
Abstract
Microbial mats growing under the permanent ice cover of Antarctic lakes occupy an exceptionally low-disturbance regime. Constant temperature, the absence of bioturbation or physical disturbance from wind action or ice formation allow mats to accumulate, as annual growth layers, over many decades or even centuries. In so doing they often assume decimetre scale, three-dimensional morphologies such as elaborate pinnacle structures and conical mounds. Here we combine existing and new information to describe microbial structures in three Antarctic lakes—simple prostrate mats in Lake Hoare, emergent cones in Lake Untersee and elaborate pinnacles in Lake Vanda. We attempt to determine whether structures emerge simply from uncoordinated organism-environment interactions or whether they represent an example of “emergent complexity”, within which some degree of self-organisation occurs to confer a holistic functional advantage to component organisms. While some holistic advantages were evident from the structures—the increase in surface area allows greater biomass and overall productivity and nutrient exchange with overlying water—the structures could also be understood in terms of potential interactions between individuals, their orientation and their environment. The data lack strong evidence of coordinated behaviour directed towards holistic advantages to the structure, though hints of coordinated behaviour are present as non-random distributions of structural elements. The great size of microbial structures in Antarctic lakes, and their relatively simple community composition, makes them excellent models for more focused research on microbial cooperation.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
4.1 Introduction
Photosynthetic microbial mats (biofilms) frequently show consistent physical patterns that hint at organisation. At one level, organisation is exemplified by metabolic zonation, which typically occurs on mm length scales (Franks and Stolz 2009). Such zonation results in organisms capable of oxygenic photosynthesis concentrated at the top of a mat, where light is most abundant, while deeper in the mats, lower irradiance and depleted oxygen enable the growth of organisms capable of a cascade of anaerobic metabolisms (Kühl and Fenchel 2000). Layering is often visible as colour changes, with an orange surface from abundant light-protective carotenoid pigments, transitioning to green light-harvesting phycobilin and chlorophyll pigments and pink layers corresponding to photosynthetic sulphur bacteria in anoxic zones. There are many examples of this metabolic “organisation” in the literature, and the close proximity of interacting metabolisms allows diffusion to be an effective coupling mechanism that provides key advantages to this mat structure (Seckbach and Oren 2010; Nadell et al. 2016). Viewed as an integrated unit, the microbial mat shows elegant metabolic coupling, facilitated by a one-dimensional, organised structure that allows a more efficient utilisation of resources than any one organism could do alone (Paerl and Pinckney 1996; Stahl 1995; Franks and Stolz 2009). Indeed, some argue that the organised complexity and physiological cooperativity of a microbial mat is analogous to that of eukaryotic tissues (Webb et al. 2003; Guerrero and Berlanga 2007; Stahl et al. 2018).
When microbial mat communities grow in low-disturbance environments, a higher level of organisation can occur, whereby elaborate three-dimensional structures emerge on scales much larger than those of either component organisms or metabolic zonation (Bosak et al. 2013). Such emergent structures are most frequently described from environments where conditions preclude metazoa large enough to physically disrupt developing microbial communities (Fenchel 1998). The most frequently described emergent microbial structures are from quiescent geothermal or hypersaline waters, where physical disruption is minimal and insects and other invertebrates are temperature- or salinity-excluded (Des Marais 1995; Oren 2010). There, a variety of macroscopic mat forms occur, overlain on metabolic zonation, which in quiescent locations often take the “ridge-pinnacle” form of regular-spaced cones of cm-scale size and spacing, connected by narrow ridges (Walter et al. 1976).
Microbial mats thus show emergent organisation on at least two levels: a three-dimensional arrangement of organisms within the mat that appears to produce an efficient metabolic unit and a three-dimensional organisation of the mat matrix to form spatially complex structures. In modern concepts of emergent organisation (Halley and Winkler 2008), the former would be termed a simple emergence, computable through a reductionist understanding of organism-environment interactions and close to an (dynamic) equilibrium (Corning 2002; de Wolf and Holvoet 2005). Whether the three-dimensional organisation is an example of complex emergence, within which some degree of self-organisation occurs to convey a holistic functional advantage, or whether it too is driven by simple organism-environment responses is not clear. At small scales, there is accumulating evidence for holistic, coordinated behaviour in biofilms (Stahl et al. 2018), perhaps controlled by cell to cell signalling and “programmed death” that would allow a mechanism for complex emergence of structure, though to date we are aware of no definitive examples from nature where coordinated behaviour for holistic benefit is shown (Battin et al. 2007). One of the few recent studies to suggest a holistic function for macroscale ridge-pinnacle structure suggests that a mechanism exists to space pinnacles to optimise overall access to water column resources by minimising inter-pinnacle competition (Petroff et al. 2010). This degree of self-organisation requires information transfer amongst pinnacles to achieve the overall optimum condition, which the authors propose via resource depletion “shells” around structures. Other authors argue for mechanisms of complex structure emergence that require much less self-organisation, suggesting that cones initiate as surface irregularities that are consequences of un-oriented movement (Shepard and Sumner 2010) that then propagate vertically either through phototaxis (Stahl 1995) or through differential growth of tips due to enhanced access to limiting resources (Tice et al. 2011). The question of whether 3-D microbial mat structures derive from internal or external drivers, or a mix of the two, extends beyond semantics; it relates to whether ridge-pinnacle structure is the inevitable outcome of individual behaviour in response to environmental drivers and has no holistic function or whether individual behaviour within microbial mats has, over time, been structured to facilitate enhanced overall function through a self-organised structure.
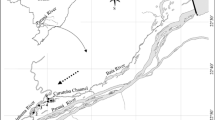
In this contribution, we examine the extent to which microbial mat structures are organised and consider whether this organisation is essentially mechanistic, with organisms responding to environmental stimuli or whether there is evidence that organised structures contribute to holistic functions that enhance overall performance. As case studies, we focus on similarities and differences between mats in three perennially ice-covered Antarctic lakes (Fig. 4.1). Antarctic lakes are particularly favourable locations for the development of complex microbial structures and contain some of the best developed modern examples (Hawes et al. 2011, 2013; Mackey et al. 2015; Sumner et al. 2016). They share with geothermal and hypersaline waters the lack of bioturbation from macro-invertebrates, due in this case to the combination of extreme environmental conditions and geographic isolation (see discussion in Karanovic et al. 2014), but furthermore, the ice cover ensures that lake water is exceptionally still, over long time periods, with no wind-induced turbulence (Wharton et al. 1993; Andersen et al. 2011; Hawes et al. 2013) that could generate sufficient shear stress to disturb mat communities (Tice et al. 2011). Consequently, large-scale structures, overlain on an internal mat zonation, reach decimetre proportions in these lakes (Fig. 4.2; Andersen et al. 2011; Hawes et al. 2011, 2013). Microbial mats with obvious internal one-dimensional structure are also common in shallow Antarctic habitats (Howard-Williams et al. 1989; Vincent 2000; Jungblut et al. 2012), but their prolonged winter freezing and summer water movement appear to prevent the development of large-scale three-dimensional organisation, and thus our focus remains on lake mats developing in perennially liquid, under-ice habitats.

The location in Antarctica of the three lakes primarily referred to in this chapter. Public domain image produced by the US National Aeronautics and Space Administration

Examples of contrasting microbial mats in perennially ice-covered Antarctic lakes. (a) Complex emergent structures at 22 m depth in Lake Vanda, Antarctica. (b) Microbial mat with little emergent complexity at 11 m depth in Lake Hoare. The checkers are 1 cm in both images, which are by courtesy of Tyler Mackey
We focus on three lakes that allow comparison of three distinctly different macroscale mat topographies: Lake Hoare where prostrate forms prevail (Hawes et al. 2014, 2016); Lake Untersee where a unique conical growth form is widespread (Andersen et al. 2011); and Lake Vanda, where dense fields of tall cuspate pinnacles dominate (Fig. 4.2; Hawes et al. 2013; Sumner et al. 2016). Although early literature refers to Antarctic lake mats rather generically as “modern stromatolites” (e.g. Parker et al. 1981), for the most part, this is misleading. Though laminated, they are mostly unlithified (Mackey et al. 2015), being entirely built from alternating annual laminations of sediment and microbially derived organic material (Hawes et al. 2001; Sutherland and Hawes 2009). Thus, the potential structuring effects of spatially constrained mineral precipitation (Petroff et al. 2013) are not in play, and Antarctic lakes instead present some of the best developed microbe-soft sediment accretion structures on modern Earth. Lake Vanda has the added advantage that it has undergone recent increases in lake level, with newly inundated areas of lake bed providing dated underwater landscapes allowing an examination of the evolution of structural complexity over time (Hawes et al. 2013; Sumner et al. 2016).
4.2 Lake Environments
4.2.1 Lake Hoare
Lake Hoare (77.63°S, 162.88°E) is a closed-basin lake near the eastern end of Taylor Valley in southern Victoria Land, Antarctica. The lake is 4.2 km long and 1.0 km wide and has maximum and mean depths of 34 and 14 m (Spigel and Priscu 1998). It is dammed to the north-east by the Canada Glacier, which provides an inflow of glacial meltwater (Wharton et al. 1992). Other sources of inflow come from Andersen Creek entering the north-east corner of Lake Hoare and drainage from Lake Chad in the south-east. No outflows from Lake Hoare exist, and water loss is restricted to sublimation of ice and evaporation of meltwater during summer (Doran et al. 1994). The ice cover of Lake Hoare is perennial, except for small areas at the lake margins, which melt during summer. The ice cover was 3.5 m thick in 1983 (Wharton et al. 1992), but the thickness has increased to ~5 m since that time (Doran et al. 2002). There are 3 months of complete darkness during winter and 3 months of continuous light during summer (Dana et al. 1998). Howard-Williams et al. (1998) summarised the optical properties of the ice cover and the water column. Net transmission of photosynthetically active radiation (PAR) in the lake centre ranged from <1% to 3%, with a spectral transmission peak at wavelengths between 450 and 550 nm. Vertical extinction coefficients for downwelling PAR within the water column from beneath the ice to a depth of 33 m were typically 0.12–0.22 m−1 (Howard-Williams et al. 1998), resulting in a 1% irradiance depth of between just below the ice cover and 9 m.
Lake Hoare shows weak density stratification in the upper waters, and although there is controversy over the extent of mixing of surface water (Spigel and Priscu 1998; Tyler et al. 1998), the density gradient is continuous to at least 13 m depth. There is a pronounced inflexion in the density-depth profile at 13–15 m from the surface (the depth of the lake varies temporally), which divides the lake into upper and lower compartments. The upper compartment is characterised by low concentrations of dissolved nutrients, particularly nitrate, which is probably limiting planktonic production (Lizotte and Priscu 1992). Upper waters also contain lower bicarbonate, with a higher pH (up to 8.6, Cathey et al. 1981) than the lower compartment (pH 7.9). The lake is anoxic below 25–26 m.
Benthic microbial mats, comprised primarily of cyanobacteria, diatoms and other bacteria, with various microbial eukaryotes at low abundance, line the lake from the edge into the anoxic zone at 25 m (Wharton et al. 1983; Hawes and Schwarz 1999). Mats in Lake Hoare are often prostrate, with little surface elaboration, though in some shallow areas rounded pinnacles and lift-off structures can form (Wharton et al. 1983). The matrices of the mats are dominated by narrow, filamentous cyanobacteria, attributable on morphological and molecular bases to the genera Leptolyngbya and Pseudanabaena with occasional Phormidium (Sutherland and Hawes 2009; Zhang et al. 2015). Diatoms comprise 10–15% of the mat community, though this increases at depth. Sixteen taxa of diatoms have been recognised, with Psammothidium chlidanos, Diadesmis contenta var. parallela and Navicula gregaria the most frequent (Sutherland and Hawes 2009). These mats are adapted to low irradiance through efficient light harvesting and utilisation, and compensation points of <1 μmol photons m−2 s−1 allow net production to occur to >20 m depth (Hawes and Schwarz 1999, 2001; Vopel and Hawes 2006).
4.2.2 Lake Untersee
Lake Untersee (Fig. 4.1) is located at 71.33°S 13.75° E in the Otto-von-Gruber-Gebirge (Gruber Mountains) of central Dronning Maud Land. The lake is 563 m above sea level, with an area of 11.4 km2, and is the largest surface lake in East Antarctica. Lake Untersee has two subbasins; the largest, 160 m deep, lies adjacent to the Anuchin Glacier and is separated by a sill at 50 m depth from a smaller, 100-m-deep basin in the southwest corner (Wand et al. 1997). The deep basin is close to homothermal and well oxygenated to the bottom. In contrast the shallow basin is density stratified below the sill depth (c. 50 m) and anoxic at its base. Profiles of conductivity, temperature and dissolved oxygen indicate that the deep basin and upper part of the anoxic basin are part of the same body of well-mixed water (Kaup et al. 1988; Haendel et al. 1995; Wand et al. 1997, 2006; Andersen et al. 2011). The unusually high pH of this mixed water mass (pH 10.4) has been attributed to weathering of the predominant anorthosite rock in the lake’s catchment in the absence of an effective connection to the atmosphere (Kaup et al. 1988; Andersen et al. 2011). The lake ice transmits approximately 5% of incident irradiance and vertical extinction coefficient for scalar PAR of 0.033 m−1, (Andersen et al. 2011) resulting in a 1% surface irradiance depth of ~50 m.
Benthic microbial mats occur to at least 100 m in Lake Untersee and comprise mostly oscillatorian cyanobacteria primarily as Phormidium autumnale and Leptolyngbya spp., with some unicellular Chamaesiphon and nitrogen-fixing Nostoc, with few chlorophytes and almost no diatoms (Andersen et al. 2011; authors’ unpublished molecular data). These microbial mats form two distinct macroscale morphologies, pinnacles and large cones, with the latter, which can be up to 50 cm tall, being most widespread. Cones are built from sub-mm scale laminations of fine glacially derived sediments and organic material, and radiocarbon dating suggests that they accumulate over centuries (Andersen et al. 2011). As in lakes Vanda and Hoare, these cyanobacteria are rich in phycoerythrin, and mats have an overall purple-pink appearance.
4.2.3 Lake Vanda
Lake Vanda (Fig. 4.1) lies in the Wright Valley, one of the McMurdo Dry Valleys of southern Victoria Land (77.50°S, 161.67°E). It occupies a closed basin with a perennial ice cover 3.5–4.0 m thick; though for several weeks each summer, the ice around parts of the lake shore melts to produce a discontinuous, open-water moat. In recent years inflow has exceeded outflow, and the lake level has risen by 12 m between 1960 and 2012 (Hawes et al. 2013). The endorheic nature of the lake and historical changes in water balance have left Lake Vanda with an unusual physical structure, with an inverse temperature gradient stabilised by increasing solute content with depth. In December 2013, temperature and conductivity were constant from 4 m to 23 m depth at ~4 °C and ~560 μS cm−1, increasing abruptly as a pycnocline from 23 to 28 m depth, then stabilising at 6 °C and 1020 μS cm−1 between 28 and 45 m. These two upper cells are individually mixed by thermohaline convection (Spigel and Priscu 1998), while below 45 m, there is a continuous gradual increase in temperature and conductivity. The ice cover transmits 15–20% of incident photosynthetically active radiation (PAR), biased to wavelengths below 550 nm (Hawes and Schwarz 2001). The lake water is extraordinarily clear, with a vertical extinction coefficient for downwelling scalar PAR of 0.06 m−1 (Howard-Williams et al. 1998), resulting in a 1% surface irradiance depth of 48–63 m. Low concentrations of dissolved reactive phosphorus (DRP) appear to limit phytoplankton growth (Vincent and Vincent 1982).
In Lake Vanda, benthic microbial mats are widespread, attain high biomass, form elaborate pinnacles and extend to at least 50 m depth (Figs. 4.2 and 4.3; Wharton 1994; Hawes and Schwarz 2001; Hawes et al. 2013). These mats comprise mostly cyanobacteria, particularly species of Phormidium and Leptolyngbya (Sumner et al. 2016), with pennate diatoms (species of Navicula, Nitzschia, Caloneis and Stauroneis) throughout and occasional strands of moss below the 25 m pycnocline (Love et al. 1983). Lake Vanda mats appear to have a higher cyanobacterial diversity than Lake Hoare, with up to 12 compared to 8 operational taxonomic units (OTUs) based on clone library analysis (Zhang et al. 2015). Vanda mats are annually laminated, with the upper 2–4 laminae constituted an orange-brown zone, rich in myxoxanthophyll and dominated by intertwined Leptolyngbya trichomes. Below the upper zone, green-/pink-pigmented subsurface zones are present, containing up to six phycobilin-rich laminae, also colonised by Leptolyngbya, Oscillatoria and Phormidium morphotypes (Hawes et al. 2013, 2016; Sumner et al. 2016). Recent, well-constrained, increases in lake level have allowed mats from different depths to be examined as a proxy for rate of development (Hawes et al. 2013). At depths inundated for <30 years, mats were accreting, adding one lamina (0.3 mm) and ~0.2 μg cm−2 of chlorophyll-a per year. At depths that have been submerged for >40 years, mat chlorophyll-a appeared to have reached an equilibrium and did not increase over time. The older mats formed tall pinnacles that were only beginning to emerge in shallower mats.

A vertical section of microbial mat from 10 m depth in Lake Hoare, 2010. (a) A photograph showing mm-scale, annual laminations and the zonation from pink-brown at the surface, though pink, to colourless. (b) A false-colour composite fluorescence image (blue excitation, red emission) of a similar section. False colour scale is in arbitrary units of chlorophyll fluorescence. The scale bars in (a) and (b) represent 10 mm. (c) An in situ dissolved oxygen profile through a mat at the same depth from which the sections were obtained, shown on a similar vertical scale
4.3 One-Dimensional Structures
Lake Hoare microbial mats frequently lack well-developed, complex three-dimensional structures (Fig. 4.2), though one-dimensional organisation is well developed. The properties of 1-D structure in these mats take two forms; lamination and zonation (Hawes et al. 2014). Lamination, composed of alternating sediment-rich and sediment-poor bands, is a product of annual accumulation (Hawes et al. 2001). Sediments enter the lake in summer via streams that typically flow for 1–2 months, and settle as a distinct optically dense band over autumn-winter when darkness prevents photosynthetic growth. The resumption of growth the following spring, when light returns but inflows are not running, forms a hyaline band (organic accumulation without sediment) overlying the relatively opaque (sediment without growth) band formed in winter (Fig. 4.3a). Hawes et al. (2001) used confocal laser scanning microscopy to show that trichomes are relatively sparse and oriented mostly vertically in the hyaline bands, including at the surface in summer, but tend to be more abundant, intertwined and oriented parallel to the mat surface in the opaque bands. Such laminations through trapping and binding of episodic depositions of sediment are reported from microbial mat communities in other sedimentary environments such as marine stromatolites (e.g. Reid et al. 2000).
Zonation is overlain on this annual banding pattern and in Antarctic lakes is recognisable by pigmentation bands that comprise multiple laminae. The orange-brown surface zone contains a higher proportion of carotenoids and live diatoms than the lower zone, where carotenoids are rare and cyanobacterial phycobilin pigments are responsible for pink coloration (Hawes et al. 2016). However, cyanobacteria dominate biovolume throughout the pigmented zone, with a similar morphotypic composition, dominated by Leptolyngbya in both zones, with Pseudanabaena present in both but most abundant in the upper pink-brown zone (Sutherland and Hawes 2009; Hawes et al. 2016).
In addition, Lake Hoare mats have two unusual features that are not common in temperate microbial mats: (1) the >18 mm thickness of the pigmented zone, which occupies 9 years of vertical accumulation (Fig. 4.3a, b) and (2) the persistence of dissolved oxygen concentrations exceeding that of the ambient water column to more than 15 mm into the mat (Fig. 4.3c). These are despite the mat receiving <1% of lake-incident irradiance (Vopel and Hawes 2006; Hawes et al. 2014). Fluorescence imaging (Fig. 4.3b, see Vopel and Hawes 2006 for methods) shows that chlorophyll-a is present throughout the pigmented zone of the mat, only reaching the limits of detection at 15 mm or more below the mat surface. The bulge in oxygen concentration associated with the pigmented zones is indicative of a broad photosynthetic zone, but also that oxic conditions extend well below the region containing pigments associated with oxygenic photosynthesis. This is different to what is normally seen in temperate mats, and even in shallow water (seasonally frozen) mats in Antarctica (Hawes et al. 1999), where oxygenic photosynthesising organisms are usually concentrated in the upper 1–2 mm, and a sharp oxycline is present at the base of the oxygenic, pigment-containing zone. In temperate system, the oxycline within the mat can be associated with an increase in sulphide and a shift in dominant metabolism from oxygen- to sulphur-based and moves vertically in response to diel changes in irradiance (e.g. Denis et al. 2012). Indeed, in temperate mats, this redox change is the basis of the 1-D metabolic structure that is often interpreted as having the functional role of facilitating nutrient cycling by positioning all components of nutrient cycling pathways within diffusion distance of each other (e.g. Paerl and Pinckney 1996).
The unusual oxygenation features of Lake Hoare mats may in part reflect the 24-h daily irradiance that they receive, albeit at low photon fluxes. Estimates of photosynthesis in Lake Hoare show that photic zones of microbial mats export oxygen to the water column and to underlying mats during summer and that oxygen consumption in mats below the photic zone quickly falls to close to zero despite an abundance of organic carbon (Hawes et al. 2014). Vopel and Hawes (2006) demonstrated that the rate of photosynthesis is linearly related to irradiance and argued for a compensation photon flux as low as 0.1 μmol m−2 s−1. Hawes et al. (2014) went on to confirm persistent light-limitation of photosynthesis but also showed that net flux of oxygen to the water column continued through the 24-h light cycle. However, the export of dissolved oxygen at low irradiances around midnight was due to ongoing effusion of dissolved oxygen produced during brighter parts of the day: under the night-time photon flux of ~1 μmol photon m−2 s−1. Gross photosynthesis was close to zero, and a net consumption of oxygen was actually occurring when flux was integrated through the mat, implying that ~1 μmol photon m−2 s−1 may be closer to the effective community compensation point than 0.1 μmol photon m−2 s−1.
The ability of the Lake Hoare microbial mat to be photosynthetic at low irradiance is enhanced by efficient absorption and utilisation of light. Hawes and Schwarz (2001) showed how the spectral absorption characteristics of ice (which absorbs red light strongly) results in light that penetrates to 10 m depth in Lake Hoare having a dominant wavelength of close to 500 nm and an overall blue-green hue. They showed how phycoerythrin allowed benthic mats to capture effectively these green wavebands and absorb up to 50% of light incident to the mat using these photosynthetic pigments. A variety of methodologies have shown how the cyanobacteria function at close to the maximum quantum yield of oxygenic photosynthesis, fixing the equivalent of 1 mol carbon per 12 mol photons absorbed (Hawes and Schwarz 1999, 2001; Vopel and Hawes 2006; Hawes et al. 2014). Low respiration rate is, however, also essential to both low-compensation irradiance and deep-penetrating oxygen (Hawes and Schwarz 1999; Hawes et al. 2014). The low respiration rate, despite an apparent abundance of oxygen and organic carbon in the mat matrix (Sutherland and Hawes 2009), is unexpected. It has been suggested that low respiration rate is due to the recalcitrant nature of the cyanobacterial sheaths and extracellular polymeric material that is suspected to form much of the organic laminae (Sutherland and Hawes 2009). What happens to dissolved oxygen concentration in mat communities over the 4 months of winter darkness is not yet known.
While we can hypothesise that the unusual oxygen profiles in Lake Hoare mats reflect the absence of release of labile organics from the photosynthetic mat components to the underlying mat, it is clear that the 1-D structure of these Antarctic lake mats is not based around redox gradients that facilitate coupling of microbial processes. The absence of an oxycline within the mats implies the absence of processes that link aerobic and anaerobic metabolisms to the carbon acquisition processes at diffusion-efficient distance scales—at least during the summer months. Nutrient accumulation does occur in these mats, with concentrations of all inorganic- and organic-dissolved forms of N and P increasing severalfold from the water column to the mat (Quesada et al. 2008), but the absence of abrupt redox gradients means that benefits from closely linked, diverse metabolisms are unlikely, and the enigma of apparently abundant carbon resources in oxygen-rich, aphotic lower zones of microbial mats remains incompletely resolved.
So does the one-dimensional arrangement of cyanobacteria in the mat have another function? It seems unlikely that the structure of the mat, with pigments arranged within a sequence of laminae representing up to 10 years of growth (Fig. 4.3), has the function of enhancing the ability to harvesting light. The most effective arrangement of pigments to intercept light would be to concentrate them in a narrow layer at the mat surface, as this would minimise attenuation due to non-photosynthetic mat components (Kühl and Fenchel 2000). Thus, one might expect these potentially motile organisms to migrate vertically to the upper, better illuminated part of the mat. At present the reason for the absence of migration is not clear, but we have previously argued (Hawes et al. 2014) that the only available cue for migration is light and that the rapid scattering of light within mats (Kühl and Fenchel 2000) and the low component of the red wavelengths that appear to be important in phototaxis (Ng et al. 2003) prevent coordinated vertical migration from occurring. The alternate hypothesis then is that pigments in deeper, older laminae are persistent legacies of previous growth and that the 1-D structure seen in these mats—annual lamination and some zonation of pigments and dominant morphotypes—has no holistic function; rather it is simply a consequence of how these mats grow and age.
4.4 Three-Dimensional Structure
4.4.1 Lake Untersee
Above we argue that the 1-D structure that is evident in vertical sections of microbial mat from Lake Hoare, at 10 m depth, is largely a consequence of the way that the mat grows and confers no advantage in the acquisition of the key-limiting resource at that location. In the second example considered here, Lake Untersee, Dronning Maud Land, the mats have a 1-D laminated structure that superficially resembles that of Lake Hoare, but this 1-D structure is folded into the emergence of conical macroscale structures (Andersen et al. 2011). Here we will consider how these structures may form and how this formation may create a holistic advantage for the capture of otherwise limiting resources.
Lake Untersee actually contains two types of emergent structure, pinnacles and cones (Andersen et al. 2011; Fig. 4.4). Surveys using drop cameras and divers have shown that the cones are the most frequent structures seen in the lake and that these are present from less than 15 m depth (the minimum depth surveyed) to in excess of 100 m, equivalent to optical depths of ~3–0.25% of surface incident irradiance (author’s unpublished data). This encompasses the optical depth of the mats discussed above from Lake Hoare (~1% surface light). Cones themselves are all sizes, but often large, up to 50 cm tall, and rise steeply (average slope 60°) from the surrounding sediment with a spacing length of metres (Andersen et al. 2011). A characteristic feature is a fibrous cluster of a broad-trichome cyanobacterium (identified by sequence analysis of 16S rRNA gene as Phormidium autumnale; author’s unpublished data) on the apex of the cones, particularly evident when viewed from above (Fig. 4.4b). In fact, 16S high-throughput sequencing showed that Phormidium dominates the cyanobacterial assemblages on the sides and top of the cone from where the fibrous cluster of a broad-trichome cyanobacterium protrudes (authors’ unpublished data). This is quantifiable as an elevated concentration of photosynthetic pigments at the cone apex (Table 4.1).

(a) A field of conical mounds at 20 m depth in Lake Untersee. For comparison with other lakes, 20 m physical depth in Lake Untersee is the depth to which 2–3% of incident irradiance penetrates. (b) A close-up of a small conical mound, with a nascent structure in the upper right. The scale bar shows cm. (c) Cross section of the cone imaged in (b) after return to the surface, section close to “shoulder” of the cone. All images are taken with no artificial lighting
Vertical sections of cones reveal that the cone structure is built as a modification of the simple laminar accrual process described in Lake Hoare, but the internal structure is quantitatively different to that seen in Lake Hoare (Fig. 4.4c). In particular, zonation is restricted to a single pink-pigmented, upper zone less than 1 mm thick, overlying an unpigmented organosedimentary zone, and the scale of lamination in Lake Untersee is much finer, sub-mm scale laminated clay-organic sediment. Although arranged differently, the absolute abundance of pigments per unit area was similar to lakes Vanda and Hoare (Table 4.1). Potential benefits of the conical geometry include an increase in mat surface area for a given area of lake bed. Given that the concentration of pigments on the cone walls are similar to those on the adjacent flat areas (Table 4.1), the increase in surface area of cones relative to flats will inevitably increase the overall biomass per unit lake bed. A regular-truncated cone that is 20 cm tall, with a 60° wall slope and a flat apex 6 cm across would have more than twice the surface area of an equivalent flat disc (996 vs. 380 cm2). Allowing for enhanced pigment concentration at the apex, the overall average chlorophyll-a concentration per unit bed area of a cone would be approximately 35.5 μg cm−2 compared to 9.5 μg cm−2 for flat mat.
The very thin laminations in Lake Untersee compared to Lake Hoare suggest a lower rate of sediment and organic carbon accrual, even though the irradiance in Lake Untersee is higher than in Lake Hoare. New estimates (author’s unpublished data) of sedimentation in Lake Untersee, based on four replicate passive traps at 20 m depth, deployed for 2 years, yield an accumulation rate of 6 g m−2, which compares to 41 g m−2 for Lake Hoare (Wharton et al. 1989). New in situ measurements of oxygen concentration profiles at 20 m depth and a photon flux of 30 μmol m−2 s−1 in Lake Untersee (Fig. 4.5), obtained as described in Vopel and Hawes (2006), show that the zone of net oxygenic photosynthesis is restricted in Lake Untersee to ~1 mm, much narrower than Lake Hoare at 10 m depth (Fig. 4.5). However, the flux of oxygen to the overlying water calculated from this profile was 190 μmol m−2 h−1 or approximately half of that in Lake Hoare at 10 m depth with an irradiance of 3 μmol photons m−2 s−1 (Vopel and Hawes 2006). Thus Lake Untersee mats show lower gross photosynthesis and photosynthetic efficiency than Lake Hoare mats. As in Lake Hoare, there is no evidence for a sharp oxycline below the zone of oxygenic photosynthesis; instead there is a steady decline deep into the mat, repeating the apparent paradox of coincident oxygen and organic carbon in the aphotic mat seen in Lake Hoare.

Profiles of dissolved oxygen and pH through the surface layer of a cone from Lake Untersee. Profile obtained using a manual micromanipulator, and freshly calibrated Unisense Ltd. (http://www.unisense.com/) microelectrodes connected to a Unisense UW-M underwater picoammeter
Two likely reasons for the lower accumulation of extracellular organic carbon in Lake Untersee compared to Lake Hoare are geochemical and taxonomic differences. Lake Untersee has an unusual water chemistry, with a pH of >10.4 generated by weathering of anorthosite rock floor under an ice cover that effectively eliminates atmospheric contact (Kaup et al. 1988; Andersen et al. 2011). High pH persists some distance into the sediment, with a maximum coincident with the photosynthesis-induced dissolved oxygen maximum at the mat surface (Fig. 4.5). Recently, using techniques described by Hawes et al. (2011), we measured concentrations of dissolved inorganic carbon (DIC) in water from Lake Untersee at 12 μmol L−1, amongst the lower values recorded in natural waters. At pH 10.4, less than 50% of DIC would be in a photosynthetically available form, e.g. as bicarbonate. Thus the difference between the rate of accumulation of lamina thickness between Lake Untersee and Lake Hoare may be due to limitation by both sediment deposition and carbon accrual rates.
A second substantial difference between the lakes is the dominance of Phormidium taxa in the phototrophic community in Lake Untersee, whereas in Lake Hoare mats are dominated by a mix of Leptolyngbya and diatoms. Phormidium tends to be a highly motile group, occurs at high pH in Antarctic lakes (Mackey et al. 2015) indicating an affinity for bicarbonate and is characterised as a “surface-smoothing” taxon (Mackey et al. 2015). In contrast Leptolyngbya is less motile, but is associated with topographical enhancement through vertically oriented growth (Reyes et al. 2013; Mackey et al. 2015). The surface topography, with filaments of Phormidium autumnale projecting from the top of each mound in a loose, woolly surficial layer, may represent a behavioural response to optimise carbon accrual at low DIC availability; the ability of cyanobacteria to move along light and DIC gradients is well known (see review by Stahl 1995). At low irradiance a tendency for trichomes to accumulate at the surface of topographic highs, through orientation towards the light, and then to extend vertically through a pH and DIC gradient, is consistent with known behaviours. Enhanced accumulation of sediment, on the flattened apices of cones is also likely due to the enhanced trapping ability of the “woolly cap” of P. autumnale trichomes and may also contribute to the differential accumulation of mass on the apex of the cone compare to the cone sides and surrounding lake floor.
Once a biomass maximum is established on a cone apex, the morphology is likely to become a self-reinforcing feature. Bosak et al. (2012) showed that the tips of pinnacles grow faster than the surrounding bases because of higher density of active cells and, like many other authors, suggested that this reflects enhanced access to water column nutrients for vertical elements projecting though the hydraulic laminar sub-layer. This advantage is most effective in situations where near-laminar flow occurs at the lake bed. Unlike many ice-covered lakes, most of the water column of Lake Untersee is mixed at least some of the time (Wand et al. 1997). Mixing in the lake is indicated by homogenous conductivity profiles with depth through the >140 m water column and is thought to be driven by a mix of solar heating and the generation of cold, dilute and buoyant water through subsurface melting of the Anuchin Glacier that forms the northern boundary of the lake (Wand et al. 1997; Andersen et al. 2011). Thus water movement, albeit slight, occurs in the lake, and sufficient water movement may occur to result in significant near-bed velocity gradients. Evidence in favour of slow flow at the lake bed that interacts with cone apices is provided by the apparent unidirectional drift of trichomes from apical concentrations of P. autumnale evident in vertical images of the lake floor taken with a drop camera (Fig. 4.6). Divers operating in the lake have never detected any evidence of water movement, even when disturbing bottom sediments during sample collection, and we suspect that at most flows are slow and laminar rather than turbulent. However, it is possible that the presence of tall, smooth cones within moving water may enhance nutrient supply to taller “canopy” elements of benthic communities over what would accrue on flats as they would be placed in higher velocity parts of the laminar flow gradient (Ghisalberti et al. 2014).

Vertical image of the floor of Lake Untersee at 45 m depth obtained with a low-resolution SeaView (http://www.seaview.com/) drop camera lowered through a hole in the ice. The dark spots are the apices of mounds, and the streaks trailing from the apices are trails of cyanobacteria. The bar represents ~2 m
Nepf (2012) reviewed the hydrodynamics of vegetated channels, providing an understanding of the role of canopy density on near-bed turbulence regime. She noted that sparse canopies, as in Lake Untersee, resemble a simple boundary layer regime with a velocity gradient predicted by distance from the channel bed, enhancing the probability that the cone apices would experience higher velocity environments than cone bases. While the conical structures allow an increase in surface area for uptake of nutrients, a possible specific functional advantage of the conical structures in Lake Untersee may thus emerge as placing the most actively growing part of the microbial population in the location where they are most hydrodynamically favoured to exploit otherwise limiting nutrients, in this case perhaps DIC.
The growth of large conical structures in Lake Untersee may be the consequence of the responses of the constituent organisms growing under unusual field conditions. Cones may naturally develop on local topographic irregularities, which propagate through enhanced growth and sediment accumulation on the apex. This model implies no complex, holistic organisation of structure. However, one observation challenges this simple interpretation. The spacing of cones shows marginally significant regularity, based on analysis of six down-looking images of cones, using nearest neighbour analysis (Clarke and Evans 1954). Cone spacing was on scales of decimetres or more, yet the R n statistic (n-sample nearest neighbour risk) of the nearest neighbour analysis for the image in Fig. 4.6 was 1.29 (where 1 is random and >1.2 indicates a significant element of regularity—analysis was at N > 200 and p > 0.05).
Overall the R n for the six analysed images was 1.36 ± 0.06 (average ± s.d.). While this statistic suggests a low level of ordering, any emergence of regular spacing may imply information transfer between cones, and how this may occur is not clear. The potential for competitive interaction for water column resources to result in regular spacing in pinnacle mats has been argued for on the basis of elegant experimentation and modelling by Petroff et al. (2010). Those arguments suggest that under still conditions, spacing on cm distance scales will result from pulsed activity on a 24 periodicity (i.e. photosynthesis). Even in Antarctic lakes, with 24 h of daylight, sufficient day-night variation in irradiance exists to drive a diel photosynthetic pattern (Hawes et al. 2014). Thus in still water, this mechanism might explain a cm scale of spacing of pinnacles (Andersen et al. 2011) but not a greater than dm-scale spacing of large cones, a point noted by Petroff et al. (2010). However, as discussed above, laminar flow of water is suspected in Lake Untersee, and Petroff et al. (2010) went on to argue that larger-scale spacing patterns may be propagated by the competitive interaction mechanism in directional flowing water. They suggest that laminar flow would result in longitudinal regularity, or ridges, rather than the distribution of cones seen in Lake Untersee.
As with Lake Hoare, the emergent structures in Lake Untersee, here a mixture of one-dimensional laminations elaborated to large conical forms, appear to largely result from uncoordinated responses of organisms to growth conditions. However, the enigmatic tendency for cones to be spaced in a non-random pattern over dm distances is unexplained. A mechanism whereby communication between cones could occur over such distances is not clear, and given the expectation of minimal competitive interaction at such scales, neither is any holistic advantage that such communication may confer.
4.4.2 Lake Vanda
If there is a “model” emergent structure for microbial mats that has dominated recent literature, it is that of cm-scale spaced, cuspate pinnacles, associated with narrow-trichome cyanobacterial taxa (Petroff et al. 2010; Tice et al. 2011; Bosak et al. 2012). This morphology and taxonomic dominance is common in Lake Vanda (Hawes et al. 2013; Zhang et al. 2015; Sumner et al. 2016). For logistic reasons, published information is largely limited to the upper part of the lake, from 6 to 26 m depth where a time series of mat age that has resulted from a gradual increase in lake level allows insights into the development of these microbial pinnacles (Hawes et al. 2013; Sumner et al. 2016). The number of annual growth laminations and accumulation of biomass along the depth gradient in the upper water column of Lake Vanda was consistent with an accumulation of mat material over the duration of inundation calculated from rising water level (Hawes et al. 2013). The gradual increase in height and spacing of pinnacles with depth (Fig. 4.7) was therefore consistent with temporal development. Recently, we deployed drop cameras across Lake Vanda and found that pinnacles extend to depths of at least 55 m (Fig. 4.8), though most pinnacles at this depth appear to have collapsed, perhaps a result of declining irradiance over time as the lake level has risen. The potential for insights into the mechanisms of pinnacle growth from Lake Vanda are high because pinnacles can be ordered by age, are particularly large and are readily measured. Thus, we focus on both the organisation of fields of pinnacles and possible functions of the pinnacle structure itself.

(a) Spacing and (b) height of pinnacles along a depth gradient in Lake Vanda. (c) The relationship between spacing and height at each sample depth. Spacing and height are not normally distributed, and the median and quartiles are shown. Modified after Hawes et al. (2013)

The appearance of pinnacle mats from a range of depths in Lake Vanda. The physical depth and optical depth, as % incident irradiance at noon, are indicated. Where visible, the red laser dots are 3 cm apart
4.4.2.1 Size and Spacing
Distributions of pinnacle height at any given depth are skewed by the presence of exceptionally large pinnacles at all depths (Fig. 4.9). There was a close, positive correlation between median size at each depth and median spacing (Fig. 4.7c, n = 7, r 2 = 0.96, p < 0.0005). Nearest neighbour analysis of the Vanda pinnacles showed, as in Lake Untersee, a slight tendency away from random, towards even spacing. For three quadrats taken at 21 m depth, R n was calculated from 80 measures of nearest neighbour difference as 1.25 ± 0.01, while for three quadrats at 8 m depth R n was 1.25 ± 0.02 (N = 3, mean ± s.d.), suggesting a weakly significant effect (p > 0.05). When pinnacle spacing was examined more closely, it was apparent that deviation from randomness was largely driven by truncation of the distribution at the closely spaced end. This was evident for both the larger, well-spaced pinnacles at 22 m depth (Fig. 4.9) and also the small, closely packed pinnacles at 8 m (8 m data not shown).

Log-normal distributions of pinnacle height at three depths in Lake Vanda and of spacing distance at 21 m depth
Two simple ways to explain this observation are that (1) at short separation, it is not possible to detect accurately small spacing between pinnacles, due to their finite basal diameter, or (2) interactions lead either to divergence or merging of pinnacles at short range. Figure 4.10 shows how closely associated tall pinnacles can tend to both merge at the base and lean away from each other, suggesting that both mechanisms may be in play at least some of the time. Divergence of closely associated pinnacles is consistent with the concept of competition for nutrients from the water column, or perhaps to minimise mutual shading. In contrast, the correlation between spacing and size is consistent with the basal merging of pinnacles to produce fewer, larger individuals as the microbial mat ages. The distribution of pinnacles in Lake Vanda provides only partial support for the view of emergence of structural complexity driven by inter-pinnacle competition for water-column resources. Rather the simplest explanation is of random initiation (see Sumner et al. 2016), merging as basal area increases, with direct inter-pinnacle interactions only being evident at close range. This type of distribution pattern can be fully accommodated by simple interactions between organisms and environment rather than needing to invoke a more holistic mechanism for pinnacle growth.

Pinnacle mats at 22 m water depth in Lake Vanda. At the centre, a cluster of large pinnacles shows a tendency to merge at the base but for the pinnacles themselves to diverge
4.4.2.2 Structure
While data from Lake Vanda suggest that pinnacle fields tend to be structured by simple mechanisms that occur at the individual pinnacle level, the prevalence of the pinnacle structure across a range of optical depths spanning at least 1–15% incident irradiance implies that it has some functional advantage. It is noteworthy to compare the pinnacles of microbial mats in Lake Vanda at 1.5% surface irradiance (45 m depth, Fig. 4.8), with the much flatter structure of Lake Hoare at similar optical depth (Fig. 4.2a). It appears that, based on molecular taxonomy, the two lakes share dominant cyanobacteria (Zhang et al. 2015), and thus behavioural differences, as proposed for Lake Untersee, are less likely. What are the functional advantages that the formation of pinnacles confer on the microbial communities in Lake Vanda, and why is this advantage enhanced for a small subset of the pinnacle population resulting in the frequent occurrence of a few “outsize” pinnacles?
The evolution of pinnacle morphology and zonation in Lake Vanda is described by Sumner et al. (2016). In brief, all pinnacles and flat mats share a laminated and zoned structure; the outer 3–4 laminae are orange-brown in colour, with green and purple subsurface laminae. The transition from the surface orange-brown zone to the green and purple zones is abrupt and only occasionally crosses lamina boundaries (Fig. 4.11). The outer laminae were typically less than 0.5 mm thick each (except at pinnacle tips), whereas the thicknesses of green/purple subsurface laminae ranged from less than 1 to greater than 3 mm thick. The shapes of the green/purple subsurface laminae towards the tips of pinnacles differed from those of the orange-brown laminae; the tops of these laminae were conical to geniculate (Fig. 4.11), rather than cuspate. Thus, the overall thickness of a single green/purple lamina was more uniform across the pinnacle than any orange-brown laminae were, and no green/purple laminae mimicked the cuspate morphology of pinnacle tips. Sumner et al. (2016) noted that at young stages, pinnacles only comprised the outer orange-brown zone, with the green and purple zones only extending into the pinnacles at later stages of development. However, once the pinnacles contain green and purple zones, the interior laminae grow and thicken, which causes the lateral expansion of pinnacles and results in a columnar pinnacle morphology (Fig. 4.12; Sumner et al. 2016). 16S rDNA clone library techniques show the orange-brown laminae in the pinnacles contain almost exclusively Leptolyngbya ribotypes (80% L. antarctica), while the green and pink zones contain 40–50% L. antarctica and the remainder larger ribotypes, primarily Phormidium and Tychonema (Sumner et al. 2016).

Pinnacles from 19 m depth in Lake Vanda. (a) A pinnacle developing “columnar” attributes with the exterior dominated by orange-brown laminae. The vertical section on the right shows the thick purple and green subsurface zones that cause the columnar shape. (b) A smaller, cuspate pinnacle that has similar exterior orange-brown laminae. The vertical section on the right illustrates the near absence of purple and green subsurface zones except near the base and near sediment at the crest

Sketch of pinnacle geometry and growth history. After several years of prostrate growth, microbial tufts nucleate in orange-brown laminated prostrate mats and grow into small cuspate pinnacles. Once thick enough, a green/pink cyanobacterial community colonises the interior of the mat, and over time more deeply buried layers begin to degrade. Apical growth exceeds the flat surrounding mat, and as pinnacles continue to grow, the green/pink community colonises the interiors of pinnacles and begins to swell. Formerly cuspate laminae become geniculate as biomass increases within the laminae. A shoulder develops on the pinnacle and propagates upward through time. Eventually, the cores of pinnacles begin to degrade. Modified after Sumner et al. (2016)
As indicated in Fig. 4.12, tall, columnar pinnacles take considerable time to emerge, and it is important to recall that pinnacle growth is a function of the differential between growth of the apex and the surrounding base. Hawes et al. (2013) calculated that during the first decades of growth, prostrate mat accumulated at approximately 0.33 mm year−1, while pinnacle apices extended at 0.25 mm year−1, a net growth rate of 0.48 mm year−1 for pinnacles. Thus the differential between the rate of growth of pinnacles and the base mat is slight—on average. However, some pinnacles increased in height by more than 2 mm year−1 over a decade (Sumner et al. 2016). Thus, the large size of pinnacles in Lake Vanda attests to both their age and variations in their growth rate. Hawes et al. (2013) calculated that the pinnacles at 20 m depth may be the result of 60–80 years of undisturbed growth.
Positive differential growth relative to background mat requires that the pinnacle morphology must provide a growth advantage to the apex. This may relate to interactions between flow of water around the pinnacle fields and nutrient supply, as discussed above for Lake Untersee. As in Untersee, parts of Lake Vanda are convectively mixed, and currents of up to 1 cm s−1 have been reported in the lake (Ragotzkie and Likens 1964). While DIC is not depleted in Lake Vanda as in Lake Untersee, dissolved reactive phosphorus is thought to be limiting (Vincent and Vincent 1982). Ghisalberti et al. (2014), like others, argue that tall organisms—or in this case pinnacles—benefit by extracting more nutrients from water columns due to the effect of the near-bed velocity gradient.
Nepf (2012) points out that the movement of water within a canopy decreases proportionally to the density of the canopy elements, enhancing this effect. Canopies significantly affect turbulence, and when the product of volume-specific frontal area of canopy elements and their height exceeds 0.23, the canopy slows water sufficient to generate a shear layer at the top of the canopy (Nepf 2012). The product of volume-specific frontal area and height in Lake Vanda pinnacle fields at 20 m is 0.3—exceeding the critical value of Nepf (2012). Thus, a shear layer is likely near the tops of pinnacles. If limiting nutrients are being derived from the water column, the density of the pinnacle field in Lake Vanda is sufficient to affect water turbulence, leading to enhanced nutrient supply and thus growth to pinnacle apices. Indeed, the presence of a velocity gradient displaced from the lake bed to the level of the median pinnacle height may also provide a growth advantage to isolated extra-tall pinnacles and thus reinforce the differential between median and quartile sizes.
While this model is attractive, there is little evidence to suggest that enhanced nutrient availability is effective. The N:P ratios of pigmented outer layers taken from pinnacles and from adjacent flat mats are both high, consistent with P limitation, but there are no significant differences that would suggest an enhanced nutrient supply, relative to growth, for pinnacles over flat mats (Table 4.2). Conditions in Lake Vanda, where currents, though measured, are slight (no visible movement of disturbed sediment has ever been seen by divers operating in Lake Vanda), may not be well suited to hydraulic enhancement of growth of pinnacles. Indeed, it has been shown that nutrient concentrations in microbial mat interstitial waters can exceed that in overlying water, suggesting that internal recycling is a significant potential source of nutrients (Quesada et al. 2008) and this may be expected to be most effective over short-length scales rather than in elongated pinnacles.
An alternative growth model is differential photosynthesis rates as observed by Bosak et al. (2012). To determine whether net photosynthetic capacity per unit area increased towards the apices of the pinnacles, we collected three pinnacles and returned them to a lakeside laboratory and maintained them at ambient photon flux (40 μmol m−2) and temperature (4 °C) in static lake water for 3 h to reach equilibrium with respect to oxygen concentration profile. Oxygen profiles were measured with a Unisense microelectrode system and the gradient of dissolved oxygen at equilibrium was measured at approximately 1 cm intervals along the length of each pinnacle, and this used to calculate the diffusive flux of oxygen out of the microbial mat using Fick’s law (methods fully described in Vopel and Hawes 2006). No consistent evidence emerged for any enhanced net rate of photosynthesis along the length of the pinnacles (Fig. 4.13). Similarly, Sumner et al. (2016) did not observe increased photosynthetic potential near pinnacle tops using PAM fluorescence techniques.

Oxygen efflux rates at simulated ambient light and temperature conditions at approximately 1 cm intervals along three representative pinnacles from Lake Vanda
While photosynthesis potential per unit area is not enhanced towards pinnacle apices, there is no doubt that surface area is. As calculated for Lake Untersee, the increase in surface area conveyed by the pinnacle morphology results in an increase in pigment biomass per unit of lake bottom (Table 4.1) and, if sufficient irradiance is received, will result in a greater overall accumulation of carbon per unit lake bottom area than for flat mats. The simplest interpretation of cumulative data on pinnacles in Lake Vanda is that the functional benefit of forming elaborate pinnacle structures is to enhance surface area. The pinnacles are physiologically and compositionally simply flat mats that have been stretched and with a more vertical orientation. Photosynthetic tissue is thus spread over a larger area than in flat oriented mats, resulting in the same number of thinner laminae each accumulating less falling sediment per unit area, due to steeper slope. Reduced sediment load minimises competition for light absorption between sediment and photosynthetic pigments, high surface area allows ready access to water column nutrients, and thin laminae allow ready access to nutrients regenerated in decomposing material below. Within these stretched outer pinnacle laminae, trichomes of Leptolyngbya are oriented parallel to the axis of the pinnacle (authors unpublished data and Sumner et al. 2016), often intertwined to form the network of meshlike skeletons that if growing by extension will naturally result in enhanced apical growth. We argue that the small growth advantage that pinnacles have over flat mats relates to this process of increased surface area and oriented extension.
The surface area mechanisms will only be effective if the steep sides of pinnacles receive adequate illumination. The angular distribution of light in Lake Vanda at 10 m depth is moderately diffuse due to the scattering nature of the lake ice cover (Sumner et al. 2016). Radiance is maximal at across a 60° arc about the vertical, and substantial light is still present at angles close to 60° from zenith. The light reaching the steeply sloping faces of pinnacles is only slightly less than that reaching horizontal surfaces, particularly as the thinning of pigmented laminae will increase the penetration of light through the pinnacle walls (c.f. Hawes and Schwarz 2001). As in Lake Hoare and Lake Untersee, the pinnacles in Lake Vanda appear to be predictable and to emerge on the basis of simple organism response to environmental variables, over long periods of stable conditions.
4.5 Conclusions
In this brief review of the function of structure in microbial mats in Antarctic lakes, we have focused on features of these unusual systems that may provide insights into general features of microbial mats. Structures formed in Antarctic lakes can be particularly well developed, due to the extreme lack of disturbance that allows them to accumulate over very long time periods (decades perhaps to centuries). Structure is evident on both 1-D and 3-D bases. 1-D structure comprises two elements, an annual banding due to temporal separation of sedimentation and carbon accrual and a pigment zonation superficially similar to the metabolic zonation seen in some more typical mats, though lacking any linkage to oxygen and sulphide gradients. Instead the pigment zonation can be reconciled to the persistence of organisms, particularly cyanobacteria, in buried laminae for many years, and penetration of just enough light to allow these to survive and even grow in some instances. There is no clear indication in data available to date that this zonation provides any holistic advantage to the mat components through, for example, cascades of nutrient cycling elements. Further investigations are needed to better understand the bacterial and microbial eukaryote assemblages as well as functional potential of the communities, using in-depth environmental genomics analyses.
Antarctic lakes provide excellent opportunities to increase understanding of the emergence of complex 3-D structures in microbial mats, because they display a wide range of such structure most of which are non-lithifying and which attain substantial sizes across environmental gradients. Limited data available to date confirm findings from elsewhere that the interaction of dominant organisms present and growth conditions are important in determining the outcome of 3-D emergence. We note that elaboration occurs slowly, resulting from small differences in growth rate in various parts of a microbial mat community. For example the spectacular pinnacles in Lake Vanda, which reach over 10 cm high, accrue at annual rates on the order of mm or less, but the rate of emergence of complex features in other less stressful locations may scale with growth rate. Thus rather small differentials in growth may be responsible for emergence of structures. Functional benefits of these structures may relate to enhanced resource availability through increased surface area, and possibly interactions of structures with water movement to enhance delivery of resources.
While we were able to show how the increase in surface area of elaborated structures over flat ones allows greater biomass and overall productivity on a lake bed area basis, the structures that formed could be easily understood in terms of potential interactions between individuals, their orientation and their environment. The data lack strong evidence of coordinated behaviour, directed towards holistic advantages to the structure. The possibility of some level of coordinated behaviour is, however, suggested by the spacing of the structural elements. Initiation of structures in the lakes considered here was close to random, and the growth of pinnacles in Lake Vanda appears to result in a gradual increase in spacing, following a log-normal distribution, due to merging of near neighbours. However, though a hint of regular spacing in both Lake Vanda and Lake Untersee does argue for some inter-communication between pinnacles, more focused research will be required to investigate this intriguing possibility. The large size of structures in Antarctic lakes make them particularly suitable to investigation of any role of cell to cell signalling over short or long spatial scales in structural emergence that occur on small spatial scales in biofilms (Battin et al. 2007; Decho et al. 2010).
References
Andersen DT, Sumner DY, Hawes I, Webster-Brown J, McKay CP (2011) Discovery of large conical stromatolites in Lake Untersee, Antarctica. Geobiology 9:280–293
Battin TJ, Sloan WT, Kjellberg S, Damis H, Head IM, Curtis TO, Eberl L (2007) Microbial landscapes: new paths to biofilm research. Nat Rev Microbiol 5:76–81
Bosak T, Liang B, Wu T-D, Templer SP, Evans A, Vali H, Guerquin-Kern J-L, Klepac-Ceraj V, Sim MS, Mui J (2012) Cyanobacterial diversity and activity in modern conical microbialites. Geobiology 10:384–401
Bosak T, Knoll AH, Petroff AP (2013) The meaning of stromatolites. Annu Rev Earth Planet Sci 41:21–44
Cathey DD, Parker BC, Simmons GM, Yongue WH, Van Brunt MR (1981) The microfauna od algal mats and artificial substrates in Southern Victoria Land lakes of Antarctica. Hydrobiologia 85:3–15
Clark PJ, Evans FC (1954) Distance to nearest neighbour as a measure of spatial relationships in populations. Ecology 35:445–453
Corning PA (2002) The re-emergence of “emergence”: a venerable concept in search of a theory. Complexity 7:18–30
Dana GL, Wharton RR, Dubayah R (1998) Solar radiation in the McMurdo Dry Valleys, Antarctica. In: Priscu JC (ed) Ecosystem dynamics in a polar desert: The McMurdo Dry Valleys, Antarctica. American Geophysical Union, Washington, DC, pp 38–64
De Wolf T, Holvoet T (2005) Emergence versus self-organisation: different concepts but promising when combined. In: Brueckner S, Di Marzo Serugendo G, Karegeoros A, Nagpal R (eds) Engineering self organising systems: methodologies and applications. lecture notes in computer science. Springer, Berlin, pp 1–15
Decho AW, Norman RS, Visscher PT (2010) Quorum sensing in natural environments: emerging views from microbial mats. Trends Microbiol 18:73–80
Denis L, Gevaert F, Spilmont N (2012) Microphytobenthic production estimated by in situ oxygen microprofiling: short-term dynamics and carbon budget implications. J Soils Sediments 12:1517–1529
Des Marais DJ (1995) The biogeochemistry of hypersaline microbial mats. Adv Microb Ecol 14:251–274
Doran PT, Wharton RA, Lyons WB (1994) Paleolimnology of the McMurdo Dry Valleys, Antarctica. J Paleoliomnol 10:85–114
Doran PT, Priscu JC, Lyons WB, Welsh JE, Fountain AG, McKnight DM, Moorhead DL, Virginia RA, Wall DH, Clow GD, Fritsen CH, McKay CP, Parsons AN (2002) Antarctic climate cooling and terrestrial ecosystem response. Nature 415:517–520
Fenchel T (1998) Formation of laminated cyanobacterial mats in the absence of benthic fauna. Aquat Microb Ecol 14:235–240
Franks J, Stolz JF (2009) Flat laminated microbial mat communities. Earth Sci Rev 96:163–172
Ghisalberti M, Gold DA, Laflamme M, Clapham ME, Narbonne GM, Summons RE, Johnston DT, Jacobs DK (2014) Canopy flow analysis reveals the advantage of size in the oldest communities of multicellular eukaryotes. Curr Biol 24:305–309
Guerrero R, Berlanga M (2007) The hidden side of the prokaryote cell: rediscovering the microbial world. Int Microbiol 10:157–168
Haendel D, Kaup E, Loopman A, Wand U (1995) Physical and hydrochemical properties of water bodies. In: Bormann O, Fritzsche D (eds) The Schirmacher Oasis, Queen Maud Land, East Antarctica, and its surroundings. PGM Ergh 289. Perthes, Gotha, pp 259–319
Halley JD, Winkler DA (2008) Classification of emergence and its relation to self-organization. Complexity 13:10–15
Hawes I, Schwarz A-M (1999) Photosynthesis in an extreme shade environment: benthic microbial mats from Lake Hoare, a permanently ice-covered Antarctic lake. J Phycol 35:448–459
Hawes I, Schwarz A-M (2001) Absorption and utilization of irradiance by cyanobacterial mats in two ice-covered Antarctic lakes with contrasting light climates. J Phycol 37:5–15
Hawes I, Schwarz A-M, Smith R, Howard-Williams C (1999) Environmental conditions during freezing, and response of microbial mats in ponds of the McMurdo Ice Shelf, Antarctica. Antarct Sci 11:198–208
Hawes I, Moorhead D, Sutherland D, Schmeling J, Schwarz A-M (2001) Benthic primary production in two perennially ice-covered Antarctic lakes: patterns of biomass accumulation with a model of community metabolism. Antarct Sci 13:18–27
Hawes I, Sumner DY, Andersen DT, Mackey TJ (2011) Legacies of recent environmental change in the benthic communities of Lake Joyce, a perennially ice covered, Antarctic lake. Geobiology 9:394–410
Hawes I, Sumner D, Andersen D, Jungblut A, Mackey T (2013) Timescales of growth response of microbial mats to environmental change in an ice-covered Antarctic lake. Biology 2:151–176
Hawes I, Giles H, Doran PT (2014) Estimating photosynthetic activity in microbial mats in an ice-covered Antarctic lake using automated oxygen microelectrode profiling and variable chlorophyll fluorescence. Limnol Oceanogr 59:674–688
Hawes I, Jungblut AD, Obryk MK, Doran PT (2016) Growth dynamics of laminated microbial mats in response to variable irradiance in an Antarctic lake. Freshw Biol 61:396–410
Howard-Williams C, Pridmore RD, Downes MT, Vincent WF (1989) Microbial biomass, photosynthesis and chlorophyll a related pigments in the ponds of the McMurdo Ice Shelf, Antarctica. Antarct Sci 1:125–131
Howard-Williams C, Schwarz A-M, Hawes I (1998) Optical properties of the McMurdo Dry Valley Lakes, Antarctica. In: Priscu JC (ed) Ecosystem dynamics in a Polar Desert: the McMurdo Dry Valleys, Antarctica. American Geophysical Union, Washington, DC, pp 189–205
Jungblut AD, Wood S, Hawes I, Webster-Brown J, Harris C (2012) The Pyramid Trough Wetland: environmental and biological diversity in a newly created Antarctic protected area. FEMS Microb Ecol 82:356–366
Karanovic T, Gibson J, Hawes I, Andersen D, Stevens M (2014) Three new species of Diacyclops (Copepods: Cyclopoida) from continental Antarctica. Antarct Sci 26:250–260
Kaup E, Loopman A, Klokov V, Simonov I, Haendel D (1988) Limnological investigations in the Untersee Oasis during the summer season 1983/84. In: Martin J (ed) Limnological studies in Queen Maud Land. Academy of Sciences, Tallinn, Estonia, pp 43–56
Kühl M, Fenchel T (2000) Bio-optical characteristics and the vertical distribution of photosynthetic pigments and photosynthesis in an artificial Cyanobacterial mat. Microb Ecol 40:94–103
Lizotte MP, Priscu JC (1992) Photosynthesis-irradiance relationships in phytoplankton from the physically stable water column of a perennially ice-covered lake (Lake Bonney, Antarctica). J Phycol 28:179–185
Love FG, Simmons GM, Parker BC, Wharton RA, Seaburg KG (1983) Modern conophyton-like microbial mats discovered in Lake Vanda, Antarctica. Geomicrobiol J 3:33–48
Mackey TJ, Sumner DY, Hawes I, Jungblut A, Andersen DT (2015) Branched columnar stromatolites in Lake Joyce. Antarct Geobiol 13:373–390
Nadell D, Drescher K, Foster KR (2016) Spatial structure, cooperation and competition in biofilms. Nat Rev Microbiol 14:589–600
Nepf HM (2012) Hydrodynamics of vegetated channels. J Hydraul Res 50:262–279
Ng WO, Grossman AR, Bhaya D (2003) Multiple light inputs control phototaxis in Synechocystis sp. strain PCC6803. J Bacteriol 185:1599–1607
Oren A (2010) Mats of filamentous and unicellular cyanobacteria in hypersaline environments. In: Seckbach J, Oren A (eds) Microbial mats: modern and ancient microorganisms in stratified systems. Springer, Dordrecht, pp 387–400
Paerl HW, Pinckney JL (1996) A mini-review of microbial consortia: their roles in aquatic production and biogeochemical cycling. Microb Ecol 31:225–247
Parker BC, Simmons GM, Love FG, Wharton RA, Seaburg KG (1981) Modern stromatolites in Antarctic Dry Valley lakes. Bioscience 31:656–661
Petroff AP, Sim MS, Maslov A, Krupenin M, Rothman DH, Bosak T (2010) Biophysical basis for the geometry of conical stromatolites. Proc Natl Acad Sci 107:9956–9961
Petroff AP, Beukes NJ, Rothamn DH, Bosak T (2013) Biofilm growth and fossil form. Phys Rev. https://doi.org/10.1103/PhysRevX.3.041012
Quesada A, Fernández-Valiente E, Hawes I, Howard-Williams C (2008) Benthic primary production in polar lakes and rivers. In: Vincent WF, Laybourn-Parry J (eds) Polar lakes and rivers–Arctic and Antarctic aquatic ecosystems. Oxford University Press, Oxford, pp 179–196
Ragotzkie RA, Likens GE (1964) The heat balance of two Antarctic lakes. Limnol Oceanogr 9:412–425
Reid RP, Visscher PT, Decho AW, Stolz JF, Beboutk BM, Dupraz C, Macintyre IG, Paerl HW, Pinckney JL, Prufert-Bebout L, Steppe TF, DesMarais DJ (2000) The role of microbes in accretion, lamination and early lithification of modern marine stromatolites. Nature 406:989–992
Reyes K, Gonzalez NI, Stewart J, Ospino F, Nguyen D, Cho DT, Ghahremani N, Spear JR, Johnson HA (2013) Surface orientation affects the direction of cone growth by Leptolyngbya sp. strain C1, a likely architect of coniform structures Octopus Spring (Yellowstone National Park). Appl Environ Microbiol 79:1302–1308
Seckbach J, Oren A (2010) Microbial mats: modern and ancient microorganisms in stratified systems. Cellular origin, life in extreme habitats and Astrobiology 14. Springer
Shepard RN, Sumner DY (2010) Undirected motility of filamentous cyanobacteria produces reticulate mats. Geobiology 8:179–190
Spigel RH, Priscu JC (1998) Physical limnology of the mcmurdo dry valley lakes. In: Priscu JC (ed) Ecosystem dynamics in a Polar desert: the mcmurdo dry valleys, Antarctica. AGU, Washington, DC, pp 153–189
Stahl LJ (1995) Physiological ecology of cyanobacteria in microbial mats and other communities. New Phytol 131:1–32
Stahl LJ, Bolhuis H, Cretoiu MM (2018) Phototrophic marine benthic microbiomes: the ecophysiology of these biological entities. Environ Microbiol. https://doi.org/10.1111/1462-2920.1449
Sumner DY, Jungblut A-D, Hawes I, Andersen DT, Mackey TJ, Wall K (2016) Growth of elaborate microbial pinnacles in Lake Vanda, Antarctica. Geobiology 14(6):559–574. https://doi.org/10.1111/gbi.12188
Sutherland DL, Hawes I (2009) Annual growth layers as proxies of past growth conditions for benthic microbial mats in a perennially ice-covered Antarctic lake. FEMS Microb Ecol 67:279–292
Tice MM, Thornton DCO, Pope MC, Olszewski TD, Gong J (2011) Archean microbial mat communities. Annu Rev Earth Planet Sci 39:297–319
Tyler SW, Cook PG, Butt AZ, Thomas JM, Doran PT, Lyons WB (1998) Evidence of deep circulation in two deep perennially ice-covered Antarctic Lakes. Limnol Oceanogr 43:625–635
Vincent WF (2000) Cyanobacterial dominance in polar regions. In: Whitton BD, Potts M (eds) The ecology of cyanobacteria. Kluwer Academic Publishers, Dordrecht, pp 321–340
Vincent WF, Vincent CL (1982) Factors controlling phytoplankton production in Lake Vanda (77°S). Can J Fish Aquat Sci 39:1602–1609
Vopel K, Hawes I (2006) Photosynthetic performance of benthic microbial mats in Lake Hoare, Antarctica. Limnol Oceanogr 51:1801–1812
Walter MR, Bauld J, Brock TD (1976) Distribution and structure of recent stromatolitic algal mats. In: Walter MR (ed) Stromatolites. Elsevier, Amsterdam, pp 273–310
Wand U, Schwarz G, Brüggemann E, Aüuer K (1997) Evidence for physical and chemical stratification in Lake Untersee (central Dronning Maud Land, East Antarctica). Antarct Sci 9:43–45
Wand U, Samarkin V, Nitzsche H, Hubberten H (2006) Biogeochemistry of methane in the permanently ice-covered Lake UNtersee, central Dronning Maud Land, East Antarctica. Limnol Oceanogr 51:1180–1194
Webb JS, Givskov M, Kjelleberg S (2003) Bacterial biofilms: prokaryotic adventures in multicellularity. Curr Opin Microbiol 6:578–585
Wharton RA Jr, Parker BC, Simmons GM Jr (1983) Distribution, species composition and morphology of algal mats in Antarctic dry valley lakes. Phycologia 22:355–365
Wharton RA Jr, McKay CP, Clow GD, Andersen DT (1993) Perennial ice covers and their influence on Antarctic lake ecosystems. In: Green WJ, Friedmann EI (eds) Physical and biogeochemical processes in Antarctic lakes. Antarctic research series. American Geophysical Union, Washington, DC, pp 53–70
Wharton RA Jr (1994) Stromatolitic mats in Antarctic lakes. In: Bertrand-Sarfati J, Monty C (eds) Phanerozoic stromatolites II. Kluwer, Dordrecht, pp 53–70
Wharton RA Jr, Simmons GM, McKay CP (1989) Perennially ice-covered Lake Hoare, Antarctica: physical environment, biology and sedimentation. Hydrobiologia 172:305–320
Wharton RA Jr, McKay CP, Clow GD, Andersen DT, Simmons GM, Love FG (1992) Changes in ice cover thickness and lake level of Lake Hoare, Antarctica: implications for local climate change. J Geophys Res 97:3503–3513
Zhang L, Jungblut AD, Hawes I, Andersen DT, Sumner DY, Mackey TJ (2015) Cyanobacterial diversity in benthic mats of the McMurdo Dry Valley Lakes, Antarctica. Polar Biol 38:1097–1110
Acknowledgments
The information presented in this contribution is derived from field and laboratory work that would have been impossible without the support of many people and organisations. Logistic support was provided by Antarctica New Zealand, the US Antarctic Program and Antarctic Logistics Centre International. We thank all of our Antarctic field colleagues, in particular Drs Dale Andersen and Tyler Mackey, without whom the underwater research would not have been possible. We also thank the reviewers and editors for helpful comments and hard work to see the volume to production.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Ethics declarations
Funding
This study was funded by NASA Astrobiology: Exobiology and Evolutionary Biology (NNX08AO19G and NN13AE77A), the New Zealand Ministry of Business Innovation and Employment (CO1605 and UOWX1401), the National Science Foundation (MCM-LTER grant number 1115245) and the Tawani Foundation.
Conflict of Interest
Ian Hawes declares that he has no conflict of interest. Dawn Sumner declares that she has no conflict of interest. Anne D. Jungblut declares that she has no conflict of interest.
Ethical Approval
This article does not contain any studies with human participants or animals performed by any of the authors.
Rights and permissions
Copyright information
© 2019 Springer Nature Switzerland AG
About this chapter

Cite this chapter
Hawes, I., Sumner, D., Jungblut, A.D. (2019). Complex Structure but Simple Function in Microbial Mats from Antarctic Lakes. In: Hurst, C. (eds) The Structure and Function of Aquatic Microbial Communities. Advances in Environmental Microbiology, vol 7. Springer, Cham. https://doi.org/10.1007/978-3-030-16775-2_4
Download citation
DOI: https://doi.org/10.1007/978-3-030-16775-2_4
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-16773-8
Online ISBN: 978-3-030-16775-2
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)