
Abstract
The Littoral Acoustic Demonstration Center (LADC) was formed in early 2001 to utilize Environmental Acoustic Recording System (EARS) buoys developed by the Naval Oceanographic Office (NAVOCEANO) which has provided technical guidance and support to LADC. The purpose of LADC is to make environmental measurements, which is not part of the mission of NAVOCEANO. This chapter describes the Gulf of Mexico marine mammal measurements and related data analysis of LADC. LADC has also used the buoys to characterize the three-dimensional acoustic field of a seismic airgun array and to analyze the noise due to nearby storms. LADC is a consortium of scientists from universities and the U.S. Navy. The following institutions are or have been represented: initially, the University of New Orleans, the University of Southern Mississippi, and the Naval Research Laboratory-Stennis Space Center; and then the University of Louisiana at Lafayette, the Applied Research Laboratories at the University of Texas at Austin, and Oregon State University. The scientists are listed in the first section of the chapter. A technical overview of EARS technology is given in Sect. 6.2. The current Generation 2 EARS buoys can record four channels of up to 25 kHz each or one channel up to 96 kHz.
LADC has conducted marine mammal experiments in the Gulf of Mexico in 2001, 2002, 2007, and 2010. The 2007 experiments were at sites 9 and 23 miles from the Macondo Well Oil Spill. These sites as well as the 2001 and 2002 sites were recorded in the 2010 experiment to measure changes related to earlier measurements. LADC has also done seismic airgun array experiments in 2003 and 2007. The marine mammal experiments are summarized in Sect. 3, where experiments in the Mediterranean Sea, which had LADC participation, are also listed.
The remaining Sects. 6.4 through 6.11 describe the analysis to date of LADC data and also the analysis by LADC scientists of workshop data for detection, classification, and localization purposes. Section 6.4 describes sperm whale click structure analysis for click-train demarcation and identification of individual whales. The tendency of whales diving together to establish different cadences for their echolocation clicks to keep from interfering with each other is presented in Sect. 6.5. The identification of individual whales by clustering echolocation clicks of sperm and beaked whales and coda clicks of sperm whales is discussed in Sect. 6.6. The application of click change detection to know if the same or a different whale are speaking on successive clicks is described in Sect. 6.7. This method allows one to follow a turning sperm whale. A technique for localizing individual clicking whales is presented in Sect. 6.8. The integration of the above techniques is discussed in Sect. 6.9, which also suggests how whales might identify each other. Sperm whale coda classification and repertoire analysis are the subject of Sect. 6.10. Finally, statistical modeling for population estimation is given in Sect. 6.11.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
6.1 Littoral Acoustic Demonstration Center (LADC)
The Littoral Acoustic Demonstration Center was formed in early 2001 to utilize U. S. Navy technology to advance the science of underwater acoustics. Specifically, the technology is the autonomous Environmental Acoustic Recording System (EARS) buoy, developed by the Naval Oceanographic Office (NAVOCEANO) at Stennis Space Center , MS. EARS are described in Sect. 6.2 of this chapter. It was decided that LADC would be a consortium of scientists from Gulf State Universities and the U. S. Navy.
Mr. Craig Peterson represented the Naval Oceanographic Office , which supplied technical support and guidance. The cooperating scientists included the following, almost all of whom are still participants: Dr. Grayson Rayborn of the Department of Physics and Astronomy (and later also of the Signal Research Center) of the University of Southern Mississippi (USM), Dr. Stan Kuczaj of the Department of Psychology of USM, Dr. George Ioup and Dr. Juliette Ioup of the Department of Physics of the University of New Orleans (UNO), Dr. Joal Newcomb of the Naval Research Laboratory at Stennis Space Center (NRL -Stennis) (now at NAVOCEANO ) and Mr. Robert Field of NRL-Stennis (now retired). Later these scientists were joined by Dr. Natalia Sidorovskaia of the Physics Department at the University of Louisiana at Lafayette (UL Lafayette), Dr. Christopher Tiemann of the Applied Research Laboratories of the University of Texas at Austin (ARL-UT) (now at R2Sonic), Dr. James Stephens of the Department of Physics and Astronomy and the Signal Research Center of USM (now at the Southwest Mississippi Community College), Dr. Arslan Tashmukhambetov of the Department of Physics at UNO (later at G Geophysics and now at LLOG), and Dr. Azmy Ackleh of the Department of Mathematics at UL Lafayette. Recently Dr. David Mellinger and Ms. Sara Heimlich of the Cooperative Institute for Marine Resources Studies at Oregon State University and Adm. Ken Barbor and Dr. Danielle Greenhow of the Department of Marine Science of USM have joined the group. These scientists have joined over the years for specific exercises and for equipment development and maintenance by many other scientists and engineers, technicians, graduate students, and crew members, to whom an immense debt of gratitude is owed. LADC ’s development is described in the remainder of this section. EARS technology is elucidated in Sect. 6.2 and LADC Marine Mammal Experiments are described in Sect. 6.3. The remaining Sects. 6.4 through 6.12 contain the results of data analysis.
The initial scientific purpose of the group was to study ocean ambient noise and upslope propagation. When it was learned in early 2001 that the Minerals Management Service (MMS) , with other sponsors, intended to have an exercise conducted in the Northern Gulf of Mexico (GoM) in the summer of 2001 to study endangered sperm whale s, LADC added marine mammal bioacoustics to its areas of interest and determined to deploy EARS moorings that summer to record sperm whales as well as ambient noise and upslope propagation. An area was chosen that had a very large number of sperm whale sightings , and EARS technology was extended to higher frequencies (~6000 Hz) by NAVOCEANO to record the whales. (Originally EARS was developed to measure ambient noise, and its upper frequency limit was 1000 Hz.) Some of the highest quality recordings of sperm whales up to that time were made. The deployment was repeated in the summer of 2002. These exercises are referred to as LADC01 and LADC02 and were supported by ONR.
When LADC learned that MMS intended to conduct a controlled exposure experiment (CEE) for sperm whale s in the Northern GoM in 2003, using a seismic source vessel supplied by the Industry Research Funders Coalition (IRFC) through the International Association of Geophysical Contractors (IAGC), it approached the IRFC through the IAGC for funding to measure the acoustic field of the seismic airgun array , adding an additional LADC research interest. LADC deployed a co-located sensitive hydrophone buoy and a desensitized hydrophone buoy (to avoid clipping), and the plan was to have the source vessel run lines over this mooring if its schedule permitted. On the last afternoon of its availability, the vessel had time to shoot five parallel lines over the mooring. This experiment, called Airgun03, is described and the data are analyzed and modeled in “Three-dimensional seismic array characterization study: Experiment and modeling,” Arslan M. Tashmukhambetov et al. (2008).
EARS technology was extended to measure up to 25 kHz for the airgun experiment. Because of increasing U. S. Navy interest in beaked whale s, the technology was further developed so that EARS buoys could measure up to 96 kHz in single-channel mode or 25 kHz per channel in four-channel mode. These are Generation 2 buoys. In 2007, LADC conducted the Source Characterization Study 2007 (SCS07 ) to completely characterize the three-dimensional (3-D) primary acoustic field of a seismic array , using Generation 2 EARS buoys in the four-channel mode. These were deployed as 20 pairs of moored hydrophones (one each, sensitive and desensitized) at different depths. Four pairs of hydrophones were suspended from a ship. This work was sponsored by the Joint Industry Programme (JIP) through the International Association of Oil and Gas Producers (OGP). Its findings will impact regulations for the protection of marine mammals and other marine life.
Also in 2007, LADC conducted an exercise, sponsored by SPAWAR, to record primarily not only beaked whale s but also sperm whale s and dolphins in the Northern GoM . The Generation 2 EARS buoys were used in the single-channel mode. It made the first recordings of beaked whales in the GoM. By coincidence, the two sites selected for deployment , based on a high density of beaked whale sightings, are 9 and 23 miles from the location of the BP oil spill in 2010. Therefore, LADC received funding in 2010 from NSF and Greenpeace to go back to the two sites and to LADC01 and LADC02 sites to measure marine mammal activity after the spill and compare it to the measurements made in LADC07 and earlier. Some abundance analysis results appear in Ackleh et al. (2012) and in this Chapter. More recently, LADC has received 3 years of funding (2015 through 2017) from the Gulf of Mexico Research Initiative (GoMRI) RFP IV to conduct two exercises to compare sperm and beaked whales and dolphin measurements using moored buoys , autonomous surface vehicles , and gliders .
All of the existing marine mammal recordings are still being analyzed. Many of our results are summarized in the following sections of this Chapter.
6.2 Environmental Acoustic Recording System (EARS) Buoys
Environmental Acoustic Recording System (EARS) buoys were developed as autonomous moored underwater recording systems by the Naval Oceanographic Office (NAVOCEANO) to make long-term ocean ambient noise measurements. When LADC was formed, the buoys were capable of measuring up to 1000 Hz for 1 year. When LADC added listening to sperm whale s to its noise and propagation measurement missions, NAVOCEANO quickly modified the buoys to measure up to 5859 Hz for 36 days. The buoys, moored at depths from 550 to 950 m in the Gulf of Mexico, produced exceptionally clear recordings of sperm whale echolocation and coda clicks and recordings of other whales. EARS Generation 2 buoys are now capable of recording one channel to 96 kHz, or 4 channels to 25 kHz. All buoy designs include high quality omnidirectional hydrophones (Figs. 6.1 and 6.2).

Components of an EARS mooring. These include flotation, the EARS buoys, dual acoustic releases, an anchor chain, and an anchor. Aside from the chain the remaining components are connected by faired cable, to minimize cable strum

EARS mooring components on deck, before deployment . The floats are yellow. The faired cable is white. The EARS buoys are black cylinders
Overview: (Environmental Acoustic Recording System (EARS) BUOY
-
Developed by NAVOCEANO
-
Autonomous Self-recording Buoy
-
Small, Easily Deployable Package
-
16-bit Sigma-Delta A/D
-
First-Generation Attributes (at peak of development)
-
1–4 channels (limited spatial separation)
-
Sampling rates from 200 Hz to 25 kHz
-
4 Disk Drives
-
Hard disk data storage from 80 to 128 GB per disk >66 days recording
-
-
Second-Generation Attributes
-
1–4 channels
-
Sampling rates from 78 to 200 kHz
-
4 Disk Drives
-
Hard disk storage from 240 GB to 1 TB per disk
-
Choice of hydrophone sensitivity/System gains
-
Delayed start available
-
More compact due to higher energy efficiency
-
Recording time dependent on disk size
-
Summary of Current Technology
-
Microprocessor:
-
Two stereo a/d boards (total of four 16-bit sigma-delta a/d)
-
-
Maximum sample rate:
-
50 kHz/channel in 4-channel mode
-
100 kHz/channel in 2-channel mode
-
192 kHz in single-channel mode
-
-
Storage media:
-
2.5 in. hard drives (four each)
-
Flash drives (future)
-
-
Max storage:
-
Hard drive dependent
-
Current: 4 × 120 GB drives (480 GB)
-
Available soon: 4 × 250 GB (1 TB) up to 4 × 1 TB
-
-
Number of bits: full 16 bit
-
Size: 2 m long, 0.425 m diameter, 110 lbs
-
Average power supply/consumption:
-
Currently the EARS are storage limited, not battery limited
-
Cost of EARS buoys
Approximate cost of Generation 2 EARS buoys considering different configurations
LADC paid $23.4K per buoy with a single hydrophone in our configuration (group of 10)
Add $950 per hydrophone and $33/m for cabling to build two or four channel arrays
6.3 LADC Marine Mammal Acoustic Experiments
Experiments in the Gulf of Mexico and the Ligurian Sea have targeted both sperm and beaked whale s. Audio results and visualizations of these recordings reveal rich detail of Odontocete clicks and enable new analyses such as the identification of individual whales from the properties of their clicks. Beginning with experiments in 2001, LADC scientists have studied sperm whale clicks and clicking behavior. In 2007, the study was extended to beaked whale clicks.
LADC has led or participated in seven experiments for the study of marine mammal acoustics. They are:
-
1.
LADC 01—Gulf of Mexico
-
2.
SIRENA 02—Ligurian Sea w/NURC
-
3.
LADC 02—Gulf of Mexico
-
4.
SIRENA 03—Ligurian Sea w/NURC
-
5.
ZIPHIO 06—Ligurian Sea w/NURC
-
6.
LADC 07—Gulf of Mexico
-
7.
LADC 10—Gulf of Mexico
Experiments 1, 3, 6, and 7 were performed by LADC , and the remaining experiments were led by the NATO Undersea Research Centre (NURC) (now the Centre for Maritime Research and Experimentation (CMRE)) with LADC participation. All experiments used EARS buoys.
1. LADC 01
-
Northeastern Gulf of Mexico.
-
July 16 through Aug 21, 2001. The acoustic record ings spanned 36 days.
-
Buoys moored upslope off mouth of Mississippi River in water depths of 600, 800, and 1000 m.
-
Buoys moored 50 m above bottom.
-
During the experiment, Tropical Storm Barry passed within 100 nmi of the EARS buoys.
3. LADC 02
-
North central Gulf of Mexico.
-
August 19 through September 15 planned, retrieval 23–24 October, 2002; 22–57 days recorded.
-
Buoys moored upslope off mouth of Mississippi River in water depths of 600, 800, and 1000.
-
Buoys moored 50 m above bottom.
-
Tropical Storm Isidore passed with 73 nmi of the EARS buoys, and Hurricane Lili passed within 116 nmi.
LADC 01 and LADC 02
-
Three Single-Channel G1 EARS Buoys.
-
11.7 kHz sampling rate.
6. LADC 07
-
Gulf of Mexico south of Gulfport, MS, at 1550 m contour.
-
July 3 to July 14, 2007.
-
Six Single-Channel G2 EARS buoys on bottom-mounted moorings.
-
Three moorings at north site 3 km separation.
-
Three moorings at south site 5 km separation.
-
750–800 m hydrophone depths.
-
192 kHz sampling rate.
-
2.2 TB recorded (9 days, ~¾ capacity).
7. LADC 10
-
Six Single-Channel G2 EARS buoys on bottom-mounted moorings.
-
Two buoys at LADC 07 north site buoy locations, two buoys at LADC 07 south site buoy locations, and two buoys at deepest site for LADC 01 and 02.
-
192 kHz sampling rate.
-
The buoys recorded for 12 days.
-
Deployments were done from 9 September through 12 September 2010.
-
Recording depths of 1000 m for the LADC 07 sites and 800 m for the LADC 01-02 site.
Recently, LADC has received funding from the Gulf of Mexico Research Initiative (GoMRI) RFP IV for two exercises to compare sperm and beaked whale s and dolphin measurements using ten single-channel Generation 2 moored EARS buoys, hydrophone arrays towed by autonomous surface vehicles (ASV ’s), and hydrophone arrays towed by gliders , and a test cruise in the off year between the two exercises. All measurements will be made at the three sites of the LADC 10 experiments. The first exercise will begin with a cruise to deploy the hydrophones and operate the ASV’s and glider in June of 2015. A retrieval cruise will take place in October 2015. Tests of equipment upgrades will be made on a cruise in July 2016. The second field exercise will begin with deployment in April of 2017 and retrieval in August of that year. Again, ASV’s and gliders will be tested. Funding is also included for equipment upgrades, experimental planning, and data analysis.
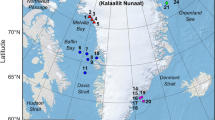
Figure 6.3 shows the locations of all GoM exercises. The green circles are the LADC 01 locations and the yellow circles are the LADC 02 locations; the yellow squares show the north and south buoy sites of the LADC 07 exercises, and the pink pushpins show the LADC 10 locations. The red flame shows the site of the BP oil spill .

The locations of all GoM exercises. The green circles are the LADC 01 locations, the yellow circles are the LADC 02 locations, the yellow squares show the north and south buoy sites of the LADC 07 exercises, and the pink pushpins show the LADC 10 locations. The red flame shows the site of the BP oil spill
Acoustic data from any single LADC experiment can support a variety of marine mammal related studies, often in parallel and complementary to each other. For example, several techniques were explored for individually identifying clicking sperm whale s, and simultaneous results compared. Other research regarding acoustic behavior and population estimation is summarized below as well.
6.4 Click Structure Analysis and Sperm Whale Identification
The ability to acoustically discriminate one clicking sperm whale from another is a challenging problem yet an important one to solve if acoustics are to be used to passively study these animals’ behavior. For example, associating all the clicks from just one individual whale while multiple animals are clicking concurrently is an important first step in the passive acoustic localization and tracking of sperm whales. There is also the question of whether these animals can distinguish each other through the individual characteristics of their clicks. The LADC group explored several techniques in parallel for grouping click sequence s from individual whales, as described below and in subsequent sections.
6.4.1 Click Structure Analysis
The click production mechanism of the sperm whale results in a “click” that is not just a single impulse but rather has a structure from the sum of multiple reflection s internal to the whale’s head (called the p0, p1, and p2 pulses), and the recorded click structure will vary depending on the animal’s aspect to the receiver . It has been LADC ’s experience that any change in recorded click structure as an animal moves relative to the receiver happens slowly compared to the rate in which clicks are made. Thus, when trying to match click event s from the same individual, adjacent clicks that have a consistent periodicity and structure (and frequency spectra) are likely matches from the same individual. As an example, when time series of clicks assumed to be from the same individual are time aligned and stacked, their persistent, or slowly evolving, shapes become visually apparent. Figure 6.4 illustrates this in two waterfall plots of evolving click shapes from two different sperm whales with unique click structures. These two click trains overlapped in time and had slightly different average interclick interval s.

Time-aligned and stacked time series of clicks from two different whales clicking concurrently, as recorded by sensor N2 during LADC07. The slowly evolving shapes of the click structures and consistent interclick interval allow association of each click to an individual whale when manually inspected
While manually associating clicks via visual inspection and periodicity clues is possible over short time spans, it is too laborious a method to apply to hours or days of acoustic data, suggesting the need for an automated solution, as explored in the following sections.
6.5 Cadence Frequency Analysis and Identification of Individual Clicking Whales
The Cadence Frequency Analysis (CFA) algorithm was developed as a part of the LADC integrated software package to identify individual sperm whale s. The algorithm allows the determination of the number of simultaneously phonating animals in a group and the identification of the click train s of an individual animal in the group of interleaving click trains. It also tests the hypothesis that simultaneously phonating animals in a group will adjust their clicking rhythms to efficiently scan the environment and distinguish among clicks emitted by different group members (Sidorovskaia et al. 2009a, b, c, 2010; Tiemann et al. 2011).
The CFA method can quickly and reliably associate clicks with individuals in a group of whales who are phonating simultaneously. It can be of interest also for developing technology associated with real-time passive acoustic monitoring from autonomous mobile platforms. The method is robust against environmental changes, signal-to-noise ratio, and surface and bottom reflections , and is species independent. The algorithm performance is independently validated by identifying individual click trains using two other techniques: the Passive Acoustic Localization using the Time Difference of Arrival (TDA) method (Tiemann et al. 2006; Tiemann et al. 2008; Van der Schaar et al. 2009) and cluster analysis. These techniques are described in Sects. 6.8 and 6.6 of this chapter.
Diving marine mammals produce echolocation click s to orient themselves in the ocean, find prey , and perform short-range localization and tracking for prey capture. Many types of marine mammals dive in groups and echolocate simultaneously. The development of the CFA algorithm is based on the hypothesis that each animal in a group has the ability to slightly vary its interclick interval (frequency of clicks) to avoid interference with signals produced by nearby divemates. Logically it would also allow them to utilize clicks produced by other animals in the group to extract information about the surroundings with higher efficiency. The concept of rhythmic identity measurements was previously discussed by Andre and Kamminga (2000). They also mentioned anecdotal evidence that drummers from African tribes, trained at early ages to play individual rhythms in a group, could easily determine how many rhythmic themes were preset in a playback of overlapping click trains . The authors proposed that marine mammal click production can be rhythmically modulated to prevent interference among individuals, and such rhythmic patterns could serve as acoustic signatures of individuals. Andre and Kamminga used a cross-correlation analysis of time domain signals for sperm whale clicks and codas to reveal rhythmic modulation. Despite the similarity of initial hypotheses, the CFA algorithm is fundamentally different and originated from an algorithm developed for human motion analysis (Ekimov and Sabatier 2008, 2011). The CFA algorithm does not have some common shortcomings of the other two methods.
-
1.
The CFA algorithm is species and environment independent. Reflections do not degrade the algorithm performance.
-
2.
The CFA algorithm is dynamic and follows the rhythm evolution during a dive.
-
3.
The CFA algorithm allows for multiple frequency band selections and can serve as a simultaneous detector and classifier .
-
4.
The CFA algorithm is robust to low signal-to-noise ratio due to band selectivity .
-
5.
The implementation of a concurrent “cleaning” procedure reveals rhythmic patterns of strong and faint click trains simultaneously.
The algorithm consists of two main steps: (1) formation of a band-limited energy function , and (2) frequency analysis of the band-limited energy function (Fig. 6.5). Figure 6.5a shows the raw temporal PAM signal recorded by the Environmental Acoustic Recording System (EARS) Buoy during July 2007 in the Northern Gulf of Mexico. The signal contains multiple sperm whale echolocation click s. The sampling frequency of the EARS buoy continuous recordings was 192 kHz and provided reliable discrimination among different marine mammal species present in the area (sperm whales, beaked whale s, pilot whales , and dolphins ). A sliding window Fourier transform is applied to the temporal data to obtain the spectrogram shown in Fig. 6.5b. The sliding window length is approximately the duration of the expected clicks (4 ms for sperm whales). The bright vertical lines on the spectrogram represent sperm whale clicks. The main energy of the sperm whale clicks is concentrated in the 5–10 kHz band; this band is chosen for energy function formation. The energy of all spectral components in the chosen band is summed for each temporal point of the spectrogram. The temporal evolution of the normalized energy functions for three different frequency bands are shown in Fig. 6.5c. The 5–10 kHz band energy function characteristic of sperm whale clicks is analyzed next to reveal different rhythmic patterns of simultaneously phonating sperm whales.

The Cadence Frequency Analysis algorithm is applied to LADC Gulf of Mexico 2007 PAM data. (a) Raw temporal broadband acoustic signal of 25 s duration. (b) Spectrogram of the acoustic signal with vertical lines corresponding to sperm whale clicks. (c) Normalized band-limited energy function vs time obtained by incoherently summing spectrogram frequency components over the chosen band. (d) Spectrogram of the energy function for the 5–10 kHz band with sliding window of 11 s. The arrows indicate two whales phonating with offset interclick frequencies of 1.2 and 1.6 Hz. The temporal evolution of the interclick interval for the 1.2 Hz individual can be also seen
The second step of the CFA algorithm consists of obtaining a spectrogram of the band-limited energy function , as shown in Fig. 6.5d. The cadence frequency spectrogram in Fig. 6.5d shows two sperm whale s clicking simultaneously at 1.2 and 1.6 Hz (as indicated by arrows). The dynamic evolution of the lower frequency to 1 Hz at time marker 16 s can also be seen. The higher frequency content of the cadence frequency spectrogram (above 2 Hz) does not represent any new information. The prominent frequency components above 2 Hz are amplitude-modulated harmonics of the fundamental cadence frequencies due to properties of the Fourier Transform. It should be noted that a careful record of time axis resolutions has been kept for associating time points in the raw signal , energy function , and cadence frequency spectrograms. The algorithm is not affected by the multipulse structure of sperm whale clicks or reflections because they will have the same cadence frequency. These may appear in later iterations of the CFA algorithm when the strong clicks of an individual whale (following the cadence spectral line) are removed from an original raw signal to reveal low amplitude phonations. The number of whales in a group and the click association with a particular whale will remain unchanged.
The method was applied to a 90-s segment of the experimental dataset collected in the Northern Gulf of Mexico in 2007 by the Littoral Acoustic Demonstration Center (LADC) and verified by comparison with results provided by two independent algorithms: manual association of clicks from the time difference of arrival maps (Tiemann et al. 2006), and self-organizing map clustering based on three click attributes (temporal structure , spectral structure , and wavelet transform structure) (Ioup et al. 2004). Figure 6.6 shows the cadence analysis final time-frequency maps for three hydrophones N1, N2, and N3 (N1 and N2 are about 2 km apart, N1 and N3 are about 2 km apart, and N2 and N3 are 4 km apart). Eight iterations of CFA are applied to the data from each hydrophone. The iteration number is identified by a colored arrow, and the same color-coded symbols correspond to the click time and cadence frequency pair. Hydrophone N2 clearly shows three whales phonating at 1.0, 1.2, and 1.6 Hz (1, 0.83, and 0.62 interclick interval s (ICI), respectively) during the first 30 s. The same cadence frequency pattern is seen on hydrophone N1. The distant N3 hydrophone does not pick up all three whales, probably due to click directionality. To verify the cadence frequency-whale association made based on the results produced by the CFA, we compared it with the manual click association from the TDA and the self-organizing map methods for the same segment of data. Results show over 80 % agreement among all three methods in associating clicks with individuals and their click production times (see Fig. 6.7). Due to the cleaning algorithm and iterative approach, the CFA algorithm is much more sensitive to weak signals, which explains why many CFA click detections are unmatched by the TDA and self-organizing map algorithms in Fig. 6.7.

Eight iterations of Cadence Frequency Analysis are applied to the data from three recording EARS buoys of the 2007 experiment. Star symbols identify the time and interclick frequency of sperm whale clicks. Three individuals phonating with slightly offset interclick interval s are easily identified on N11 and N21 buoys in the first 30-s of the analyzed segment. Only two individuals are identified on the N31 buoy

Comparison of CFA individual animal click production time results with self-organizing map clusters (a), and TDA results (b). Horizontal lines correspond to the individuals identified by the algorithms. Symbols are located at times of individual clicks
In summary, the CFA algorithm shows good agreement with other algorithms targeting identification of individuals in a group of simultaneously phonating marine mammals. It also supports the hypothesis of “polite speakers ,” i.e., sperm whale s adjust their interclick interval s so that they do not overlap with the interclick intervals of divemates.
6.6 Identification of Individual Whales from Click Properties by Clustering
The initial motivation for working with individual clicks came when G. Ioup and J. Ioup noticed that all the clicks in a sperm whale coda were similar both in the time and frequency domains, but that they could differ from coda to coda. Coda clicks are used for communication, and they occur in groups of 6–15 clicks with an interclick interval of approximately 40 ms and a click duration of 3 to 6 ms. It has been shown that the time difference between peaks (intraclick interval) within a sperm whale echolocation click is related to the size of the whale (Norris and Harvey 1972; Møhl et al. 1981; Gordon 1991; Goold 1996; Rhinelander and Dawson 2004; Teloni et al. 2007; Growcott et al. 2011). The observation that coda clicks from a given whale are similar to each other but differ from the clicks of other whales is consistent with the connection of click properties to the size of the whale.
Baggenstoss (2011, 2013) has developed sophisticated mathematical approaches to associate echolocation click s into click trains originating from individual sperm and beaked whale s. The methods can break down if there are too many overlapping click trains and/or if some of the click trains are sparse. Baggenstoss has advanced his method by using the cross-correlations among the clicks to assist in associating the clicks into click trains. His work provides another independent confirmation that properties of clicks differ from one individual whale to another.
Figure 6.8 displays one minute of EARS recorded underwater acoustic data from 2001. The top graph is an amplitude proportional to pressure plotted versus time. The middle figure is a data spectrogram showing frequencies up to 6000 Hz on the vertical axis and time on the horizontal axis. Color gives the intensity of the transform, with red indicating the highest intensity. The bottom figure is the 0–1000 Hz portion of the spectrogram. The 6000 Hz spectrogram shows that there are many echolocation click s and some codas. The lower spectrogram shows clearly the seismic airgun firing every 11 s. The source ship was 107 km from the EARS buoy.

One minute of recorded data. The top graph is an amplitude proportional to pressure plotted versus time. The middle figure is a data spectrogram showing frequencies up to 6000 Hz on the vertical axis and time on the horizontal axis. Color gives the intensity of the transform, with red indicating the highest intensity. The bottom figure is the 0–1000 Hz portion of the spectrogram. The 6000 Hz spectrogram shows that there are many echolocation click s and some codas. The lower spectrogram shows clearly the seismic airgun firing every 11 s. Reprinted from G.E. Ioup et al. (2009)
Figures 6.9a, b show the time domain signals and the spectrograms of two segments of four seconds of sperm whale clicks; each contains a coda. As in Fig. 6.8, the top figure in each group of three is proportional to the pressure plotted versus time. The middle figure shows the spectrogram with frequency to 6000 Hz on the vertical axis and time on the horizontal axis. Color indicates the magnitude of the transform. The bottom figure shows the frequency to 1000 Hz.

Two 4-s segments of acoustic record ings from the Gulf of Mexico, containing sperm whale codas. (a, b) Time domain signals and the spectrograms of two different segments of four seconds of sperm whale clicks; each contains a coda. The top figure in each group is an amplitude proportional to the pressure plotted versus time. The middle figure shows the spectrogram with frequency to 6000 Hz on the vertical axis and time on the horizontal axis. Color indicates the magnitude of the transform. The bottom figure shows the frequency to 1000 Hz. Reprinted from Tiemann et al. (2011)
Figure 6.10 shows an overplot of the magnitude spectra of all the clicks in a coda for five different codas. The similarity for clicks within a coda is striking. Figure 6.11 shows an amplitude versus time overplot of the clicks within a coda offset in the vertical axis, with similarity among the clicks evident.

Overplot of the magnitude Fourier spectra versus frequency for all the clicks in five sperm whale codas. Each panel is one coda, with individual clicks in each coda in a different color. Note that the shapes for the clicks in one coda are quite similar to each other, but that they can be distinctly different from those in other codas. A few overlapping echolocation click s from other whales are also included in these plots. Reprinted from Tiemann et al. (2011)

As has been pointed out by Tiemann (see, for example, Sect. 6.4), identification of sperm whale echolocation click trains by humans is far too laborious when needed for many days or even hours of recordings. Baggenstoss’ (2011, 2013) automated methods are one approach to solving this problem for sperm and beaked whale s echolocation trains. The method of cadence analysis, Sect. 6.5, is another. A method of associating clicks with individuals, which avoids the need for click-train identification, and also applies to sperm whale coda clicks, is clustering of clicks. Clustering (Hartigan 1975; Seber 1984; Spath 1985; Estivill-Castro 2002) means putting clicks which closely resemble each other into individual clusters (classes ), each of which presumably represents the clicks from one whale. Clustering is normally performed by computer analysis. The fact that the whale clicks have some natural biological variation from click to click for the same animal, and also changing physical parameters, such as whale depth and aspect between the whale and the receiver (the latter especially important for sperm whale echolocation clicks), means that the received signal from one whale will change over time and that the clustering must be designed with these factors in mind.
Clustering methods, mainly K-means (Lloyd 1982; Seber 1984; Spath 1985) and Self Organizing Maps (SOM ) (Kohonen 1989; Pandya and Macy 1995), have been compared for application to whale clicks. SOM has been found to be more useful. It is iterative and allows straightforward incorporation of stopping criteria such as minimizing the Euclidean distance s within a cluster and maximizing the Euclidean distances between cluster centers. It adds new clusters as it iterates. If adding a new cluster worsens the quality measures, the iterations are terminated and SOM gives the number of clusters needed. This number has turned out to be a good first estimate of the number of whales present.
SOM has been applied to both sperm whale coda and echolocation click s and beaked whale echolocation clicks. LADC scientists have been using cluster analysis since 2004 (Ioup et al. 2005, 2009, 2010; Tiemann et al. 2011). The coda clustering results are shown in Fig. 6.12, which gives the results of clustering 43 sperm whale codas occurring over a 3-min interval for data collected in 2001 in the northern Gulf of Mexico. The clustering is done by taking an average click for each coda and clustering the 43 codas using the average click for each. Figure 6.12a shows the clustering results based on the time data. Figure 6.12b is based on clustering the magnitude of the discrete Fourier transform (DFT), and Fig. 6.12c is obtained by clustering the discrete wavelet transform s (DWT) of the time signals. As stated above, SOM algorithms determine how many clusters are needed to obtain optimum results. The time and wavelet clustering found that four clusters (four whales) were optimal, whereas the Fourier clustering put two codas in a fifth cluster. The time and wavelet clustering agreed completely; the Fourier clustering disagreed for three codas out of the 43 total codas. More recent work has shown that if clustering is done with the complex DFT instead of simply using the magnitude, better agreement is obtained with the time and wavelet clustering.


The results of clustering 43 average coda clicks, one from each of 43 sperm whale codas. (a) displays the clicks in four clusters determined by SOM using the time signal; (b) shows the five classes found by SOM based on the DFT magnitude; and (c) gives the four classes resulting from applying SOM to the DWT, which are the same results as in (a)
Tiemann used 46 sperm whale echolocation click s from the LADC07 data to do a click-train analysis. He concluded that there were two whales clicking. When the same 46 clicks were clustered using SOM , the output showed three clusters. This is still a grey area in clustering, as sometimes minor differences in clicks will lead to the creation of an additional cluster when none is needed. It is hoped that incorporating click change detection (Sect. 6.7) into clustering will make the cluster count more accurate. In this case, to compare the clustering to Tiemann’s manual click-train analysis, the maximum number of clusters was set to two in SOM. The results are shown in Fig. 6.13a, b, c. Figure 6.13a gives the two clusters for the time signals, (b) displays the two clusters for the complex DFT (showing only the magnitude), and (c) shows DWT clusters. Figure 6.14 shows the average click for each cluster, overplotted. The top figure is the time signal, the middle is DFT magnitude, and the bottom is the DWT. All amplitudes are normalized to one. Figure 6.15 shows the comparison of the SOM clustering to Tiemann’s identification . The top figure shows the DFT classes (clusters) and the bottom shows the classes of Tiemann. There is 85 % agreement.

SOM clustering results for 46 sperm whale echolocation click s, which were also analyzed by Tiemann to determine click trains present. (a) Shows the two clusters resulting from using the time signal, (b) gives the results of clustering based on the complex DFT, with the magnitude shown in the graphs, and (c) displays the results of clustering the DWT. Clicks from LADC07 data sets

Average clicks for each of the two SOM clusters found for 46 sperm whale echolocation click s. The top graph gives the cluster average time signals; the middle plot shows the magnitudes of the cluster average complex DFT; and the lower figure contains the cluster average DWT’s

Comparison of SOM clustering with manual click-train identification . Two classes or clusters (two colors) represent two whales. The top figure gives the SOM identifications and the bottom figure the click-train identifications of the two whales. There is 85 % agreement. Reprinted from G. Ioup et al. (2009)
Figure 6.16 shows the clustering of Cuvier’s beaked whale data from the Third International Workshop on Detection and Classification of Marine Mammals using Passive Acoustics, Boston, July 2007. The data were provided by Johnson of Woods Hole Oceanographic Institution and the reference for the data is Zimmer et al. (2005b). SOM found that six clusters were needed for these clicks, implying that six whales are present. Although the figures are quite busy, it can be seen that the clicks within each cluster generally match in shape. Clustering on the linear amplitude of the DFT proved to be much more satisfactory than clustering on the DFT given in dB.

SOM clustering for Cuvier’s beaked whale clicks. Clustering is based on the magnitude of the DFT. Six clusters resulted. Data are from the Third International Workshop on Detection and Classification of Marine Mammals using Passive Acoustics, Boston, July 2007, and were supplied by Johnson of Woods Hole Oceanographic Institution. Data are filtered with a 20–60 kHz Butterworth filter
6.7 Click Change Detection
Tiemann has analyzed clicks collectively by doing manual click-train identification (see, for example, Sect. 6.4). This involves following the click sequence of an echolocating whale, including the identification of multiple reflections , and distinguishing it from overlapping click sequences of other whales. He has developed several tools to facilitate the difficult work.
Meanwhile G. Ioup and J. Ioup have led the investigation of click change detection (CCD) (Ioup et al. 2010, 2011) to deal with the problem of turning sperm whales , which change their aspect with respect to the detector and therefore the properties of their received clicks (Møhl et al. 2003). Starkhammar et al. (2011) have independently developed a click change approach. Click change detection is also helpful in dealing with the change of clicks with depth (pressure) for diving whales and with the natural click-to-click variation for an individual whale.
The click change detection method used to determine whether the successive clicks are from the same or a different whale is based on taking successive cross-correlation maximum values and comparing these values to a threshold . Proper normalization of the cross-correlation is essential. For this purpose, division by the product of the square root of the energy of each click gives the normalization needed. The normalization expression is
and CCmax value is between 0 and 1. The threshold value, based on analysis of the data and on comparison with Tiemann’s results, is 0.4.
The data used by Tiemann for sperm whale click-train identification is from data given in conjunction with the 4th International Workshop on Detection, Classification and Localization of Marine Mammals Using Passive Acoustics, University of Pavia, Italy, in September 2009. Dataset 3—NEMO ONDE deep-sea platform—was Sperm whale sounds recorded at 96 K sampling rate—four hydrophones on a tetrahedron placed at 2000 m depth , 25 km off Catania (Eastern coast of Sicily, Italy). Most files contain sperm whale sounds; one contains other clicks, similar to those emitted by Cuvier’s beaked whale s.
Tiemann has selected 550 successive sperm whale echolocation click s for analysis. By grouping the clicks into trains, he is able to identify which whale is producing any given click. In particular, his analysis can be used to determine whether any click comes from the same whale as the preceding click in time.
The normalized cross-correlation was then calculated between each click and the preceding click and the maximum value of these cross-correlations is plotted versus time, as shown in Fig. 6.17. The plotted points are also color-coded. Green is used when, according to Tiemann’s analysis, class (i) = class(i − 1) (where i is the time index), and red is used when class(i) ≠ class(i − 1).

Normalized cross-correlation value plotted versus click time. Chris Tiemann click-train analysis results indicated by red and green star colors. Click of speaking whale unchanged from previous click colored green, and click of speaking whale changed from previous click colored red, summarized as green: CT class (i) = CT class(i − 1); red: CT class(i) not equal to CT class(i − 1). For more than 98.5 % of the clicks, green stars are above the threshold and red stars are below. Reprinted from Tiemann et al. (2011)
As can be seen in Fig. 6.17, almost all the green points are above the selected threshold of 0.4 and almost all the red points are below the threshold. Click change detection agrees with manual click-train analysis 98.5 % of the time. This is strong evidence that the cross-correlation analysis does a very good job identifying whether the same or a different whale produces a succeeding click.
As part of his analysis, Tiemann identified about 30 successive clicks, as shown in Fig. 6.18, between 125 and 145 s, which appear to be coming from a turning whale. Although there is a significant amount of change in click structure over that period, because sperm whale s turn slowly, the amount of change from one click to the next is small. Therefore, click change detection applied to these clicks shows all clicks to be from the same whale. Not only are the cross-correlations above the minimum of 0.4, they are all greater than or equal to 0.5. Click change detection , at least in this case, has successfully dealt with the problem of a turning sperm whale.

A sequence of about 30 clicks (part of the 550 clicks analyzed) identified by Tiemann as being emitted by a turning sperm whale
While the success of click change detection is notable, it is not by itself capable of identifying which whale is clicking. It remains to be combined with clustering, or otherwise advanced, to accomplish identification .
6.8 Passive Acoustic Localization
Passive acoustic localization includes various methods to estimate the position of a phonating marine mammal, relative to a receiver or array , using only an acoustic record without the benefit of tags or visual observations (Tiemann et al. 2006, 2011; Tiemann 2008; Baggenstoss 2011, 2013). Benefits of acoustic localization include the ability to monitor whale behavior continuously and relatively inexpensively, even in times of reduced visibility. While localization and tracking are useful in studies of marine mammal behavior, they can also be considered another identification tool complementary to the others; acoustic localization is a way to sort or associate acoustic events geographically. For example, the estimated source position s for several consecutive sperm whale clicks should form a continuous track of animal motion. If they do not, there has likely been an error during click association . When used in that manner, localization serves as a way to check the results of any click associations derived through the methods above.
The passive acoustic localization of marine mammals is frequently accomplished by measuring the time of arrival of a given animal phonation at different acoustic sensors . The differences in arrival time s, or time lag , can then be used to geometrically estimate an animal’s position at the time it made the sound. A common technique called hyperbolic fixing uses a measured time lag to trace out candidate source position s on hyperbolic path s relative to a receiver pair . As time lags for the same sound event are measured by other receiver pairs, more candidate source position s can be defined. Finally, a candidate source position shared by all time lags is declared the most likely whale position . As an example, Fig. 6.19 shows the candidate locations for a clicking sperm whale estimated by time lags measured at two receiver pairs for a single click event . The intersection of the hyperbolic paths of possible source positions indicates the most likely whale location approximately 6 km from the hydrophone array .

Hyperbolic curves trace out candidate whale position s associated with a single sperm whale click recorded by the northern receiver array during LADC07. The intersection of the curves represents the most likely whale position
6.9 Identification Cues and Their Integration
The developments summarized in Sects. 6.4 through 6.8 can all be used to help identify individual clicking whales. Consideration of these results has led to these research questions:
-
1.
How can the methods of 6.5, 6.6, and 6.7 be verified ?
-
2.
How can the methods be made more robust and perhaps more efficient ?
-
3.
What do the results tell us about how whales identify each other?
Investigation of these topics is still very much work in progress, but some observations can be made at this stage.
Although coupling the methods of Sects. 6.5, 6.6, and 6.7 with visual identification may provide verification in the future, this has not been possible for LADC cruises to date. The most promising approaches for verifying cadence analysis, clustering, and click change detection have been manual (or, in the future, automated) click-train analysis and click localization . The work of Tiemann and colleagues (Sects. 6.4 and 6.8, and Tiemann and Porter 2003; Tiemann et al. 2006, 2011, Tiemann 2008), Baggenstoss (2011a, b, 2013, 2014, 2015), and Nosal and colleagues (2006, 2007a, b, 2008, 2013a, b; Young et al. 2013) is the key for progressing in this direction. One of the main challenges with LADC data is having enough sequential clicks from one whale (especially for beaked whale s) to identify click trains , and having detection on more than one sensor to do tracking. (Single sensor tracking, Tiemann et al. 2006, is not applicable unless there are enough multiple reflections , usually present with a hard ocean bottom and not in the Gulf of Mexico.) LADC deployments have been mainly for detection, but future exercises will have one site with six clustered hydrophones, to achieve better localization and click-train identification . Thus far, click-train analysis by Tiemann (Sect. 6.4) has been compared to cadence analysis (Sect. 6.5), clustering (Sect. 6.6) and to click change detection (CCD) (Sect. 6.7) for echolocating sperm whale s. The results are excellent for CCD and very good for cadence analysis and clustering.
Increasing the robustness and perhaps the efficiency of the identification methods will be important, but it will be difficult to do until verification is used to get confidence in the accuracy of the methods. Verification will also guide the combining of methods for increased accuracy.
It is likely that whales use elements of all of the methods described in Sects. 6.4 through 6.8 to identify and locate each other. Binaural hearing can give good enough localization to distinguish separated speakers. If better localization is needed, a few directed echolocation click s, coupled with the whales’ superb echolocation abilities, could locate companions.
It seems fairly certain that whales know which whale is speaking. Mothers and calves identify each other, and whales synchronize their dives. Whales have highly developed cochlea and copious ganglia (Ketten 1994; and references cited therein). As mentioned, their ability to echolocate prey is advanced. Although humans might be misled by the shortness of whale clicks, it is quite possible that whales know which one is phonating just from listening to individual clicks.
It is instructive to consider an analogy with human hearing , which may offer guidance, although the analogy could break down for several reasons. Humans are known to need about three cycles to identify a low frequency tone (50 Hz), about 20 cycles to identify a mid-frequency tone (2000 Hz), and about 250 cycles to identify a high frequency tone (10 kHz) (Bürck et al. 1935; Rossing et al. 2002). Sperm whale clicks can have energy up to 25 or 30 kHz, but the bulk of the energy is below 17 kHz. At 10 kHz, which is roughly mid-frequency, a 6 ms click contains 60 cycles. For beaked whale s, which have click duration between 240 and 900 μs and a click frequency range from 20 to 60 kHz, a mid-frequency 40 kHz signal will have 24 cycles in 600 μs. Therefore, the possibility that sperm and beaked whales can identify individuals from the properties of single clicks merits investigation. Data analysis by LADC scientists and others provides important supporting evidence.
For diving whales , whose click properties change due to changing pressure (Thode et al. 2002), it is plausible that the whales use some form of click change detection (CCD) to keep track of which companion is phonating . The same is possible for turning sperm whales , whose click properties change slowly with time at a receiving point.
Since whales foraging together appear to have the ability to adopt different cadences for their echolocation click trains probably to reduce interference with each other, the cadence of an individual can be a component of identifying echolocating companions for each whale.
6.10 Sperm Whale Coda Classification and Repertoire Analysis
As noted in Sect. 6.3 and 6.1, during the summer of 2001, the Littoral Acoustic Demonstration Center (LADC) conducted the acquisition , deployment , and retrieval of three bottom-moored environmental acoustic record ing system (EARS ) buoys in the Northern Gulf of Mexico (GoM). The original focus of the project was to investigate ambient noise and upslope propagation in the area, but after consultation with the Minerals Management Service (MMS) Sperm Whale Acoustic Monitoring Program (SWAMP) , the focus was expanded to include an examination of sperm whale acoustic behavior. The data collected during the summer of 2001 have been used in several studies examining the contributions of marine mammals and offshore drilling to the ambient noise level at the edge of the continental shelf in the Northern Gulf of Mexico (e.g., Newcomb et al. 2004, 2002a, b, c; Snyder et al. 2003). The project presented here is focused on identifying the coda repertoire of sperm whales in the area.
6.10.1 Location
EARS buoys, as described in Sect. 6.2, were used for the measurements in 2001, which produced the data analyzed in this section. The buoys were deployed from a ship on July 16th–19th. The deployment site was chosen to optimize LADC ’s goal of measuring noise propagation up the continental slope , as well as maximizing exposure to the largest concentration of previous sperm whale sightings in the general area. This area has been identified as an area rich in sperm whale activity , specifically around the 1000 m depth contour south/southeast of the Mississippi River delta (Mate et al. 1994; Würsig et al. 2000). The first buoy, EARS 1, was moored at approximately 28° 15′ N and 88° 50′ W. The other two buoys, EARS 2 and EARS 3, were deployed along a 43 km line extending from the 200 m contour to just beyond the 1000 m contour, along which oceanographic data were collected. Data were also collected along a cross track. The LADC01 and 02 buoy locations shown in Fig. 6.3. are specified in greater detail in Fig. 6.20. Total separation between buoys was approximately 25 km with roughly 7 km separating EARS 2 and EARS 3. The buoy closest to land, EARS 3, was located approximately 55 km from the Louisiana shore. EARS 1 was moored at a depth of approximately 1000 m, EARS 2 at 800 m, and EARS 3 at 600 m.

The EARS deployment sites in the Northern Gulf of Mexico. Adapted from image provided by Joal Newcomb
Each EARS buoy continuously recorded acoustic signals with frequencies up to 5859 Hz for a period of approximately 36 days (digital sampling rate of 11.7 kHz). Each buoy gathered 72 GB of acoustic data for a three-buoy total of 216 GB. A total of 2592 h of audio data were recorded and then inspected visually and acoustically using Raven 1.1 (Bioacoustics Research Program 2003).
6.10.2 Classification and Repertoire Identification
Codas are stereotyped sequences of sperm whale clicks lasting from approximately 0.2 to 5 s, which occur in a pattern of about 3–20 clicks (Whitehead 2003). Some of the codas we observed are shown in Fig. 6.21. Codas are sometimes produced at the end of a usual echolocation click train, so in following musical terminology , Watkins and Schevill (1977a) called them “codas.”

Spectrograph image illustrating three codas (between solid black arrows) and a creak (between dashed arrows) from the EARS 3 buoy recording file 08652121
Codas are most frequently heard when whales are at or near the surface and are moving slowly in and around one another (Weilgart and Whitehead 1997; Whitehead and Weilgart 1991), but are also heard in small numbers during dives (Madsen et al. 2002). Furthermore, they seem to occur only between whales that are in close physical proximity to one another (Watkins and Schevill 1977b).
Codas appear to be used primarily as intragroup communication, rather than in communication between groups since codas lose their acoustic integrity after approximately 2 km and groups are usually separated by much greater distances (Madsen 2002; Weilgart and Whitehead 1997; Whitehead 2003). In addition, there is regional geographic variation in coda type s (Weilgart and Whitehead 1997). Weilgart and Whitehead (1997) suggest that this variation results from the distinctive coda dialects of particular social units , which have preferred geographic ranges. Strong group-specific dialects that persist over a number of years seem to exist (Weilgart and Whitehead 1997). Furthermore, differences in repertoire have been found in different geographical locations (Apple 2002; Moore et al. 1993; Pavan et al. 2000; Weilgart and Whitehead 1993, 1997).
To determine the coda repertoire of the whales recorded in this study, codas were classified based on methodology used in previous studies (Apple 2002; Moore et al. 1993; Weilgart and Whitehead 1993, 1997). Using both the number of clicks in the coda and the structure of the click pattern , a coda type was determined. For example, 5-click codas were labeled as type: 5R (Regular: with equal spaces between clicks), 5V (Variable: unevenly spaced clicks), 5 + 1 (plus-one: contained a double interval between the last two clicks), or 5 1+ (one-plus). The one-plus structure is a deviation of the protocol used in previous studies and was created here due to the frequency of codas with a double interval after the first click of the coda.
A total of 5035 codas were identified from the EARS recordings and were classified into 34 types (Table 6.1). Representing 19.09 % of all codas, the type 6V was the most prevalent, with 4V (13.45 %) and 7R (11.88 %) ranking second and third.
The identification of the coda repertoire from sperm whale s in a given area can provide information not only about vocal behavior, but also about group structure and group affiliation . This LADC project recorded a large number of codas over the course of the study period. Because codas from known breeding grounds are often produced by socializing females (Goold 1999; Gordon et al. 1992; Marcoux et al. 2006), and this study area is known to be inhabited by a mixed group of mature female s and immatures (Watwood et al. 2006; Würsig et al. 1999), it is likely that there were mature females in or passing through the area for much of the study period.
6.11 Statistical Modeling and Population Estimation
6.11.1 Modeling Acoustic Data
Monitoring deepwater marine mammal abundance based on acoustic record ings has been introduced as a new tool when visual observations are limited or unavailable (Marques et al. 2009; Barlow and Taylor 2005). However, only a few case studies to estimate population densities based on acoustic cue s have been published in the literature. In this section, we describe a statistical methodology that was recently developed (Ackleh et al. 2012) which utilizes passive acoustic data collected before and after the 2010 Deepwater Horizon oil spill incident to assess its impact on the population abundance of endangered sperm whale s.
We begin discussion of the statistical model s for the acoustic data collected from buoys in the northern and southern sites in years 2007 and 2010. The click rate histogram s imply a “power law " pattern. Therefore, we adopt a general power law model (Johnson et al. 1992) for probability density function s P, to fit the histograms formed for selected datasets for a specific experiment and location.
where k(θ, b) is a normalizing constant, and \( k\left(\theta, b\right)={\left({\displaystyle \sum}_{x=0}^{\infty }{\left(b+x\right)}^{-\theta}\right)}^{-1} \).
The log-likelihood function L for a given dataset with random observations, x 1, x 2, …, x n , of the number of click s recorded at n consecutive minutes is given by
\( L\left(\theta, b\Big|{x}_1,{x}_2,\cdots, {x}_n\right)= \ln \left({\varPi}_{i=1}^nk\left(\theta, b\right)/{\left(b+{x}_i\right)}^{\theta}\right) \).
Estimates of the parameters b and θ (denoted by \( \widehat{b} \) and \( \widehat{\theta} \)) are obtained by maximizing the above log-likelihood function over a parameter space \( \varOmega =\left\{\left(\theta, b\right)\Big|\theta >1,b>0\right\}. \) However, maximization of L with respect to θ and b simultaneously runs into the “saddle point” type problem, where L is monotonically increasing with respect to θ and b. Thus, to resolve this problem, a different approach is used in Ackleh et al. (2012). In particular, for a fixed b, L is maximized with respect to θ and the optimal value θ, which depends on b, is obtained and denoted θ(b).
All power law fitting curves for buoys in the northern and southern sites are overlaid in Fig. 6.22. The power law functions indicate a sharp difference between years 2007 and 2010 in the northern area but similar fittings in the southern area. Acoustic activity of sperm whale s at the closest northern site (9 miles away from the incident site) shows a decrease between 2007 and 2010, but no obvious differences are observed at the southern site (23 miles away).

Comparison of the power law fittings for the northern (Left) and southern (Right) sites. N1–N3 and S1–S3 correspond to data collected in 2007, A1–A3 correspond to data collected in 2010. (Extracted from earlier work (Ackleh et al. 2012))
6.11.2 Estimating Population Density
The objective of this section is twofold: (1) to formulate a statistical methodology for the point and interval population abundance estimation based on passive acoustics, and (2) to apply this model to data collected before and after the oil spill in the Gulf of Mexico (GoM) and to assess its impact on the sperm whale s population.
Following Marques et al. (2009), a point estimate of population density \( \widehat{D} \) based on the number of detected cues n c over a time period T can be given by
\( \widehat{D}=\frac{n_{\mathrm{c}}\left(1-\widehat{c}\right)}{K\pi {w}^2\widehat{P}\left(T\widehat{r}\right)} \).
Here, ĉ is the estimated proportion of false positive detections and K is the number of replicate sensors used in the experiment (\( K=1 \) in our experiment). A target region is considered to be a circular area centered at the buoy location with w representing the maximum detection radius. The expected number of cues per unit time by a single mammal is denoted by r. The estimated average probability of detecting a cue is given by \( \widehat{P} \).
Different approaches have been applied to obtain a suitable value of \( \widehat{P} \). In the first procedure (Marques et al. 2009), parameters p y and h(y) can be evaluated by relating sensor detection events to the sounds produced by tagged animals. This requires tagging a considerable number of animals in a survey area (which was not conducted during LADC deployments). The second approach (Tiemann et al. 2006) is computationally and operator-time costly since it involves a localization of an animal producing each detected cue. The third approach (Zimmer et al. 2005; Küsel et al. 2011) is based on modeling utilizing environmental data and the animal’s beam pattern that are absent for GoM . We adopt a different approach and assumptions to obtain \( \widehat{P} \) (Ackleh et al. 2012). In particular, let h(y) be the probability of the whale being y units away from the buoy location, and p y be the probability a cue is detected provided that the cue is generated at a distance y. Then
\( \widehat{P}=\underset{0}{\overset{w}{{\displaystyle \int }}}{p}_{\mathrm{y}}h(y)\mathrm{d}y \).
We further assume that p y is one of two general types:
We generalize h(y) used in Marques et al. (2009) to \( h(y)=\left(d+1\right){y}^d/{w}^{d+1} \).
The most appropriate values of parameters β and d are unknown and probably hard to obtain. For various combinations of β and d this methodology is applied to data collected before and after the 2010 spill, as the acoustic detection techniques used are the same, to see if there is a significant difference between 2007 and 2010 models.
Due to difficulties encountered in estimating the variance of \( \widehat{D} \) directly, we apply the following nonparametric bootstrap method for the interval estimates of population density .
-
1.
Specify the location and survey of interest, obtain the k consecutive hourly point estimates of D for the given survey, and denote them by \( {\widehat{D}}_1,\cdots, {\widehat{D}}_k \).
-
2.
Draw with replacement a bootstrap sample of size k from \( \left\{{\widehat{D}}_1,,\cdots,, {\widehat{D}}_k\right\} \), denote this sample by \( {{\widehat{D}}^{*}}_1,\cdots, {{\widehat{D}}^{*}}_k \), and then calculate the sample average \( {\overline{D}}^{*}={\displaystyle \sum}_{i=1}^k\frac{{{\overline{D}}_i}^{*}}{k} \).
-
3.
Repeat step (2) M times, where M is a large number, and obtain M bootstrap replicates \( {\overline{D}}^{*} \).
-
4.
Sort the bootstrap replicates \( {\overline{D}}^{*} \) from the smallest to the largest as \( {{\overline{D}}^{*}}_{(1)}\le {{\overline{D}}^{*}}_{(2)}\le \cdots \le {{\overline{D}}^{*}}_{(M)} \).
-
5.
Calculate the lower and upper tail \( \frac{\alpha }{2} \)-probability (empirical) cut-off points of \( {{\overline{D}}^{*}}_{(1)},{{\overline{D}}^{*}}_{(2)},\cdots, {{\overline{D}}^{*}}_{(M)} \) and denote them by \( {{\overline{D}}^{*}}_{\left(\left(\alpha /2\right)M\right)} \) and \( {{\overline{D}}^{*}}_{\left(\left(1-\alpha /2\right)M\right)} \), respectively. Then the approximation of \( \left(1-\alpha \right) \) level confidence interval for the hourly point estimates of D is \( \left({{\overline{D}}^{*}}_{\left(\left(\alpha /2\right)M\right)},{{\overline{D}}^{*}}_{\left(\left(1-\alpha /2\right)M\right)}\right) \).
By a comparison of manual and automatic detections we obtain \( \widehat{c}=0.059 \). From the literature on sperm whale s (Watwood et al. 2006: Whitehead and Weilgart 1990) we take the cue production rate to be 1.22 clicks per second per whale and the maximum detection radius as \( w=20 \) km. The values \( \beta =2.5,\kern0.5em d=1 \) are chosen so that the estimated sperm whales density before the spill matches the NOAA reported population of 1665 in the Northern GoM (Waring et al. 2009). The confidence level is set to be \( 1-\alpha =0.95 \), and the number of replications is \( M=5000 \). Based on the values and assumptions chosen above, we obtain estimates of population density as presented in Fig. 6.23. These results show that there is a considerable decrease of the population abundance from 2007 to 2010 at the northern site closest to the DWH incident site and an apparent increase at the southern site. One can further observe that the decrease in the population density at the northern site nearest to the DWH site exceeds statistical uncertainties and can be accepted as an existing trend.

The 95 % confidence interval of the average hourly density for all survey sites in northern and southern locations. Light gray denotes northern sites and dark gray denotes southern sites. (Extracted from our earlier work (Ackleh et al. 2012))
6.12 Summary
This chapter presents the work of the Littoral Acoustic Demonstration Center (LADC), a consortium comprising University, private industry, and U.S. Navy scientists, formed in early 2001. The Universities currently represented are the University of Louisiana at Lafayette (UL Lafayette), the University of Southern Mississippi (USM), the University of New Orleans (UNO), and Oregon State University (OSU). LADC was formed to utilize technology developed by the Naval Oceanographic Office (NAVOCEANO) to make environmental underwater acoustic measurements. Specifically, the technology consists of buoys containing electronics and power and having an external hydrophone(s), which are moored to the ocean bottom to make acoustic measurements over extended periods of time. The buoys are named Environmental Acoustic Recording System (EARS). LADC is using EARS to make measurements which are not part of NAVOCEANO’s mission. NAVOCEANO provides technical guidance and support to LADC.
A list of LADC scientists and their affiliations is in 6.1. This section also elucidates the developments which led LADC into recording marine mammals.
Initially the EARS technology used by LADC recorded to about 6000 Hz. Then buoys which measured to 25,000 Hz were developed. Now our Generation 2 EARS can measure to 25 kHz on four channels or to 96 kHz on one channel. The latter configuration makes possible the measurement of beaked whale clicks and much of the spectral band of offshore dolphin clicks and whistles. A description of the features of EARS buoys which are important for LADC applications is given in 6.2.
After recording sperm whales in 2001 and 2002, and a seismic airgun array on one mooring in 2003 (Tashmukhambetov et al. 2008), LADC made the first recordings of beaked whales in the Gulf of Mexico (GoM) in 2007. These measurements were at sites which are located 9 and 23 miles from the Macondo well. Then in September 2007, LADC conducted a large exercise to produce detailed measurements of the acoustic field of a seismic airgun array. In 2010, after the oil spill at Macondo, LADC went back to the 2007 sites close to Macondo and to a 2001–2002 site, 50 miles from Macondo. This enabled before and after comparisons to be made. An article comparing before and after abundances for sperm whales has been published (Ackleh et al. 2012). Important details of all LADC marine mammal experiments and experimental collaborations are given in 6.3.
Since the early 1970s, significant progress has been made in understanding the bioacoustics of sperm and beaked whales. Much of the research is reported in the references given in this chapter and in references cited therein. Various facets of click generation and the properties of the clicks themselves are becoming well characterized. Those aspects of the bioacoustics of these whales which are not yet well understood are mainly related to identification of and communication among the whales and the relation of these to their behavior and the environment. Although these topics are difficult to study because there are not animals in captivity, the clarity and volume of the recordings available have permitted advances to be made, even though the science is still in its early stages. Sections 6.4 through 6.10 describe LADC research in these areas. Section 6.8 on Passive Acoustic Localization also stands on its own, since it can be applied to understanding other areas of whale science.
Section 6.4, Click Structure Analysis and Sperm Whale Identification, discusses the structure of the clicks of an individual sperm whale and how this structure can differ from whale to whale. These differences are used to separate the clicks from different whales and associate them over time into click trains, one for each whale. These identified click trains allow several very important analyses to be made, such as determining interclick intervals for each animal. These are found to differ from animal to animal. They also facilitate identifying the click structure for each animal. It is important that they allow the determination of how that click structure can change as the whale changes its aspect with respect to the receiver. While manual click train identification is too laborious to perform on large sets of data, the analysis that has been done has greatly facilitated the development of related methods to study interclick intervals and identify individual whales from the properties of their clicks.
The idea of an interclick interval or an individual click frequency for echolocation clicks, referred to as a cadence frequency, is explored in great detail in Sect. 6.5, Cadence Frequency Analysis and Identification of Individual Clicking Whales. A robust method is developed and applied to identify all different cadence frequencies present among the group of phonating whales. The method allows the association of clicks with individuals in a group of whales. It has been compared to passive acoustic localization in Sect. 6.8 and cluster identification in Sect. 6.6. The cadence frequency analysis algorithm is based on an approach developed for human motion analysis and is a significant improvement on other methods which have been developed to identify cadences. The important advantages of this approach are given in the section. The details of application of the algorithm are discussed. It has been used for sperm whales, beaked whales, pilot whales, and dolphins. It is found that simultaneously diving whales adopt somewhat different cadences from each other, presumably to keep from interfering with each other as they echolocate. The method shows good agreement with the other approaches mentioned.
In the analysis of the coda clicks of sperm whales, it became obvious that all the clicks in a coda were similar to each other in their time and spectral properties. In comparing codas, it was seen that some codas had the same click properties as some others, while other codas had different click properties. This led to the idea that clustering of coda clicks could be used to identify individual whales. Each cluster or class identified by the clustering method would be associated with an individual. Although several different clustering methods were tried, the one that led to the greatest success is a neural nets-based technique called self-organizing maps. After achieving separation into clusters that were differentiated from each other and could identify sperm whales based on these codas, the method was extended to sperm whale echolocation clicks and beaked whale echolocation clicks. Good agreement was obtained when the clustering of sperm whale echolocation clicks was compared with click train identifications when two whales were present.
In order to deal with the change of received click properties from a turning sperm whale which is changing its aspect with respect to the receiver, a click change detection method was developed. This method is used to determine whether the clicks are from the same or a different whale. It is based on taking successive normalized cross-correlation maximum values and comparing these to a threshold. For the work with sperm whale echolocation clicks reported here, the threshold for the normalized cross-correlation is 0.4. Tiemann has analyzed 550 successive echolocation clicks into click trains. From his analysis it is known for each successive click whether the same whale is clicking or a different whale. The same thing can be determined using click change detection. When the results of manual click train analysis are compared to click change detection, the agreement is 98.5 %, which is very strong supporting evidence for the click change detection method. A sequence of clicks from a turning sperm whale was isolated over a 20 s interval in these data. Click change detection showed a correlation with a minimum of 0.5, indicating that all clicks came from the same whale. In the future click change detection can possibly be used to deal with changing clicks due to changing pressure for a diving whale or the simply natural click to click variation for a single whale.
As stated previously, Sect. 6.8, Passive Acoustic Localization, is important for many applications in studying whale identification, communication, and behavior. Its special importance in this chapter is that it has given a way to check the results of cadence frequency analysis. In the future it can help verify the identifications given by clustering. It is one of the most important ground truth methods available for LADC analysis.
In Sect. 6.9, Identification Cues and Their Integration, an investigation of the combination of the methods of Sects. 6.4 through 6.8 is given. Questions of verification, robustness, and efficiency are considered, as well as how the results of this section can further the science of understanding how whales identify each other. In the absence of sufficient visual identifications, the main bases for verifying the methods have been click train analysis and click localization. It is suggested that whales use elements of all the methods described in Sects. 6.4 through 6.8 to identify and locate each other. Consideration is also given as to whether whales can identify each other from individual clicks. An analogy with human hearing is given to illuminate the possibilities for identification from a single click.
Section 6.10, Sperm Whale Coda Classification and Repertoire Analysis, presents a detailed study of 5035 codas selected from the data of LADC01. It was found that there are 34 different rhythms identified, all given with their frequency of occurrence in the table in the section. Three coda rhythms accounted for almost 45 % of all the codas. These results can be used to give information about group structure and group affiliation among the sperm whales in the region, which are most likely to be mature females and immatures.
Section 6.11, Statistical Modeling and Population Estimation, concerns estimating the population density for sperm whales at the northern and southern sites for LADC07 and LADC10. The estimation is based on using a formula given in the literature in 2009. Because some of the parameters needed for the formula were not measured in the LADC experiments, new techniques have been developed to deduce these quantities. Standard approaches were used to fit probability density functions to the histograms of the data, although different functional forms were needed than those used previously. One of the difficulties is knowing the estimated average probability of detecting a cue. None of the methods for estimating this probability used by others was available for the LADC data so a novel technique was developed. Estimating the variance of the density is also difficult, and a nonparametric bootstrap method was developed to get interval estimates of the population density. Based on the assumptions given in the section, results show a considerable decrease in the abundance of sperm whales from 2007 to 2010 (before and after the Macondo well oil spill) at the northern site, 9 miles away from the spill, and an apparent increase at the southern site, 23 miles away.
References
A.S. Ackleh, G.E. Ioup, J.W. Ioup, B. Ma, N. Pal, N.A. Sidorovskaia, C.O. Tiemann, Assessing the deepwater horizon oil spill impact on marine mammal population through acoustics: endangered sperm whales. J. Acoust. Soc. Am. 131, 2306–2314 (2012)
M. Andre, C. Kamminga, Rhythmic dimension in the echolocation click trains of sperm whales: a possible function of identification and communication. J. Mar. Biol. Assoc. UK 80, 163–169 (2000)
T.C. Apple, Temporal patterns and types of sperm whale (Physeter macrocephalus) codas in the Northern Gulf of Mexico: a metadata analysis, Unpublished M.S. thesis, Texas A&M, (2002)
P.M. Baggenstoss, Separation of sperm whale click-trains for multipath rejection. J. Acoust. Soc. Am. 129, 3598–3609 (2011a)
P.M. Baggenstoss, An algorithm for the localization of multiple interfering sperm whales using multi-sensor time difference of arrival. J. Acoust. Soc. Am. 130, 102–112 (2011b)
P.M. Baggenstoss, Processing advances for localization of beaked whales using time difference of arrival. J. Acoust. Soc. Am. 133, 4065–4076 (2013)
P.M. Baggenstoss, The Jonker-Volgenant algorithm applied to click-train separation. J. Acoust. Soc. Am. 135, 2485–2488 (2014)
P.M. Baggenstoss, A multi-hypothesis tracker for clicking whales. J. Acoust. Soc. Am. 137, 2552–2562 (2015)
J. Barlow, B. Taylor, Estimates of sperm whale abundance in the Northeastern Temperate Pacific from a combined acoustic and visual survey. Mar. Mamm. Sci. 21, 429–445 (2005)
Bioacoustics Research Program, Raven Pro: Interactive Sound Analysis Software (Version 1.1) [Computer software]. (The Cornell Lab of Ornithology, Ithaca, NY, 2003), http://www.birds.cornell.edu/raven
W. Bürck, P. Kotowski, H. Lichte, Die Horbarkeit von Laufzeitdifferenenzen. Elektrotechnol. Nachr. Technol. 12, 355 (1935)
A.M. Dougherty, Acoustic identification of individual sperm whales (Physeter macrocephalus), M.S. Thesis, University of Washington, 1999, 52 pages
A. Ekimov, J.M. Sabatier, Human motion analyses using footstep ultrasound and Doppler ultrasound. J. Acoust. Soc. Am. 123, EL149–EL153 (2008)
A. Ekimov, J.M. Sabatier, Rhythm analysis of orthogonal signals from human walking. J. Acoust. Soc. Am. 129, 1306–1314 (2011)
V. Estivill-Castro, Why so many clustering algorithms—a position paper. ACM SIGKDD Explor. Newsl. 4, 65–67 (2002). doi:10.1145/568574.568575
J.C. Goold, Signal processing techniques for acoustic measurement of sperm whale body lengths. J. Acoust. Soc. Am. 100, 3431–3441 (1996)
J.C. Goold, Behavioural and acoustic observations of sperm whales in Scapa Flow, Orkney Islands. J. Mar. Biol. Assoc. UK 79, 541–550 (1999)
J.C. Gordon, Evaluation of a method for determining the length of sperm whales from their vocalizations. J. Zool. 224, 301–314 (1991)
J. Gordon, R. Leaper, F.G. Hartley, O. Chappell, Effects of Whale-Watching Vessels on the Surface and Underwater Acoustic Behaviour of Sperm Whales of Kaikoura New Zealand, Science and Conservation Series, No. 52 (Department of Conservation, Wellington, 1992)
A. Growcott, B. Miller, P. Sirguey, E. Slooten, S. Dawson, Measuring body length of male sperm whales from their clicks: The relationship between inter-pulse intervals and photogrammetrically measured lengths. J. Acoust. Soc. Am 130, 568–573 (2011)
J.A. Hartigan, Clustering Algorithms (John Wiley & Sons, New York, NY, 1975)
J.W. Ioup, G.E. Ioup, Sperm whale identification using self organizing maps. J. Acoust. Soc. Am. 115, 2556 (2004)
J.W. Ioup, G.E. Ioup, Identifying individual sperm whales acoustically using self-organizing maps. J. Acoust. Soc. Am. 118, 2001 (2005)
G.E. Ioup, J.W.J. Ioup, P. Larue, N.A. Sidorovskaia, S.A. Kuczaj, G.H. Rayborn, C.D. Walker, Spectrogram analysis of low to mid frequency marine mammal clicks. J. Acoust. Soc. Am. 115, 2556 (2004)
G.E. Ioup, J.W. Ioup, N.A. Sidorovskaia, R.T. Walker, S.A. Kuczaj, C.D. Walker, G.H. Rayborn, B. Brack, A. Wright, J. Newcomb, R. Fisher, Analysis of Bottom-Moored hydrophone measurements of Gulf of Mexico sperm whale phonations. Proceedings: 23rd Ann Gulf of Mex Info Transf Meet. MMS 66, 109–136 (2005)
G.E. Ioup, J.W. Ioup, L.A. Pflug, A.M. Tashmukhambetov, N.A. Sidorovskaia, Acoustic identification of beaked and sperm whales. J. Acoust. Soc. Am. 122, 3003 (2007)
G.E. Ioup, J.W. Ioup, L.A. Pflug, A.M. Tashmukhambetov, N.A. Sidorovskaia, C.H. Thompson, C.O. Tiemann, Identifying individual clicking whales acoustically amid the clutter of other clicking whales, invited. J. Acoust. Soc. Am. 124, 2500 (2008)
G.E. Ioup, J.W. Ioup, L.A. Pflug, C.O. Tiemann, A. Bernstein, N.A. Sidorovskaia, Identifying individual clicking whales acoustically I. From click properties. J. Acoust. Soc. Am. 126, 2165 (2009)
G.E. Ioup, J.W. Ioup, L.A. Pflug, C.O. Tiemann, N.A. Sidorovskaia, Using click change detection to modify cluster analysis for acoustically identifying individual sperm whales with changing aspect. J Acoust Soc. Am 127, 2004 (2010a)
J.W. Ioup, G.E. Ioup, L.A. Pflug, N.A. Sidorovskaia, P. Schexnayder, C.O. Tiemann, A. Bernstein, Acoustic identification of individual beaked whales using self-organizing maps for automated classification of clicks in LADC data. J. Acoust. Soc. Am. 127, 2004 (2010b)
G.E. Ioup, J.W. Ioup, C.O. Tiemann, N.A. Sidorovskaia, Complementary techniques for robust acoustic identification of individual sperm and beaked whales. J. Acoust. Soc. Am. 129, 2536 (2011)
J.S. Jaffe, P.L.D. Roberts, Estimating fish orientation from broadband, limited-angle, multiview acoustic reflections. J. Acoust. Soc. Am. 129(2), 670–680 (2011)
N.L. Johnson, S. Kotz, A.W. Kemp, Univariate Discrete Distributions (Wiley Series in Probability and Statistics) (John Wiley & Sons, New York, NY, 1992), p. 565
D.R. Ketten, Functional analyses of whale ears: adaptations for underwater hearing. OCEANS’04: ‘Oceans Engineering for Today’s Technology and Tomorrow’s Preservation”. Proceedings, vol 1, 1994, p I/264–I/270
T. Kohonen, Self-Organization and Associative Memory, vol. 8 (Springer, New York, NY, 1989)
E.T. Küsel, D.K. Mellinger, L. Thomas, T.A. Marques, D.J. Moretti, J. Ward, Cetacean population density estimation from single fixed sensors using passive acoustics. J. Acoust. Soc. Am. 129, 3610–3622 (2011)
J. Larue, G.E. Ioup, J.W. Ioup, Detecting sperm whale clicks in the presence of ambient and shipping noise using higher order moments. J. Acoust. Soc. Am. 115, 2487 (2004)
S.P. Lloyd, Least squares quantization in PCM. IEEE Trans. Inform. Theor. 28, 129–137 (1982)
P.T. Madsen, Sperm whale sound production, Unpublished Ph.D. dissertation, University of Aarhus, Denmark, 2002
P.T. Madsen, M. Wahlberg, B. Møhl, Male sperm whale (Physeter macrocephalus) acoustics in a high-latitude habitat: Implications for echolocation and communication. Behav. Ecol. Sociobiol. 53, 31–41 (2002)
M. Marcoux, H. Whitehead, L. Rendell, Coda vocalizations recorded in breeding areas are almost entirely produced by mature female sperm whales (Physeter macrocephalus). Can. J. Zool. 84, 609–614 (2006)
T.A. Marques, L. Thomas, J. Ward, N. DiMarzio, P.L. Tyack, Estimating cetacean population density using fixed passive acoustic sensors: an example with Blainville’s beaked whales. J. Acoust. Soc. Am. 125, 1982–1994 (2009)
B.R. Mate, K.M. Stafford, D.K. Ljungblad, A change in sperm whale (Physeter macrocephalus) distribution correlated to seismic surveys in the Gulf of Mexico. J. Acoust. Soc. Am. 96, 3268–3269 (1994)
B. Møhl, E. Larsen, M. Amundin, Sperm whale size determination: outlines of an acoustic approach. FAO Fish Ser. 5, 327–332 (1981)
B. Møhl, M. Wahlberg, P.T. Madsen, A. Heerfordt, A. Lund, The monopulsed nature of sperm whale clicks. J. Acoust. Soc. Am. 114, 1143–1154 (2003)
K.E. Moore, W.A. Watkins, P.L. Tyack, Pattern similarity in shared codas from sperm whales (Physeter catodon). Mar. Mamm. Sci. 9, 1–9 (1993)
J. Newcomb, R. Fisher, A. Turgut, R. Field, G. Ioup, J. Ioup, G. Rayborn, S. Kuczaj, J. Caruthers, R. Goodman, N. Sidorovskaia, Modeling and Measuring the Acoustic Environment of the Gulf of Mexico. Proceedings of the Information Transfer Meeting Minerals Management Service, 2002, pp. 509–521
J. Newcomb, R. Fisher, A. Field, G. Rayborn, S. Kuczaj, G. Ioup, J. Ioup, A. Turgut, Measurements of ambient noise and sperm whale vocalizations in the Northern Gulf of Mexico using near bottom hydrophones. Proceedings of the MTS/IEEE OCEANS2002, 2002, pp. 1365–1371
J. Newcomb, R. Fisher, R. Field, A. Turgut, G. Ioup, J. Ioup, G. Rayborn, S. Kuczaj, J. Caruthers, R. Goodman, N. Sidorovskaia, Near-bottom hydrophone measurements of ambient noise and sperm whale vocalizations in the northern Gulf of Mexico. J. Acoust. Soc. Am. 111, 2371 (2002c)
J. Newcomb, R. Fisher, R. Field, G. Ioup, J. Ioup, J. Larue, G. Rayborn, S. Kuczaj, J. Caruthers, R. Goodman, C. Walker, R. Paulos, J. Ramos, R. Thames, S. Vinogradov, N. Sidorovskaia, I. Udovydchenkov, I. Rypina, and M. Snyder, Using Acoustic Buoys to Assess Ambient Noise and Sperm Whale Vocalizations, invited. Proceedings of the Information Transfer Meeting of the Minerals Management Service, 2004, pp. 269–276 and 578–588
K.J. Norris, G.W. Harvey, A theory for the function of the spermaceti organ of the sperm whale. NASA SP-262, 397–417 (1972)
E.M. Nosal, Methods for tracking multiple marine mammals with wide-baseline passive acoustic arrays. J. Acoust. Soc. Am. 134, 2383–2392 (2013a)
E.V. Nosal, Model-based marine mammal localization methods, in Detection Classification and Localization of Marine Mammal Using Passive Acoustics – 10 Years of Progress, ed. by O. Adam, F. Samaran (Dirac NGO, Paris, 2013b)
E.M. Nosal, L.N. Frazer, Track of a sperm whale from delays between direct and surface-reflected clicks. Appl. Acoust. 87, 1187–1201 (2006)
E.M. Nosal, L.N. Frazer, Sperm whale 3D track, swim orientation, beam pattern, and click levels observed on bottom-mounted hydrophones. J. Acoust. Soc. Am. 122, 1969–1978 (2007a)
E.M. Nosal, L.N. Frazer, Modified pair-wise spectrogram processing for localization of unknown broadband sources. IEEE J. Ocean Eng. 32, 721–728 (2007b)
E.M. Nosal, L.N. Frazer, Pair-wise spectrogram processing used to track a sperm whale. Can. Acoust. 36, 132–138 (2008)
A.S. Pandya, R.B. Macy, Pattern Recognition with Neural Networks in C++ (CRC Press, Boca Raton, FL, 1995)
G. Pavan, T.J. Hayward, J.F. Borsani, M. Priano, M. Manghi, C. Fossati, J. Gordon, Time patterns of sperm whale codas recorded in the Mediterranean Sea 1985–1986. J. Acoust. Soc. Am. 107, 3487–3495 (2000)
M.Q. Rhinelander, S.M. Dawson, Measuring sperm whales from their clicks: Stability of interpulse intervals and validation that they indicate whale length. J. Acoust. Soc. Am. 115, 1826–1831 (2004)
T.D. Rossing, F.R. Moore, P.A. Wheeler, The Science of Sound, 3rd edn. (Addison Wesley, Boston, MA, 2002). Figure 7.4
G.A.F. Seber, Multivariate Observations (John Wiley & Sons, Hoboken, NJ, 1984)
N.A. Sidorovskaia, P. Schexnayder, A. Ekimov, J. Sabatier, G.E. Ioup, J.W. Ioup, Rhythmic analysis of sperm whale broadband acoustic signals. J. Acoust. Soc. Am. 125, 2738 (2009a)
N.A. Sidorovskaia, P. Schexnayder, G.E. Ioup, J.W. Ioup, C.O. Tiemann, A. Bernstein, A. Ekimov, J. Sabatier, Identifying individual clicking whales acoustically II: from click rhythms. J. Acoust. Soc. Am. 126, 2165–2166 (2009b)
N. Sidorovskaia, P. Schexnayder, G. E. Ioup, J. W. Ioup, C. O. Tiemann, J. Sabatier, and A. Ekimov, Integrated passive acoustic approach for detection, localization, and identification of individual marine mammals as applied to workshop data, Proceedings of the 4th International Workshop on Detection, Classification and Localization of Marine Mammals Using Passive Acoustics (2009c).
N.A. Sidorovskaia, C.O. Tiemann, G. Ioup, J. Ioup, Integrating techniques for identifying individual sperm whales acoustically. J. Acoust. Soc. Am. 128, 2299–2300 (2010)
N. A. Sidorovskaia, G. E. Ioup, J. W. Ioup, and J. W. Caruthers, 2004, Acoustic propagation studies for sperm whale vocalization analysis during LADC experiments, High-Frequency Ocean Acoustics, M. B. Porter, M. Siderius, and W. A. Kuperman, editors, AIP Conference Proceedings, pp. 288–295
N.A. Sidorovskaia, I.A. Udovydchenkov, I.I. Rypina, G.E. Ioup, J.W. Ioup, J.W. Caruthers, J. Newcomb, R. Fisher, Propagation modeling for sperm whale acoustic clicks in the northern Gulf of Mexico. J. Acoust. Soc. Am. 115, 2556 (2004b)
N.A. Sidorovskaia, G.E. Ioup, J.W. Ioup, Studies of waveguide propagation effects on frequency properties of sperm whale communication signals. J. Acoust. Soc. Am. 116, 2614 (2004c)
N. A. Sidorovskaia, A. Tashmukhambetov, G. E. Ioup, J. W. Ioup, J. J. Newcomb, J. M. Stephens, C. D. Walker, and G. H. Rayborn, Littoral Acoustic Demonstration Center: Experimental and Modeling Capabilities for Assessment of the Environmental Impact of Acoustic Sources in the Gulf of Mexico, invited, Proceedings of the 9th World Multiconference on Systemics, Cybernetics and Informatics (WMSCI 2005), 2005, vol. IX, pp. 113–114
N.A. Sidorovskaia, B. Richards, G.E. Ioup, J.W. Ioup, Spectral identification of sperm whales from Littoral Acoustic Demonstration Center passive acoustic recordings. J. Acoust. Soc. Am. 118, 2001 (2005b)
N. Sidorovskaia, P. Schexnayder, G. E. Ioup, J. W. Ioup, C. O. Tiemann, J. Sabatier, and A. Ekimov, Integrated passive acoustic approach for detection, localization, and identification of individual marine mammals as applied to workshop data. Proceedings of the 4th International Workshop on Detection, Classification and Localization of Marine Mammals Using Passive Acoustics, 2009
Сидоровская, Н.А., Ioup, G.E., Ioup, J.W., Tashmukhambetov, A.M., Ekimov, A.E. (2011). Использование пассивных акустических методов для изучения влияния индустриальной деятельности человека на морскую мегафауну, [N.A. Sidorovskaia, G.E. Ioup, J.W. Ioup, A.M. Tashmukhambetov, A.E. Ekimov, Using passive acoustic methods to study anthropogenic noise impact on oceanic megafauna,] In Ocean acoustics, Proceedings of the XIII the L.M. Brekhovskikh’s conference, Москва, ГЕОС 2011 г., pp. 270–276 [In Russian with summary translated in English]
M. Snyder, P. Orlin, A. Schulte, J. Newcomb, Ambient noise analysis of underwater acoustic data. J. Acoust. Soc. Am. 113, 2320 (2003)
H. Spath, Cluster Dissection and Analysis: Theory, FORTRAN Programs, Examples (Halsted, New York, NY, 1985). Transl. J. Goldschmidt
J. Starkhammar, J. Nilsson, M. Amundin, S.A. Kuczaj, M. Almqvist, H.W. Persson, Separating overlapping click trains originating from multiple individuals in echolocation recordings. J. Acoust. Soc. Am. 129, 458–466 (2011)
A.M. Tashmukhambetov, G.E. Ioup, J.W. Ioup, N.A. Sidorovskaia, J.J. Newcomb, Three-dimensional seismic array characterization study: experiment and modeling. J Acoust. Soc. Am. 123, 4094–4108 (2008)
V. Teloni, W.M.Z. Zimmer, M. Wahlberg, P.T. Madsen, Consistent acoustic size estimation of sperm whales using clicks recorded from unknown aspects. J. Cet. Res. Manag. 9, 127–136 (2007)
A. Thode, D. K. Mellinger, S. Stienessen, A. Martinez, K. Mullin, Depth-dependent acoustic features of diving sperm whales (Physeter macrocephalus) in the Gulf of Mexico, J. Acoust. Soc. Am. 112, 308–321 (2002)
C.O. Tiemann, Three-dimensional single-hydrophone tracking of a sperm whale demonstrated using workshop data from the Bahamas. Can. Acoust. 36, 67–73 (2008)
C.O. Tiemann, M.B. Porter, Automated model-based localization of sperm whale clicks. Oceans Proc. 2, 821–827 (2003)
C.O. Tiemann, S.W. Martin, J.R. Mobley, Aerial and acoustic marine mammal detection and localization on navy ranges. IEEE J. Ocean Eng. 31, 107–119 (2006a)
C.O. Tiemann, A.M. Thode, J. Straley, V. O’Connell, K. Folkert, Three-dimensional localization of sperm whales using a single hydrophone. J. Acoust. Soc. Am. 120, 2355–2365 (2006b)
C.O. Tiemann, J.S. Jaffe, P.L.D. Roberts, N.A. Sidorovskaia, G.E. Ioup, J.W. Ioup, A. Ekimov, S. Lehman, Signal and image processing techniques as applied to animal bioacoustics problems. Acoust. Today 7, 35–43 (2011). Errata: Acoust. Today 7, 57
M. Van der Schaar, E. Delory, M. André, Classification of sperm whale clicks (Physeter Macrocephalus) with Gaussian-Kernel-based networks. Algorithms 2, 1232–1247 (2009)
G.T. Waring, E. Josephson, K. Maze-Foley, P.E. Rosel, U.S. Atlantic and Gulf of Mexico Marine Mammal Stock Assessments – 2009, in NOAA Technical Memorandum NMFS-NE-213, 2009, ed. by K. Barry, B. Byrd, T.V.N. Cole, C. Dias, L. Engleby, C. Fairfield, L.P. Garrison, A. Glass, L. Hansen, J. Litz, C. Orphanides, R.M. Pace, D.L. Palka, M. Rivera, M.C. Rossman, C. Sinclair, J. Valade, F.W. Wenzel (NOAA, Washington, DC, 2009), p. 528
W.A. Watkins, W.E. Schevill, Sperm whale codas. J. Acoust. Soc. Am. 66, 1485–1490 (1977a). plus phonograph record
W.A. Watkins, W.E. Schevill, Spatial distribution of Physeter catodon (sperm whales) underwater. Deep-Sea Res. 24, 693–699 (1977b)
S.L. Watwood, P.J.O. Miller, M. Johnson, P.T. Madsen, P.L. Tyack, Deep-diving foraging behaviour of sperm whales (Physeter macrocephalus). J. Anim. Ecol. 75, 814–825 (2006)
L. Weilgart, H. Whitehead, Coda communication by sperm whales (Physeter macrocephalus) off the Galápagos Islands. Can. J. Zool. 71, 744–752 (1993)
L. Weilgart, H. Whitehead, Group-specific dialects and geographical variation in coda repertoire in South Pacific sperm whales. Behav. Ecol. Sociobiol. 40, 277–285 (1997)
H. Whitehead, Sperm Whales: Social Evolution in the Ocean (University of Chicago Press, Chicago, 2003)
H. Whitehead, L. Weilgart, Click rates from sperm whales. J. Acoust. Soc. Am. 87, 1798–1806 (1990)
H. Whitehead, L. Weilgart, Patterns of visually observable behavior and vocalizations in groups of female sperm whales. Behaviour 118, 275–296 (1991)
B. Würsig, D.W. Weller, S.K. Lynn, J.G. Ortega-Ortiz, A.J. Schiro, T.D. Sparks, W.E. Evans, Behavior and site fidelity of sperm whales in the northern Gulf of Mexico, in Cetaceans, Sea Turtles and Seabirds in the Northern Gulf of Mexico: Distribution, Abundance and Habitat Associations, Vol II: Technical Report, OCS Study MMS, ed. by R.W. Davis, W.E. Evans, B. Würsig (Gulf of Mexico OCS Region Publications, Santa Barbara, CA, 1999)
B. Würsig, T.A. Jefferson, D.J. Schmidley, Marine Mammals of the Gulf of Mexico (Texas A & M University Press, College Station, TX, 2000)
J. Young, A. Host-Madsen, E.M. Nosal, Impulsive source separation with application to sperm whale clicks. Proceedings of the 2013 IEEE Digital Signal Processing workshop, Napa California, 2013
W.M.X. Zimmer, P.L. Tyack, M.P. Johnson, P.T. Madsen, Three-dimensional beam pattern of regular sperm whale clicks confirms bent-horn hypothesis. J. Acoust. Soc. Am. 117, 1473–1485 (2005a)
W.M.X. Zimmer, M.P. Johnson, P.T. Madsen, P.L. Tyack, Echolocation clicks of free-ranging Cuvier’s beaked whales (Ziphius cavirostris). J. Acoust. Soc. Am. 117, 3919–3927 (2005b)
W.M.X. Zimmer, P.T. Madsen, V. Teloni, M.P. Johnson, P.L. Tyack, Off-axis effects on the multipulse structure of sperm whale usual clicks with implications for sound production. J. Acoust. Soc. Am. 118, 3337–3345 (2005c)
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2016 Springer Science+Business Media New York
About this chapter
Cite this chapter
Ioup, G.E. et al. (2016). Environmental Acoustic Recording System (EARS) in the Gulf of Mexico. In: Au, W., Lammers, M. (eds) Listening in the Ocean. Modern Acoustics and Signal Processing. Springer, New York, NY. https://doi.org/10.1007/978-1-4939-3176-7_6
Download citation
DOI: https://doi.org/10.1007/978-1-4939-3176-7_6
Published:
Publisher Name: Springer, New York, NY
Print ISBN: 978-1-4939-3175-0
Online ISBN: 978-1-4939-3176-7
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)