
Abstract
The lack of baseline data has hindered the assessment of impacts from large-scale oil spills throughout their history. Baseline data collected before an adverse event such as an oil spill are critical for quantifying impacts and understanding recovery rates to pre-spill levels. In the case of the two largest oil spills in the Gulf of Mexico (GoM), Deepwater Horizon and Ixtoc 1, the lack of comprehensive contaminant baselines limits our ability to project when the ecosystem will return to pre-spill conditions and assess the short- and long-term impacts of contamination on ecosystems. Beginning in 2011, we initiated comprehensive sampling in the GoM to develop broad-scale and Gulf-wide hydrocarbon contaminant baselines primarily targeting continental shelf fishes in the USA, Mexico, and Cuba. We also developed a time series of collections over 7 years from the region in which DWH occurred. In the event there is another oil spill in the GoM, the samples from these baselines will provide broad-scale but not installation-specific baseline information for the assessment of impact and recovery. This chapter provides a summary of historical sampling and current baseline data for pelagic, mesopelagic, and demersal fish in the GoM. Further, we outline the importance of ongoing and more specific collection of monitoring data for hydrocarbon pollution.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
1 Introduction
Assessments evaluating the extent, duration of elevated concentrations, and impacts of oil spills are heavily reliant on data collected prior to, during, and after such events. A chronological sedimentary history of oil deposition can be recorded in cores (e.g., dated through radioisotopes or other means), assuming oil is deposited proportionally to the quantity released, bioturbation does not disturb the chronological order, and bacterial degradation is relatively slow. However, for living resources, depending upon how long after the event sampling occurs, contaminants within the animals may be reduced over time (via decontamination) and transformed into other compounds (via metabolism), some of which may be more toxic than the original exposure. For long-duration, post-spill monitoring assessments (e.g., several decades post-oil spill), few animals will record such events owing to their life spans. The most direct way to assess the effects and impacts of such spills is to have obtained pre-spill baselines beforehand with which to compare contamination levels and toxic biomarkers post-spill. Assessments of the impacts of the subsurface well blowout of the Mexican oil well Ixtoc 1 in 1979 were inconclusive due in part to the lack of pre-spill contaminant data (Amezcua-Linares et al. 2015). Ten years later, pre-spill data was dubbed one of the “rarest of all commodities” for researchers following the 1989 Exxon Valdez oil spill (Shigenaka 2014). Furthermore, 21 years subsequent to EVOS, researchers assessing the Deepwater Horizon oil spill (DWH, NAS 2013) lamented the virtual lack of relevant comprehensive pre-spill baselines, especially for living resources (Murawski and Hogarth 2013; Murawski et al. 2014). For decades, the unavailability of baseline data and reference sites in the Gulf of Mexico (GoM) has hindered the evaluation and health assessments of both the biota and the environment (Kennicutt et al. 1988; Lewis et al. 2002; Ward and Tunnell Jr. 2017). This chapter summarizes the availability and adequacy of PAH contamination data from fish tissues collected before and after the two largest spills in the GoM, the DWH and Ixtoc 1.
2 Pre-DWH Polycyclic Aromatic Hydrocarbon Baselines in Fish
In the USA, all 50 US states, the District of Columbia, the US territories of American Samos and Guam, and 5 Native American tribes issue fish consumption guidelines for potential health risks from consuming contaminated fish caught in their waters (US EPA 2009). Only five bioaccumulative chemicals (mercury, PCBs, chlordane, dioxins, and DDT) constitute 97% of all advisories in effect. Additionally, all five states bordering the GoM have coastal fish consumption guidelines, yet given the amount of oil and gas activity in the GoM, there are no active advisories or closures for petroleum-derived compounds. Fishery closures were in place during and just after the DWH accident, but most closures were lifted following seafood safety surveillance testing (Ylitalo et al. 2012). Importantly, while most fish tested post-spill were below levels of concern (LOC ), there were little data with which to judge if contamination levels had risen beyond pre-spill levels after the spill or to assess species most at risk.
Several programs have collected PAH contaminant data prior to DWH either as part of surveillance programs or in response to specific events. The National Oceanographic and Atmospheric Administration’s (NOAA ) National Status and Trends (NS&T) program undertakes nationwide sampling (i.e., Mussel Watch) to describe the current status and trends in the environmental quality of our estuarine and near-coastal waters (Farrington et al. 2016). Sampling included biliary PAH data for six inshore fish species (hardhead catfish, Ariopsis felis; Atlantic croaker, Micropogonias undulatus; red drum, Sciaenops ocellatus; spot, Leiostomus xanthurus; black drum, Pogonias cromis; and sand seatrout, Cynoscion arenarius) collected from 1985 to 1991 from along the US Gulf coast. Total biliary PAHs ranged from 7,000 to 1,000,000 ng FACs/g bile for both low- and high-molecular-weight fluorescent aromatic compounds (FACs ; NCCOS 2017). The highest concentrations were found in Atlantic croaker collected in 1989 from Barataria Bay, Louisiana. In general, the samples collected across all years from the northwest region of the GoM, along the Texas coast, had significantly higher biliary PAHs (300,000 ± 240,000 ng/g bile) than those collected in the north central region (p = 0.0046; Louisiana, Mississippi, Alabama: 160,000 ± 220,000 ng/g bile) and west Florida (p = 0.0054; 94,000 ± 100,000 ng/g bile).
In a separate study, hardhead catfish, Gulf killifish (Fundulus grandis), longnose killifish (F. majalis), and red drum were collected during 1990–1991 from Tampa and Sarasota bays to compare levels of organic contaminants between industrialized (Tampa Bay) and nonindustrialized areas (Sarasota Bay) (McCain et al. 1996). Mean biliary naphthalene equivalents ranged from 11,000 to 120,000 ng FAC/mg of biliary protein and 6,800 to 150,000 ng FAC/mg of biliary protein in the Tampa Bay and Sarasota Bay, respectively. Mean concentrations of the biliary naphthalene equivalents in hardhead catfish collected in Tampa Bay were all significantly higher than those collected from within Sarasota Bay. In addition, the biliary PAHs measured in fish collected within the more industrialized areas of Tampa Bay (i.e., Hillsborough Bay) were significantly higher than nearby nonindustrialized sites.
Edible muscle samples from hardhead catfish, channel catfish (Ictalurus punctatus), and largemouth bass (Micropterus salmoides) were surveyed in 1996 to compare the tissue quality between fish collected from wastewater-impacted areas and reference locations in northwestern Florida and southwestern Alabama (Lewis et al. 2002). Average muscle concentrations of PAHs ranged from 0.6 to 3.5 ng/g wet weight (w.w.). The results from this study indicated that total PAH concentrations averaged across fish collected from the wastewater-impacted areas (2.2 ± 3.3 ng/g w.w.) were similar to those collected at the reference sites (1.8 ± 1.6 ng/g w.w.). A regional assessment of whole-body concentrations in fish and shellfish across the west, northeast, southeast, and Gulf coasts were analyzed for chemical contaminants to evaluate compliance with the EPA fish meal recommended guidelines (Harvey et al. 2008). In general, PCBs accounted for the greatest percentage (31%) of exceedances across all regions, followed by mercury (29%), PAHs (21%), and total DDTs (11%). In the Gulf region, mercury accounted for the highest percentage of exceedances (19%), followed by PCBs (11%), total DDTs (7%), PAHs (6%), toxaphene (2%), and cadmium (1%).
In the early 1990s, a study supported by the Minerals Management Service (MMS) evaluated biliary PAHs at several offshore gas rigs in the western GoM (McDonald et al. 1996). This study evaluated the impacts of oil contamination from specific rigs by evaluating fish collected near the rigs and those collected from several kilometers away. Biliary PAHs were evaluated for a variety of bottom-dwelling fishes, generally showing weak negative correlation with distances from the rigs (McDonald et al. 1996).
In the aftermath of Hurricane Katrina in 2005, Hom et al. (2008) evaluated the safety of seafood primarily in Mississippi Sound and offshore waters, examining Atlantic croaker, shrimps, and a few other species. No samples were found to exceed LOCs despite the strong storm surge from that hurricane.
Mesopelagic species were collected in 2007 over the slope of the north central GoM near cold-seep habitats to characterize species composition as well as distributions and determine PAH content in muscle tissues (Ross et al. 2010; Romero et al. 2018). Because of their proximity in time and space, PAH determinations from this dataset represent a serendipitous PAH baseline for DWH-based contamination of mesopelagic fishes in the northern GoM. Pre-spill PAH values ranged from 24 to 555 ng/g w.w., containing a few PAH compounds (mostly naphthalene). These values are relatively high compared to other more shallow-water communities, this trend of organic pollutants accumulating in the deep ocean (Froescheis et al. 2000; Koenig et al. 2013) is a pattern potentially explained by the natural seeps in the study area and likely due to a combination of biological factors intrinsic to mesopelagic communities (e.g., vertical migration, PAH metabolism).
3 Post-DWH and Ixtoc 1 Baselines in Fish
3.1 Seafood Safety
With the release of more than 200 million gallons of crude oil from DWH, there were concerns regarding the integrity and safety of the seafood supply for human consumption, resulting in intensive testing, federal and state fishery closures, risk assessments, and the inclusion of petroleum-related compounds in the 2011 National Listing of Fish Advisories (Ylitalo et al. 2012; US EPA 2013). Between April 28, 2010, and March 31, 2011, more than 8,000 seafood samples were collected and analyzed from within and around closure areas across the GoM for PAHs and the dispersant component dioctyl sodium sulfosuccinate (DOSS). Primarily edible tissues from fish (snappers, groupers, porgies, tuna, etc.) and shellfish contained low concentrations of PAH (<1 ng/g w.w.) and DOSS (0.05–0.29 μg/g w.w.) with both at least two orders of magnitude lower than the LOCs for human health risk (Ylitalo et al. 2012).
An independent assessment of seafood safety evaluated fishes obtained from within fishing closures along the Mississippi coast (Xia et al. 2012). Samples were collected and analyzed weekly from May 27, 2010, until October 2010 and then monthly until August 2011. Eleven fish species consisting of Atlantic croaker, black drum, red drum, cobia (Rachycentron canadum), Gulf menhaden (Brevoortia patronus), red snapper (Lutjanus campechanus), southern flounder (Paralichthys lethostigma), spotted seatrout (Cynoscion nebulosus), striped mullet (Mugil cephalus), tripletail (Lobotes surinamensis), and sand seatrout were analyzed for PAHs. Edible muscle composites were analyzed for 25 PAHs for each species. Overall, all PAHs were at least three orders of magnitude below the LOCs established by NOAA, the FDA, and the Gulf states. LOCs ranged from 35 ng/g for benzo[a]pyrene and dibenzo[a,h]anthracene to 490,000 ng/g w.w. for anthracene and phenanthrene. Maximum concentrations detected in fillets (muscle) ranged from non-detect to 20.1 ng/g for benzo[a]pyrene and anthracene/phenanthrene, respectively. The levels of total PAHs (average, 16 ng/g w.w.) were consistent until January 2011, and thereafter concentrations significantly declined ~44% to an average of 9 ng/g (w.w.).
3.2 Hepatobiliary and Extrahepatic PAH Levels in Fish
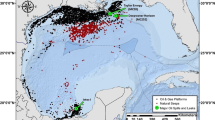
The most comprehensive Gulf-wide survey of the health of GoM fishes conducted to date is that by the Center for Integrated Modeling and Analysis of Gulf Ecosystems (C-IMAGE) wherein ~15,000 fishes were evaluated for disease frequencies and a subset sampled for PAH levels. The C-IMAGE sampling campaign focused on reef and deep-dwelling shelf species (e.g., snappers, tilefishes, groupers) collected from 2011 to 2017 (Fig. 15.1; Murawski et al. 2014, 2018). To date, over 1800 bile samples from 75 offshore species of finfish and sharks have been analyzed for fluorescent aromatic compounds (FACs) in fish bile (Snyder et al., 2015; Pulster et al. 2017a). These samples were collected between 2011 and 2017 from coastal regions from southwest Florida to the Yucatán Peninsula and Cuba. Preliminary biliary PAHs ranged from <1 to 1,900,000 ng/g bile for naphthalene and benzo[a]pyrene FACs (Pulster et al. 2017a). This represents a 90% increase in the maximum total biliary PAHs compared to those measured by the earlier studies conducted in the northern GoM between 1985 and 1991 (Pulster et al. 2017a; NCCOS 2017). Overall for the years 2011–2016, the north central region (39–1,900,000 ng FACs/g bile; 125,000 ± 190,000 ng FAC/g bile) of the GoM remained significantly higher in average total biliary FACs than the southwest region (<1–960,000 ng FAC/g; 73,000 ± 97,000 ng FAC/g bile; p < 0.0001), the west Florida shelf (1,500–360,000 ng FAC/g; 37,000 ± 59,000 ng FAC/g bile; p < 0.0001), and the northwest region (130–370,000 ng FAC/g; 90,000 ± 79,000 ng FAC/g bile; Fig. 15.2; Pulster et al. 2017a). The 2011–2016 preliminary biliary naphthalene and benzo[a]pyrene concentrations ranged from 410 to 1,900,000 ng/g and 0.073 to 9,600 ng/g, respectively (Fig. 15.2). The highest concentrations of biliary naphthalene and benzo[a]pyrene were found in the north central region of the GoM (where DWH occurred). Additional “hot spots” of elevated levels of biliary FACs occurred near major shipping ports (i.e., Brownsville, Corpus Christi, Louisiana, Tampa), vessel shipping lanes, and rivers (i.e., Rio Grande, Mississippi, Coatzacoalcos).

Standardized catch rates (catch in numbers per 1000 hook-hours) of fishes sampled with demersal longlines, 2011–2017 (Murawski et al. 2018)

Mean 2011–2016 biliary naphthalene (Nap ng/g) and benzo[a]pyrene (BaP ng/g) in fish collected in the Gulf of Mexico by the C-IMAGE Consortium (Pulster et al. 2017a). Black dots indicate station locations where fish were collected
Following the 2010 DWH oil spill, skin lesion frequencies and biliary FACs declined in a number of species collected from the north central region of the GoM (where DWH occurred), and this decline provides compelling evidence of the acute and episodic nature of PAH exposures from the DWH oil spill (Murawski et al. 2014; Snyder et al. 2015; Pulster et al. 2017a, 2020) and the impacts such exposures may have on fish health. Lesion frequencies in red snapper (Lutjanus campechanus) declined 53% between 2011 and 2012 (Murawski et al. 2014). In addition, 12 species that were repeatedly collected in the north central region of the GoM also demonstrated declines over time in total biliary PAHs in the years following DWH (2011–2015, Fig. 15.3, Pulster et al. 2017a). Total biliary PAHs declined by 15% in little gulpers (Centrophorus uyato) (2014–2015) to 90% in yellow conger eels (Rhynchoconger flavus) (2011–2015). As of 2015, 5 years after DWH, biliary PAH levels had continued to decline suggesting that levels were not yet reduced back to baseline or pre-spill levels.

Trends (2011–2015) in preliminary mean biliary PAHs (ng FACs/g bile) for multiple fish species collected from the north central Gulf of Mexico. Percent declines in biliary PAHs were calculated between the first and last year sampled for each species. Mean biliary PAHs are the sum of the naphthalene and benzo[a]pyrene fluorescent aromatic compounds (FACs). Error bars are ±1 standard error of the mean
Golden tilefish (Lopholatilus chamaeleonticeps) and a number of other species, including Gulf hake (Urophycis cirrata), snowy grouper (Epinephelus niveatus), yellowedge grouper (Hyporthodus flavolimbatus), red snapper, and red grouper (Epinephelus morio), all exhibited an increase in biliary PAHs between 2012/2013 and 2014 (Struch et al. unpublished; Pulster et al. 2017a, 2018; Snyder et al. 2015). This pattern of increasing PAH levels during the same time period (2012–2014) has also been observed in common loons, seaside sparrows, and sediments (Perez-Umphrey et al. 2018; Paruk et al. 2016; Turner et al. 2014). In Gulf menhaden, body burdens of PAH concentrations decreased, while benzo[a]pyrene toxic equivalents increased between 2012 and 2013 suggesting resuspension of oil residues rather than a new source (Olson et al. 2016). Changes in redox-sensitive elements, radioisotope analyses, and in situ data from the GoM also provide supporting evidence for bioturbation, sedimentation pulses, and resuspension events during a similar time frame (Brooks et al. 2015; Hastings et al. 2016; Diercks et al. 2018). Resuspension events can be caused by a number of physical processes such as variation in subsurface currents, surface and internal waves, and storm-driven surges. An episodic large-scale resuspension event as the result of Hurricane Isaac in 2012 was identified from current meter data and sediment records in the northern GoM (Diercks et al. 2018). This is not surprising considering the GoM experiences frequent tropical storms, for instance, Tropical Storm Debby and Hurricane Isaac in 2012 and Tropical Storm Andrea and Tropical Storm Karen in 2013. The resuspension of sediments reintroduces contaminants into the water column (e.g., freely dissolved or sediment bound), thereby increasing bioavailability to biota. Suspended particles with associated sediment-bound contaminants, including PAHs, enhance the uptake rates and bioaccumulation in fish and other organisms (Zhai et al. 2018; Peterson et al. 1996; Leppänen and Kukkonen 1998; Menon and Menon 1999; Zhang et al. 2015).
Post-DWH, tissues from a number of fish species were also analyzed for PAHs from multiple regions within the GoM as part of the C-IMAGE Gulf-wide comprehensive fish survey (Murawski et al. 2014; Snyder et al. 2015, 2017; Carr et al. 2018; Pulster et al. 2018; Murawski et al. 2018). In 2011, concentrations of muscle and liver PAHs in red snapper collected in the north central region of the GoM were low (<35 ng/g w.w.) compared to the relatively high concentrations of PAH equivalents measured in the bile (41,000–470,000 ng FAC/g bile, Murawski et al. 2014). Despite the ability of fishes to quickly metabolize most PAHs, the PAH composition profiles in the liver samples had a significantly strong positive relationship (r2 = 0.82, p < 0.001) with the profiles of the crude oil from the DWH wellhead (Fig. 15.4, Murawski et al. 2014).

Concentrations of some PAH parent compounds and alkylated homologues sampled from the Macondo (Deepwater Horizon) wellhead (Reddy et al. 2012; gray bars) and 2011 composite samples of red snapper livers collected from the Gulf of Mexico (red bars). (Reprinted with permission from Murawski et al. 2014)
Muscle tissues collected in 2011 from mesopelagic fish species in the north central region of the GoM contained PAH levels (330–2,350 ng/g w.w.; Romero et al. 2018) more than an order of magnitude higher than other species during the same time period (Murawski et al. 2014; Romero et al. 2018; Ylitalo et al. 2012; Xia et al. 2012). Similar to other species, the muscle concentrations in mesopelagic fish demonstrated significant increases between pre- and post-DWH levels, followed by a significant decline by 2015 (Romero et al. 2018). Romero et al. suggest the higher tissue concentrations in mesopelagic fish than shallower water species may be due to the increased frequency and duration of exposure periods due to their vertical migratory behaviors throughout the water column (Romero et al. 2018). However, in addition to species-specific differences in metabolic capacity, a number of additional critical environmental and physiochemical factors may partially explain the increased concentrations observed in mesopelagic fish. First, the fish in this study were collected (50–1,000 m) within similar depths of the continuous subsurface plume of oil residues (~1,000–1,200 m) that was identified post-DWH (Camilli et al. 2010). Although there is evidence that chemical cues can elicit habitat avoidance in fish (Pulster et al. 2020), the potential for increased and extended periods of exposure in 2010 exists due to the subsurface plume which persisted for months and continued water column contamination persisted in 2010, likely due to resuspended sediments. Furthermore, water temperatures decrease significantly with depth and are 15–20° lower at 1,000 m than at the surface, increasing the persistence and bioavailability of contaminants at deep pelagic depths. Specifically to PAHs, low water temperatures promote their persistence in water due to lower volatilization and biodegradation which consequently decreases elimination rates further resulting in the retention of PAHs in fish tissues (Varanasi et al. 1981; Egaas and Varanasi 1982; Varanasi 1989). For example, starry flounders (Platichthys stellatus) held at temperatures (4 °C) similar to the temperatures at the depth of DWH in the GoM were found to have 26–34 times higher naphthalene concentrations than those held at 12 °C (Varanasi et al. 1981). The preliminary total PAHs in liver tissues collected ranged from 7.7 to 407 ng/g w.w. for hakes (Urophycis sp., 2012–2015; Struch et al. unpublished), 1200 to 195,000 ng/g w.w. for golden tilefish (2012–2016; Snyder et al. 2017, 2018), and 67.6 to 17,300 ng/g w.w. for groupers (Epinephelus sp., 2012–2016; Pulster et al. 2018). While there were a few species and site differences, in general the groupers collected within the north central region (2,350 ± 2,800 ng/g w.w.) had similar levels of total PAHs to those collected in the northwest region (1580 ± 2470 ng/g w.w.) yet significantly higher than those collected in the southwest region (983 ± 2,260 ng/g w.w.; p < 0.0001). In contrast to biliary PAHs, there appears to be an increasing trend over time in the liver concentrations of PAHs for a number of species. For instance, between 2012 and 2015, liver concentrations in yellowedge groupers from the north central region increased to 362% (p = 0.036) (Fig. 15.5). Similar trends were also observed in the southwest region for both snowy (237% increase) and yellowedge groupers (177% increase) from 2015 to 2016. In addition, PAH levels increased to 102% in hake livers between 2012 and 2015.

Trends in preliminary mean liver concentrations of PAHs (ng/g wet weight) in grouper species collected from the southwest (blue bars), north central (red bars), and northwest (green bars) regions of the Gulf of Mexico (2011–2016). Percent increases in liver PAHs were calculated between the first and last year sampled for each species. The ∑PAHs is the sum of 46 parental PAHs and their alkylated homologues. Error bars are ±1 standard error of the mean
Preliminary concentrations of total PAHs in liver tissues from golden tilefish sampled between 2015 and 2016 were similar among regions but significantly higher in the northwest region offshore of Texas (31,000 ± 25,000 ng/g w.w.) than the southwest (21,000 ± 28,000 ng/g w.w.) and north central (16,000 ± 11,000 ng/g w.w.) regions (Snyder et al. 2017, 2018). The opposite pattern occurred for biliary PAHs. Biliary PAHs are higher in the north central region (470,000 ± 400,000 ng/g) than the southwest (170,000 ± 140,000 ng/g) and northwest regions (230,000 ± 88,000 ng/g). These results suggest the highest recent exposure to PAHs in the north central region, perhaps more efficient metabolism and elimination, and higher accumulation in the liver tissue in the southwest and northwest regions. As mentioned above, many species, including golden tilefish from the north central GoM, demonstrated increased biliary naphthalene metabolite concentrations between 2012 and 2015, possibly related to the resuspension of sedimented oil as discussed above. In contrast, total liver PAHs have remained relatively constant for golden tilefish sampled in 2012–2015.
In the southwest region, hardhead catfish collected in 2008 from along the Tabasco coast in Mexico had liver concentrations of PAHs ranging from 7,340 to 119,000 ng/g w.w., 1 year after the Kab-121 oil spill and 28 years post-Ixtoc 1 (Gold-Bouchot et al. 2014). Liver concentrations significantly declined to 1390–9470 ng/g w.w. within 3 months of the first sampling event. In 2010, Gracia (2010) analyzed the muscle content from 29 fish species collected at 30 sampling locations in the southwestern GoM near Ixtoc I spill area (off Veracruz, Tabasco, and Campeche states, Gracia 2010). The most frequent fish species sampled included hardhead catfish, Mexican flounder, sheepshead (Archosargus probatocephalus), gray triggerfish (Balistes capriscus), Atlantic croaker, and southern kingfish (Menticirrhus americanus). The highest value of PAH muscle concentrations (474 ng/g d.w.) was found in goliath grouper (Epinephelus itajara); however, relatively high concentrations of PAHs (100–474 ng/g d.w.) were also found in spotted weakfish (Cynoscion nebulosus), sand seatrout, sheepshead (Archosargus probatocephalus), and goliath grouper which were mainly found in coastal areas (Fig. 15.6; Gracia 2010). Anthracene (99 ng/g d.w.), fluoranthene (100 ng/g d.w.), and phenanthrene (98 ng/g d.w.) were the dominant PAHs in tissue content.

Mean concentrations of PAHs (μg/g d.w.) in muscle tissues from fish (29 species) collected in 2010 from the southern Gulf of Mexico near oil infrastructure within the Ixtoc 1 zone
Fish surveys conducted in 2015 and 2016 by C-IMAGE in the southern GoM evaluated muscle tissues for PAH concentrations in 107 organisms from 16 species. The muscle concentrations of PAHs in red snapper and yellowedge grouper ranged from 10 to 493 ng/g d.w. (Gracia et al. 2018). Red snapper (59–83.4 ng/g d.w.) had higher mean muscle PAH concentrations than yellowedge grouper (53–37 ng/g d.w.), although not significantly so (Fig. 15.7). Muscle concentrations of PAHs in red snapper collected within the exclusion zone near where the Ixtoc 1 wellhead was located were relatively high; however, the highest levels were located off the Yucatán Peninsula where there are no known oil rigs or natural oil seeps (Fig. 15.8). The high levels of PAHs observed in multiple species in this location may be the result of proximity to major shipping channels and strong current systems (Snyder et al. 2017; Gracia et al. 2018). Yellowedge grouper showed high PAH muscle values in three main areas; however, similar to the red snapper, the concentrations off the Yucatán Peninsula were higher than those samples near the exclusion zone (Fig. 15.9; Gracia et al. 2018). However, the mean PAH muscle concentration of both species was very low with respect to US LOCs.

Muscle concentrations of PAHs (μg/kg d.w.) measured in red snapper and yellowedge grouper collected in the southwestern Gulf of Mexico, 2015–2016

Muscle concentrations of PAHs (μg/kg d.w.) in red snapper collected in the southwestern Gulf of Mexico, 2015–2016

Distributions of muscle concentrations of PAHs (μg/kg d.w.) in yellowedge grouper collected in the southwestern Gulf of Mexico, 2015–2016
4 Conclusions
A complex interaction of physiochemical properties (e.g., molecular weight and octanol-water partition coefficients), species-specific life histories (e.g., habitat, diet), metabolic capacities, and environmental factors (e.g., temperature, salinity) regulates the chemical bioavailability, uptake, metabolic rates, and excretion rates of oil contaminant dynamics in fishes. Although fish have a relatively high capacity to rapidly metabolize PAHs, there is a potential for increased toxicity and bioaccumulation during the metabolic process as PAHs are hydrolyzed, reabsorbed, and returned through circulation (Lech and Vodicnik 1985; Kleinow et al. 1987; Varanasi et al. 1989).
Studies have demonstrated that exposure routes have little effect on the biotransformation products of some compounds and the distribution of PAHs within tissues (Stein et al. 1984; Varanasi 1989; Pulster et al. 2017b). Rather, differences observed between muscle and liver concentrations are more likely due to chronic exposures and accumulation rates that are faster than the fishes’ ability to metabolize and excrete xenobiotics. Importantly, chronically exposed fish have slower elimination and increased PAH levels than those exposed acutely (Varanasi 1989). This is evident in the increasing trend in baseline levels of biliary and tissue concentrations of PAHs in a number of fish species between pre-2000 and post-DWH data in the GoM that raises concern for the overall health of the Gulf ecosystem and its fish populations. The increasing oil production (especially in the northern GoM) and the higher fraction of oil derived from ultra-deep waters (>1500 m) mean that fish communities not traditionally exposed to oil contamination from oil extraction activities might now be at a higher risk (Romero et al. 2018). The increasing trend in oil accumulation in deepwater fishes and expansion of drilling into deeper waters warrants increased monitoring and fish health assessments and intensifies the need for more extensive and comprehensive offshore baseline data, since the majority of all baseline studies in the GoM prior to the DWH were for nearshore and shelf fishes.
Given the importance of the GoM to energy production in both the USA and Mexico (Murawski et al. 2020), the lack of coordinated, comprehensive, and continuous monitoring of oil-related pollution in sediments, fishes, and waters from these facilities is remarkable. Other than the post-DWH assessment activities, primarily supported by private foundation funding, there is no specific program in the USA to understand and assess pollution related to oil production and transportation activities. The advantages of developing such a program are numerous. Assessing the impacts of pollution from a specific infrastructure failure is the obvious primary advantage, but such data can also identify the interactions between pollution from natural sources (e.g., cold seeps) in relation to the entirety of the marine oil budget.
Pre-Ixtoc 1 data from Mexican waters are virtually nonexistent, and there is at least a 10-year data gap between the pre- and post-DWH data for PAH concentrations in fish from US waters. Given that the GoM is a highly dynamic system with numerous and expanding sources of anthropogenically derived pollution, collection of baseline data at shorter time intervals is critical for assessing and evaluating sources of pollution. Baseline data and long-term trends are essential to monitor the health of the GoM, changing environmental conditions, and the interactions of natural systems with anthropogenic activities. In the context of a future oil spill and changing environmental conditions in the GoM, continued monitoring is required for the assessment of impact, resilience, and recovery from future events.
References
Amezcua-Linares F, Amezcua F, Gil-Manrique B (2015) Effects of the Ixtoc 1 oil spill on fish assemblages in the Southern Gulf of Mexico. In: Alford JB, Peterson MS, Green CC (eds) Impacts of Oil Spill disasters on marine habitats and fisheries in North America. CRC Press, Boca Raton, pp 209–236
Brooks GR, Larson RA, Schwing PT, Romero I, Moore C, Reichart G-J, Jilbert T, Chanton JP, Hastings DW, Overholt WA, Marks KP, Kostka JE, Holmes CW, Hollander D (2015) Sedimentation pulse in the NE Gulf of Mexico following the 2010 DWH Blowout. PLoS One 10(7):e0132341. https://doi.org/10.1371/journal.pone.0132341
Camilli R, Reddy CM, Yoerger DR, Van Mooy BAS, Jakuba MV, Kinsey JC, McIntyre CP, Sylva SP, Maloney JV (2010) Tracking hydrocarbon plume transport and biodegradation at Deepwater Horizon. Science 330:201–204. https://doi.org/10.1126/science.1195223
Carr BE, Pulster EL, Gracia A, Armenteros M, Murawski SA (2018) A spatiotemporal analysis of hepatic and biliary PAHs in groupers from around the Gulf of Mexico. In: Gulf of Mexico Oil Spill and ecosystem science, New Orleans
Diercks AR, Dike C, Asper VL, DiMarco SF, Chanton JP, Passow U (2018) Scales of seafloor sediment resuspension in the northern Gulf of Mexico. Elementa Sci Anthropocene 6. https://doi.org/10.1525/elementa.285
Egaas E, Varanasi U (1982) Effects of polychlorinated biphenyls and environmental temperature on in vitro formation of benzo[a]pyrene metabolites by liver of trout (Salmo gairdneri). Biochem Pharmacol 31:561–566. https://doi.org/10.1016/0006-2952(82)90160-5
Farrington JW, Tripp BW, Tanabe S, Subramanian A, Sericano JL, Wade TL, Knap AH (2016) Edward D. Goldberg’s proposal of “the Mussel Watch”: reflections after 40 years. Mar Pollut Bull 110:501–510
Froescheis O, Looser R, Cailliet GM, Jarman WM, Ballschmiter K (2000) The deep-sea as a final global sink of semivolatile persistent organic pollutants? Part I: PCBs in surface and deep-sea dwelling fish of the North and South Atlantic and the Monterey Bay Canyon (California). Chemosphere 40:651–660. https://doi.org/10.1016/S0045-6535(99)00461-0
Gold-Bouchot G, Ceja-Moreno V, Chan-Cocom E, Zapata-Perez O (2014) Petroleum hydrocarbons, fluorescent aromatic compounds in fish bile and organochlorine pesticides from areas surrounding the spill of the Kab121 well, in the Southern Gulf of Mexico: a case study. J Environ Biol 35:147–156
Gracia A (2010) Campaña Oceanográfica (SGM-2010). Informe Final. Gerencia de Seguridad Industrial, Protección Ambiental y Calidad Región Marina Noreste, PEMEX - EXPLORACIÓN – PRODUCCIÓN. Instituto de Ciencias del Mar y Limnología, UNAM, México
Gracia A, Murawski SA, Alexander-Valdés RM, Vázquez-Bader AR, Snyder S, LópezDurán IM, Ortega-Tenorio P, Pulster EL, Frausto-Castillo JA (2018) Fish stock resiliency to environmental PAH. Presentation at: Gulf of Mexico Oil Spill & ecosystem science conference, New Orleans, LA, February 6–9 2018
Harvey J, Harwell L, Summers JK (2008) Contaminant concentrations in whole-body fish and shellfish from US estuaries. Environ Monit Assess 137:403–412. https://doi.org/10.1007/s10661-007-9776-1
Hastings DW, Schwing PT, Brooks GR, Larson RA, Morford JL, Roeder T, Quinn KA, Bartlett T, Romero IC, Hollander DJ (2016) Changes in sediment redox conditions following the BP DWH blowout event. Deep Sea Res II Top Stud Oceanogr 129:167–178. https://doi.org/10.1016/j.dsr2.2014.12.009
Hom T, Collier TK, Krahn MM, Strom MS, Ylitalo GM, Nilsson WB, Paranjpye RN, Varanasi U (2008) Assessing seafood safety in the aftermath of Hurricane Katrina. Am Fish Soc Symp 64:73–93
Kennicutt MC, Brooks JM, Atlas EL, Giam CS (1988) Organic compounds of environmental concern in the Gulf of Mexico: a review. Aquat Toxicol 11:191–212. https://doi.org/10.1016/0166-445X(88)90013-6
Kleinow KM, Melancon MJ, Lech JJ (1987) Biotransformation and induction: implications for toxicity, bioaccumulation and monitoring of environmental xenobiotics in fish. Environ Health Perspect 71:105–119. https://doi.org/10.2307/3430418
Koenig S, Fernández P, Company JB, Huertas D, Solé M (2013) Are deep-sea organisms dwelling within a submarine canyon more at risk from anthropogenic contamination than those from the adjacent open slope? a case study of Blanes canyon (NW Mediterranean). Prog Oceanogr 118:249–259. https://doi.org/10.1016/j.pocean.2013.07.016
Lech JJ, Vodicnik MJ (1985) Biotransformation. In: Rand GM, Petrocelli SR (eds) Fundamentals of aquatic toxicology: methods and applications. Hemisphere, New York, pp 526–557
Leppänen MT, Kukkonen JVK (1998) Relative importance of ingested sediment and pore water as bioaccumulation routes for pyrene to oligochaete (Lumbriculus variegatus, Müller). Environ Sci Technol 32:1503–1508. https://doi.org/10.1021/es970941k
Lewis MA, Scott GI, Bearden DW, Quarles RL, Moore J, Strozier ED, Sivertsen SK, Dias AR, Sanders M (2002) Fish tissue quality in near-coastal areas of the Gulf of Mexico receiving point source discharges. Sci Total Environ 284:249–261. https://doi.org/10.1016/S0048-9697(01)00891-9
McCain BB, Brown DW, Hom T, Myers MS, Pierce SM, Collier TK, Stein JE, Chan SL, Varanasi U (1996) Chemical contaminant exposure and effects in four fish species from Tampa Bay, Florida. Estuaries 19:86–104. https://doi.org/10.2307/1352655
McDonald SJ, Willett KL, Thomsen J, Beatty KB, Connor K, Narasimhan TR, Erickson CM, Safe SH (1996) Sublethal detoxification responses to contaminant exposure associated with offshore oil production. Can J Fish Aquat Sci 53:2606–2617
Menon NN, Menon NR (1999) Uptake of polycyclic aromatic hydrocarbons from suspended oil borne sediments by the marine bivalve Sunetta scripta. Aquat Toxicol 45:63–69. https://doi.org/10.1016/s0166-445x(98)00092-7
Murawski SA, Hogarth WT (2013) Enhancing the ocean observing system to meet restoration challenges in the Gulf of Mexico. Oceanography 26:10–16
Murawski SA, Hogarth WT, Peebles EB, Barbieri L (2014) Prevalence of external skin lesions and polycyclic aromatic hydrocarbon concentrations in Gulf of Mexico Fishes, post-Deepwater Horizon. Trans Am Fish Soc 143:1084–1097. https://doi.org/10.1080/00028487.2014.911205
Murawski SA, Peebles EB, Gracia A, Tunnell JW, Armenteros M (2018) Comparative abundance, species composition, and demographics of continental shelf fish assemblages throughout the Gulf of Mexico. Mar Coast Fish 10:325–346. https://doi.org/10.1002/mcf2.10033
Murawski SA, Hollander D, Gilbert S, Gracia A (2020) Deep-water oil and gas production in the Gulf of Mexico, and related global trends (Chap. 2). In: Murawski SA, Ainsworth C, Gilbert S, Hollander D, Paris CB, Schlüter M, Wetzel D (eds) Scenarios and responses to future Deep Oil Spills – fighting the next war. Springer, Cham
NAS (2013) An ecosystem services approach to assessing the impacts of the Deepwater Horizon oil spill in the Gulf of Mexico. National Academies Press, Washington, DC
NCCOS (2017) NOAA’s National status and trends data. National Centers for Coastal Ocean Science. 2018
Olson GM, Meyer BM, Portier RJ (2016) Assessment of the toxic potential of polycyclic aromatic hydrocarbons (PAHs) affecting Gulf Menhaden (Brevoortia patronus) harvested from waters impacted by the BP Deepwater Horizon Spill. Chemosphere 145:322–328. https://doi.org/10.1016/j.chemosphere.2015.11.087
Paruk JD, Adams EM, Uher-Koch H, Kovach KA, Long D, Perkins C, Schoch N, Evers DC (2016) Polycyclic aromatic hydrocarbons in blood related to lower body mass in common loons. Sci Total Environ 565:360–368. https://doi.org/10.1016/j.scitotenv.2016.04.150
Perez-Umphrey AA, Bergeon Burns CM, Stouffer PC, Woltmann S, Taylor SS (2018) Polycyclic aromatic hydrocarbon exposure in seaside sparrows (Ammodramus maritimus) following the 2010 Deepwater Horizon oil spill. Sci Total Environ 630:1086–1094. https://doi.org/10.1016/j.scitotenv.2018.02.281
Peterson CH, Kennicutt MC II, Green RH, Montagna P, Harper DE Jr, Powell EN, Roscigno PF (1996) Ecological consequences of environmental perturbations associated with offshore hydrocarbon production: a perspective on long-term exposure in the Gulf of Mexico. Can J Fish Aquat Sci 53:2637–2654
Pulster EL, Snyder S, Struch R, Carr B, Toro-Farmer GA, Gracia A, Murawski SA (2017a) What can bile tell us about the environmental health of the Gulf of Mexico? Presentation at: Gulf of Mexico Oil Spill and Ecosystem Science, New Orleans, LA
Pulster EL, Main K, Wetzel D, Murawski S (2017b) Species-specific metabolism of naphthalene and phenanthrene in 3 species of marine teleosts exposed to Deepwater Horizon crude oil. Environ Toxicol Chem 36:3168–3176. https://doi.org/10.1002/etc.3898
Pulster EL, Carr B, Murawski SA (2018, October) Spatiotemporal trends in hepatobiliary levels of polycyclic aromatic hydrocarbons in Gulf of Mexico groupers. University of South Florida, Presentation at: MARCUBA 2018, Havana Cuba
Pulster EL, Gracia A, Snyder SM, Deak K, Fogleson S, Murawski SA (2020) Chronic sublethal effects observed in wild caught fish following two major oil spills in the Gulf of Mexico: Deepwater Horizon and Ixtoc 1 (Chap. 24). In: Murawski SA, Ainsworth C, Gilbert S, Hollander D, Paris CB, Schlüter M, Wetzel D (eds) Deep Oil Spills: facts, fate and effects. Springer International, Cham
Reddy CM, Arey JS, Seewald JS, Sylva SP, Lemkau KL, Nelson RK, Carmichael CA, McIntyre CP, Fenwick J, Ventura GT, Van Mooy BAS, Camilli R (2012) Composition and fate of gas and oil released to the water column during the Deepwater Horizon oil spill. Proc Natl Acad Sci 109(50):20229–20234
Romero IC, Sutton T, Carr B, Quintana-Rizzo E, Ross SW, Hollander DJ, Torres JJ (2018) Decadal assessment of polycyclic aromatic hydrocarbons in mesopelagic fishes from the Gulf of Mexico reveals Exposure to oil-derived Sources. Environ Sci Technol. https://doi.org/10.1021/acs.est.8b02243
Ross SW, Quattrini AM, Roa-Varón AY, McClain JP (2010) Species composition and distributions of mesopelagic fishes over the slope of the north-central Gulf of Mexico. Deep Sea Res II Top Stud Oceanogr 57:1926–1956. https://doi.org/10.1016/j.dsr2.2010.05.008
Shigenaka G (2014) Twenty-five years after the Exxon Valdez Oil Spill: NOAA's Scientific Support, Monitoring, and Research. NOAA Office of Response and Restoration, Seattle
Snyder S, Pulster EL, Murawski SA (2017) Gulf-wide analysis of PAH exposure and accumulation in Golden Tilefish. Presented at: American Fisheries Society Annual Meeting, Tampa, FL
Snyder SM, Pulster EL, Fogelson SB (2018) Murawski SA Hepatic accumulation of PAHs and prevalence of hepatic lesions in Golden Tilefish from the northern Gulf of Mexico. Presentation at: Gulf of Mexico Oil Spill & Ecosystem Science Conference, New Orleans
Snyder SM, Pulster EL, Wetzel DL, Murawski SA (2015) PAH exposure in Gulf of Mexico demersal fishes, post-Deepwater Horizon. Environ Sci Technol 49:8786–8795. https://doi.org/10.1021/acs.est.5b01870
Stein JE, Hom T, Varanasi U (1984) Simultaneous exposure of English sole (Parophrys vetulus) to sediment-associated xenobiotics: part 1—uptake and disposition of 14C-polychlorinated biphenyls and 3H-benzo[a]pyrene. Mar Environ Res 13:97–119. https://doi.org/10.1016/0141-1136(84)90021-7
Turner RE, Overton EB, Meyer BM, Miles MS, McClenachan G, Hooper-Bui L, Engel AS, Swenson EM, Lee JM, Milan CS, Gao H (2014) Distribution and recovery trajectory of Macondo (Mississippi Canyon 252) oil in Louisiana coastal wetlands. Mar Pollut Bull 87:57–67. https://doi.org/10.1016/j.marpolbul.2014.08.011
US EPA (2009) 2008 Biennial National Listing of Fish Advisories. Washington, D.C.
US EPA (2013) 2011 National Listing of Fish Advisories. United States Environmental Protection Agency, Washington, D.C.
Varanasi U (ed) (1989) Metabolism of polycyclic aromatic hydrocarbons in the aquatic environment. CRC Press, Boca Raton
Varanasi U, Gmur DJ, Reichert WL (1981) Effect of environmental temperature on naphthalene metabolism by Juvenile Starry flounder (Platichthys stellatus). Arch Environ Contam Toxicol 10:203–214. https://doi.org/10.1007/bf01055622
Varanasi U, Stein JE, Nishimoto M (1989) Biotransformation and disposition of polycyclic aromatic hydrocarbons (PAH) in fish. In: Varanasi U (ed) Metabolism of polycyclic aromatic hydrocarbons in the aquatic environment. CRC Press, Boca Raton, pp 94–140
Ward CH, Tunnell JW Jr (2017) Habitats and biota of the Gulf of Mexico: An overview. In: Ward CH (ed) Habitats and Biota of the Gulf of Mexico: before the Deepwater Horizon Oil Spill, vol I. Springer Nature, New York
Xia K, Hagood G, Childers C, Atkins J, Rogers B, Ware L, Armbrust K, Jewell J, Diaz D, Gatian N, Folmer H (2012) Polycyclic aromatic hydrocarbons (PAHs) in Mississippi seafood from areas affected by the Deepwater Horizon oil spill. Environ Sci Technol 46:5310–5318. https://doi.org/10.1021/es2042433
Ylitalo GM, Krahn MM, Dickhoff WW, Stein JE, Walker CC, Lassitter CL, Garrett ES, Desfosse LL, Mitchell KM, Noble BT, Wilson S, Beck NB, Benner RA, Koufopoulos PN, Dickey RW (2012) Federal seafood safety response to the Deepwater Horizon oil spill. Proc Nat Acad Sci USA 109:20274–20279. https://doi.org/10.1073/pnas.1108886109
Zhai Y, Xia X, Xiong X, Xia L, Guo X, Gan J (2018) Role of fluoranthene and pyrene associated with suspended particles in their bioaccumulation by zebrafish (Danio rerio). Ecotoxicol Environ Saf 157:89–94. https://doi.org/10.1016/j.ecoenv.2018.03.065
Zhang X, Xia X, Li H, Zhu B, Dong J (2015) Bioavailability of pyrene associated with suspended sediment of different grain sizes to Daphnia magna as investigated by passive dosing devices. Environ Sci Technol 49:10127–10135. https://doi.org/10.1021/acs.est.5b02045
Acknowledgments
This research was made possible by a grant from the Gulf of Mexico Research Initiative/C-IMAGE I, II, and III. Preliminary data will be publicly available once validated and finalized. Final data are publicly available through the Gulf of Mexico Research Initiative Information and Data Cooperative (GRIIDC) at https://data.gulfresearchinitiative.org (R6.x805.000:0048, R6.x805.000:0052, R6.x805.000:0049, R6.x805.000:0051).
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2020 Springer Nature Switzerland AG
About this chapter

Cite this chapter
Pulster, E.L. et al. (2020). Polycyclic Aromatic Hydrocarbon Baselines in Gulf of Mexico Fishes. In: Murawski, S., et al. Scenarios and Responses to Future Deep Oil Spills. Springer, Cham. https://doi.org/10.1007/978-3-030-12963-7_15
Download citation
DOI: https://doi.org/10.1007/978-3-030-12963-7_15
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-12962-0
Online ISBN: 978-3-030-12963-7
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)