
Abstract
Herein, general aspects of the relationship between lymnaeid snails and their parasites are reviewed from a mechanistic point of view. It includes an outlook to encounter filters whilst emphasizing on compatibility filters driven by the different components of the internal defense system within Lymnaeidae, and to the immunobiological interactions that determined the recognition and elimination of the pathogen upon contact, or the establishment of the dynamic equilibrium characteristic of a successful infection. The negative effects caused by parasitism on the physiology and life history traits of the lymnaeid hosts are briefly reviewed, as well as the role of ecological factors on modifying encounter and compatibility filters and the ultimate infection outcome. Finally, insights into the significance of building up knowledge on the biology of host–parasite interaction will be discussed to introduce the opportunities and challenges that lies ahead concerning the family Lymnaeidae and their most significant pathogens. Most of the published works on the subject deals with Lymnaeidae–trematode interaction and consequently, this represents the main topic reviewed in this chapter.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
11.1 Introduction
Lymnaeid snails are known hosts of a plethora of pathogens, mainly within the class Trematoda, among which there are some veterinary and/or medically-relevant parasites, e.g. Fasciola spp., Diplostomum spp., Trichobilharzia spp., Echinostoma spp.. For instance, in a family of around 100 species described, the species Radix natalensis is involved in the life cycles of at least 71 trematodes belonging to 13 different families that affect birds and both domestic and wild mammals as definitive hosts (Brown 1978), and this without counting digeneans of lower vertebrates such as amphibians. Therefore, most of the works published deals with trematode–Lymnaeidae interactions and consequently, this represents the main topic reviewed within the present chapter.
In the next sections, we will review some general aspects of the relationship between lymnaeid snails and their parasites from a mechanistic point of view. In this sense, several features, mechanisms, and conditions which involve not only the parasite and the lymnaeid snail but also the environment relevantly contribute in shaping the landscape of snail host specificity and infection success; from the pre-penetration step to the final outcome of each interaction. Two “filters” have been proposed by Combes (2001) aiming to distinguish the main components encompassing parasite success: the encounter filter and the compatibility filter. The first one comprises the elements affecting the probability of contact between the parasite and the host and includes appropriate spatial and temporal overlap as well as the ability to enter the potential host. The compatibility filter determines whether the parasite succeeds in infecting and stablishing itself in the host. Among the determining elements of the compatibility filter, the morphological, biochemical, immunological, and physiological interplays between both protagonists can be referred as they permit or impede the development and transmission of the parasite (Combes 2001). Selection will act on these filters to increase or to decrease their permeability and specificity, changing the diameter of the encounter filter or the angle of the compatibility filter (Poulin 2007). Furthermore, the development of human civilization imposes new challenges to our understanding of the host–parasite interplay as factors such as the global effects of urbanization, climate change, or increased pollution heavily influenced environmental dynamics which are known to affect both encounter and compatibility filters, and to ultimately influence infection outcome (Fig. 11.1).

Integrative scheme representing the encounter and compatibility filters and some extrinsic biotic processes and environmental factors all of which primarily and ultimately govern the probability of infection of a Lymnaeidae snail by any given parasite (exemplified by a trematode parasite), and the balance of the interaction and its outcome
Herein, emphasis will be given to compatibility and immunobiological interactions, and to ecological features affecting infection outcome and transmission. Insights into the significance of building up knowledge on the biology of host–parasite–environment interactions will be also discussed to introduce the opportunities and challenges that lies ahead concerning the family Lymnaeidae and their most significant pathogens.
11.2 How Do Parasites Meet Their Lymnaeid Hosts?
Whilst the nature of the compatibility filter is often amenable to experimentation as infective stages of parasites are provided access to potential hosts, direct assessment of the encounter filter is difficult to quantify, particularly in the field where the use of sentinel snails may be required (see Allan et al. 2013, for example). However, no successful parasite infection can occur without a successful contact of the parasite with the host. In this sense, the encounter filter includes the large- and small-scale spatial distribution of the host and parasite (i.e., geography and habitat use), and their behavior (e.g., circadian rhythms/chronobiology) and biology (e.g., for trophically-transmitted parasites, whether the host ever eats the next host of the transmission chain; see Combes 2001).
The family Lymnaeidae is present in all inhabited continents with species occurring in different climatic conditions, from tropical to temperate to artic conditions (Vázquez et al. 2018). Certain species such as Galba cousini and G. truncatula are known to occur at high altitudes (Celi-Erazo et al. 2020; Roldán et al. 2020). Representatives within this family are also commonly found in a wide variety of freshwater environments; from rivers, lakes and streams to ponds, ditches, puddles, and even, on man-made water systems, and they present a tendency to occur above or near the shore and to inhabit ecosystems with slow-moving currents that could be even temporal (Utzinger and Tanner 2000; Kock et al. 2003; Schweizer et al. 2007; Vázquez et al. 2009; Dida et al. 2014). These ecosystems usually present high productivity and biomass content, and a diversity of other species within or associated to them, including parasites (Soldánová et al. 2011; Preston et al. 2013, 2021; Schmid-Araya et al. 2020). In such habitats lymnaeid snails also become important members of the biotic community due to their propensity for feeding on numerous species of algae, and macrophytes, foraging behavior when food is scarce and certain resilience to drought and to playing roles in structuring the plant and macrophyte communities in the environment (Hunter 1980). These ecological features of the family Lymnaeidae should contribute to the establishment of ecological equivalences for a number of parasites. For instance, snail feeding ecology and habitat preference were suggested to be important factors in the infection dynamics of Echinostoma spp. cercariae among the snail second intermediate hosts, with Physa spp. and the lymnaeid Pseudosuccinea columella showing the highest prevalence and intensity of infection, respectively, when compared to compatible sympatric planorbid snails in the field. The pattern of infection displayed by P. columella was related to its extensive vagility while foraging, in order to acquire sustainable food resources, and to an increased probability of autoinfection as they are more likely to harbor Echinostoma spp. as first intermediate hosts. Furthermore, its amphibious tendency was speculated to aid transmission of the parasite to the definitive host as they can locate closer to the surface (Zimmermann et al. 2014).
Besides ecological affinities between host and parasites, both entities can “try” to open or close the encounter filters by modifying their behavior to chase or to avoid each other. Several parasites like water-associated digeneans present free-living stages that seek hosts in the environment primarily moving toward suitable microhabitats based on cues like gravity, temperature, or light intensity when the snail host is not present within their vicinity, all of which have been linked to a selection driven by the ecology of the host and by the probability of encounter (Christensen 1980; Combes 2001). Once in the proximity of a host, molecular sensors that are believed to exist in the parasitic larva facilitate a discriminatory chemotaxis toward the suitable snail through the gradient of biochemical cues segregated by the host (Wilson and Denison 1970a, b). Chemical attraction of Fasciola hepatica and Trichobilharzia szidati [European isolates have been erroneously classified as Trichobilharzia ocellata; see (Rudolfova et al. 2005; Aldhoun et al. 2016)] miracidia by species-specific mucin-type glycoproteins released by Galba truncatula and Lymnaea stagnalis is supposed to influence host finding in these snail–parasite systems (Kalbe et al. 1997, 2000).
There are different degrees of “openness” of encounter (and of compatibility) filters irrespective of the biological level (species, population, individuals; see Combes, 2001), and even known host snails vary their degrees of attractiveness through parasite chemo-orientation behavior at the species or individual levels (Christensen et al. 1976; Langeloh and Seppälä 2018). In this sense, an “open” encounter filter does not determine that the contact will be followed by infection success as physiological, biochemical, and immunological interactions with the host (determined by compatibility filters), as well as environmental factors will ultimately dictate the overall success of the parasite in infecting the snail host (Combes 2001; Langeloh and Seppälä 2018).
11.3 How Does the Immunobiological Interaction Between the Lymnaeid Host and the Parasite Take Place?
-
i.
The immune system of Lymnaeidae snails
Lymnaeid snails, as all invertebrates, rely on an internal defense system (IDS) to protect themselves against pathogens. Even though lymphocytes, antibodies, and adaptive responses to specific antigens are lacking, their defense strategies are far from being simple or non-effective (Loker et al. 2004). Homologs of molecules and immune mechanisms between vertebrate and snail (invertebrate) immune defenses exist although, there are unique immune features that are specifics to their lineage and are the result of a long independent evolutionary process (see Schultz and Adema 2017 for details). In general, three basic components from the snail host’s IDS are included in the multistep infection processes determining the outcome of any lymnaeid–parasite interaction: (1) physical, chemical, and biological barriers (e.g., shell, mucus, microbiota), which impose the first challenge to overcome for any given pathogen, (2) elements to discriminate self from dangerous non-self (e.g., pattern recognition receptors, PRR), and (3) an arsenal of cellular and humoral effectors (e.g., hemocytes, reactive oxygen and nitrogen species (ROS, RNS), opsonins, proteases) and defense mechanisms which are triggered to actively control the injury/infection. Cellular and humoral elements co-operate in the process of self-non-self-recognition as well as in the killing of the non-self, whereas the communication and integration between the different immune pathways allow to orchestrating a multifactorial defense response of the snail host toward pathogens (Pila et al. 2017; Alba et al. 2020). Evidences of specificity and innate immune memory within snail’s immune system also exist (Sire et al. 1998; Portela et al. 2013; Pinaud et al. 2016; Seppälä et al. 2021). Significantly, comparative studies between juveniles and adult lymnaeid snails have brought forward some functional differences of the IDS between both age groups as the former showed aspects related to immunological immaturity, e.g. lower and/or delayed capacity concerning phagocytosis, humoral defenses (namely opsonization and agglutination), and elimination of pathogens (Rondelaud and Barthe 1980a; McReath et al. 1982; Dikkeboom et al. 1985).
Barriers
Mucus itself is an excellent physical barrier to cells, microorganisms, and pathogens; its visco-elastic properties may act as an adhesive providing an initial physical trap created by cross-linked glycoproteins (mucoprotein matrices) and preventing colonization by microorganism and pathogens (Vieira et al. 2004; Allam and Espinosa 2015). In addition to representing an efficient physical barrier, mucus matrices contain various cells and bioactive molecules and have gained prominence in the last few decades as main components of the innate and acquired immune systems (Allam and Espinosa 2015). The biochemical nature of the mucus may have anti-pathogenic activities, whether by the excretion-secretion of host-produced factors or by the exogenous products secreted by the microbiota (Ehara et al. 2002; Guo et al. 2009; Allam and Espinosa 2015). Particularly, the mucus has been pointed as a mediator of snail–digenean compatibility when contact is established. The in vitro effect of snail mucus of two different species on Fascioloides magna miracidia was assayed (Coyne et al. 2015). While no damage was observed when parasite larvae were exposed to the mucus of the lymnaeid species Ladislavella elodes (compatible host), significant tegument damage and/or larval death were recorded in the miracidia when tested against the mucus of an incompatible snail (Helisoma trivolvis) and related to the presence of a cytotoxic protein-like factor (Coyne et al. 2015).
In addition to immunological and enzymatic factors such as PRRs, humoral effectors, heat shock proteins, etc., the diverse mucus matrices in mollusks are also enriched in specialized microbiota that are recruited and maintained by the mucus layer and kept in check by the IDS. However, microbiota is believed to also influence and be influenced by snail–pathogen interactions (Allam and Espinosa 2015) and for instance, differences regarding the diversity and composition of bacterial microbiota exist between resistant and susceptible strains of Biomphalaria glabrata to the trematode Schistosoma mansoni (Allan et al. 2018; Huot et al. 2020). Furthermore, a shift in the composition of B. glabrata microbiota during early infection by S. mansoni (Portet et al. 2021) or by the nematode Angiostrongylus cantonensis (Osorio et al. 2020) has been observed. Changes in the expression of some antimicrobial peptides were recorded in accordance to the changes in the structure of the microbial community in B. glabrata–S. mansoni system (Portet et al. 2021). While this topic will deserve further investigations in lymnaeid snails, evidences exist of modulation of the bactericidal activity from in vitro bacterial clearance assays following infection of L. stagnalis with T. szidati (van der Knaap et al. 1987; Núñez et al. 1994).
Recognition Elements
A prerequisite for the functioning of any internal defense system is the recognition of foreignness and therefore, recognition molecules are pivotal in orchestrating the immune response of the host that follows parasite’s penetration. In molluscs, recognition of dangerous non-self and subsequent immune activation is significantly mediated through lectins and, as other PRRs, they can occur freely in the plasma or on the surface of immune phagocytic cells named hemocytes (Schultz and Adema 2017). These molecules are non-enzymatic, non-antibody proteins that function as PRRs by binding to repetitive carbohydrate surface determinants that characterize groups of pathogens and by activating immune responses (Horák and van der Knaap 1997). When soluble, lectins can function as agglutinins/opsonins; directly neutralizing the target element or creating carbohydrates bridges between the hemocyte receptors and the target element (Horak and Deme 1998).
Little is still known about the molecular arsenal of PRRs of lymnaeid snails and thorough functional and molecular characterizations are surely needed to elucidate the level of diversity within such family of molecules. However, it has been proved that recognition of and defense response against several degrees of foreignness is possible in lymnaeid snails; from abiotic materials to xenografts, septic injuries and parasitic infections, (e.g., Sminia et al. 1974; van der Knaap et al. 1981; Gutiérrez et al. 2003). In this sense, several lectins have been found in L. stagnalis and P. columella and some of their properties and/or binding abilities have been identified (e.g., van der Knaap et al. 1983b; Horak and Deme 1998; Alba et al. 2019a, 2020; Seppälä et al. 2021). The presence of most on the hemocytes of lymnaeid snails has been recognized by means of saccharide-dependent inhibition of phagocytosis/encapsulation, as saturation of lectins with sugars reduces lectin recognition (Horak and Deme 1998; Horák et al. 1998; Alba et al. 2020), whereas evidence of reversed bound to hemocytes of cell-free lectins has been also found (van der Knaap et al. 1983b). Recently, the transcriptome of L. stagnalis under different biotic and abiotic stressors was elucidated and several transcripts within different types of lectins were identified in the reference transcriptome; i.e. galectin, Chi-lectin, L-type lectin, M-type lectin (Seppälä et al. 2021). Specifically, non-self-recognition lectin-like soluble factors belonging to the immunoglobulin superfamily, i.e. fibrinogen-related proteins of the variable immunoglobulin and lectin domain-containing molecules (FREP; a C-terminal fibrinogen-related domain and one or two Ig domains, see Seppälä et al. 2021), and a molluscan defense molecule (MDM; five tandemly arranged Ig domains; see Hoek et al. 1996) have been described in L. stagnalis. In P. columella, the characterization of the differential transcriptome between susceptible and resistant snails to F. hepatica infection resulted in the identification of several lectins; including variants of C-type lectins, different mannose and galactose binding molecules, galectin, and CD109, which were found to be overrepresented in the resistant phenotype (Alba et al. 2019a). Although no specific molecules have been related to P. columella–F. hepatica recognition, the role of lectins in this interaction was brought forward as the in vitro encapsulation of the parasite larvae by the snail’s hemocytes was significantly diminished by adding sugars; with D-mannose exerting the strongest inhibition (Alba et al. 2020).
Furthermore, stimulation of L. stagnalis hemocytes with pathogen-associated molecular patterns such as laminarin (a β-glucan of brown algae), zymosan (β-glucan of fungi), and bacteria lipopolysaccharide (LPS) leads to the activation of the immune cells (Plows et al. 2004; Lacchini et al. 2006; Wright et al. 2006) suggesting the presence of specific β-1, 3-glucan-binding proteins (BGBP) and LPS receptors on the surface of the immune cells that has yet to be characterized. Furthermore, several transcripts of gram-negative binding protein have been identified in the reference transcriptome of L. stagnalis (Seppälä et al. 2021) and proteins containing peptidoglycan binding domains and Toll-like receptors have been found in both L. stagnalis and P. columella snails (Alba et al. 2019a; Seppälä et al. 2021).
Cellular Effectors
At least four types of cells participate in snail defenses; three are “fixed” cells that have been poorly studied concerning snail–pathogen interactions; i.e. antigen-trapping endothelial cells, reticulum cells, and pore cells, whereas the fourth type comprises macrophage-like cells that dwell in the tissues or circulate with the blood fluid (hemolymph) of mollusks and are called hemocytes or amebocytes (van der Knaap and Loker 1990). Hemocytes move freely to and from the tissues continuously sensing the internal environment, and prompt to cellular immune responses and cytotoxic reactions involving lysosomal enzymes and the production of ROS and RNS when stimulated by immune elicitors (van der Knaap and Loker 1990; Adema et al. 1991; Lacchini et al. 2006; Wright et al. 2006; Zelck et al. 2007). These cells originate from connective tissues or specialized organs (e.g., the amebocyte producing organ (APO) in some gastropods, or the white body organ in cephalopods) and their continuous and regulated hematopoiesis is important not only for immune functions, but also for the survival of the animal as a whole as it replenish the number of circulating hemocytes that are depleted from defense reactions, migration to tissues or cellular turn-over or senescence (Pila et al. 2016b).
The localization and structure of the APO varies among snail species and some lymnaeid snails may exhibit a diffuse hematopoietic site as it is believed to be the case in L. stagnalis; the hemocytes are possibly formed throughout the connective tissue and the vascular system of this snail species (Sminia 1974). Contrastingly, a lymphoid organ located beneath the mantle, between the pericardial cavity and the saccular kidney, and presenting a central lumen that is continuous with the kidney lumen has been described in G. truncatula (Rondelaud and Barthe 1981, 1982). In this species, the APO involves the proximal part of the kidney sac and presents a dual structure: the main part, containing an epithelial layer of podocytes that separates the broad hemal spaces adapted for hemolymph filtration from the urinary space, and the apical portion, adhering to the mantle and pericardial epithelia, and consisting in thicker connective-tissue frame in which hemocyte nodules may develop (Monteil and Matricon-Gondran 1991a). An ill-defined tissue located in the region of the lung roof close to the heart of Ladislavella catascopium contained massive numbers of hemocytes but whether it is the site for hemocyte production remains to be clearly demonstrated (Loker 1979).
During infection, the APO can increase its size compared to non-infected snails (Rondelaud and Barthe 1981). However, in lymnaeid snails, proliferation of hemocytes can occur outside of the hematopoietic organ such as in the connective tissue (Sminia 1974), in various scattered nodular regions (Monteil and Matricon-Gondran 1991a), and even in the hemolymph (Sminia 1974; Alba et al. 2020).
Another feature of hemocytes within Mollusca is their biochemical, functional, and morphological heterogeneity, whilst the variety of approaches used for their characterization (as there is a lack of specific universal molecular markers) has resulted in a non-unified classification system (Pila et al. 2016b). From a rough morphological perspective, based on their spreading capabilities, two main types of hemocytes can be defined: cells that typically spread, forming filopodia or lamellipodia in the process, are believed to be the mature effectors in phagocytosis and encapsulation processes and constitute the majority (if not all) of hemocyte populations, and smaller cells that do not spread or spread minimally when placed on artificial surfaces and present high nucleus/cytoplasm ratio, which are often called round cells or blast-like cells (Loker 2010). Spreading cells have been further segregated, mainly based on their granular content, into two groups: (i) granulocytes, which present conspicuous granules located mainly in the endoplasm and produce extensive pseudopodia in all directions, and the (ii) hyalinocytes which are smaller in size, agranular or less granular compared to the granulocytes, generally spherical or slightly oval with lobose and shorted pseudopodia (Pila et al. 2016b). Granulocytes are believed to be more active (e.g., phagocytic and enzymatic activities) than hyalinocytes (Mahilini and Rajendran 2008; Travers et al. 2008). Two hypothesis exist behind the morphological and functional heterogeneity of hemocytes; whereas some researchers consider that the different morphotypes represent different lineages of the cells, others state that they reflect different developmental stages of a single cell type (Pila et al. 2016b).
In G. truncatula, the ultrastructure of circulating hemocytes revealed a preponderance of spreading cells at different stages of differentiation, whereas a few round cells were also observed and suggested as a distinct population. Endogenous peroxidase activity was localized in secretory granules as well as in multivesicular bodies of the spreading hemocytes (Monteil and Matricon-Gondran 1993).
Fluorescence staining of cytoplasm and nucleus, and the estimation of size/cytoplasmic complexity by flow cytometry allowed the description of two main morphotypes in P. columella: (i) large adherent cells and (ii) non-adherent small, roundly-shaped hemocytes with low cytoplasmic complexity and high nucleus-to-cytoplasm ratio; i.e. blast-like cells (Alba et al. 2020). In P. columella, the blast-like cells were related to proliferation reactions in the hemolymph, particularly in resistant snails exposed to F. hepatica (Alba et al. 2020), endorsing the hypothesis of the pluripotency of this morphotype related with certain stem-cell capacity (Wang et al. 2012; Rebelo et al. 2013).
In S. palustris, three populations of circulating hemocytes, morphologically and cytochemically distinct, were observed; (i) round non-adherent cells showing a central spherical nucleus and a low nucleus-to-cytoplasm ratio, (ii) adherent irregular hyalinocytes showing an eccentric kidney-shaped nucleus surrounded by a thin ring of cytoplasm, a light basophilic cytoplasm containing small vacuoles and few basophilic, rarely neutrophilic, granules, and (iii) large granulocytes producing long, filamentous pseudopodia and characterized by an acidophilic cytoplasm containing many acidophilic granules (Russo and Lagadic 2000).
In L. stagnalis, size differentiation following a density gradient centrifugation segregated the hemocytes in: small cells (3.6–4.8 μm; 4.4 ± 0.6) with high nucleus/cytoplasm ratio, and two types of spreading cells; medium-size cells (6–12 μm; 9.04 ± 2.00) displaying some pseudopodia, and large hemocytes (13–17 μm; 14.22 ± 1.00). Although it is worth mentioning, that no complete separation among cellular fractions was achieved. Moreover, cytochemical assays for lysosomal enzymes revealed α-naphthyl acetate esterase activity in all three size groups, whereas acid phosphatase activity was weak or absent in most of the large-size cells (Adema et al. 1994a). The previous size/appearance based-segregation was more recently observed by (Boisseaux et al. 2016) in side-scattered signal outputs as three clusters, differing in size and granulometry, were also resolved; a small cells cluster (clearly segregated from the others), and intermediate and large cells clusters that sometimes overlapped. However, differences on intermediate and large cells clusters observed in the cytograms were believed to be caused by the variety of nucleus lobulation (hemocytes with one up to four nuclear lobules were observed) rather than to a cytoplasm granulation gradient as light microscopy made difficult to discriminate granulocytes from hyalinocytes (Boisseaux et al. 2016). Further ultrastructural analysis showed only one cell population, evidences of high cellular activity and the presence of two interesting organelles, so far, only described from L. stagnalis; (i) a large organelle with a spectacular fibrillary arrangement called lysosomal-like structures and (ii) a massive structure, usually located within the perinuclear regions, with a dense aspect and vesicles sprouting out from the internal nuclear membrane was also depicted filling intracellular compartments and occupying a vast cytosolic space in the vast majority of hemocytes (Boisseaux et al. 2016). The hypothesis that hemocytes from L. stagnalis originate from a single cell lineage that mature progressively as a differentiation/maturation continuum reflected in the cytograms clusters and in the small differences observed in microscopy, is strongly suggested (Boisseaux et al. 2016).
Phagocytosis (Plows et al. 2006b), encapsulation (Loker et al. 1982), and production of cytotoxic molecules (Lacchini et al. 2006; Wright et al. 2006) involved in pathogen killing and elimination are among the significant roles of hemocytes in mollusks. However, they also participate in other vital processes such as wound healing (Franchini and Ottaviani 2000) and nerve repair (Hermann et al. 2005), as well as in the production of a plethora of molecules involved in signal transduction, defense regulation, or effector capacities (Baeza-Garcia et al. 2010; Galinier et al. 2013; Pila et al. 2016a). All these functions ensure the homeostasis of the organism and require the activation of the cells through different signaling pathways and regulatory molecules, many of which occurs in concert with humoral factors.
Signaling/Regulatory Molecules and Pathways, and Defense Reactions
Hemocytes can phagocytize small particles and cells, whereas encapsulation is reserved for larger elements or organism (such as metazoan parasites like digenean larvae). Both processes involve recognition and adherence to the surface of non-self-particles through cellular receptors and, in the case of encapsulation, recognition and adherence must be extended to other hemocytes that must significantly spread to form layers of overlapping cells that will eventually surround and isolate the element (Loker et al. 1982). The involvement of the mitogen-activated protein kinase (MAPK) pathways has been particularly described in several snail species in association to adhesion, motility and spreading cellular events required for phagocytosis, encapsulation, and regulation of cytotoxic molecules/reactions (e.g., Humphries and Yoshino 2003, 2008; Iakovleva et al. 2006). In L. stagnalis, the pivotal role of the MAPK and phosphatidylinositol 3-kinase (PI3-K) signal cascades in the immune responses of the snail’s hemocytes was elucidated using different specific inhibitors (Zelck et al. 2007). To note, the extracellular signal-regulated kinase (ERK) inhibitor, PD098059, substantially impaired hemocyte spreading and aggregation, phagocytosis, encapsulation and H2O2 production, making patent the involvement of the ERK cascade in all phases of the cellular processes. The MEK/ERK inhibitor showed the strongest effect when added before hemocyte attachment, supporting the significant role for this signaling pathway in the initial processes of cell migration and cell defense responses (Zelck et al. 2007).
Upstream events involved in the defense activity of L. stagnalis hemocytes have been related to the activation of cell-surface bound αvβ3 and β1-like integrins mediating Ca2+/Mg2+-dependent cell adhesion; their inhibition with blocking peptides resulted in a significant diminished of the spreading capacity of the cells and in a significant inhibition of the phagocytosis of Escherichia coli bioparticles. It has been proposed that integrin engagement promotes outside-in signaling events in L. stagnalis hemocytes (Plows et al. 2006a; Walker et al. 2010) as it mediates association of protein kinase C (PKC) with focal adhesion kinase (FAK)/tyrosine kinase Src complexes to promote focal adhesion assembly, and to possibly modulate hemocyte early adhesion to pathogens (Walker et al. 2010). This model has been endorsed by the findings that phosphorylated (activated) PKC colocalized with focal adhesion sites (where a FAK-like protein was also found phosphorylated; (Plows et al. 2006a)), whereas specific inhibitors of PKC and of Src (which phosphorylates and associates with FAK) attenuated hemocyte spreading (Walker et al. 2010). Moreover, phosphatidyl inositol-3 phosphate kinase (PI3-K) and ERK have been also pointed out to strongly regulate cell motility and spreading of L. stagnalis hemocytes as the addition, prior to hemocyte attachment, of the MEK/ERK inhibitor, PD98059, or the PI3-K inhibitor, wortmannin, severely suppressed hemocyte spreading and aggregate formation in a concentration-dependent manner. No evidences of a significant participation of c-Jun N-terminal kinase (JNK) inhibitor, SP600125, and the p38 kinase inhibitor, SB203580, on hemocyte spreading were observed (Zelck et al. 2007).
A variety of targets such as zymosan (Dikkeboom et al. 1987), gram-positive and gram-negative bacteria (Dikkeboom et al. 1987; Plows et al. 2006b), red blood cells (Zelck et al. 2007), or latex particles (Dikkeboom et al. 1987; Russo and Lagadic 2000) have been used to elicit phagocytosis by lymnaeid snails’ hemocytes. In vitro phagocytosis assays of E. coli by L. stagnalis hemocytes in the presence of inhibitors of PI3-K and of protein kinase A (PKA) demonstrated that this reaction is dependent on the activation of the first enzyme (Plows et al. 2006b). In another study, the stimulation of L. stagnalis hemocytes with bacterial LPS resulted in a transient activation of ERK, whereas inhibition of MEK activity impaired phosphorylation of ERK, its colocalization in the nuclear and perinuclear regions after LPS challenge, as well as the phagocytosis of E. coli bioparticles (Plows et al. 2004). Furthermore, addition of GF109203X, a PKC inhibitor, or of a FTase inhibitor I, which blocks farnesylation of the upstream MAPK component Ras, resulted in a reduced phagocytosis suggesting that activation of ERK can be driven by both Ras-dependent (with MEK as upstream kinase of ERK) and Ras-independent mechanisms (which may involve PKC, PKA or PI-3-kinase; Plows et al. 2004). ERK, and JNK, and p38 Map kinase in a lesser extent, were also found to contribute in signal transduction mediating the phagocytosis of sheep red blood cells by L. stagnalis hemocytes, indicating that regulation of this process may depend on the activation of and the cross-talk among all three MAPK pathways. Moreover, trematode encapsulation by L. stagnalis was also impaired by inhibitors for MEK/ERK and PI3-K (Zelck et al. 2007).
Activated hemocytes trigger cytotoxic reactions toward phagocytized or encapsulated elements. Contact between the immune elicitor and the hemocyte’s surface stimulates a membrane-bound system to generate and to release oxygen radicals that can be sensed in the extracellular milieu (Adema et al. 1991). The encapsulation reaction allows the hemocytes to isolate the menace and to neutralize it within the formed capsule without seriously endanger the integrity of the surrounded tissues (Loker et al. 1982; McReath et al. 1982). When internalization of the immune elicitor occurs through phagocytosis, the production of oxygen radicals localized inside the phagolysosome (Dikkeboom et al. 1987; Adema et al. 1991).
The mechanism for production of toxic ROS in L. stagnalis (as well as in other mollusk species) has been proposed to be homologous to that of the respiratory burst of vertebrate phagocytes as both are sensitive to catechol-like phenol inhibitors of the vertebrate NADPH-oxidase enzyme complex that generates superoxide (Adema et al. 1993; Lacchini et al. 2006). It is postulated that, in L. stagnalis hemocytes, an active NADPH-oxidase enzyme complex has to be assembled from putative cytosolic and membrane-associated components (Adema et al. 1993). At the molecular level, ERK appears to play a major role in the activation of the oxidative burst of L. stagnalis hemocytes following phagocytosis or encapsulation reactions (Zelck et al. 2007). Challenge of freshly collected hemocytes with 1,3-glucan laminarin promoted PKC-mediated signaling and downstream MEK/ERK activation and H2O2 release (Lacchini et al. 2006). Moreover, ERK1/2 and PKC (to a lesser extend) signaling seems to play a pivotal role in mediating nitric oxide synthase (NOS) activation and the subsequent NO production following stimulation of L. stagnalis hemocytes by laminarin and the phorbol ester, phorbol myristate acetate (Wright et al. 2006). However, as with phagocytosis, observations that PD098059, a specific inhibitor of ERK/MEK, did not completely inhibit H2O2 generation suggest that multiple signaling cascades, possibly other MAPK pathways, may contribute to the orchestration of cellular defense responses in lymnaeid snails (Zelck et al. 2007).
Another significant pathway mediating immune reactions in different phyla is the Toll-like receptor (TLR) pathway. In mollusks, TLRs can be activated and their expression can be upregulated after septic injury to promote hemocyte activation (Nie et al. 2018), whereas the positive influence of TLRs and their activation in the protective response toward pathogens have been experimentally stated in B. glabrata–S. mansoni system (see Pila et al. 2016c). One Toll-like receptor (TLR) with transmembrane domain and several transcripts belonging to downstream mediators within the TLR pathway (e.g., MyD88, IkB, and NF-kBp65) have been recorded in the reference transcriptome of L. stagnalis. It was also observed that IκB, a regulator of TLR signaling, showed increased transcription to different immune elicitors (gram-negative and gram-positive bacteria, trematode, injury) except wounding and even though this effect was not seen in all individuals, it suggests that the TLR pathway generally participates in the control of the defense responses in L. stagnalis (Seppälä et al. 2021). In P. columella, PKC, leucine-rich repeat-containing molecules, and several Toll-like receptors (TLR) and some of their downstream molecules, were found to be overrepresented in F. hepatica-resistant snails (Alba et al. 2019a).
Other transcripts involved in regulation/signaling such as granulocyte colony stimulatory factor receptor (G-CSFR), transforming growth factor 1-beta (TGF1β), members of the superfamily of tumoral necrosis factor (TNF) receptors and interferon regulatory factors (IRF) have been recorded in P. columella transcriptome (Alba et al. 2019a). Similarly, several variants of the cytokine macrophage migration inhibitory factor (MIF) have been found in L. stagnalis and P. columella (Alba et al. 2019a; Seppälä et al. 2021), whereas three variants of the TNF and 23 variants of interleukin 17 families were depicted in the reference transcriptome of L. stagnalis (Seppälä et al. 2021). Furthermore, expression of granulin, a cytokine with pleiotropic functions in mammals (Jian et al. 2013) that also promotes proliferation, differentiation, and activation of B. glabrata hemocytes in response to S. mansoni (Pila et al. 2016a; Hambrook et al. 2019), was specifically studied in P. columella by qRT-PCR following exposure to F. hepatica (Alba et al. 2020). In this study, the granulin gene was found to be significantly upregulated in resistant P. columella snails during infection in consistency with the increase of hemocyte proliferation and spreading that was also associated to this host phenotype (Alba et al. 2020).
Apart from the mentioned cellular reactions, the phenoloxidase (PO) system is another immune trait with a major role in invertebrates and their defenses against pathogens and damaged tissues that eventually leads to melanization (Cerenius and Söderhäll 2004). In L. stagnalis, PO-like activity, measured as an increase in optical density of a solution in which PO enzymes from snail hemolymph oxidize the substrate L-dopa, has been assessed after experimental challenges with different immune elicitors or environmental conditions (e.g., Seppälä and Jokela 2010; Seppälä and Leicht 2013). Although, this assay may also measure the combined activity of different factors (see Le Clec’h et al. 2016), the finding of several variants of laccase and tyrosinase, enzymes involved in phenoloxidase/melanization type-reactions, in the reference transcriptome of L. stagnalis warrants further investigations about their significance in protective immune defenses (Seppälä et al. 2021).
In addition, hemocytes of the lymnaeid snails L. stagnalis and Radix lagotis, and of the planorbid Planorbarius corneus have recently been shown to generate extracellular trap-like (ET-like) fibers in vitro, although their induction following stimulation of hemocytes with immune elicitors was rare (Skála et al. 2018). This mechanism was firstly described in mammalian neutrophils, which generate extracellular chromatin fibers that facilitates immobilization and elimination of unicellular pathogens (Kaplan and Radic 2012). In terrestrial slugs and snails, ET-like fibers were observed to be extruded from hemocytes following exposure to metastrongyloid parasites and to participate in the contact, entrapment, and immobilization of the larva (Lange et al. 2017). Further characterization of this phenomenon in the family Lymnaeidae is needed to elucidate its significance in the IDS.
Humoral Effectors and Antioxidant System
The role of plasma (cell-free hemolymph) in digenean–snail compatibility has proved crucial since significant damage to sporocyst (digenean intramolluscan larva) tegument and diminishing of its survival can be seen after parasite’s exposure to plasma from incompatible hosts. In 9 of 10 combinations tested, lymnaeid plasma was toxic to the parasites of planorbid snails and in 2 of 4 combinations, planorbid plasma was toxic to the parasites of lymnaeid snails. The effect was not associated to the physic-chemical conditions of the plasma and thus, the existence of particular humoral factors dictating patterns of snail specificity was speculated (Sapp and Loker 2000).
Further studies have described the presence of humoral factors with potential defense activity against infection in lymnaeid snails, such as different variants within families of antimicrobial peptides, i.e. hydramacin in P. columella (Alba et al. 2019a) and macin in L. stagnalis (Seppälä et al. 2021). Soluble lectins in the plasma can function as opsonins or agglutinins neutralizing pathogens, or promoting recognition and effector capacity of cellular defenses (Sminia et al. 1979; van der Knaap et al. 1981, 1983b; Horak and Deme 1998). Moreover, an opsonin named granularin, a cysteine-rich protein that shares amino acid sequence identity with domains present in Von Willebrand factor and thrombospondin, has been described in L. stagnalis to stimulate phagocytosis of foreign particles by hemocytes (Smit et al. 2004). Several proteins with potential activity in antimicrobial defenses were identified in the reference transcriptome of L. stagnalis; i.e. lipopolysaccharide-binding protein/bactericidal permeability-increasing proteins (LBP/BPI), lysozymes and L-amino acid oxidases. In addition, different transcripts encoding for cytolytic β pore-forming toxins, called as Lymnaea-lysin and stagnalysin, were also found (Seppälä et al. 2021). In P. columella, a G-type lysozyme and a LBP/BPI were only observed in the differential proteome of resistant P. columella to F. hepatica (Alba et al. 2019a). It is worth mentioning that, whereas hemocytes are believed to play a crucial role in the synthesis and secretion of humoral factors (Baeza-Garcia et al. 2010; Galinier et al. 2013), other cellular types and organs have been proved to be involved in this activity in lymnaeid snails, such as the granular cells (Hoek et al. 1996; Smit et al. 2004) and the albumen gland (Alba et al. 2019a).
Enzymes and proteins with roles in regulating the oxidative damage, i.e. superoxide dismutases (Cu/Zn and Fe/Mn families; SOD), glutathione peroxidases, glutathione reductase, peroxiredoxins, glutathione S-transferases (GST) and catalase, were represented in a transcriptomic study on L. stagnalis focalized on responses to environmental stress (Bouétard et al. 2012). In naïve resistant P. columella, transcripts identified as catalase, superoxide dismutase and probable deferrochelatase peroxidase have been identified as overrepresented, as well as molecules involved in acute phase response like ferritin and NOS (Alba et al. 2019a). Such antioxidant potential is essential to maintain the redox balance of the host prior and during infection and has proved significant to resist bacterial infection in the clam Meretrix meretrix (Wang et al. 2013) or to be positively associated to anti-parasite resistance in B. glabrata snails as higher constitutive levels of Cu/Zn SOD make killing of the trematode larva more effective (Goodall et al. 2004; Lockyer et al. 2012).
To summarize the main content of the present section, a representation of some of the elements, signaling pathways, and immune reactions described to be involved in the IDS of Lymnaeidae is depicted in Fig. 11.2.

Overview on Lymnaea internal defense system. Snail defense system could be separated into 4 steps, (1) physical, chemical, and biological barriers dedicated to block pathogens and avoid their penetration in snail tissues; (2) immune recognition to discriminate self from non-self based on Pattern Recognition Receptors (PRR) able to recognize Pathogen Associated Molecular Patterns (PAMPs) or Danger Associated Molecular Patterns (DAMPs) associated with pathogen-dependent wounding or tissue lysis; (3) signaling pathways regulate immune and stress response (cytokine, TLR, MAP kinase, …), and (4) an arsenal of cellular and humoral effectors (e.g., hemocytes, reactive oxygen and nitrogen species (ROS, RNS), opsonins, proteases) are produced as humoral hemolymphatic factors or degranulated inside hemocytic capsule (encapsulation process) to kill and eliminate pathogens. Endosymbiotic microbiota composition changed during early infection, microbiota is believed to influence the internal defense system of the host but also to be influenced by snail–pathogen interactions, this resulting in changes in host fitness and defense capabilities. Abbreviations: ROS: reactive oxygen species, RNS: reactive nitrogen species, AMP: antimicrobial peptide, NOS: nitric oxide synthase, SOD: super oxide dismutase, MOP: myeloperoxidase, MIF: macrophage migration inhibitory factor, PKC: protein kinase C, MAPK: mitogen-activated protein kinase, ERK: extracellular-signaling regulated kinases, MEK: mitogen-activated protein kinase kinase, PI3K: phosphatidylinositol kinase, NF-kB: nuclear factor-kappa B, Rap1: Ras family GTPase, Ras: Rat sarcoma virus small GTPase, FAK: focal adhesion kinase, Src: steroid receptor coactivator, RAM: regulation of Ace2p activity and cellular morphogenesis, Akt: protein kinase B
-
ii.
The interplay between the lymnaeid host and the parasite: defenses vs. counter defenses
The penetration of a pathogen into the lymnaeid snail and its colonization stimulate the IDS of the host through the recognition of the somatic components or the excretion-secretion products (ESP) of the parasite (Núñez et al. 1994; Skála et al. 2014). However, whether these events will result in the destruction of the pathogen or in the dynamic equilibrium that characterized an established infection will depend on the interplay between host’s defenses and pathogen’s counter defenses at each step of the interaction (van der Knaap and Loker 1990). The interplay is, therefore, dependent on (epi)genetic factors that control the infectivity and pathogenicity of the parasite and the level of resistance of the host (Mitta et al. 2017).
Destruction of the Pathogen
The sensing of a trematode occurs early in the infection and it may lead to the activation of the IDS of the snail host as shown by the enhanced in vitro phagocytosis of zymosan particles or live bacteria by L. stagnalis hemocytes between 1.5 and 3 h post-exposure (p.e.) to T. szidati (van der Knaap et al. 1987; Núñez et al. 1994). It can be featured by the recruitment of hemocytes to the site of infection as it was proposed when a decrease of hemocytes counts in the hemolymph of P. columella was observed as early as 30 minutes p.e. to F. hepatica (Ribeiro et al. 2017). In R. lagotis infected by Trichobilharzia regenti, histological sections showed that circulating hemocytes migrate and accumulate nearby the developing parasite between 2 and 36 h p.e. (Skála et al. 2014). In S. palustris, hemocytes gathered around F. hepatica sporocyst between 24 and 72 h p.e., although the length of the mobilization period and of the defense reactions decreases as the age of the snail increases (McReath et al. 1982).
An increase of circulating hemocytes (Ribeiro et al. 2017; Alba et al. 2020), changes in the dynamic of the different morphotypes (Russo and Lagadic 2000) and even qualitative alterations of their morphology and functionality can also result from the contact with trematodes (Alba et al. 2020). In resistant P. columella challenged with F. hepatica, an increase of the proliferative activity of blast-like cells and of the spreading of large hemocytes, which tended to aggregate into a sheet-like spreading pattern and to display an increased length of broad-based cytoplasmic projections, was observed at 24 h p.e. (Alba et al. 2020). In G. truncatula, the APO is significantly enlarged upon exposure to F. hepatica and showed an increase of hemocyte proliferation and density during early infection compared to uninfected snails (Rondelaud and Barthe 1981).
The regulation of the expression of several immune molecules also occurs following IDS activation, which results in mounting and controlling the immune response during all the infection process. In resistant P. columella challenged with F. hepatica, the transcription of the cytokine granulin is significantly upregulated at 6 h p.e. compared to naïve snails, whilst at 96 h p.e. a higher increase was recorded in exposed snails irrespective of the phenotype (Alba et al. 2020). In L. stagnalis snails infected with T. szidati, the gene encoding the opsonin granularin is upregulated from 1.5 h p.e. onwards (Smit et al. 2004). Upregulation of laccase transcription following injection of soluble extracts from trematode infected snails has been also reported in L. stagnalis (Seppälä et al. 2021).
All the reactions elicited in the snail by the infective trematodes might eventually lead to an increase of the phagocytosis/encapsulation activity specifically directed toward the parasite and to the activation of the respiratory burst. Highly antigenic miracidial ciliated plates shed by the parasite upon penetration have been observed within phagosomes of hemocytes of R. lagotis after infection with T. regenti (Skála et al. 2014). On the other hand, stimulation of L. stagnalis with zymosan or trematode larvae triggered the generation of elevated amounts of H2O2 in the snail hemocytes 2 to 48 h p.e.; maximal inductions were observed at 45 min during phagocytosis and at 12 h during encapsulation reactions (Zelck et al. 2005). The respiratory burst and the concomitant production of ROS and RNS should follow the proper activation of hemocytes (Adema et al. 1991; Lacchini et al. 2006; Zelck et al. 2007) and the cytotoxic effect exerted by reactive oxygen intermediates like superoxide (O2−) and hydrogen peroxide (H2O2) has been proved effective against schistosomes (Adema et al. 1994b).
In particular, the encapsulation and further killing of the encapsulated parasite are the effective snail defense that is generally associated to protection against trematodes (Loker 1979; McReath et al. 1982; Gutiérrez et al. 2003). The velocity and effectiveness of mounting such a response significantly increases with the age of the snails; in 6 to 9 days old S. palustris, encapsulation of 50% of F. hepatica sporocysts was observed at 96 h p.e and the infection was resolved within 7 days p.e., whereas in 18–21 days old snails, 50% of parasite larva were already encapsulated within the first 24 h and total clearance was attained at day 5 p.e. (McReath et al. 1982).
On the other hand, even though most studies have encompassed the immune response toward trematodes, defense reactions against other types of pathogens have been explored in lymnaeid snails through experimental challenges using model bacteria species. As with digenean trematodes, evidences of certain specificity of the immune response can be observed as distinctive molecular patterns have emerged following exposure. In this sense, experimental challenge of L. stagnalis with the gram-negative E. coli bacteria results in the differential expression of the TLR and of several molecules of the TLR signaling pathway (IκB, NF-κB) as well as of the antibacterial cytotoxins Lymnaea-lysins. In contrast, exposure to the gram-positive bacteria Micrococcus lysodeikticus leads to the overexpression of the lectin FREP and the cytokine IL-17. The enzyme laccase and the component of the TLR pathway IκB were found overexpressed 6 h after both bacterial challenges (Seppälä et al. 2021). At the phenotypic level, injection of M. lysodeikticus significantly increases antibacterial activity of the hemolymph of L. stagnalis at 6 h p.e. (Seppälä and Leicht 2013). Exposure to Staphylococcus saprophyticus or E. coli results in early phagocytosis of the bacteria by hemocytes with the number of fusions of lysosomes and phagosomes as well as of the fusions among phagosomes increasing from 30 min p.e. onwards, and 99% of bacterial clearance attained as early as 2 h p.e. (van der Knaap et al. 1981).
Nevertheless, all the previous immunobiological events will result in the protective response only when resistant or incompatible snail–pathogen combinations take place (Núñez and De Jong-Brink 1997; Gutiérrez et al. 2003; Alba et al. 2020). In P. columella, two contrasting phenotypes concerning infection by the trematode F. hepatica exist; naturally resistant and susceptible snail populations (Gutiérrez et al. 2003; Alba et al. 2018, 2019a, b). Although defense reactivity seems to be constitutively enhanced in naïve resistant snails compared to susceptible ones (Alba et al. 2019a, 2020), the experimental challenge with F. hepatica ends up boosting the encapsulation capacity of the hemocytes toward the sporocysts particularly in resistant snails, both in vitro (Alba et al. 2020) and in vivo (Gutiérrez et al. 2003). Apparently, the defense response elicited by F. hepatica in resistant P. columella is potent enough to eliminate the infection regardless of the infective dose, infection scheme, or the geographic origin of the parasite (Calienes et al. 2004; Vázquez et al. 2014; Alba et al. 2018). However, when analyzing susceptible P. columella and other compatible lymnaeid snails high among-individual variation in terms of the immune response toward trematodes has been observed at the molecular (Seppälä et al. 2021) and phenotypical levels (Vázquez et al. 2014; Alba et al. 2020). Even within a single snail host individually infected by multiple miracidia, evidences of effective encapsulation of certain larva whilst other remain free to develop and multiply attest for this mosaic of phenotypic responses to infection within susceptible (= compatible) hosts (e.g., (Loker 1979; Thèron and Coustau 2005; Coustau et al. 2015). This is the result of a polymorphism of compatibility, which is driven by genotype/genotype interaction from each single host–parasite combination (Thèron and Coustau 2005).
Dynamic Equilibrium of a Successful Infection
In snail–trematode systems, although all intramolluscan developmental stages are continuously confronted with a potentially lethal host response, compatible parasites are left unharmed as the countermeasures displayed by them are effective enough as to impair the orchestration of a protective response and to succeed in guarantying the infection and development within the host. Two main strategies; evasion of immunorecognition, and selective and direct interference of the antiparasitic activities elicited upon infection, are used by compatible parasites to overcome host defenses, and can act independently or concomitantly depending on the host–parasite system (van der Knaap and Loker 1990).
The avoidance of recognition by the host’s IDS can be achieved through antigenic variation or molecular disguise (i.e., molecular masking and molecular mimicry; see van der Knaap and Loker 1990). The miracidium-to-sporocyst (-to rediae) transformation of digenean trematodes involves considerable changes in the molecular composition of the larvae, including the main targets for non-self-recognition via host’s lectins, i.e. the carbohydrate moieties, as demonstrated on the surface of T. szidati and F. hepatica by the use of a variety of lectin probes (Gerhardus et al. 1991; Horák 1995; Georgieva et al. 2016). Moreover, the intramolluscan stages of T. szidati (miracidium, mother sporocyst, and cercaria) contain a polypeptide exhibiting N-acetyl-galactosaminyl-transferase activity that controls the synthesis of N’N-diacetyllactosediamino LacdiNAc-type glycans that are also common to L. stagnalis and might, therefore, contribute to molecular mimicry (Neeleman et al. 1994). In F. hepatica–P. columella/Galba cubensis, cross-reactivity of anti-rediae polyclonal antibodies raised in mice and rabbits with antigens of naïve snail is suggestive of the existence of some immunogenic determinants of the parasite similar to those of the snail hosts, mainly associated with low molecular weight proteins that appear to be parasite-specific (Alba et al. 2014). In addition, it has been observed that some glycan domains are shared between F. hepatica and some of the tissues of G. truncatula where the parasite establishes itself (Georgieva et al. 2016). Ultrastructural observations of Haplometra cylindracea sporocysts revealed that G. truncatula hemocytes, which are modified upon contact with the parasite, end up covering the sporocyst in a structure called “paletot,” an intermediate state between typical free hemocytes and paletot cells derived from the transformation of the formers upon contact with the parasite larva. This results in hemocytes becoming fixed cells, losing most pseudopodia, endogenous peroxidase activity and secretory granules, acquiring a basal lamina and presenting a cytoplasm gradually invaded by glycogen and lipid inclusions and in a final structure that fails to encapsulate the larva but could aid in interfering with IDS recognition and/or defense while providing nutrition for the larvae (Monteil and Matricon-Gondran 1991b).
Direct interference from the parasite with defense activities of the host can be also cited from Lymnaeidae–trematode interactions. In compatible L. stagnalis–T. szidati combinations, snail hemocytes displayed an initial enhance of phagocytic activity that is followed by a reduced capacity to eliminate foreign particles from 12 to 96 h p.e. (e.g., Núñez et al. 1994). In this sense, glycoproteins present in the high molecular weight fraction of T. szidati ESPs and produced between 33 and 72 h of culture inhibited the phagocytosis of bacteria suggesting an immunosuppressive role (Núñez and De Jong-Brink 1997). Exposure to Plagiorchis mutationis causes a severe immunosuppression of the cellular immune response in L. stagnalis. In this study, hemocytes of both uninfected and infected snails did not form a capsule around live cercariae, whereas all dead P. mutationis larva were encapsulated by hemocytes in the uninfected snails but only 20% of all dead cercariae were completely surrounded in vitro by hemocytes from infected snails (Kryukova et al. 2014). Similarly, only few hemocytes of L. catascopium occasionally adhere to the body wall of compatible Schistosomatium douthitti mother sporocyst that typically neither flatten nor accumulate in any numbers and thus, no effective host responses are observed (Loker 1979). On the other hand, the early recruitment of hemocytes of R. lagotis infected by T. regenti does not result in the destruction of the parasite. From 20 to 36 h p.e., hemocytes did not accumulate in layers and appear scattered around the parasite without contacting parasite’s tegument, whilst at 44–49 h p.e. no hemocytes were further observed in proximity to the sporocysts. In the same study, phagocytic activity and H2O2 production of these cells were found diminished during the patent period of infection compared to those from non-infected snails, showing also lower levels of phosphorylation of PKC and of ERK (Skála et al. 2014). In G. truncatula infected by F. hepatica, interference with hemocyte functions was suggested as in the few capsules that occurred in parasitized snails the hemocytes did not spread normally and their peroxidase granules were resorbed into multivesicular bodies (Monteil and Matricon-Gondran 1993). Evidences of a putative F. hepatica driven inhibition of host immunity in susceptible P. columella snails were recorded by Alba et al. (2020). No significant increase of total hemocyte counts, no patent proliferation of blast-like cells, and a shrinkage of large adherent cells which appeared more scattered, without a clear tendency to aggregate and showing a discrete spreading morphology with shorter pseudopodia when compared to naïve conditions, were recorded following experimental infection by the parasite. In addition, a significant decrease of production of the cytokine granulin at 12 and 24 h p.e. compared with the constitutive level in uninfected snails also features the dynamic of the interplay in F. hepatica-susceptible P. columella (Alba et al. 2020).
Even though compatibility filters are early gauged, countermeasures and immune regulation associated with parasite development are also present during mid and late-infections in compatible host–parasite systems. An increase of the density of G. truncatula hemocytes at the sites where F. hepatica occurs is significantly patent at day 7 p.e., reached a maximum between days 35 and 49 p.e, peaking simultaneously with the redial multiplication, to disappear almost completely at day 70 p.e. (Rondelaud and Barthe 1980a). In the APO, the number of cells reached a maximum at day 21 p.e. (Rondelaud and Barthe 1981). However, although a certain hemocyte reaction can be observed, no encapsulation of the parasitic larva occurred (Rondelaud and Barthe 1980a). In this sense, an increase in the number of hemocytes has been detected in advanced infections in different Lymnaeidae–trematode models; e.g. L. stagnalis–T. szidati (van der Knaap et al. 1987), S. palustris–Metaleptocephalus sp. (Russo and Lagadic 2000), R. lagotis–T. regenti (Skála et al. 2014), L. catascopium–S. douthitti (Loker 1979) possibly to cope with the effects of parasite multiplication and/or cercarial shedding. In L. stagnalis, the expression of the MDM receptor is suppressed by the avian trematode parasite T. szidati with a significant downregulation to 76% of the non-parasitized level observed at 2 weeks p.e. that continues at least up to 8 week p.e. where a 21% decrease of expression was still observable (Hoek et al. 1996). Moreover, a decrease on the phagocytic activity of the hemocytes was recorded from 5 weeks p.e. onwards (van der Knaap et al. 1987), whereas Horak and Deme (1998) reported just minor differences between infected and uninfected snails in terms of hemocyte phagocytic activity at 10 weeks p.e. Hemocytes from infected L. stagnalis shedding P. mutationis cercaria showed a significant diminish of their phagocytic and encapsulation capacities against E. coli bioparticles and Sephadex beads, respectively, compared to uninfected snails (Kryukova et al. 2014). Similarly, patent infection of T. regenti in R. lagotis results in a significant reduction of hemocyte phagocytic activity and H2O2 production when compared to uninfected counterparts, an effect that was associated with a lower PKC and ERK activity in cell adhesion assays (Skála et al. 2014).
11.4 What Could Be the Effects of a Parasitic Infection in the Lymnaeid Host?
In compatible host–parasite combinations, the successful infection of the snail host by a pathogen, once established, concomitantly leads to the development and/or reproduction of the parasite and to the continuity of its transmission cycle. This causes profound effects in several aspects within the host; from the physical damage to organs and tissues where the pathogen settled to changes in the metabolism and physiology of the host that ultimately result in changes in life history traits. Examples involving infection-associated effects that somehow contributes to reproduction and transmission of the parasite, particularly within snail–digenean systems, e.g. modulation of energy reserves, energetic budget and neuroendocrine metabolism (de Jong-Brink et al. 1997), behavioral changes (Żbikowska and Żbikowski 2015), host castration (Schallig et al. 1991), gigantism (Sorensen and Minchella 1998), or growth stunting (Muñoz-Antoli et al. 2007), are reported.
-
i.
On the integrity of internal organs and tissues
The development of an infection, particularly those characterized by high parasite burdens can result in a significant damage to the internal systems of the snail hosts. In the case of digenean trematodes, those involving the production of rediae within the host can inflict serious injuries to the snail’s tissues partly associated with the voracious behavior of this actively-feeding larva (Esch et al. 2002). For instance, structural modifications, degeneration, and necrosis have been described in the digestive and reproductive tissues of G. truncatula infected by F. hepatica, in association to the mechanic trauma exerted by the establishment and multiplication of the parasite in these organs (Rondelaud and Barthe 1980b; Wilson and Denison 1980). The lesions relate to the intensity of the evolutive infection: necrosis of the digestive gland is found in moderate to low infections, whereas gonadal atrophy with epithelial necrosis is usually found in highly infected snails (Sindou et al. 1991). Mid and late-infections by F. hepatica in G. truncatula also cause the destruction of renal epithelial cells followed by epithelial hyperplasia (Rondelaud and Barthe 1983), whereas in Omphiscola glabra, S. palustris, and Ampullaceana balthica, epithelial necrosis of the kidney has been recorded (Sindou et al. 1991). In addition, in O. glabra, multifocal and generalized epithelial necrosis, and epithelial reconstitution with cell hyperplasia can be observed in the digestive gland of F. hepatica-infected snails, with the latter event occurring more frequently and appearing sooner in individuals showing bigger shells (Sindou et al. 1991). In Galba viator infected with F. hepatica, severe epithelial damage and leakage of hemolymph into the vicinity of the digestive gland leading to a persistent degeneration and necrotic process of the organ, was observed and related to the presence the parasite rediae (Magalhães et al. 2008). Edema, cellular degeneration, and loss of cytoplasmic content of the hepatic cells and glycogen clumps in the luminal boundary of the hepatopancreatic cells were noted in Radix auricularia infected with Fasciola gigantica (Patnaik 1968).
It was observed that infection by Echinostoma revolutum in young S. palustris snails partially inhibited the development of the hepatopancreas, and the hepatopancreatic epithelium was reduced to a syncytium showing necrosis and loss of tissue. In infected adult snails, the hepatopancreas was crowded to the periphery and the epithelium commonly underwent vacuolar degeneration, whereas gonads were degenerated (Zischke 1972). In L. elodes, the growth of the rediae population of E. revolutum had a strong negative effect on the ovotestis and the digestive gland of the snail host; the size and integrity of the organs decreased with the progression of the infection due to their apparent consumption by the parasitic larvae (Sorensen and Minchella 1998).
In L. catascopium–S. douthitti, the migration and establishment of daughter sporocyst results in atrophy of the digestive gland and, in consequence, many smaller lobules of the digestive gland disappear, whilst the remaining lobules became more widely separated. In occasions, the pressure resulting from growth of mother sporocysts has proved sufficient to rupture the transverse membrane that normally separates the cephalopedal and visceral sinuses in lymnaeid snails (Loker 1979). In the kidney of L. stagnalis parasitized by T. ocellata (North American isolate), vacuolization was observed within the cells of the tubule epithelium, although no evidences of mechanical trauma related to the development and multiplication of the sporocysts were recorded. Significantly, the reproductive tract of infected snails failed to develop, remaining rudimentary with the albumen and muciparous glands often being indiscernible and the ovotestis regressed (McClelland and Bourns 1969).
Experimental infection of P. columella with the entomopathogenic nematode Heterorhabditis baujardi results in severe damage of the cephalopodal mass and the digestive gland due to the intense cell disorganization and the formation of granulomatous reactions caused by the inflammatory response occurring in the infected snails (Tunholi et al. 2017).
-
ii.
On the physiology and the metabolism
Pathogens profit from host resources to support their development which causes significant physiological and metabolic alterations aiming at balancing the energy budget (Patnaik 1968; de Jong-Brink et al. 1997; Humiczewska and Rajski 2005). Furthermore, parasitism can also alter the biological rhythms, foraging behavior, and responses to abiotic stimuli of the host, usually benefiting parasite development and/or transmission (Voutilainen 2010; Żbikowska and Żbikowski 2015). For instance, changes in ionic balance and calcium deposition can result from infection. Significant hypercalcification of the shell has been observed in infected compared to uninfected L. stagnalis when inhabiting lakes with low calcium content (Żbikowska 2003), whereas the calcium content of the shells and soft parts of Radix natalensis shedding F. gigantica cercariae was lower compared to uninfected snails (Mostafa 2008). In the latter study, the concentration of lead and sodium ions in infected R. natalensis showed a significant decrease during the patent period whilst higher concentrations of zinc, potassium, and copper were found in infected compared to non-infected snails.
Infection of R. auricularia by E. revolutum results in depletory changes in glycogen reserves within host tissues which are shifted toward the development of or deposition in the larval trematode (Patnaik 1968). Lower rates of oxygen consumption have been reported in E. revolutum-infected S. palustris in association with the parasite burden (Zischke 1972), whereas the galactogen concentration in the albumen gland of P. columella exposed to the nematode Heterorhabditis baujardi declined significantly as infection progressed in relation to non-exposed control group (Tunholi et al. 2017). Infection of P. columella by Echinostoma paraensei significantly altered the contents of carbohydrates (60% reduction) and total proteins (76.4% reduction) in the digestive gland during the prepatent period, whereas total protein amounts were also 80% lower in the hemolymph of parasitized snails compared to uninfected individuals. Nitrogen catabolism was estimated by measuring uric acid and urea; the former was reduced in both organs after 10 days p.e, whereas urea increased in the hemolymph, peaking at day 10 to decrease afterwards, and it also increased in the albumen gland by 20 days p.e. when the larvae were well-established. A shift in infected host from using carbohydrates to proteins as main energy source was suggested possibly in response to the depletion of snail glycogen deposits caused by the parasites (Pinheiro et al. 2009).
In G. truncatula, infection by F. hepatica resulted in increased cellular vacuolization and lysosomal enzyme activity that apparently leads to an increase in intracellular digestive processes, including autolysis, most likely as a result of a host’s attempt to maintain its nutritional requirements under parasitic stress (Moore and Halton 1973). Furthermore, it leads to a significant decrease of up to 80% of the lipids reserves in the digestive gland of the snail host from early infection onwards, which was associated to a mobilization of lipid energy reserves to compensate for the deficiency of carbohydrates as they are used by F. hepatica, and to the direct accumulation of the lipid content by the different parasitic larvae (Humiczewska and Rajski 2005). An increase of 38% in the free fatty acid content probably correlated to a decrease of 56% and 52% in the monoglyceride and the triglyceride fractions has been observed in the hepatopancreas of F. hepatica-infected G. truncatula compared to uninfected snails, which was linked to a direct increase of hydrolysis of host’s triglycerides and diglycerides. Moreover, phospholipids were reduced by 13% compared to uninfected snails, with the most significant change being the 80% decrease of the phosphatidyl ethanolamine fraction (Southgate 1970).
On the other hand, thermo-behavioral study of L. stagnalis shedding Notocotylus attenuatus cercariae showed an induced anapyrexia that contrasted with the behavior of non-infected snails, which preferred the warmer end of the thermal gradient for longer periods of time. It was speculated that such behavioral change favored parasite transmission as snail hosts kept at a decreased temperature present increased life span and release significantly more cercariae (Żbikowska and Żbikowski 2015). In the case of Diplostomum pseudospathaceum infection, it has been suggested that, even though parasitized L. stagnalis snails display fewer feeding events per individual than their uninfected counterparts, they are more active in the presence of a fish predator (parasite’s second intermediate host) than in the absence of other non-fish predators. As D. pseudospathaceum cercariae have low motility and a short infective span once release in the water, this change of the foraging pattern of the infected snail enhances the success of transmission of the parasite to its next host, whether it is a direct result of parasite’s manipulation or a by-product of infection (Voutilainen 2010).
One of the most studied examples of parasite-mediated regulation of the neuroendocrine system of the host is that of T. szidati when infecting L. stagnalis; its effect resulting in the castration of infected hosts (Schallig et al. 1991). Schistosomin, a host neuropeptide elicited upon infection (Schallig et al. 1991), has been strongly associated to physiological and metabolic effects observed on parasitized snails. They are featured by the inhibition of the bioactivity of female gonadotropic hormones, i.e. the caudodorsal cell hormone, calfluxin, and dorsal body hormone, via the antagonizing effect of schistosomin on target organs, the changes in the electrophysiological properties and the release growth- and reproduction-controlling peptides by neuroendocrine centers, and the changes in the gene expression in the central nervous system of infected L. stagnalis snails (reviewed in de Jong-Brink et al. 1997).
-
iii.
On the life history traits
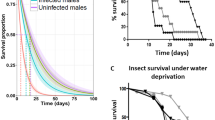
Energy constrains during infection affect host’s life history traits, as both, the host and the pathogen attempt to use the same limited resources to maximize their own growth, reproduction, and survival. At the light of host–parasite interactions, host’s traits must be interpreted in terms of the life history strategies of both, the parasite and of the host (Sorensen and Minchella 2001).
In rediae-producing infections, the strongest negative effect on host survival usually occurs during the prepatent period, whilst infections characterized by the development of mother and daughter sporocysts generally decrease host survival during patency (Sorensen and Minchella 2001). Moreover, survival usually decreases as the infective dose or the number of successive exposures increases (Rondelaud and Barthe 1982; Alba et al. 2018). In any case, although survival of the snail host can be significantly affected by infection (see (Sorensen and Minchella 1998; Gutiérrez et al. 2000, 2002; Salazar et al. 2006; Muñoz-Antoli et al. 2007) for examples on parasitized lymnaeid snails), when analyzing this parameter it should be considered that the degree of damage caused by the parasite is kept in check by the requirement that the host survives long enough until the time comes for the pathogen to be transmitted (Sorensen and Minchella 1998; Żbikowska 2011). For instance, in one study, infection of L. stagnalis by T. ocellata resulted in an increase of the life span of infected snails; 90% of the exposed group survived at week 28 of age when all uninfected snails were already dead (McClelland and Bourns 1969). In addition, snail age and host density also appear to affect host survival as well as compatibility. Death observed during mid and latter infections presumably indicates excessive energy demands or tissue damage imposed by the parasite, whilst mortality of exposed snail, especially during early infection may reflect compatibility issues (Sorensen and Minchella 2001). Therefore, survival has been used as an additional element to depict gradients within compatible host–parasite systems (Alba et al. 2018; Vázquez et al. 2019). The dynamic balance allowing parasite’s exploitation of host resources without severely endangered critical resources for host’s survival, at least before transmission is possible, generally occurs at the expenses of other hosts traits such as reproduction (Sorensen and Minchella 2001).
Reduction in the reproductive parameters of the host can result from parasitism. The entomopathogenic nematode H. baujardi negatively affects oviposition, the number of eggs and eggs masses per snail, and the hatching rate of infected P. columella, significantly impairing reproduction (Tunholi et al. 2017). Some studies have shown that infection of P. columella by the digeneans trematodes F. magna and F. hepatica is accompanied by a reduction in egg mass production and by a decrease in the number of eggs per egg mass (Salazar et al. 2006; Pankrác et al. 2016). In G. cubensis–F. hepatica, infected snails showed lower fecundity parameters than non-exposed individuals (Gutiérrez et al. 2000) whereas, egg laying progressively declined during infection of S. palustris snails by E. revolutum until complete castration occurred (Zischke 1972). During experimental infection of Lymnaea olulla by F. gigantica, a consistently lower levels of egg and embryo production was observed throughout a 7 weeks observation period in parasitized snails (Kumkate et al. 2009). A fecundity compensation can occur with parasitic castrators in which large numbers of eggs are produced by infected individuals before a significant impairment of oviposition ensues (Schallig et al. 1991; Sorensen and Minchella 2001; Gutiérrez et al. 2002). In any case, the ultimate reduction in the reproductive outputs of parasitized snails may be directly linked to the redirection of limited host resources to better support infection, as in L. stagnalis–T. ocellata/T. szidati systems (McClelland and Bourns 1969; Schallig et al. 1991), or to a direct result of biochemical, histopathological, and/or mechanical damage caused by the development of the parasite in the snail’s reproductive organs, as observed in G. truncatula when heavily infected with F. hepatica (Wilson and Denison 1980). Regarding the first cause, there is debate concerning whether parasite-mediated fecundity reduction reflects “strategy” directed by the host or the parasite. However, parasitic castrators that actively divert host resources away from reproduction can catalyze another phenomenon: enhanced growth of infected snails (Sorensen and Minchella 2001; Hall et al. 2007). Thus, it is not surprising that examples of both, a significant decrease of reproductive traits and an increase of growth rate and shell size in parasitized snails have been observed, e.g. L. stagnalis–T. szidati/D. pseudospathaceum (Schallig et al. 1991; Seppälä et al. 2013), L. elodes–E. revolutum/Plagiorchis elegans (Sorensen and Minchella 1998; Zakikhani and Rau 1999). The overall increase on the shell length on infected snails may vary with factors like food availability (see Sorensen and Minchella 2001), the infective dose or the age of the snail host (Zakikhani and Rau 1999). Conversely to the previous examples of gigantism in infected hosts, it has been reported that the infection of Peregriana peregra by Echinostoma friedi not only produces a total parasitic castration but also reduction of normal development (stunting) of infected snails (Muñoz-Antoli et al. 2007).
Independently of reproductive constrains, evidence of parasitism by digenean trematodes on the size and shape of lymnaeid snails has been observed in both, field and experimental studies (see Sorensen and Minchella 2001 for review). A decrease of the growth rate during early and mid-infection and a reduction of the rate of shell formation in young snails have been reported in E. revolutum-infected S. palustris in association with the parasite burden (Zischke 1972). In addition, it has been observed that field-occurring L. stagnalis snails infected with Echinoparyphium aconiatum, E. revolutum, D. pseudospathaceum, and Opisthioglyphe ranae differed in shell shape compared to uninfected individuals; a slenderer shell, with a more elongate spiral was recorded in the former group and suggested to be associated with infection (Żbikowska and Żbikowski 2005).
11.5 How Can the Environmental Factors Affect the Infection Outcome of Lymnaeid Snails: Parasite Interaction?
Host–parasite interactions result from the conjunction of the interplay between biological factors (encounter filter and compatibility filter), which are primarily determined by the genetic and epigenetic structure of each partner, and a plethora of environmental factors, as changes in environmental variables (e.g., temperature, droughts, pollution, co-infections) may affect the exposure to parasites and the outcome of the infection, even in highly compatible host–parasite systems (Sandland and Minchella 2003; Wolinska and King 2009). It is important to note, that the same factor could have positive and negative effects on the different encounter and compatibility filters and thus, it is the final balance between the multiple influences what will decide the faith or the extent of the contact/infection. In this sense, environmental factors can affect crucial features of parasite’s infectivity and/or survival (e.g., Rondelaud and Barthe 1980b; Ford et al. 1998; Morley et al. 2007) or the immunological and/or life history traits in the host (e.g., Sandland and Minchella 2003; Leicht et al. 2013), all of which could modify encounter and compatibility filters and the ultimate phenotype resulting from exposure to the pathogen.
In addition, ecological features of the host population (e.g., density, abundance, aggregation, demographic structure, population genetics), by themselves, are also significant influencers of encounter and compatibility filters. For instance, although parasitism, in general, should have a negative effect on host population density, a positive correlation between L. stagnalis density and Diplostomum sp. prevalence was reported as the parasite was fairly common in L. stagnalis populations in small lakes but rare in large lakes (Voutilainen et al. 2009). The demographic structure of the host population can also influence contact and compatibility events with parasites; adult snails showing higher sizes could be better in attracting parasites as they may produce higher amounts of chemical clues (Langeloh and Seppälä 2018) although, contrastingly, susceptibility to infection usually decreases with age (e.g., Zakikhani and Rau 1999). Moreover, the high transmission of F. hepatica through G. truncatula snails in the Bolivian Altiplano has been related to a depleted genetic diversity within the host population featured by the occurrence of a single and unique genotype that is highly compatible with the parasite (Meunier et al. 2001). Similar as to what occurred at the individual level, the ecological features of host populations are also constantly and ultimately influenced and regulated by a plethora of environmental factors and their dynamics.
-
i.
Abiotic factors
The degree of openness of the encounter filter ultimately depends on host availability which is determined by environmental factors that define the temporal and spatial occurrence of a given host–parasite system, as well as its meeting rates (Detwiler and Minchella 2009). Factors such as altitude (Walker et al. 2008), pH and water hardness (Alba et al. 2019b) have been reported as significant in determining the occurrence and habitat preference of different lymnaeid snails. Furthermore, certain seasonal patterns of transmission have been informed for different lymnaeid-borne trematodes species in correspondence to fluctuations of abiotic factors such as rainfall pattern, temperature, water flow/level, and soil evapotranspiration driving variations on parasite’s infectivity and/or intermediate hosts populations (e.g., Luzón-Peña et al. 1995; Yurlova et al. 2006; Żbikowska et al. 2006). In this sense, although many lymnaeid species show resilience to desiccation, estivation of infected snails during drought can result in a significant degeneration of the intramolluscan parasitic larvae most likely related to an increased stress and to the negative effect on host nutrition, as observed in G. truncatula–F. hepatica (Rondelaud and Barthe 1980b). In addition, water with high turbidity levels negatively influence host-finding capacity of F. hepatica miracidium (Christensen 1980), whereas the oxygen content and the pH of the water were identified to be associated with the probability of infection by trematodes of field-occurring Radix auricularia and L. stagnalis (Soldánová et al. 2010). Negative and positive correlations between the mean abundance and the water level have been reported for Cotylurus cornutus and Molinella anceps trematodes in L. stagnalis inhabiting a Siberian lake (Yurlova et al. 2006). An increased risk of infection of G. truncatula by F. hepatica has been associated with the type of habitat; higher risk was related to snail populations originating from spring swamps, wells, and reeds compared to populations from streams (Schweizer et al. 2007).
The influence of temperature on hosts, parasites and their interplay has been studied from different points of view, particularly in the case of ectothermic hosts such as snails. Overall, an increase of temperature within the optimal range usually favors higher metabolic rates and physiological process such as the motility of free-living parasitic stages (Morley et al. 2007; Selbach and Poulin 2020), and the growth rate and fecundity of the snail host (Vaughn 1953; Leicht et al. 2013). However, a decrease or elevation of the temperature away from the biological range of tolerance for each partner will produce negative effects on host’s life traits, as well as on parasite’s infectivity and survival (e.g., Vaughn 1953; Luzón-Peña et al. 1995; McCarthy 1999). Furthermore, intramolluscan colonization, development and further transmission of the parasite are also influenced by temperature (Wilson and Draskau 1976; Abrous et al. 1999; Morley et al. 2007) in a species-specific manner (see Selbach and Poulin 2020) for examples among trematodes). In this sense, immune responsiveness of the host is another parameter heavily affected by changes in this environmental factor. For instance, an increased transcription of the cytokine MIF was recorded in L. stagnalis snails exposed to high temperature (Seppälä et al. 2021), whereas a long heat wave (i.e., for more than 7 days) was found to decrease hemocyte concentration and PO-like activity (Leicht et al. 2013). It has been referred that phagocytosis in this snail species is optimal between 20–25 °C range (van der Knaap et al. 1983b). An increased susceptibility of L. stagnalis snails to E. aconiatum was observed when the host population was exposed to a heat wave for 7 days, in association to the impairment of host’s immune reactiveness known to occur under such treatment (Leicht and Seppälä 2014).
Chemical pollution, particularly those associated to human practices such as agriculture, can also have potential effects on parasite transmission. Several studies have experimentally assessed the influence of toxicants on the biology of the lymnaeid snails. A myriad of effects on their life history traits and population genetic structure (see Coutellec et al. 2013; Amorim et al. 2019 for examples on Lymnaea stagnalis), on the IDS and snail immunoreactivity (e.g., Russo and Lagadic 2000; Russo et al. 2008), and on host–parasite interactions, in particular (e.g., in R. natalensis–F. gigantica model, see Soliman 2009); P. peregra/L. stagnalis–Diplostomum spathaceum, see Morley et al. 2005), have been recorded. Others discuss about a potential relation between the use of herbicides and chemical pollutants on food crops and the punctual decline of lymnaeid snails’ density in the field (Nguyen et al. 2017). Moreover, it has been demonstrated that pollution by the herbicide Paraquat of the aquatic environment not only adversely affects R. natalensis metabolism but also resulted in a significant reduction of the infectivity of F. gigantica miracidia to the snail (Bakry et al. 2016).
-
ii.
Biotic factors
Biotic factors add complexity to host–parasite interplays as other organisms, co-occurring with the system, can significantly influence encounter and compatibility filters and transmission success through a variety of interactions (predation, competence, dilution effect, decoy effect, invasive species, etc.). This implies that biodiversity, in general, should have significant effects on parasite transmission (Thieltges et al. 2008). In this sense, food resources have proved significant in host–parasite interactions as factors like the quantity and quality of the diet may significantly affect the encounter filter and can modify the physiological characteristics and conditions of the host. For instance, strong long-term, but not short-term food deprivation reduced L. stagnalis chemical attractiveness to E. aconiatum cercariae and thus, may negatively affect the probability of encounter (Seppälä and Leicht 2015; Langeloh and Seppälä 2018). Furthermore, starvation can directly modify the IDS and in consequence, the immune reactivity and the ultimate compatibility of the host. In this sense, food deprivation downregulates the transcription of an M-type lectin, and GST, and enhances the transcription of MIF, LBP/BPI and lysozyme in L. stagnalis (Seppälä et al. 2021). Concerning hemocytes, those derived from 14 days-starved L. stagnalis snails phagocytized less avidly than cells of individuals fed ad libitum and showed significantly higher titers of agglutinin in the hemolymph (van der Knaap et al. 1983b). Moreover, E. revolutum-infected L. elodes snails subject to a low-protein diet exhibited lower survival than infected snails fed on a high-protein diet and thus, influencing transmission to the next host (Sandland and Minchella 2003). Food deprivation constrained the overall parasite within-host reproduction in L. stagnalis–D. pseudospathaceum system, as the release of parasite transmission stages was reduced (Seppälä et al. 2015). In G. truncatula, the cercarial production of F. hepatica, F. gigantica, and Calicophoron daubneyi varied depending on the different food sources used (Rondelaud et al. 2002a).
In addition, an increased diversity and abundance of co-occurring non-host species could reduce the risk of contact and further infection of suitable snails, a phenomenon commonly known as “dilution effect” (Civitello et al. 2015) where non-host species can disrupt parasite–host encounter via several mechanisms like predation of hosts and of free-living stages of parasites (Johnson et al. 2010), or by acting as decoys attracting the pathogen (Marszewska et al. 2018). In particular, besides directly reducing the abundance of the host, it was observed through experimental infections of G. truncatula with F. hepatica that the stress caused by the presence of predators (sciomyzid larvae or zonitid snails) in the host during infection significantly influences survival of parasitized snails; the duration of the cercarial shedding and the number of emitted cercariae were reduced compared to controls (infected snails not subjected to predators; see Rondelaud et al. 2002b).
As each snail host presents limited nutrient and spatial resources, a structuration of the parasite community, driven by complex interactions among infecting pathogens (particularly trematodes) related to competition, facilitation, or predation, can occur within the host, during a multi-infection process (Lie et al. 1973; Abrous et al. 1996; Soldánová et al. 2012). For instance, infection synergism between C. daubneyi followed by F. hepatica, and between Trichobilharzia brevis followed by Echinostoma hystricosum has being reported when successive cross-exposures, in the given order, were performed in G. truncatula or L. rubiginosa, as higher infection rates were attained compared to each mono species-infection (Lie et al. 1973; Augot et al. 1996). Conversely, if T. brevis followed a primary infection by E. hystricosum, the predatory rediae of E. hystricosum consumed T. brevis sporocysts impairing the establishment of the second parasite (Lie et al. 1973). A field study on the trematode infracommunity of L. stagnalis reported two pairs of trematodes, i.e. P. elegans/T. szidati and M. anceps/Notocotylus attenuatus, that were more frequently found infecting in combination than would be expected by chance suggesting facilitation. Moreover, competitive relationships among trematode species were hypothesized as a dominance hierarchy in which seven top dominant species with putatively similar competitive abilities reduced the prevalence of the other trematode species found (which develop in sporocysts only; see Soldánová et al. 2012).
11.6 What Can the Study of Interaction Between Lymnaeid Snails and Their Parasite Bring to Evolutionary Biology and Disease Control, and Where Do We Go from Here?
Since some snail-borne parasites cause significant diseases in humans and domestic animals, the study of the host snails and of their specific interaction may have a great impact on veterinary and human health sciences. In particular, given that it is mandatory for digenean parasites to develop inside their snail hosts, controlling the populations of the latter is a suitable strategy for directly tackling snail-borne trematodiasis. However, the mechanisms and factors promoting the emergence or re-emergence of these diseases are still poorly understood making any predictions on their establishment and spreading, in a context of environmental changes, and successfully preventing their propagation, particularly challenging. In this sense, a thorough look into snail–parasite interaction is mandatory (Adema et al. 2012). Aspects such as the factors determining the population structure and geographical distribution of snails and thus, possible transmission foci, the effects of environmental perturbations on the distribution and transmission of parasites through their vector snails, and the extent of the influence of encounter and compatibility filters between hosts and parasites versus the ecological factors that limit infections, are among the significant questions that can be solved by investigating the biology of snail–parasite interactions. Moreover, the deep study of the immunobiological features of each particular digenean–mollusk system could allow to identify molecules with potential biomedical relevance, and could also serve as model to study the impact of pollutants or other stressors on immune function, to monitor climate change and to record its impact, or ultimately to predict their effects on parasite transmission and vectorial competence (Loker 2010). The knowledge on how immunity and virulence have evolved to influence the snail–parasite compatibility or which are the genetic markers determining such compatibility or the regulators of parasite transmission in natural snail populations can only be attained through more investigations encompassing snail–parasite dynamics (Adema et al. 2012). They will also bring light on evolutionary and environmental immunology.
Nevertheless, in spite of the crucial contribution of this field to move securely toward the control of human trematodiasis or a major understanding of invertebrate’s biology and immunology, and the progresses achieved, significant knowledge gaps remain, specially within Lymnaeidae–parasites systems. Encounter filter and its association to susceptibility/virulence of host and parasites needs to be deeply characterized under experimental as well as natural conditions. The role of microbiota in lymnaeid snails–parasite dynamics has been largely overlooked, although the host genome and the microbiota meta-genome are a real unit of selection, one that is not only associated to the host genotype and/or physiological status but also to environmental variations, and that affects host plasticity and adaptive responses (Zilber-Rosenberg and Rosenberg 2008). Therefore, identifying microbial communities more or less prone to modify parasite transmission in lymnaeid host would have critical consequences in terms of snail control strategies for fighting Lymnaeidae-borne diseases in the field and to understand evolutionary dynamics. Furthermore, and despite the advances, our knowledge of PRRs, receptor–ligand interactions, downstream mechanisms of signaling transduction and molecular and mechanistic regulators, humoral effectors and defense reactions to convey an appropriate immune response is currently poor in snails in general, and in the family Lymnaeidae in particular. Therefore, it warrants further investigations by multi-“omics” methods and functional validations as it is crucial for depicting the (epi)genetic and molecular bases of the defense responses. In this sense, although RNA interference is a powerful transient gene-expression repression system widely employed in snails including the Lymnaeidae (e.g., Korneev et al. 2002; Lu and Feng 2011), the potential of the CRISPR-Cas9 gene editing technique offers a wider variety of applications (from complete gene knock-out to gene editing), appears to be a more determinant approach that it is already under development to generate transgenic mollusc (Perry and Henry 2014). A recent report demonstrates its successful application for the genome editing of L. stagnalis (Abe and Kuroda 2019) and opens up significant opportunities for functional genomics to investigate the role of specific genes in snail immunobiology. Moreover, the lack of molecular markers to properly classify hemocytes, its ontogeny and functional differences deserves further efforts in order to properly elucidate the different lineages or types, and their specific role during infection dynamics. In-depth characterization of hemocyte populations and their interaction with trematodes to discriminate transcriptional infra-populations, trajectories of differentiation and specific genes related to each cellular stage/lineage can now be performed using Single-Cell RNA sequencing technology (scRNA-seq; see Fu et al. 2020).
It is important to note, that most of the immunobiological information concerning lymnaeid snails comes primarily from the study of a single species, the pond snail L. stagnalis. Nonetheless, as the response of different species of snails to parasitic infections is greatly varied, the information about one snail host/parasite model cannot be strictly applied to other systems. Hence, it is crucial for future investigations to encompass other species in order to widen up the view of the IDS of the Lymnaeidae and its interaction with pathogens. In parallel, further characterization of virulence factors of pathogens affecting lymnaeid snails should be carried out in order to completely elucidate the evolutionary bases of each host–parasite interaction. Topics such as the specificity of the defense response (e.g., (van der Knaap et al. 1983a; Seppälä et al. 2021), immune memory (van der Knaap et al. 1983a), immune priming (Alba et al. 2018), and transgenerational priming (Vorontsova et al. 2019) have been only modestly embraced within the Lymnaeidae and further efforts should be directed toward them.
In addition, investigations of the effects of host–parasite interplay on lymnaeid snail physiology, metabolism and ultimately, on life history traits must be emphasized and approached by integrative studies. Lastly, as the Anthropocene imposes significant challenges for environmental dynamics, it is pivotal to develop a more mechanistic understanding of how anthropic disturbances and environmental degradation, habitat transformation, biodiversity loss and invasive species, and climate change will affect hosts and parasites communities and their interplay if we aimed at predicting changes in transmission dynamics and disease epidemiology. In this sense, field studies, although difficult to carried out, should be particularly stimulated not only with the finality of building up basic knowledge on host–parasite–environment interplay, but also of exploring ways to practically exploit snail-based control procedures of parasite transmission. Vector mediation by transgenesis strategies either using introgression of selected snail genotypes refractory to trematodiasis infection into, or direct genetic modification by CRISPR-Cas9 (transfection with a multi-resistant gene) of natural populations might be envisioned for increasing snail resistance, reducing their vectorial competence and tackling pathogen transmission. Profiting on snail’s microbiota for potentially controlling snail-borne diseases, either through chemical modification or using paratransgenesis solutions (using transgenic bacteria expressing foreign gene products; see Coutinho-Abreu et al. 2010) to favor snail’s microbial communities rendering the host more refractory or resistant to parasite infection, could be also projected.
The incorporation of additional research efforts, new competences, technology, and funding is as challenging as it is essential since global attention is traditionally diverted toward other host–parasite systems mostly driven by the notoriety of other vectors and diseases in global health agendas. The scarce resources and interest devoted to tackle snails and the diseases they transmit should be always considered, as it is a major limitation for exploring related scientific and practical opportunities. In addition, the list of scientists working on snails and the diseases they transmit continues to grow thinner (Adema et al. 2012). Nevertheless, it must be brought forward that the variety of models to study, the exciting investigations and discoveries that lay ahead, the applicability of such knowledge in different disciplines and their usefulness for science and society, present to scientist and decision-makers an attractive road deserving to be explored.
References
Abe M, Kuroda R (2019) The development of CRISPR for a mollusc establishes the formin Lsdia1 as the long-sought gene for snail dextral/sinistral coiling. Development 146:175976. https://doi.org/10.1242/dev.175976
Abrous M, Rondelaud D, Dreyfuss G (1996) Paramphistomum daubneyi and Fasciola hepatica: the effect of dual infection on prevalence and cercarial shedding in preadult Lymnaea glabra. J Parasitol 82:1026–1029
Abrous M, Rondelaud D, Dreyfuss G (1999) Influence of low temperatures on the cercarial shedding of Paramphistomum daubneyi from the snail Lymnaea truncatula. Parasite 6:85–88. https://doi.org/10.1051/parasite/1999061085
Adema CM, van Deutekom-Mulder EC, van der Knaap WP et al (1991) Generation of oxygen radicals in hemocytes of the snail Lymnaea stagnalis in relation to the rate of phagocytosis. Dev Comp Immunol 15:17–26
Adema CM, van Deutekom-Mulder EC, van der Knaap WP et al (1993) NADPH-oxidase activity: the probable source of reactive oxygen intermediate generation in hemocytes of the gastropod Lymnaea stagnalis. J Leukoc Biol 54:379–383. https://doi.org/10.1002/jlb.54.5.379
Adema CM, Mohandas A, van der Knaap WP et al (1994a) Separation of Lymnaea stagnalis hemocytes by density gradient centrifugation. Dev Comp Immunol 18:25–31
Adema CM, van Deutekom-Mulder EC, Van der Knaap WP et al (1994b) Schistosomicidal activities of Lymnaea stagnalis hemocytes: the role of oxygen radicals. Parasitology 109:479–485
Adema CM, Bayne CJ, Bridger JM et al (2012) Will all scientists working on snails and the disease they transmit please stand up? PLoS Negl Trop Dis 6:e1835. https://doi.org/10.1371/journal.pntd.0001835
Alba A, Hernández H, Marcet R et al (2014) Exploring the antigenic features of Fasciola hepatica (Trematoda: Digenea) rediae through the evaluation of three different antigenic candidates for further monoclonal antibody development. Parasitol Res 113:3185–3193
Alba A, Vázquez AA, Sánchez J et al (2018) Fasciola hepatica - Pseudosuccinea columella interaction: effect of increasing parasite doses, successive exposures and geographic origin on the infection outcome of naturally-resistant and susceptible snails from Cuba. Parasite Vector 11:559. https://doi.org/10.1186/s13071-018-3155
Alba A, Tetrau G, Chaparro C et al (2019a) Field-occurring resistance to Fasciola hepatica (Trematoda) in Pseudosuccinea columella snails: a review from literature and insights from comparative “omic” analyses. Dev Comp Immunol 101:103463. https://doi.org/10.1016/j.dci.2019.103463
Alba A, Vázquez AA, Sánchez J et al (2019b) Patterns of distribution, population genetics and ecological requirements of field-occurring resistant and susceptible Pseudosuccinea columella snails to Fasciola hepatica in Cuba. Sci Rep 9:14359. https://doi.org/10.1038/s41598-019-50894-7
Alba A, Duval D, Sánchez J et al (2020) The immunobiological interplay between Pseudosuccinea columella resistant/susceptible snails with Fasciola hepatica: hemocytes in the spotlight. Dev Comp Immunol 102:103485. https://doi.org/10.1016/j.dci.2019.103485
Aldhoun J, Kment P, Horak P (2016) Historical analysis of the type species of the genus Trichobilharzia Skrjabin et Zakharov, 1920 (Platyhelminthes: Schistosomatidae). Zootaxa 4084:593–595. https://doi.org/10.11646/zootaxa.4084.4.10
Allam B, Espinosa E (2015) Mucosal immunity in mollusks. In: Beck BH, Peatman E (eds) Mucosal health in aquaculture. Academic Press, Amsterdam, pp 325–370. https://doi.org/10.1016/B978-0-12-417186-2.00012-1
Allan F, Dunn AM, Emery AM, Stothard JR et al (2013) Use of sentinel snails for the detection of Schistosoma haematobium transmission on Zanzibar and observations on transmission patterns. Acta Trop 128:234–240
Allan ER, Tennessen JA, Sharpton TJ et al (2018) Allelic variation in a single genomic region alters the microbiome of the snail Biomphalaria glabrata. J Hered 109:604–609. https://doi.org/10.1093/jhered/esy014
Amorim J, Abreu I, Rodrigues P et al (2019) Lymnaea stagnalis as a freshwater model invertebrate for ecotoxicological studies. Sci Total Environ 669:11–28
Augot D, Abrous M, Rondelaud D et al (1996) Paramphistomum daubneyi and Fasciola hepatica: the redial burden and cercarial shedding in Lymnaea truncatula subjected to successive unimiracidial cross-exposures. Parasitol Res 82:623–627
Baeza-Garcia A, Pierce RJ, Gourbal B et al (2010) Involvement of the cytokine MIF in the snail host immune response to the parasite Schistosoma mansoni. PLoS Pathog 6:e1001115. https://doi.org/10.1371/journal.ppat.1001115
Bakry FA, Eleiwa ME, Taha SA et al (2016) Comparative toxicity of Paraquat herbicide and some plant extracts in Lymnaea natalensis snails. Toxicol Ind Health 32:143–153. https://doi.org/10.1177/0748233713498457
Boisseaux P, Delignette-Muller ML, Abbaci K et al (2016) Analysis of hemocytes in Lymnaea stagnalis: characterization and effects of repeated hemolymph collections. Fish Shellfish Immunol 57:116–126
Bouétard A, Noirot C, Besnard AL et al (2012) Pyrosequencing-based transcriptomic resources in the pond snail Lymnaea stagnalis, with a focus on genes involved in molecular response to diquat-induced stress. Ecotoxicology 21:2222–2234. https://doi.org/10.1007/s10646-012-0977-1
Brown DS (1978) Pulmonate molluscs as intermediate hosts for digenetic trematodes. In: Fretter V, Peake J (eds) Pulmonates, vol 2A. Academic Press, London, pp 287–333
Calienes AF, Fraga J, Pointier JP et al (2004) Detection and genetic distance of resistant populations of Pseudosuccinea columella (Mollusca: Lymnaeidae) to Fasciola hepatica (Trematoda: Digenea) using RAPD markers. Acta Trop 92:83–87
Celi-Erazo M, Alda P, Montenegro-Franco M et al (2020) Prevalence of Fasciola hepatica infection in Galba cousini and Galba schirazensis from an Andean region of Ecuador. Vet Parasitol Reg Stud Rep 20:100390. https://doi.org/10.1016/j.vprsr.2020.100390
Cerenius L, Söderhäll K (2004) The prophenoloxidase-activating system in invertebrates. Immunol Rev 198:116–126. https://doi.org/10.1111/j.0105-2896.2004.00116.x
Christensen NO (1980) A review of the influence of host- and parasite-related factors and environmental conditions on the host-finding capacity of the trematode miracidium. Acta Trop 37:303–318
Christensen NO, Nansen P, Frandsen F (1976) Molluscs interfering with the capacity of Fasciola hepatica miracidia to infect Lymnaea truncatula. Parasitology 73:161–167. https://doi.org/10.1017/s0031182000046849
Civitello DJ, Cohen J, Fatima H et al (2015) Biodiversity inhibits parasites: broad evidence for the dilution effect. Proc Natl Acad Sci USA 28:8667–8671
Combes C (2001) Parasitism. The ecology and evolution of intimate interactions. University of Chicago Press, Chicago, IL
Coustau C, Gourbal B, Duval D et al (2015) Advances in gastropod immunity from the study of the interaction between the snail Biomphalaria glabrata and its parasites: a review of research progress over the last decade. Fish Shellfish Immunol 46:5–16
Coutellec MA, Besnard AL, Caquet T (2013) Population genetics of Lymnaea stagnalis experimentally exposed to cocktails of pesticides. Ecotoxicology 22:879–888
Coutinho-Abreu IV, Zhu KY, Ramalho-Ortigao M (2010) Transgenesis and paratransgenesis to control insect-borne diseases: current status and future challenges. Parasitol Int 59:1–8. https://doi.org/10.1016/j.parint.2009.10.002
Coyne K, Laursen JR, Yoshino TP (2015) In vitro effects of mucus from the mantle of compatible (Lymnaea elodes) and incompatible (Helisoma trivolvis) snail hosts on Fascioloides magna miracidia. J Parasitol 101:351–357
de Jong-Brink M, Bergamin-Sassen M, Solis Soto M (1997) Multiple strategies of schistosomes to meet their requirements in the intermediate snail host. Parasitology 123:S129–S141
Detwiler JT, Minchella DJ (2009) Intermediate host availability masks the strength of experimentally-derived colonisation patterns in echinostome trematodes. Int J Parasitol 39:585–590
Dida GO, Gelder FB, Anyona DN et al (2014) Distribution and abundance of schistosomiasis and fascioliasis host snails along the Mara River in Kenya and Tanzania. Infect Ecol Epidemiol 4(1):24281. https://doi.org/10.3402/iee.v4.24281
Dikkeboom R, Van der Knaap WP, Meuleman EA et al (1985) A comparative study on the internal defence system of juvenile and adult Lymnaea stagnalis. Immunology 55:547–553
Dikkeboom R, Tunagel JMGH, Mulder EC et al (1987) Hemocytes of the pond snail Lymnaea stagnalis generate reactive forms of oxygen. J Invert Pathol 49:321–331
Ehara T, Kitajima S, Kanzawa N et al (2002) Antimicrobial action of achacin is mediated by l-amino acid oxidase activity. FEBS Lett 531:509–512
Esch GW, Barger MA, Fellis J (2002) The transmission of digenetic trematodes: style, elegance, complexity. Integr Comp Biol 42:304–312. https://doi.org/10.1093/icb/42.2.304
Ford DM, Nollen PM, Romano MA (1998) The effects of salinity, pH and temperature on the half-life and longevity of Echinostoma caproni miracidia. J Helminthol 72:325–330
Franchini A, Ottaviani E (2000) Repair of molluscan tissue injury: role of PDGF and TGF-beta1. Tissue Cell 32:312–321. https://doi.org/10.1054/tice.2000.0118
Fu Y, Huang X, Zhang P et al (2020) Single-cell RNA sequencing identifies novel cell types in Drosophila blood. J Genet Genomics 47:175–186. https://doi.org/10.1016/j.jgg.2020.02.004
Galinier R, Portela J, Moné Y et al (2013) Biomphalysin, a new β pore-forming toxin involved in Biomphalaria glabrata immune defense against Schistosoma mansoni. PLoS Pathog 9:e1003216. https://doi.org/10.1371/journal.ppat.1003216
Georgieva K, Georgieva L, Mizinska-Boevska Y et al (2016) Study of surface carbohydrates in Galba truncatula tissues before and after infection with Fasciola hepatica. Mem Inst Oswaldo Cruz 111:475–483
Gerhardus MJT, Baggen JMC, van der Knaap WP et al (1991) Analysis of surface carbohydrates of Trichobilharzia ocellata miracidia and sporocyst using lectin binding techniques. Parasitology 103:51–59
Goodall CP, Bender RC, Broderick EJ et al (2004) Constitutive differences in Cu/Zn superoxide dismutase mRNA levels and activity in hemocytes of Biomphalaria glabrata (Mollusca) that are either susceptible or resistant to Schistosoma mansoni (Trematoda). Mol Biochem Parasitol 137:321–328
Guo F, Huang ZB, Huang MQ et al (2009) Effects of small abalone, Haliotis diversicolor, pedal mucus on bacterial growth, attachment, biofilm formation and community structure. Aquaculture 293:35–41
Gutiérrez A, Perera G, Yong M et al (2000) Life-history traits of Fossaria cubensis (Gastropoda: Lymnaeidae) under experimental exposure to Fasciola hepatica (Trematoda: Digenea). Mem Inst Oswaldo Cruz 95:747–752
Gutiérrez A, Perera G, Yong M et al (2002) Fasciola hepatica (Trematoda: Digenea): its effect on the life history traits of Pseudosuccinea columella (Gastropoda: Lymnaeidae), an uncommon interaction. Parasitol Res 88:535–539
Gutiérrez A, Pointier JP, Yong M et al (2003) Evidence of phenotypic differences between resistant and susceptible isolates of Pseudosuccinea columella (Gastropoda: Lymnaeidae) to Fasciola hepatica (Trematoda: Digenea) in Cuba. Parasitol Res 90:129–134
Hall SP, Becker C, Cáceres CE (2007) Parasitic castration: a perspective from a model of dynamic energy budgets. Int Comp Biol 47:295–309
Hambrook JR, Gharamah AA, Pila EA et al (2019) Biomphalaria glabrata Granulin increases resistance to Schistosoma mansoni infection in several Biomphalaria species and induces the production of reactive oxygen species by Haemocytes. Genes 11:38. https://doi.org/10.3390/genes11010038
Hermann PM, Nicol JJ, Nagle GT et al (2005) Epidermal growth factor-dependent enhancement of axonal regeneration in the pond snail Lymnaea stagnalis: role of phagocyte survival. J Comp Neurol 492:383–400
Hoek RM, Smit AB, Frings H et al (1996) A new Ig-superfamily member, molluscan defence molecule (MDM) from Lymnaea stagnalis, is down-regulated during parasitosis. Eur J Immunol 26:939–944
Horák P (1995) Developmentally regulated expression of surface carbohydrate residues on larval stages of the avian schistosome Trichobilharzia szidati. Folia Parasitol 42:255–265
Horak P, Deme R (1998) Lectins and saccharides in Lymnaea stagnalis haemocyte recognition. Comp Haematol Int 8:210–218
Horák P, van der Knaap WP (1997) Lectins in snail-trematode immune interactions: a review. Folia Parasitol 44:161–172
Horák P, Neeleman AP, van der Knaap WP et al (1998) Effect of saccharides on plasma mediated haemagglutination and in vitro phagocytosis by haemocytes of the snail Lymnaea stagnalis. Comp Haematol Int 8:43–49
Humiczewska M, Rajski K (2005) Lipids in the host-parasite system: digestive gland of Lymnaea truncatula infected with the developmental stages of Fasciola hepatica. Acta Parasitol 50:235–239
Humphries JE, Yoshino TP (2003) Cellular receptors and signal transduction in molluscan hemocytes: connections with the innate immune system of vertebrates. Integr Comp Biol 43:305–312
Humphries JE, Yoshino TP (2008) Regulation of hydrogen peroxide release in circulating haemocytes of the planorbid snail Biomphalaria glabrata. Dev Comp Immunol 32:554–562
Hunter R (1980) Effects of grazing on the quantity and density of freshwater aufwuchs. Hydrobiologia 69:251–259. https://doi.org/10.1007/BF00046800
Huot C, Clerissi C, Gourbal B et al (2020) Schistosomiasis vector snails and their microbiota display a phylosymbiosis pattern. Front Microbiol 10:3092. https://doi.org/10.3389/fmicb.2019.03092
Iakovleva NV, Gorbushin AM, Storey KB (2006) Modulation of mitogen-activated protein kinases (MAPK) activity in response to different immune stimuli in haemocytes of the common periwinkle Littorina littorea. Fish Shellfish Immunol 21:315–324
Jian J, Konopka J, Liu C (2013) Insights into the role of progranulin in immunity, infection, and inflammation. J Leukoc Biol 93:199–208. https://doi.org/10.1189/jlb.0812429
Johnson PT, Dobson A, Lafferty KD et al (2010) When parasites become prey: ecological and epidemiological significance of eating parasites. Trends Ecol Evol 25:362–371
Kalbe M, Haberl B, Haas W (1997) Miracidial host-finding in Fasciola hepatica and Trichobilharzia ocellata is stimulated by species-specific glycoconjugates released from the host snails. Parasitol Res 83:806–812
Kalbe M, Bernhard H, Haas BJ (2000) Snail host finding by Fasciola hepatica and Trichobilharzia ocellata: compound analysis of “Miracidia-attracting glycoproteins”. Exp Parasitol 96:231–242
Kaplan MJ, Radic M (2012) Neutrophil extracellular traps (NETs): double-edged swords of innate immunity. J Immunol 189:2689–2695. https://doi.org/10.4049/jimmunol.1201719
Kock KN, Wolmarans CT, Bornman M (2003) Distribution and habitats of the snail Lymnaea truncatula, intermediate host of the liver fluke Fasciola hepatica, in South Africa. J South Afr Vet Assoc 74:117–122
Korneev SA, Kemenes I, Straub V et al (2002) Suppression of nitric oxide (NO)-dependent behavior by double-stranded RNA-mediated silencing of a neuronal NO synthase gene. J Neurosci 22(11):RC227. https://doi.org/10.1523/JNEUROSCI.22-11-j0003.2002
Kryukova NA, Yurlova NI, Rastyagenko NM et al (2014) The influence of Plagiorchis mutationis larval infection on the cellular immune response of the snail host Lymnaea stagnalis. J Parasitol 100:284–287
Kumkate S, Onmek N, Boonburapong B et al (2009) Estrogen-related fecundity reduction of Lymnaea ollula following Fasciola gigantica infection. J Parasitol 95:1391–1396. https://doi.org/10.1645/GE-2080.1
Lacchini AH, Davies AJ, Mackintosh D et al (2006) beta-1, 3-glucan modulates PKC signalling in Lymnaea stagnalis defence cells: a role for PKC in H2O2 production and downstream ERK activation. J Exp Biol 209:4829–4840
Lange MK, Penagos-Tabares F, Muñoz-Caro T et al (2017) Gastropod-derived haemocyte extracellular traps entrap metastrongyloid larval stages of Angiostrongylus vasorum, Aelurostrongylus abstrusus and Troglostrongylus brevior. Parasites Vector 10:50. https://doi.org/10.1186/s13071-016-1961-z
Langeloh L, Seppälä O (2018) Relative importance of chemical attractiveness to parasites for susceptibility to trematode infection. Ecol Evol 8:8921–8929. https://doi.org/10.1002/ece3.4386
Le Clec'h W, Anderson TJC, Chevalier FD (2016) Characterisation of hemolymph phenoloxidase activity in two Biomphalaria snail species and impact of Schistosoma mansoni infection. Parasite Vector 9:32. https://doi.org/10.1186/s13071-016-1319-6
Leicht K, Seppälä O (2014) Infection success of Echinoparyphium aconiatum (Trematoda) in its snail host under high temperature: role of host resistance. Parasites Vector 7:192. https://doi.org/10.1186/1756-3305-7-192
Leicht K, Jokela J, Seppälä O (2013) An experimental heat wave changes immune defense and life history traits in a freshwater snail. Ecol Evol 3:4861–4871. https://doi.org/10.1002/ece3.874
Lie KJ, Lim HK, Ow-Yang CK (1973) Synergism and antagonism between two trematodes species in the snail Lymnaea rubiginosa. Int J Parasitol 3:729–733
Lockyer AE, Emery AM, Kane RA et al (2012) Early differential gene expression in haemocytes from resistant and susceptible Biomphalaria glabrata strains in response to Schistosoma mansoni. PLoS One 7:e51102. https://doi.org/10.1371/journal.pone.0051102
Loker ES (1979) Pathology and host responses induced by Schistosomatium douthitti in the freshwater snail Lymnaea catascopium. J Invert Pathol 33:265–273
Loker ES (2010) Gastropod immunobiology. Adv Exp Med Biol 708:17–43. https://doi.org/10.1007/978-1-4419-8059-5_2
Loker ES, Bayne CJ, Buckley PM et al (1982) Ultrastructure of encapsulation of Schistosoma mansoni mother sporocysts by hemocytes of juveniles of the 10-R2 strain of Biomphalaria glabrata. J Parasitol 68:84–94
Loker ES, Adema CM, Zhang SM et al (2004) Invertebrate immune systems – not homogeneous, not simple, not well understood. Immunol Rev 198:10–24
Lu TZ, Feng Z-P (2011) A sodium-leak current regulates pacemaker activity of adult central pattern generator neurons in Lymnaea stagnalis. PLoS One 6:e18745. https://doi.org/10.1371/journal.pone.0018745
Luzón-Peña M, Rojo-Vázquez FA, Gómez-Bautista M (1995) Seasonal availability of Fasciola hepatica metacercariae in a temperate Mediterranean area (Madrid, Spain). Zentralbl Veterinarmed B 42:577–585. https://doi.org/10.1111/j.1439-0450.1995.tb00751.x
Magalhães KC, Jannotti-Passos LK, Caldeira RL et al (2008) Isolation and detection of Fasciola hepatica DNA in Lymnaea viatrix from formalin-fixed and paraffin-embedded tissues through multiplex-PCR. Vet Parasitol 152:333–338
Mahilini H, Rajendran A (2008) Categorization of hemocytes of three gastropod species Trachea vittata (Muller), Pila globosa (Swainson) and Indoplanorbis exustus (Dehays). J Invertebr Pathol 97:e26. https://doi.org/10.1016/j.jip.2007.07.007
Marszewska A, Cichy A, Bulantová J et al (2018) Potamopyrgus antipodarum as a potential defender against swimmer’s itch in European recreational water bodies-experimental study. PeerJ 6:e5045. https://doi.org/10.7717/peerj.5045
McCarthy AM (1999) The influence of temperature on the survival and infectivity of the cercariae of Echinoparyphium recurvatum (Digenea: Echinostomatidae). Parasitology 118:383–388. https://doi.org/10.1017/s003118209900400x
McClelland G, Bourns TKR (1969) Effects of Trichobilharzia ocellata on growth, reproduction, and survival of Lymnaea stagnalis. Exp Parasitol 24:137–146
McReath AM, Reader TA, Southgate VR (1982) The development of the host-response in juvenile Lymnaea palustris to invasion by Fasciola hepatica. Z Parasitenkd 67:175–184. https://doi.org/10.1007/BF00928113
Meunier C, Tirard C, Hurtrez-Boussès S et al (2001) Lack of molluscan host diversity and the transmission of an emerging parasitic disease in Bolivia. Mol Ecol 10:1333–1340
Mitta G, Gourbal B, Grunau C et al (2017) The compatibility between Biomphalaria glabrata snails and Schistosoma mansoni: an increasingly complex puzzle. Adv Parasitol 97:111–145. https://doi.org/10.1016/bs.apar.2016.08.006
Monteil JF, Matricon-Gondran M (1991a) Hemocyte production in trematode-infected Lymnaea truncatula. Parasitol Res 77:491–497
Monteil JF, Matricon-Gondran M (1991b) Interactions between the snail Lymnaea truncatula and the plagiorchiid trematode Haplometra cylindracea. J Invert Pathol 58:127–135
Monteil JF, Matricon-Gondran M (1993) Structural and cytochemical study of the hemocytes in normal and trematode-infected Lymnaea truncatula. Parasitol Res 79:675–682. https://doi.org/10.1007/BF00932510
Moore MN, Halton DW (1973) Histochemical changes in the digestive gland of Lymnaea truncatula infected with Fasciola hepatica. Z Parasitenk 43:1–16
Morley NJ, Crane M, Lewis JW (2005) Changes in survival characteristics of Diplostomum spathaceum cercariae emerged from cadmium-exposed Lymnaea stagnalis. J Helminthol 79:55–59. https://doi.org/10.1079/joh2004271
Morley NJ, Adam ME, Lewis JW (2007) The effects of temperature on the distribution and establishment of Echinoparyphium recurvatum metacercariae in Lymnaea peregra. J Helminthol 81:311–315. https://doi.org/10.1017/S0022149X07818530
Mostafa OMS (2008) Effects of Fasciola gigantica experimental infection on some inorganic elements in the snail host Lymnaea natalensis. Exp Parasitol 118:468–471. https://doi.org/10.1016/j.exppara.2007.10.009
Muñoz-Antoli C, Marín A, Toledo R et al (2007) Effect of Echinostoma friedi (Trematoda: Echinostomatidae) experimental infection on longevity, growth and fecundity of juvenile Radix peregra (Gastropoda: Lymnaeidae) and Biomphalaria glabrata (Gastropoda: Planorbidae) snails. Parasitol Res 101:1663–1670
Neeleman AP, van der Knaap WP, van den Eijnden DH (1994) Identification and characterization of a UDP-GalNAc:GlcNAc beta-R beta-->4-N-acetylgalactosaminyl transferase from cercariae of the schistosome Trichobilharzia ocellata. Catalysis of a key step in the synthesis of N'N-diacetyllactosediamino (lacdiNAc)-type glycans. Glycobiology 4:641–651
Nguyen NT, Le TC, Vo MDC et al (2017) High prevalence of cattle fascioliasis in coastal areas of Thua Thien Hue province, Vietnam. J Vet Med Sci 79:1035–1042. https://doi.org/10.1292/jvms.16-0331
Nie L, Cai SY, Shao JZ et al (2018) Toll-like receptors, associated biological roles, and signaling networks in non-mammals. Front Immunol 9:1523–1542. https://doi.org/10.3389/fimmu.2018.01523
Núñez PE, De Jong-Brink M (1997) The suppressive excretory-secretory product of Trichobilharzia ocellata: a possible factor for determining compatibility in parasite-host interactions. Parasitology 115:193–203. https://doi.org/10.1017/s0031182097001169
Núñez PE, Adema CM, de Jong-Brink M (1994) Modulation of the bacterial clearance activity of phagocytes from the freshwater mollusc, Lymnaea stagnalis, by the avian schistosome, Trichobilharzia ocellata. Parasitology 109:299–310. https://doi.org/10.1017/s0031182000078331
Osorio JB, de Mattos PL, Giongo A et al (2020) Mollusk microbiota shift during Angiostrongylus cantonensis infection in the freshwater snail Biomphalaria glabrata and the terrestrial slug Phillocaulis soleiformis. Parasitol Res 119:2495–2503. https://doi.org/10.1007/s00436-020-06743-y
Pankrác J, Novobilský A, Rondelaud D et al (2016) Effect of Fascioloides magna (Digenea) on fecundity, shell height, and survival rate of Pseudosuccinea columella (Lymnaeidae). Parasitol Res 115:3119–3125. https://doi.org/10.1007/s00436-016-5068-4
Patnaik MM (1968) Notes on glycogen deposits in Lymnaea auricularia var. rufescens and the parasitic larval stages of Fasciola gigantica and Echinostoma revolutum. A histochemical study. Ann Parasitol Hum Comp 43:449–456. https://doi.org/10.1051/parasite/1968434449
Perry KJ, Henry JQ (2014) CRISPR/Cas9-mediated genome modification in the mollusc, Crepidula fornicata. Genesis 53:237–244
Pila EA, Gordy MA, Phillips VK et al (2016a) Endogenous growth factor stimulation of hemocyte proliferation induces resistance to Schistosoma mansoni challenge in the snail host. Proc Natl Acad Sci USA 113:5305–5310
Pila EA, Sullivan JT, Wu XZ et al (2016b) Haematopoiesis in molluscs: a review of haemocyte development and function in gastropods, cephalopods and bivalves. Dev Comp Immunol 58:119–128
Pila EA, Tarrabain M, Kabore AL et al (2016c) A novel toll-like receptor (TLR) influences compatibility between the gastropod Biomphalaria glabrata, and the digenean trematode Schistosoma mansoni. PLoS Pathog 12:e1005513. https://doi.org/10.1371/journal.ppat.1005513
Pila EA, Li H, Hambrook JR et al (2017) Schistosomiasis from a snail’s perspective: advances in snail immunity. Trends Parasitol 33:845–857
Pinaud S, Portela J, Duval D et al (2016) A shift from cellular to humoral responses contributes to innate immune memory in the vector snail Biomphalaria glabrata. PLoS Pathog 12:e1005361. https://doi.org/10.1371/journal.ppat.1005361
Pinheiro J, Maldonado Júnior A, Lanfredi RM (2009) Physiological changes in Lymnaea columella (Say, 1817) (Mollusca, Gastropoda) in response to Echinostoma paraensei Lie and Basch, 1967 (Trematoda: Echinostomatidae) infection. Parasitol Res 106:55–59. https://doi.org/10.1007/s00436-009-1630-7
Plows LD, Cook RT, Davies AJ et al (2004) Activation of extracellular-signal regulated kinase is required for phagocytosis by Lymnaea stagnalis haemocytes. Biochim Biophys Acta 1692(1):25–33. https://doi.org/10.1016/j.bbamcr.2004.03.002
Plows LD, Cook RT, Davies AJ et al (2006a) Integrin engagement modulates the phosphorylation of focal adhesion kinase, phagocytosis, and cell spreading in molluscan defence cells. Biochim Biophys Acta 1763(8):779–786. https://doi.org/10.1016/j.bbamcr.2006.04.008
Plows LD, Cook RT, Davies AJ et al (2006b) Phagocytosis by Lymnaea stagnalis haemocytes: a potential role for phosphatidylinositol 3-kinase but not protein kinase A. J Invert Pathol 91:74–77
Portela J, Duval D, Rognon A et al (2013) Evidence for specific genotype-dependent immune priming in the lophotrochozoan Biomphalaria glabrata snail. J Innate Immunol 5:261–276
Portet A, Toulza E, Lokmer A et al (2021) Experimental infection of the Biomphalaria glabrata vector snail by Schistosoma mansoni parasites drives snail microbiota dysbiosis. Microorganisms 9:1084. https://doi.org/10.3390/microorganisms9051084
Poulin R (2007) Evolutionary ecology of parasites. Princeton University Press, Princeton, NJ
Preston DL, Orlofske SA, Lambden JP et al (2013) Biomass and productivity of trematode parasites in pond ecosystems. J Anim Ecol 82:509–517. https://doi.org/10.1111/1365-2656.12030
Preston DL, Layden TJ, Segui LM et al (2021) Trematode parasites exceed aquatic insect biomass in Oregon stream food webs. J Anim Ecol 90:766–775. https://doi.org/10.1111/1365-2656.13409
Rebelo MF, Figueiredo ES, Mariante RM et al (2013) New insights from the oyster Crassostrea rhizophorae on bivalve circulating hemocytes. PLoS One 8:e57384. https://doi.org/10.1371/journal.pone.0057384
Ribeiro VMA, Coaglio AL, Pereira Oliveira FL et al (2017) Dynamics of haemocytes from Pseudosuccinea columella circulating infected by Fasciola hepatica. Braz J Vet Parasitol 26:411–418
Roldán C, Begovoeva M, López-Olvera J et al (2020) Endemic occurrence of Fasciola hepatica in an alpine ecosystem, Pyrenees Northeastern Spain. Transbound Emerg Dis 68(4):2589–2594. https://doi.org/10.1111/tbed.13865
Rondelaud D, Barthe D (1980a) Etude descriptive d'une réaction amibocytaire chez Lymnaea truncatula Müller infestée par Fasciola hepatica L. Z Parasitenkd 61:187–196. https://doi.org/10.1007/BF00925464
Rondelaud D, Barthe D (1980b) Fasciola hepatica L.: Les formes larvaires non évolutives ou en dégénérescence chez Lymnaea truncatula Müller. Z Parasitenk 62:95–104. https://doi.org/10.1007/bf00925370
Rondelaud D, Barthe D (1981) The development of the amoebocyte-produсing organ in Lymnaea truncatula Müller infected by Fasciola hepatica L. Z Parasitenkd 65:331–341. https://doi.org/10.1007/bf00926728
Rondelaud D, Barthe D (1982) Relationship of the amoebocyte-producing organ with the generalized amoebocytic reaction in Lymnaea truncatula Muller infected by Fasciola hepatica L. J Parasitol 68:967–969
Rondelaud D, Barthe D (1983) Les modifications structurales du rein chez Lymnaea truncatula Muller infestée par Fasciola hepatica. Ann Parasitol Hum Comp 58:109–116
Rondelaud D, Abrous M, Dreyfuss G (2002a) The influence of food sources on cercarial production in Lymnaea truncatula experimentally infected with Digenea. Vet Res 33:95–100
Rondelaud D, Vignoles P, Dreyfuss G (2002b) The presence of predators modifies the larval development of Fasciola hepatica in surviving Lymnaea truncatula. J Helminthol 76:175–178
Rudolfova J, Hampl V, Bayssade-Dufour C et al (2005) Validity reassessment of Trichobilharzia species using Lymnaea stagnalis as the intermediate host. Parasitol Res 95:79–89
Russo J, Lagadic L (2000) Effects of parasitism and pesticide exposure on characteristics and functions of hemocyte populations in the freshwater snail Lymnaea palustris (Gastropoda, Pulmonata). Cell Biol Toxicol 16:15–30
Russo J, Madec L, Brehélin M (2008) Effect of a toxicant on phagocytosis pathways in the freshwater snail Lymnaea stagnalis. Cell Tissue Res 333:147–158. https://doi.org/10.1007/s00441-008-0578-8
Salazar L, Estrada VE, Velásquez LE (2006) Effect of the exposure to Fasciola hepatica (Trematoda: Digenea) on life history traits of Lymnaea cousini and Lymnaea columella (Gastropoda: Lymnaeidae). Exp Parasitol 114:77–83
Sandland GJ, Minchella DJ (2003) Effects of diet and Echinostoma revolutum infection on energy allocation patterns in juvenile Lymnaea elodes snails. Oecologia 134:479–486. https://doi.org/10.1007/s00442-002-1127-x
Sapp KK, Loker ES (2000) Mechanisms underlying digenean-snail specificity: role of miracidial attachment and host plasma factors. J Parasitol 86:1012–1019
Schallig HD, Sassen MJ, Hordijk PL et al (1991) Trichobilharzia ocellata: influence of infection on the fecundity of its intermediate snail host Lymnaea stagnalis and cercarial induction of the release of schistosomin, a snail neuropeptide antagonizing female gonadotropic hormones. Parasitology 102:85–91. https://doi.org/10.1017/s0031182000060376
Schmid-Araya JM, Schmid PE, Majdi N et al (2020) Biomass and production of freshwater meiofauna: a review and a new allometric model. Hydrobiologia 847:2681–2703. https://doi.org/10.1007/s10750-020-04261-7
Schultz JH, Adema CM (2017) Comparative immunogenomics of molluscs. Dev Comp Immunol 75:3–15
Schweizer G, Meli ML, Torgerson PR et al (2007) Prevalence of Fasciola hepatica in the intermediate host Lymnaea truncatula detected by real time TaqMan PCR in populations from 70 Swiss farms with cattle husbandry. Vet Parasitol 150:164–169. https://doi.org/10.1016/j.vetpar.2007.08.006
Selbach C, Poulin R (2020) Some like it hotter: trematode transmission under changing temperature conditions. Oecologia 194:745–755
Seppälä O, Jokela J (2010) Maintenance of genetic variation in immune defense of a freshwater snail: role of environmental heterogeneity. Evolution 64:2397–2407. https://doi.org/10.1111/j.1558-5646.2010.00995.x
Seppälä O, Leicht K (2013) Activation of the immune defence of the freshwater snail Lymnaea stagnalis by different immune elicitors. J Exp Biol 216:2902–2907. https://doi.org/10.1242/jeb.084947
Seppälä K, Leicht K (2015) Quality attracts parasites: host condition-dependent chemo-orientation of trematode larvae. Funct Ecol 29:791–795
Seppälä O, Karvonen A, Kuosa M et al (2013) Are sick individuals weak competitors? Competitive ability of snails parasitized by a gigantism-inducing trematode. PLoS One 8:e79366. https://doi.org/10.1371/journal.pone.0079366
Seppälä O, Louhi KR, Karvonen A et al (2015) Relative reproductive success of co-infecting parasite genotypes under intensified within-host competition. Infect Genet Evol 36:450–455. https://doi.org/10.1016/j.meegid.2015.08.015
Seppälä O, Walser JC, Cereghetti T et al (2021) Transcriptome profiling of Lymnaea stagnalis (Gastropoda) for ecoimmunological research. BMC Genomics 22:144. https://doi.org/10.1186/s12864-021-07428-1
Sindou P, Cabaret J, Rondelaud D (1991) Survival of snails and characteristic lesions of Fasciola hepatica infection in four European species of Lymnaea. Vet Parasitol 40:47–58. https://doi.org/10.1016/0304-4017(91)90082-7
Sire C, Rognon A, Theron A (1998) Failure of Schistosoma mansoni to reinfect Biomphalaria glabrata snails: acquired humoral resistance or intra-specific larval antagonism? Parasitology 117:117–122. https://doi.org/10.1017/s0031182098002923
Skála V, Cerníková A, Jindrová Z et al (2014) Influence of Trichobilharzia regenti (Digenea: Schistosomatidae) on the defence activity of Radix lagotis (Lymnaeidae) haemocytes. PLoS One 9:e111696. https://doi.org/10.1371/journal.pone.0111696
Skála V, Walker A, Horák P (2018) Extracellular trap-like fiber release may not be a prominent defence response in snails: evidence from three species of freshwater gastropod molluscs. Dev Comp Immunol 79:137–141. https://doi.org/10.1016/j.dci.2017.10.011
Sminia T (1974) Haematopoiesis in the freshwater snail Lymnaea stagnalis studied by electron microscopy and autoradiography. Cell Tissue Res 150:443–454
Sminia T, Borghart-Reinders E, van de Linde AW (1974) Encapsulation of foreign materials experimentally introduced into the freshwater snail Lymnaea stagnalis. An electron microscopic and autoradiographic study. Cell Tissue Res 153:307–326
Sminia T, van der Knaap WP, Edelenbosch P (1979) The role of serum factors in phagocytosis of foreign particles by blood cells of the freshwater snail Lymnaea stagnalis. Dev Comp Immunol 3:37–44
Smit AB, De Jong-Brink M, Li KW et al (2004) Granularin, a novel molluscan opsonin comprising a single vWF type C domain is up-regulated during parasitation. FASEB J 18:845–847. https://doi.org/10.1096/fj.03-0590fje
Soldánová M, Selbach C, Sures B et al (2010) Larval trematode communities in Radix auricularia and Lymnaea stagnalis in a reservoir system of the Ruhr River. Parasite Vector 3:56. https://doi.org/10.1186/1756-3305-3-56
Soldánová M, Faltýnková A, Scholz T et al (2011) Parasites in a man-made landscape: contrasting patterns of trematode flow in a fishpond area in Central Europe. Parasitology 138:789–807. https://doi.org/10.1017/s0031182011000291
Soldánová M, Kuris AM, Scholz T et al (2012) The role of spatial and temporal heterogeneity and competition in structuring trematode communities in the great pond snail, Lymnaea stagnalis (L.). J Parasitol 98:460–471. https://doi.org/10.1645/GE-2964.1
Soliman MF (2009) Fasciola gigantica: cercarial shedding pattern from Lymnaea natalensis after long-term exposure to cadmium at different temperatures. Exp Parasitol 121:307–311
Sorensen RE, Minchella DJ (1998) Parasite influences on host life history: Echinostoma revolutum parasitism of Lymnaea elodes snails. Oecologia 115:188–195. https://doi.org/10.1007/s004420050507
Sorensen RE, Minchella DJ (2001) Snail-trematode life history interactions: past trends and future directions. Parasitology 123:S3–S8. https://doi.org/10.1017/s0031182001007843
Southgate VR (1970) Observations on the effect of the rediae of Fasciola hepatica on the lipid composition of the hepatopancreas of Lymnaea truncatula. Parasitology 61:293–299. https://doi.org/10.1017/s0031182000041123
Thèron A, Coustau C (2005) Are Biomphalaria snails resistant to Schistosoma mansoni? J Helminthol 79:189–191
Thieltges DW, Jensen KT, Poulin R (2008) The role of biotic factors in the transmission of free-living endohelminth stages. Parasitology 135:407–426
Travers MA, Mirella da Silva P, Le Go N et al (2008) Morphologic, cytometric and functional characterisation of abalone (Haliotis tuberculata) haemocytes. Fish Shellfish Immunol 24:400–411
Tunholi VM, Lorenzoni PO, da Silva YH et al (2017) Molluscicidal potential of Heterorhabditis baujardi (Rhabditida: Heterorhabditidae), strain LPP7, on Lymnaea columella (Gastropoda: Pulmonata): an alternative for biological control of fasciolosis. Acta Trop 173:23–29
Utzinger J, Tanner M (2000) Microhabitat preferences of Biomphalaria pfeifferi and Lymnaea natalensis in a natural and a man-made habitat in southeastern Tanzania. Mem Inst Oswaldo Cruz 95:287–294
van der Knaap WP, Loker ES (1990) Immune mechanism in trematode – snail interactions. Parasitol Tod 6:175–182
van der Knaap WP, Sminia T, Kroese FG et al (1981) Elimination of bacteria from the circulation of the pond snail Lymnaea stagnalis. Dev Comp Immunol 5:21–32
van der Knaap WP, Boots AMH, van Asselt LA et al (1983a) Specificity and memory in increased defence reactions against bacteria in the pond snail Lymnaea stagnalis. Dev Comp Immunol 7:435–443
van der Knaap WP, Sminia T, Schutte R et al (1983b) Cytophilic receptors for foreignness and some factors which influence phagocytosis by invertebrate leucocytes: in vitro phagocytosis by amoebocytes of the snail Lymnaea stagnalis. Immunology 48:377–383
van der Knaap WP, Meuleman EA, Sminia T (1987) Alterations in the internal defence system of the pond snail Lymnaea stagnalis induced by infection with the schistosome Trichobilharzia ocellata. Parasitol Res 73:57–65. https://doi.org/10.1007/BF00536337
Vaughn CM (1953) Effects of temperature on hatching and growth of Lymnaea stagnalis appressa Say. Am Midland Nat 49:214–228
Vázquez AA, Hevia Y, Sánchez J (2009) Distribución y preferencia de hábitats de moluscos hospederos intermediarios de Fasciola hepatica en Cuba. Rev Cubana Med Trop 61:248–253
Vázquez AA, Sánchez J, Pointier JP et al (2014) Fasciola hepatica in Cuba: compatibility of different isolates with two intermediate hosts, Galba cubensis and Pseudosuccinea columella. J Helminthol 88:434–440. https://doi.org/10.1017/s0022149x13000382
Vázquez AA, Pilar A, Lounnas M et al (2018) Lymnaeid snails hosts of Fasciola hepatica and Fasciola gigantica (Trematoda: Digenea): a worldwide review. CAB Rev 13:1–15. https://doi.org/10.1079/PAVSNNR201813062
Vázquez A, Vargas M, Alba A et al (2019) Reviewing Fasciola hepatica transmission in the West Indies and novel perceptions from experimental infections of sympatric vs. allopatric snail/fluke combinations. Vet Parasitol 275:108955. https://doi.org/10.1016/j.vetpar.2019.108955
Vieira TC, Costa-Filho A, Salgado NC et al (2004) Acharan sulfate, the new glycosaminoglycan from Achatina fulica Bowdich 1822. Structural heterogeneity, metabolic labeling and localization in the body, mucus and the organic shell matrix. Eur J Biochem 271:845–854
Vorontsova YL, Slepneva IA, Yurlova NI et al (2019) The effect of trematode infection on the markers of oxidative stress in the offspring of the freshwater snail Lymnaea stagnalis. Parasitol Res 118(12):3561–3564. https://doi.org/10.1007/s00436-019-06494-5
Voutilainen A (2010) Interactive effects of predation risk and parasitism on the circadian rhythm of foraging activity in the great pond snail Lymnaea stagnalis (Gastropoda: Lymnaeidae). Ann Limnol Int J Lim 46:217–223. https://doi.org/10.1051/limn/2010025
Voutilainen A, van Ooik T, Puurtinen M et al (2009) Relationship between prevalence of trematode parasite Diplostomum sp. and population density of its snail host Lymnaea stagnalis in lakes and ponds in Finland. Aquat Ecol 43:351–357. https://doi.org/10.1007/s10452-008-9203-x
Walker SM, Makundi AE, Namuba FV et al (2008) The distribution of Fasciola hepatica and Fasciola gigantica within southern Tanzania – constraints associated with the intermediate host. Parasitology 135:495–503
Walker AJ, Lacchini AH, Sealey KL et al (2010) Spreading by snail (Lymnaea stagnalis) defence cells is regulated through integrated PKC, FAK and Src signaling. Cell Tissue Res 341:131–145
Wang Y, Hu M, Chiang MW et al (2012) Characterization of subpopulations and immune-related parameters of hemocytes in the green-lipped mussel Perna viridis. Fish Shellfish Immunol 32:381–390
Wang C, Yue X, Lu X et al (2013) The role of catalase in the immune response to oxidative stress and pathogen challenge in the clam Meretrix meretrix. Fish Shellfish Immunol 34:91–99
Wilson RA, Denison J (1970a) Short-chain fatty acids as stimulants of turning activity by the miracidium of Fasciola hepatica. Comp Biochem Physiol 32:511–517
Wilson RA, Denison J (1970b) Studies on the activity of the miracidium of the common liver fluke, Fasciola hepatica. Comp Biochem Physiol 32:301–313
Wilson RA, Denison J (1980) The parasitic castration and gigantism of Lymnaea truncatula infected with the larval stages of Fasciola hepatica. Z Parasitenk 61:109–119. https://doi.org/10.1007/bf00925458
Wilson RA, Draskau T (1976) The stimulation of daughter redia production during the larval development of Fasciola hepatica. Parasitology 72:245–257. https://doi.org/10.1017/s0031182000049465
Wolinska J, King KC (2009) Environment can alter selection in host-parasite interactions. Trends Parasitol 25:236–244
Wright B, Lacchini AH, Davies AJ et al (2006) Regulation of nitric oxide production in snail (Lymnaea stagnalis) defence cells: a role for PKC and ERK signalling pathways. Biol Cell 96:265–278
Yurlova NI, Vodyanitskaya SN, Serbina EA et al (2006) Temporal variation in prevalence and abundance of metacercariae in the pulmonate snail Lymnaea stagnalis in Chany Lake, West Siberia, Russia: long-term patterns and environmental covariates. J Parasitol 92:249–259. https://doi.org/10.1645/ge-544r2.1
Zakikhani M, Rau ME (1999) Plagiorchis elegans (Digenea: Plagiorchiidae) infections in Stagnicola elodes (Pulmonata: Lymnaeidae): host susceptibility, growth, reproduction, mortality, and Cercarial production. J Parasitol 85:454–463
Żbikowska E (2003) The effect of Digenea larvae on calcium content in the shells of Lymnaea stagnalis (L.) individuals. J Parasitol 89:76–79
Żbikowska E (2011) One snail – three Digenea species, different strategies in host-parasite interaction. Anim Biol 61:1–19. https://doi.org/10.1163/157075511X554383
Żbikowska E, Żbikowski J (2005) Differences in shell shape of naturally infected Lymnaea stagnalis (l.) individuals as the effect of the activity of digenetic trematode larvae. J Parasitol 91:1046–1051
Żbikowska E, Żbikowski J (2015) Digenean larvae—the cause and beneficiaries of the changes in host snails’ thermal behavior. Parasitol Res 114:1063–1070
Żbikowska E, Kobak J, Żbikowski J et al (2006) Infestation of Lymnaea stagnalis by digenean flukes in the Jeziorak Lake. Parasitol Res 99:434–439. https://doi.org/10.1007/s00436-006-0178-z
Zelck UE, Janje B, Schneider O (2005) Superoxide dismutase expression and H2O2 production by hemocytes of the trematode intermediate host Lymnaea stagnalis (Gastropoda). Dev Comp Immunol 29:305–314
Zelck UE, Gege BE, Schmid S (2007) Specific inhibitors of mitogen-activated protein kinase and PI3-K pathways impair immune responses by hemocytes of trematode intermediate host snails. Dev Comp Immunol 31:321–331. https://doi.org/10.1016/j.dci.2006.06.006
Zilber-Rosenberg I, Rosenberg E (2008) Role of microorganisms in the evolution of animals and plants: the hologenome theory of evolution. FEMS Microbiol Rev 32:723–735
Zimmermann MR, Luth KE, Esch GW (2014) Differences in snail ecology lead to infection pattern variation of Echinostoma spp. larval stages. Acta Parasitol 59:502–509
Zischke DP (1972) The effects of Echinostoma revolutum larval infection on the histology, growth and fecundity of the snail, Stagnicola palustris. PhD Thesis, Louisiana State University and Agricultural and Mechanical College, LSU Historical Dissertations and Theses
Author information
Authors and Affiliations
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2023 The Author(s), under exclusive license to Springer Nature Switzerland AG
About this chapter

Cite this chapter
Alba, A., Gourbal, B. (2023). Biology of the Lymnaeidae–Parasite Interaction. In: Vinarski, M.V., Vázquez, A.A. (eds) The Lymnaeidae. Zoological Monographs, vol 7. Springer, Cham. https://doi.org/10.1007/978-3-031-30292-3_11
Download citation
DOI: https://doi.org/10.1007/978-3-031-30292-3_11
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-031-30291-6
Online ISBN: 978-3-031-30292-3
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)