
Abstract
Gila trout Oncorhynchus gilae historically occupied high-elevation (>2000 m) headwater streams of the Gila River in southwest New Mexico and central Arizona, USA. By the mid-twentieth century, human activities such as resource extraction and nonnative species introductions caused habitat degradation and loss, thereby restricting the trout to a few small remote streams in the upper Gila River catchment in New Mexico. Since 1989, conservation efforts have been hampered by numerous wildfires that caused elimination of some populations and greatly reduced others. But wildfires also eliminated nonnative fishes from streams thereby making them available for Gila trout repatriation. Elimination of all fishes, including nonnative trout, from upper Black Canyon (ca. 17 km), an East Fork Gila River tributary, by wildfire and associated sediment-laden flows in 1995 and subsequent colonization by native warmwater species (longfin dace Agosia chrysogaster, speckled dace Rhinichthys osculus, Sonora sucker Catostomus insignis, and desert sucker Pantosteus clarkii) provided the opportunity to study the dynamics of a mixed warm-coldwater fish assemblage. Although upper Black Canyon was thermally compromised (summer maxima >25 °C), age-0 hatchery-produced Gila trout (≈1000–2000) were released annually from 1998 through 2012. Establishment of a nonnative piscivore, brown trout Salmo trutta, added complexity to the study. By 2012, native warmwater species had thriving populations but despite limited reproduction annual stocking of Gila trout likely enabled it to persist in the stream. It was not possible to determine the effects of brown trout predation on the Gila trout population or negative interactions, if any, between native warmwater fishes and Gila trout. Regardless, warmwater fish and brown trout abundance was greatest in downstream reaches and that of Gila trout was greatest in the most upstream reach. Inter-annual differences in somatic metrics (condition and size structure) of the Black Canyon Gila trout population were as great as it were between it and reference populations. Unassisted re-population of upper Black Canyon by native warmwater species succinctly illustrated the importance connectivity within a drainage and the rapidity in which this was accomplished is testimony to the resilience of each species to major disturbance. In 2013, wildfire again eliminated all fishes from upper Black Canyon. Hatchery-produced Gila trout were annually stocked from 2013 through 2018 and sampling in 2018 yielded a large number of age-0 fish, suggesting that a Gila trout population might be established in upper Black Canyon, but in the absence of native warmwater fishes and piscivorous brown trout.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
- Mixed fish assemblages
- Oncorhynchus gilae
- Condition factor
- Size structure
- Catastrophic disturbance
- Wildfire
- Nonnative trout
- Assisted repatriation
- Conservation
1 Introduction
Gila trout Oncorhynchus gilae historically inhabited high elevation tributaries of the Gila River in southwestern New Mexico and central Arizona, USA (Fig. 1). By the late nineteenth century, extensive European settlement and accompanying habitat alteration caused by widespread domestic livestock overgrazing, timber harvest, and mineral extraction had rapidly reduced its occurrence to remote, small, upland streams. Its persistence was additionally challenged by extensive introduction of nonnative salmonids, especially rainbow trout O. mykiss and brown trout Salmo trutta. When concerted conservation efforts for the species began in the last quarter of the twentieth century, Gila trout occurred in only five small, isolated streams, all within federally managed lands (Propst et al. 2020).

Gila Trout, Oncorhynchus gilae. Photo by DJ Myers
Initially, the overarching conservation strategy for Gila trout was to secure occupied habitats and replicate each remnant population in other suitable streams. Securing a population typically involved ensuring it was not exposed to nonnative trout invasion by constructed or natural fish movement barriers and closing it to recreational angling. Because each remnant population was genetically distinct (Turner et al. this volume), multiple replicates of each strain were deemed essential to achieve genetic as well as demographic security. Initially, this approach appeared successful in that within about 10 years, each remnant population had at least one replicate. The efficacy of this approach was tested with the elimination of Gila trout from Diamond Creek, the species type locality, by wildfire in 1989 (Propst et al. 1992). Subsequent fires in the mid-1990s and elimination of several Gila trout populations further challenged the merits of Gila trout conservation in small single-strand headwater streams (Brown et al. 2001). Consequently, conservation strategies shifted to establishing Gila trout populations in large, dendritically complex drainages in the belief that such systems would be sufficiently large that if wildfire burned in the catchment Gila trout would have refuge in unburned portions and individuals from unaffected reaches could colonize reaches depopulated by the effects of wildfire.
As conservation strategies for Gila trout were evolving in the 1990s to ameliorate the risk posed by wildfires, empirical and theoretical evidence for climate change, in particular global warming, was rapidly accumulating and model projections for the American Southwest, and the entire Gila River drainage, were particularly dire (Williams et al. 2020). Collectively, these models projected increasing temperatures, increasing aridity, shifting precipitation patterns (including increased frequency of extreme events), diminishment and loss of snowpack, and altered stream flow regimes (Gutzler 2013; Dettinger et al. 2015; Udall and Overpeck 2017). The net effect of climate change would be substantially less optimal habitat for trout, but also an increase in wildfire frequency, extent, intensity, and severity (Westerling et al. 2006).
In a region having comparatively few perennial streams, even at higher elevations, and these are generally disconnected, persistence of robust trout populations is problematic even under current climatic conditions. Optimal habitat for Gila trout is limited and generally occurs >2000 m. Under conditions projected by climate models, the lower elevational limits of optimal habitat will increase (Kennedy et al. 2008). The contraction of optimal habitat will be driven mainly by elevated thermal regimes and diminished and altered flow regimes. With these changes, lower reaches of traditional Gila trout habitat will become inhabitable by warmwater species. Survival of Gila trout might therefore ultimately depend on whether it can coexist with warmwater species in thermally sub-optimal trout habitat.
2 Setting
In 1995, wildfire induced ash- and sediment-laden flows eliminated nonnative trout and native fishes (cyprinids and catostomids) from upper reaches of Black Canyon, a Gila River tributary in southwest New Mexico (Fig. 2). Over the next 2 years, native cyprinids and catostomids from downstream refugia colonized depopulated stream reaches but nonnative trout did not. In June 1998, a fish movement barrier was constructed to preclude return of nonnative salmonids to the upper-most 17 km of Black Canyon and release of hatchery-reared native Gila trout was planned for that autumn. After barrier construction, a final check of Black Canyon was made to ensure absence of nonnative trout; four brown trout and one rainbow trout were found at a single location. Because rainbow trout hybridize with Gila trout and brown trout prey upon and compete with Gila trout their presence was untenable. Surrendering Black Canyon to nonnative trout was not an option. The presence of viable populations of four native warmwater species (longfin dace Agosia chrysogaster, speckled dace Rhinichthys osculus, Sonora sucker Catostomus insignis, and desert sucker Pantosteus clarkii) precluded use of piscicides to eliminate nonnative trout. The alternative was mechanical removal of nonnative trout with electrofishing gear. From June through October 1998, Black Canyon was electro-fished on multiple occasions with multiple crews to capture nonnative trout (Brooks and Propst 1999). In addition to brown trout and rainbow trout, seven cutthroat trout Oncorhynchus clarkii individuals were collected. All brown trout collected (n = 345) were of a single cohort (age 1) and evidently of hatchery origin as scale circuli were evenly spaced, all rainbow trout (n = 24) were sub-adults or adults (>170 mm total length), and cutthroat trout were all adults (>250 mm total length). The likely provenance of brown trout, absence of age-0 rainbow trout, and first record of cutthroat trout strongly suggested their illegal release rather than their invasion from downstream reaches. Based on October sampling, it was likely all rainbow trout and cutthroat trout were removed but possible that some brown trout might remain. Nonetheless, hatchery-produced age-0 (≈100 mm total length) Gila trout were released throughout upper Black Canyon in November 1998.

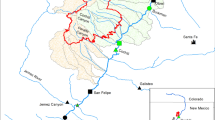
Black Canyon, New Mexico study area
Black Canyon was next sampled in July 2001 and only native fishes were captured. Gila trout was most common in the upper-most reach (Aspen-Falls) and represented by several size classes, including age-0 individuals (<90 mm TL). Speckled dace was the most common fish in both sampled reaches (Barrier-Private reach not sampled in 2001).
The absence of nonnative trout in Black Canyon in 2001 indicated the 1998 removal effort had been successful. In addition, the presence of Gila trout <90 mm TL confirmed in-stream reproduction. At this point, it appeared that Black Canyon could support a mixed warmwater-coldwater assemblage, and the decision was made to not sample Black Canyon for several years. When next sampled in 2008, multiple size classes of brown trout were found throughout Black Canyon upstream of the fish movement barrier. These fish were either progeny of individuals that survived the 1998 mechanical removal effort or individuals and progeny of brown trout that traversed the gabion basket barrier, or both. In 2009, the gabion fish movement barrier was replaced with a concrete structure that presumably was more effective in precluding fish passage to upper reaches.
Beginning in 2008, the paramount management imperative was to suppress or eliminate brown trout so that a viable Gila trout population could be maintained in Black Canyon. A second objective was to document the dynamics of a mixed native warmwater-coldwater fish assemblage. Our expectation was that in time Gila trout would occur primarily, if not exclusively, in upstream, colder reaches (i.e., Aspen-Falls), that reproduction and recruitment by Gila trout would be limited, and that reference Gila trout populations would be demonstrably more robust (e.g., abundance, condition, and size structure) than the population in Black Canyon. Among native warmwater species, we anticipated that speckled dace and desert sucker would be the most successful (e.g., distribution, abundance, and size structure) and that longfin dace and Sonora sucker might maintain smaller populations in upper Black Canyon. The presence of brown trout would provide an opportunity to characterize, if not quantify, the impact of a nonnative predator and competitor on native fishes.
In addition to the presence of a nonnative predator, global warming (Kennedy et al. 2008) was expected to present another challenge to maintenance of a viable Gila trout population in Black Canyon. Regional metrics indicate that the upper Gila River drainage climate is warming, that total annual precipitation is declining, and river flow regimes have been altered. While regional metrics do not reflect exactly the climatic and hydrologic conditions in Black Canyon over this period, they do indicate the changes that have likely occurred—increasing air temperatures, diminished summer flows, altered flow regimes, and elevated water temperatures.
Our purpose herein is to use information obtained on an assemblage of native warmwater fishes and Gila trout to determine if such communities have long-term viability. Specifically, we wished to determine if somatic and demographic attributes of Gila trout in such a mixed assemblage were demonstrably different from those attributes in solely Gila trout populations. In addition, we wished to characterize somatic and demographic attributes of the native warmwater species in the presence of Gila trout. And finally, to evaluate efficacy of mechanical removal of a nonnative salmonid.
3 Approach
Study Area—Black Canyon lies entirely within the Gila National Forest of southwestern New Mexico, USA and almost its entire course is within designated wilderness (Aldo Leopold and Gila Wildernesses). From its origins in the Black Range (elevation ca. 2450 m), Black Canyon flows east to join East Fork Gila River (elevation 1770 m) a short distance upstream of the latter’s confluence with the Gila River (Fig. 2). The study area was in the upper 17 km of Black Canyon, extending from the fish movement barrier (elevation 2074 m) upstream to the confluence of Falls Canyon (elevation 2440 m). In upper reaches, the stream was canyon-bound and bordered by mixed conifers (white fir Abies concolor and Douglas fir Pseudotsuga menziesii) and ponderosa pine (Pinus ponderosa). Near Aspen Canyon confluence, the stream entered a broad valley with scattered ponderosa pine stands and Gamble oak (Quercus gambellii) (Fig. 3). Downstream of Bonner Canyon, the stream progressed through more constricted terrain to the fish movement barrier. Riparian vegetation consisted of ponderosa pine, narrow leaf cottonwood (Populus angustifolia), and willow (Salix sp) was scattered among ponderosa pines along the stream. Three tributaries joined Black Canyon in its course within the study area; Falls Canyon was perennial, and Aspen and Bonner canyons were intermittent. Fish did not occur above the waterfalls a short distance (ca. 50 m) upstream of the confluence of Black and Falls canyons.

Black Canyon near confluence of Aspen Canyon, New Mexico
Methods—The study area was divided into four sampling reaches. The most upstream and canyon-bound reach extended from Aspen Canyon to Falls Canyon confluence (8.7 km). The middle valley reach was from the upstream boundary of a small private inholding to Aspen Canyon (5.8 km). The penultimate downstream reach was from the fish movement barrier to the downstream boundary of the private property (1.3 km). The stream was not sampled in the 1.1 km private property reach. And the lowest was a 3-km reach downstream of the fish movement barrier. Nonnative salmonids were not removed from this reach, and it was accessible to fishes from downstream reaches and tributaries (Whitney et al. 2017; Hedden et al. 2022). Species codes (first three letters of genus and species, e.g., AGOCHR = Agosia chrysogaster longfin dace and ONCGIL = Oncorhynchus gilae Gila trout) were used on graphs and tables to conserve space.
Following release of Gila trout in 1998, Black Canyon was not sampled until 2001. After a 7-year hiatus, it was again sampled in 2008. Thence, the stream was sampled annually in July or August through 2012. The extent of the study area sampled in a year was dependent upon personnel availability and weather; the Barrier-Private reach was sampled in all years (June in 2008), Private-Aspen reach in all years, and Aspen-Falls reach in 2008, 2011, and 2012. The fish assemblage downstream of the barrier was sampled multiple times prior to, during, and after the 2008–2012 sampling period. The Below Barrier reach data are presented to provide a contrast to above barrier data. Two or three crews, each composed of 3-5 individuals and sampling in different stream sections, used battery-powered backpack shockers to stun fish. Stunned fish were netted and mass (±1 g) and total length (±1 mm, TL) of large-bodied individuals (≥100 mm TL) determined. Small-bodied individuals were typically only counted, but in several years, TL was measured for subsamples of small-bodied fishes. In 2011, small-bodied fishes were not counted; elevated water temperatures and handling stress necessitated rapid processing of fish to reduce mortality. All fish, except nonnative brown trout, were returned to stream near point of capture. Abundance was the number of individuals captured/elapsed time electrofishing (#/min).
Water temperature data were obtained from temperature loggers deployed by New Mexico Surface Water Quality Bureau (NM Environment Department). Regional precipitation and temperature data 1980 through 2020 were obtained from four weather stations in the upper Gila River drainage (USGS HUC # 15040001) NOAA website (www.ncdc.noaa.gov/cdo-web). Two stations (Lookout Mountain and Signal Peak) were at high elevations (2591 m and 2548 m, respectively) and the remaining two (Beaverhead and Gila Hot Springs) were somewhat lower (2033 m and 1718 m, respectively). Gila River discharge data were obtained from the USGS Gila River near Gila gage (# 09430500).
Three metrics were used to characterize the well-being of Gila trout, brown trout, Sonora sucker, and desert sucker: condition, size structure, and abundance (catch-per-unit-effort: #/min elapsed electrofishing time). Because Gila trout have near isometric growth (i.e., W = aL3) throughout their lives, Fulton’s Condition Factor was used to estimate general physiological condition of Gila and brown trout (Pope and Kruse 2007). The factor is determined by KTL = (Mass/Total Length3) * 105. Because condition was normally distributed for each year and reach, ANOVA was used to compare mean condition between years, reaches, and populations. The Kolmogorov–Smirnov Goodness of Fit two-sample test was used to compare annual intra-reach, inter-reach, and population size structure (Neumann and Allen 2007). Specimens were grouped in 10-mm TL classes for K-S analysis. Gila trout specimens <70 mm TL (age 0) and brown trout specimens <100 mm TL were excluded from analyses. All statistical procedures were done with XLSTAT (Addinsoft 2021).
Reference Gila trout populations were in McKnight and Diamond creeks. Diamond Creek (elevation ca. 2430 m) was about 14 air km north of Black Canyon while McKnight Creek (elevation ca. 2340 m) was about 14 air km south of Black Canyon. Both streams were roughly comparable to Black Canyon in habitat dimensions (stream width, depth, and discharge) but upper Black Canyon was substantially longer (≈18 km) than Diamond Creek (≈6 km) and McKnight Creek (≈8 km). There was no stream unaffected by wildfire and drought within the current range of Gila trout that also had a mixed warmwater-coldwater assemblage that might serve as a reference for mixed assemblage or warmwater species comparisons. Black Canyon downstream of the fish movement barrier was sampled (nonnative salmonids not removed) relatively frequently from 1996 through 2021 and thus provided some perspective on the changes in the fish assemblage in upstream reaches.
Following an initial stocking of age-0 (70–115 mm TL, number not recorded) hatchery-produced Gila trout in Black Canyon upstream of the barrier in late 1998, it was stocked annually thereafter in autumn. Numbers released annually ranged from several hundred to several thousand. Most releases were of age-0 fishes (80–115 mm TL), but two releases were of larger and older fish (105 individuals averaging about 300 mm TL in 2009 and about 1000 individuals averaging 150 mm TL in 2016). Individuals <100 mm TL were considered juveniles, those between 101 and 150 sub-adults, and >150 adults (Propst and Stefferud 1997).
4 Results
Regional Climatic, Discharge, and Water Temperature Patterns (1980–2020)—Total annual precipitation over the past 40 years has declined significantly at four upper Gila River catchment weather stations (Fig. 4). Although total annual precipitation declined, mean daily discharge at the Gila River near Gila gage did not (Fig. 5). However, extreme flow events (floods and minimum flows) increased. Since 1980, there have been 10 annual peak flows exceeding 283 m3/s (=104 ft3/s), whereas in the preceding 51 years there were only 4 years with peak flows greater than 283 m3/s. Annual minimum daily discharge and summer (June and July) mean daily discharge decreased over the 1980–2020 period. At the two weather stations that recorded mean daily air temperature, mean annual temperature increased significantly (Fig. 6). Temperature loggers deployed upstream of Aspen Canyon confluence and at Aldo Leopold Wilderness boundary recorded mean daily water temperatures in 2012 approaching 20 °C, but maximum daily water temperatures exceeded 25 °C frequently in June at both locations and less frequently in July and August (Fig. 7).

Total annual precipitation at four weather stations in the upper Gila River catchment. Dashed lines indicate significant (α = 0.05) relationship

Discharge attributes of Gila River at USGS Gila River near Gila gage (09430500), New Mexico. Dashed lines indicate significant (α = 0.05) relationship

Mean annual air temperature at two weather stations in upper Gila River catchment, New Mexico. Dashed lines indicate significant (α = 0.05) relationship

Mean, maximum, and minimum daily water temperature in Black Canyon at Aldo Leopold Wilderness boundary (a) and just upstream of Aspen Canyon confluence (b), New Mexico, May–October 2012
Temporal Changes in Fish Assemblage—Although the upper Black Canyon fish assemblage was greatly reduced, if not eliminated, by ash- and sediment-laden flows associated with the 1995 Bonner Fire, native cyprinids and catostomids were comparatively common in the Below Barrier reach a year after the fire and especially in 1998 (Fig. 8). Speckled dace was the most common species in all collections and nonnative salmonids were rare. Although abundance was variable across collections, fish numbers, especially speckled dace, increased substantially in 2012 and 2013, crashed following ash and sediment flows caused by the 2013 Silver Fire, and no fish was collected in this reach until October 2014 when 12 speckled dace and a single Gila trout were collected. Thereafter, fish abundance increased slightly through 2021 but remained considerably less than in the collections preceding the Silver Fire.

Abundance (#/min) of fishes in Black Canyon Below Barrier reach, New Mexico, 1996–2021. Month of sampling in parentheses
Within a year of the Bonner Fire, native fishes moved from downstream refugia to the upper reaches of Black Canyon. Thereafter, fish abundance in the reaches upstream of the barrier generally increased through 2012 (Fig. 9). Ash and sediment flows associated with the Aspen Fire contributed to the comparatively low abundance of fishes in the Barrier-Private and Private-Aspen reaches in 2010. Speckled dace was typically the most common fish in all reaches in all years; low abundance of fishes in 2011 was largely a reflection of partially counting small-bodied fishes in the Barrier-Private reach and not counting them in the Private-Aspen and Aspen-Falls reaches. Neither brown nor Gila trout was common in any reach in 2011 but desert sucker was moderately common in the Aspen-Falls reach. In 2012, fishes attained their greatest abundance in the Barrier-Private and Private-Aspen reaches while that in the Aspen-Falls reach was moderately high. The 2013 Silver Fire eliminated fishes from all reaches (Fig. 10). In 2013 and subsequent years, hatchery-reared Gila trout were released in autumn at several locations in the above-barrier reaches. Most, if not all, Gila trout captured in the Aspen-Falls reach in 2016 were hatchery-produced fish. But in August 2018, a large portion of the sample in the Private-Aspen reach was age-0 fish (<90 mm TL), which were almost certainly spawned in the stream. The fish movement barrier precluded the natural colonization of the upper reaches of Black Canyon by native warmwater species.

Abundance (#/min) of fishes in Black Canyon reaches upstream of fish movement barrier, 1996–2018. Aspen Fire not indicated on Aspen-Falls reach because it did not burn in Aspen-Falls catchment. Note different abundance axes

Debris and fine sediment deposits in Black Canyon in Barrier-Private reach following the 2013 Silver Fire
Longitudinal Abundance of Fishes—Although variable from year to year, abundance of warmwater species generally declined from down- to upstream (Fig. 11). In average, longfin dace and speckled dace were roughly equally abundant in the Barrier-Private and Private-Aspen reaches and rare or uncommon in the most upstream reach. Sonora sucker was most common in the Barrier-Private reach and absent in the most upstream. Desert sucker abundance was greatest in the Barrier-Private reach, and it was more common in the most upstream reach than other warmwater species. Gila trout abundance increased from lowest to most upstream reach. In contrast, brown trout abundance was greatest in the middle reaches, and there its abundance was low compared to Gila trout. Since the 2013 Silver Fire only Gila trout occupy the reaches upstream of the fish movement barrier.

Abundance of fishes from most downstream reach to most upstream. The Below Barrier and Aspen-Falls reaches were not sampled in 2009 and 2010. The Below Barrier reach was sampled one (2008), three (2011), and four (2012) times. In 2011, small-bodied fishes in Private-Aspen and Aspen-Falls reaches were not counted. Note different scales on Abundance axis
Size structure of Black Canyon Gila Trout Population and Reference Populations—During 2008–2012, the number of Gila trout captured varied considerably across reaches and years within each reach (Table 1). Age-0 individuals (≤90 mm TL) were rarely collected in any reach in any year (Fig. 12). An unknown portion of the fishes collected each year in each reach in July–August were hatchery-produced fish. When stocked (typically October or November) most releases were comprised of age-0 (<130 mm TL) but in 3 years larger fish (157–196 mm TL) were released and in 2009 about 100 >275 mm TL were released (Table 2). Some stocked fish likely survived to be collected the following year as age-1 individuals between 100- and 150-mm TL. And some stocked fish survived for multiple years. Most collections were comprised largely of fish <200 mm TL, but in several years fish >200 mm was a substantial portion of the collection (e.g., 2010 and 2011 in Private-Aspen reach).

Size structure of Black Canyon Gila trout population, 2008–2012
Size structure of Gila trout in each reach across years was generally similar. Between year differences in size structure were different for only two comparisons: Barrier-Private 2011–2012 and Aspen-Falls 2011–2012 samples (Table 3). Size structure was different for all Barrier-Private/Private-Aspen comparisons, except 2012 (Table 4). The size structure of the 2012 Private-Aspen/Aspen-Falls samples was different.
The size structure of the Black Canyon Gila trout population was different from that of the Diamond population in 2008 and McKnight population in 2011 (Table 5). Figure 13 illustrates the differences in size structure of the populations. In both Diamond and McKnight creeks maximum specimen TL rarely exceeded 220 mm whereas in Black Canyon several individuals in most years exceeded >300 mm TL.

Size structure of Black Canyon Gila trout population and that of its reference populations. 2008 Black Canyon—Diamond Creek and 2011 Black Canyon—McKnight Creek were significantly different (see Table 5). Note different proportion axis scale on Black Canyon July 2001 graph
Post-Silver Fire monitoring of the above barrier Gila trout population was initiated in 2015. Most, if not all fish, collected in 2015 and 2016 were hatchery-produced individuals (Fig. 14). In 2018, however, a substantial number of age-0 (<90 mm TL) individuals were collected.

Post-Silver Fire size structure of Gila trout population in Black Canyon upstream of fish movement barrier. The September 2015 and August 2018 samples were taken in Private-Aspen and Aspen-Falls reaches whereas the May 2016 sample was from the Aspen-Falls reach. Note the different proportion axis scale for the 2016 collection
Condition (KTL,) of Black Canyon Gila Trout and Reference Populations—Mean Gila trout condition was >0.9000 in all years in all Black Canyon reaches, except Barrier-Private in 2009 when it was 0.8341 (Table 6). Most variation in mean annual condition was in the Barrier-Private reach. Mean condition of Gila trout in the Barrier-Private reach in 2009 was significantly less than that in 2008 and 2010 (Table 7). In 2011, mean condition in the reach was greater than that in 2012 but not 2010. In the Private-Aspen reach, annual mean condition differences from 2008–2009 through 2010–2011 were significant but that of 2011–2012 was not. Only two Barrier-Private/Private-Aspen condition comparisons were different; condition was less in Barrier-Private reach in 2009, but greater in 2011 (Table 8). All Private-Aspen/Aspen-Falls comparisons were non-significant; mean condition of the 2011 Private-Aspen/Aspen-Falls samples was virtually identical. In two of four population condition comparisons, KTL of the Black Canyon population was greater than that of the reference population and less in the other two (Table 9). All population comparisons yielded significant differences.
Brown Trout Population Attributes—Although absent in 2001, brown trout occurred throughout the upper reaches of Black Canyon in 2008. Despite annual efforts (2008–2012) to suppress their numbers by removing all individuals captured, the number captured generally increased over time (Table 10). The size structure of the population (reaches combined) indicated that reproduction and recruitment occurred each year from 2007 through 2011, except 2008 (Fig. 15). Most individuals collected were <300 mm TL, but larger individuals were found each year. No between year size-structure differences were detected (Table 11). Mean condition of brown trout each year >1.0000 in all years (Table 12). Between year condition was different in 2009–2010 and 2010–2011 (Table 13).

Size structure of the Black Canyon brown trout population upstream of fish movement barrier (reaches combined), 2008–2012. Aspen-Falls reach not sampled in 2009 and 2010
Mass of Gila and Brown Trouts—Total mass of collected Gila trout (excluding individuals <70 mm TL) was variable across years and reaches (Table 14). It generally increased in Barrier-Private and Aspen-Falls reaches from 2008 through 2012. In Private-Aspen reach, total mass was greatest in 2008 and steadily declined to its lowest in 2012. In years all reaches were sampled, total Gila trout mass was least in 2011 and greatest in 2012. Brown trout total mass (excluding individuals <70 mm TL) was likewise variable among reaches and year to year. It was greatest in Barrier-Private and Aspen-Falls in 2012. Total brown trout mass increased from 2008 through 2012. Brown trout mass exceeded that of Gila trout in all years in Barrier-Private, all but one in Private-Aspen, and never in Aspen-Falls.
Warmwater Species Population Attributes—Because large numbers of desert sucker were often collected, TL and mass were obtained from a representative subsample of desert suckers collected in each reach each year. For example, in the Private-Aspen reach in 2008, 595 desert suckers were captured, TL was obtained from 131 and mass determined for 126. Desert sucker abundance generally increased from 2008 through 2012 (Table 15). Although the range of TL was similar across years, mean TL was more variable (Fig. 16). Size structure of the Black Canyon desert sucker population, however, did not differ from one year to the next (Table 16). Mean annual condition of desert suckers exceeded 1.1 in all years and was greatest in 2010 (Table 17). Between year mean condition differences were different for all years (Table 18).

Black Canyon desert sucker population size structure (reaches combined), 2008–2012
The TL range of Sonora suckers captured each year indicated multiple age classes and annual reproduction (Table 19). Most individuals were between 100- and 250-mm TL but several larger fish were captured in all years (Fig. 17). Size structure of the population did not differ from year to year (Table 20). Mean condition of the population exceeded 1.2 in all years but 2009 when it was slightly less (Table 21). Mean condition was different only between 2009 and 2010 (Table 22).

Black Canyon Sonora sucker population size structure (reaches combined), 2008–2012
In 2008 and 2011, small subsamples of speckled dace were measured, and a larger subsample was measured in 2012 (Fig. 18a). Several individuals >100 mm TL were captured in 2008 and 2012. The size range of individuals collected indicated successful reproduction and recruitment each year. The abundance of speckled dace varied considerably from year to year but was greatest in 2012 (Fig. 18b).

(a) Black Canyon speckled dace population size structure (reaches combined), 2008, 2011, and 2012. (b) Abundance of speckled dace and longfin dace in Black Canyon, 2008–2012. A subsample of specimens of each species was tallied only in Barrier-Private reach in 2011. (c) Black Canyon longfin dace population size structure in Black Canyon (reaches combined), 2011 and 2012
A subsample of longfin dace in the 2011 and 2012 collections was measured. Most specimens were <65 mm TL (Fig. 18c). The presence of longfin dace <40 mm TL indicated successful reproduction in both years. Like speckled dace, longfin dace abundance varied considerably from year to year.
5 Implications and Conclusions
For species restricted to high elevation cool-coldwater habitats, climate change presents multiple challenges to their persistence. For Gila trout that naturally had a comparatively limited distribution in headwater reaches of streams draining the Mogollon Rim of central Arizona and southwest New Mexico (Behnke 2002), the contraction (within stream shrinkage of habitat) and reduction (loss of perennial streams) of suitable habitat imposes an additional layer of difficulty to its survival. Over past 20 years or so, climate models have predicted altered flow regimes and more extreme flow events (Stewart et al. 2005; Hamlet and Lettenmaier 2007; Wenger et al. 2011), increasing aridity (Seager et al. 2007; Seager and Vecchi 2010; Udall and Overpeck 2017; Overpeck and Udall 2020), elevated stream thermal regimes (Schultz et al. 2017), and more large and intense wildfires (Chikamoto et al. 2017; Brown et al. 2021). Exacerbating the changes wrought by extended drought (Cook et al. 2015; Ault 2020) is the increasing frequency and intensity of wildfire (Dennison et al. 2014; Abatzoglou and Williams 2016; Singleton et al. 2019). These projections and events are playing out over the upper Gila River catchment as illustrated by regional temperature, precipitation, stream flow records, and multiple large wildfires. Since 2000, 21 wildfires have each burned over 4050 ha on the Gila National Forest, including the 56,175 ha Silver Fire (Gila National Forest 2021). Over the past 40 years, total annual precipitation has decreased and mean annual ambient temperature has increased in the Gila River catchment. Both contribute to warming of streams that historically provided habitat for Gila trout. For a sibling species, Apache trout Oncorhynchus apache, median temperature for optimal growth must be below 19 °C and lower if daily fluctuations are >12 °C (Recsetar et al. 2014). Median 2012 summer (June–July) water temperatures in Black Canyon were 17.2 and 16.8 °C at Aldo Leopold Wilderness boundary and above Aspen Canyon confluence, respectively, but daily fluctuations at both locations were >12° almost every day in June 2012. Mean annual daily discharge in the Gila River has not declined since 1980, but frequency of days with extremely low mean discharge has increased substantially. Collectively, these metrics (wildfire frequency, declining annual precipitation, increasing ambient temperature, and altered flow regimes) indicate that many, if not all, streams within the upper Gila River catchment are becoming less suitable for maintaining Gila trout populations.
The elimination of fishes from upper Black Canyon by the 1995 Bonner Fire provided an opportunity to determine the potential for Gila trout to persist in a thermally compromised stream. And the movement of native warmwater species into upper Black Canyon added another dimension to evaluate: the potential for coexistence in a mixed warmwater-coldwater fish assemblage. An additional challenge for Gila trout and fellow native fishes was the return of nonnative piscivorous brown trout, especially individuals >175 mm TL (Jonsson et al. 1999; Budy and Gaeta 2018), to upper Black Canyon.
The rapidity with which the four native warmwater species recolonized upper Black Canyon was impressive. Although the specific location of the refugia they survived ash and sediment flows in is uncertain, it is likely colonizers moved from perennial tributaries (Squaw and Apache creeks) about 12 river km downstream of the fish movement barrier and not affected by recent wildfires. Small-scale movement (≈2 km) of Sonora sucker and desert sucker was documented by Booth and Shipley (2012) and Booth et al. (2013). Based on otolith microchemistry, Whitney et al. (2017) provided information indicating that speckled dace colonizers were likely from downstream Black Canyon tributaries or survived in the most downstream reaches of Black Canyon near the East Fork Gila River confluence. Although Whitney et al. (2017) did not have data to support longfin dace, Sonora sucker, or desert sucker movement from downstream tributaries, it is likely these refugia were the source of upstream colonizers. Presence of each native warmwater species in Squaw and Apache creeks and lower Black Canyon was confirmed in 2020 (Paggen, unpubl. data). If Squaw and Apache creeks were the source of colonizers, speckled dace moved over 20 km to the mid-reach of upper Black Canyon (about confluence of Aspen Canyon) in a year or less (July 1995–May 1996). The other species were all present in upper Black Canyon by 2001. By 2008, each species had substantial populations composed of multiple size classes in the lower two study reaches (Barrier-Private and Private-Aspen) and desert sucker regularly occurred in the upper-most reach (Aspen-Falls). Both speckled dace and desert sucker met our expectations that each would establish populations in the study reach but we did not expect that both longfin dace and Sonora sucker would also have multiple size-class populations in upper Black Canyon. Abundance of longfin dace was quite variable from year-to-year, indicating its status in upper Black Canyon was more tenuous than that of other native warmwater species.
Gila trout lacked a pathway for a natural return to Black Canyon. The nearest Gila trout population, Diamond, was over 70 river km distant and where streams were perennial provided habitat for only warmwater species, including piscivorous smallmouth bass Micropterus dolomieu (Johnson et al. 2008). Thus, the annual release of hatchery-produced Gila trout in upper Black Canyon from 1998 through 2012 was necessary to establish and potentially maintain a population. Population augmentation, however, made determination of whether Black Canyon could sustain a naturally recruiting Gila trout population problematic. Using the presence of individuals <100 mm TL in August (i.e., Fig. 12) as an indicator of successful in-stream reproduction, there was limited survival of age-0 individuals in 3 years between 2008 and 2012. Nonetheless, from 2008 through 2012, multiple size classes were present in each above-barrier reach. When compared to reference populations, the size structure of Black Canyon was different in two comparisons (Black-Diamond 2008 and Black-McKnight 2011), which was likely due to the number of large Gila trout (>250 mm TL) in Black Canyon and absence of any fish >250 mm TL in Diamond or McKnight creeks. Abundance of Gila trout in each Black Canyon reach was <1.0/min and varied considerably across years, especially in the Barrier-Private reach. Mean annual condition (KTL) exceeded 1.0 only once in each Barrier-Private and Private-Aspen. Comparisons of the Black Canyon population to reference populations did not yield consistent differences. For example, mean KTL of Black Canyon fishes was greater than reference population in two comparisons, but in two others the reference population KTL was greater. In 2012, the longitudinal distribution of Gila trout indicated that if Black Canyon was to support a viable Gila trout population it would be centered in the most upstream reaches.
The long-term potential for native warmwater species and Gila trout coexistence appeared possible based on the longitudinal distribution of native fishes in upper Black Canyon and the robust warmwater species populations. But could the Gila trout population persist without frequent augmentation and the continued suppression of nonnative piscivorous brown trout? The low numbers and irregular collection of age-0 Gila trout might reflect brown trout predation and the need for continued Gila trout augmentation. Brown trout tended to be more common in Barrier-Private and Private-Aspen reaches and Gila trout was more common in the Aspen-Falls reach. Whether this pattern was a consequence of brown trout occurring where there was abundant warmwater fish prey or other factor(s) is impossible to discern. Regardless of the specific reason(s) for the observed distribution pattern of brown trout, it was apparent that suppression efforts had to continue and likely be more frequent and intensive, similar to successful suppression efforts elsewhere (Healy et al. 2020), to enable a mixed warm-coldwater assemblage to coexist in Black Canyon.
Our study ended abruptly in early summer 2013 when monsoon rains following the Silver Fire washed ash and fine sediments into Black Canyon eliminating all fishes upstream of the fish movement barrier. Although a number of physical and chemical factors, including displacement by elevated flows, might have contributed to elimination of fishes (Gresswell 1999), it is likely a combination of scoured stream channel in upper-most reaches and deposition of large volumes of fine sediments in middle and lower reaches were major contributors to elimination of fishes, similar to what Rust et al. (2019) reported for upper Rio Grande streams following wildfire and late summer storms.
Following the Silver Fire hatchery-produced Gila trout were released annually in upper Black Canyon and in August 2018 the collection of large numbers of individuals <90 mm TL indicated successful in situ spawning and recruitment. In the absence of brown trout, and native warmwater species, it appears Gila trout might establish a viable population in upper Black Canyon. Clearly, native speckled dace, desert sucker, and Sonora sucker could thrive in upper Black Canyon, and it is likely the fourth, longfin dace, could also. What is less certain is whether Gila trout could in the presence of native warmwater fishes. Nonetheless, the results of our study are encouraging in that native warmwater species maintained robust populations and Gila trout maintained mixed size classes, albeit with annual augmentation, in Black Canyon despite the presence of piscivorous brown trout. With brown trout eliminated, we believe that a coexisting warm-coldwater fish assemblage in upper Black Canyon is possible. Other studies (e.g., Dunham et al. 2003; Gido et al. 2019; Jager et al. 2021) have reported the resilience of native fishes to catastrophic wildfire and this study provides yet another example of the resilience of native warmwater species to such events. But this resilience cannot be manifested without connectivity among refugia and depopulated streams, as demonstrated by native warmwater fishes rapidly colonizing depopulated reaches of Black Canyon and the inability of Gila trout to populate Black Canyon without human assistance.
References
Abatzoglou JT, Williams AP (2016) Impact of anthropogenic climate change on wildfire across western US forests. Proc Natl Acad Sci 113:11770–11775
Addinsoft (2021) XLSTAT statistical and data analysis solution, New York. https://www.xlstat.com
Ault TR (2020) On the essentials of drought in a changing climate. Science 368:256–260
Behnke RJ (2002) Trout and Salmon of North America. The Free Press, New York
Booth MT, Shipley AJ (2012) Spatial dynamics and growth of two native species of catostomids: are movements restricted? Southwest Nat 57:248–256
Booth MT, Hairston NG Jr, Flecker AS (2013) How mobile are fish populations? Diel movement, population turnover, and site fidelity in suckers. Can J Fish Aquat Sci 70:666–677
Brooks JE, Propst DL (1999) Nonnative salmonid removal from the Black Canyon drainage, East Fork Gila River, June-October 1998. U.S. Fish Wildl Serv, New Mex Fish Resource Office, Albuquerque
Brown DK, Echelle AA, Propst DL, Brooks JE, Fisher WL (2001) Catastrophic wildfire and number of populations as factors influencing risk of extinction for Gila trout (Oncorhynchus gilae). West N Am Nat 61:139–148
Brown EK, Wang J, Feng Y (2021) US wildfire potential: a historical view and future projection using high-resolution climate data. Environ Res Lett 16(2021):034060
Budy P, Gaeta JW (2018) Brown trout as an invader: a synthesis of problems and perspectives in North America. In: Lobon-Cervia J, Sanz N (eds) Brown trout: biology, ecology, and management. Wiley, Hoboken, NJ, pp 525–543
Chikamoto Y, Timmermann A, Widlansky MJ et al (2017) Multi-year predictability of climate, drought, and wildfire in southwestern North America. Sci Rep – UK 7:6568. https://doi.org/10.1038/s41598-017-06869-7
Cook BI, Ault TR, Smerdon JE (2015) Unprecedented 21st century drought risk in the American Southwest and Central Plains. Sci Adv 1:e1400082
Dennison PE, Brewer SC, Arnold JD et al (2014) Large wildfire trends in the western United States, 1984-2011. Geophys Res Lett 41:2928–2933
Dettinger M, Udall B, Georgakakos A (2015) Western water and climate change. Ecol Appl 25:2069–2093
Dunham JB, Young MK, Gresswell RE et al (2003) Effects of fire on fish populations: landscape perspectives on persistence of native fishes and nonnative fish invasions. Forest Ecol Manag 178:183–196
Gido KB, Propst DL, Whitney JE et al (2019) Pockets of resistance: response of arid-land fish communities to climate, hydrology, and wildfire. Freshw Biol 64:761–777
Gila National Forest (2021) Fire history 1974-2020-Gila National Forest. MAP, Silver City, NM
Gutzler D (2013) Regional climatic considerations for borderlands sustainability. Ecosphere 4(1):7. https://doi.org/10.1890/ES12-00283.1
Gresswell RE (1999) Fire and aquatic ecosystems in forested biomes of North America. J Am Fish Soc 128:193–221
Hamlet AF, Lettenmaier DP (2007) Effects of 20th century warming and climate variability on flood risk in the western U.S. Water Resour Res 43:W06427. https://doi.org/10.1029/2006WR005099
Healy BD, Schelly RC, Yackulic CB, Omana Smith EC, Budy P (2020) Remarkable response of native fishes to invasive trout suppression varies with trout density, temperature, and annual hydrology. Can J Fish Aquat Sci 77:1446–1462. https://doi.org/10.1139/cjfas-2020-0028
Hedden C, Propst DL, Gido KB, Hedden SC, Whitney JE (2022) Differential responses of native fishes in two headwater tributaries of the Gila River following severe wildfires. West N Am Nat 82:201–207
Jager HI, Long JW, Malison RL et al (2021) Resilience of terrestrial and aquatic fauna to historical and future wildfire regimes in western North America. Ecol Evol 00:1–26. https://doi.org/10.1002/ece3.8026
Johnson BM, Martinez PJ, Hawkin JA et al (2008) Ranking predatory threats by nonnative fishes in the Yampa River, Colorado, via bioenergetics modeling. N Am J Fish Manage 28:1941–1953
Jonsson N, Naesje TF, Jonsson B et al (1999) The influence of piscivory on life history traits of brown trout. J Fish Biol 55:1129–1141
Kennedy TL, Gutzler DS, Leung RL (2008) Predicting future threats to the long-term survival of Gila trout using a high-resolution simulation of climate change. Clim Change 94:503–515
Neumann RM, Allen MS (2007) Size structure. In: Guy CS, Brown ML (eds) Analysis and interpretation of freshwater fisheries data. American Fisheries Society, Bethesda, MD, pp 375–421
Overpeck JT, Udall B (2020) Climate change and the aridification of North America. Proc Natl Acad Sci U S A 117:11856–11858
Pope KL, Kruse CG (2007) Condition. In: Guy CS, Brown ML (eds) Analysis and interpretation of freshwater fisheries data. American Fisheries Society, Bethesda, MD, pp 423–471
Propst DL, Stefferud JA (1997) Population dynamics of Gila trout in the Gila River drainage of the south-western United States. J Fish Biol 51:1137–1154
Propst DL, Stefferud JA, Turner PR (1992) Conservation and status of Gila trout, Oncorhynchus gilae. Southwest Nat 37:117–125
Propst DL, Turner TF, Monzingo JA et al (2020) Ecology, politics, and conservation of Gila trout. In: Propst DL, Williams JE, Bestgen KR, Hoagstrom CW (eds) Standing between life and extinction: ethics and ecology of conserving aquatic species in North American Deserts. The University of Chicago Press, Chicago, pp 285–294
Recsetar MS, Bonar SA, Feuerbacher OG (2014) Growth and survival of Apache trout under static and fluctuating temperature regimes. Trans Am Fish Soc 143:1247–1254. https://doi.org/10.1080/00028487.2014.931298
Rust AJ, Randell J, Todd AS et al (2019) Wildfire impacts on water quality, macroinvertebrate, and trout populations in the upper Rio Grande. Forest Ecol Manag 453(2019):117636. https://doi.org/10.1016/j.foreco.2019.117636
Schultz LD, Heck MP, Hockman-Wert D et al (2017) Spatial and temporal variability in the effects of wildfire and drought on thermal habitat for a desert trout. J Arid Environ 145:60–66
Seager R, Vecchi GA (2010) Greenhouse warming and the 21st century hydroclimate of southwestern North America. Proc Natl Acad Sci U S A 107:21277–21282
Seager R, Ting M, Held I, Kushnir Y, Lu J, Vecchi G, Huang H-P, Harnik N, Leetmaa A, Lau N-C, Li C, Velez J, Naik N (2007) Model projections of an imminent transition to a more arid climate in southwestern North America. Science 316:1181–1184
Singleton MP, Thode AE, Sanchez Meador AJ et al (2019) Increasing trends in high-severity fire in the southwestern USA from 1984 to 2015. Forest Ecol Manag 433:709–719
Stewart IT, Cayan DR, Dettinger MD (2005) Changes towards earlier streamflow timing across western North America. J Climate 18:1136–1155
Udall B, Overpeck J (2017) The twenty-first century Colorado River drought and implications for the future. Water Resour Res 53:2404–2418. https://doi.org/10.1002/2016WR019638
Wenger SJ, Isaack DJ, Luce CH et al (2011) Flow regime, temperature, and biotic interactions drive differential declines of trout species under climate change. Proc Natl Acad Sci U S A 108:14175–14180
Westerling AL, Hidalgo HG, Cayan DR et al (2006) Warming and earlier spring increase western U.S. forest wildfire activity. Science 313:940–943
Whitney JE, Gido KB, Hedden SC et al (2017) Identifying the source population of fish re-colonizing an arid-land stream following wildfire-induced extirpation using otolith microchemistry. Hydrobiologia 797:29–45. https://doi.org/10.1007/s10750-017-3143-1
Williams AP, Cook ER, Smerdon JE et al (2020) Large contribution from anthropogenic warming to an emerging North American megadrought. Science 368:314–318
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2024 This is a U.S. government work and not under copyright protection in the U.S.; foreign copyright protection may apply
About this chapter

Cite this chapter
Propst, D.L., Myers, D.J., Wick, J.M., Paggen, R.J. (2024). Dynamics of a Warmwater-Coldwater Fish Assemblage in a Wildfire-Prone Landscape. In: Lobon-Cervia, J., Budy, P., Gresswell, R. (eds) Advances in the Ecology of Stream-Dwelling Salmonids. Fish & Fisheries Series, vol 44. Springer, Cham. https://doi.org/10.1007/978-3-031-44389-3_27
Download citation
DOI: https://doi.org/10.1007/978-3-031-44389-3_27
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-031-44388-6
Online ISBN: 978-3-031-44389-3
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)