and an energy calculation for each fire. 8-day and monthly day-and-night composite fire occurrence (full resolution) and gridded 10-km and 0.5° summaries (including counts) per fire class (daily/8-day/monthly) are also available. The Level 2 products include various fire related parameters including the occurrence of day and night time thermal anomalies, flagged and grouped into different temperature classes associated with different levels of emitted energy from the fire. These parameters are retrieved daily at 1 km spatial resolution.
An ASTER Global Digital Elevation Model (GDEM) was used (http://asterweb.jpl.nasa.gov/gdem-wist.asp) to establish the topography of the two national parks, with this released at 30 m resolution and available for 1 × 1 tiles (geographic projection). Other data layers were also available to support understanding the movement of large cats, including continuous distance from roads and the locations of anti-poaching camps and settlements. The park boundary, roads and rivers were digitized from topographic sheets and Google Earth images, although the boundary location was revised using the Landsat sensor data available for 2010.
Methods of Data Analysis
Analysis of Camera Trap Data
Each day (24 h) was defined as one sampling occasion (Otis et al. 1978) and this was repeated over a period of 57 days (Otis et al. 1978; Karanth 1995). Individual tigers were identified with reference to their documented stripe pattern, with this undertaken by carefully examining the position and shape of stripes on the flanks, limbs, forequarters and sometimes the tail (Schaller 1967; Karanth and Nichols 1998). Each tiger captured in a photograph was then assigned a unique identification number (e.g. TM1M, TM2F for the first and second tiger of male and female gender). Similar procedures were undertaken for leopard and clouded leopard. Individual capture histories for tigers, leopards and clouded leopards were developed in a standard ‘X-matrix format’ (Otis et al. 1978) and these were analyzed using models developed for closed populations within the programs CAPTURE (Rexstad and Burnham 1991) and MARK (White 2008). The abundance of all three species for the effective sampled area was estimated by using a buffer around the locations, determined using half the mean of the maximum distance moved (HMMDM) model, and the area associated also with a trapping grid polygon (Karanth and Nichols 1998). A habitat mask was also used to remove non-tiger habitat (e.g., deep water, human settlements; Karanth and Nicols 2002). The density (per 100 km2) was then estimated by dividing this area by the population size (number of individuals). The density of individual prey species, which were also recorded through the camera traps, was estimated using the program DISTANCE (Thomas et al. 2010). This online software works by using detection functions that model the probability of detecting an animal given its distance from the transect. Further details of both methods can be found in Jhala et al. (2010).
Spatial Analysis of Field Data
The spatial configuration of home ranges of individual cats is an important property of their social systems and has been defined as an area traversed by individuals during normal activities of food gathering, mating and caring for young (Dillon and Kelly 2007). For this study, the GPS locations for each capture event and for identified individuals were extracted and plotted spatially. Where an animal was photographed, the distance from the camera trap was determined and a minimal-bounding polygon was constructed around each of the individual point locations where the same individual was sighted and also for all species of the same type. The resulting spatial datasets were then used to quantify ranging behavior, niche overlap and movement in relation to observed land cover types and landscape dynamics. Home range size is considered an important predictable aspect of an animal’s feeding strategy and has been related to food density, metabolic needs and the efficiency of movement. The metric also indicates the degree of overlap as a function of territory and can reflect the variation in population and community characteristics. For example, home-range size is frequently correlated inversely with population density and home-range exclusiveness can indicate significant inter and intra-specific competition (ibid). To calculate the ranging patterns of tigers and sympatric carnivores within the habitat, a point density tool was used which calculates a magnitude per unit area from point features falling within a neighborhood around each cell. In addition, a minimum boundary geometry tool was used, which represents a specified minimum bounding geometry enclosing each input feature for each group of input features.
Land Cover Classification
To classify land cover and associated changes between the different years, 150 training areas representing 14 land cover categories (see Table 5) were established, with a further 150 collected to subsequently assess the accuracy of the classification. A hybrid supervised classification of the Landsat ETM+ data (all spectral bands) was then undertaken within Erdas Imagine 9.3.
Carnivore Distribution in Relation to Anthropogenic Disturbance
Fires, roads and settlements were considered to be the main anthropogenic disturbances likely to influence the distribution and movement of large cats within the landscape. The point distances tool available in the ArcGIS Geospatial Modelling Environment (GSM; http://www.spatialecology.com/gme and previously known as Hawths tools) was used to calculate the distances from tiger observations to fire locations over the observation period. This tool, which was designed to produce flexible distance matrix outputs, calculates distances between points. The output of distance between each fire location and species capture point was then categorized into 500 m intervals and analyzed using simple regression analysis. The null hypothesis that there was no link between fire and carnivore occurrence was rejected where p > 0.05.
Results
Camera Trap Sightings
A total of 14 tigers (8 males and 6 females) were identified from the camera trap photographs (Table 6, Fig. 2). Other top carnivores sighted during the same survey period included 27 leopards (Panthera pardus; 11 males, 13 females, 3 unidentified) and 16 clouded leopards (Neofelis nebulosa; 4 males, 5 females, 7 unidentified) (Fig. 3). Others species that were photographed included the marbled cat (Pardofelis marmorata), Leopard Cat (Prionailurus bengalensis), Jungle Cat (Felis chaus), Dhole (Cuon alpinus), Himalayan Black Bear (Ursus thibetanus), Sloth Bear (Melursus ursinus), Jackal (Canis aureus) and Civet species. Herbivore prey species photo captured included gaur (Bos gaurus), wild pig (Sus scrofa), sambar (Rusa unicolor), hog deer (Hyelaphus porcinus) and barking deer (Muntiacus muntjac) (Borah et al. 2012).

Camera trap pictures of individual tigers in the IBMTCL (Nov 2010–Feb 2011; Source: Forest Department, Manas and Borah et al. 2012)

Camera trap pictures of some Leopards and Clouded Leopards in the IBMTCL (Nov 2010–Feb 2011; Source: Forest Department, Manas and Borah et al. 2012)
Ranging Pattern and Seasonal Home Range
Within Manas NP, tigers were found to be highly territorial (Fig. 2), with males sharing their territory with only 1 or 2 females. However, one adult male tiger (Tiger 1; and possibly the dominant) shared its territory with almost all the other individuals. The range of one female was significantly greater than that of others (Table 7), which may be attributed to the sub-optimal habitats in her territory. During the observation period, all tigers were moving to the areas of higher elevation to the north (i.e., towards Bhutan). No tigers were observed near the boundary of Manas NP, with most occurring towards the central area.
Leopards and Clouded Leopards shared much of their territory with tigers, although their movement was more restricted to the fringe areas. This also corroborates with studies elsewhere (e.g., Harihar et al. 2011) that sympatric species of carnivores co-exist in sub-optimal habitats (Fig. 4).

Ranging pattern of female tigers in relation to the movement of the dominant male v(Tiger 1) in IBMTCL (Camera trap Area = 300 km2)
Land Cover Classifications and Carnivore Preferences
The classification of land covers for the IBMTCL is illustrated in Fig. 5. The majority of the lowland was occupied by closed moist mixed deciduous forests, with subtropical evergreen forest dominating the lower slopes and progressing into temperate broadleaved and coniferous forests and then subalpine vegetation with increasing elevation. Grasslands were found primarily in the south of the IBMTCL and riverine vegetation and unconsolidated deposits occurred along the wide river channels. Based on standard confusion matrices, the overall accuracy in the classification of land covers was 81.5% (kappa coefficient of 0.78), with the majority of users’ or producers’ accuracies exceeding 80% but being as low as 40% for some classes (e.g., user’s accuracy 60% for cultivation and scrub, 56.7% for closed moist mixed forests). Manas NP supported many of the habitats suitable for tigers including the moist mixed deciduous forests (36.5%), semi-evergreen forest (19.1%), riverine sandy areas (15.1%) and moist alluvial grassland (8.4%). During the observation period, female tigers were most commonly recorded in the closed moist deciduous forests, whereas males were also recorded in sandy riverine areas close to forests. Although considered suitable because of the large numbers of prey, both sexes were not recorded within the tall savanna grasslands but rather remained on the periphery. Leopards and clouded leopards were observed primarily in the riverine areas.

Land cover classification of the IBMTCL generated from 2010 Landsat TM data
Distributions of Tigers in Relation to Fires and Anthropogenic Disturbance
The Manas NP section of the IBMTCL landscape experienced repeated and extensive burning between 2000 and 2012, with 1749 fire events recorded in the MOD 14 product. The greatest number of fires was in 2011, with 271 recorded (Fig. 6a). In all years, there was considerable intra-annual variability, with the period December to March associated with the highest frequency of fires. The transition period before and after the monsoon season (May to September) (i.e., the first and last 2 months of the dry season) experienced fewer fires. Manas NP is divided into three ranges; the western range (Panbari), central (Bansbari) and eastern (Bhuyanpara), and the density of fires (5.13 km−2) was greatest in the grassland areas of the Bansbari range, with those in the Kuribeel and Pahu field areas repeatedly burnt. The incidence of fires was greatest in the more open moist mixed deciduous forests (Fig. 6b; Table 8), with this attributed largely to human activities. Over the period of the camera trapping, 32 fires were detected in the areas within which camera traps were placed. Reference to the tiger data indicated that the highest number of counts (indicating presence) was at a distance of 5–10 km from fires and often around the margins of burnt areas (Fig. 7). In general, tigers were recorded away from roads and village settlements, with this indicating their sensitivity to human activity, including vehicular movements.

The frequency of fires in the IBMTCL between 2000 and 2012 as determined from MODIS data

The distribution of tiger observations in relation to fire density in the Manas NP section of the IBMTCL
Discussion
The Role of Earth Observation
The main benefit of using earth observation data was the ability to provide a land cover classification for a similar period as the camera trap surveys. The overall accuracy of the land cover classification for the IBMTCL was relatively high, but there was confusion between a number of classes, largely because of the high seasonal variation in the phenology of vegetation but also fires and flooding. In some areas, dry areas became inundated during the wet season, but as the waters receded, extensive areas of highly productive grassland established. These became senescent as the dry season progressed and many were then subject to extensive burning. Hence, the spectral signatures of the same area of ground varied considerably (senescent and productive grassland, water and burnt ground). For this reason, the use of dense time-series of optical remote sensing data would provide a better overview of the changing states of vegetated and non-vegetated environments within the IBMTCL. Whilst atmospheric correction of the Landsat sensor data was undertaken, smoke haze and cloud still limited reliable retrieval of surface reflectance for some areas.
A further limitation of the land cover maps was that insufficient information on the structural characteristics of forests that might influence tiger movements was obtained, particularly in relation to the tree canopy density and canopy height. A common approach to estimating canopy cover is to establish relationships between ground based measurements and combinations of reflectance data. However, the lack of ground data and seasonal changes in leaf cover prevented the generation of reliable retrieval algorithms or use of those that are existing. Information on canopy height was also not obtained although there is potential to retrieve this using NASA’s Ice, Cloud, and land Elevation (ICESat) satellite Geoscience Laser Altimeter System (GLAS). Future mapping could therefore focus on developing these products and applying classification schemes that consider their inclusion such as the Food and Agricultural Organisation’s (FAO) Land Cover Classification System (LCCS; see Lucas and Mitchell (2017; this book). The MODIS data provided invaluable insights into the frequency and distribution of fires, identifying that the majority occurred in the moist mixed deciduous forests and alluvial grasslands.
Distribution and Movement of Tigers
Although camera trapping occurred over a relatively short period, 52 tiger sightings were obtained. The resulting photographs allowed 14 different individuals to be distinguished and their gender determined. The movement of tigers was variable but several individuals moved large distances (tens of kilometers), with one male having a substantive range (232 km2). A general movement of tigers towards forested areas in the northern sections (of slightly higher elevation) may be attributable to the comparatively low availability of prey in the grassland areas during the winter months.
Reference to the land cover classification generated from Landsat sensor data indicated that the majority of tigers remained within the forest areas, although some males ventured into the riverine areas. Whilst recognizing that the observation period was relatively short, it was evident that some tigers remained in forest areas in proximity to the fires but were also sufficiently far away (typically 5–10 km) from the last known burn. It is hypothesized that tigers may stay close to areas that are burning (or may be susceptible to burning) in order to take advantage of prey species fleeing from the fire. They may also anticipate or take advantage of the prey species returning to graze on new growth in the periods following the fires. The influence of fires on the distribution of large cats and their prey has also been noted by Sarma et al. (2008) and Takahata et al. (2010) but further studies are need to specifically link occurrence and movements with fire activity. Avoidance of areas occupied or influenced by humans was noticeable, with many individuals located towards the less disturbed central areas where human activity and vehicular movements were minimal and prey was more abundant.
A practical outcome from the study is that information generated on individual movement of tigers can assist future surveillance, especially as the likelihood of tiger movements can be communicated to the forest camps. Training in the use of camera traps and photographic interpretation allows frontline staff to identify individual tigers and contribute to a better understanding of their ranging behaviour in this landscape. Whilst tigers may benefit from fires, they may also be disturbed from those that are human-induced and therefore the establishment of camps in close proximity to fire prone areas may lead to prevention of many and hence less overall disturbance of habitats.
The tigers showed minimal overlap with leopards and clouded leopards and some studies (e.g., Rabinowitz et al. 1987) have suggested that the clouded leopard may alter its range or become less nocturnal when larger competitive species occur. Further research on the movement of tigers and other large cats based on camera trap surveys is nevertheless advocated in order to better determine their interactions and the distribution of their prey. The camera traps also provide a basis for better estimating the abundance and density of large cats and to therefore monitor changes and trends in populations and their behaviour.
When used in combination with camera trapping and other monitoring techniques, earth observation data can play a role in understanding how large carnivores utilize the IBMTCL during different times of the year. These data can also be used to address connectivity issues within and between landscapes. Several new initiatives have included the development of software (e.g., MIST, SMART, M-STrIPES) that facilitate spatial monitoring of tiger populations including through the integration of field observations. Within such systems, there is considerable opportunity to make better use of the large, extensive and publically available archives of satellite sensor data.
Final Conclusions
In 1998, the global tiger population was estimated at 5000–7000 individuals (Seidensticker and McDougal 1993). By 2014, however, the global tiger population was at an historic low of approximately 3500 individuals, having declined by more than 50% of what they were just a few decades earlier (Goodrich et al. 2015). Given this situation, there is an urgent need to prioritize efforts to protect the remaining tiger populations (Stokes 2010; Walston et al. 2010). For the most part, these are found in PAs (Chapron et al. 2008; Rabinowitz 2009; Clements et al. 2010; Walston et al. 2010) but there are considerable opportunities to expand their range, including by connecting areas where they occur. This study has highlighted the potential of using earth observation data in conjunction with ground-based (including camera trap) surveys for better understanding the distribution, movement and preferences of tigers and a recommendation is to develop what is termed a Tiger Observation System (TOS) to maximize these opportunities. Whereas past use of earth observation data has been limited by availability and cost, the release of the Landsat sensor archive and the provision of freely available Sentinel-1 and -2 radar and optical sensor data respectively gives tiger conservation efforts additional impetus to achieve their long term conservation and expansion of populations.
References
Ahmed, M.F., Borah, J., Das, C., Basumatary, A., Sarma, R.N., Gogoi, D.D., Buragohain, S.N., Vasu, N.K., Talukdar, B.K., Jhala, Y.V., Qureshi, Q.: Monitoring Tigers and Prey Animals of Kaziranga National Park, Assam, India. Technical Report, Aaranyak. 50 pp (2010)
Balme, G.A., Hunter, L.T.B., Slotow, R.: Evaluating methods for counting cryptic carnivores. J. Wildl. Manag. 73, 433–441 (2009)
Blake, S., Hedges, S.: Sinking the flagship: The case of forest elephants in Asia and Africa. Conserv. Biol. 18, 1191–1202 (2004)
Borah, J., Wangchuk, D., Swargowari, A., Wangchuk, T., Sharma, T., Das, D., Rabha, N., Basumatari, A., Kakati, N., Ahmed, M.F., Sharma, A., Sarmah, A., Dutta, D.K., Lahkar, B., Dorji, T., Brahma, P.K., Ramchiary, L., Tempa, T., Wangdi, Y., Nedup, T., Wangdi, T., Tharchen, L., Dhendup, P., Bhobora, C.R., Pandav, B., Vattakaven, J.: Tigers in Indo-Bhutan Transboundary Manas Conservation Complex. 2012. WWF Technical report (2012)
Bunting, P., Clewley, D., Lucas, R.M., Gillingham, S.: The Remote Sensing and GIS Software Library (RSGISLib). Comput. Geosci. 62(2–6), 226 (2013)
Carbone, C., Mace, G.M., Roberts, S.C., Macdonald, D.W.: Energetic constraints on the diet of terrestrial carnivores. Nature. 402, 286–288 (1999)
Carbone, C., Christie, S., Conforti, K., Coulson, T., Franklin, N., Ginsberg, J.R., Griffiths, M., Holden, J., Kawanishi, K., Kinnaird, M., Laidlaw, R., Lynam, A., Macdonald, D.W., Martyr, D., McDougal, C., Nath, L., O’Brien, T., Seidensticker, J., Smith, D.J.L., Sunquist, M., Tilson, R., Shahruddin, W.N.W.: The use of photographic rates to estimate densities of tigers and other cryptic mammals. Anim. Conserv. 4, 75–79 (2001)
Carroll, C., Miquelle, D.G.: Spatial viability analysis of Amur tiger Panthera tigris altaica in the Russian Far East: the role of protected areas and landscape matrix in population persistence. J. Appl. Ecol. 43, 1056–1068 (2006)
Chapron, G., Miquelle, D.G., Lambert, A., Goodrich, J.M., Legendre, S., Clobert, J.: The impact on Tigers of poaching versus prey depletion. J. Appl. Ecol. 45, 1667–1674 (2008)
Choudhury, S.R.: Let us count our Tiger. Cheetal. 14(2), 41–51 (1970)
Clements, R., Rayan, D.M., Zafir, A.W.A.: Trio under threat: can we secure the future of Rhinos, Elephants and Tigers in Malaysia. Biodivers. Conserv. 19, 1115–1136 (2010)
Davis, B.W., Li, G., Murphy, W.J.: Supermatrix and species tree methods resolve phylogenetic relationships within the big cats, Panthera (Carnivora: Felidae). Mol. Phylogenet. Evol. 56(1), 64–76 (2010)
Dillon, A., Kelly, M.J.: Ocelot Leopardus pardalis in Belize: the impact of trap spacing and distance moved on density estimates. Oryx 41(04), 469–477 (2007)
Dinerstein, E., Loucks, C., Wikramanayake, E., Ginsberg, J., Sanderson, E., Seidensticker, J., Forrest, J., Bryja, G., Heydlauff, A., Klenzendorf, S., Leimgruber, P., Mills, J., O’Brien, T.G., Shrestha, M., Simons, R., Songer, M.: The fate of wild tigers. Bioscience. 57, 508–514 (2007)
Dorji, D.P., Santiapillai, C.: The status, distribution and conservation of the Tiger Panthera tigris in Bhutan. Biol. Conserv. 48, 311–319 (1989)
Gittleman, J.L., Harvey, P.H.: Carnivore home-range size, metabolic needs and ecology. Behav. Ecol. Sociobiol. 10, 57–63 (1982)
GOI: Joining the Dots-The report of the Tiger Task Force, Project Tiger, Government of India. 217 pp (2005)
Goodrich, J., Lynam, A., Miquelle, D., Wibisono, H., Kawanishi, K., Pattanavibool, A., Htun, S., Tempa, T., Karki, J., Jhala, Y., Karanth, U.: Panthera tigris. The IUCN Red List of Threatened Species 2015: e.T15955A50659951. http://dx.doi.org/10.2305/IUCN.UK.2015-2.RLTS.T15955A50659951.en. Downloaded on 02 June 2017 (2015)
Gratwicke, B., Bennett, E.L., Broad, S., Christie, S., Dutton, A., Gabriel, G., Kirkpatrick, C., Nowell, K.: The world can’t have wild tigers and eat them, too. Conserv. Biol. 22, 222–223 (2008)
Harihar, A., Pandav, B., Goyal, S.P.: Subsampling photographic capture-recapture data of tigers (Panthera tigris) to minimize closure violation and improve estimate precision: a case. Popul. Ecol. 51, 471–479 (2009)
Harihar, A., Pandav, B., Goyal, S.P.: Responses of leopard Panthera pardus to the recovery of a tiger Panthera tigris population. J. Appl. Ecol. 48(3), 806–814 (2011)
Homer, C.G., Ramsey, R.D., Edwards, T.C., Falconer, A.: Landscape cover type mapping and modelling using multi-scene thematic mapper mosaic. Photogramm. Eng. Remote Sens. 63, 59–67 (1997)
Imam, E., Kushwaha, S.P.S., Singh, A.: Evaluation of suitable tiger habitat in Chandoli National Park, India, using multiple logistic regression. Ecol. Model. 220, 3621–3629 (2009)
Jhala, Y. V., Gopal, R., Qureshi, Q.: Status of tigers, co-predators and prey in India. National Tiger Conservation Authority, Government of India and the Wildlife Institute of India, New Delhi and Dehra Dun (2008)
Jhala, Y.V., Qureshi, Q., Gopal, R., Sinha, P.R. (eds.): Status of the tigers, co-predators and prey in India, 2010. National Tiger Conservation Authority, Govt of India and Wildlife Institute of India, New Delhi and Dehra Dun (2011)
Jhala, Y.V., Qureshi, Q., Gopal, R.: The status of tigers in India. National Tiger Conservation Authority, Wildlife Institute of India, New Delhi and Dehra Dun (2015)
Karanth, K.U., Sunquist, M.E.: Prey selection by Tiger, Leopard and Dhole in Tropical Forests. J. Animal Ecol. 64, 439–450 (1995)
Karanth, K.U., Nichols, J.D.: Estimation of tiger densities in India using photographic captures and recaptures. Ecology 79(8), 2852–2862 (1998)
Karanth, K.U., Nichols, J.D.: Monitoring tigers and their prey: a manual for researchers, managers and conservationists in tropical Asia. Centre for Wildlife Studies, Bangalore (2002)
Karanth, K.K., Nichols, J.D., Hines, J.E., Karanth, K.U., Christensen, N.L.: Patterns and determinants of mammal species occurrence in India. J. Appl. Ecol. (2009)
Karanth, K.U., Nichols, J.D., Kumar, N.S., Hines, J.E.: Assessing tiger population dynamics using photographic capture-recapture sampling. Ecology. 87, 2925–2937 (2006)
Kelly, M.J., Noss, A.J., Di Bitetti, M.S., Maffei, L., Arispe, R.L., Paviolo, A., De Angelo, C.D., Di Blanco, Y.E.: Estimating puma densities from camera trapping across three study sites: Bolivia, Argentina, and Belize. J. Mammal. 89(2), 408–418 (2008)
Kerley, L.L., Salkina, G.P.: Using scent-matching dogs to identify individual Amur tigers from scats. J. Wildl. Manag. 71(4), 1349–1356 (2007)
Kerley, L.L., Goodrich, J.M., Miquelle, D.G., Smirnov, E.N., Quigley, H.B., Hornocker, M.G.: Reproductive parameters of wild female Amur (Siberian) tigers (Panthera tigris altaica). J. Mammal. 84, 288–298 (2003)
Linkie, M., Chapron, G., Martyr, D.J., Holden, J., Leader-Williams, N.: Assessing the viability of tiger subpopulations in a fragmented landscape. J. Appl. Ecol. 43, 576–586 (2006)
Luo, S.J., Kim, J.H., Johnson, W.E., van der Welt, J., Martenson, J., Yuhki, N., Miquelle, D.G., Uphyrkina, O., Goodrich, J.M., Quigley, H.B., Tilson, R., Brady, G., Martelli, P., Subramaniam, V., McDougal, C., Hean, S., Huang, S.Q., Pan, W.S., Karanth, U.K., Sunquist, M., Smith, J.L.D., O’Brien, S.J.: Phylogeography and genetic ancestry of tigers (Panthera tigris). PLos Biol. 2, 2275–2293 (2004)
MacKenzie, D.I., Nichols, J.D., Lachman, G.B., Sam, D., Royle, J.A., Langtimm, C.A.: Estimating site occupancy rates when detection probabilities are less than one. Ecology 83(8), 2248–2255 (2002)
Nowell, K.: Deciding the Tiger’s future: Implementation of CITES decision 14.65–14.72, In 58th meeting of the CITES Standing Committee. Geneva (2009)
O’Brien, T.G., Kinnaird, M.F., Wibisono, H.T.: Crouching tigers, hidden prey: Sumatran tiger and prey populations in a tropical forest landscape. Anim. Conserv. 6, 131–139 (2003)
Otis, D.L., Burnham, K.P., White, G.C., Anderson, D.R.: Statistical-inference from capture data on closed animal populations. Wildl. Monogr. 62, 7–135 (1978)
Rabinowitz, A.: Stop the bleeding: implementing a strategic tiger conservation protocol. CAT News. 51, 30–31 (2009)
Rabinowitz, A., Andau, P., Chai, P.P.: The clouded leopard in Malaysian Borneo. Oryx. 21(02), 107–111 (1987)
Ranganathan, J., Chan, K.M.A., Karanth, K.U., Smith, J.L.D.: Where can tigers persist in the future? A landscape-scale, density-based population model for the Indian subcontinent. Biol. Conserv. 141(1), 67–77 (2008)
Rangarajan, M.: India’s Wildlife history: an introduction. Delhi, Permanent Black (2001)
Rexstad, E., Burnham, K.P.: User’s guide for interactive program CAPTURE. Color. Cooperative Fish and Wildlife Research Unit (1991)
Royle, J.A., Karanth, K.U., Gopalaswamy, A.M., Kumar, N.S.: Bayesian inference in camera trapping studies for a class of spatial capture-recapture models. Ecology. 90, 3233–3244 (2009)
Samrat, M., Karanth, K.U., Kumar, N.S., Gopalaswamy, A.M., Andheria, A., Ramakrishnan, U.: Evaluation of non-invasive genetic sampling methods for estimating tiger population size. Biol. Conserv. 142(10), 2350–2360 (2009)
Sanderson, E., Forrest, J., Loucks, C., Ginsberg, J., Dinerstein, E., Seidensticker, J., Leimgruber, P., Songer, M., Heydlauff, A., O’Brien, T., Bryja, G., Klenzendorf, S., Wikramanayake, E.: Setting priorities for the conservation and recovery of Wild Tigers: 2005–2015. The technical assessment. WCS, WWF, Smithsonian, and NFWF-STF, New York and Washington, DC, USA (2006)
Sarma, P.K., Lahkar, B.P., Ghosh, S., Rabha, A., Das, J.P., Nath, N.K., Dey, S., Brahma, N.: Land-use and land-cover change and future implication analysis in Manas National Park, India using multi-temporal satellite data. Curr. Sci. 95, 223–227 (2008)
Schaller, G.B.: The deer and the tiger- a study of wildlife in India. University of Chicago Press, Chicago (1967)
Seidensticker, J., McDougal, C.: Tiger predatory behavior, ecology and conservation. Mamm. Predators. 65, 105–125 (1993)
Sharma, S., Jhala, Y., Sawarkar, V.B.: Identification of individual tigers (Panthera tigris) from their pugmarks. J. Zool. 267(1), 9–18 (2005)
Smith, J.L.D., Ahearn, S.C., McDougal, C.: Landscape analysis of tiger distribution and habitat quality in Nepal. Conserv. Biol. 12, 1338–1346 (1998)
Stokes, E.J.: Improving effectiveness of protection efforts in tiger source sites: Developing a framework for law enforcement monitoring using MIST. Integr. Zool. 5, 363–377 (2010). doi:10.1111/j.1749-4877.2010.00223.x
Sunquist, M., Karanth, K.U., Sunquist, F.: Ecology, behaviour and resilience of the tiger and its conservation needs. In: Seidensticker, J., Christie, S., Jackson, P. (eds.) Riding the Tiger: Tiger Conservation in Human-Dominated Landscapes, pp. 5–18. Cambridge University Press, London (1999)
Takahata, C., Amin, R., Sarma, P., Banerjee, G., Oliver, W., Fa, J.E.: Remotely-sensed active fire data for protected area management: eight-year patterns in the Manas National Park. India. Environ. Manag. 45, 414–423 (2010)
Thomas, L., Buckland, S.T., Rexstad, E.A., Laake, J.L., Strindberg, S., Hedley, S.L., Bishop, J.R., Marques, T.A., Burnham, K.P.: Distance software: design and analysis of distance sampling surveys for estimating population size. J. Appl. Ecol. 47(1), 5–14 (2010)
UNESCO: World Heritage Centre-IUCN: Mission report. Reactive monitoring mission to the Manas Wildlife Sanctuary, India From 12 to 18 February 2008. Available from: http://whc.unesco.org/en/list/338/documents/. (2008)
Walston, J., Robinson, J.G., Bennett, E.L., et al.: Bringing the tiger back from the brink: The six percent solution. PLoS Biol. 8, e1000485 (2010)
Wangda, P., Ohsawa, M.: Gradational forest change along the climatically dry valley slopes of Bhutan in the midst of humid eastern Himalaya. Plant Ecol. 186(1), 109–128 (2006)
Wegge, P., Storaas, T.: Sampling tiger ungulate prey by the distance method: lessons learned in Bardia National Park, Nepal. Anim. Conserv. 12, 78–84 (2009)
White, G.C.: Closed population estimation models and their extensions in program MARK. Environ. Ecol. Stat. 15(1), 89–99 (2008)
Wikramanayake, E.D., Dinerstein, E., Robinson, J.G., Karanth, U., Rabinowitz, A., Olson, D., Mathew, T., Hedao, P., Conner, M., Hemley, G., Bolze, D.: An ecology-based method for defining priorities for large mammal conservation: the tiger as case study. Conserv. Biol. 12, 1427–1427 (1998)
Acknowledgements
The authors would like to thank the Honourable Deputy Minister, Bodoland Territorial Areas District Shri Kampa Borgayari and his officials, Shri G.C.Basumatary, Shri A.Swargowari, Assam Forest Department for giving the requisite permissions to carry out the study. Dr. Jimmy Borah and his team from WWF, Dr. Bibhuti Lahkar from Aaranyak, and the Park Authorities of India and Bhutan are thanked for providing the field data. Dr. P.S. Roy, Dr. SPS Kushwaha and Dr. Arijit Roy from Indian Institute of Remote Sensing are also thanked for their technical help and support.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2017 Springer International Publishing AG
About this chapter

Cite this chapter
Ghosh, S., Lucas, R. (2017). Coupling Field Sampling with Earth Observation Increases Understanding of Tiger Movement and Behaviour. In: Díaz-Delgado, R., Lucas, R., Hurford, C. (eds) The Roles of Remote Sensing in Nature Conservation. Springer, Cham. https://doi.org/10.1007/978-3-319-64332-8_12
Download citation
DOI: https://doi.org/10.1007/978-3-319-64332-8_12
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-64330-4
Online ISBN: 978-3-319-64332-8
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)