
Abstract
Hypophysiotropic neurons of the paraventricular nucleus of the hypothalamus that express thyrotropin-releasing hormone (TRH) control the synthesis and release of thyrotropin, the pituitary hormone that regulates the synthesis and release of thyroid hormones. Thyroid hormones are pleiotropic hormones with multiple functions involved in growth, development, and energy homeostasis. The TRH neuroendocrine cells receive neuronal inputs from different parts of the brain, as well as local and hormonal signals, and integrate and transduce the information as a hormone (TRH) output. They are glutamatergic neurons; most co-express cocaine- and amphetamine-regulated transcripts; however, their transcriptomic characterization is still in its infancy but suggests functional diversity. Transcription of the Trh gene is rapidly but transiently increased by multiple signals, some of which also cause the release of TRH. This review summarizes the basic mechanisms involved in the generation of TRH in hypophysiotropic neurons and turnover in median eminence, and recapitulates the multiple factors that regulate Trh synthesis and the amount of TRH that reaches thyrotropes, the physiological conditions and environmental stressors that alter TRH neurons and thyroid axis status during development and in adult animals, as well as critical sex differences.
Disclosure statement: The authors have nothing to disclose.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
1 Introduction
Although evidence of hypothalamic control of anterior pituitary hormones began to accumulate in the 1940s, it was not until 1969 that the structure of a hypothalamic factor that induces the secretion of thyrotropin was solved (reviewed in Joseph-Bravo et al. 2015a); it received the name thyrotropin-releasing hormone (TRH) (Boler et al. 1969; Burgus et al. 1969). The hypothalamus has long been considered the center of homeostasis. Localized at the base of the brain, it contains nuclei that contact many areas in the brain; among these hypothalamic nuclei, the paraventricular nucleus (PVN) contains neurons whose axon terminals do not make synaptic contacts with other neurons but instead approximate fenestrated capillaries. Some of the neurons with a small somatic size, called parvocellular, project their axons to the median eminence, where they secrete releasing or inhibiting factors near portal vessels that transport them to the anterior pituitary to control synthesis and release of adenohypophysial hormones; they are called hypophysiotropic neurons (Watts 2015). Hypophysiotropic PVN neurons that express TRH control the synthesis and release of thyrotropin (TSH), the pituitary hormone that regulates the synthesis and release of thyroid hormones (TH, T4, and T3) from the thyroid gland. These neuroendocrine cells receive neuronal inputs from different parts of the brain, as well as local and hormonal signals, and transduce the integrated information as a hormone (TRH) output that controls the function of the hypothalamic-pituitary-thyroid (HPT) axis (Fig. 10.1). To study the hypophysiotropic TRH neurons it is necessary to consider the physiology of the HPT axis, including the efficient feedback mechanisms common to all neuroendocrine axes, provided by the hormones released by the target organ (in this case T4 and T3) that maintain axis homeostasis (Fekete and Lechan 2014; Hoermann et al. 2015; Joseph-Bravo et al. 2015a, b). In this chapter, we review the mechanisms that determine TRH metabolism and the activity of the hypophysiotropic TRH neurons, including findings that appeared after recent comprehensive reviews (Joseph-Bravo et al. 2015a, b, 2016; Chatzitomaris et al. 2017; Rodríguez-Rodríguez et al. 2019; Charli et al. 2020).

Hypophysiotropic TRH neurons regulate the hypothalamic-pituitary-thyroid axis. In rats, hypophysiotropic TRH neurons are localized in mid-caudal PVN and receive projections from two leptin-responsive neuronal populations located in the ARC (αMSH/CART and NPY/AgRP), from the suprachiasmatic nucleus, and from adrenergic afferents from the brainstem. Hypophysiotropic TRH neurons express αMSH receptor (MC4R) and NPY receptors (Y1, Y5). Nerve terminals of hypophysiotropic TRH neurons reside in the ME, from where TRH is released next to portal vessels that transport it to the pituitary. and stimulates synthesis and release of TSH; TSH regulates in the thyroid synthesis and secretion of T4; T4 is partially deiodinated to T3 in thyroid and in target tissues. TH processing by deiodinases and their chemical structure are depicted on the left; light blue circles represent iodine atoms. Figure created with BioRender.com, modified from Joseph-Bravo et al. (2015b, 2017)
2 Hypophysiotropic TRH Neurons
TRH is a peptide formed of three amino acids with the NH and COOH terminal ends modified: pyroglutamyl-histidyl-prolinamide. Soon after its discovery (detailed in Joseph-Bravo et al. 2015a), specific antibodies recognizing TRH allowed its detection by immunohistochemistry in nerve terminals of the median eminence, as well as in neurons of the PVN, various nuclei of the hypothalamus, brain regions, and spinal cord; it was necessary to use colchicine to prevent axonal transport and observe the peptide in cell bodies (Lechan and Jackson 1982). Elucidating the mode of TRH synthesis was a challenge that was overcome by the use of recombinant DNA techniques. These permitted characterization of the mRNA sequence encoding the protein precursor of TRH (proTRH) (Lechan et al. 1986) and the identification of the gene (Lee et al. 1988). These discoveries led to complementary immunohistochemical approaches with proTRH antibodies, and in situ hybridization with nucleotide probes against proTRH mRNA, which revealed rat PVN parvocellular and some magnocellular cell bodies containing Trh mRNA/proTRH in the PVN and in neurons of other hypothalamic nuclei (Lechan et al. 1987).
The neuroanatomical localization of TRH hypophysiotropic neurons was evidenced by the systemic administration of fluorogold into rats. The tracer was taken up by the median eminence and transported retrogradely to neurons in the PVN that express Trh mRNA (Fekete et al. 2000). Hypophysiotropic TRH neurons are thus the parvocellular cells localized in the mid and caudal zones of rat PVN. In mice, TRH neurons are localized only in the middle zone of PVN (Kádár et al. 2010). Within the mid and caudal areas of the PVN, there are also parvocellular neurons that are not hypophysiotropic, though their efferents have not been characterized. In rats, the anterior or rostral PVN contains a population of parvocellular TRH neurons that send projections into various hypothalamic and extrahypothalamic nuclei (Wittmann et al. 2009). TRH is also expressed in a few magnocellular neurons located in the mid-PVN. These neurons project to the neurohypophysis where TRH modulates oxytocin and vasopressin release (Ciosek and Izdebska 2009). This heterogeneity has to be taken into account whenever data from the whole PVN are obtained.
Additional neurotransmitters have been characterized in TRH neurons of the PVN. Eighty percent of hypophysiotropic TRH neurons express cocaine- and amphetamine-activated transcript (CART) in the rat PVN (Fekete et al. 2000), and many (not quantified) in the mouse PVN (Kádár et al. 2010). Vesicular glutamate transporter 2 is expressed in the hypophysiotropic TRH neurons, suggesting that these neurons use glutamate as neurotransmitter (Hrabovszky et al. 2005; Farkas et al. 2020). Several other peptides and neurotransmitters have been reported in parvocellular TRH neurons (Table 10.1), but a careful assignment to hypophysiotropic or non-hypophysiotropic type is lacking (Table 10.1). The variable match between parvocellular TRH neurons and some neuronal inputs (Table 10.2) suggests heterogeneity of TRH hypophysiotropic neurons, with subsets being responsive to specific stimuli (Sánchez et al. 2001). The potential heterogeneity and limited information about the proteome of hypophysiotropic TRH neurons complicates the adscription of a specific Trh cluster from published transcriptomes (Box 10.1, Fig. 10.2) (Chen et al. 2017; Zeisel et al. 2018) to hypophysiotropic TRH neurons. Furthermore, a discrepancy between the TRH cluster ascribed to the PVN in the study of Zeisel et al. 2018 and the chemical identity of hypophysiotropic neurons based on immunohistochemistry and in situ hybridization warrants further study.

Single-cell RNA sequencing provides a large quantity of data from a few samples; diverse types of hypothalamic TRH neurons have been identified. (a) To identify multiple types of cells: (1) Hypothalamus is dissected and cells disaggregated. (2) Each cell is conjugated with a barcoded bead and cDNA libraries are generated in situ. (3) Beads are subjected to next-generation DNA sequencing to obtain a matrix of gene counts per cell. (4) Each cell is represented as a vector in a space with multiple dimensions (each gene corresponds to one dimension). Dimensionality is reduced by computational analysis, resulting in grouping of cells with similar expression patterns along two components. Each group of cells or cluster represents a particular cell type or status. Clusters can be refined by subsequent analysis . (5) Cell identity and location can be confirmed with a subsequent analysis of markers with highest expression for each cluster and in situ hybridization experiments. (b) Chen et al. (2017) detected six Trh expressing clusters; violin plots refer to gene expression in log2 of the transcript count per million along the x-axis. Zeisel et al. (2018) identified three clusters of Trh expressing neurons; circles radius represent the expression value relative to peptidergic glutamatergic clusters; they also mapped the likely location in the hypothalamus with global transcriptome expression and Allen Mouse Brain Atlas. Data to generate figure b were obtained from two public datasets: Gene Expression Omnibus, with accession number GSE87544, and from curated data downloaded from http://mousebrain.org/downloads.html. Data were filtered using dplyr in R and LoomPy in an instance of JupyterLab. Graphs were made using ggplot2 library in R
Box 10.1. Can Non-spatial Information Provide Spatial Information About Molecular Marker Expression in the Hypothalamus? Retrieving Spatial Information with New Generation Tools
The brain is one of the organs with the most cellular diversity per unit volume. Obtaining spatial definition is always a challenge, and this is not an exception for the paraventricular nucleus of the hypothalamus, a nucleus that shows marked morphological differences in its neuronal populations, along with the expression of a large repertoire of neuropeptides. This information has been retrieved for many years with classical techniques as immunohistochemistry and in situ hybridization, methods that allow a high spatial definition of the molecular architecture. The incorporation of diverse fluorescent markers allowed the detection of more than one molecular marker from the same cell and subject. With the accessibility of new technologies, such as powerful computers and practically unlimited data-storage units, hypothalamic investigations are starting to take advantage of bioinformatics and sequencing technologies, delivering massive amounts of information. This is the case for single-cell RNA sequencing (scRNAseq) (Fig. 10.2a), a technique that allowed more information about cell expression in the hypothalamus to be obtained than all the information gathered in the past ~60 years. scRNAseq shows that TRH neuron phenotypes of the hypothalamus are diverse (Fig. 10.2b). With this technique, a high level of mRNA counts per cell can be retrieved, surpassing classical histological methods allowing the detection of heterogeneity among individual cells. However, many cells (and their transcriptome) may be underrepresented, as just a small percentage of the cells are sampled, and the final decision about cell types (as clusters) depends on how the clustering algorithms are refined with different information, such as previous findings and brain expression maps. It is likely that soon enough a detailed map of the hypothalamic cell types will be prepared based on transcriptomics.
3 Setting the Concentration of TRH That Reaches the Anterior Pituitary; a Multi-level Task
3.1 TRH-Gene Transcription
The Trh gene has been sequenced in several species and the consensus sequences of response elements to various transcription factors have been identified in the promoter region (Hollenberg et al. 1995; Díaz-Gallardo et al. 2010a, b; Cote-Vélez et al. 2011; Guo et al. 2004) (Fig. 10.3). Initial work centered on recognizing the binding sites for thyroid hormone receptors (THR), to explain the negative feedback TH exerts on the axis; TH inhibits Trh transcription in vivo, exclusively on hypophysiotropic neurons of the PVN (Dyess et al. 1988; Sugrue et al. 2010). Due to the low concentrations of receptors in cells, and even lower concentrations of their transcripts, the mode of synthesis was first studied with cells transfected with Thr and Trh-gene promoter linked to a reporter such as luciferase; of the two receptors THRα and THRβ, products of independent genes, THRβ binds either as a monomer, homodimer or heterodimer with retinoid acid receptor X (RXR), to the DNA sequence called Site 4, containing half of a TH-response element (THRE), previously identified in other T3-regulated genes; increased transcription was detected in absence of T3 but repressed in its presence (Hollenberg et al. 1995). Studies with KO mice allowed NCoR to be defined as the nuclear repressor involved (Astapova and Hollenberg 2013), and the isoform THRβ2 as the one responsible for feedback (Abel et al. 2001). The model of unliganded THR bound to DNA having an opposite effect to that when THR is bound to T3 is now questioned, since in transfected cells reporter transcription lacks the constraints of chromatin, and transfection can generate receptor concentrations higher than that normally found in tissues. More sensitive methodologies such as chromatin immunoprecipitation-DNA sequencing (Nakato and Sakata 2020) have recently been applied to liver of hypo- and hyperthyroid mice; various THRE with different affinities for THRβ1 (the receptor found in liver) and for either T3-positively or -negatively regulated genes have been identified together with chromatin remodeling changes (Ramadoss et al. 2014; Grøntved et al. 2015). An analysis of the problems encountered can be found in various articles (Vella and Hollenberg 2017; Flamant et al. 2017; Sasaki et al. 2018).

Regulation of Trh transcription and proTRH processing. (A) Schematic representation of a TRH hypophysiotropic neuron. Identified receptors are depicted in the membrane, and some of the regulatory elements that modulate Trh transcription (mentioned in Sect. 10.3.1) are marked in the design of the Trh-promoter region. During translation and ER translocation, the signal sequence (red) of preproTRH is cleaved. ProTRH undergoes processing by PC1/3, PC2, CPE, QC and PAM to TRH (Sect. 10.3.2). TRH is released into the extracellular space in a calcium-dependent manner. Figure modified from Joseph-Bravo et al. (2016), Perello and Nillni (2007), created with BioRender.com. (B) Co-expression pSTAT3 and Trh mRNA in neurons at the parvocellular mid-level of the PVN of an adult male control rat, revealed by a combination of radioactive in situ hybridization (for Trh mRNA) and chromogenic immunohistochemistry (for pSTAT3) on rat brain slices (Bregma -1.9; Paxinos and Watson 2004). (a) Dark field 5X micrograph of Trh mRNA-positive neurons (white dots). (b) Bright field 5× micrograph of pSTAT3-positive cells (brown staining). (c) 40X magnification of rectangle in b); white arrow: pSTAT3-positive cell (brown color); black arrow: co-expression of Trh mRNA (silver grains) and pSTAT3; empty arrow: pSTAT3 negative Trh mRNA-positive neuron
Findings on liver samples with THRβ1 still might not be representative of regulation of TRH in PVN or TSH in adenohypophysis, since THRβ2 is the isoform involved in the negative feedback of Trh and Tshb transcription, and recent studies have identified three aminoacids present in helix 10 of THRβ2 that are responsible for the formation of a homodimer, fully functional for transcription repression of Tshb (Pinto et al. 2017). This supports chromatin-immunoprecipitation analyses performed on stable cell lines expressing Trh, showing that T3 promotes THRβ and histone-deacetylase 3 (HDAC3) binding to Trh promoter; binding of HDAC3 was transient (15–60 min) (Ishii et al. 2004). Similar experiments with primary cultures of rat hypothalami showed THRβ2 and HDAC2 bound to a fragment of Trh promoter containing Site 4 (–242/+34). It remains to be determined whether chromatin compaction due to deacetylation expels the receptor and other factors from chromatin (Díaz-Gallardo et al. 2010a; Ishii et al. 2004; Sotelo-Rivera et al. 2017). Recently, the relevance of THR binding to Site 4, and to an equivalent site in Tshb, has been questioned since these are near the transcription start site; instead, it is proposed that binding of transcription factor GATA2 to site –357/–352 increases Trh transcription, and that THRβ2-bound T3 binds to GATA2 in a tethering fashion, repressing Trh transcription (Kuroda et al. 2020, Fig. 10.3). Thus, the whole picture of how T3 regulates Trh transcription remains incomplete.
Other response elements that bind transcription factors that respond to extra- and intra-cellular signals are depicted in Fig. 10.3. Foot-printing analyses demonstrated that in primary hypothalamic neurons cAMP analogs promote nuclear proteins binding to an extended area that includes cAMP, Krüppel, KEM and SP-1 recognition elements (Díaz-Gallardo et al. 2010a, b; Cote-Vélez et al. 2011; Pérez-Monter et al. 2011). Upstream of this extended CRE the binding site for STAT3 has been detected (Guo et al. 2004). The element that binds glucocorticoid receptor (GR) is a half site that is stabilized as a heterodimer with c-JUN or c-FOS; this site is called composite GRE and its activity depends on the bound partner (Díaz-Gallardo et al. 2010b; Cote-Vélez et al. 2008). Furthermore, various transcription factors may form heterodimers with THR like RXR, LXR, to name a few. The multiple combinations of transcription factors able to bind THRE, CRE, STAT, or GRE sites are likely to contribute to the multifactorial regulation of Trh transcription (Joseph-Bravo et al. 2015b, 2016; Kouidhi and Clerget-Froidevaux 2018).
3.2 Pre-pro-TRH Translation and Processing
The primary transcript, a heterologous RNA (2.6 Kb) that contains three exons and two introns, is processed to a mature mRNA that encodes preproTRH (Lechan et al. 1986). As for all secretory proteins, after translation and translocation into the lumen of the endoplasmic reticulum, the propeptide is transported to the Golgi apparatus where it may be cleaved and sorted into secretory granules in the trans-Golgi (Fig. 10.3) (Perello and Nillni 2007). The sequence glutamine-histidine-proline-glycine that leads to TRH is repeated five times in rat and six times in human proTRH; processing enzymes convert it to pGlu-His-ProNH2, which is the active form. All enzymes and cofactors are inside the secretory granule and processing continues inside it during its transport through the axon; the processed peptides are enriched at the nerve terminal. In contrast to the extensive knowledge of POMC tissue-specific processing (Cawley et al. 2016; Harno et al. 2018), the functions of the diverse cryptic peptides derived from TRH precursors have been only partially characterized (Nillni 2010).
The expression of processing enzymes is regulated by many factors, including TH (Nillni 2010) and the concentration of Cu++ ions, which are a cofactor of the enzyme peptidyl glycine alpha-amidating monooxygenase (PAM) (Giraud et al. 1992). Furthermore, processing of neuropeptides can be affected by altered calcium homeostasis or REDOX state at the level of the endoplasmic reticulum (stressed ER), disrupting the adequate folding of the precursors necessary for their transport to the trans-Golgi; for example, fasting or increased inflammatory markers induce changes in ER molecules involved in folding; their effects have been demonstrated, for example, in an inadequate processing of POMC, decreasing αMSH levels (Cakir and Nillni 2019).
3.3 Signals on Hypophysiotropic TRH Neurons Soma Regulate Synthesis and Release of TRH
Immunocytochemical analyses have revealed nerve terminals containing diverse neurotransmitters and neuropeptides in close apposition to the cell body of TRH neurons, suggesting a rich innervation (see Table 10.2), although it is not always clear that they make contact on hypophysiotropic neurons; use of CART coexistence could aid this characterization (Fekete et al. 2000). Furthermore, the expression of neurotransmitter or peptide receptor(s) in the TRH neurons suggest a functional interaction. For example, αMSH nerve terminals contact TRH neurons in the PVN (Fekete et al. 2000), and TRH neurons express melanocortin receptor 4 (MC4R) (Fekete and Lechan 2014). Nerve terminals containing other Arc peptides such as NPY or AgRP (Fekete and Lechan 2014) are observed on TRH cell bodies in the PVN and the administration of these peptides in appropriate physiological conditions modifies Trh expression (Fekete and Lechan 2014). These somatic inputs have the potential to modify the rate of action potential generation by the TRH neurons, although this has been seldom studied. One of the few studies that monitored the spiking activity of the TRH neurons demonstrated a rapid inhibitory effect of glucocorticoids, mediated by endocannabinoid-induced inhibition of glutamate release from neurons contacting parvocellular TRH neurons in the PVN (Di et al. 2003). Most evidence about TRH neurons activity is indirect, either being gained in vitro or using serum TSH concentration as a surrogate measure of TRH release. The in vitro data suggest TRH is released by exocytosis, in response to action potentials that open voltage-dependent Ca2+ channels (Joseph-Bravo et al. 1979). The coexistence of neuropeptides from distinct precursors in hypophysiotropic TRH neurons (including CART), as well as the existence of two different types of secretory granule containing TRH from the first precursor cleavage (differing in cryptic peptides) (Perello and Nillni 2007), leave unanswered questions as to how secretion is regulated. Action potential frequency dependency differs for secretory granules containing peptides and for neurotransmitter vesicles (van den Pol 2012) but, the characteristics of secretory granules exocytosis differing in peptide content are still unknown, which opens many questions regarding their function.
3.4 In the Median Eminence, Hypophysiotropic TRH Varicosities/Nerve Endings and Tanycytes Form a Functional Unit That Controls TRH Output into Portal Vessels
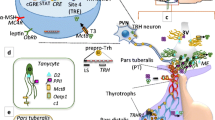
In the external layer of the median eminence, nerve varicosities and terminals of TRH neurons are localized next to other nerve terminal types, cells of the portal capillary vessels and various glial cell types including tanycytes (Fig. 10.4a, Box 10.2) (Müller-Fielitz et al. 2017; Rodríguez-Rodríguez et al. 2019). Tanycytes express TH transporters, deiodinase 2 (DIO2) and TRH-degrading ectoenzyme (TRH-DE), important elements in regulating HPT axis activity (Fig. 10.4b). In the median eminence, T4 transported from the circulation into tanycytes is deiodinated to T3 by DIO2 (Fekete and Lechan 2014) and T3 is transported to the extracellular compartment of the median eminence. T3 feedback effect on TRH synthesis is proposed to be due to T3 captured by TRH nerve terminals and retrograde axonal transport to the nucleus at the level of the cell body where it inhibits transcription (Fekete and Lechan 2014). β2-tanycytes may also control the amount of TRH released from nerve terminals, based on the activity of TRH neurons. Optogenetic activation of TRH neurons in the median eminence promotes glutamate release, which enhances intracytosolic calcium concentration and diacylglycerol lipase α activity (DAGLα) in tanycytes leading to the synthesis of 2-arachinodonoylglycerol (2-AG), which feedbacks onto TRH neurons through cannabinoid receptors 1 (CB1R), thereby inhibiting TRH release (Fig. 10.4b; Farkas et al. 2020).

End-feet of tanycytes of the median eminence control TRH release and extracellular turnover. (a) Photomicrograph showing the alignment of vimentin-expressing tanycytes of the ME along the 3V and immunodetection of TRH in terminal buttons reaching the external zone of the ME, a region irrigated by fc that allow TRH transport to the anterior pituitary. (b) Diagram of elements involved in controlling TRH release and turnover in the external layer of the ME. Released TRH released into the extracellular space may reach the capillary vessels and travel to the adenohypophysis, or it may interact with TRH-R1 localized on β2-tanycytes and increase the activity of TRH-DE, which inactivates TRH, and modify the position of tanycyte end feet, limiting TRH diffusion. In addition, when Glu, a co-transmitter in TRH neurons, binds to GluR in tanycytes, it increases the cytosolic Ca2+ concentration and the activity of DAGLα, leading to the synthesis of 2-AG. 2-AG feeds back onto TRH neurons through CB1R and inhibits TRH release. The amount of TRH that reaches anterior pituitary thyrotrophs may also be reduced by the activity of thyroliberinase, which is present in the portal capillary. Figure created with BioRender.com
TRH-DE is an ectopeptidase with high specificity towards TRH, present on the surface of tanycytes, that cleaves the pyroglutamyl residue and inactivates TRH in the extracellular space after its release from nerve terminals of TRH neurons. Although the portal capillaries are fenestrated (they have transcellular pores that allow diffusion of macromolecules, including TRH released from nerve terminals), the amount of TRH that enters the lumen of the portal vessels is likely to be limited by TRH-DE activity. T3 enhances Trhde expression, probably contributing to a decrease in serum TSH concentration (Sánchez et al. 2009). In addition, TRH released into the median eminence interacts with TRH-receptor 1 (TRH-R1) also localized on β2-tanycyte surface and increases the activity of TRH-DE in tanycytes and the size of tanycyte end-feet, likely reducing TRH flux into the portal vessels (Müller-Fielitz et al. 2017).
Therefore, both the somatic and terminal compartments of hypophysiotropic TRH neurons dictate TRH output into portal capillaries. Furthermore, TRH output may also be further reduced by the activity of a serum isoform of TRH-DE (thyroliberinase) in the portal circulation, which is regulated by multiple factors (Charli et al. 2020).
Box 10.2. Tanycytes
Brain ventricles are lined with a simple layer of ependymocytes that allow a free exchange of molecules between cerebrospinal fluid and parenchymal tissue (neurons and other glial cells). However, selected portions of the basal medial hypothalamus have a modified type of ependymal cell with an elongated cell body whose shape resembles that of radial glia. These glial cells are called tanycytes; in contrast to ependymocytes, tanycytes are joined through tight junctions that limit the passage of substances from brain parenchyma to cerebrospinal fluid by pericellular diffusion. Cytoplasmic tanycyte extensions establish contact with neurons of nearby hypothalamic nuclei or basement membranes surrounding capillaries; various subtypes are distributed according to their dorsoventral position in the third ventricle wall. The most basic classification divides tanycytes into four subtypes: α1 tanycytes, adjacent to the ventromedial nucleus of the hypothalamus and part of the dorsomedial nucleus; α2 tanycytes, which mostly contact the arcuate nucleus parenchyma; β1 tanycytes, which localize to the lateral zone of the median eminence, and contact the external limiting membrane of the brain and the portal capillaries; and finally, in the ventral part of the third ventricle/parenchyma interface, β2 tanycytes, which send end feet that approximate portal capillaries in the medial zone of the median eminence. Multiple cellular properties explain the influence of tanycytes in hypothalamic signaling (Rodríguez-Rodríguez et al. 2019). They serve as a bidirectional conduit for the transfer of signaling molecules from the systemic circulation and the hypothalamus. In addition, tanycytes can sense metabolites and hormones from the cerebrospinal fluid, peripheral circulation, or arcuate nucleus, process and transmit this information to adjacent hypothalamic nuclei, through the release of multiple types of signals. Tanycytes are dynamic; they can retract their body to turn the blood–brain barrier leaky and regulate the secretion of neurohormones. Finally, tanycytes are progenitors that can differentiate into neurons and integrate into hypothalamic circuits (Goodman and Hajihosseini 2015; Prevot et al. 2018; Ebling and Lewis 2018; Bolborea et al. 2020).
4 Regulation of the Activity of Hypophysiotropic TRH Neurons; Impact on Thyroid Axis Function
4.1 Feedback Regulation of HPT Axis Occurs at Multiple Levels
As the activity of TRH hypophysiotropic neurons triggers the activity of the HPT axis, regulation of TRH-neuronal activity has been studied in many paradigms that show alterations in the circulating levels of TSH or TH. HPT axis activity is also regulated at additional levels of the axis and at targets. TH are pleiotropic hormones with many effects in almost every cell; they regulate basal metabolic rate, energy expenditure, thermogenesis, and autonomic function, to mention just a few examples (Fliers et al. 2014; Mullur et al. 2014; Giammanco et al. 2020).
The negative feedback effects of TH provide a good example of their concerted action on the HPT axis at all levels; T3 inhibits Trh and TSH synthesis, stimulates degradation of released TRH, TH-metabolism, and cellular transport in a tissue-specific manner (Mendoza and Hollenberg 2017; Nillni 2010; Joseph-Bravo et al. 2016; Sánchez et al. 2009; Lazcano et al. 2015); at the peripheral level, deiodinases have an exquisite control of the cellular and circulating concentration of T3 (Bianco et al. 2019). Low T3 or T4 and high TSH serum concentrations are the clinical references that define the hypothyroid condition, with the opposite for hyperthyroidism: high serum T3 and low serum TSH concentrations. These pathological conditions are accompanied by disturbances that affect the function of many organs. Hypothyroid individuals present fatigue, weight gain, constipation, dry hair and skin, cardiovascular risk factors, dyslipidemia and increased atherosclerosis, which are many of the symptoms related to metabolic syndrome (Sanchez Jimenez and De Jesus 2020; Biondi and Cooper 2019).
4.2 Hypophysiotropic TRH Neurons and Nutrient Status
Alterations of nutrient status during fasting and food restriction, or during hypercaloric diet intake, change HPT axis activity by the concerted action of circulating leptin concentrations and the activity of the Arc-peptidergic neurons (Fekete and Lechan 2014; Joseph-Bravo et al. 2017). Neurons of the Arc, with a “loose” blood–brain barrier, sense hormones and metabolites that reflect the nutritional status of the organism. Two main populations of Arc neurons are those synthesizing POMC or NPY, which have opposite effects on food intake (anorectic and orexigenic respectively) and on PVN Trh expression (POMC neurons stimulate while NPY neurons inhibit) (Fig. 10.1; Fekete and Lechan 2014). Leptin is a hormone released by adipose tissue that stimulates the synthesis of TRH directly in PVN neurons via STAT3 activation, and indirectly through activation of αMSH (a product of POMC processing) release and CREB activation (Perello et al. 2010) (Figs. 10.3 and 10.5). Fasting produces tertiary hypothyroidism, with decreases in Trh expression in the PVN, TSH serum and pituitary concentrations, and circulating thyroid hormones concentrations, in a sex-specific manner (Joseph-Bravo et al. 2020). The activity of the HPT axis is inhibited by fasting because of the low levels of leptin and the high levels of corticosterone that stimulate tanycyte DIO2 activity (Fig. 10.4, Box 10.2), increasing levels of T3 in PVN neurons (Coppola et al. 2005; Fekete and Lechan 2014). In contrast, pituitary DIO2 is inhibited by fasting, decreasing available T3, and DIO1 activity is reduced in liver decreasing circulating T3. Caloric restriction, even as low as 20% of normal consumption (Uribe et al. 2014), or protein malnutrition (Pałkowska-Goździk et al. 2017), also diminish PVN Trh expression and HPT axis activity. Other nutritional deficiencies (Joseph-Bravo et al. 2017), or pathological conditions such as non-thyroidal illness syndrome or cachexia present tertiary hypothyroidism caused also by increased DIO2 activity in tanycytes due to increased interleukins and nuclear factor κB (Fliers and Boelen 2020).

Hormonal regulation of HPT axis activity occurs at various levels. (a) T3 exerts negative feedback through THRβ2 inhibiting of Trh synthesis and TRH output (stimulation of ME TRH-DE activity), reducting TRH signaling at the pituitary by inhibiting Trhr and Tshb expression and TSH synthesis. (b) Leptin stimulates Trh transcription directly (see Fig. 10.3) or, via regulation of ARC neuropeptides, and stimulates pituitary Tshb expression. (c) Corticosterone inhibits the expression of Trh in the PVN and of Pomc in the ARC and increases ARC-Npy expression. At the pituitary level, corticosterone decreases Tshb mRNA levels. The serum T3 concentration is also reduced by inhibition of hepatic Dio1. (d) E2 administration to ovariectomized rats reduces Trh synthesis in the PVN and secretion from the ME by as yet unidentified mechanisms; in contrast, E2 stimulates Tshr, and Tpo expression in the thyroid, increasing TSH and T3 concentrations in serum. (e) In orchidectomized rats, treatment with testosterone increases TRH secretion, TSH concentration in serum and Tshr mRNA levels in the thyroid gland. Continuous red arrows in panel a refer to T3 negative feedback; black arrows indicate a stimulatory effect, dashed red lines indicate inhibition. Figure created with BioRender.com
4.3 Hypophysiotropic TRH Neurons and Thermogenesis
As well as their role in controlling basal metabolic rate, TH are crucial in thermogenesis (Mullur et al. 2014). In response to cold exposure, TRH neurons are activated, Trh synthesis and release, as well as TSH and T4 release, are stimulated within minutes (Uribe et al. 1993; Zoeller et al. 1995); the increase in Trh expression and TRH release is rapid and transient (30–60 min); Trh mRNA levels peak at 1 h, normalizing by 2 h of exposure at 5 °C, even if cold stress is extended (Uribe et al. 1993; Zoeller et al. 1995). The thermogenic effect of HPT axis activation is co-regulated by the concerted action of the sympathetic system that activates DIO2, which transforms T4 to T3 in the main thermogenic organ, the brown adipose tissue (BAT) (Nedergaard and Cannon 2018). In BAT, the uncoupling protein 1 (UCP-1) produces heat by uncoupling ATP synthesis from oxidative phosphorylation in the mitochondria; T3 stimulates the synthesis of UCP-1, DIO2 and the adrenergic receptor ADRβ3 (Bianco et al. 2019). During cold stress, the increase in serum TSH concentration is amplified when TRH-DE activity is reduced by the injection of a specific inhibitor (Sánchez et al. 2009) or after ablation of tanycytes that does not change PVN Trh expression (Yoo et al. 2020), suggesting that tanycyte TRH-DE activity is limiting TSH release during a cold stress.
4.4 Hypophysiotropic TRH Neurons and Stress
4.4.1 Acute Stress
An adequate response of the HPT axis to metabolic cues, in time and intensity, guarantees energy homeostasis. Situations of stress, however, can affect this response (Joseph-Bravo et al. 2015b). Acute psychological stress such as restraint inhibits PVN Trh expression, pituitary TSH release and lowers the serum concentration of T3 because of inhibited DIO1 in liver (Bianco et al. 1987); since in vitro or in vivo, corticosterone administration increases Trh expression within 1 h, inhibition of Trh expression by acute stress is probably caused by neuronal inputs or by corticosterone-induced endocannabinoid inhibition of TRH neuron activity, as mentioned in Sect. 10.3.3 (Cote-Vélez et al. 2008; Di et al. 2003; Sotelo-Rivera et al. 2014).
Other types of acute stress inhibit the activity of the HPT axis (Joseph-Bravo et al. 2015b) and may also affect its response to acute cold exposure. If an animal is exposed to a short period of stress just before cold exposure, the expected increase in PVN Trh expression or serum TSH concentration in response to cold is not observed, an effect mimicked by injection of corticosterone 30 min before cold exposure (Sotelo-Rivera et al. 2014). The mechanism involved has been unraveled at the molecular level: cold increases the amount of pCREB in TRH neurons of the PVN in control rats but not in those injected with corticosterone; in hypothalamic cell culture, forskolin, a drug that activates protein kinase A (PKA) by releasing the catalytic subunit (PKAc), increases phosphorylation of CREB, increases pCREB in TRH neurons and pCREB binding to CRE (Díaz-Gallardo et al. 2010b; Sotelo-Rivera et al. 2014); this effect is not detected if cells are co-incubated with forskolin + dexamethasone (DEX, an analog of corticosterone that specifically binds GR). Nuclear transport of PKAc and of GR is repressed in cells incubated with forskolin and DEX, compared to the drugs alone, suggesting an interaction that impedes each other transport; in cells incubated with forskolin and DEX, physical interaction of PKAc with GR is demonstrated by co-immunoprecipitation. Since coincubation of forskolin and DEX does not stimulate binding to Trh promoter of a transcription factor that would recruit deacetylases and impede the binding of GR or pCREB to their response elements (Sotelo-Rivera et al. 2017), the data support a model whereby activated GR traps PKAc in the cytosol, impeding its transport to the nucleus and GR or PKAc binding to Trh promoter. If forskolin is added 1 h before DEX, no interference with Trh expression is observed (Sotelo-Rivera et al. 2014). Timing is of utmost importance since it takes few minutes for PKAc to translocate to the nucleus after its activation whereas ligand-bound GR might take longer (Gervasi et al. 2007; Vandevyver et al. 2012; Sotelo-Rivera et al. 2017; Kim and Iremonger 2019); new techniques such as functional fluorescence microscopy imaging (Krmpot et al. 2019) and combined optogenetic approaches (Nomura et al. 2020), reduce calculated time. However, what remains irreducible is the time for a neuronal signal to activate PKA compared to that required for GR to be activated in the brain in vivo, since approximately 20–50 min are required for corticosterone to enter the brain (Joëls et al. 2012; Kim and Iremonger 2019); this is due to the time taken for CRH release, ACTH release from pituitary, ACTH signaling at the adrenal cortex and induction of synthesis and release of corticosterone since, in contrast to neurotransmitters or peptide hormones, steroids are not stored in granules and are synthesized in response to stimuli (Payne and Hales 2004); depending on the stressor, an increased serum concentration of corticosterone may be detected 15 min later, with a peak at 20–60 min (Koolhaas et al. 1997). The blunting effect of a previous corticosterone injection on the response of the hypothalamus-pituitary-adrenal (HPA) axis to a stressor was first reported as restraint-induced expression of PVN Crh requiring at least 30 min but no more than 180 min of corticosterone injection previous to the stressor to repress the response (Osterlund and Spencer 2011); in vitro also, DEX must be added before forskolin to stimulate Crh expression (van der Laan et al. 2009). GR interference on CREB phosphorylation holds for PKA and not for other kinases such as MAPK or ERK (Cote-Vélez et al. 2008).
4.4.2 Chronic Stress
The effects of chronic stress on the HPT axis depend on the type of stress, since the axis habituates to a homotypic stressor as daily intermittent restraint (Uribe et al. 2014). Chronic stress also inhibits the response of the HPT axis to acute cold exposure (Castillo-Campos et al. 2020) but in contrast to the response to an acute stressor, which is attenuated after homotypic chronic stress (as intermittent restraint) and hyperactivated after a heterotypic one (as daily variable stress), the HPT axis response to cold is similarly blunted with both types of stress. The neuronal circuits involved in controlling CRH neurons in these two types of stressor have been characterized and involve GABAergic and glutamatergic neurons projecting from the bed nucleus of stria terminalis to the PVN (Radley and Sawchenko 2015) but whether these circuits participate in the inhibitory effect of chronic stress on the cold-response of hypophysiotropic TRH neurons to cold remains to be elucidated.
4.5 Hypophysiotropic TRH Neurons and Exercise
Another energy-demanding situation that activates the HPT axis is physical activity. Exercise requires an adequate supply of fuel to active tissues such as muscle, provided by endogenous reserves through glycolysis and lipolysis; TH regulates several of the enzymes involved in these reactions as well as the supply of metabolic substrates (Mullur et al. 2014; Klieverik et al. 2009). TH are also involved in the adequate functioning of skeletal and cardiac muscle and in respiration, which explains why hypothyroid individuals have a low exercise performance (Ylli et al. 2020). Increased serum TSH and TH concentrations are detected in animals or humans if taken during or at short times after exercise, before exhaustion or when the individual achieves a negative energy balance (Ylli et al. 2020; Chatzitomaris et al. 2017). Activation of HPT axis activity also occurs in rats undertaking voluntary exercise; Trh expression in the PVN increases, compared to pair-fed controls, proportionally to the amount of exercise performed and to the loss of fat mass (Uribe et al. 2014); likewise, serum T4 concentration increases during treadmill exercise (Fortunato et al. 2008). In humans, only peripheral hormones can be quantified, and results are controversial, depending on the time of sampling and nature of exercise; increased serum TSH, T4, or T3 concentrations are detected after submaximal exercise (running or swimming) (Ylli et al. 2020). A recognized feature of chronic exercise is the loss of adipose tissue. Two weeks of voluntary wheel running, which is a non-stressful form of exercise for rodents, diminish fat depots in abdominal and subcutaneous regions, more in males than in females, and increase Trh expression in PVN of males, and of TSH concentration in serum of females; these responses are curtailed by chronic restraint stress (Parra-Montes de Oca et al. 2019) and by chronic variable stress (CVS) (Parra-Montes de Oca unpublished) which reduce the quantity of exercise performed and block increased Trh expression in the PVN and fat loss.
The inhibitory effect of either acute or chronic stress on the response of PVN TRH neurons or TH concentration to energy demands could explain some of the symptoms of subclinical hypothyroidism, such as fatigability, cold intolerance, and low exercise performance. The efficient mechanisms of HPT axis regulation allow a rapid return to homeostasis; only drastic or pathological situations cause low detectable serum concentrations of TH, in particular T3 (McAninch and Bianco 2014). Knowledge of the mechanisms involved in regulating PVN TRH neurons has allowed a better understanding of situations that stimulate the axis in a transient manner. One can envisage that HPT axis intermittent activation promotes the signals required by energy demands, and that repression of this response produces situations of energy deficit that may be felt as hunger or fatigue, causing increased food consumption and diminished physical activity and eventually leading to obesity and metabolic syndrome.
4.6 Sex Differences in the Activity of Hypophysiotropic TRH Neurons
Energy balance depends on mechanisms that regulate food intake, metabolic substrate distribution, and the different components of energy expenditure, including basal metabolism, physical activity, and thermogenesis, all of which are profoundly affected by HPT axis function (Mullur et al. 2014). Energy homeostasis is regulated differently in males and females; during exercise, for example, males first utilize glycogen reserves while females utilize fat first (Mauvais-Jarvis 2015). Fat distribution also differs, males storing more fat in abdominal depots which promotes the development of metabolic syndrome while females store more in subcutaneous spaces which secrete higher quantities of leptin than abdominal depots (Mauvais-Jarvis 2015; Chusyd et al. 2016).
Parameters of the HPT axis activity and some of those involved in its regulation show sexual dimorphism in their basal state (Fig. 10.5d, e); in some cases, a direct effect of sex hormones has been identified. In the hypothalamus, sex steroids regulate the neuroendocrine and autonomic activity of the PVN; androgen (AR) and estrogen (ER) receptors are expressed in various hypophysiotropic parvocellular neurons and in neurons that project to the brainstem (Bingham et al. 2006), though co-expression of either kind of ER (α or β) with TRH is scant (Suzuki and Handa 2005); using in situ hybridization for Trh, which is more sensitive than immunocytochemistry for the TRH precursor, we also detected few cells co-expressing ERα (Uribe et al. 2009), but ERβ has yet to be studied. Trh expression in the PVN of ovariectomized rats is higher than in controls in the caudal part of the PVN and is decreased by estradiol administration in a dose-dependent manner (Uribe et al. 2009); in these animals, serum concentration of T3 increases in a dose-dependent manner; in contrast, serum TSH concentration is not affected by the higher dose and serum concentrations of T3 and 17β-oestradiol (E2) correlate positively, whereas both correlate negatively with PVN Trh mRNA levels. This illustrates the concerted response of the HPT axis and suggests that the negative effect of E2 on Trh expression could in fact be due to the T3 feedback effect. The response of serum TSH to E2 could be explained by the higher levels of pituitary TRH receptors (Donda et al. 1990; Minakhina et al. 2020). Furthermore, E2 directly stimulates the activity of thyroid peroxidase (TPO) (Lima et al. 2006), an enzyme involved in TH synthesis. The direct effect of testosterone on PVN Trh expression has not been studied, though it has positive effects on other parameters. The TRH content of median eminence decreases in orchidectomized rats but normalizes after testosterone replacement (Pekary et al. 1990), and testosterone induces the expression of Tshb in pituitary (Christianson et al. 1981; Ross 1990) and of Thsr in the thyroid gland (Banu et al. 2001).
Previous sections described how HPT axis activity responds to situations that alter energy homeostasis or stress. Most work was performed in males, but a few studies compared males and females within the same experimental paradigm. For example, HPT axis inhibition in response to starvation occurs more intensely and earlier in males than in females and PVN Trh expression and TSH serum concentration are reduced in males after 24 h of starvation, whereas in females these changes take place only after 48 h of fasting and to a lesser degree (Joseph-Bravo et al. 2020). The response to voluntary exercise also differs; Trh expression in PVN increases in males compared to pair-fed controls proportionally to running but not in females, which show increased TSH serum concentration instead; and females lose less fat than males, in spite of their exercising three times more than males (Parra-Montes de Oca et al. 2019). These results suggest that PVN TRH neurons in females are not activated as in males, but more work is needed to be able to elucidate the mechanism involved; whether it relates to the effects of estradiol on metabolism (Xu and López 2018) awaits resolution.
4.7 Developmental Programming of Hypophysiotropic TRH Neurons
Thyroid hormones play a crucial role in fetal and postnatal neurodevelopment and metabolism (Mullur et al. 2014; Moog et al. 2017a); therefore, any insult that may affect serum TH concentration of the mother, such as maternal disease and diet, exposure to toxins, or disrupting chemicals may have many deleterious effects in offspring development (Mughal et al. 2018; Miranda et al. 2020). Effects may be long-lasting and sex-specific, affecting different tissues depending on their developmental window and time of perturbation (Fall and Kumaran 2019; Miranda et al. 2020).
Before the onset of thyroid function in the human and rat fetus, which occurs around 16–20 weeks of gestation and at embryonic (E) day 17.5, respectively, the fetus is completely dependent on the maternal supply of TH; however, a significant transfer of TH from the mother to the fetus persists after the onset of the fetal thyroid function (Moog et al. 2017a; Morreale de Escobar et al. 1987). While hypothalamic TRH neurons appear by E13 in the rat (Markakis and Swanson 1997), TRH is mainly produced postnatally, showing a peak at day 21 after birth (Burgunder and Taylor 1989). Gestation, lactation, and adolescence are thus critical developmental windows that can alter the function of the HPT axis.
4.7.1 Nutrition
Maternal obesity is associated with decreased hypothalamic Trh expression only in rat male pups, and increased hypothalamic Dio2 expression only in female pups (Dias-Rocha et al. 2018). In non-human primates, maternal obesity induced by a pregestational high-fat diet decreases hypothalamic Trh expression of the fetus at the beginning of the first trimester (Suter et al. 2012).
Maternal protein or energy restriction during gestation/lactation programs the adult rat offspring for thyroid dysfunction, provoking low TSH in vitro TRH-induced release (Lisboa et al. 2008), low body weight, and a decreased resting metabolic rate (Ayala-Moreno et al. 2013). Inadequate micronutrient intake during gestation and lactation is another factor implicated in the development and HPT axis function of the progeny. In particular, necessary elements for TH synthesis (iodine) or enzyme function (e.g., Se for deiodinases and Zn for various metalloenzymes and transcription factors), if insufficient in the mother’s diet during gestation or suckling periods, produce deleterious effects (see reviews Rezaei et al. 2019; Hubalewska-Dydejczyk et al. 2020). Zn deficiency in the mother’s diet and during adolescence decreases TRH-DE activity in median eminence and in pituitary and increases TSH concentration in serum with no effect on TH levels (Alvarez-Salas et al. 2015). During pregnancy, adequate iodine intake is required to meet maternal and fetal needs and to account for increased maternal losses; iodine deficiency in early life impairs cognition and growth (Pearce et al. 2016). Excessive iodine intake during pregnancy and lactation increases the mother’s susceptibility to thyroid dysfunction; this can affect cognitive development of the offspring and increases the risk of infant hypothyroidism induced by an excessive concentration of iodine in breast-milk (Pearce et al. 2016; Farebrother et al. 2019). Maternal ingestion of high concentrations of iodine alters the function of the HPT axis of male rat offspring in adulthood; Trh and Tsh expression in the hypothalamus and pituitary are increased, along with elevated TSH secretion.
During the suckling period pups are also susceptible; overnutrition programs adult male rats for central hypothyroidism, reflected by decreased mRNA levels of Trh in the PVN and protein expression in the hypothalamus, and lower basal and TRH-stimulated TSH secretion (Aréchiga-Ceballos et al. 2014; Lisboa et al. 2015). Neonatal and postnatal exposure to a diet high with soy content leads to increased hypothalamic Trh expression in adult mice, although TH levels are not affected (Cederroth et al. 2007). In contrast, undernourishment during suckling mice alters postnatal development and long-term hypothalamic gene expression, including that of Trh, expression of which is decreased in 21-day-old male and female offspring (Kaczmarek et al. 2016).
4.7.2 Stress
Fetal exposure to synthetic GC as DEX or postnatal chronic stress results in behavioral and metabolic disturbances later in life and permanent alterations of gene expression of neuropeptides. Sex-specific effects are observed in adult rats exposed to DEX during late gestation; core body temperature is reduced in females, but not males, and although preproTRH-ir fibers are reduced in the PVN of both male and female offspring, only adult females present a reduced number of preproTRH-ir neurons in the PVN as well as mRNA levels (Carbone et al. 2012). The higher susceptibility of females to prenatal GC exposure is confirmed in guinea pigs (Moisiadis et al. 2017). Postnatal stress, such as maternal separation (MS) during the suckling period or childhood maltreatment, is associated with reduced thyroid activity (decreased serum levels of TSH and/or T3) in male rats, adolescents and non-pregnant women (Jaimes-Hoy et al. 2016, 2019; Sinai et al. 2014; Machado et al. 2015), increasing their risk of exhibiting subclinical hypothyroidism during pregnancy (Moog et al. 2017b) and putting the child at risk of adverse neurodevelopmental outcomes. In contrast to the inhibitory effects of GC administration during gestation on Trh expression in females, MS increases it; PVN-Trh expression in males is not affected but the expression of its degrading enzyme is increased, resulting in a low TSH serum concentration in adult male rats. MS affected not only the basal activity of the HPT axis but its response to various challenges such as food deprivation or cold exposure is blunted in males, but not in females (Jaimes-Hoy et al. 2016, 2021). The response to hypercaloric palatable diet and psychological stress (restraint) is also modified, depending on whether the diet is started in puberty or adulthood. For example, restrained MS male rats exposed to a high-fat/high-carbohydrate diet from puberty have increased Trh expression in PVN and decreased concentrations of TSH and TH in serum, whereas females do have increased PVN-Trh mRNA but serum TH levels are also increased, suggesting that males are more susceptible to interference with the adaptive response of this neuroendocrine axis to a metabolic stressor (Jaimes-Hoy et al. 2019).
4.7.3 Tobacco and Endocrine Disrupting Chemicals
Prenatal and infant exposure to toxins or pollutants may have persistent effects throughout life. Tobacco smoking during pregnancy/lactation exerts numerous short- and long-term adverse effects on the neonate’s health, increasing the risk of developing obesity, hypertension and metabolic and lung-related diseases, including altered thyroid function and development (Banderali et al. 2015; Miranda et al. 2020). Maternal nicotine exposure leads to microgliosis in the PVN (Younes-Rapozo et al. 2015), reduced hypothalamic TRH content (Younes-Rapozo et al. 2013) and secondary hypothyroidism is induced in the PVN of male offspring at adulthood, with low serum levels of TSH and TH. This is due in part to in vivo TRH-TSH suppression and decreased sensitivity to TRH (Miranda et al. 2020).
Plastics and other chemicals that have contaminated water and soil produce substances that affect the endocrine system, called endocrine-disrupting chemicals (EDCs) (Gore et al. 2019). EDCs may act alone or in combination, impairing estrogenic/androgenic and thyroid function, the latter acting at multiple levels of the HPT and gonadal axes; dysfunction of these axes have been associated with obesity, reproductive alterations, breast, ovarian and thyroid cancer, hypothyroidism and cognitive impairment (Mughal et al. 2018). Perinatal exposure to triclosan (TCS), a common chemical present in household and personal products, reduces expression levels of Trh, Thra and TH transporters in the brains of mouse offspring (Tran et al. 2020). In gestating mice dams, acute treatment on the day of delivery with the organotin tributyltin (TBT) dose-dependently increases Trh transcription in pups’ hypothalamic, independent of T3 and mediated by hypothalamic overexpression of Rxra; in contrast, chronic exposure of dams to the flame-retardant tetrabromobisphenol A (TBBPA) during late gestation diminishes Trh and Mc4r transcription in pups’ hypothalami in the absence of T3 (Mughal et al. 2018). Poly-brominated diphenyl ethers (PBDEs), used as flame retardant additives, have been banned in several countries but persist in the environment (Sharkey et al. 2020); they reduce whole-body T4 content accompanied by down-regulation of Trh and Tshb in offspring (Han et al. 2017), which may contribute to the associated neurodevelopmental alterations.
The list of endocrine disruptors will probably keep growing, and with uncontrollable stress situations cause long-term effects on HPT axis function, but most worrying are those affecting the gestation period, which might be the basis of the increasing number of patients with diseases related to thyroid effects on brain development.
5 Hypophysiotropic TRH Neurons Regulate Prolactin Secretion
Soon after its discovery, it was shown that TRH stimulates prolactin (PRL) secretion either in vivo or in vitro (Jacobs et al. 1971; Tashjian et al. 1971) although controversies about male responses soon appeared (Watanobe et al. 1985). PRL secretion is controlled directly by hypothalamic tuberoinfundibular dopamine neurons, which exert an inhibitory drive, but also by various hypothalamic neurons releasing stimulatory factors (Grattan 2015). During suckling, dopamine (DA) release into portal blood is inhibited while TRH biosynthesis in the PVN and release from the ME are stimulated (Fink et al. 1982; Uribe et al. 1993; Van Haasteren et al. 1996; Sánchez et al. 2001), implicating the hypophysiotropic PVN TRH neurons. Furthermore, TRH antisera inhibit suckling-induced PRL release (de Greef et al. 1987), and studies with TRH and TRH-R1 KO mice have shown that TRH is necessary to maintain maximal prolactin output in lactating mice (Rabeler et al. 2004; Yamada et al. 2006).
TSH is not released by suckling, nor is PRL by cold exposure (Uribe et al. 1993; Van Haasteren et al. 1996; Sánchez et al. 2001), suggesting that post-secretory processes likely refine the specificity of TRH action in the anterior pituitary. Although the hypophysiotropic TRH neurons regulating prolactin may differ in part from those controlling TSH secretion (Sánchez et al. 2001), there is no evidence yet for an anatomical pathway that may segregate TRH released by each kind of hypophysiotropic neuron. The stimulus-dependent response may originate from other aspects of PRL secretion control. CART, co-expressed in TRH hypophysiotropic neurons, is upregulated by 1 h exposure to cold but not by suckling (Sánchez et al. 2007). Since CART inhibits prolactin release in vitro and cold exposure does not induce the release of PRL, CART may serve as a modulator of TRH actions in these physiological circumstances, stimulating TSH release while blocking prolactin release (Sánchez et al. 2001, 2007; Raptis et al. 2004).
DA withdrawal during suckling, or in primary cultures of hypophysial cells, potentiates TRH-induced PRL secretion (Martinez de la Escalera and Weiner 1992). In lactotrophs the intensity of TRH action is under TRH-DE control: in primary cultures of female rat anterior pituitary cells, Trhde is expressed in some lactotrophs, and inhibition of Trhde expression or activity enhances TRH-induced prolactin secretion (Bauer et al. 1990; Cruz et al. 2008). In anterior pituitary cells, TRH-DE activity is rapidly enhanced by the removal of DA and addition of TRH (Bourdais et al. 2000). These results suggest that lactotrophs TRH-DE activity is controlled by signals that shape PRL secretion in response to TRH and that TRH-DE regulation may in turn alter PRL release.
Finally, although many studies have shown that TRH acts directly on lactotrophs, it is relevant to note that numerous TRH fibers enter the Arc (Péterfi et al. 2018). TRH terminals abut on tuberoinfudibular dopaminergic (TIDA) neurons in rats (Lyons et al. 2010) and humans (Dudas and Merchenthaler 2020), coincident with the expression of the TRH receptor in Arc (Heuer et al. 2000), but the TRH neurons of origin are not known. In hypothalamic slices, TRH provokes a transition from phasic to tonic firing of the TIDA neurons that control PRL secretion (Lyons et al. 2010). The relative importance of direct and indirect control of prolactin secretion by TRH remains to be elucidated.
6 Perspectives
In this review, we have attempted to present a summary of the characteristics of TRH hypophysiotropic neurons. These parvocellular neurons are mixed with multiple cell types in the PVN, including various subtypes of TRH neurons, making this characterization daunting. We have information about a small set of genes; although single-cell transcriptomes of hypothalamic TRH neurons have been published, a definitive assignation of a TRH cluster to the hypophysiotropic neurons is lacking.
Delineation of the neuronal circuits that modulate hypophysiotropic TRH neurons activity under different paradigms is not complete. Advances have been made in defining how they sense metabolic information, including the involvement of Arc inputs. However, knowledge of the circuits involved in other events is only partial. Afferents from the suprachiasmatic nucleus contact PVN TRH neurons (Kalsbeek et al. 2000), but the input and target neuronal types involved are still unknown. Cold exposure activates noradrenaline neurons that contact hypophysiotropic TRH neurons, but whether these inputs arise from locus coeruleus, nucleus tractus solitarius or other noradrenergic nuclei is still under investigation. How chronic stress affects the activity of TRH neurons, and whether it depends on the type of stress as for CRH neurons (Radley and Sawchenko 2015) is also unknown. Another unknown is the identity of the neuronal circuit that activates the hypophysiotropic TRH neurons in response to suckling.
We also reviewed the most important data on TRH mode of synthesis, release, and inactivation. Knowledge of these metabolic steps has been used to obtain subrogate measures of TRH neurons activity, since quantification of TSH or TH serum concentrations does not precisely reflect changes in TRH release into portal blood nor the activity of HPT axis as peripheral metabolism of TH is strictly regulated in a time- and tissue-dependent manner (Bianco et al. 2019). Together, these measurements allowed the identification of an array of regulators of TRH metabolism and neuronal activity that are intimately linked to the activity of the whole HPT axis since basal levels of Trh expression depend on TH feedback and nutrition status. In addition, TRH neurons and HPT axis respond to energy-demanding situations according to previous exposure to stress (immediate or during development).
The design of experiments that analyze the activity of the TRH neurons must take into account various considerations mentioned in Box 10.3, as well as conditions of stress and metabolic changes imposed by the paradigm (Castillo-Campos et al. 2020). Consideration of stress conditions during the experiment and at sacrifice is essential since even taking rats from a cage causes the immediate release of corticosterone, which depends on the order of animal removal from its cage (Ferland and Schrader 2011). Another factor is the animal species under study; the rat was the preferred animal for research in physiology, but since the development of transgenesis the mouse became a common object of study; because, these species differ in many ways (Ellenbroek and Youn 2016), extrapolating results from one species to another could produce false hypotheses. For translational studies, it is relevant to note that most work on rodents is performed during the light period that corresponds to the inactive period of rodents, with low serum corticosterone and high thyroid hormone concentrations, according to circadian status. Another very important task is a comparison of results from males and females, ideally within the same experiment to control most variables.
Finally, until recently many studies have been limited to measuring activity at one (or sometimes a few) sampling time; it is like a snapshot that does not explain the processes under study, which are dynamic, and causal relationships have been difficult to investigate. However, the development of genetic techniques in the recent past, including transgenesis, CRISPR-Cas9, chemogenetic and optogenetic methods, provide an ample portfolio of tools that will undoubtedly allow precise monitoring of TRH neurons activity in vivo, using for example real-time calcium fiber photometry recordings, and manipulation of their activity with chemical and optical tools (Müller-Fielitz et al. 2017; Farkas et al. 2020).
The information gathered so far on TRH neuron activity could help to better diagnose subclinical hypothyroidism and suggests that it is relevant to consider the stress level of the patient. It appears that measurements of serum TSH, TH and cortisol concentrations before and after a bout of exercise or cold exposure may be more appropriate than evaluation only at basal state, which utility is limited for diagnosis and treatment (Biondi et al. 2019).
Box 10.3. Tools to Study Activity of TRH Neurons
To evaluate the activation of TRH neurons, measurement of the levels of mRNA has been used as a good index since they are rapidly increased in response to stimulation; an increase can be detected at 45–60 min (Uribe et al. 1993; Zoeller et al. 1995), near the time required to measure immediate early gene expression, such as c-fos. In many events, the same effector stimulates both synthesis and release; for example, cold-induced activation of noradrenaline neurons signals to TRH hypophysiotropic neurons, and increases release as well as the synthesis of TRH (Perello et al. 2007). However, not all effectors affect both processes; for example, acute corticosterone administration increases Trh mRNA levels but decreases TRH release through the endocannabinoid pathway at PVN level (Sotelo-Rivera et al. 2014; Di et al. 2003). The activity of the hypophysiotropic neurons can be studied either through sampling most TRH neurons in a large volume of the PVN or with cellular resolution. For rapidly sampling the activity of most hypophysiotropic TRH neurons, a common strategy is to measure Trh mRNA levels in punches of the PVN with RT-PCR. Alternatively, levels of proTRH measured by Western blotting, or levels of TRH by RIA, may be used to infer the status of TRH biosynthesis in the PVN. For these strategies to be most specific for hypophysiotropic neurons, an important consideration is the adequate dissection of the PVN. Reports on single-cell transcriptomes that do not report how dissection was performed may have limited usefulness; likewise, analysis of the whole hypothalamus to deduce regulation of HPT axis activity is meaningless. Within the hypothalamus, apart from the PVN, several nuclei express TRH (Joseph-Bravo et al. 2015b). Caudal to the PVN, the dorsomedial hypothalamus contains a large population of non-endocrine TRH neurons that are activated for example by exercise, like those of PVN (Uribe et al. 2014); this localization is essential to consider when injecting directly into the PVN, as well as the size of cannulas. In the ventrolateral directions, the lateral hypothalamus expresses many TRH neurons too. In the PVN proper, many TRH neurons of the anterior PVN are not hypophysiotropic, and project to other hypothalamic nuclei and brain areas (Wittmann et al. 2009), whereas TRH magnocellular neurons located in mid-PVN can only be differentiated from the hypophysiotropic neurons under the microscope. Finally, other parvocellular TRH neurons of mid-caudal PVN (in the rat) are not hypophysiotropic (Simmons and Swanson 2009) although a good correlation has been obtained between punches and histochemical data in various paradigms. On the other hand, immunohistochemical and/or in situ hybridization techniques generate cellular resolution, but to guarantee that data refers to hypophysiotropic parvocellular TRH neurons, additional information, such as CART expression (Fekete et al. 2000) is required. A direct measure of PVN TRH neuron (identified afterward by immunohistochemistry or single-cell transcriptomics) activity can be obtained by electrophysiology in hypothalamic slices (Di et al. 2003). The distinct localization of the PVN and median eminence along the anteroposterior axis in coronal slices allows separate quantifications of PVN Trh expression and processed TRH in nerve endings. Techniques designed to measure in vivo release of TRH are cumbersome and not precise. Measuring TRH content in median eminence extracts, less than 2 h after a stimulus, may indicate release if the content is reduced, although processing could also be affected. At the nerve terminal of TRH neurons, cleaved products of TRH precursor are enriched (Lechan et al. 1987), thus for TRH quantification adequate antibodies are required, which usually recognize pGlu and ProNH2 moieties, so those raised against precursor forms will not recognize it. Another indirect measure of TRH release is the measurement of serum TSH concentration. The i.v. injection of anti-TRH antibodies can reduce serum TSH concentration, which validates this approach in short-term studies. However, secretion of TSH is also regulated by other hypothalamic (SRIF) or peripheral influences (TH, corticosterone). The development of recombinant DNA techniques and the creation of transgenic mice have provided information on the role of multiple molecules involved in signaling and transcription of Trh, or of the elements involved in the HPT axis as deiodinases. The most interesting data are those obtained with conditional transgenesis, so that expression of the gene of interest is only altered in a specific tissue and/or in a defined window of time, which avoids the problems produced by indirect effects through other cell types (Fonseca et al. 2013) and/or the lack of that protein during development.
Abbreviations
- 2-AG:
-
2-arachinodonoylglycerol (Italics are used for gene or mRNA names, and capital letters, for peptides/proteins)
- 3V:
-
Third ventricle
- αMSH:
-
α-melanocyte stimulating hormone
- a:
-
Anterior
- A2 or 6:
-
Group 2 or 6 noradrenergic neurons
- ACTH:
-
Adrenocorticotropin hormone
- ADRB3:
-
Beta 3 adrenergic receptor
- AgRP:
-
Agouti related protein
- Arc/ARC:
-
Hypothalamic arcuate nucleus
- BAT:
-
Brown adipose tissue
- BDE-209:
-
Decabromodiphenyl ether
- BNST:
-
Bed nucleus of the stria terminalis
- C1–3:
-
Catecholaminergic neurons
- CART:
-
Cocaine and amphetamine regulated transcript
- CB1R:
-
Cannabinoid receptor 1
- CPE:
-
Carboxy-peptidase E
- CRE:
-
Cyclic AMP response element
- CREB:
-
cAMP-response element binding protein
- CRH:
-
Corticotropin releasing hormone
- DA:
-
Dopamine
- DAGLα:
-
Diacylglycerol lipase α
- DBH:
-
Dopamine beta-hydroxylase
- DEX:
-
Dexamethasone
- DIO1:
-
Deiodinase type 1
- DIO2:
-
Deiodinase type 2
- DIO3:
-
Deiodinase type 3
- DMH:
-
Dorsomedial hypothalamic nucleus
- E:
-
Embryonic day
- E2:
-
17β-oestradiol
- EDCs:
-
Endocrine disrupting chemicals
- EM66:
-
Secretogranin II-derived peptide
- ER:
-
Endoplasmic reticulum
- ERK:
-
Extracellular signal-regulated kinase
- ERα or βb:
-
Estrogen receptor α or β
- fc:
-
Fenestrated capillaries
- GABA:
-
γ-aminobutyric acid
- GAD:
-
Glutamic acid decarboxylase
- GATA2:
-
GATA-binding factor 2
- GC:
-
Glucocorticoid
- GLRA1,2,3:
-
Glycine receptor alpha 1, 2 or 3
- GLRB:
-
Glycine receptor beta
- Glu:
-
Glutamate
- GluR:
-
Glutamate receptor
- GLYT2:
-
Glycine transporter 2
- GR:
-
Glucocorticoid receptor
- GRE:
-
Glucocorticoid response element
- HDAC2 or 3:
-
Histone-deacetylase 2 or 3
- HLHP-2:
-
Helix-loop-helix protein 2
- HPA:
-
Hypothalamic-pituitary adrenal
- HPT:
-
Hypothalamic-pituitary-thyroid
- Jun:
-
AP-1 transcription factor subunit p39
- KLF10:
-
Krüppel like factor 10
- KO:
-
Knockout
- LC:
-
Locus coeruleus
- LH:
-
Lateral hypothalamus
- LXR:
-
Liver X receptor
- m:
-
Medial
- MAPK:
-
Mitogen-activated protein kinase
- MBH:
-
Mediobasal hypothalamus
- MC4R:
-
Melanocortin 4 receptor
- MCT8:
-
Monocarboxylate transporter 8
- ME:
-
Median eminence
- MS:
-
Maternal separation
- n:
-
Undetermined position
- NCoR:
-
Nuclear receptor co-repressor
- NE:
-
Norepinephrine
- NPY:
-
Neuropeptide Y
- NTS:
-
Nucleus of solitary tract
- p:
-
Posterior
- PAM:
-
Peptidylgycine alpha-amidating monooxygenase
- PBDEs:
-
Poly-brominated diphenyl ethers
- PC:
-
Prohormone convertase
- pCREB:
-
Phosphorylated cAMP-response element binding protein
- PKA:
-
Protein kinase A
- PKAc:
-
Catalytic subunit of PKA
- PLZF:
-
Promyelocytic leukemia zinc finger protein
- PND:
-
Postnatal day
- PNMT:
-
Phenylethanolamine n-methyl transferase
- POMC:
-
Proopiomelanocortin
- PRL:
-
Prolactin
- pSTAT3/5:
-
Phosphorylated signal transducer and activator of transcription 3/5
- PVN:
-
Hypothalamic paraventricular nucleus
- QC:
-
Glutaminyl cyclase
- RMg:
-
Raphe magnus
- rT3:
-
Reverse T3
- RVLM:
-
Rostral ventrolateral medulla
- RXR:
-
Retinoid X receptor
- SCN:
-
Suprachiasmatic nucleus
- scRNAseq:
-
Single cell RNA sequencing
- SIM2:
-
Single-minded homolog 2
- SOCS-3:
-
Suppressor of cytokine signaling-3
- STAT3:
-
Signal transducer and activator of transcription 3
- T2:
-
3,5-diiodothyronine
- T3:
-
3,3′,5 triiodo-l-thyronine
- T4:
-
Thyroxine
- TBBPA:
-
Tetrabromobisphenol A
- TBT:
-
Tributyltin
- TCS:
-
Triclosan
- TH:
-
Thyroid hormone
- THR:
-
Thyroid hormone receptor
- THRE:
-
Thyroid hormone response element
- THRα or β:
-
Thyroid hormone receptor α or β
- TIDA:
-
Tuberoinfudibular dopaminergic
- TPO:
-
Thyroid peroxidase
- TRH:
-
Thyrotropin releasing hormone
- TRH-DE:
-
TRH-degrading ecto-enzyme
- Trhr, TRH-R1:
-
TRH receptor-1
- TSH:
-
Thyrotropin
- TSHR:
-
Thyrotropin receptor
- UCP-1:
-
Uncoupling protein 1
- VGLUT2:
-
Vesicular glutamate transporter 2
- VLPAG:
-
Ventrolateral periaqueductal gray
- VMH/VMN:
-
Hypothalamic ventromedial nucleus
- Y1 or Y5 receptor:
-
NPY receptor 1 or 5
References
Abel ED, Ahima RS, Boers ME, Elmquist JK, Wondisford FE (2001) Critical role for thyroid hormone receptor beta2 in the regulation of paraventricular thyrotropin-releasing hormone neurons. J Clin Invest 107:1017–1023
Alkemade A, Friesema EC, Unmehopa UA, Fabriek BO, Kuiper GG, Leonard JL, Wiersinga WM, Swaab DF, Visser TJ, Fliers E (2005) Neuroanatomical pathways for thyroid hormone feedback in the human hypothalamus. J Clin Endocrinol Metab 90:4322–4334
Alvarez-Salas E, Alcántara-Alonso V, Matamoros-Trejo G, Vargas MA, Morales-Mulia M, de Gortari P (2015) Mediobasal hypothalamic and adenohypophyseal TRH-degrading enzyme (PPII) is down-regulated by zinc deficiency. Int J Dev Neurosci 46:115–124
Aréchiga-Ceballos F, Alvarez-Salas E, Matamoros-Trejo G, Amaya MI, García-Luna C, de Gortari P (2014) Pro-TRH and pro-CRF expression in paraventricular nucleus of small litter-reared fasted adult rats. J Endocrinol 221:77–88
Astapova I, Hollenberg AN (2013) The in vivo role of nuclear receptor corepressors in thyroid hormone action. Biochim Biophys Acta 1830:3876–3881
Ayala-Moreno R, Racotta R, Anguiano B, Aceves C, Quevedo L (2013) Perinatal undernutrition programmes thyroid function in the adult rat offspring. Br J Nutr 110:2207–2215
Banderali G, Martelli A, Landi M, Moretti F, Betti F, Radaelli G, Lassandro C, Verduci E (2015) Short and long term health effects of parental tobacco smoking during pregnancy and lactation: a descriptive review. J Transl Med 13:327
Banu KS, Govindarajulu P, Aruldhas MM (2001) Testosterone and estradiol modulate TSH-binding in the thyrocytes of Wistar rats: influence of age and sex. J Steroid Biochem Mol Biol 78:329–342
Bauer K, Carmeliet P, Schulz M, Baes M, Denef C (1990) Regulation and cellular localization of the membrane-bound thyrotropin-releasing hormone-degrading enzyme in primary cultures of neuronal, glial and adenohypophyseal cells. Endocrinology 127:1224–1233
Bianco AC, Nunes MT, Hell NS, Maciel RM (1987) The role of glucocorticoids in the stress-induced reduction of extrathyroidal 3,5,3′-triiodothyronine generation in rats. Endocrinology 120:1033–1038
Bianco AC, Dumitrescu A, Gereben B, Ribeiro MO, Fonseca TL, Fernandes GW, Bocco BMLC (2019) Paradigms of dynamic control of thyroid hormone signaling. Endocr Rev 40:1000–1047
Bingham B, Williamson M, Viau V (2006) Androgen and estrogen receptor-beta distribution within spinal-projecting and neurosecretory neurons in the paraventricular nucleus of the male rat. J Comp Neurol 499:911–923
Biondi B, Cooper DS (2019) Thyroid hormone therapy for hypothyroidism. Endocrine 66:18–26
Biondi B, Cappola AR, Cooper DS (2019) Subclinical hypothyroidism: a review. JAMA 322:153–160
Bolborea M, Pollatzek E, Benford H, Sotelo-Hitschfeld T, Dale N (2020) Hypothalamic tanycytes generate acute hyperphagia through activation of the arcuate neuronal network. Proc Natl Acad Sci USA 117:14473–14481
Boler J, Enzmann F, Folkers K, Bowers CY, Schally AV (1969) The identity of chemical and hormonal properties of the thyrotropin releasing hormone and pyroglutamyl-histidyl-proline amide. Biochem Biophys Res Commun 37:705–710
Bourdais J, Romero F, Uriostegui B, Cisneros M, Joseph-Bravo P, Charli JL (2000) [3-Me-His(2)]-TRH combined with dopamine withdrawal rapidly and transiently increases pyroglutamyl aminopeptidase II activity in primary cultures of adenohypophyseal cells. Neuropeptides 34:83–88
Broberger C (1999) Hypothalamic cocaine- and amphetamine-regulated transcript (CART) neurons: histochemical relationship to thyrotropin-releasing hormone, melanin-concentrating hormone, orexin/hypocretin and neuropeptide Y. Brain Res 848:101–113
Burgunder JM, Taylor T (1989) Ontogeny of thyrotropin-releasing hormone gene expression in the rat diencephalon. Neuroendocrinology 49:631–640
Burgus R, Dunn TF, Desiderio D, Guillemin R (1969) Molecular structure of the hypothalamic hypophysiotropic TRF factor of ovine origin: mass spectrometry demonstration of the PCA-His-Pro-NH2 sequence. C R Acad Hebd Seances Acad Sci D 269:1870–1873
Cakir I, Nillni EA (2019) Endoplasmic reticulum stress, the hypothalamus, and energy balance. Trends Endocrinol Metab 30:163–176
Campos AMP, Teixeira PDS, Wasinski F, Klein MO, Bittencourt JC, Metzger M, Donato J Jr (2020) Differences between rats and mice in the leptin action on the paraventricular nucleus of the hypothalamus: implications for the regulation of the hypothalamic-pituitary-thyroid axis. J Neuroendocrinol 32:e12895
Carbone DL, Zuloaga DG, Lacagnina AF, McGivern RF, Handa RJ (2012) Exposure to dexamethasone during late gestation causes female-specific decreases in core body temperature and prepro-thyrotropin-releasing hormone expression in the paraventricular nucleus of the hypothalamus in rats. Physiol Behav 108:6–12
Castillo-Campos A, Gutiérrez-Mata A, Charli JL, Joseph-Bravo P (2020) Chronic stress inhibits hypothalamus-pituitary-thyroid axis and brown adipose tissue responses to acute cold exposure in male rats. J Endocrinol Invest. https://doi.org/10.1007/s40618-020-01328-z
Cawley NX, Rathod T, Young S, Lou H, Birch N, Loh YP (2016) Carboxypeptidase E and secretogranin III coordinately facilitate efficient sorting of proopiomelanocortin to the regulated secretory pathway in AtT20 Cells. Mol Endocrinol 30:37–47
Ceccatelli S, Cintra A, Hökfelt T, Fuxe K, Wikström AC, Gustafsson JA (1989) Coexistence of glucocorticoid receptor-like immunoreactivity with neuropeptides in the hypothalamic paraventricular nucleus. Exp Brain Res 78:33–42
Cederroth CR, Vinciguerra M, Kühne F, Madani R, Doerge DR, Visser TJ, Foti M, Rohner-Jeanrenaud F, Vassalli JD, Nef S (2007) A phytoestrogen-rich diet increases energy expenditure and decreases adiposity in mice. Environ Health Perspect 115:1467–1473
Charli JL, Rodríguez-Rodríguez A, Hernández-Ortega K, Cote-Vélez A, Uribe RM, Jaimes-Hoy L, Joseph-Bravo P (2020) The thyrotropin-releasing hormone-degrading ectoenzyme, a therapeutic target? Front Pharmacol 11:640
Chatzitomaris A, Hoermann R, Midgley JE, Hering S, Urban A, Dietrich B, Abood A, Klein HH, Dietrich JW (2017) Thyroid allostasis-adaptive responses of thyrotropic feedback control to conditions of strain, stress, and developmental programming. Front Endocrinol (Lausanne) 8:163
Chen R, Wu X, Jiang L, Zhang Y (2017) Single-cell RNA-seq reveals hypothalamic cell diversity. Cell Rep 18:3227–3241
Cheng H, Pablico SJ, Lee J, Chang JS, Yu S (2020) Zinc finger transcription factor Zbtb16 coordinates the response to energy deficit in the mouse hypothalamus. Front Neurosci 14:592947
Christianson D, Roti E, Vagenakis AG, Braverman LE (1981) The sex-related difference in serum thyrotropin concentration is androgen mediated. Endocrinology 108:529–535
Chusyd DE, Wang D, Huffman DM, Nagy TR (2016) Relationships between rodent white adipose fat pads and human white adipose fat depots. Front Nutr 3:10
Cintra A, Fuxe K, Solfrini V, Agnati LF, Tinner B, Wikström AC, Staines W, Okret S, Gustafsson JA (1991) Central peptidergic neurons as targets for glucocorticoid action. Evidence for the presence of glucocorticoid receptor immunoreactivity in various types of classes of peptidergic neurons. J Steroid Biochem Mol Biol 40:93–103
Ciosek J, Izdebska K (2009) Thyrotropin-releasing hormone modulates vasopressin and oxytocin synthesis and release from the hypothalamo-neurohypophysial system of different age male rats. J Physiol Pharmacol 60:63–70
Coppola A, Meli R, Diano S (2005) Inverse shift in circulating corticosterone and leptin levels elevates hypothalamic deiodinase type 2 in fasted rats. Endocrinology 146:2827–2833
Cote-Vélez A, Pérez-Martínez L, Charli JL, Joseph-Bravo P (2008) The PKC and ERK/MAPK pathways regulate glucocorticoid action on TRH transcription. Neurochem Res 33:1582–1591
Cote-Vélez A, Pérez-Maldonado A, Osuna J, Barrera B, Charli JL, Joseph-Bravo P (2011) Creb and Sp/Krüppel response elements cooperate to control rat TRH gene transcription in response to cAMP. Biochim Biophys Acta 1809:191–199
Cruz R, Vargas MA, Uribe RM, Pascual I, Lazcano I, Yiotakis A, Matziari M, Joseph-Bravo P, Charli JL (2008) Anterior pituitary pyroglutamyl peptidase II activity controls TRH-induced prolactin release. Peptides 29:1953–1964
de Greef WF, Voogt JL, Visser TJ, Lamberts SW, van der Schoot P (1987) Control of prolactin release induced by suckling. Endocrinology 121:316
Di S, Malcher-Lopes R, Halmos KC, Tasker JG (2003) Nongenomic glucocorticoid inhibition via endocannabinoid release in the hypothalamus: a fast feedback mechanism. J Neurosci 23:4850–4857
Dias-Rocha CP, Almeida MM, Santana EM, Costa JCB, Franco JG, Pazos-Moura CC, Trevenzoli IH (2018) Maternal high-fat diet induces sex-specific endocannabinoid system changes in newborn rats and programs adiposity, energy expenditure and food preference in adulthood. J Nutr Biochem 51:56–68
Díaz-Gallardo MY, Cote-Vélez A, Carreón-Rodríguez A, Charli JL, Joseph-Bravo P (2010a) Phosphorylated cyclic-AMP-response element-binding protein and thyroid hormone receptor have independent response elements in the rat thyrotropin-releasing hormone promoter: an analysis in hypothalamic cells. Neuroendocrinology 91:64–76
Díaz-Gallardo MY, Cote-Vélez A, Charli JL, Joseph-Bravo P (2010b) A rapid interference between glucocorticoids and cAMP-activated signalling in hypothalamic neurones prevents binding of phosphorylated cAMP response element binding protein and glucocorticoid receptor at the CRE-Like and composite GRE sites of thyrotrophin-releasing hormone gene promoter. J Neuroendocrinol 22:282–293
Donda A, Reymond F, Rey F, Lemarchand-Béraud T (1990) Sex steroids modulate the pituitary parameters involved in the regulation of TSH secretion in the rat. Acta Endocrinol (Copenh) 122:577–584
dos-Santos RC, Grover HM, Reis LC, Ferguson AV, Mecawi AS (2018) Electrophysiological effects of ghrelin in the hypothalamic paraventricular nucleus neurons. Front Cell Neurosci 12:275
Dudas B, Merchenthaler I (2020) Thyrotropin-releasing hormone axonal varicosities appear to innervate dopaminergic neurons in the human hypothalamus. Brain Struct Funct 225:2193–2201
Dyess EM, Segerson TP, Liposits Z, Paull WK, Kaplan MM, Wu P, Jackson IM, Lechan RM (1988) Triiodothyronine exerts direct cell-specific regulation of thyrotropin-releasing hormone gene expression in the hypothalamic paraventricular nucleus. Endocrinology 123:2291–2297
Ebling FJP, Lewis JE (2018) Tanycytes and hypothalamic control of energy metabolism. Glia 66:1176–1184
Ellenbroek B, Youn J (2016) Rodent models in neuroscience research: is it a rat race? Dis Model Mech 9:1079–1087
El Yamani FZ, Yon L, Guérin M, El Ouezzani S, Alaoui A, Chartrel N, Anouar Y, Magoul R (2013) Immunocytochemical distribution of EM66 within the hypothalamic parvocellular paraventricular nucleus: colocalization with CRH and TRH but no plasticity related to acute stress and thyroidectomy in the rat. Reg Pept 182:28–34
Fall CHD, Kumaran K (2019) Metabolic programming in early life in humans. Philos Trans R Soc Lond B Biol Sci 374:20180123
Farebrother J, Zimmermann MB, Andersson M (2019) Excess iodine intake: sources, assessment, and effects on thyroid function. Ann N Y Acad Sci 1446:44–65
Farkas E, Varga E, Kovács B, Szilvásy-Szabó A, Cote-Vélez A, Péterfi Z, Matziari M, Tóth M, Zelena D, Mezriczky Z, Kádár A, Kővári D, Watanabe M, Kano M, Mackie K, Rózsa B, Ruska Y, Tóth B, Máté Z, Erdélyi F, Szabó G, Gereben B, Lechan RM, Charli JL, Joseph-Bravo P, Fekete C (2020) A glial-neuronal circuit in the median eminence regulates thyrotropin-releasing hormone-release via the endocannabinoid system. iScience 23:100921
Fekete C, Lechan RM (2014) Central regulation of hypothalamic-pituitary-thyroid axis under physiological and pathophysiological conditions. Endocr Rev 35:159–194
Fekete C, Mihály E, Luo LG, Kelly J, Clausen JT, Mao Q, Rand WM, Moss LG, Kuhar M, Emerson CH, Jackson IM, Lechan RM (2000) Association of cocaine- and amphetamine-regulated transcript-immunoreactive elements with thyrotropin-releasing hormone-synthesizing neurons in the hypothalamic paraventricular nucleus and its role in the regulation of the hypothalamic-pituitary-thyroid axis during fasting. J Neurosci 20:9224–9234
Fekete C, Sarkar S, Rand WM, Harney JW, Emerson CH, Bianco AC, Lechan RM (2002a) Agouti-related protein (AGRP) has a central inhibitory action on the hypothalamic-pituitary-thyroid (HPT) axis; comparisons between the effect of AGRP and neuropeptide Y on energy homeostasis and the HPT axis. Endocrinology 143:3846–3853
Fekete C, Wittmann G, Liposits Z, Lechan RM (2002b) GABA-ergic innervation of thyrotropin-releasing hormone-synthesizing neurons in the hypothalamic paraventricular nucleus of the rat. Brain Res 957:251–258
Ferland CL, Schrader LA (2011) Cage mate separation in pair-housed male rats evokes an acute stress corticosterone response. Neurosci Lett 489:154–158
Fink G, Koch Y, Ben Aroya N (1982) Release of thyrotropin releasing hormone into hypophysial portal blood is high relative to other neuropeptides and may be related to prolactin secretion. Brain Res 243:186–189
Flamant F, Cheng SY, Hollenberg AN, Moeller LC, Samarut J, Wondisford FE, Yen PM, Refetoff S (2017) Thyroid hormone signaling pathways: time for a more precise nomenclature. Endocrinology 158:2052–2057
Fliers E, Boelen A (2020) An update on non-thyroidal illness syndrome. J Endocrinol Invest. https://doi.org/10.1007/s40618-020-01482-4
Fliers E, Boelen A, van Trotsenburg AS (2014) Central regulation of the hypothalamo-pituitary-thyroid (HPT) axis: focus on clinical aspects. Handb Clin Neurol 124:127–138
Fonseca TL, Correa-Medina M, Campos MP, Wittmann G, Werneck-de-Castro JP, Arrojo e Drigo R, Mora-Garzon M, Ueta CB, Caicedo A, Fekete C, Gereben B, Lechan RM, Bianco AC (2013) Coordination of hypothalamic and pituitary T3 production regulates TSH expression. J Clin Invest 123:1492–1500
Fortunato RS, Ignácio DL, Padron AS, Peçanha R, Marassi MP, Rosenthal D, Werneck-de-Castro JP, Carvalho DP (2008) The effect of acute exercise session on thyroid hormone economy in rats. J Endocrinol 198:347–353
Füzesi T, Wittmann G, Lechan RM, Liposits Z, Fekete C (2009) Noradrenergic innervation of hypophysiotropic thyrotropin-releasing hormone-synthesizing neurons in rats. Brain Res 1294:38–44
Gervasi N, Hepp R, Tricoire L, Zhang J, Lambolez B, Paupardin-Tritsch D, Vincent P (2007) Dynamics of protein kinase A signaling at the membrane, in the cytosol, and in the nucleus of neurons in mouse brain slices. J Neurosci 27:2744–2750
Giammanco M, Di Liegro CM, Schiera G, Di Liegro I (2020) Genomic and non-genomic mechanisms of action of thyroid hormones and their catabolite 3,5-diiodo-L-thyronine in mammals. Int J Mol Sci 21:4140
Giraud P, Maltèse JY, Boudouresque F, Salers P, Ouafik L, Renard M, Pelen F, Oliver C (1992) Peptidylglycine alpha-amidating monooxygenase activity and TRH and CRF biosynthesis. Role of copper. Biol Trace Elem Res 32:293–301
Goodman T, Hajihosseini MK (2015) Hypothalamic tanycytes-masters and servants of metabolic, neuroendocrine, and neurogenic functions. Front Neurosci 9:387
Gore AC, Krishnan K, Reilly MP (2019) Endocrine-disrupting chemicals: effects on neuroendocrine systems and the neurobiology of social behavior. Horm Behav 111:7–22
Goshu E, Jin H, Lovejoy J, Marion JF, Michaud JL, Fan CM (2004) Sim2 contributes to neuroendocrine hormone gene expression in the anterior hypothalamus. Mol Endocrinol 18:1251–1262
Grattan DR (2015) 60 Years of neuroendocrinology: the hypothalamo-prolactin axis. J Endocrinol 226:T101–T122
Grøntved L, Waterfall JJ, Kim DW, Baek S, Sung MH, Zhao L, Park JW, Nielsen R, Walker RL, Zhu YJ, Meltzer PS, Hager GL, Cheng SY (2015) Transcriptional activation by the thyroid hormone receptor through ligand-dependent receptor recruitment and chromatin remodelling. Nat Commun 6:7048
Guo F, Bakal K, Minokoshi Y, Hollenberg AN (2004) Leptin signaling targets the thyrotropin-releasing hormone gene promoter in vivo. Endocrinology 145:2221–2227
Han Z, Li Y, Zhang S, Song N, Xu H, Dang Y, Liu C, Giesy JP, Yu H (2017) Prenatal transfer of decabromodiphenyl ether (BDE-209) results in disruption of the thyroid system and developmental toxicity in zebrafish offspring. Aquat Toxicol 190:46–52
Harno E, Gali Ramamoorthy T, Coll AP, White A (2018) POMC: the physiological power of hormone processing. Physiol Rev 98:2381–2430
Harris M, Aschkenasi C, Elias CF, Chandrankunnel A, Nillni EA, Bjøorbaek C, Elmquist JK, Flier JS, Hollenberg AN (2001) Transcriptional regulation of the thyrotropin-releasing hormone gene by leptin and melanocortin signaling. J Clin Invest 107:111–120
Heuer H, Schäfer MK, O’Donnell D, Walker P, Bauer K (2000) Expression of thyrotropin-releasing hormone receptor 2 (TRH-R2) in the central nervous system of rats. J Comp Neurol 428:319–336
Hoermann R, Midgley JEM, Larisch R, Dietrich JW (2015) Homeostatic control of the thyroid–pituitary axis: perspectives for diagnosis and treatment. Front Endocrinol 6:177
Hollenberg AN, Monden T, Flynn TR, Boers ME, Cohen O, Wondisford FE (1995) The human thyrotropin-releasing hormone gene is regulated by thyroid hormone through two distinct classes of negative thyroid hormone response elements. Mol Endocrinol 9:540–550
Hrabovszky E, Wittmann G, Turi GF, Liposits Z, Fekete C (2005) Hypophysiotropic thyrotropin-releasing hormone and corticotropin-releasing hormone neurons of the rat contain vesicular glutamate transporter-2. Endocrinology 146:341–347
Hubalewska-Dydejczyk A, Duntas L, Gilis-Januszewska A (2020) Pregnancy, thyroid, and the potential use of selenium. Hormones 19:47–53
Huo L, Münzberg H, Nillni EA, Bjørbaek C (2004) Role of signal transducer and activator of transcription 3 in regulation of hypothalamic trh gene expression by leptin. Endocrinology 145:2516–2523
Ishii S, Yamada M, Satoh T, Monden T, Hashimoto K, Shibusawa N, Onigata K, Morikawa A, Mori M (2004) Aberrant dynamics of histone deacetylation at the thyrotropin-releasing hormone gene in resistance to thyroid hormone. Mol Endocrinol 18:1708–1720
Jacobs LS, Snyder PJ, Wilber JF, Utiger RD, Daughaday WH (1971) Increased serum prolactin after administration of synthetic thyrotropin releasing hormone (TRH) in man. J Clin Endocrinol Metab 33:996–998
Jaimes-Hoy L, Gutiérrez-Mariscal M, Vargas Y, Pérez-Maldonado A, Romero F, Sánchez-Jaramillo E, Charli JL, Joseph-Bravo P (2016) Neonatal maternal separation alters, in a sex-specific manner, the expression of TRH, of TRH-degrading ectoenzyme in the rat hypothalamus, and the response of the thyroid axis to starvation. Endocrinology 157:3253–3265
Jaimes-Hoy L, Romero F, Charli JL, Joseph-Bravo P (2019) Sex dimorphic responses of the hypothalamus-pituitary-thyroid axis to maternal separation and palatable diet. Front Endocrinol (Lausanne) 10:445
Jaimes-Hoy L, Pérez-Maldonado A, Narváez Bahena E, de la Cruz GN, Rodríguez-Rodríguez A, Charli JL, Soberón X, Joseph-Bravo P (2021) Sex dimorphic changes in trh gene methylation and thyroid-axis response to energy demands in maternally separated rats. Endocrinol 162:1–18
Jing E, Nillni EA, Sanchez VC, Stuart RC, Good DJ (2004) Deletion of the Nhlh2 transcription factor decreases the levels of the anorexigenic peptides alpha melanocyte-stimulating hormone and thyrotropin-releasing hormone and implicates prohormone convertases I and II in obesity. Endocrinology 145:1503–1513
Joëls M, Sarabdjitsingh RA, Karst H (2012) Unraveling the time domains of corticosteroid hormone influences on brain activity: rapid, slow, and chronic modes. Pharmacol Rev 64:901–938
Joseph-Bravo P, Charli JL, Palacios JM, Kordon C (1979) Effect of neurotransmitters on the in vitro release of immunoreactive thyrotropin-releasing hormone from rat mediobasal hypothalamus. Endocrinology 104:801–806
Joseph-Bravo P, Jaimes-Hoy L, Uribe RM, Charli JL (2015a) 60 Years of neuroendocrinology: TRH, the first hypophysiotropic releasing hormone isolated: control of the pituitary-thyroid axis. J Endocrinol 226:T85–T100
Joseph-Bravo P, Jaimes-Hoy L, Charli JL (2015b) Regulation of TRH neurons and energy homeostasis-related signals under stress. J Endocrinol 224:R139–R159
Joseph-Bravo P, Jaimes-Hoy L, Charli JL (2016) Advances in TRH signaling. Rev Endocr Metab Disord 17:545–558
Joseph-Bravo P, Gutiérrez-Mariscal M, Jaimes-Hoy L, Charli JL (2017) Thyroid axis and energy balance: focus on animals and implications for humankind. In: Preedy V, Patel V (eds) Handbook of famine, starvation, and nutrient deprivation. Springer, Cham. https://doi.org/10.1007/978-3-319-40007-5_76-1
Joseph-Bravo P, Lazcano I, Jaimes-Hoy L, Gutierrez-Mariscal M, Sanchez-Jaramillo E, Uribe RM, Charli JL (2020) Sexually dimorphic dynamics of thyroid axis activity during fasting in rats. Front Biosci (Landmark Ed) 25:1305–1323
Kaczmarek MM, Mendoza T, Kozak LP (2016) Lactation undernutrition leads to multigenerational molecular programming of hypothalamic gene networks controlling reproduction. BMC Genomics 17:333
Kádár A, Sánchez E, Wittmann G, Singru PS, Füzesi T, Marsili A, Larsen PR, Liposits Z, Lechan RM, Fekete C (2010) Distribution of hypophysiotropic thyrotropin-releasing hormone (TRH)-synthesizing neurons in the hypothalamic paraventricular nucleus of the mouse. J Comp Neurol 518:3948–3961
Kalló I, Mohácsik P, Vida B, Zeöld A, Bardóczi Z, Zavacki AM, Farkas E, Kádár A, Hrabovszky E, Arrojo E, Drigo R, Dong L, Barna L, Palkovits M, Borsay BA, Herczeg L, Lechan RM, Bianco AC, Liposits Z, Fekete C, Gereben B (2012) A novel pathway regulates thyroid hormone availability in rat and human hypothalamic neurosecretory neurons. PLoS One 7:e37860
Kalsbeek A, Fliers E, Franke AN, Wortel J, Buijs RM (2000) Functional connections between the suprachiasmatic nucleus and the thyroid gland as revealed by lesioning and viral tracing techniques in the rat. Endocrinology 141:3832–3841
Kim JS, Iremonger KJ (2019) Temporally tuned corticosteroid feedback regulation of the stress axis. Trends Endocrinol Metab 30:11
Klieverik LP, Coomans CP, Endert E, Sauerwein HP, Havekes LM, Voshol PJ, Rensen PC, Romijn JA, Kalsbeek A, Fliers E (2009) Thyroid hormone effects on whole-body energy homeostasis and tissue-specific fatty acid uptake in vivo. Endocrinology 150:5639–5648
Kohno D, Nakata M, Maejima Y, Shimizu H, Sedbazar U, Yoshida N, Dezaki K, Onaka T, Mori M, Yada T (2008) Nesfatin-1 neurons in paraventricular and supraoptic nuclei of the rat hypothalamus coexpress oxytocin and vasopressin and are activated by refeeding. Endocrinology 149:1295–1301
Koolhaas JM, Meerlo P, De Boer SF, Strubbe JH, Bohus B (1997) The temporal dynamics of the stress response. Neurosci Biobehav Rev 21:775–782
Kouidhi S, Clerget-Froidevaux MS (2018) Integrating thyroid hormone signaling in hypothalamic control of metabolism: crosstalk between nuclear receptors. Int J Mol Sci 19:2017
Krmpot AJ, Nikolić SN, Oasa S, Papadopoulos DK, Vitali M, Oura M, Mikuni S, Thyberg P, Tisa S, Kinjo M, Nilsson L, Terenius L, Rigler R, Vukojević V (2019) Functional fluorescence microscopy imaging: quantitative scanning-free confocal fluorescence microscopy for the characterization of fast dynamic processes in live cells. Anal Chem 91:11129–11137
Kuroda G, Sasaki S, Matsushita A, Ohba K, Sakai Y, Shinkai S, Nakamura HM, Yamagishi S, Sato K, Hirahara N, Oki Y, Ito M, Suzuki T, Suda T (2020) GATA2 mediates the negative regulation of the prepro-thyrotropin-releasing hormone gene by liganded T3 receptor β2 in the rat hypothalamic paraventricular nucleus. PLoS One 15:e0242380
Lazcano I, Cabral A, Uribe RM, Jaimes-Hoy L, Perello M, Joseph-Bravo P, Sánchez-Jaramillo E, Charli JL (2015) Fasting enhances pyroglutamyl peptidase II activity in tanycytes of the mediobasal hypothalamus of male adult rats. Endocrinology 156:2713–2723
Lechan RM, Jackson IM (1982) Immunohistochemical localization of thyrotropin-releasing hormone in the rat hypothalamus and pituitary. Endocrinology 111:55–65
Lechan RM, Wu P, Jackson IM, Wolf H, Cooperman S, Mandel G, Goodman RH (1986) Thyrotropin-releasing hormone precursor: characterization in rat brain. Science 231:159–161
Lechan RM, Wu P, Jackson IM (1987) Immunocytochemical distribution in rat brain of putative peptides derived from thyrotropin-releasing hormone prohormone. Endocrinol 121:1879–1891
Lechan RM, Qi Y, Jackson IM, Mahdavi V (1994) Identification of thyroid hormone receptor isoforms in thyrotropin-releasing hormone neurons of the hypothalamic paraventricular nucleus. Endocrinology 135:92–100
Lee SL, Stewart K, Goodman RH (1988) Structure of the gene encoding rat thyrotropin releasing hormone. J Biol Chem 263:16604–16609
Lee SK, Lee S, Shin SY, Ryu PD, Lee SY (2012) Single cell analysis of voltage-gated potassium channels that determines neuronal types of rat hypothalamic paraventricular nucleus neurons. Neuroscience 205:49–62
Légrádi G, Lechan RM (1998) The arcuate nucleus is the major source for neuropeptide Y-innervation of thyrotropin-releasing hormone neurons in the hypothalamic paraventricular nucleus. Endocrinology 139:3262–3270
Légrádi G, Lechan RM (1999) Agouti-related protein containing nerve terminals innervate thyrotropin-releasing hormone neurons in the hypothalamic paraventricular nucleus. Endocrinology 140:3643–3652
Légrádi G, Hannibal J, Lechan RM (1997) Association between pituitary adenylate cyclase-activating polypeptide and thyrotropin-releasing hormone in the rat hypothalamus. J Chem Neuroanat 13:265–279
Liao N, Bulant M, Nicolas P, Vaudry H, Pelletier G (1991) Anatomical interactions of proopiomelanocortin (POMC)-related peptides, neuropeptide Y (NPY) and dopamine beta-hydroxylase (D beta H) fibers and thyrotropin-releasing hormone (TRH) neurons in the paraventricular nucleus of rat hypothalamus. Neuropeptides 18:63–67
Liao N, Vaudry H, Pelletier G (1992) Neuroanatomical connections between corticotropin-releasing factor (CRF) and somatostatin (SRIF) nerve endings and thyrotropin-releasing hormone (TRH) neurons in the paraventricular nucleus of rat hypothalamus. Peptides 13:677–680
Lima LP, Barros IA, Lisbôa PC, Araújo RL, Silva AC, Rosenthal D, Ferreira AC, Carvalho DP (2006) Estrogen effects on thyroid iodide uptake and thyroperoxidase activity in normal and ovariectomized rats. Steroids 71:653–659
Lisboa PC, Fagundes AT, Denolato AT, Oliveira E, Bonomo IT, Alves SB, Curty FH, Passos MC, Moura EG (2008) Neonatal low-protein diet changes deiodinase activities and pituitary TSH response to TRH in adult rats. Exp Biol Med (Maywood) 233:57–63
Lisboa PC, Conceição EP, de Oliveira E, Moura EG (2015) Postnatal overnutrition programs the thyroid hormone metabolism and function in adulthood. J Endocrinol 226:219–226
Lyons DJ, Horjales-Araujo E, Broberger C (2010) Synchronized network oscillations in rat tuberoinfundibular dopamine neurons: switch to tonic discharge by thyrotropin-releasing hormone. Neuron 65:217–229
Machado TD, Salum GA, Bosa VL, Goldani MZ, Meaney MJ, Agranonik M, Manfro GG, Silveira PP (2015) Early life trauma is associated with decreased peripheral levels of thyroid-hormone T3 in adolescents. Int J Dev Neurosci 47:304–308
Markakis EA, Swanson LW (1997) Spatiotemporal patterns of secretomotor neuron generation in the parvicellular neuroendocrine system. Brain Res Brain Res Rev 24:255–291
Martínez-Armenta M, Díaz de León-Guerrero S, Catalán A, Alvarez-Arellano L, Uribe RM, Subramaniam M, Charli JL, Pérez-Martínez L (2015) TGFβ2 regulates hypothalamic Trh expression through the TGFβ inducible early gene-1 (TIEG1) during fetal development. Mol Cell Endocrinol 400:129–139
Martinez de la Escalera G, Weiner RI (1992) Dissociation of dopamine from its receptor as a signal in the pleiotropic hypothalamic regulation of prolactin secretion. Endocr Rev 13:241–255
Mauvais-Jarvis F (2015) Sex differences in metabolic homeostasis, diabetes, and obesity. Biol Sex Differ 6:14
McAninch EA, Bianco AC (2014) Thyroid hormone signaling in energy homeostasis and energy metabolism. Ann N Y Acad Sci 1311:77–87
Mendoza A, Hollenberg AN (2017) New insights into thyroid hormone action. Pharmacol Ther 173:135–145
Minakhina S, Bansal S, Zhang A, Brotherton M, Janodia R, De Oliveira V, Tadepalli S, Wondisford FE (2020) A direct comparison of thyroid hormone receptor protein levels in mice provides unexpected insights into thyroid hormone action. Thyroid 30:1193–1204
Miranda RA, Gaspar de Moura E, Lisboa PC (2020) Tobacco smoking during breastfeeding increases the risk of developing metabolic syndrome in adulthood: lessons from experimental models. Food Chem Toxicol 144:111623
Moisiadis VG, Constantinof A, Kostaki A, Szyf M, Matthews SG (2017) Prenatal glucocorticoid exposure modifies endocrine function and behaviour for 3 generations following maternal and paternal transmission. Sci Rep 7:11814
Moog NK, Entringer S, Heim C, Wadhwa PD, Kathmann N, Buss C (2017a) Influence of maternal thyroid hormones during gestation on fetal brain development. Neuroscience 342:68–100
Moog NK, Heim CM, Entringer S, Kathmann N, Wadhwa PD, Buss C (2017b) Childhood maltreatment is associated with increased risk of subclinical hypothyroidism in pregnancy. Psychoneuroendocrinology 84:190–196
Morreale de Escobar G, Obregon MJ, Escobar del Rey F (1987) Fetal and maternal thyroid hormones. Horm Res 26:12–27
Mughal BB, Fini JB, Demeneix BA (2018) Thyroid-disrupting chemicals and brain development: an update. Endocr Connect 7:R160–R186
Müller-Fielitz H, Stahr M, Bernau M, Richter M, Abele S, Krajka V, Benzin A, Wenzel J, Kalies K, Mittag J, Heuer H, Offermanns S, Schwaninger M (2017) Tanycytes control the hormonal output of the hypothalamic-pituitary-thyroid axis. Nat Commun 8:484
Mullur R, Liu YY, Brent GA (2014) Thyroid hormone regulation of metabolism. Physiol Rev 94:355–382
Nakato R, Sakata T (2020) Methods for ChIP-seq analysis: a practical workflow and advanced applications. Methods. https://doi.org/10.1016/j.ymeth.2020.03.005
Nedergaard J, Cannon B (2018) Brown adipose tissue as a heat producing thermo effector. Handb Clin Neurol 156:137–152
Nillni EA (2010) Regulation of the hypothalamic thyrotropin releasing hormone (TRH) neuron by neuronal and peripheral inputs. Front Neuroendocrinol 31:134–156
Nomura S, Tricoire L, Cohen I, Kuhn B, Lambolez B, Hepp R (2020) Combined optogenetic approaches reveal quantitative dynamics of endogenous noradrenergic transmission in the brain. iScience 23:101710
Osterlund C, Spencer RL (2011) Corticosterone pretreatment suppresses stress-induced hypothalamic-pituitary-adrenal axis activity via multiple actions that vary with time, site of action, and de novo protein synthesis. J Endocrinol 208:311–322
Pałkowska-Goździk E, Lachowicz K, Rosołowska-Huszcz D (2017) Effects of dietary protein on thyroid axis activity. Nutrients 10:5
Parra-Montes de Oca MA, Gutiérrez-Mariscal M, Salmerón-Jiménez MF, Jaimes-Hoy L, Charli JL, Joseph-Bravo P (2019) Voluntary exercise-induced activation of thyroid axis and reduction of white fat depots is attenuated by chronic stress in a sex dimorphic pattern in adult rats. Front Endocrinol (Lausanne) 10:418
Paxinos G, Watson C (2004) The rat brain in stereotaxic coordinates—the new coronal set, 5th edn. Elsevier Academic
Payne AH, Hales DB (2004) Overview of steroidogenic enzymes in the pathway from cholesterol to active steroid hormones. Endocr Rev 25:947–970
Pearce EN, Lazarus JH, Moreno-Reyes R, Zimmermann MB (2016) Consequences of iodine deficiency and excess in pregnant women: an overview of current knowns and unknowns. Am J Clin Nutr 104:918S–923S
Pekary AE, Knoble M, Garcia NH, Bhasin S, Hershman JM (1990) Testosterone regulates the secretion of thyrotrophin-releasing hormone (TRH) and TRH precursor in the rat hypothalamic-pituitary axis. J Endocrinol 125:263–270
Perello M, Nillni EA (2007) The biosynthesis and processing of neuropeptides: lessons from prothyrotropin releasing hormone (proTRH). Front Biosci 12:3554–3565
Perello M, Stuart RC, Vaslet CA, Nillni EA (2007) Cold exposure increases the biosynthesis and proteolytic processing of prothyrotropin-releasing hormone in the hypothalamic paraventricular nucleus via beta-adrenoreceptors. Endocrinology 148:4952–4964
Perello M, Cakir I, Cyr NE, Romero A, Stuart RC, Chiappini F, Hollenberg AN, Nillni EA (2010) Maintenance of the thyroid axis during diet-induced obesity in rodents is controlled at the central level. Am J Physiol Endocrinol Metab 299:E976–E989
Pérez-Monter C, Martínez-Armenta M, Miquelajauregui A, Furlan-Magaril M, Varela-Echavarría A, Recillas-Targa F, May V, Charli JL, Pérez-Martínez L (2011) The Krüppel-like factor 4 controls biosynthesis of thyrotropin-releasing hormone during hypothalamus development. Mol Cell Endocrinol 333:127–133
Péterfi Z, Farkas E, Nagyunyomi-Sényi K, Kádár A, Ottó S, Horváth A, Füzesi T, Lechan RM, Fekete C (2018) Role of TRH/UCN3 neurons of the perifornical area/bed nucleus of stria terminalis region in the regulation of the anorexigenic POMC neurons of the arcuate nucleus in male mice and rats. Brain Struct Funct 223:1329–1341
Pinto VMS, Minakhina S, Qiu S, Sidhaye A, Brotherton MP, Suhotliv A, Wondisford FE (2017) Naturally occurring amino acids in helix 10 of the thyroid hormone receptor mediate isoform-specific TH gene regulation. Endocrinology 158:3067–3078
Prevot V, Dehouck B, Sharif A, Ciofi P, Giacobini P, Clasadonte J (2018) The versatile tanycyte: a hypothalamic integrator of reproduction and energy metabolism. Endocr Rev 39:333–368
Rabeler R, Mittag J, Geffers L, Rüther U, Leitges M, Parlow AF, Visser TJ, Bauer K (2004) Generation of thyrotropin-releasing hormone receptor 1-deficient mice as an animal model of central hypothyroidism. Mol Endocrinol 18:1450–1460
Radley JJ, Sawchenko PE (2015) Evidence for involvement of a limbic paraventricular hypothalamic inhibitory network in hypothalamic-pituitary-adrenal axis adaptations to repeated stress. J Comp Neurol 523:2769–2787
Ramadoss P, Abraham BJ, Tsai L, Zhou Y, Costa-e-Sousa RH, Ye F, Bilban M, Zhao K, Hollenberg AN (2014) Novel mechanism of positive versus negative regulation by thyroid hormone receptor β1 (TRβ1) identified by genome-wide profiling of binding sites in mouse liver. J Biol Chem 289:1313–1328
Raptis S, Fekete C, Sarkar S, Rand WM, Emerson CH, Nagy GM, Lechan RM (2004) Cocaine- and amphetamine-regulated transcript co-contained in thyrotropin-releasing hormone (TRH) neurons of the hypothalamic paraventricular nucleus modulates TRH-induced prolactin secretion. Endocrinology 145:1695–1699
Rezaei M, Javadmoosavi SY, Mansouri B, Azadi NA, Mehrpour O, Nakhaee S (2019) Thyroid dysfunction: how concentration of toxic and essential elements contribute to risk of hypothyroidism, hyperthyroidism, and thyroid cancer. Environ Sci Pollut Res Int 26:35787–35796
Rodríguez-Rodríguez A, Lazcano I, Sánchez-Jaramillo E, Uribe RM, Jaimes-Hoy L, Joseph-Bravo P, Charli JL (2019) Tanycytes and the control of thyrotropin-releasing hormone flux into portal capillaries. Front Endocrinol (Lausanne) 10:401
Ross DS (1990) Testosterone increases TSH-β mRNA, and modulates α-subunit mRNA differentially in mouse thyrotropic tumor and castrate rat pituitary. Horm Metab Res 22:163–169
Sánchez E, Charli JL, Morales C, Corkidi G, Seidah NG, Joseph-Bravo P, Uribe RM (1997) Expression of the proprotein convertases PC1 and PC2 mRNAs in thyrotropin releasing hormone neurons of the rat paraventricular nucleus of hypothalamus. Brain Res 761:77–86
Sánchez E, Uribe RM, Corkidi G, Zoeller RT, Cisneros M, Zacarias M, Morales-Chapa C, Charli JL, Joseph-Bravo P (2001) Differential responses of thyrotropin-releasing hormone (TRH) neurons to cold exposure or suckling indicate functional heterogeneity of the TRH system in the paraventricular nucleus of the rat hypothalamus. Neuroendocrinology 74:407–422
Sánchez E, Fekete C, Lechan RM, Joseph-Bravo P (2007) Cocaine- and amphetamine-regulated transcript (CART) expression is differentially regulated in the hypothalamic paraventricular nucleus of lactating rats exposed to suckling or cold stimulation. Brain Res 1132:120–128
Sánchez E, Vargas MA, Singru PS, Pascual I, Romero F, Fekete C, Charli JL, Lechan RM (2009) Tanycyte pyroglutamyl peptidase II contributes to regulation of the hypothalamic-pituitary-thyroid axis through glial-axonal associations in the median eminence. Endocrinology 150:2283–2291
Sanchez Jimenez JG, De Jesus O (2020) Hypothalamic dysfunction. In: StatPearls [Internet]. StatPearls, Treasure Island, FL. PMID: 32809578
Sasaki S, Matsushita A, Kuroda G, Nakamura HM, Oki Y, Suda T (2018) The mechanism of negative transcriptional regulation by thyroid hormone: lessons from the thyrotropin β subunit gene. Vitam Horm 106:97–127
Sharkey M, Harrad S, Abou-Elwafa Abdallah M, Drage DS, Berresheim H (2020) Phasing-out of legacy brominated flame retardants: the UNEP Stockholm Convention and other legislative action worldwide. Environ Int 144:106041
Simmons DM, Swanson LW (2009) Comparison of the spatial distribution of seven types of neuroendocrine neurons in the rat paraventricular nucleus: toward a global 3D model. J Comp Neurol 516:423–441
Sinai C, Hirvikoski T, Nordström AL, Nordström P, Nilsonne A, Wilczek A, Asberg M, Jokinen J (2014) Hypothalamic pituitary thyroid axis and exposure to interpersonal violence in childhood among women with borderline personality disorder. Eur J Psychotraumatol 5. https://doi.org/10.3402/ejpt.v5.23911
Smith MA, Makino S, Kim SY, Kvetnansky R (1995) Stress increases brain-derived neurotropic factor messenger ribonucleic acid in the hypothalamus and pituitary. Endocrinology 136:3743–3750
Sotelo-Rivera I, Jaimes-Hoy L, Cote-Vélez A, Espinoza-Ayala C, Charli JL, Joseph-Bravo P (2014) An acute injection of corticosterone increases thyrotrophin-releasing hormone expression in the paraventricular nucleus of the hypothalamus but interferes with the rapid hypothalamus pituitary thyroid axis response to cold in male rats. J Neuroendocrinol 26:861–869
Sotelo-Rivera I, Cote-Vélez A, Uribe RM, Charli JL, Joseph-Bravo P (2017) Glucocorticoids curtail stimuli-induced CREB phosphorylation in TRH neurons through interaction of the glucocorticoid receptor with the catalytic subunit of protein kinase A. Endocrine 55:861–871
Sugrue ML, Vella KR, Morales C, Lopez ME, Hollenberg AN (2010) The thyrotropin-releasing hormone gene is regulated by thyroid hormone at the level of transcription in vivo. Endocrinology 151:793–801
Suter MA, Sangi-Haghpeykar H, Showalter L, Shope C, Hu M, Brown K, Williams S, Harris RA, Grove KL, Lane RH, Aagaard KM (2012) Maternal high-fat diet modulates the fetal thyroid axis and thyroid gene expression in a nonhuman primate model. Mol Endocrinol 26:2071–2080
Suzuki S, Handa RJ (2005) Estrogen receptor-beta, but not estrogen receptor-alpha, is expressed in prolactin neurons of the female rat paraventricular and supraoptic nuclei: comparison with other neuropeptides. J Comp Neurol 484:28–42
Tashjian AH Jr, Barowsky NJ, Jensen DK (1971) Thyrotropin releasing hormone: direct evidence for stimulation of prolactin production by pituitary cells in culture. Biochem Biophys Res Commun 43:516–523
Toni R, Jackson IM, Lechan RM (1990a) Neuropeptide-Y-immunoreactive innervation of thyrotropin-releasing hormone-synthesizing neurons in the rat hypothalamic paraventricular nucleus. Endocrinol 126:2444–2453
Toni R, Jackson IMD, Lechan RM (1990b) Thyrotropin-releasing-hormone-immunoreactive innervation of thyrotropin releasing-hormone-tuberoinfundibular neurons in rat hypothalamus: anatomical basis to suggest ultrashort feedback regulation. Neuroendocrinol 52:422–428
Tran DN, Jung EM, Yoo YM, Lee JH, Jeung EB (2020) Perinatal exposure to triclosan results in abnormal brain development and behavior in mice. Int J Mol Sci 21:4009
Uribe RM, Redondo JL, Charli JL, Joseph-Bravo P (1993) Suckling and cold stress rapidly and transiently increase TRH mRNA in the paraventricular nucleus. Neuroendocrinology 58:140–145
Uribe RM, Zacarias M, Corkidi G, Cisneros M, Charli JL, Joseph-Bravo P (2009) 17β-Oestradiol indirectly inhibits thyrotrophin-releasing hormone expression in the hypothalamic paraventricular nucleus of female rats and blunts thyroid axis response to cold exposure. J Neuroendocrinol 21:439–448
Uribe RM, Jaimes-Hoy L, Ramírez-Martínez C, García-Vázquez A, Romero F, Cisneros M, Cote-Vélez A, Charli JL, Joseph-Bravo P (2014) Voluntary exercise adapts the hypothalamus-pituitary-thyroid axis in male rats. Endocrinology 155:2020–2030
van den Pol AN (2012) Neuropeptide transmission in brain circuits. Neuron 76:98–115
van der Laan S, de Kloet ER, Meijer OC (2009) Timing is critical for effective glucocorticoid receptor mediated repression of the cAMP-induced CRH gene. PLoS One 4:e4327
Vandevyver S, Dejager L, Libert C (2012) On the trail of the glucocorticoid receptor: into the nucleus and back. Traffic 13:364–374
van Haasteren GA, van Toor H, Klootwijk W, Handler B, Linkels E, van der Schoot P, van Ophemert J, de Jong FH, Visser TJ, de Greef WJ (1996) Studies on the role of TRH and corticosterone in the regulation of prolactin and thyrotrophin secretion during lactation. J Endocrinol 148:325–336
Varga E, Farkas E, Zséli G, Kádár A, Venczel A, Kővári D, Németh D, Máté Z, Erdélyi F, Horváth A, Szenci O, Watanbe M, Lechan RM, Gereben B, Fekete C (2019) Thyrotropin-releasing-hormone-synthesizing neurons of the hypothalamic paraventricular nucleus are inhibited by glycinergic inputs. Thyroid 29:1858–1868
Vella KR, Hollenberg AN (2017) The actions of thyroid hormone signaling in the nucleus. Mol Cell Endocrinol 458:127–135
Watanobe H, Endo K, Kitaoka M, Takebe K (1985) Sexual differentiation of prolactin responsiveness to thyrotropin releasing hormone (TRH) in the rat. Effects of postnatal testosterone on adenohypophyseal TRH receptor ontogenesis in male rats. J Endocrinol Invest 8:459–464
Watts AG (2015) 60 Years of neuroendocrinology: the structure of the neuroendocrine hypothalamus: the neuroanatomical legacy of Geoffrey Harris. J Endocrinol 226:T25–T39
Wittmann G, Liposits Z, Lechan RM, Fekete C (2002) Medullary adrenergic neurons contribute to the neuropeptide Y-ergic innervation of hypophysiotropic thyrotropin-releasing hormone-synthesizing neurons in the rat. Neurosci Lett 324:69–73
Wittmann G, Liposits Z, Lechan RM, Fekete C (2004a) Medullary adrenergic neurons contribute to the cocaine- and amphetamine-regulated transcript-immunoreactive innervation of thyrotropin-releasing hormone synthesizing neurons in the hypothalamic paraventricular nucleus. Brain Res 1006:1–7
Wittmann G, Sarkar S, Hrabovszky E, Liposits Z, Lechan RM, Fekete C (2004b) Galanin- but not galanin-like peptide-containing axon terminals innervate hypophysiotropic TRH-synthesizing neurons in the hypothalamic paraventricular nucleus. Brain Res 1002:43–50
Wittmann G, Lechan RM, Liposits Z, Fekete C (2005) Glutamatergic innervation of corticotropin-releasing hormone- and thyrotropin-releasing hormone-synthesizing neurons in the hypothalamic paraventricular nucleus of the rat. Brain Res 1039:53–62
Wittmann G, Füzesi T, Singru PS, Liposits Z, Lechan RM, Fekete C (2009) Efferent projections of thyrotropin-releasing hormone-synthesizing neurons residing in the anterior parvocellular subdivision of the hypothalamic paraventricular nucleus. J Comp Neurol 515:313–330
Xu Y, López M (2018) Central regulation of energy metabolism by estrogens. Mol Metab 15:104–115
Yamada M, Shibusawa N, Ishii S, Horiguchi K, Umezawa R, Hashimoto K, Monden T, Satoh T, Hirato J, Mori M (2006) Prolactin secretion in mice with thyrotropin-releasing hormone deficiency. Endocrinology 147:2591–2596
Ylli D, Klubo-Gwiezdzinska J, Wartofsky L (2020) Exercise and thyroid function. In: Hackney A, Constantini N (eds) Endocrinology of physical activity and sport. Contemporary endocrinology. Humana, Cham. https://doi.org/10.1007/978-3-030-33376-8_6
Yoo S, Cha D, Kim S, Jiang L, Cooke P, Adebesin M, Wolfe A, Riddle R, Aja S, Blackshaw S (2020) Tanycyte ablation in the arcuate nucleus and median eminence increases obesity susceptibility by increasing body fat content in male mice. Glia 68:1987–2000
Younes-Rapozo V, Moura EG, Manhães AC, Pinheiro CR, Santos-Silva AP, de Oliveira E, Lisboa PC (2013) Maternal nicotine exposure during lactation alters hypothalamic neuropeptides expression in the adult rat progeny. Food Chem Toxicol 58:158–168
Younes-Rapozo V, Moura EG, Manhães AC, Pinheiro CR, Carvalho JC, Barradas PC, de Oliveira E, Lisboa PC (2015) Neonatal nicotine exposure leads to hypothalamic gliosis in adult overweight rats. J Neuroendocrinol 27:887–898
Zeisel A, Hochgerner H, Lönnerberg P, Johnsson A, Memic F, van der Zwan J, Häring M, Braun E, Borm LE, La Manno G, Codeluppi S, Furlan A, Lee K, Skene N, Harris KD, Hjerling-Leffler J, Arenas E, Ernfors P, Marklund U, Linnarsson S (2018) Molecular architecture of the mouse nervous system. Cell 174:999–1014
Zoeller RT, Kabeer N, Albers HE (1990) Cold exposure elevates cellular levels of messenger ribonucleic acid encoding thyrotropin-releasing hormone in paraventricular nucleus despite elevated levels of thyroid hormones. Endocrinology 127:2955–2962
Zoeller RT, Simonyi A, Butnariu O, Fletcher DL, Rudeen PK, McCrone S, Petersen SL (1995) Effects of acute ethanol administration and cold exposure on the hypothalamic-pituitary-thyroid axis. Endocr 3:39–47
Acknowledgments
The authors thank the technical help of BSc F. Romero, BSc M. Cisneros, BSc E. Mata, BSc R. Rodríguez Bahena, B.Sc. A. Ocadiz, BA S. Ainsworth, BSc J.O. Arriaga, BSc G. Cabeza, M. Villa and S.V.Serrano (UNAM). Supported in part by grants from CONACYT (284883 to PJB, 254960 and PN562 to JLC, 128665 to RMU), and DGAPA-UNAM (IN213419 to PJB, IN208515 and IN209018 to JLC, IA201519 to LJH, IN215420 to RMU). MSc. Adair Rodríguez-Rodríguez, MSc. Marco Parra-Montes de Oca, fellows of the Postgraduate Program in Biochemical Sciences (UNAM), were supported by CONACYT and DGAPA fellowships.
Key References
-
Bianco et al. (2019) A comprehensive review of thyroid hormone signaling and the role of deiodinases.
-
Sánchez et al. (2009) Demonstration of the role of the TRH-degrading ectoenzyme in tanycytes of the median eminence.
-
Farkas et al. (2020) Regulation of TRH release by a glial-neuronal circuit in the median eminence.
-
Fekete et al. (2000) Characterization of TRH hypophysiotropic neurons in rat PVN.
-
Harris et al. (2001) Transcriptional regulation of TRH expression by leptin and αMSH.
-
Joseph-Bravo et al. (2015a, b) Historic account of TRH discovery and its role in HPT axis physiology.
-
Lechan et al. (1986) Characterization of pro-TRH sequence and localization of neurons expressing Trh mRNA in the brain.
-
Sotelo-Rivera et al. (2017) Identification of the mechanism of acute glucocorticoid interference on neuronal stimulation of TRH synthesis.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2021 The Author(s), under exclusive license to Springer Nature Switzerland AG
About this chapter

Cite this chapter
Joseph-Bravo, P., Jaimes-Hoy, L., Rodríguez-Rodríguez, A., Parra-Montes de Oca, M., Uribe, R.M., Charli, JL. (2021). Multifactorial Regulation of the Activity of Hypophysiotropic Thyrotropin-Releasing Hormone Neurons. In: Grinevich, V., Dobolyi, Á. (eds) Neuroanatomy of Neuroendocrine Systems. Masterclass in Neuroendocrinology, vol 12. Springer, Cham. https://doi.org/10.1007/978-3-030-86630-3_10
Download citation
DOI: https://doi.org/10.1007/978-3-030-86630-3_10
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-86629-7
Online ISBN: 978-3-030-86630-3
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)