
Abstract
Initial experiments using Mediterranean Ni-hyperaccumulator plants for the purpose of phytomining were carried out in the 1990s. In order to meet commercial phytoextraction requirements, a technology has been developed using hyperaccumulator species with adapted intensive agronomic practices on natural Ni-rich soils. Ultramafic soils in the Balkans display a great variability in Ni concentrations and available Ni levels, both in Albania and the Pindus Mountains of Greece. In Albania, Vertisols are currently being used for low-productivity agriculture (pasture or arable land) on which phytomining could be included in cropping practices. Alyssum murale occurs widely on these ultramafic Vertisols and is a spontaneous weed that grows among other crops. This review chapter presents the different steps that were investigated during the study of soil suitability, and selection of plants up to optimization of agronomic practices, at field scale, as recently developed to reach the implementation stage of Ni agromining in Albania. During a 7-year study we addressed the following questions: (i) what are the optimal soils for Ni agromining in terms of fertility and Ni availability? (ii) what is the phytoextraction potential of local populations of Ni hyperaccumulator species? (iii) what should be the agronomical practices used to optimize the cropping of A. murale for extensive phytomining adapted to a Balkan agricultural setting?
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others


1 Introduction
Ultramafic soils cover large areas in the Balkans, more than in any other part of Europe. Therefore, this region is a potential target for agromining activities (van der Ent et al. 2015). However, no woody and perennial species such as Rinorea bengalensis or Phyllanthus securinegioides from Southeast Asia can be cropped under Mediterranean conditions (van der Ent et al. 2015; Nkrumah et al. 2016). Moreover, the Balkans also has the highest diversity in Ni hyperaccumulator plants in Europe and is home to the widespread plant A. murale, one of the most studied species worldwide for phytomining (e.g. Nkrumah et al. 2016). Initial experiments using Mediterranean Ni-hyperaccumulators for the purpose of phytomining were carried out in the 1990s. In order to meet commercial phytoextraction requirements, technology has been developed using hyperaccumulator plant species with adapted intensive agronomic practices on natural Ni-rich soils. In a first stage, several studies have been carried out in order to identify the best soil conditions to establish field trials. Then the native species were evaluated for their potential in phytomining; finally, trials were set to optimize cropping conditions in the Balkan context.
2 Agromining Nickel in The Balkans
2.1 Properties of Ultramafic Soils the Balkans and Suitability for Agromining
Outcrops of ultramafic rocks in the Balkans (Dilek and Furnes 2009) extend towards central Bosnia, western and central Serbia, southern Bulgaria, and north-central and southeastern Albania to the ultramafic formations of Epirus and Thessalia in Greece (Fig. 1; Stevanovič et al. 2003). The number of Balkan endemics growing on ultramafic-derived soils comprises approximately 335 taxa, of which 123 are obligate. The most floristically rich areas are situated in northwestern Greece (Epirus), the island of Evvia, and northern Albania, together with southwestern Serbia and northern Greece (Mount Vourinos). The Pindus Mountains are located in northwestern Greece and extend about 160 km from the southern border of Albania. Ultramafic terranes in Albania and the Pindus Mountains constitute the largest blocks of ultramafic outcrops in the Balkans. The ouctcrops are also part of similar floristic assemblages: northeastern and southeastern Albania to northwestern Greece (Reeves et al. 1983; Shallari et al. 1998; Tan et al. 1999; Stevanovič et al. 2003), located in similar climate zones (a Mediterranean climate with montane influences). The ultramafic soils of Albania and the Pindus Mountains (Greece) contain elevated levels of metals such as Ni, Cr, Co and Fe. Most soil cover on these ultramafic materials falls into the Cambisol type (Echevarria, Chap. 8 in this book). Surface horizons of the soils are characterized by extremely high Mg/Ca quotients of up to 30 in Albania and up to 17.7 in Greece (Bani et al. 2009, 2010), typical of ultramafic soils worldwide (Reeves et al. 1997). Concentrations of Co in these soils are relatively high, also typical of Mediterranean ultramafic soils (Reeves et al. 1997; Wenzel and Jockwer 1999). Cobalt concentrations vary between 93 μg g−1 (Malakasi-Soil 2) and 280 μg g−1 (Pojska Soil 2); Cr concentrations in soils range from 667 μg g−1 (Pojska-Soil 1) to 3250 μg g−1 (Pishkash). The soils with the highest Ni contents come from eastern Albania and have ca. 3000 μg g−1. The Ni concentrations in soil samples from the Gjegjan area, in northern Albania, vary from 1070 to 2580 μg g−1. In the Greek soils, Ni is also lower than in eastern Albania: between 1280 to 2660 μg g−1. The soils of both areas have a similar geological setting, varying from partly serpentinized peridotite (harzburgite) to serpentinite (Bani et al. 2014; Estrade et al. 2015).
The nature of Ni-bearing phases and the overall soil mineralogy strongly influence Ni availability. Because soil mineralogy depends on pedogenetic properties, one can say that Ni availability depends on soil genesis. As observed for other soils, the fate of Ni is strongly related to the ‘free’ status of Fe in the soils (Massoura et al. 2006; Chardot et al. 2007; Cheng et al. 2011; Raous et al. 2013); ‘free’ Ni partition is strongly related to the content of amorphous Fe-oxides in the soils. In Albanian ultramafic soils, DTPA-Ni ranges from 81 to 285 μg g−1. In the Pindus Mountain Cambisols, DTPA-extracted Ni in soils varies from 48 to 126 μg g−1 (Bani et al. 2009). The pH values of the Greek ultramafic soils (6.3–7.0) are in general lower than those in the Albanian soils (6.7–8.2), probably because of the higher altitudes of the former (Bani et al. 2009). In this case, there is a direct effect of pH on DTPA-Ni extractability. However, Ni partitioning among mineral phases and its speciation play the most important roles within these ultramafic soils (Massoura et al. 2006). In particular, the association of Ni with amorphous Fe-oxides (Cambisols and other Cambic soils) and with secondary high-exchange clays (Vertisols and Saprolites) are the main factors determining Ni phytoavailability in these ultramafic soils (Massoura et al. 2006; Bani et al. 2014). In our study, soils from the Pindus are shown to be the least weathered from the point of view of Fe speciation. Nickel is clearly associated with unweathered clay minerals and amorphous Fe-oxides in the Cambisols of the entire region. The soil having the highest amount of Ni associated with the amorphous Fe-oxides is unsurprisingly the soil with the highest chemical availability of Ni (Pojska, Albania). Vertisols were selected among all candidates as the target soils for agromining in these regions, because of high Ni phytoavailability, and position in the landscape (downslope soils and alluvial valleys) that conditions water availability to crops and easy access to agricultural machinery. In Albania, ultramafic outcrops cover 10% of the surface, and the Mg-rich arable Vertisols have been estimated to cover about 10,000 ha of the 700,000 ha of the total agricultural land available in the country (Zdruli 1997). These Vertisols are currently being used for low-productivity agriculture on which agromining could be included in management practices (Bani et al. 2007).
2.2 Nickel Hyperaccumulation by Plant Species of the Balkans
Ultramafic flora and vegetation have attracted much interest from botanists in those parts of the world where significant ultramafic terranes occur. The largest number of Ni-hyperaccumulators is found in the Brassicaceae family in temperate climates, especially Mediterranean Europe and Turkey (Reeves and Adıgüzel 2008). Hyperaccumulation of Ni is defined as accumulation in aerial parts up to concentrations of >1000 μg g−1 on a dry-matter (DM) basis (Brooks et al. 1977; Reeves 1992; van der Ent et al. 2013). This natural process was first discovered in Alyssum bertolonii (Brassicaceae) in Italy (Minguzzi and Vergnano 1948). Subsequently, similar reports have been given for A. murale in Armenia (Doksopulo 1961), and A. serpyllifolium ssp. lusitanicum in Portugal (Menezes de Sequeira 1969). Later investigations revealed that Ni hyperaccumulation is widespread among Alyssum species on ultramafics of Mediterranean Europe, Turkey, and adjacent countries (Brooks et al. 1979; Reeves and Adıgüzel 2008). Nearly 50 taxa of Alyssum, all in Sect. Odontarrhena (now accepted as a separate genus by Španiel et al. 2015), act as Ni hyperaccumulators (Brooks 1987; Baker and Brooks 1989, Reeves and Adıgüzel 2008). Nickel hyperaccumulation in the Brassicaceae of the Balkan countries occurs not only in Alyssum species, but also in species of the genera Thlaspi s.l. (Noccaea), Bornmuellera, and Leptoplax (Peltaria). Two additional Ni hyperaccumulators are reported from other families: the Asteraceae (Centaurea thracica and the Violaceae (Viola vourinensis) (Psaras and Constantinidis 2009).
Thirteen Alyssum hyperaccumulator taxa are distributed on the Balkan Peninsula (Bani et al. 2010, 2013). This number is a significant proportion of Alyssum hyperaccumulators recorded in Europe (Cecchi et al. 2010), suggesting that the Balkan Peninsula is a ‘hotspot’ for ultramafic flora of the continent. Although this number of Ni hyperaccumulators will be reduced after new molecular evidence and accepted taxonomy of the genus (Cecchi et al. 2010; Španiel et al. 2015), it is higher than in other parts of Europe. The majority of Ni hyperaccumulators distributed in the Balkans are ultramafic endemics; Greece has the largest number of local endemics. The number of facultative hyperaccumulator species, i.e. hyperaccumulator species that occur naturally on both metalliferous and non-metalliferous soils (Pollard et al. 2014) in the Balkans is small. They occur on a variety of substrates having widely varying Ni concentrations, and show a large range of plant Ni concentrations as a result (Reeves and Adıgüzel 2008). Data for Ni accumulation carried out over the last 30 years on a wide variety of herbarium and field-collected Alyssum species on the Balkan Peninsula are listed in Table 1. The highest Ni concentrations are recorded in A. murale and A. heldreichii (Bani et al. 2010, 2013). The taxonomically complex genus Thlaspi s.l. (divided into many separate genera, Noccaea being the largest) has numerous Ni hyperaccumulator species. The highest Ni concentrations are measured in N. ochroleuca (15–23,400 μg g−1 according to Bani et al. 2010). Facultative Ni-hyperaccumulators (N. ochroleuca, N. praecox and N. kovatsii have Ni-accumulating potential only where growing on ultramafic soils. Measured Ni concentrations in the leaves of the above species from the Balkans are approximately four times higher than those reported for Turkey (Reeves and Adigüzel 2008). Local species endemic to ultramafic soils in Greece are N. epirota, N. graeca and N. tymphaea. Noccaea species are known to hyperaccumulate more than one metal (Cd, Ni, Pb, Zn). Noccaea ochroleuca grown on various substrates shows a wide range of concentrations of both Ni and Zn, each element exceeding 1000 μg g−1 (Reeves and Adigüzel 2008). The genus Bornmuellera is restricted to Greece, Kosovo, Serbia, S. Albania, and Turkey (Marhold 2011). Some of its species are ultramafic-endemics. The highest Ni concentration is reported for Bornmuellera tymphaea from 1590 to 31,200 μg g−1 (Bani et al. 2010). Recently, in the light of available molecular data (Rešetnik et al. 2013) and a re-evaluation of morphological characters, the monotypic genus Leptoplax (previously considered closely related to and even included in Peltaria) has been merged with Bornmuellera (Rešetnik et al. 2014). A local ultramafic-endemic species, L. emarginata, occurring mainly in northern Greece and on the island of Evvia, are reported to have varying Ni concentrations of 2040 to 34,400 μg g−1 (Bani et al. 2010). This species typically forms hybrids with B. baldaccii and B. tymphaea. Psaras and Constantinidis (2009) described two more hyperaccumulator species from the flora of Greece, Centaurea thracica and Viola vourinensis, with 3830 μg g−1 and 1023 μg g−1 Ni, respectively. The leaves remain the most important plant part to harvest for phytomining purposes, although stems have intermediate Ni concentrations and potential for high biomass production (Zhang et al. 2014).
The percentage of leaves in the biomass of different hyperaccumulator species is highly variable, even within a given species, thus being crucial for defining the potential phytoextraction yield. In an Albanian Vertisols site, the mean height of A. murale plants at flowering stage varied from 45 to 93 cm, and for one plant, the percentage of leaves in the whole biomass ranged from 11.6 to −17.3% by weight (Bani et al. 2015b). The highest Ni contents in plants were recorded for the leaves of L. emarginata with more than 30,000 μg g−1 Ni (Zhang et al. 2014). Alyssum murale with more than 20,000 μg g−1 Ni in leaves (Bani et al. 2014) is the predominant species on all ultramafic sites in Albania and Greece (and many other Balkan countries). The highest Ni concentrations recorded in A. murale leaves collected from ultramafic sites of Bulgaria (Rhodope massif) ranged from 5000 μg g−1 (Kardzali) to 1.5 μg g−1 (Kazak) (Bani et al. 2010), whereas for ultramafic sites in Serbia the leaf-hosted Ni contents were 700 to 13,000 μg g−1 (Tumi et al. 2012). No relevant effect of pH on Ni accumulation in Alyssum could be deduced from our results in ultramafic sites of the Balkans (Bani et al. 2009, 2010). The Ni concentration of this species is highly dependent on the site of collection, and greatest in the area of Prrenjas and Pojska in Albania where shoots contain >10,000 μg g−1 (Bani et al. 2010, 2013, 2015a, b). Other Alyssum species have lower Ni concentrations (Bani et al. 2009; Zhang et al. 2014). The concentration of Ni in tissues of A. markgrafii collected at 10 sites in ultramafics of Kosovo was moderately high, ranging from 1586 μg g−1 to 7564 μg g−1 (Salihaj et al. 2016). Distribution of Ni among the different organs of hyperaccumulator plants depends on the biology of the species and can be influenced by edaphic factors such as climate, soil pH, etc. It is clear that the endemic A. markgrafii, several efficient populations of A. murale, L. emarginata, and B. tymphaea (Greece), displayed the best Ni-efficiency for use in phytomining (Chardot et al. 2005; Bani et al. 2009; Zhang et al. 2014).
3 Case Study in Albania: 7-Year Agromining Field Experiments with Alyssum murale
Alyssum murale occurs widely on ultramafic Vertisols (Bani et al. 2009) and is a spontaneous weed to other crops. We consequently started investigating the performance of a phytoextraction system with some agronomic practices to evaluate whether semi-extensive phytoextraction of Ni is feasible on Mg-rich Vertisols. A. murale was the easiest crop to plant, especially because of its high rate of seeding and high frequency of occurrence in ultramafic agricultural lands of Albania. Experiments were undertaken from 2005–2009 and from 2012–2014 on in situ experimental plots. Field experiments (Bani et al. 2015a, b) were aimed at studying (i) the distribution of Ni and other elements in plant organs, (ii) plant nutrition and fertilization, (iii) weed control, (iv) the crop establishment technique (natural cover vs. sown crop), and (v) plant density. Firstly, the cropping of native stands of A. murale was studied for 3 years. After this period, the A. murale cover was renewed by sowing native seeds without density control. Finally, in the last set of experiments, the effect of density was studied from data on 1 to 6 plants per m2. The final aim was to propose a cropping system for A. murale that would optimize agromining in the Balkans.
3.1 Fertilization Regime and Nutrient Requirements
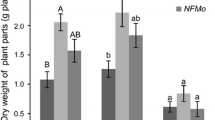
At first, native stands of A. murale were cropped in Pojska from 2005 until 2007 (Bani et al. 2015a). In March 2005, six 36 m2 plots were designed, three of which were fertilized in April with 120 kg ha−1 of N, P and K and 77 kg ha−1 of Ca (ammonium nitrate, potassium sulfate, and calcium phosphate). The three other plots were not fertilized (Bani et al. 2007). In 2006, we used the same fertilization protocol as above, but also treated the three plots with anti-monocot herbicide (FocusTM ultra 33 mL applied in 3 L water sprayed onto 108 m2) (FH); a further three were not treated (NFNH). The optimal fertilization pattern for A. murale had been carefully studied on the same soil in glasshouse pot experiments (Bani et al. 2007; Nkrumah et al. 2016). Plants were harvested for the first and second year at the end of June (before studies on Ni distribution in plant parts according to the phenological stage). In 2007, the fertilization protocol involved 100 kg ha−1 P and K, and for N there was a split application of 120 kg ha−1. In early April, 60 kg ha−1 were brought into early vegetation and a further 60 kg ha−1 were fertilized at the end of April, during the stem elongation stage. Following phenological studies (Bani et al. 2015a; Estrade et al. 2015), plants were harvested in mid-June for the last 3 years of the experiment. The overall vegetation responded dramatically to fertilization, by doubling the biomass and phytoextraction yield. For the first year (2005) in the fertilized plots we obtained a total biomass for all species of 6.3 t ha−1 (dry weight) and a Ni phytoextraction yield of 22.6 kg Ni ha−1; in unfertilized plots, it was only 3.2 t ha−1 (Figs. 2 and 4) with a Ni phytoextraction yield of 1.7 kg Ni ha−1. These data thus show a highly significant difference (p < 0.01) between fertilized and unfertilized plots (Fig. 4). In total phytoextraction yield, the main contributory species was A. murale. In the third year (2007) after we had changed the fertilization protocol and the date of harvest, the biomass yield in the fertilized plots was 6.0 t ha−1, whereas in unfertilized plots, it was only 0.5 t ha−1 (Fig. 2) (Bani et al. 2015a).

Ultramafic sites of Balkans (Stevanovič et al. 2003)

Evolution of biomass-production of Alyssum murale during 7 years of field-study in Pojska, Pogradec, Albania

Photo of Alyssum murale in Albanian ultramafic sites. (a) Pojska site 2013, (b) Pojska site 2014, (c) A. murale 1 plant for m2, (d) Harvesting of A. murale in Pojska site (June 2016)

Evolution of phytoextraction yield kg Ni ha−1 of Alyssum murale during 7 years of field-study in Pojska, Pogradec, Albania
3.2 Sowing Alyssum murale
In 2007–2008, the soils of the six initial 36 m2 plots were tilled to prepare appropriate seedbeds (Bani et al. 2015a). Seeds had previously been collected from the immediate vicinity of the experimental plots. All plots were hand sown in September 2007. Fertilized plots received 100 kg ha−1 P and K, 65 kg ha−1Ca, and 50 kg ha−1 N in April, and were treated with anti-monocot herbicide. A second N fertilization was performed (50 kg ha−1) 2 weeks later. The N addition was lowered in order to take into account the mineralization of plant residues. The other three plots were not treated (neither fertilized nor herbicide-treated). During the growth period the treated experimental plots were irrigated by water from a natural stream in the area. The harvesting date was early July 2008, when the biomass was considered high enough (little flowering had occurred in the newly planted plots). In 2009, the protocol of fertilization and irrigation was repeated exactly as carried out in 2008, and the plots were harvested on June 12 at the mid-flowering stage. In 2008, with split-N fertilization, irrigation, and the herbicide treatment, the biomass production was 4.6 t ha−1 in the fertilized and herbicide-treated plots; the Ni phytoextraction yield was 55 kg ha−1 (Figs. 2 and 4), in contrast to only 0.4 t and 4.3 kg Ni ha−1, respectively, in the unfertilized plots with a significant difference (p < 0.05). In 2009, in the fertilized plots the biomass production was 9.0 t ha−1 and Ni phytoextraction yield was 105 kg ha−1, which subsequently proved to be optimal in such edaphic conditions (Bani et al. 2015a, b).
3.3 Competition with Other Species and Density Effects
When enhancing natural vegetation stands by fertilization (Bani et al. 2015a), A. murale, Chrysopogon gryllus (Poaceae), and Trifolium nigrescens (Fabaceae) were the most abundant species identified on this site in the first year (2005). Other species were reported on the plots, although their contribution to total biomass production was negligible (Fig. 2). Because C. gryllus was identified as the most competitive species and T. nigrescens tended to disappear with fertilization, a good weed control was obtained on the fields by addition of 120-kg ha−1 N fertilization and a single spraying with an anti-monocot herbicide at the end of April of 2005 (Bani et al. 2015a).
During 2012–2014 we evaluated the influence of plant density on Ni phytoextraction yield in the case of sown/planted crops of A. murale on two representative ultramafic Vertisol bodies at Pojska (POJ) in the municipality of Pogradec (Fig. 3), and Domosdova (DOM) in the municipality of Prrenjas (Bani et al. 2015b). Three different densities were studied: (i) at the Domosdova site, A. murale was either transplanted without density constraints (ca. 1.5 plant m−2) (DOM-1) or a density of 6 plants m−2 on an adjacent plot (DOM-6). It was grown at a density of 4 plants m−2 in Pojska (POJ-4). In 2014, the Domosdova field was inadvertently ploughed, and was therefore unavailable for more cropping, hence only data for 2013 were available for this site.
The biomass of DOM-1 (10.6 t ha−1) was much higher, nearly twice as much as for DOM-6 (4.8 t ha−1). Nickel yield followed the same trend: DOM-1 (77 kg ha−1) was higher than DOM-6 (41 kg ha−1) (Table 2). With a density of 4 plants m−2 (POJ-4), biomass was the highest (10.2 t ha−1 in 2013 and 8.8 t ha−1 in 2014); in 2014, A. murale was in competition with its own spontaneous recruits. The Ni yields in the biomass of A. murale for 2013 (112 kg per ha−1) were higher than in 2014 (95.9 kg ha−1) (Table 2 and Figs. 2 and 4). According to the process for producing ammonium nickel sulfate hexahydrate (ANSH) from A. murale ash (Barbaroux et al. 2012; Zhang et al. 2014), it is crucial to evaluate the ash-producing yield and the percent recovery of Ni at 550 °C (Zhang et al. 2014). The total mass of ash that could be recovered was 1.3 t per ha for DOM-1 and 0.4 t per ha for DOM-6. Consequently, the total Ni mass in the ash of A. murale in the DOM-1 treatment was 76 kg ha−1 with almost no loss of Ni during the combustion process. The quantity of ash of A. murale produced in the DOM-6 treatment was only 41 kg ha −1, also without significant loss of Ni during ashing. Using the optimized process to produce ANSH from A. murale ash (Barbaroux et al. 2012), at least 45 g of ANSH (close to 100% purity) was obtained from one plant of A. murale, which represents 450 kg ANSH ha−1 in the DOM-1 plot.
Nickel phytoavailability is thought to have a stronger influence on uptake of this metal in shoots than does plant density (Bani et al. 2015b). We also stress the fact that the biomass yield of each plant does not impair Ni uptake, and that no strong dilution effect is observed in cropped A. murale (Bani et al. 2015a). Therefore, on a given site, it is important to maximize the yield of plant biomass. Biomass yield at harvest under these edaphic conditions (i.e. ultramafic Vertisols of southeastern Albania) seems to reach a maximum of ca. 10 t ha−1 when using seeds of native populations (Bani et al. 2015a, b). Only genetic selection can further improve biomass yields (i.e. 20 t ha−1), through more production of performing cultivars (Li et al. 2003), which are keys to increased Ni phytoextraction yield (Bani et al. 2015b).
3.4 Elemental Balance in Fields Cropped with A. murale
Calcium and potassium are strongly exported by Alyssum crops (Bani et al. 2015a, b). It seems that the Ca/Ni quotient varies between soils (Zhang et al. 2014) and hence the soils from Pojska produce biomass of better quality for Ni recovery (Zhang et al. 2014; Bani et al. 2015a, b) as they show lower Ca contents than at Domosdova. Based on biomass yields and on the Ca and K contents of A. murale at Pojska (Alyssum 6 in Zhang et al. 2014), Ca and K exports can reach approximately 135 and 100 kg ha−1 y−1, respectively, when assuming a biomass yield of 9 t ha−1. These results are somewhat balanced by the fertilization pattern that was validated in previous pot (Bani et al. 2007) and field (Bani et al. 2015a, b) experiments. Potassium uptake is fully compensated by fertilization, but Ca losses could appear over time due to the imbalance (77 kg added only). Calcium is not as essential as K, and this may possibly lead to regulation by A. murale and thus a decrease in Ca uptake. It is crucial that the phosphate fertilizer used in Ni agromining not be the common Di-ammonium phosphate (DAP), widely in use worldwide, because maintaining Ca fertility in soils is imperative over the long term. However, as a consequence of a lower Ca uptake, Ni uptake could be increased in A. murale crops, as suggested by our field data for these specific soils and native populations (Bani et al. 2009). Nevertheless, other studies have pointed out that Ca concentrations enhance Ni uptake under controlled conditions (Chaney et al. 2008). In any case, when agromined soils are returned for cultivation of other ‘normal’ crops, and therefore Ca fertility will be of great importance.
4 Conclusion and Perspectives
Our experimental data should help in the development of an improved growing system for Ni agromining in the Balkans. Results obtained so far demonstrate that it is necessary to sow or transplant A. murale at a density of 4 plants m2 on well-structured (ploughed) soil under conditions of non-limiting Ni availability on ultramafic Vertisols (e.g. Pojska). The fertilization pattern is now efficiently set for these soils, at 120 kg N ha−1 with a split application and additional P, K, S, and Ca fertilization (K2SO4, and Ca(H2PO4)2) at 100 kg ha−1 for K and P, and 77 kg ha−1 for Ca. Crops are likely to export ca. 100 kg of Ni and K and more than 135 kg of Ca annually. Calcium and K should be returned by fertilization over the years to keep fertility of the soil suitable for agromining. Nickel availability had not changed after 3 years of cropping (Bani et al. 2015a), but probably would decrease eventually. Weeding is crucial and targets competing species as well as young recruits of A. murale to maintain an optimum density. Harvest can be carried out at the mid-flowering stage, which maximizes the recovery of Ni. Alyssum murale is harvested by hand and allowed to dry on the field as a hay crop. It can then be baled, ready for combustion. The technology can thus be applied to all ultramafic terranes of the Balkans, where agromining has the potential to become an economically justifiable agricultural cropping system and hence enter traditional crop rotations. Other suitable edaphic situations (including Cambisols) should also be tested under field conditions in order to potentially widen the applications of agromining. Other nickel products (e.g. ANSH) with high purity can easily be obtained from A. murale shoots, some of which may have a much higher value than gross nickel metal (more than five times for a given Ni yield). In this case, the profit from the crop would be substantially higher (Barbaroux et al. 2012; Zhang et al. 2016).
References
Baker AJM, Brooks RR (1989) Terrestrial higher plants which hyper-accumulate metallic elements—a review of their distribution, ecology and phytochemistry. Biorecovery 1:81–126
Bani A, Echevarria G, Sulce S, Morel JL, Mullai A (2007) In-situ phytoextraction of Ni by a native population of Alyssum murale on an ultramafic site (Albania). Plant Soil 293:79–89
Bani A, Echevarria G, Mullaj A, Reeves R, Morel JL, Sulce S (2009) Nickel hyperaccumulation by Brassicaceae in serpentine soils of Albania and northwestern Greece. Northeast Nat 16:385–404
Bani A, Pavlova D, Echevarria G, Mullaj A, Reeves RD, Morel JL, Sulçe S (2010) Nickel hyperaccumulation by species of Alyssum and Thlaspi (Brassicaceae) from ultramafic soils of the Balkans. Bot Serb 34:3–14
Bani A, Imeri A, Echevarria G, Pavlova D, Reeves RD, Morel JL, Sulce S (2013) Nickel hyperaccumulation in the serpentine flora of Albania. Fresenius Environ Bull 22:1792–1801
Bani A, Echevarria G, Pelletier EM, Gjoka F, Sulce S, Morel JL (2014) Pedogenesis and nickel biogeochemistry in a typical Albanian ultramafic toposequence. Environ Monit Assess 186:4431–4442
Bani A, Echevarria G, Sulçe S, Morel JL (2015a) Improving the agronomy of Alyssum murale for extensive phytomining: a five-year field study. Int J Phytoremediation 17:117–127
Bani A, Echevarria G, Zhang X, Benizri E, Laubie B, Morel JL (2015b) The effect of plant density in nickel—phytomining field experiments with Alyssum murale in Albania. Aust J Bot 63:72–77
Barbaroux R, Plasari E, Mercier G, Simonnot M-O, Morel JL, Blais JF (2012) A new process for nickel ammonium disulfate production from ash of the hyperaccumulating plant Alyssum murale. Sci Total Environ 423:111–119
Brooks RR (1987) Serpentine and its Vegetation, Discorides, Portland, Oregon, USA
Brooks RR, Lee J, Reeves RD, Jaffré T (1977) Detection of nickeliferous rocks by analysis of herbarium specimens of indicator plants. J Geochem Explor 7:49–57
Brooks RR, Morrison RS, Reeves RD, Dudley TR, Akman Y (1979) Hyperaccumulation of nickel by Alyssum L. (Cruciferae). Proc R Soc Lond B 203:387–403
Cecchi L, Gabbrielli R, Arnetoli M, Gonnelli C, Hasko A, Selvi F (2010) Evolutionary lineages of nickel hyperaccumulation and systematics in European Alysseae (Brassicaceae): evidence from nrDNA sequence data. Ann Bot 106:751–767
Chaney RL, Li Y-M, Chen K-Y, Angle JS, Baker AJM (2008) Effect of calcium on nickel tolerance and accumulation in Alyssum species and cabbage grown in nutrient solution. Plant Soil 311:131–140
Chardot V, Massoura ST, Echevarria G, Reeves RD, Morel JL (2005) Phytoextraction potential of the nickel hyperaccumulators Leptoplax emarginata and Bornmuellera tymphaea. Int J Phytoremediation 7:323–335
Chardot V, Echevarria G, Gury M, Massoura ST, Morel JL (2007) Nickel bioavailability in an ultramafic toposequence in the Vosges Mountains (France). Plant Soil 293:7–21
Cheng CH, Jien SH, Iizuka Y, Tsai H, Chang YH, Hseu ZY (2011) Pedogenic chromium and nickel partitioning in serpentine soils along a toposequence. Soil Sci Soc Am J 75:659–668
Dilek Y, Furnes H (2009) Structure and geochemistry of Tethyan ophiolites and their petrogenesis in subduction rollback systems. Lithos 113:1–20
Doksopulo EP (1961) Nickel in rocks, soils, water and plants adjacent to the talc deposits of the Chorchanskaya group. Izdatelvo Tbiliskovo Universitet, Tbilisi
Estrade N, Cloquet C, Echevarria G, Sterckeman T, Deng T, Tang Y, Morel J-L (2015) Weathering and vegetation controls on nickel isotope fractionation in surface ultramafic environments (Albania). Earth Planet Sci Lett 423:24–35
Li YM, Chaney R, Brewer E, Roseberg R, Angle JS, Baker AJM, Reeves RD, Nelkin J (2003) Development of a technology for commercial phytoextraction of nickel: economic and technical considerations. Plant Soil 249:107–115
Marhold K (2011) Brassicaceae. – In: Euro+Med Plantbase—the information resource for Euro-Mediterranean plant. http://ww2.bgbm.org/EuroPlusMed/PTaxonDetail.asp?NameCache=Brassicaceae&PTRefFk=7200000
Massoura ST, Echevarria G, Becquer T, Ghanbaja J, Leclerc-Cessac E, Morel JL (2006) Nickel-bearing phases and availability in natural and anthropogenic soils. Geoderma 136:28–37
Menezes de Sequeira E (1969) Toxicity and movement of heavy metals in serpentinitic rocks (north-eastern Portugal). Agron Lusit 30:115–154
Minguzzi C, Vergnano O (1948) Il contenuto di nichel nelle ceneri d’Alyssum bertolonii Desv. Atti della Società Toscana di Scienze Naturali, Memorie Serie A 55:49–77
Nkrumah PN, Baker AJM, Chaney RL, Erskine PD, Echevarria G, Morel JL, van der Ent A (2016) Current status and challenges in developing nickel phytomining: an agronomic perspective. Plant Soil 406:55–69
Pollard AJ, Reeves RD, Baker AJM (2014) Facultative hyperaccumulation of heavy metals and metalloids. Plant Sci 217–218:8–17
Psaras GK, Constantinidis T (2009) Two new nickel hyperaccumulators from the Greek serpentine flora. Fresenius Environ Bull 18:798–803
Raous S, Echevarria G, Sterckeman T, Hanna K, Thomas F, Martins ES (2013) Potentially toxic metals in ultramafic mining materials: identification of the main bearing and reactive phases. Geoderma 192:111–119
Reeves RD (1992) The hyperaccumulation of nickel by serpentine plants. In: AJM B, Proctor J, Reeves RD (eds) The Vegetation of Ultramafic (Serpentine) Soils. Intercept Ltd., Andover, pp 253–277
Reeves RD, Adigüzel N (2008) The nickel hyperaccumulating plants of the serpentines of Turkey and adjacent areas: a review with new data. Turk J Biol 32:143–153
Reeves RD, Brooks RR, Dudley TR (1983) Uptake of nickel by species of Alyssum, Bommuellera, and other genera of Old World tribus Alysseae. Taxon 32:184–192
Reeves RD, Baker AJM, Kelepertsis A (1997) The distribution and biogeochemistry of some serpentine plants of Greece. In: Jaffré T, Reeves RD, Becquer T (eds) Écologie des milieux sur roches ultramafi ques et sursols métallifères, ORSTOM, Nouméa, pp 205–207. Documents Scientifi ques et Techniques No. III/2
Rešetnik I, Satovic Z, Schneeweiss GM, Liber Z (2013) Phylogenetic relationships in Brassicaceae tribe Alysseae inferred from nuclear ribosomal and chloroplast DNA sequence data. Mol Phylogenet Evol 69:772–786
Rešetnik I, Schneeweiss G, Liber Z (2014) Two new combinations in the genus Bornmuellera (Brassicaceae). Phytotaxa 159:298–300
Salihaj M, Bani A, Echevarria G (2016) Heavy metals uptake by hyperaccumulating flora in some serpentine soils of Kosovo. Global Nest J 18(1):214–222
Shallari S, Schwartz C, Hasko A, Morel JL (1998) Heavy metals in soils and plants of serpentine and industrial sites of Albania. Sci Total Environ 209:133–142
Španiel S, Kempa M, Salmeron-Sanchez E, Fuertes-Aguilar J, Mota J, Al-Shehbaz I, German D, Olsavska K, Barbora Singliarova B, Lihova J, Marhold K (2015) AlyBase: database of names, chromosome numbers, and ploidy levels of Alysseae (Brassicaceae), with a new generic conceptof the tribe. Plant Syst Evol 301(10):2463–2491
Stevanović V, Tan K, Iatrou G (2003) Distribution of the endemic Balkan Flora on serpentine I 309 obligate serpentine endemics. Plant Syst Evol 242:149–170
Tan K, Mullaj A, Ruci B, Iatrou G (1999) Serpentine endemics in Albania and NW Greece. In Abstacts of the Vth Conference on Plant Taxonomy. Lisboa 1999 s.68. A. Correia et al. (eds) Museu, Laboratorio, e Jardim Botanico da Universida de Lisboa, Lisboa, Portugal
Tumi A, Mihailović N, Gajić B, Niketić M, Tomović G (2012) Comparative study of hyperaccumulation of nickel by Alyssum murale s.l. populations from the ultramafics of Serbia. Pol J Environ Stud 21:1855–1866
van der Ent A, Baker AJM, Reeves RD, Pollard AJ, Schat H (2013) Hyperaccumulators of metal and metalloid trace elements: facts and fiction. Plant Soil 362:319–334
van der Ent A, Baker AJM, Reeves RD, Chaney RL (2015) Agromining: farming for metals in the future? Environ Sci Technol 49:4773–4780
Wenzel WW, Jockwer F (1999) Accumulation of heavy metals in plants grown on mineralized soils of the Austrian Alps. Environ Pollut 104:145–155
Zdruli P (1997) Benchmark Soils of Albania: Resource Assessment for Sustainable Land Use. PhD thesis. Published by the USDA Natural Resources Conservation Service (NRCS), Washington DC and the International Fertilizer Development Center (IFDC), Muscle Shoals, Alabama, Two Vols, 293 p
Zhang X, Houzelot V, Bani A, Morel JL, Echevarria G, Simonnot M-O (2014) Selection and 306 combustion of Ni-hyperaccumulators for the phytomining process. Int J Phytoremediation 16:1058–1072
Zhang X, Laubie B, Houzelot V, Plasari E, Echevarria G, Simonnot M-O (2016) Increasing purity of ammonium nickel sulfate hexahydrate and production sustainability in a nickel phytomining process. Chem Eng Res Des 106:26–32
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2018 Springer International Publishing AG
About this chapter

Cite this chapter
Bani, A., Echevarria, G., Pavlova, D., Shallari, S., Morel, J.L., Sulçe, S. (2018). Element Case Studies: Nickel. In: Van der Ent, A., Echevarria, G., Baker, A., Morel, J. (eds) Agromining: Farming for Metals. Mineral Resource Reviews. Springer, Cham. https://doi.org/10.1007/978-3-319-61899-9_12
Download citation
DOI: https://doi.org/10.1007/978-3-319-61899-9_12
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-61898-2
Online ISBN: 978-3-319-61899-9
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)