
Abstract
Heparanase is upregulated in various tumors, and its expression is closely associated with tumor growth, angiogenesis and metastasis, which accomplishes this mainly through degrading heparan sulfate and releasing heparin-binding growth factors thereby influencing multiple signaling pathways. In addition to its enzymatic degrading activity, heparanase can act via its non-enzymatic mechanisms that directly regulate various signaling. This review mainly focuses on the expression levels and role of heparanase in gastric cancer, and multiple genes and mechanisms regulating heparanase expression in gastric cancer. Furthermore, the development of heparanase-targeted immunotherapy and its potential application for treating gastric cancer are discussed.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


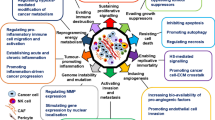
Keywords
1 Introduction
Heparanase is a 61.2 kDa protein with 543 amino acids encoded by its mRNA, and then this pro-enzyme is post-translationally cleaved into 8 and 50 kDa subunits that non-covalently associate to form the active heparanase [1, 2]. Heparanase is an endo-β-glucuronidase that cleaves heparan sulfate (HS) side chains, regulating the structure and function of heparan sulfate proteoglycans (HSPG) and remodeling cell surfaces and the extracellular matrix [3,4,5,6]. HSPGs mainly inhibit cellular invasion through promoting tight cell-cell, cell-ECM interactions, and self-assembly of the ECM [7, 8], which facilitate the biological activity of bound ligands (i.e., FGF, HGF, VEGF). Cleavage of HSPGs by heparanase could release these bound ligands and convert them into bioactive mediators, ensuring rapid tissue response.
In normal cells and tissues, heparanase is kept tightly regulated at transcriptional and post-translational levels as well [9], and the gene promoter of which is constitutively inhibited and the gene is not transcribed, largely due to promoter methylation [10,11,12]. Nevertheless, heparanase expression is enhanced in almost all cancers including ovarian, stomach, pancreas, colon, bladder, brain, prostate, breast, liver, myeloma and rhabdomyosarcoma and so on [13,14,15,16,17,18,19]. Multiple evidence indicate that heparanase could not only promote the breakdown of ECM but is also is involved in regulating the bioavailability and activity of growth factors and cytokines. Briefly, cleavage of HS by heparanase could promotes tumor progression via disassembly of extracellular barriers for cell invasion, release of HS-bound angiogenic and growth promoting factors, and induction of signal transduction pathways bound to heparan sulfate to promote growth and metastasis signaling [20,21,22] (see chapters xyz). Various studies showed that enhanced heparanase expression correlates with increased tumor size, tumor progression, advanced metastasis and poor prognosis [23,24,25]. Moreover, knockdown or inhibition of heparanase markedly impairs tumor progression further highlighting the role of heparanase in cancer progression and the potential role of anti-heparanase therapy for multiple types of cancer [26,27,28,29,30,31,32,33]. Recent studies demonstrate that a major function of heparanase in various cancers. This review will mainly focus on the role of heparanase in gastric cancer progression and immunotherapy.
2 Heparanase in Gastric Cancer Progression
2.1 Heparanase Expression in Gastric Cancer
The role of heparanase in gastric cancer has not been well elucidated. To date, several studies investigated the expression levels of heparanase in gastric cancers. Chen et al. investigated the expression of heparanase in gastric cancer tissues and non-cancerous gastric tissues and found that the expression of heparanase mRNA was positive in 29 cases of gastric cancer with a positive rate of 67.4%, while its expression rate was only 10% in non-cancerous gastric tissues [34]. And the Expression was correlated with the tumor size, serosal infiltration, lymph node metastasis, distant metastasis and TNM staging of gastric carcinomas [32]. Tang et al. [35] investigated the expression levels of heparanase mRNA of heparanase in 116 cases of gastric carcinomas and showed that heparanase mRNA was positive in 83% of cases, while no positive labeling was identified in normal gastric epithelium. Endo et al. [36] evaluated the heparanase mRNA expression in gastric cancer tissues and normal gastric tissues using qPCR analysis, and the positive rate in gastric cancer tissues was significantly higher than that in normal tissues. Our research group also examined the expression levels of heparanase by immunohistochemistry and found that the expression levels of heparanase were significantly higher in gastric cancer tissues than those in adjacent normal tissues [37]. Thus, various studies validated the higher expression of heparanase in gastric cancer tissues .
2.2 Heparanase in Gastric Cancer Metastasis and Progression
The prognosis of gastric cancer is poor due to the early invasion and metastasis which are the most common causes of death in gastric cancer [38]. It is generally known that the invasion of the basement membrane and extracellular matrix is one of the critical steps for cancer cell metastasis [39]. As an important component of the extracellular matrix (ECM), heparan sulfate proteoglycans (HSPGs) can serve as extracellular barrier and functional receptor coupling with various growth factors. Thus, degradation of HSPGs may plays a critical role in cancer cell invasion and metastasis. Heparanase is an endo-β-glucuronidase that cleaves heparan sulfate (HS) side chains, thus facilitating disassembly of the ECM and enhancing cell invasion, suggesting heparanase plays an important role in tumor invasion and metastasis. So far, The role of heparanase in tumor development and progression is well documented. Wang et al. [40, 41] reported that heparanase expression in primary gastric carcinoma cells was related to the metastatic behavior of gastric cancer. Similarly, Xie et al. [42] found that heparanase mRNA expression was significantly correlated with invasion and TNM stage of gastric cancer. Our research group also reported that heparanase is weakly expressed in the normal gastric tissues but is significantly increased in gastric cancer tissues. and the Higher expression of heparanase was associated with advanced TNM stage and depth of invasion [43] and, importantly, shorter survival time post-operation [43]. Nevertheless Notably, specific silencing of the heparanase gene significantly suppressed the adhesion, invasion, and metastasis of gastric cancer cells in vitro [44, 45]. Taken together, heparanase is overexpressed in gastric cancer and greatly associated with the invasion and metastasis of gastri cancer metastasis and poor prognosis. While inhibition of heparanase could suppress gastric cancer invasion.
2.3 Regulation of Heparanase Expression in Gastric Cancer
The vast majority of studies showed that heparanase was overexpressed in gastric cancer cells and played a key role in tumor invasion and metastasis. Various studies also focused on the regulatory mechanisms of heparanase expression in gastric cancer. Cao et al. [46] found that NF-ĸB signaling was significantly activated in gastric cancer tissues, and the activation was related to increased heparanase gene expression and correlated with poor clinicopathological characteristics such as lymphatic invasion, pathological stage, and depth of invasion, suggesting that NF-ĸB signaling is a major controller for regulator of heparanase expression in gastric cancer. Moreover, our research group further reported that HGF, a growth factor that binds HSPGs, could significantly increase heparanase expression at both mRNA and protein levels through PI3K/Akt/NF-ĸB signaling pathway and finally promote gastric cancer metastasis [43]. We also found that telomerase reverse transcriptase (TERT) could act as a co-activator of c-Myc to transactivate heparanase promoter activity, upregulate heparanase expression in gastric cancer cells, and promote invasion and metastasis of gastric cancer [37]. MicroRNA (miRNA) as small non-coding RNA molecules with 18–25 nucleotides post-transcriptionally regulate gene expression in various cancer types [47, 48]. Shi et al. [49] reported that miR-1258 could act as a tumor suppressor to inhibit invasion and metastasis by inhibiting heparanase expression. miR-429 also acts as a tumor-suppressor gene to inhibit transcription and translation of the heparanase gene, and reduce the invasion ability of gastric cancer cells by downregulating heparanase expression [50]. On the other hand, miR-558 recognizes its complementary site within the heparanase promoter to decrease the binding of Smad4, and hence activate the transcription and expression of heparanase in gastric cancer cell lines [51]. These findings demonstrate that different genes facilitate the progression of gastric cancer through directly regulating heparanase expression.
3 Heparanase as an Immunotherapeutic Target in Gastric Cancer
Nowadays, immunotherapy has emerged as a novel strategy for cancer therapy because of its weak side effects and targeting characteristics. One of the key components of immunotherapy is using the immune cells to be loaded with the tumor-associated antigens (TAAs) to induce antigen-specific anti-tumor immunity [52,53,54]. An ideal TAA is supposed to be uniquely expressed in tumors, which can induce not only antitumor immunity but also have crucial functional roles in tumor development [55, 56]. It is well documented that heparanase is overexpressed in almost all malignant tumors, and higher expression is associated with tumor progression and poor prognosis [57, 58]. Therefore, heparanase has been recognized as a suitable universal TAA in because of its crucial role in progression and metastasis of multiple tumors.
3.1 Heparanase Gene-Based Immunotherapy
Dendritic cells (DCs), the most efficient antigen-presenting cells (APCs), have an important role in the initiation and regulation of tumor-specific immune responses, leading to the rapid development of DC-based cancer immunotherapy. Genetic modification of DCs with TAA genes is an effective strategy for activating DCs in tumor immunotherapy [59, 60]. Our research group firstly used the recombinant adenovirus vector containing the full-length cDNA of heparanase (rAd-Hpa) to transfect DCs from peripheral blood mononuclear cells of healthy HLA-A2-positive donors to generate heparanase gene-modified DC vaccine [61]. Then, this genetically modified DC vaccine was used to activate T lymphocytes from the same donors to generate heparanase- specific cytotoxic T lymphocytes (CTLs) . The study showed that the modified DCs activate heparanase-specific CTLs, resulting in specific lysis of human gastric cancer KATO-III cells that were heparanase positive and HLA-A2 matched, while there was no killing effect on SGC-7901 cells that were heparanase positive but not HLA-A2 matched . Meanwhile, the studies revealed that the modified DCs can increase interferon IFN-γ secretion by the CTL cells to enhance non-specific immunological killing [61].
3.2 Heparanase Peptide-Based Immunotherapy
CTLs play key roles in tumor immunosurveillance through recognition of TAAs expressed on the surface of tumor cells [62, 63]. It is well known that CTL epitopes binding to MHC, rather than integral TAA, induce CTL reactions [64]. These epitope peptides usually are comprised of eight to ten amino acids, with two to three primary anchor residues that interact with the MHC-I molecules and two to three amino acid residues that bind to the T cell receptor (TCR) [64,65,66]. Therefore, identification of suitable CTL epitopes from TAA is extremely important for targeted immunotherapy.
Heparanase, as a suitable universal TAA, was firstly predicted Three epitopes derived from the human heparanase amino acid sequence were first predicted by Sommerfeldt et al. [67]. Their results showed that DCs loaded with the three predicted peptides of human heparanase (hHpa) could promote heparanase-specific CTLs to lyse cancer cells [67]. Our research group used super motif and quantitative motif methods to predict another three HLA-A2-restricted heparanase epitopes including hHpa277 (277–285, KMLKSFLKA), hHpa405 (405–413, WLSLLFKKL), and hHpa525 (525–533, PAFSYSFFV), which were found to elicit HLA-A2-restricted CTL responses specific for KATO-III gastric cancer cells, SW480 colorectal cancer cells and U2OS osteogenic sarcoma cells [68]. To investigate the in vivo immune response elicited by heparanase CTL epitopes, we further predicted candidate CTL epitopes derived from the mouse heparanase protein (mHpa) [69]. In vitro experiments showed that the predicted peptides could activate heparanase-specific CTLs to lyse three kinds of carcinoma cells expressing both heparanase and H-2Kb. In vivo experiments further indicated that the predicted peptides could immunize against tumors and successfully treat tumor-bearing hosts [69]. We further evaluated the in vivo immune response elicited by the above human heparanase CTL epitopes including hHpa277, hHpa405 and hHpa525 using HLA-A2 transgenic C57BL/6 mice and showed that these peptides could be presented naturally in vivo and also elicited heparanase-specific lysis of various gastric cancer cells [70]. These results suggest that the predicted heparanase peptides are novel CTL epitopes capable of inducing heparanase-specific CTLs in vitro and in vivo, serving as valuable targets for immunotherapy.
3.3 Multiple Antigen Peptide (MAP)-Based Immunotherapy
Synthetic epitope peptides are inadequate to clinical use due to their small molecular weight, single structure, weak immunogenicity, and rapid degradation, which cannot elicit an ideal immune response in the body [71, 72]. Nowadays, the multiple antigen peptides (MAP) can increase the molecular weight of the peptide, elevate immunogenicity and boost its activity against tumors [73, 74]. Yang et al. [75] firstly designed 3 MAP vaccines of human heparanase based on B-cell epitopes and found that these MAP vaccines could inhibit the invasiveness of tumor cells in vitro. We then designed three 4-branched MAPs based on the human leukocyte antigen (HLA)-A2-restricted CTL epitopes of human heparanase. The results showed that the MAP vaccines could induce heparanase-specific CTL and much stronger lysis of gastric cancer cells compared with their corresponding linear peptides without killing effect on heparanase-expressing autologous lymphocytes and dendritic cells [72]. Zhang et al. [76] designed 8-branched MAPs comprising FLNPDVLDI and found that it could induce specific CTLs for human heparanase in vitro, which effectively secreted IFN-γ and potently lysed human tumor cells. These findings indicate that MAP vaccines based on CTL epitopes of human heparanase might be valuable for cancer immunotherapy.
3.4 Heparanase in CAR T-Cell Therapy
The generation of chimeric antigen receptor (CAR) T-cell therapy has revolutionized T cell-based immunotherapy for the treatment of some cancers [77,78,79]. CAR T-cells (which are genetically engineered T cells expressing CARs on their surface) therapy is a form of adoptive cell therapy which has recently gained attention due to success in clinical trials and FDA approval [80,81,82]. CAR T-cell therapy is mainly used in treating hematological malignancies particularly in infants, achieving up to 90% clinical response rates in acute lymphoblastic leukemia [83], which leads to numerous clinical trials of CAR T-cell therapy against multiple hematological antigens such as CD19, CD20 and CD22 [84, 85]. However, the clinical efficacy of CAR T-cell therapy in solid tumors has been greatly limited with side effects and lack of therapeutic response [86,87,88]. Multiple factors are responsible for limiting the efficacy of CAR T-cell therapy in solid tumors such as gastric cancer, colorectal cancer and breast cancer. Among these factors, the extracellular matrix (ECM) around solid tumors may hinder T-cell penetration [89]. Heparanase is the only known mammalian β-D-endoglycosidase capable of cleaving the heparan sulfate chains of HSPGs, thereby degrading ECM. Based on this, Ignazio et al. [90] firstly engineered CAR-T cells to express heparanase and showed improved capacity to degrade ECM, which promoted tumor T-cell infiltration and antitumor activity. The results suggest that modification of CAR T-cells to express high levels of heparanase may be of benefit for the application of CAR-T cells in individuals with stroma-rich solid tumors.
4 Conclusions and Perspectives
Since the cloning of the heparanase gene, its effect was mainly associated with tumor metastasis and angiogenesis, both major aspects of tumor progression [91,92,93]. In addition to its enzymatic HS-degrading activity, heparanase also acts via non-enzymatic mechanisms that regulate various signal transduction, exosome formation [94,95,96], autophagy [97, 98], inflammation [99, 24, 100] and chemoresistance [101]. Nevertheless, the role and mode of heparanase action in gastric cancer remain to be better elucidated. Increasing studies focusing on the expression and role of heparanase in gastric cancer demonstrated that heparanase is overexpressed in gastric cancer and closely associated with cancer metastasis and poor prognosis. It is therefore essential to elucidate the mechanisms regulating heparanase expression in gastric cancer, including the involvement of TERT, HGF, and miRNAs. Our understanding of how heparanase is upregulated in gastric cancer is still incomplete. Another challenge in the field is the development of clinically effective heparanase-targeted therapy to treat cancer. Nowadays, immunotherapy has emerged as a novel strategy for cancer therapy because of its weak side effects, specificity and targeting characteristics. Heparanase has been recognized as a suitable universal TAA because of its crucial role in progression of multiple tumors. Several heparanase-based CTL epitopes have been shown to elicit specific antitumor immunity in vitro and in vivo against various tumors, which could be further enhanced by creating MAP vaccines. Moreover, generation of CAR-T cells with high expression levels of heparanase was found to promote tumor T-cell infiltration and antitumor activity. Further studies are needed to unravel the mechanisms of heparanase action in gastric cancer and optimize heparanase-targeted immunotherapy of gastric cancer.
References
Levy-Adam, F., Miao, H. Q., Heinrikson, R. L., Vlodavsky, I., & Ilan, N. (2003). Heterodimer formation is essential for heparanase enzymatic activity. Biochemical and Biophysical Research Communications, 308(4), 885–891.
McKenzie E, Young K, Hircock M, Bennett J, Bhaman M, Felix R, Turner P, Stamps A, McMillan D, Saville G, Ng S, Mason S, Snell D, Schofield D, Gong H, Townsend R, Gallagher J, Page M, Parekh R, Stubberfield C. Biochemical characterization of the active heterodimer form of human heparanase (Hpa1) protein expressed in insect cells. The Biochemical Journal 2003; 373(Pt 2):423–435.
Sanderson, R. D., Elkin, M., Rapraeger, A. C., Ilan, N., & Vlodavsky, I. (2017). Heparanase regulation of cancer, autophagy and inflammation: New mechanisms and targets for therapy. The FEBS Journal, 284(1), 42–55.
Bernfield, M., Götte, M., Park, P. W., Reizes, O., Fitzgerald, M. L., Lincecum, J., & Zako, M. (1999). Functions of cell surface heparan sulfate proteoglycans. Annual Review of Biochemistry, 68, 729–777.
Ilan, N., Elkin, M., & Vlodavsky, I. (2006). Regulation, function and clinical significance of heparanase in cancer metastasis and angiogenesis. The International Journal of Biochemistry & Cell Biology, 38(12), 2018–2039.
Iozzo, R. V., & Sanderson, R. D. (2011). Proteoglycans in cancer biology, tumour microenvironment and angiogenesis. Journal of Cellular and Molecular Medicine, 15(5), 1013–1031.
Sanderson, R. D. (2001). Heparan sulfate proteoglycans in invasion and metastasis. Seminars in Cell & Developmental Biology, 12(2), 89–98.
Tímár, J., Lapis, K., Dudás, J., Sebestyén, A., Kopper, L., & Kovalszky, I. (2002). Proteoglycans and tumor progression: Janus-faced molecules with contradictory functions in cancer. Seminars in Cancer Biology, 12(3), 173–186.
Meirovitz, A., Goldberg, R., Binder, A., Rubinstein, A. M., Hermano, E., & Elkin, M. (2013). Heparanase in inflammation and inflammation-associated cancer. The FEBS Journal, 280(10), 2307–2319.
Ateeq, B., Unterberger, A., Szyf, M., & Rabbani, S. A. (2008). Pharmacological inhibition of DNA methylation induces proinvasive and prometastatic genes in vitro and in vivo. Neoplasia, 10(3), 266–278.
Shteper, P. J., Zcharia, E., Ashhab, Y., Peretz, T., Vlodavsky, I., & Ben-Yehuda, D. (2003). Role of promoter methylation in regulation of the mammalian heparanase gene. Oncogene, 22(49), 7737–7749.
Ogishima, T., Shiina, H., Breault, J. E., Terashima, M., Honda, S., Enokida, H., Urakami, S., Tokizane, T., Kawakami, T., Ribeiro-Filho, L. A., Fujime, M., Kane, C. J., Carroll, P. R., Igawa, M., & Dahiya, R. (2005). Promoter CpG hypomethylation and transcription factor EGR1 hyperactivate heparanase expression in bladder cancer. Oncogene, 24(45), 6765–6772.
Friedmann, Y., Vlodavsky, I., Aingorn, H., Aviv, A., Peretz, T., Pecker, I., & Pappo, O. (2000). Expression of heparanase in normal, dysplastic, and neoplastic human colonic mucosa and stroma. Evidence for its role in colonic tumorigenesis. The American Journal of Pathology, 157(4), 1167–1175.
Gohji, K., Okamoto, M., Kitazawa, S., Toyoshima, M., Dong, J., Katsuoka, Y., & Nakajima, M. (2001). Heparanase protein and gene expression in bladder cancer. The Journal of Urology, 166(4), 1286–1290.
Koliopanos, A., Friess, H., Kleeff, J., Shi, X., Liao, Q., Pecker, I., Vlodavsky, I., Zimmermann, A., & Büchler, M. W. (2001). Heparanase expression in primary and metastatic pancreatic cancer. Cancer Research, 61(12), 4655–4659.
Hong, X., Nelson, K. K., deCarvalho, A. C., & Kalkanis, S. N. (2010). Heparanase expression of glioma in human and animal models. Journal of Neurosurgery, 113(2), 261–269.
El-Assal, O. N., Yamanoi, A., Ono, T., Kohno, H., & Nagasue, N. (2001). The clinicopathological significance of heparanase and basic fibroblast growth factor expressions in hepatocellular carcinoma. Clinical Cancer Research, 7(5), 1299–1305.
Masola, V., Maran, C., Tassone, E., Zin, A., Rosolen, A., & Onisto, M. (2009). Heparanase activity in alveolar and embryonal rhabdomyosarcoma: Implications for tumor invasion. BMC Cancer, 9, 304.
Cohen, I., Pappo, O., Elkin, M., San, T., Bar-Shavit, R., Hazan, R., Peretz, T., Vlodavsky, I., & Abramovitch, R. (2006). Heparanase promotes growth, angiogenesis and survival of primary breast tumors. International Journal of Cancer, 118(7), 1609–1617.
Kato, M., Wang, H., Kainulainen, V., Fitzgerald, M. L., Ledbetter, S., Ornitz, D. M., & Bernfield, M. (1998). Physiological degradation converts the soluble syndecan-1 ectodomain from an inhibitor to a potent activator of FGF-2. Nature Medicine, 4(6), 691–697.
Ilan, N., Elkin, M., & Vlodavsky, I. (2006). Regulation, function and clinical significance of heparanase in cancer metastasis and angiogenesis. The International Journal of Biochemistry & Cell Biology, 38(12), 2018–2039.
Parish, C. R., Freeman, C., & Hulett, M. D. (2001). Heparanase: A key enzyme involved in cell invasion. Biochimica et Biophysica Acta, 1471(3), M99–M108.
Kelly, T., Miao, H. Q., Yang, Y., Navarro, E., Kussie, P., Huang, Y., MacLeod, V., Casciano, J., Joseph, L., Zhan, F., Zangari, M., Barlogie, B., Shaughnessy, J., & Sanderson, R. D. (2003). High heparanase activity in multiple myeloma is associated with elevated microvessel density. Cancer Research, 63(24), 8749–8756.
Vlodavsky, I., Beckhove, P., Lerner, I., Pisano, C., Meirovitz, A., Ilan, N., & Elkin, M. (2012). Significance of heparanase in cancer and inflammation. Cancer Microenvironment, 5(2), 115–132.
Mahtouk, K., Hose, D., Raynaud, P., Hundemer, M., Jourdan, M., Jourdan, E., Pantesco, V., Baudard, M., De Vos, J., Larroque, M., Moehler, T., Rossi, J. F., Rème, T., Goldschmidt, H., & Klein, B. (2007). Heparanase influences expression and shedding of syndecan-1, and its expression by the bone marrow environment is a bad prognostic factor in multiple myeloma. Blood, 109(11), 4914–4923.
Edovitsky, E., Elkin, M., Zcharia, E., Peretz, T., & Vlodavsky, I. (2004). Heparanase gene silencing, tumor invasiveness, angiogenesis, and metastasis. Journal of the National Cancer Institute, 96(16), 1219–1230.
Cassinelli, G., Lanzi, C., Tortoreto, M., Cominetti, D., Petrangolini, G., Favini, E., Zaffaroni, N., Pisano, C., Penco, S., Vlodavsky, I., & Zunino, F. (2013). Antitumor efficacy of the heparanase inhibitor SST0001 alone and in combination with antiangiogenic agents in the treatment of human pediatric sarcoma models. Biochemical Pharmacology, 85(10), 1424–1432.
Dredge, K., Hammond, E., Handley, P., Gonda, T. J., Smith, M. T., Vincent, C., Brandt, R., Ferro, V., & Bytheway, I. (2011). PG545, a dual heparanase and angiogenesis inhibitor, induces potent anti-tumour and anti-metastatic efficacy in preclinical models. British Journal of Cancer, 104(4), 635–642.
Ritchie, J. P., Ramani, V. C., Ren, Y., Naggi, A., Torri, G., Casu, B., Penco, S., Pisano, C., Carminati, P., Tortoreto, M., Zunino, F., Vlodavsky, I., Sanderson, R. D., & Yang, Y. (2011). SST0001, a chemically modified heparin, inhibits myeloma growth and angiogenesis via disruption of the heparanase/syndecan-1 axis. Clinical Cancer Research, 17(6), 1382–1393.
Meirovitz, A., Hermano, E., Lerner, I., Zcharia, E., Pisano, C., Peretz, T., & Elkin, M. (2011). Role of heparanase in radiation-enhanced invasiveness of pancreatic carcinoma. Cancer Research, 71(7), 2772–2780.
Xiong, Z., Lü, M. H., Fan, Y. H., Cao, Y. L., Hu, C. J., Wu, Y. Y., Wang, S. M., Luo, G., Fang, D. C., Li, C., & Yang, S. M. (2012). Downregulation of heparanase by RNA interference inhibits invasion and tumorigenesis of hepatocellular cancer cells in vitro and in vivo. International Journal of Oncology, 40(5), 1601–1609.
Masola, V., Secchi, M. F., Gambaro, G., & M, O. (2014). Heparanase as a target in cancer therapy. Current Cancer Drug Targets, 14(3), 286–293.
Pisano, C., Vlodavsky, I., Ilan, N., & Zunino, F. (2014). The potential of heparanase as a therapeutic target in cancer. Biochemical Pharmacology, 89(1), 12–19.
Chen, J. Q., Zhan, W. H., He, Y. L., Peng, J. S., Wang, J. P., Cai, S. R., & Ma, J. P. (2004). Expression of heparanase gene, CD44v6, MMP-7 and nm23 protein and their relationship with the invasion and metastasis of gastric carcinomas. World Journal of Gastroenterology, 10(6), 776–782.
Tang, W., Nakamura, Y., Tsujimoto, M., Sato, M., Wang, X., Kurozumi, K., Nakahara, M., Nakao, K., Nakamura, M., Mori, I., & Kakudo, K. (2002). Heparanase: A key enzyme in invasion and metastasis of gastric carcinoma. Modern Pathology, 15(6), 593–598.
Endo, K., Maejara, U., Baba, H., Tokunaga, E., Koga, T., Ikeda, Y., Toh, Y., Kohnoe, S., Okamura, T., Nakajima, M., & Sugimachi, K. (2001). Heparanase gene expression and metastatic potential in human gastric cancer. Anticancer Research, 21(5), 3365–3369.
Tang, B., Xie, R., Qin, Y., Xiao, Y. F., Yong, X., Zheng, L., Dong, H., & Yang, S. M. (2016). Human telomerase reverse transcriptase (hTERT) promotes gastric cancer invasion through cooperating with c-Myc to upregulate heparanase expression. Oncotarget, 7(10), 11364–11379.
Shah, M. A., & Ajani, J. A. (2010). Gastric cancer—an enigmatic and heterogeneous disease. Journal of the American Medical Association, 303(17), 1753–1754.
Eccles, S. A. (1999). Heparanase: Breaking down barriers in tumors. Nature Medicine, 5(7), 735–736.
Wang, Z., Zhang, X., Xu, H., Zhou, X., Jiang, L., & Lu, C. (2005). Detection of peritoneal micrometastasis by reverse transcriptase-polymerase chain reaction for heparanase mRNA and cytology in peritoneal wash samples. Journal of Surgical Oncology, 90(2), 59–65.
Wang, Z., Xu, H., Jiang, L., Zhou, X., Lu, C., & Zhang, X. (2005). Positive association of heparanase expression with tumor invasion and lymphatic metastasis in gastric carcinoma. Modern Pathology, 18(2), 205–211.
Xie, Z. J., Liu, Y., Jia, L. M., & He, Y. C. (2008). Heparanase expression, degradation of basement membrane and low degree of infiltration by immunocytes correlate with invasion and progression of human gastric cancer. World Journal of Gastroenterology, 14(24), 3812–3818.
Hao, N. B., Tang, B., Wang, G. Z., Xie, R., Hu, C. J., Wang, S. M., Wu, Y. Y., Liu, E., Xie, X., & Yang, S. M. (2015). Hepatocyte growth factor (HGF) upregulates heparanase expression via the PI3K/Akt/NF-κB signaling pathway for gastric cancer metastasis. Cancer Letters, 361(1), 57–66.
Zheng, L., Jiang, G., Mei, H., Pu, J., Dong, J., Hou, X., & Tong, Q. (2010). Small RNA interference-mediated gene silencing of heparanase abolishes the invasion, metastasis and angiogenesis of gastric cancer cells. BMC Cancer, 10, 33.
Ma, X. M., Shen, Z. H., Liu, Z. Y., Wang, F., Hai, L., Gao, L. T., & Wang, H. S. (2014). Heparanase promotes human gastric cancer cells migration and invasion by increasing Src and p38 phosphorylation expression. International Journal of Clinical and Experimental Pathology, 7(9), 5609–5621.
Cao, H. J., Fang, Y., Zhang, X., Chen, W. J., Zhou, W. P., Wang, H., Wang, L. B., & Wu, J. M. (2005). Tumor metastasis and the reciprocal regulation of heparanase gene expression by nuclear factor kappa B in human gastric carcinoma tissue. World Journal of Gastroenterology, 11(6), 903–907.
Jiang, C., Chen, X., Alattar, M., Wei, J., & Liu, H. (2015). MicroRNAs in tumorigenesis, metastasis, diagnosis and prognosis of gastric cancer. Cancer Gene Therapy, 22(6), 291–301.
Lim, L. P., Lau, N. C., Garrett-Engele, P., Grimson, A., Schelter, J. M., Castle, J., Bartel, D. P., Linsley, P. S., & Johnson, J. M. (2005). Microarray analysis shows that some microRNAs downregulate large numbers of target mRNAs. Nature, 433(7027), 769–773.
Shi, J., Chen, P., Sun, J., Song, Y., Ma, B., Gao, P., Chen, X., & Wang, Z. (2017). MicroRNA-1258: An invasion and metastasis regulator that targets heparanase in gastric cancer. Oncology Letters, 13(5), 3739–3745.
Sheng, N., Zhang, L., & Yang, S. (2018). MicroRNA-429 decreases the invasion ability of gastric cancer cell line BGC-823 by downregulating the expression of heparanase. Experimental and Therapeutic Medicine, 15(2), 1927–1933.
Zheng, L., Jiao, W., Song, H., Qu, H., Li, D., Mei, H., Chen, Y., Yang, F., Li, H., Huang, K., & Tong, Q. (2016). miRNA-558 promotes gastric cancer progression through attenuating Smad4-mediated repression of heparanase expression. Cell Death & Disease, 7(9), e2382.
Zhang, Y. F., Tang, X. D., Gao, J. H., Fang, D. C., & Yang, S. M. (2011). Heparanase: A universal immunotherapeutic target in human cancers. Drug Discovery Today, 16(9–10), 412–417.
Liu, C. C., Yang, H., Zhang, R., Zhao, J. J., & Hao, D. J. (2017). Tumour-associated antigens and their anti-cancer applications. European Journal of Cancer Care, 26(5).
Martinez, M., & Moon, E. K. (2019). CAR T cells for solid Tumors: New strategies for finding, infiltrating, and surviving in the tumor microenvironment. Frontiers in Immunology, 10, 128.
van den Broek, M., von Boehmer, L., & Knuth, A. (2010). Developments in cancer immunotherapy. Digestive Diseases, 28(1), 51–56.
Zhou, G., Lu, Z., McCadden, J. D., Levitsky, H. I., & Marson, A. L. (2004). Reciprocal changes in tumor antigenicity and antigen-specific T cell function during tumor progression. The Journal of Experimental Medicine, 200(12), 1581–1592.
Vlodavsky, I., Elkin, M., Abboud-Jarrous, G., Levi-Adam, F., Fuks, L., Shafat, I., & Ilan, N. (2008). Heparanase: One molecule with multiple functions in cancer progression. Connective Tissue Research, 49(3), 207–210.
Arvatz, G., Weissmann, M., Ilan, N., & Vlodavsky, I. (2016). Heparanase and cancer progression: New directions, new promises. Human Vaccines & Immunotherapeutics, 12(9), 2253–2256.
Janikashvili, N., Larmonier, N., & Katsanis, E. (2010). Personalized dendritic cell-based tumor immunotherapy. Immunotherapy, 2(1), 57–68.
Banchereau, J., & Palucka, A. K. (2005). Dendritic cells as therapeutic vaccines against cancer. Nature Reviews. Immunology, 5(4), 296–306.
Cai, Y. G., Fang, D. C., Chen, L., Tang, X. D., Chen, T., Yu, S. T., Luo, Y. H., Xiong, Z., Wang, D. X., & Yang, S. M. (2007). Dendritic cells reconstituted with a human heparanase gene induce potent cytotoxic T-cell responses against gastric tumor cells in vitro. Tumour Biology, 28(4), 238–246.
Andersen, M. H., Schrama, D., Thor Straten, P., & Becker, J. C. (2006). Cytotoxic T cells. The Journal of Investigative Dermatology, 126(1), 32–41.
Lundegaard, C., Nielsen, M., & Lund, O. (2006). The validity of predicted T-cell epitopes. Trends in Biotechnology, 24(12), 537–538.
Rammensee, H. G., Falk, K., & Rötzschke, O. (1993). Peptides naturally presented by MHC class I molecules. Annual Review of Immunology, 11, 213–244.
Lundegaard, C., Nielsen, M., & Lund, O. (2006). The validity of predicted T-cell epitopes. Trends in Biotechnology, 24(12), 537–538.
Eisenbach, L., Bar-Haim, E., & El-Shami, K. (2000). Antitumor vaccination using peptide based vaccines. Immunology Letters, 74(1), 27–34.
Sommerfeldt, N., Beckhove, P., Ge, Y., Schütz, F., Choi, C., Bucur, M., Domschke, C., Sohn, C., Schneeweis, A., Rom, J., Pollmann, D., Leucht, D., Vlodavsky, I., & Schirrmacher, V. (2006). Heparanase: A new metastasis-associated antigen recognized in breast cancer patients by spontaneously induced memory T lymphocytes. Cancer Research, 66(15), 7716–7723.
Chen, T., Tang, X. D., Wan, Y., Chen, L., Yu, S. T., Xiong, Z., Fang, D. C., Liang, G. P., & Yang, S. M. (2008). HLA-A2-restricted cytotoxic T lymphocyte epitopes from human heparanase as novel targets for broad-spectrum tumor immunotherapy. Neoplasia, 10(9), 977–986.
Tang, X. D., Wan, Y., Chen, L., Chen, T., Yu, S. T., Xiong, Z., Fang, D. C., Liang, G. P., & Yang, S. M. (2008). H-2Kb-restricted CTL epitopes from mouse heparanase elicit an antitumor immune response in vivo. Cancer Research, 68(5), 1529–1537.
Tang, X. D., Liang, G. P., Li, C., Wan, Y., Chen, T., Chen, L., Yu, S. T., Xiong, Z., Fang, D. C., Wang, G. Z., & Yang, S. M. (2010). Cytotoxic T lymphocyte epitopes from human heparanase can elicit a potent anti-tumor immune response in mice. Cancer Immunology, Immunotherapy, 59(7), 1041–1047.
van der Burg, S. H., Bijker, M. S., Welters, M. J., Offringa, R., & Melief, C. J. (2006). Improved peptide vaccine strategies, creating synthetic artificial infections to maximize immune efficacy. Advanced Drug Delivery Reviews, 58(8), 916–930.
Wang, G. Z., Tang, X. D., Lü, M. H., Gao, J. H., Liang, G. P., Li, N., Li, C. Z., Wu, Y. Y., Chen, L., Cao, Y. L., Fang, D. C., & Yang, S. M. (2011). Multiple antigenic peptides of human heparanase elicit a much more potent immune response against tumors. Cancer Prevention Research (Philadelphia, Pa.), 4(8), 1285–1295.
Pini, A., Falciani, C., & Bracci, L. (2008). Branched peptides as therapeutics. Current Protein & Peptide Science, 9(5), 468–477.
Iriemenam, N. C., Khirelsied, A. H., Nasr, A., ElGhazali, G., Giha, H. A., Elhassan A-Elgadir, T. M., Agab-Aldour, A. A., Montgomery, S. M., Anders, R. F., Theisen, M., Troye-Blomberg, M., Elbashir, M. I., & Berzins, K. (2009). Antibody responses to a panel of Plasmodium falciparum malaria blood-stage antigens in relation to clinical disease outcome in Sudan. Vaccine, 27(1), 62–71.
Yang, J. M., Wang, H. J., Du, L., Han, X. M., Ye, Z. Y., Fang, Y., Tao, H. Q., Zhao, Z. S., & Zhou, Y. L. (2009). Screening and identification of novel B cell epitopes in human heparanase and their anti-invasion property for hepatocellular carcinoma. Cancer Immunology, Immunotherapy, 58(9), 1387–1396.
Zhang, J., Yang, J., Fan, D., Tao, H., Wang, H., & Yu, T. (2013). Peptide FLNPDVLDI of heparanase is a novel HLA-A2-restricted CTL epitope and elicits potent immunological antitumor effects in vitro with an 8-branched design. Oncology Reports, 29(5), 1955–1961.
Zhang, J., Yang, J., Fan, D., Tao, H., Wang, H., & Yu, T. (2013). Peptide FLNPDVLDI of heparanase is a novel HLA-A2-restricted CTL epitope and elicits potent immunological antitumor effects in vitro with an 8-branched design. Oncology Reports, 29(5), 1955–1961.
DeSelm CJ, Tano ZE2, Varghese AM3, Adusumilli PS1,2. CAR T-cell therapy for pancreatic cancer. Journal of Surgical Oncology 2017; 116(1):63–74.
Benmebarek, M. R., Karches, C. H., Cadilha, B. L., Lesch, S., Endres, S., & Kobold, S. (2019). Killing mechanisms of chimeric antigen receptor (CAR) T cells. International Journal of Molecular Sciences, 20(6).
Swann, J. B., & Smyth, M. J. (2007). Immune surveillance of tumors. The Journal of Clinical Investigation, 117(5), 1137–1146.
Neelapu, S. S., Locke, F. L., Bartlett, N. L., Lekakis, L. J., Miklos, D. B., Jacobson, C. A., Braunschweig, I., Oluwole, O. O., Siddiqi, T., Lin, Y., Timmerman, J. M., Stiff, P. J., Friedberg, J. W., Flinn, I. W., Goy, A., Hill, B. T., Smith, M. R., Deol, A., Farooq, U., McSweeney, P., Munoz, J., Avivi, I., Castro, J. E., Westin, J. R., Chavez, J. C., Ghobadi, A., Komanduri, K. V., Levy, R., Jacobsen, E. D., Witzig, T. E., Reagan, P., Bot, A., Rossi, J., Navale, L., Jiang, Y., Aycock, J., Elias, M., Chang, D., Wiezorek, J., & Go, W. Y. (2017). Axicabtagene Ciloleucel CAR T-cell therapy in refractory large B-cell lymphoma. The New England Journal of Medicine, 377(26), 2531–2544.
Zinzani, P. L., Ribrag, V., Moskowitz, C. H., Michot, J. M., Kuruvilla, J., Balakumaran, A., Zhang, Y., Chlosta, S., Shipp, M. A., & Armand, P. (2017). Safety and tolerability of pembrolizumab in patients with relapsed/refractory primary mediastinal large B-cell lymphoma. Blood, 130(3), 267–270.
Maude, S. L., Frey, N., Shaw, P. A., Aplenc, R., Barrett, D. M., Bunin, N. J., Chew, A., Gonzalez, V. E., Zheng, Z., Lacey, S. F., Mahnke, Y. D., Melenhorst, J. J., Rheingold, S. R., Shen, A., Teachey, D. T., Levine, B. L., June, C. H., Porter, D. L., & Grupp, S. A. (2014). Chimeric antigen receptor T cells for sustained remissions in leukemia. The New England Journal of Medicine, 371(16), 1507–1517.
Porter, D. L., Levine, B. L., Kalos, M., Bagg, A., & June, C. H. (2011). Chimeric antigen receptor-modified T cells in chronic lymphoid leukemia. The New England Journal of Medicine, 365(8), 725–733.
Holzinger, A., Barden, M., & Abken, H. (2016). The growing world of CAR T cell trials: A systematic review. Cancer Immunology, Immunotherapy, 65(12), 1433–1450.
Maus, M. V., Haas, A. R., Beatty, G. L., Albelda, S. M., Levine, B. L., Liu, X., Zhao, Y., Kalos, M., & June, C. H. (2013). T cells expressing chimeric antigen receptors can cause anaphylaxis in humans. Cancer Immunology Research, 1(1), 26–31.
Kershaw MH, Westwood JA, Parker LL, Wang G, Eshhar Z, Mavroukakis SA, White DE, Wunderlich JR, Canevari S, Rogers-Freezer L, Chen CC, Yang JC, Rosenberg SA, Hwu P. A phase I study on adoptive immunotherapy using gene-modified T cells for ovarian cancer. Clinical Cancer Research 2006; 12(20 Pt 1):6106–6115.
Morgan, R. A., Yang, J. C., Kitano, M., Dudley, M. E., Laurencot, C. M., & Rosenberg, S. A. (2010). Case report of a serious adverse event following the administration of T cells transduced with a chimeric antigen receptor recognizing ERBB2. Molecular Therapy, 18(4), 843–851.
Tumeh, P. C., Harview, C. L., Yearley, J. H., Shintaku, I. P., Taylor, E. J., Robert, L., Chmielowski, B., Spasic, M., Henry, G., Ciobanu, V., West, A. N., Carmona, M., Kivork, C., Seja, E., Cherry, G., Gutierrez, A. J., Grogan, T. R., Mateus, C., Tomasic, G., Glaspy, J. A., Emerson, R. O., Robins, H., Pierce, R. H., Elashoff, D. A., Robert, C., & Ribas, A. (2014). PD-1 blockade induces responses by inhibiting adaptive immune resistance. Nature, 515(7528), 568–571.
Caruana, I., Savoldo, B., Hoyos, V., Weber, G., Liu, H., Kim, E. S., Ittmann, M. M., Marchetti, D., & Dotti, G. (2015). Heparanase promotes tumor infiltration and antitumor activity of CAR-redirected T lymphocytes. Nature Medicine, 21(5), 524–529.
Barash, U., Cohen-Kaplan, V., Dowek, I., Sanderson, R. D., Ilan, N., & Vlodavsky, I. (2010). Proteoglycans in health and disease: New concepts for heparanase function in tumor progression and metastasis. The FEBS Journal, 277(19), 3890–3903.
Fux, L., Ilan, N., Sanderson, R. D., & Vlodavsky, I. (2009). Heparanase: Busy at the cell surface. Trends in Biochemical Sciences, 34(10), 511–519.
Levy-Adam, F., Ilan, N., & Vlodavsky, I. (2010). Tumorigenic and adhesive properties of heparanase. Seminars in Cancer Biology, 20(3), 153–160.
Thompson, C. A., Purushothaman, A., Ramani, V. C., Vlodavsky, I., & Sanderson, R. D. (2013). Heparanase regulates secretion, composition, and function of tumor cell-derived exosomes. The Journal of Biological Chemistry, 288(14), 10093–10099.
David, G., & P, Z. (2015). Heparanase tailors syndecan for exosome production. Molecular & Cellular Oncology, 3(3), e1047556.
David, G., & P, Z. (2015). Heparanase tailors syndecan for exosome production. Molecular & Cellular Oncology, 3(3), e1047556.
Shteingauz, A., Boyango, I., Naroditsky, I., Hammond, E., Gruber, M., Doweck, I., Ilan, N., & Vlodavsky, I. (2015). Heparanase enhances tumor growth and Chemoresistance by promoting autophagy. Cancer Research, 75(18), 3946–3957.
Ilan, N., Shteingauz, A., & Vlodavsky, I. (2015). Function from within: Autophagy induction by HPSE/heparanase--new possibilities for intervention. Autophagy, 11(12), 2387–2389.
Wang, L., Fuster, M., Sriramarao, P., & Esko, J. D. (2005). Endothelial heparan sulfate deficiency impairs L-selectin- and chemokine-mediated neutrophil trafficking during inflammatory responses. Nature Immunology, 6(9), 902–910.
Vlodavsky, I., Singh, P., Boyango, I., Gutter-Kapon, L., Elkin, M., Sanderson, R. D., & Ilan, N. (2016). Heparanase: From basic research to therapeutic applications in cancer and inflammation. Drug Resistance Updates, 29, 54–75.
Ramani, V. C., Vlodavsky, I., Ng, M., Zhang, Y., Barbieri, P., Noseda, A., & Sanderson, R. D. (2016). Chemotherapy induces expression and release of heparanase leading to changes associated with an aggressive tumor phenotype. Matrix Biology, 55, 22–34.
Author information
Authors and Affiliations
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2020 Springer Nature Switzerland AG
About this chapter

Cite this chapter
Tang, B., Yang, S. (2020). Involvement of Heparanase in Gastric Cancer Progression and Immunotherapy. In: Vlodavsky, I., Sanderson, R., Ilan, N. (eds) Heparanase. Advances in Experimental Medicine and Biology, vol 1221. Springer, Cham. https://doi.org/10.1007/978-3-030-34521-1_13
Download citation
DOI: https://doi.org/10.1007/978-3-030-34521-1_13
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-34520-4
Online ISBN: 978-3-030-34521-1
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)