
Abstract
Background
Numerous previous studies have investigated the effects of grazing or mowing on plant community diversity and productivity in grasslands; however, few have deliberately made sound comparison between the effects of paralleled grazing and mowing in terms of biomass removal on plant community diversity and productivity in semi-arid grasslands. Using a 4-year field manipulative experiment, we investigated how moderate intensity of domestic cattle (Bos taurus) grazing and mowing can affect plant community diversity and productivity in the semi-arid grassland in northeastern China, with the attempt to find a better management practice.
Results
Our results showed that grazing significantly increased plant species richness by 9% but did not change plant biomass, whereas mowing did not alter plant species richness but significantly reduced total plant biomass and root biomass by 18% and 12%, respectively, and significantly altered plant community composition, reflected by 32% increase of grass to forb biomass ratio.
Conclusions
Cattle grazing exerted a neutral effect on plant biomass and a positive effect on plant species richness, suggesting that cattle grazing is a better management practice compared to the paralleled mowing, but longer-term experiments are needed to explore the lasting influences of grazing vs. mowing on grassland productivity, plant diversity and the sustainability.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Introduction
Managements, like grazing and mowing, are needed to maintain plant community diversity and productivity in grassland ecosystems (Tälle et al. 2016; Henn and Damschen 2022; Hassan et al. 2023). Based on previous researches, the effect of stocking rates was quantified based on the changes in diversity and species richness of grassland ecosystems under diverse environmental conditions, as plant diversity and species richness would become more severely affected when stocking rate increased, while most the responses were uniform across environmental changes. For example, highly negative impacts were observed in low productivity system (McNaughton and Coughenur 1983; Loeser et al. 2004; Gebremedhn et al. 2023).
The effect of grazing on plant biomass may be negative (Song et al. 2013; Zhao et al. 2011; Cao et al. 2024), positive and neutral (Milchunas and Lauenroth 1993; McNaughton et al. 1989) However, this fluctuation under grazing management might help to describe shifts in species dominance in grasslands open to long-term livestock grazing (Loeser et al. 2004). Besides, plant species may respond to grazing by increasing or decreasing their relative biomass or reproductive fitness, which remains a debatable issue. These diverse effects of grazing on grassland biomass might depend on resource supply, herbivore type, management type, species pool size, plant dominance and spatial scale (Milchunas and Lauenroth 1988; Olff et al. 2002; Bai et al. 2012; Hassan et al. 2023).
Globally, mowing is an effective practice for grassland management, and mowing is similar to grazing in terms of its effect on biomass removal (Hassan et al. 2023). Like grazing, mowing has also been found to exert positive, negative or neutral effects on plant community diversity and productivity, structure and composition in grassland ecosystems (Tälle et al. 2015; Hejcman et al. 2011; Gillespie et al. 2022; Hassan et al. 2023). Mowing with low frequency may increase litter accumulation, which in turn delays seedling germination, while mowing with high frequency may suppress the seed set of some plant species, which will then decrease species richness and abundance and affect community composition (Milberg et al. 2017; Poschlod et al. 2011). However, by minimizing competition for resources, regular mowing could also increase seed establishment which in turn elevated species richness and biological diversity (Hansson and Fogelfors 2000; Smith et al. 2018). On other occasions, plant diversity was negatively affected due to lowered germination and retarded seed bank development on account of grassland fragmentation as consequences of trespass and mechanical disturbance (Valkó et al. 2012; Socher et al. 2012).
Mowing can negatively affect plant biomass both above- and below-ground through the alteration in vegetation structure and soil nutrient pool in grassland ecosystems (Billeter et al. 2007; Huhta et al. 2001; Bonanomi et al. 2013; Li et al. 2022). Regular mowing with high frequency reduces biomass and plant litter accumulation and thus keeps grassland open and decreases organic nutrient transport to the soil system (Shao et al. 2012; Reisch and Poschlod 2009; Mao et al. 2023). However, long-term mowing degraded natural grassland and led to worse community properties and structure by affecting many community processes such as plant compensatory growth (Liu et al. 2007). Generally under mowing, the aboveground biomass of forbs was decreased to a much larger extent compared to that of dominant grasses in grassland ecosystems (Hejcman et al. 2011; Billeter et al. 2007). Moreover, the removal of apical buds due to mowing may inhibit flowering, fruiting and seeding to a larger extent for forb species than for grasses, leading to alteration of community composition, reflected by the decrease of forb to grass ratio (Valkó et al. 2012). These diverse effects of mowing might be due to the variation of several abiotic and biotic factors such as mowing intensity, species pool, mowing timing, soil condition, litter accumulation, climate, light intensity and so on (Tälle et al. 2016; Socher et al. 2012; Huhta et al. 2001; Schmid et al. 2022; Hassan et al. 2023).
Most of the previous studies have been focused on the effect of either mowing or grazing on grassland vegetation (Bai et al. 2012; Wang et al. 2019; Hassan et al. 2023), and very few have compared the effects of paralleled mowing and grazing in terms of biomass removal, therefore there is still ambiguity in selecting grassland management practice. The purpose of this study was to investigate how livestock grazing and mowing affect plant communities in grassland ecosystems and to choose an optimum management practice. We produced the following three hypotheses. First, grazing can increase species richness and biomass, whereas paralleled mowing can increase plant density but decrease biomass. Second, grasses have higher plasticity than forbs in terms of biomass reversibility in response to grazing or mowing. Third, annual grazing with moderate intensity can stabilize grassland plant communities and thus can be a sustainable management practices in northeastern China.
Materials and methods
Study area
The study was carried out in a semi-arid meadow steppe at Grassland Ecological Research Station of Northeast Normal University, Jilin Province, China (44° 45′ N, 123° 47′ E). It is 292 m above sea level. The climate of the locality is semi-arid continental, with an annual mean temperature ranging from 4.6 to 6.5 °C, whereas the summer temperature was recorded from 25 to 38 °C in July and August and –29 °C in January with annual precipitation from 280 to 400 mm. The soil is mixed salt-alkali (Salid Aridisol, US Soil Taxonomy) composed of 40% silt, 29% sand, and 31% clay. The soil is nutrient-poor in total nitrogen content ranging from 2.2 to 2.5 mg g−1, and total phosphorous content from 0.23 to 0.27 mg g−1. The dominant plant species is Leymus chinensis (a perennial grass), accounting for ≥ 60% of the total annual aboveground biomass. The other grass species include Phragmites australis, Calamagrostis epigeios, Setaria viridis, Chloris virgata and the forbs include Artemisia scoparia, Artemisia anethifolia, Kalimeris integrtifolia, and Messerschmidias ibirica. The study area has a long history of grazing and mowing, but our study site was fenced in 2005 to shield against uncontrolled anthropogenic disturbances.
Experimental design and treatment
We established a gazing and mowing experiment to test the different effects of paralleled grazing and mowing on plant community diversity and productivity from 2016 to 2019 (Fig. 1). Within the study site comparatively flat land area with uniform soil conditions was selected as our experimental arena and was fenced in 2016. In June 2016, we randomly established six 400 m × 150 m blocks. The distance between every two adjacent blocks was kept 100–250 m apart. Each block was comprised of three sub-enclosure plots of 100 m × 100 m size, which were randomly assigned to the three treatments including control, domestic cattle grazing and mowing. Adjacent plots were at least 50 m apart. The plots were grazed by cattle and mowed with hay mower at moderate intensity.

The experimental layout
Grazing treatment was performed from 6:00 am to 8:00 am and from 4:00 pm to 6:00 pm every day during the first two weeks of each month from June to September each year, which mimicked local grazing habits. Mowing treatment was started in early August, the peak of the growing season each year. The plant material higher than 20 cm was removed with hay mowers, which is about equivalent to the grazing treatment in terms of biomass removal.
Plant and litter sampling
Plant and litter sampling were carried out in mid-August each year. We surveyed plant diversity, biomass of each plant species, total plant biomass and plant litter biomass. We set up two parallel transects at a distance of 30 m within each plot, and within each of them, we set up five 1 m × 1 m quadrats, every adjacent two of which were 15 m apart. We measured species richness by visually counting the number of plant species, and clipped all the plant material to ground level in the five 1 m × 1 m quadrats within each transect (Fig. 1). We also collected plant litter in the same locations. Aboveground plant material and litter were then dried for 48 h at 65 °C and weighed to estimate aboveground plant biomass (Bai et al. 2012; Hassan et al. 2023).
Statistical analyses
All statistical analyses were performed in R 3.1.0 (R Development Core Team 2014). To study the effects of grazing and mowing on plant community and diversity, we used linear mixed effect models with “treatment” (grazing, mowing and control) included as the fixed effects and “replicate block” included as the random effect. The specific response variables assessed were: plant diversity, density, root biomass and shoot biomass. We also used the same method to assess the effects of grazing on grass and forb biomass. Models were fitted using the function lmer from the package lme 4 and the package lmer Test was used to calculate P-values (Kuznetsova et al. 2017). Prior to the above analyses, all the data were tested for homogeneity of residual variances, using Levene’s test. If needed, data were normalized by arcsine square root transformations.
Results
Cattle grazing and mowing did not significantly affect plant species evenness (F2,15 = 1.08, P = 0.720; Fig. 2a), but significantly affect plant species richness (F2,15 = 25.33, P = 0.026; Fig. 2b). Species richness in grazed plots was higher than that in the mowed plots (P = 0.034), but was not different from that in control plots (P = 0.361). Compared to grazing, mowing significantly decreased total biomass (P = 0.047; Fig. 3a) and root biomass (P = 0.037, Fig. 3c). However, shoot biomass did not show any significant responses to grazing or mowing (Fig. 3b). Grazing significantly decreased shoot:root biomass ratio (P = 0.037, Fig. 3d), but mowing did not. Cattle grazing and mowing did not affect grass biomass (F2,15 = 2.13, P = 0.256; Fig. 3e), but significantly altered forb biomass (F2,15 = 29.77, P = 0.042; Fig. 3f). Mowing significantly decreased forb biomass (P = 0.019) and thus increased grass:forb biomass ratio (P = 0.011) (Fig. 3g).

Effects of grazing and mowing on plant species evenness (a) and species richness (b). Letters “a” and “b” indicate a significant difference between treatments (P < 0.05)

Effects of grazing and mowing on plant total biomass (a), shoot biomass (b), root biomass (c), shoot:root biomass ratio (d), grass biomass (e), forb biomass (f), grass:forb biomass ratio (g) and litter mass (h). Letters “a” and “b” indicate a significant difference between treatments (P < 0.05)
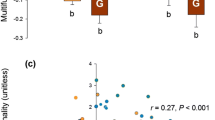
Both cattle grazing and mowing significantly decreased litter biomass (F2,15 = 96.23, P = 0.002; Fig. 3h). Litter biomass in mowed and grazed plots were not different from each other (P = 0.658, Fig. 3h), but were significantly lower than that in the control plots (P = 0.001). Regardless of experimental treatments, there was no significant relationship between species richness and shoot biomass (F1,16 = 0.782, P = 0.146, Fig. 4a), root biomass (F1,16 = 6.268, P = 0.369, Fig. 4b), or total biomass (F1,16 = 1.36, P = 0.694, Fig. 4c).

Effect of grazing and mowing on the relationship of shoot biomass (a), root biomass (b) and total biomass (c) with species richness. In the panels, the open, green filled and orange filled circles denote control, grazing and mowing treatments, respectively
Mowing significantly increased grass (P = 0.021) and overall (P = 0.007) plant density while decreased forb density (P = 0.001), but grazing did not (P > 0.05; Fig. 5a–c). The density of both grass and forb and the community in the grazed plots was halfway between but did not differ significantly from that in the control or mowed plots. Grass:forb density ratio in mowed plots was significantly higher than in the control and grazed plots (P = 0.005, Fig. 5f). Both grass (P = 0.003) and forb (P = 0.009) height in mowed plots was significantly lower than the control plots but did not differ from that in the grazed plots (P > 0.05, Fig. 5d, e).

Effect of grazing and mowing on total density (a), grass density (b), forb density (c), grass height (d), forb height (e) and grass:forb density ratio (f). Letters “a” and “b” indicate a significant difference between treatments (P < 0.05)
Discussion
Unlike mowing, grazing was found to significantly increase plant species richness (Fig. 2a, b), which is in line with our hypothesis and also supporting previous studies claiming that grazing has positive effects on plant species richness (Pykälä 2005; Anderson and Minor 2020; Gavrichkova et al. 2022). This grazing-induced elevation of plant species richness was assumed to be attributed to several mechanisms. First, as compared to mowing, grazing increased the soil nutrient pool, through the addition of N and P in the form of faces and urine into the soil, hence the subordinate species would usually be facilitated disproportionally more than the dominant species (Hassan et al. 2021; Wang et al. 2019). Second, trampling and wallowing by livestock enhanced litter disintegration and decomposition, thus leading to enriched soil, which would facilitate subordinate species to a larger extent than dominant species as the latter were inhibited more severely by livestock (Risch et al. 2015). Third, more subordinate species would be fostered under the competitive release from the dominant species, L. chinensis in our case, which was preferentially foraged by livestock (Anderson and Minor 2020; Hassan et al. 2021). In contrast to grazing, mowing decreased species richness, probably because mowing increased the openness of vegetation, thereby increasing soil temperature and decreasing soil moisture. Consequently, seed germination and bud outgrowth were retarded, ultimately leading to a reduction of species richness (Méndez and Karlsson 2005; Ågren 2008; Pykälä 2005; Karami et al. 2021; Hassan et al. 2024).
In our study, mowing significantly decreased total biomass and root biomass whereas grazing significantly lowered shoot:root biomass ratio (Fig. 3a–d). First, along with above-ground biomass removal, mowing also reduced the accumulation of plant litter, which would disturb the balance of soil nutrients between soil and organic matter. Second, mowing shrank soil seed banks, delayed seed development and decreased plant species richness (Ziter and MacDougall 2013; Yang et al. 2020; Kotas et al. 2017). Third, mowing decreased the production of photosynthate by removing plant photosynthetic organs (Zhao et al. 2023). All these three mechanisms could account for the reduction of plant community biomass under mowing. On the other hand, mowing slightly but insignificantly decreased shoot biomass, probably the negative effects on shoot biomass were offset by the increase in stem density.
Grazing did not affect shoot, root or total biomass, probably because the possible negative effects of such short-term grazing with moderate intensity were offset by the fertilizing effect of cattle faeces and urine. Generally, an increase in grazing duration and intensity might significantly affect the biomass. However, grazing during the non-growing season may significantly increase aboveground biomass compared to that during the growing season. These findings may have implications for grassland management especially from perspective of seasonal pattern of grazing practice, i.e., grazing duration should not be concentrated on the growing season, but could be lengthened to cover the entire or at least a major part of non-growing season (Hao and He 2019). On the other hand, the decreased shoot:root ratio might be the consequence of shoot biomass removal by cattle and also of the adaptation of plants to grazing pressure.
It is commonly believed that both grazing and mowing differentially affect forb and grass biomass and thus their ratios (Liu et al. 2018; Fynn et al. 2004; Hassan et al. 2023). In our study, mowing significantly decreased forb biomass and thereby increased the grass:forb biomass ratio (Fig. 3f, g). Indeed, in most cases, mowing was found to suppress the growth of forbs more than of grasses, hence altering their ratio (Valkó et al. 2012; Billeter et al. 2007; Hejcman et al. 2011). Probably, grasses have basal meristems, which may be less susceptible to mowing compared with aerial meristems of forbs (Briske and Richards 1994; Adhikari and Russell 2014). In contrast, the removal of apical meristems may prevent forbs from flowering, fruiting, and seed setting, ultimately leading to a decrease in forb biomass (Hejcman et al. 2011; Valkó et al. 2012).
Compared to grazing, mowing was found to significantly increase plant total density, grass density and grass:forb density ratio while decreasing forb density, grass and forb height (Fig. 5a–f), which is consistent with previous studies (Gibson 2009; Davies et al. 2012; Deng et al. 2014; Karami et al. 2021). The increase in grass density might be attributed to the fact that underground rhizomes of rhizomatous grasses such as L. chinensis, the predominant species, were not easily affected by mowing (Briske and Richards 1994) and even the bud outgrowth and tillering would be stimulated by shoot injury under mowing (Wang et al. 2004). However, forbs were inert relative to grasses in terms of being stimulated by mowing, and would thus further be suppressed more in response to denser grasses. In contrast, despite that cattle grazing has a similar effect by feeding on grasses and forbs, it would exert greater grazing pressure on grasses than on forbs due to their preference for grasses, i.e., selective foraging for grasses (Fynn et al. 2004; Díaz et al. 2007; Deng et al. 2014). In addition, cattle trampling might have created gaps for seedlings of forbs rather than of grasses, which more relied on clonal propagation. Taken together, it is understandable that, unlike mowing, grazing has more complex and even antagonistic effects on the growth and propagation of offspring plants for grasses and forbs, thus leading to little difference in its impact on such many characteristic features of both plant functional types.
Since grazing and mowing decreased above-ground biomass, they decreased litter production (Fig. 3h), which consolidates previous findings that management practices such as grazing and mowing have negative impacts on litter biomass (Shao et al. 2012; O’Neill et al. 2003; Zhang et al. 2022). However, the effect on litter biomass may vary with grazing and mowing intensity, timing and soil type (Grime 1973). Unexpectedly, we failed to find a significant relationship between shoot, root and total biomass with species richness (Fig. 4a–c). This failure might be attributed to the moderate intensity with short duration of grazing and mowing treatment and likely also to the absence of interspecific compensatory effects due to low species richness in the study area (Beltman et al. 2003).
Conclusions
This study suggests that, compared to mowing, cattle grazing is a better management practice in the semi-arid grassland in northeastern China, on condition that they are equivalent in terms of biomass removal. Annual grazing of moderate intensity can maintain the stability of grassland plant community and thus can be a sustainable management practice. Therefore, grazing was a better management practice than mowing. There is still ambiguity that, how it will persist in the long run, particularly under climate change. Despite that, our study provides baseline information from the locality, which can help researchers step further and inform herdsmen to adjust their strategy of grassland use. Taken together, the present study was a short-term one with moderate intensity of grazing and mowing, and longer-term experiments are needed to explore the influences of grazing vs. mowing on grassland productivity, plant diversity and sustainability.
Availability of data and materials
The data that support the findings of this study are available from the corresponding author upon reasonable request.
References
Adhikari S, Russell FL (2014) Effects of apical meristem mining on plant fitness, architecture, and flowering phenology in Cirsium altissimum (Asteraceae). Am J Bot 101:2079–2087
Ågren GI (2008) Stoichiometry and nutrition of plant growth in natural communities. Annu Rev Ecol Evol Syst 39:153–170
Anderson EC, Minor ES (2020) Management effects on plant community and functional assemblages in Chicago’s vacant lots. Appl Veg Sci 23:266–276
Bai Y, Wu J, Clark CM, Pan Q, Zhang L, Chen S (2012) Grazing alters ecosystem functioning and C:N:P stoichiometry of grasslands along a regional precipitation gradient. J Appl Ecol 49:1204–1215
Beltman B, Van Den Broek T, Martin W, Ten Cate M, Güsewell S (2003) Impact of mowing regime on species richness and biomass of a limestone hay meadow in Ireland. Bull Geobot Inst ETH 69:17–30
Billeter R, Peintinger M, Diemer M (2007) Restoration of montane fen meadows by mowing remains possible after 4–35 years of abandonment. Bot Helv 117:1–13
Bonanomi G, Incerti G, Allegrezza M (2013) Assessing the impact of land abandonment, nitrogen enrichment and fairy-ring fungi on plant diversity of Mediterranean grasslands. Biodivers Conserv 22:2285–2304
Briske D, Richards JH (1994) Physiological responses of individual plants to grazing: current status and ecological significance. In: Vavra M, Laycock WA, Pieper RD, eds. Ecological implications of herbivory in the west. Society for Range Management, Denver, CO. pp 147–176
Cao F, Li W, JiangY GX, Zhao C, Ma J (2024) Effects of grazing on grassland biomass and biodiversity: a global synthesis. Field Crops Res 306:109204
Davies K, Bates J, Nafus A (2012) Vegetation response to mowing dense mountain big sagebrush stands. Rangel Ecol Manag 65:268–276
Deng L, Sweeney S, Shangguan ZP (2014) Grassland responses to grazing disturbance: plant diversity changes with grazing intensity in a desert steppe. Grass Forage Sci 69:524–533
Díaz S, Lavorel S, McIntyre S, Falczuk V, Casanoves F, Milchunas DG (2007) Plant trait responses to grazing: a global synthesis. Glob Change Biol 13:313–341
Fynn RW, Morris CD, Edwards TJ (2004) Effect of burning and mowing on grass and forb diversity in a long-term grassland experiment. Appl Veg Sci 7:1–10
Gavrichkova O, Pretto G, Brugnoli E, Chiti T, Ivashchenko KV, Mattioni M (2022) Consequences of grazing cessation for soil environment and vegetation in a subalpine grassland ecosystem. Plants 11:2121
Gebremedhn HH, Ndiaye O, Mensah S, Fassinou C, Taugourdeau S, Tagesson T, Salgado P (2023) Grazing effects on vegetation dynamics in the savannah ecosystems of the Sahel. Ecol Process 12:54
Gibson DJ (2009) Grasses and grassland ecology. Oxford University Press, New York
Gillespie MA, Buckley HL, Condron L, Wratten SD (2022) Grassland plant and invertebrate species richness increases from mowing are mediated by impacts on soil chemistry. Basic Appl Ecol 63:152–163
Grime J (1973) Control of species density in herbaceous vegetation. J Environ Manage 1:151–167
Hansson M, Fogelfors H (2000) Management of a semi-natural grassland: results from a 15-year-old experiment in southern Sweden. J Veg Sci 11:31–38
Hao Y, He Z (2019) Effects of grazing patterns on grassland biomass and soil environments in China: a meta-analysis. PLoS ONE 14:e0215223
Hassan N, Li X, Wang J, Zhu H, Nummi P, Wang D (2021) Effects of grazing on C:N:P stoichiometry attenuate from soils to plants and insect herbivores in a semi-arid grassland. Oecologia 195:785–795
Hassan N, Zhong Z, Wang D, Zhu Y, Naeem I, Ahungu AB (2023) Effects of long-term mowing on species diversity, biomass and composition of plant community in a semi-arid grassland in northeastern China. Appl Veg Sci 26:e12743
Hassan N, Abdullah I, Khan W, Khan A, Ahmad N, Iqbal B et al (2024) Effect of grazing and mowing on soil physiochemical properties in a semi-arid grassland of Northeast China. Pol J Environ Stud 33:1725–1735
Hejcman M, Pavlů SJ (2011) Competitive ability of Rhinanthus minor L. in relation to productivityin the Rengen Grassland Experiment. Plant Soil Environ 57:45–51
Henn JJ, Damschen EI (2022) Grassland management actions influence soil conditions and plant community responses to winter climate change. Ecosphere 13:e4270
Huhta AP, Rautio P, Tuomi J, Laine K (2001) Restorative mowing on an abandoned semi-natural meadow: short-term and predicted long-term effects. J Veg Sci 12:677–686
Karami P, Bandak I, Karaji MG, Dragovich D (2021) Effects of seasonal grazing and annual mowing on floristic composition and plant diversity in the Saral rangeland, Kurdistan, Iran. Glob Ecol Conserv 27:e01515
Kotas P, Choma M, Šantrůčková H, Lepš J, Tříska J, Kaštovská E (2017) Linking above- and belowground responses to 16 years of fertilization, mowing, and removal of the dominant species in a temperate grassland. Ecosystems 20:354–367
Kuznetsova A, Brockhoff PB, Christensen RHB (2017) lmerTest package: tests in linear mixed effects models. J Stat Softw 82:1–26
Li L, Liu H, Baoyin T (2022) Mowing increases root-to-shoot ratio but decreases soil organic carbon storage and microbial biomass C in a semiarid grassland of North China. Agriculture 12:1324
Liu Z, Baoyin T, Sun J, Minggagud H, Li X (2018) Plant sizes mediate mowing-induced changes in nutrient stoichiometry and allocation of a perennial grass in semi-arid grassland. Evol Ecol 8:3109–3118
Liu M, Baoyin T, Yang C, Zhang X (2007) Effect of mowing systems on composition of Stipa grandis community in Inner Mongolia steppe. J Beijing Norm Univ Nat Sci 43:83–87
Loeser MR, Crews TE, Sisk TD (2004) Defoliation increased above-ground productivity in a semi-arid grassland. Rangel Ecol Manag 57:442–447
Mao Z, Wang Y, Li Q, Li W, Wang H, Li Y, Yue M (2023) Deep mowing rather than fire restrains grassland Miscanthus growth via affecting soil nutrient loss and microbial community redistribution. Front Plant Sci 13:1105718
McNaughton WLL, Coughenour MB (1983) Plant adaptation in an ecosystem context: effects of defoliation, nitrogen, and water on growth of an African C4 sedge. Ecology 64:307–318
McNaughton OM, Frank DA, Williams K (1989) Ecosystem-level patterns of primary productivity and herbivory in terrestrial habitats. Nature 341:142–144
Méndez M, Karlsson PS (2005) Nutrient stoichiometry in Pinguicula vulgaris: nutrient availability, plant size, and reproductive status. Ecology 86:982–991
Milberg P, Tälle M, Fogelfors H, Westerberg L (2017) The biodiversity cost of reducing management intensity in species-rich grasslands: mowing annually vs. every third year. Basic Appl Ecol 22:61–74
Milchunas DG, Lauenroth WK (1993) Quantitative effects of grazing on vegetation and soils over a global range of environments. Ecol Monogr 63:327–366
Milchunas SOE, Lauenroth WK (1988) A generalized model of the effects of grazing by large herbivores on grassland community structure. Am Nat 132:87–106
O’Neill KM, Olson BE, Rolston MG, Wallander R, Larson DP, Seibert CE (2003) Effects of livestock grazing on rangeland grasshopper (Orthoptera: Acrididae) abundance. Agric Ecosyst Environ 97:51–64
Olff H, Ritchie ME, Prins HH (2002) Global environmental controls of diversity in large herbivores. Nature 415:901–904
Poschlod P, Hoffmann J, Bernhardt-Römermann M (2011) Effect of grassland management on the age and reproduction structure of Helianthemum nummularium and Lotus corniculatus populations. Preslia 83:421–435
Pykälä J (2005) Cattle grazing increases plant species richness of most species trait groups in mesic semi-natural grasslands. Plant Ecol 175:217–226
Reisch C, Poschlod P (2009) Land use affects flowering time: seasonal and genetic differentiation in the grassland plant Scabiosa columbaria. Evol Ecol 23:753–764
Risch AC, Schütz M, Vandegehuchte ML, van der Putten WH, Duyts H, Raschein U (2015) Aboveground vertebrate and invertebrate herbivore impact on net N mineralization in subalpine grasslands. Ecology 96:3312–3322
Schmid JS, Huth A, Taubert F (2022) Impact of mowing frequency and temperature on the production of temperate grasslands: explanations received by an individual-based model. Oikos 2022:e09108
Shao C, Chen J, Li L, Zhang L (2012) Ecosystem responses to mowing manipulations in an arid Inner Mongolia steppe: an energy perspective. J Arid Environ 82:1–10
Smith AL, Barrett RL, Milner RN (2018) Annual mowing maintains plant diversity in threatened temperate grasslands. Appl Veg Sci 21:207–218
Socher SA, Prati D, Boch S, Muller J, KlausVH HN (2012) Direct and productivity-mediated indirect effects of fertilization, mowing and grazing on grassland species richness. J Ecol 100:1391–1399
Song Y, Zhou D, Zhang H, Li G, Jin Y, Li Q (2013) Effects of vegetation height and density on soil temperature variations. Chin Sci Bull 58:907–912
Tälle M, Fogelfors H, Westerberg L, Milberg P (2015) The conservation benefit of mowing vs grazing for management of species-rich grasslands: a multi-site, multi-year field experiment. Nord J Bot 33:761–768
Tälle M, Deák B, Poschlod P, Valkó O, Westerberg L, Milberg P (2016) Grazing vs. mowing: a meta-analysis of biodiversity benefits for grassland management. Agr Ecoyst Environ 222:200–212
Valkó O, Török P, Matus G, Tóthmérész B (2012) Is regular mowing the most appropriate and cost-effective management maintaining diversity and biomass of target forbs in mountain hay meadows? Funct Ecol 207:303–309
Wang ZW, Li LH, Han XG, Dong M (2004) Do rhizome severing and shoot defoliation affect clonal growth of Leymus chinensis at ramet population level? Acta Oecol 26:255–260
Wang L, Delgado-Baquerizo M, Wang D, Isbell F, Liu J, Feng C (2019) Diversifying livestock promotes multidiversity and multifunctionality in managed grasslands. Proc Nat Acad Sci 116:6187–6192
Yang Z, Minggagud H, Baoyin T, Li FY (2020) Plant production decreases whereas nutrients concentration increases in response to the decrease of mowing stubble height. J Environ Manag 253:109745
Zhang S, Wei Y, Liu N, Wang Y, Manlike A, Zhang Y (2022) Mowing facilitated shoot and root litter decomposition compared with grazing. Plants 11:846
Zhao Y, Peth S, Reszkowska A, Gan L, Krümmelbein J, Peng X (2011) Response of soil moisture and temperature to grazing intensity in a Leymus chinensis steppe, Inner Mongolia. Plant Soil 340:89–102
Zhao T, Zhang F, Suo R, Zhen J, Qiao J, Zhao M, Bai K, Zhang B (2023) The importance of functional diversity in regulating forage biomass and nutrition: evidence from mowing in semiarid grasslands. Restor Ecol 31:e13742
Ziter C, MacDougall AS (2013) Nutrients and defoliation increase soil carbon inputs in grassland. Ecology 94:106–116
Acknowledgements
Two anonymous reviewers are appreciated for their insightful comments on our manuscript.
Funding
This project was supported by the National Natural Science Foundation of China (No. 32061143027) and the National Key Research and Development Program of China (No. 2022YFF1300600), and the Program for Introducing Talents to Universities (B16011), the Natural Science Foundation of Jilin Province Science and Technology Department (20220101283JC) and NSFC grant of China (42230515).
Author information
Authors and Affiliations
Contributions
N.H. designed the research work and performed experiments, N.H. and Z.W. analyzed the data and wrote the manuscript; Z.W. reframed the structure of the earlier version of the manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Hassan, N., Wang, Z. Paralleled grazing and mowing differentially affected plant community diversity and productivity in a semi-arid grassland. Ecol Process 13, 62 (2024). https://doi.org/10.1186/s13717-024-00543-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13717-024-00543-3