
Abstract
One of the fundamental properties of living organisms is the ability to sense and respond to changes in their environment by movement. If a motile cell senses soluble molecules and follows along a concentration gradient to the source, or if it moves away from a source of undesirable chemicals, e.g. repellent, toxin, it is displaying a directional movement called positive or negative chemotaxis, respectively. This phenomenon is well-known to biologists and intensively studied in living systems. In contrast chemokinesis is a change in movement due to environmental input but the resulting movement is non-vectorial and can be considered directionally random. Recently, in the last ten years, few laboratories started to focus on the movement properties of artificial constructs, including the directional movement of non-living objects in chemical gradients. This chapter will focus on chemotaxis and chemokinesis of natural and synthetic systems that may provide chemical platforms for unconventional computing.
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others

Keywords
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
11.1 Cellular Movement in Biological Systems
Living cells can physically move through several mechanisms. Due to the length scales of most living cells, viscosity will dominate over inertia. Therefore at such low Reynolds numbers, motion of the cell will require the expenditure of energy. However, some cells rely entirely upon passive flotation and Brownian motion for dispersal. Under the microscope non-motile (and also dead) cells seem to move in a purposeful way, though they may frequently change direction, but this is due to random molecular bombardment of cells by the molecules of the solvent. This type of undirected motion demands no energy nor sophisticated machinery from the cells and therefore it is necessary to distinguish the effects of passive Brownian motion from active cellular motility [12]. There are several primary mechanisms of active cell motion: swimming using rotating flagella in prokaryotes, e.g. E. coli, or cilia in eukaryotes, Tetrahymena; cytoskeleton polarization, e.g. Dictyostelium; and gliding, cyanobacteria. Overall, cell motility can be divided into three types [25]:
-
1.
basal random motility, which takes place in the absence of chemical stimuli,
-
2.
chemokinesis, which corresponds to increased random motility in response to chemical stimuli,
-
3.
chemotaxis, which corresponds to stimulated migration towards an increasing (or decreasing) chemical gradient.
Cells can move independently or collectively, and they can migrate as single unattached cells or multicellular groups (see Table 2 in [60]). For example, single tumor cells can perform amoeboid migration or mesenchymal migration. Some carcinoma cells with an amoeboid morphology can move at high speeds inside the tumors (\(\sim \)4 \(\upmu \)m/min), whereas mesenchymal migration is characterized by an elongated cell morphology with established cell polarity and relatively low speeds of cell migration (0.1–1 \(\upmu \)m/min). Directed multicellular migration can be either collective migration, in which the cells are in tight contact with each other (also known as cohort migration) or cell streaming, in which the coordinated cell migration involves cells that are not always in direct physical contact.
11.2 Sensing in Biology
Most organisms (even bacteria) can sense hearing, light, pressure, gravity and chemicals and as a response to them can display specific movements (phototaxis-response to light [11], magnetotaxis-response to magnetic field [58], thermotaxis-movement along thermal gradient [50], galvanotaxis-movement along the electric field [5], haptotaxis-movement along the gradient of adhesion sites [15], and gravitaxis-movement along the direction of the gravitational force [59]). Intelligent animals can find food by coordinating their senses (smell, sight, and sound) with their central nervous system. Small organisms with no sense-specific organs or central nervous system find food by chemotaxis. The bacterium E. coli as a well-studied organism can sense and respond to changes in temperature, osmolarity, pH, noxious chemicals, DNA-damaging agents, mineral abundance, energy sources, electron acceptors, metabolites, chemical signals from other bacteria, and parasites [49]. E. coli has five chemotaxis sensors: Trg, for ribose and galactose; Tar, for aspartate; Tsr, for serine; Tap, for peptides; and the newly discovered Aer, which may be a redox detector [31].
Depending on the type and size of a cell, there are two modes of chemical gradient sensing: temporal and spatial. Bacteria are very small organisms and due to their length scale it is thought that the cells are not able to detect gradients spatially, therefore they detect the chemicals temporally. In other words chemotaxis of bacteria involves memory. Bacteria detect the gradient by comparing their currently sensed chemical environment with that sensed in recent past. If the conditions are improving, then the cell will continue with running along its current path. However, if the conditions are not improving because the cell is moving away from the source of nutrients, then the cell will tumble and try to find a new path that yields a more favorable trajectory. By this trial-and-error approach, the bacterial cell would eventually move up the gradient. This mechanism is based on sensory adaptation as the bacterium responds only to temporal changes in signal molecule concentration rather than their absolute values [19].
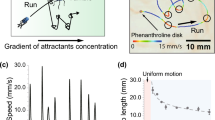
Eukaryotic cells detect differences in the chemicals concentrations across their cell body because of different receptor occupancy on opposite sides resulting on polarization of the molecular distributions in the cell [38]. Such a spatial gradient sensing mechanism leads to more or less straight line migration, as opposed to the running and tumbling of flagellated bacteria. One of the best-studied examples of eukaryotic chemotactic system is the migration of amoeba Dictyostelium discoideum along an increasing concentration of cyclic adenosine-3\('\),5\('\)-monophosphate (cAMP) [43, 61]. A Dictyostelium cell can measure about 1 % of concentration difference over its total length (10–20 \(\upmu \)m) in a spatial cAMP gradient. In the absence of cAMP, the cells are not at rest, but perform a random type of motion with the average motility 4.19 \(\upmu \)m/min [62]. Cells show no directional response to negligible stationary, linear cAMP concentration gradients (of less than 10\(^{-3}\) nM/\(\upmu \)m) and exhibited a constant basal motility. In steeper gradients, cells move up the gradient with an average motility of \(\sim \)9 \(\upmu \)m/min directly towards the cAMP source. In very steep gradients (above 10 nM/\(\upmu \)m) the cells lose directionality and the motility returned to random motion (see Fig. 6 in [62]). From this it is evident that the chemotaxis can occur only in a certain range of chemoattractant concentration. Extremely low or conversely too high concentrations do not trigger the chemotactic migration.
The true slime mold Physarum polycephalum in the plasmodial stage also shows the chemotactic behavior [6]. Several works focused on the effect of various compounds on Physarum chemotaxis. For example, carbohydrates [14, 18, 44], amino acids and nucleotides [18], and volatile organic chemicals [20] have been found as chemoattractants, whereas inorganic salts [3] were found to be chemorepellents.
11.3 Importance of Chemotaxis in Multicellular Organisms
In multicellular organisms, chemotaxis is very important in physiological processes, such as the recruitment of inflammatory cells to sites of infection and in organ development during embryogenesis [26]. Neuronal and embryonic cells migrate during development. During angiogenesis, endothelial cells undergo chemotaxis to form blood vessels, while epithelial cells and fibroblasts chemotaxis during wound healing [48]. In addition, chemotaxis governs the motion of sperm towards the egg during mammalian fertilization [24].
However in addition to a role in obviously beneficial processes in multicellular organisms, chemotaxis is also involved in each crucial step of tumor cell dissemination and metastatic progression (invasion, intravasation and extravasation). Many cancer cells such as breast cancer cells are known to preferentially metastasize into certain tissues and organs. This preference is correlated with the production of chemoattractants by the target tissues and organs and up-regulation of chemoattractant receptors in the cancer cells [60, 71].
11.4 Chemotaxis Versus Chemokinesis
There are two different terms in biology that link chemical input with motion output: chemotaxis and chemokinesis [56]. Whereas chemotaxis results in oriented movement, chemokinesis occurs through the increase or decrease of speed, but the directionality is not biased. Both types of movement have been shown in both healthy and malignant cells. For example, multiple growth factors stimulate chemotaxis and/or chemokinesis in malignant mesothelioma cells [47]. For healthy neurons and epithelial cells, micromolar concentrations of gamma-aminobutyric acid (GABA) induce chemokinesis in rat cortical neurons during development, while femtomolar concentrations induce chemotaxis [10]. Furthermore, vascular endothelial growth factor A (VEGFA) was shown, using microfluidic chemotactic chambers, to increase chemotaxis of epithelial cells while decreasing chemokinesis [37]. Although both chemotaxis and chemokinesis may play essential roles in biology, modes of cellular motion employing one or both mechanisms will be different.
11.5 Artificial Chemotaxis and Chemokinesis
The spontaneous motion of liquid droplets, solid particles, and gels under non-equilibrium conditions has been investigated both experimentally and theoretically in several works. Self-propelled systems are studied in broad range of scales and can be called nano-/micro-robots [21, 45], nano-/micro-motors [72], nano-/micro-swimmers [23, 57], and some such systems can be studied at the macroscale [73]. The self-propelled movement of non-biological objects can mimic the chemotactic or chemokinetic behavior of living cells. Usually such examples are based on mechanisms completely different from biological movement, but a link is established between chemical energy and mechanical work.
Camphor grains in water represent the classic non-living chemical system studied for autonomous motion [67] and one of the most coherent examples of chemokinesis. Starting far from equilibrium, the solid camphor spreads as a surfactant at the air water interface and at the same time sublimates. Therefore surface tension gradients are established in the system and fluid can flow through Marangoni-type motion. Due to sublimation of the camphor from the interface, the system will not become blocked with surfactant but the motion can be sustained as long as the camphor grains persist.
Lateral motion of oil or alcohol droplets in aqueous solution containing surfactants are reported in several papers [8, 34, 65, 68]. The droplets are usually propelled by unbalanced surface tension resulting in fluid dynamics through Marangoni-type instabilities. This behavior could be compared to the chemokinesis of living cells, because droplets perform a movement without any orientation towards the source of chemoattractant or from the chemorepellent.
As with motile organelles and cells that exhibit polarity, spontaneous symmetry breaking is needed for self-propelled movement of non-biological objects. A droplet on a surface of a substrate could move when the underlying surface is asymmetrically patterned creating a difference in the interfacial energy between the leading edge and the trailing one of the droplet [22, 63]. Furthermore a droplet could break symmetry through a coupled chemical reaction that occurs at the interface between the droplet and its surrounding medium. The chemical reaction produces a symmetry breakage due to the accumulation and release of the products [35] and the droplets swim through the aqueous media without the need of an air-water or solid-liquid interface.
We have presented several works describing a system of oil droplet movement based on fatty acid chemistry. In [34], the oil containing oleic anhydride precursor was introduced into an aqueous environment that contains oleate micelles. The products of the precursor hydrolysis were coupled to the movement of the oil droplet and the production of waste vesicles. The oil droplet successfully moves away from this waste product into fresh unmodified solution and even displays a primitive form of chemotaxis. This example mimics the cells that move away from their metabolic products and waste into regions with fresh nutrients. In another system, the catalyst solvated in the anhydrous droplet reacts with surfactant substrate in the surrounding aqueous environment that results in symmetry breakage and fuelled droplet motion [68].
Several trajectory patterns of self-propelling droplets that mimic chemokinesis can be observed: irregular [53], circular [52], back-and-forth, knot-forming, and irregular [66]. They may or may not depend on the shape of the boundary. The center of trajectory either corresponds to the dish center (global trajectory) or it does not correspond to the dish center (local trajectories) [66]. We have studied the movement of oleic anhydride-loaded nitrobenzene droplets in oleate micelle solution and we have found that droplet size (volume) correlates with various behavioral modes, namely fluctuating, directional, vibrational, and circular [36].
Research on asymmetric bimetallic catalytic rods or spheres in the concentration gradient of a chemoattractant shows oriented movement of artificial particles that could be compared to chemotaxis. Typically, bimetal nanorod contains one segment made of a metal that functions as a catalyst for the decomposition of H2O2.
During the decomposition, oxygen is produced at the surface of the metallic segment according to the reaction 2H2O2 (l) \(\rightarrow \) O2(g) + 2H2O (l) and provides the driving force for nanorod propulsion. The other segment can be made of any material that is not reactive towards hydrogen peroxide, such as an inorganic, organic or polymeric metal, semiconductor or insulator; often this segment is made of gold. Bidoz et al. [27] studied Ni-Au nanorods in the H2O2 gradient. The group of Ayusman Sen [55] studied the artificial chemotaxis of platinum-gold rods (2 \(\upmu \)m in length, 370 nm in diameter) in the H2O2 gradient. Sundararajan with colleagues [64] used also Pt-Au rods in H2O2, in this case connected with the polystyrene bead cargo. Howse et al. [37] prepared polystyrene 1.6 \(\upmu \)m microspheres half-coated in platinum and studied their movement in the H2O2 gradient, later Ke et al. [40] performed similar experiments with silica 1 \(\upmu \)m particles half-coated in Pt by providing a chemical mechanism of motion and imposed asymmetric design, such systems display or may be capable of artificial chemotaxis.
In several works, it was shown that the diffusio-phoretic phenomenon shares similarities with chemotaxis [1]. When a rigid colloidal particle is placed in a solution which is not uniform in the concentration of a solute that interacts with the particle, the particle will be propelled in the direction of lower concentration of the solute. The resulting locomotion is called diffusio-phoresis [7].
Artificial systems can be created to mimic also other kind of induced directional movement, such as magnetotaxis. Dreyfus et al. [23] showed that a linear chain of colloidal magnetic particles linked by DNA and attached to a red blood cell can act as a flexible artificial flagellum. The filament was aligned with an external uniform magnetic field and was readily actuated by oscillating a transverse field. They found that the actuation induced a beating pattern that propels the structure, and that the external fields can be adjusted to control the velocity and the direction of motion, reproducing one magnetotaxis like behavior. Chaturvedi et al. [17] fabricated microscale catalytic motors using particles of various material compositions, including metal and magnetic polymer. Such particles have shown autonomous movement, in random directions, in the presence of H2O2 and UV light, but if an external magnetic field was also present, they showed directed movement, forming exclusion regions around them. The magneto-phototactic response shown by the magnetic colloidal chains opens a new path for their use as delivery vehicles.
Several methods are used in biology to study chemotaxis. In the most direct method, cells and their trails can be directly visualized with a camera and then analyzed with image analysis software. Chemotaxis of populations of cells is often measured by counting cells at the attractant source over time [39]. Similar methods are used for the non-biological chemotaxis studies. Chemotaxis studies require a way to deliver chemicals to the observed object (cell, particle, droplet) in a controlled way. A wide range of techniques is available to evaluate chemotactic activity of cells, such as generation of gradient with micropipette [70], using of special chambers (Boyden chamber [13], Dunn chamber [74], Zigmond chamber [75]), and microfluidic devices. Microfluidic devices are ideal tools for creating and controlling chemotactic gradients and are widely used and popular in recent times. Microfabrication methods allow arbitrary microchannel designs to be created through which cells can move. Microchannel dimensions can be created small enough that diffusion occurs in minutes to seconds, thus reducing the waiting time for a gradient to become established [69]. Other methods for studying chemotaxis have been reviewed in [30, 32, 41]. For the study of artificial chemotaxis the most common methods are direct addition of signal molecules by pipette [15, 34], addition of gels soaked with chemoattractant [35, 46], and microfluidics devices [9, 29].
11.6 Motile Systems in Mazes
Living cells and non-living objects can move in mazes towards chemoattractant targets. Many examples of directional movement of single cells in natural maze-like environments can be found. For example, leukocytes, in the case of inflammation, move towards the inflamed site through the intricate maze of the circulatory system following chemoattractants released from the injured tissue [28]. During axonal growth, neurons move through the intricate maze of the brain and create new synapses responding to specific neutrophic factors gradients [51].
More complex organisms, such as slime molds [54], solve mazes doped with chemoattractants. For example, in [4] slime mould is inoculated in a maze’s peripheral channel and an oat flake (source of attractants) in a the maze’s central chamber. The slime mould grows toward target oat flake and connects the flake with the site of original inoculation with a pronounced protoplasmic tube. The protoplasmic tube represents a path in the maze. The plasmodium solves maze in one pass because it is assisted by a gradient of chemo-attractants propagating from the target oat flake.
The method for solving the maze involves the search of several alternative solutions in parallel. When one or more of the searches finds the source of food then these successful paths are reinforced in time and the non-successful branches in the slime mold are diminished. Currently Physarum is used for unconventional computation and for fabrication of novel and emerging non-silicon computers and robots. How to create experimental Physarum machines—a green and environmentally friendly unconventional computers—has been shown in Adamatzky’s book titled ‘Physarum Machines: Computers from Slime Mould’ [2].
In our recent work we have shown that even nonliving objects such as decanol droplets in decanoate solutions are able to move chemotactically in salt gradients [16]. It has been demonstrated that this artificial chemotactic system bears many qualitative similarities with natural chemotactic systems, namely:
-
1.
the ability to perform chemotaxis repeatedly when the chemoattractant gradients are recreated,
-
2.
to perform chemotaxis in topologically complex environments (Fig. 11.1),
-
3.
to select the chemotaxis direction based on the relative strength of alternative chemoattractant sources,
-
4.
to rest in a dormant state and later respond to a stimuli-responsive chemoattractant release, and finally,
-
5.
to deliver a reactive payload at the chemoattractant source.

Maze solving decanol droplet. (Supplementary movie in youtube http://youtu.be/P5uKRqJIeSs). Reprinted with permission from [16]. ©2014, American Chemical Society
Lagzi and his colleagues [46] showed a droplet of mineral oil and 2-hexyldecanoic acid in a labyrinth with a pH gradient, where the droplets were able to find the shortest path away from the maze. They prepared a maze and filled it with a KOH solution. Due to the presence of a gel soaked with HCl in the ‘goal’, a pH gradient was established in the maze channels. Then a 1 \(\upmu \)l droplet of mineral oil and 2-hexyldecanoic acid placed into the ‘start’ position moved toward the source of HCl. Such droplets have shown chemotactic movement in the pH gradient (pH-taxis).
11.7 Chemotactic Decanol Droplets as ‘Chemo-Taxi’
Chemotactic droplets could serve as transporters for chemicals or small objects, a sort of ‘chemo-taxi’. In our previous work [16], it was shown that the decanol droplet can carry a reactive payload and deliver it towards the source of chemoattractant (see Fig. 9 in [16]). Here it will be shown the use of decanol droplets for transport of small objects.

‘Chemo-taxi’-decanol droplets transporting small objects in salt gradients. a fly, b hair, c polystyrene and d paper. The size of a rectangular slide is 25 \(\times \) 75 mm
Generally, a taxi is a type of vehicle that conveys passengers between locations of their choice. Let us consider that our decanol droplet could serve as a taxi and as our model passenger a dead fly was chosen. Figure 11.2 represents the transport of a fly from the right hand side of the glass slide to the left hand side of the glass into the place where in the beginning of experiment small amount of salt was added. In principle this kind of chemo-taxi could serve for the transport of any other small object. Further examples with piece of paper, polystyrene and hair are shown in Fig. 11.2. In addition, a piece of conductive wire can be transported through chemotaxis. Then the environmental conditions can be changed such that the droplet releases the cargo on demand, in this case by adding 25 mM decanoate in water (pH 12) to the system. See movie [78] and Fig. 11.3.

‘Chemo-taxi’—decanol droplet transporting and dropping a piece of conductive copper wire of 1.5 mm in length. The diameter of the glass petri dish is 60 mm. 0 s: droplet with wire placed in experiment; 8 s: salt gradient added; 15 s–1 min: droplet migration towards salt source; 1 min–3 min 30 s: 25mM decanoate addition; 3 min 30 s: second salt addition; 3 min 30 s–4 min: droplet migration towards second salt source; 4–6 min 25 mM decanoate addition to trigger release of wire from droplet; 6 min third salt addition; 6 min–6 min 30 s migration towards third salt source and wire left behind. See movie at 8\(\times \) real time [78]
The idea to have such a ‘chemo-taxi’ could be used for several potential applications in the future. This chemotactic system could be developed to help machine engineers maintain their equipment, because droplets could track down pH, salt or heat gradients to get to the desired destination and lubricate a specific joint or axle. Another application of chemotactic droplets could be for chemical reaction control. The motile reaction containers in the form of the oil droplets could move to another place or closer to other reaction container via the concentration gradient. Gradients are thus one way of controlling reactions and for manipulation of reactants or products. Chemotactic droplets can fulfil various tasks in response to chemical signals in hazardous areas or limited spaces beyond the control of external power sources.
11.8 Chemotactic Technology with Design Applications
The pattern of movement of the chemotactic decanol system depends on the environmental context. Usually we perform such experiments by placing a decanol droplet with dye in a glass petri dish or glass slide containing 10 mM decanoate pH 12 surfactant. We then see chemotaxis of the droplet towards the high salt source with little or no observable distortion in droplet shape. However if we use a more challenging environmental context, we see a different pattern. For example, when we saturate glass fiber paper with the decanoate solution, then place a droplet of decanol with dye on the surface, the decanol will migrate towards the salt source by producing multiple protruding ends through the fiber matrix. Those ends that successfully manage the fiber network and find the salt will be reinforced over ends that find little or no salt. In this way we see a pattern of migration and chemotaxis similar in phenomenology to the searching behavior of slime mold [76]. Figure 11.4 shows such an experiment. Two droplets of decanol are placed with a space of about 1 cm between them on decanoate saturated glass fiber paper. Concentrated NaCl is added in the gap between them. After about one minute the salt gradient reaches the decanol droplets and the initial protuberances form (see Fig. 11.4, arrows A and B), These protuberances extend, bifurcate and they migrate through the glass fiber matrix. The figure demonstrates the chemotactic movement with the leading edges of the droplets growing towards the chemoattractant and the trailing edges retracting, resulting the net migration towards the source.

Chemotaxis of decanol through glass fiber paper. Time series including initial state, then every minute time point post NaCl addition. Star indicates point of NaCl addition. Arrows A, B indicate initial protrusions from red and black droplets, respectively, shown here in grey scale as black (right) and grey (left). Arrow C indicates retraction of red droplet. Area of glass fiber paper shown is 3 \(\times \) 1.8 cm [76]
The decanol system on fiber glass is a novel system utilizing the design potential of chemotactic technology as its principle criterion [33]. The decanol system moves to towards a salt source over the substrate [77]. Three design conditions which establish the success of the movement are gravity, buoyancy of the decanol system and the design of the structural matrix of the glass fibre substrate. By infusing the decanol system with dyes the scale of the droplets is enhanced in order to observe its interaction across a substrate. It was observed that its manoeuvrability around obstacles solve complex paths to reach the salt solution.
This process shares similar problem solving criteria to the design of architecture, a theme also explored through the 200 Year Continuum [42]. By observing the decanol system, facets of architectural design are developed using site-specific environmental conditions affecting the outcome of successful architectural solutions. By designing the decanol system to move through the substrate we observe behavioral characteristics in relation to its environment-that is, moving between fiber glass matrixes or navigating across or around grains of sand etc. [77]. The initial conditions are set and the experiment unfolds through movement to explore and solve a complex chemical and physical landscape.
11.9 Conclusion
We have discussed the role of movement in natural cellular and artificial systems. Cellular movement is important in single cell nutrition and survival but also in the larger macroscopic healthy and disease states in multicellular organisms. Sensing of the immediate environment by cells affects the rate of motion through chemokinesis and/or the direction of motion through chemotaxis. Some cells are capable of both types of motion which often depends on the amount of chemical stimulant present. Interest in the design of self-moving artificial systems on the same size scale as cells is rapidly developing. Such systems either with imposed asymmetry or spontaneously breaking symmetry are already capable of chemokinesis and chemotaxis and are sensitive to different chemical signals such as salt concentration, light, and pH. In some cases, as with droplets, the artificial system can move and leave a chemical pattern on the environment but also pick up and transfer physical cargo. Basic computational aspects of such motile artificial systems are being demonstrated in maze solving tasks. The future development of self-motile artificial systems towards applications in unconventional computing is intriguing and worthy of further study and experimentation.
References
Abecassis, B., Cottin-Bizonne, C., Ybert, C., Ajdari, A., Bocquet, L.: Boosting migration of large particles by solute contrasts. Nature Mater. 7, 785–789 (2008)
Adamatzky, A.: Physarum Machines: Computers from Slime Mould. Series on Nonlinear Science Series A, vol. 74. World Scientific, Singapore (2010)
Adamatzky, A.: Routing Physarum with repellents. Eur. Phys. J. 31, 403–410 (2010)
Adamatzky, A.: Slime mold solves maze in one pass, assisted by gradient of chemo-attractants. IEEE Trans. NanoBioscience 11(2), 131–134 (2012)
Adler, J., Shi, W.: Galvanotaxis in a bacteria. Cold Spring Harbor Symp. Quant. Biol. 53, 23–25 (1988)
Aldrich, H.: Cell Biology of Physarum and Didymium V1: Organisms, Nucleus, and Cell Cycle. Elsevier Science, Amsterdam (2012)
Anderson, J.L., Prieve, D.C.: Diffusiophoresis: migration of colloidal particles in gradients of solute concentration. Sep. Purif. Methods 13, 67–103 (1984)
Ban, T., Yamagami, T., Nakata, H., Okano, Y.: pH-Dependent motion of self-propelled droplets due to marangoni effect at neutral pH. Langmuir 29, 2554–2561 (2013)
Baraban, L., Harazim, S.M., Sanchez, S., Schmidt, O.G.: Chemotactic behavior of catalytic motors in microfluidic channels. Angewandte Chemie International Edition 52, 5552–5556 (2013)
Behar, T.N., Li, Y.X., Tran, H.T., Ma, W., Dunlap, V., Scott, C., Barker, J.L.: GABA stimulates chemotaxis and chemokinesis of embryonic cortical neurons via calcium-dependent mechanisms. J. Neurosci. 16(5), 1808–1818 (1996)
Bennett, R.R., Golestanian, R.: A steering mechanism for phototaxis in chlamydomonas. J. R. Soc. Interface 12, (2015)
Berg, H.C.: Random Walks in Biology. Princeton University Press, Princeton (1993)
Boyden, S.: The chemotactic effect of mixtures of antibody and antigen on polymorphonuclear leucocytes. J. Exp. Med. 115, 453–466 (1962)
Carlile, M.J.: Nutrition and chemotaxis in the myxomycete Physarum polycephalum: the effect of carbohydrates on the plasmodium. J. Gen. Microbiol. 63, 221–226 (1970)
Carter, S.B.: Haptotaxis and the mechanism of cell motility. Nature 213, 256–260 (1967)
Čejková, J., Novák, M., Štěpánek, F., Hanczyc, M.M.: Dynamics of chemotactic droplets in salt concentration gradients. Langmuir 30, 11937–11944 (2014)
Chaturvedi, N., Hong, Y., Sen, A., Velegol, D.: Magnetic enhancement of phototaxing catalytic motors. Langmuir 26, 6308–6313 (2010)
Chet, I., Naveh, A., Henis, Y.: Chemotaxis of Physarum polycephalum towards carbohydrates, amino acids and nucleotides. J. Gen. Microbiol. 102, 145–148 (1977)
Collin, M., Schuch, R.: Bacterial Sensing and Signaling. Karger, Switzerland (2009)
de Lacy Costello, B.P.J., Adamatzky, A.I.: Assessing the chemotaxis behavior of Physarum polycephalum to a range of simple volatile organic chemicals. Commun. Integr. Biol. 6, (2013)
Dey, K.K., Bhandari, S., Bandyopadhyay, D., Basu, S., Chattopadhyay, A.: The pH taxis of an intelligent catalytic microbot. Small (2013)
Dos Santos, F.D., Ondarcuhu, T.: Free-running droplets. Phys. Rev. Lett. 75, 2972–2975 (1995)
Dreyfus, R., Baudry, J., Roper, M.L., Fermigier, M., Stone, H.A., Bibette, J.: Microscopic artificial swimmers. Nature 437, 862–865 (2005)
Eisenbach, M.: Sperm chemotaxis. Rev. Reprod. 4, 56–66 (1999)
Eisenbach, M.: Chemotaxis, vol. 499. Imperial College Press, London (2004)
Entschladen, F., Zänker, K.S.: Cell Migration: Signalling and Mechanisms. Karger, Switzerland (2009)
Fournier-Bidoz, S., Arsenault, A.C., Manners, I., Ozin, G.A.: Synthetic self-propelled nanorotors. Chem. Commun., 441–443 (2005)
Fox, R.B., Hoidal, J.R., Brown, D.M., Repine, J.E.: Pulmonary inflammation due to oxygen toxicity: involvement of chemotactic factors and polymorphonuclear leukocytes. Am. Rev. Respir. Dis. 123, 521 (1981)
Francis, W., Fay, C., Florea, L., Diamond, D.: Self-propelled chemotactic ionic liquid droplets. Chem. Commun. 51, 2342–2344 (2015)
Genzer, J., Bhat, R.R.: Surface-bound soft matter gradients. Langmuir 24, 2294–2317 (2008)
Grebe, T.W., Stock, J.: Bacterial chemotaxis: the five sensors of a bacterium. Curr. Biol. 8 (1998)
Guan, J.L.: Cell migration: developmental methods and protocols. Methods in Molecular Biology. Humana, New York (2004)
Hanczyc, M.M., Ikegami, T.: Protocells as smart agents for architectural design. Technoetic Arts J. 7(2), 117–120 (2009)
Hanczyc, M.M., Toyota, T., Ikegami, T., Packard, N., Sugawara, T.: Fatty acid chemistry at the oil-water interface: self-propelled oil droplets. J. Am. Chem. Soc. 129, 9386–9391 (2007)
Hiroki, M., Hanczyc, M.M., Ikegami, T.: Self-maintained movements of droplets with convection flow. Progress in Artificial Life. Springer, Berlin (2007)
Horibe, N., Hanczyc, M.M., Ikegami, T.: Mode switching and collective behavior in chemical oil droplets. Entropy 13, 709–719 (2011)
Howse, J.R. et al. Self-motile colloidal particles: From directed propulsion to random walk. Phys. Rev. Lett. 99 (2007)
Jeon, K.W.: International Review of Cytology: A Survey of Cell Biology. Elsevier Science, Amsterdam (2007)
Jin, T., Hereld, D.: Chemotaxis: Methods and Protocols. Humana Press, New York (2009)
Ke, H., Ye, S., Carroll, R.L., Showalter, K.: Motion analysis of self-propelled Pt-Silica particles in hydrogen peroxide solutions. J. Phys. Chem. A 114, 5462–5467 (2010)
Keenan, T.M., Folch, A.: Biomolecular gradients in cell culture systems. Lab Chip 8, 34–57 (2008)
Kerrigan, C.: The 200 Year Continuum. Leonardo, Massachusetts Institute of Technology Press, 42(4), 314–323 (2009)
Kessin, R.H.: Dictyostelium: Evolution, Cell Biology, and the Development of Multicellularity. Cambridge University Press, Cambridge (2001)
Knowles, D.J.C., Carlile, M.J.: The chemotactic response of plasmodia of the myxomycete physarum polycephalum to sugars and related compounds. J. Gen. Microbiol. 108, 17–25 (1978)
Lagzi, I.: Chemical robotics-chemotactic drug carriers. Cent. Eur. J. Med., 1–6 (2013)
Lagzi, I., Soh, S., Wesson, P.J., Browne, K.P., Grzybowski, B.A.: Maze solving by chemotactic droplets. J. Am. Chem. Soc. 132, 1198–1199 (2010)
Liu, Z., Klominek, J.: Chemotaxis and chemokinesis of malignant mesothelioma cells to multiple growth factors. Anticancer Res. 24, 1625–1630 (2004)
Martin, P.: Wound healing-aiming for perfect skin regeneration. Science 276, 75–81 (1997)
Meyers, L.A., Bull, J.J.: Fighting change with change: adaptive variation in an uncertain world. Trends Ecol. Evol. 17, 551–557 (2002)
Mori, I., Ohshima, Y.: Neural regulation of thermotaxis in Caenorhabditis elegans. Nature 376, 344–348 (1995)
Mortimer, D., Fothergill, T., Pujic, Z., Richards, LJ., Goodhill, GJ.: Growth cone chemotaxis. Trends Neurosci. 31, 90–98 (2008)
Nagai, K., Sumino, Y., Kitahata, H., Yoshikawa, K. Mode selection in the spontaneous motion of an alcohol droplet. Phys. Rev. 71 (2005)
Nagai, K.H., Takabatake, F., Ichikawa, M., Sumino, Y., Kitahata, H., Yoshinaga N.: Rotational motion of a droplet induced by interfacial tension. Phys. Rev. 87 (2013)
Nakagaki, T., Yamada, H., Toth, A.: Intelligence: maze-solving by an amoeboid organism. Nature 407, 470–470 (2000)
Paxton, W.F., Kistler, K.C., Olmeda, C.C., Sen, A., St Angelo, S.K., Cao, Y., Mallouk, T.E., Lammert, P.E., Crespi, V.H.: Catalytic nanomotors: autonomous movement of striped nanorods. J. Am. Chem. Soc. 126, 13424–13431 (2004)
Petrie, R.J., Doyle, A.D., Yamada, K.M.: Random versus directionally persistent cell migration. Nat. Rev. Mol. Cell Biol. 10, 538–549 (2009)
Peyer, K.E., Zhang, L., Nelson, B.J.: Bio-inspired magnetic swimming microrobots for biomedical applications. Nanoscale 5, 1259–1272 (2013)
Polyakova, T., Zablotskii, V.: Magnetization processes in magnetotactic bacteria systems. J. Magn. Magn. Mater. 293, 365–370 (2005)
Roberts, A.M.: Mechanisms of gravitaxis in chlamydomonas. Biol. Bull. 210, 78–80 (2006)
Roussos, E.T., Condeelis, J.S., Patsialou, A.: Chemotaxis in cancer. Nat. Rev. Cancer 11, 573–587 (2011)
Ševčíková, H., Čejková, J., Krausová, L., Přibyl, M., Štěpánek, F., Marek, M.: A new traveling wave phenomenon of Dictyostelium in the presence of cAMP. Phys. D: Nonlinear Phenom. 239, 879–888 (2010)
Song, L., Nadkarnia, S.M., Boedeker, H.U., Beta, C., Bae, A., Francka, C., Rappele, W.J., Loomisf, W.F., Bodenschatz, E.: Dictyostelium discoideum chemotaxis: threshold for directed motion. Eur. J. Cell Biol. 85, 981–989 (2006)
Sumino, Y., Magome, N., Hamada, T., Yoshikawa, K.: Self-running droplet: emergence of regular motion from non equilibrium noise. Phys. Rev. Lett. 94 (2005)
Sundararajan, S., Lammert, P.E., Zudans, A.W., Crespi, V.H., Sen, A.: Catalytic motors for transport of colloidal cargo. Nano Lett. 8, 1271–1276 (2008)
Takabatake, F., Magome, N., Ichikawa, M., Yoshikawa, K.: Spontaneous mode-selection in the self-propelled motion of a solid/liquid composite driven by interfacial instability. J. Chem. Phys. 134 (2011)
Tanaka, S., Sogabe, Y., Nakata, S.: Spontaneous change in trajectory patterns of a self-propelled oil droplet at the air-surfactant solution interface. Phys. Rev. 91 (2015)
Tomlinson, C.: On the motions of camphor on the surface of water. Proc. R. Soc. Lond. 11, 575–577 (1860)
Toyota, T., Maru, N., Hanczyc, M.M., Ikegami, T., Sugawara, T.: Self-propelled oil droplets consuming “fuel” surfactant. J. Am. Chem. Soc. 131, 5012–5013 (2009)
Walker, G.M., Sai, J., Richmond, A., Stremler, M., Chung, C.Y., Wikswo, J.P.: Effects of flow and diffusion on chemotaxis studies in a microfabricated gradient generator. Lab Chip 5, 611–618 (2005)
Wang, F., Herzmark, P., Weiner, O.D., Srinivasan, S., Servant, G., Bourne, H.R.: Lipid products of PI(3)Ks maintain persistent cell polarity and directed motility in neutrophils. Nat. Cell Biol. 4, 513–518 (2002)
Wu, D.: Signaling mechanisms for regulation of chemotaxis. Cell Res. 15, 52–56 (2005)
Yamamoto, D., Shioi, A.: Self-propelled nano/micromotors with a chemical reaction: underlying physics and strategies of motion control. KONA Powder Part. J. 32, 2–22 (2015)
Zhao, G., Pumera, M.: Macroscopic self-propelled objects. Chem.- Asian J. 7, 1994–2002 (2012)
Zicha, D., Dunn, G.A., Brown, A.F.: A new direct-viewing chemotaxis chamber. J. Cell Sci. 99, 769–775 (1991)
Zigmond, S.H.: Ability of polymorphonuclear leukocytes to orient in gradients of chemotactic factors. J. Cell Biol. 75, 606–616 (1977)
Hong, Y., Blackman, N.M.K., Kopp, N.D., Sen, A., Velegol, D.: Chemotaxis of nonbiological colloidal rods. Phys. Rev. Lett. 99 (2007)
Acknowledgments
This work was supported by the European Commission FP7 Future and Emerging Technologies Proactive: 318671 (MICREAgents) and 611640 (EVOBLISS).
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2017 Springer International Publishing Switzerland
About this chapter
Cite this chapter
Čejková, J., Holler, S., Nguyenová, T.Q., Kerrigan, C., Štěpánek, F., Hanczyc, M.M. (2017). Chemotaxis and Chemokinesis of Living and Non-living Objects. In: Adamatzky, A. (eds) Advances in Unconventional Computing. Emergence, Complexity and Computation, vol 23. Springer, Cham. https://doi.org/10.1007/978-3-319-33921-4_11
Download citation
DOI: https://doi.org/10.1007/978-3-319-33921-4_11
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-33920-7
Online ISBN: 978-3-319-33921-4
eBook Packages: EngineeringEngineering (R0)