
Abstract
Background
Western Ghats is a biodiversity treasure trove with reports of indigenous leishmaniasis cases. Hence, systematic sand fly surveillance was carried out among the tribal population. The present study reports a novel sand fly species, Phlebotomus (Anaphlebotomus) ajithii n. sp. (Diptera: Psychodidae), discovered in the Western Ghats of India.
Methods
A comprehensive sand fly survey was conducted across the Kollam, Thrissur, Idukki, Kasaragod and Malappuram districts of Kerala, India. The survey spanned both indoor and outdoor habitats using standard collection methods over a 3-year, 3-month period. DNA barcoding of samples was performed targeting mitochondrial cytochrome c oxidase subunit I (COI) gene, and the sequence generated was subjected to phylogenetic analysis.
Results
Phlebotomus (Anaphlebotomus) ajithii, a new sand fly species, is recorded and described in this communication. The morphological relationship of the new species to other members of the subgenus Anaphlebotomus is discussed. Mitochondrial COI barcode followed by phylogenetic analysis confirmed that specimens of Ph. ajithii belong to the same taxonomic group, while a genetic distance of 11.7% from congeners established it as a distinct species.
Conclusions
The Western Ghats, known for its rich biodiversity, has lacked systematic entomological surveys focusing on sand flies. This study aims to fill this gap and reports and describes a new species of sand fly.
Graphical abstract

Similar content being viewed by others


Background
Phlebotomine sand flies are classified within the Psychodidae family under the order Diptera, and they are diminutive, hairy insects that feed on blood. These insects are of immense public health importance since they play a role as vectors of leishmaniasis. Leishmaniasis, a neglected tropical disease (NTD), is caused by a flagellate protozoan parasite from the Leishmania genus. It has been endemic in many countries like India, Brazil, Sudan, Bangladesh, Ethiopia and Nepal for centuries [1].
The Western Ghats, also known as the Sahyadri, is a biological treasure trove that boasts a rich biodiversity comprising numerous endemic species of plants, fishes, amphibians, reptiles and mammals [2, 3]. In India, the Western Ghats expands from Gujarat to Tamil Nadu with an area of 160,000 km2 and is considered one of 34 biodiversity hotspots worldwide [4]. Several tribal populations belonging to different ethnic groups are inhabitants of this zone. Human inhabitation and the practice of agriculture have made several alterations in the ecology and landscape of the Western Ghats [5]. In addition, this region is also relevant in terms of different vector-borne diseases, transmitted by insects, ticks, mites, etc. [6,7,8].
Western Ghats is emerging as an endemic belt for visceral leishmaniasis (VL) and cutaneous leishmaniasis (CL) in the last few decades, with many new indigenous case records [9]. Over the last 2 decades, nearly 50 cases of CL and VL were reported from the tribal population of these zones [10, 11]. Sand flies are the only proven vector of the Leishmania parasite involved in the transmission of leishmaniasis. The availability of resting and breeding habitats and the many blood meals via various hosts have aided in the abundant survival of sand flies in this region [12]. From 1990 till 2022, many new species and country records of sand flies have been documented from the Western Ghats through various entomological survey carried out by a few researchers [13,14,15,16,17,18,19,20,21,22]. Later, all these species were combined and included in a review article with a total of the 69 sand fly species recorded in India; almost 50% those species are found in Kerala [23]. Hence, considering the rich biodiversity and endemicity of the disease, systematic sand fly surveillance has been carried out across the Western Ghats of Kerala, especially in the tribal belts. A new species of sand fly was identified during this survey, Phlebotomus (Anaphlebotomus) ajithii n. sp. Taxonomic and phylogenetic characterizations of the species are documented in this article.
Methods
Study area
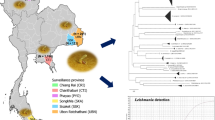
The entomological investigation was conducted within the various tribal communities situated in Western Ghats of Kollam, Thrissur, Idukki, Kasaragod and Malappuram districts in Kerala from January 2021–March 2024 (Table 1 and Fig. 1).

Sand fly specimen collection area in Thiruvananthapuram, Kollam, Thrissur, Idukki, Kasaragod and Malappuram districts of Kerala, India, along with species composition
These tribal settlements are scattered throughout the mountainous forest expanse of the Western Ghats in southern India. Referred to locally as the Sahyadri, this elevated area in Kerala lies on the periphery of the Deccan Plateau, demarcating it from the coastline strip along the Arabian Ocean. The Ghats boasts diverse vegetation, encompassing grasslands, arid and humid temperate forests, evergreen and semi-evergreen woodlands, scrub jungles and more. Coconut, rubber, pepper, jackfruit and teak plantations within these forests serve as major sources of livelihood for the indigenous tribes. The rich cultural heritage of the region is safeguarded by its intricate topography and abundant rainfall. Moreover, the forests within these ranges are designated as reserve forests and are patrolled by forest rangers appointed by the Kerala Government.
The tribes residing within these mountain ranges now inhabit dwellings constructed of concrete blocks, a measure introduced by the Kerala Government as part of an initiative aimed at the development of the tribal population. Reliant on the forest for their livelihood, these tribes often migrate to the remote reaches of the forest. Consequently, the abandoned houses, left vacant amid the moisture and humidity brought by the rains, create an ideal microhabitat for the breeding of sand flies. Within these settlements, houses are spaced roughly 100–200 m apart, totaling around 50–100 dwellings. Sand fly collection was conducted using standard entomological collection methods such as mechanical aspirators, light and sticky traps both indoors (cattle sheds and human dwellings) and outdoors (rodent burrows, tree holes, termite mounts etc.). On the other hand, resting collections employing mechanical aspirators were carried out in the dawn hours, between 9:00 a.m. and 12:00 noon. Trap collections, on the other hand, were undertaken from 6:00 p.m. (the preceding day) to 6:00 a.m. (the following day), covering the evening and night hours.
Morphological identification
Sand fly specimens were brought to Indian Council of Medical Research-Vector Control Research Centre (ICMR-VCRC), Field Station at Kottayam, and preserved in 70% ethanol. The samples were dissected under stereomicroscope (Weswox Optik-SZM-100, India) and mounted in Hoyer’s medium on microscopic slides. Identification at species level was carried out by examining specimens under binocular microscope (Primostar 3, Carl Zeiss Suzhou Co., Ltd., China) with reference to standard taxonomic keys (Table 2) [13, 24, 25]. Some specimens (female and male) did not match the characters of the reported species in the keys and published literature. Taxonomic features of these specimens are distinct from the congeners sand fly species (subgenus Anaphlebotomus) such as Phlebotomus (Ana.) colabaensis Young and Chalam, 1927, Ph. (Ana.) stantoni Newstead, 1914, and Ph. (Ana.) hoepplii Tang and Maa, 1945, which have been previously explained [13]. The morphometric analysis of the possible new sand fly species was carried out using a Zeiss binocular microscope aided with a micrometer. Every measurement was logged in micrometers (µm). Images of unique identifying features of these specimens were taken using a camera mounted over the same compound microscope. A holotype female and allotype male along with nine paratype female samples were used for morphometric analysis. Terminologies of the attributes for description were adopted from Galati et al. [26]. Nomenclature was adopted following the guidelines given by the International Code of Zoological Nomenclature (ICZN) [27]. Measurements (µm) of Ph. (Ana.) ajithii n. sp. (holotype female and allotype male) are described below (Table 3).
Molecular identification
The genomic DNA was extracted from both the legs and entire body of individual sand flies using the QIAmp DNeasy Kit (Qiagen, Germany), adhering to the manufacturer's instructions. The sand fly specimen was homogeneously crushed with a mortar and pestle, and final DNA elution was done in 30 μl molecular-grade nuclease-free water. DNA barcoding of both female and male samples was performed targeting mitochondrial cytochrome c oxidase subunit I (COI) gene (~ 720 bp). The gene was amplified according to the procedure outlined by Kumar et al. [28]. Sequencing of the amplicons was done bi-directionally using the same set of primers. The nucleotide sequences generated were deposited in GenBank.
Phylogenetic analyses
For phylogenetic analysis, the sequences were blasted with nucleotide repository (GenBank), and the sequences most similar to resulting sequences were aligned using MEGA 7.0. The sequences aligned with other congeners were analysed for phylogenetic tree construction using neighbor-joining statistical method with Kimura 2.0 parameter and 10,000 bootstraps. The genetic distance and other related parameters were estimated using MEGA 7.0.
Natural infection assessment of Leishmania parasite
Since the possible new species of sand fly belongs to the Phlebotomus genus, assessment for natural infection of Leishmania parasite was carried out. The whole genomic DNA was subjected to real-time detection of Leishmania kinetoplast minicircle DNA (kDNA). The protocol for kDNA detection was followed as described by Castelli et al. [29] using primers LEISH-1 (5ʹ GGCGTTCTGCGAAAACCG 3ʹ), LEISH-2 (5ʹ AAAATGGCATTTTCGGGCC 3ʹ) and TaqMan probe (5ʹ FAM-TGGGTGCAGAAATCCCGTTCA 3ʹ-BHQ1).
Results
Family Psychodidae Newman, 1834.
Subfamily Phlebotominae Rondani & Berté, in Rondani 1840.
Genus Phlebotomus Rondani & Berté, in Rondani 1840.
Subgenus Anaphlebotomus Theodor, 1948.
Species Phlebotomus (Anaphlebotomus) ajithii n. sp. Shah, Fathima, Jicksy & Saini (Figs. 2 and 3).

Phlebotomus (Anaphlebotomus) ajithii n. sp. (female). A Whole body without head and dissected terminalia; B (1) Palps, (2) flagellomere 1 to 3; C f2 with ascoid; D maxillary external and internal teeth; E cibarium with distinct horizontal teeth; F pharynx; G wing; H spermatheca with long and highly coiled individual duct

Phlebotomus (Anaphlebotomus) ajithii n. sp. (male). A Dissected terminalia; B sperm pump, aedeagal duct and parameral sheath; C gonostylus with spines arranged on it; D gonocoxite with internal setae on median tuft; E complex paramere with setae on upper lobe and a smaller ventral lobe; F pharynx and cibarium
Female
Holotype female: general colour of the specimen is consistently golden brown. Body size: 1920 µm. Head: length 442 µm, width 386 µm. Interocular distance: 163 µm. Labrum is 268 µm long. Hypopharynx with about 18 teeth on each side. Number of maxillary internal teeth: 15–18; external teeth: 9–11. Palps: formula 5, 3 = 2, 1, 4 (p5 > p3 = p2 > p1 > p4). Palpomere measurements: p1 128 µm, p2 155 µm, p3 155 µm, p4 75 µm and p5 180 µm. A group of about 20 club-like Newstead’s sensilla observed on middle of third palpal segment (p3). No such structure found on other palpal segments. One distal spiniform seta observed on p3, two setae on p4 and six setae on p5. Antenna: f1 255 µm, f2 106 µm, f3 106 µm (f1 > f2 + f3). Ascoidal formula: each antenna (f1–f13) has a pair of ascoids (almost reaching the next antennal segment), one on each sides. Ascoid length in f2: 68 µm. No simple setae (SS) present on f1 to f10. Three SS on f11, eight on f12, ten on f13 and 20 on f14 were observed. Single distal papilla observed on f1 to f3. Papilla absent from f4 to f11. One papilla on f11, three on f12, five on f13 and three on f14 were observed.
Cibarium: the ventral area bears a few distinct horizontal teeth but no denticles or fore-teeth; pigment patch was absent on the dorsal plate of the cibarium. Cibarium bears about 4–5 varying distinct horizontal teeth. All the teeth are irregularly arranged, tapering to fine points. Pharynx: nearly slender in shape with relatively broad base, armed heavily with a small group of minute spicules, length 189 µm, width 74 µm, pharyngeal armature depth 63 µm. Pharynx is about 2.5 times as long as wide. Wings: length 1786 µm, width 567 µm. Length of principal vein sections: alpha 430 µm, beta 283 µm, gamma 232 µm, delta (R1 overlap) 102 µm, R5 length 1252 µm. Wing index (alpha/beta) 1.51.
Fore leg: coxa 275 µm, trochanter 79 µm, femur 763 µm, tibia 1013 µm, tarsomeres: T1 642 µm, T2 263 µm, T3 188 µm, T4 163 µm, T5 100 µm.
Genitalia: Spermathecae: tubular shaped, slightly narrow towards duct end with 13–15 clear distinct segmentations. Length 39 µm, width 16 µm. Apical segment of the spermatheca is not enlarged (short neck). Spermatheca with secretary cells at the distal end and narrow individual spermathecal duct with striations. The length of the individual spermathecal duct is about 15–20 times (585–780 µm) the length of the spermatheca, and the individual ducts were highly coiled. The common spermathecal duct is short, 54 µm long. Cerci is simple and 146 µm long. Genital furca is 63 µm long (Fig. 2).
Male
Allotype male: same colour as the female specimen, i.e. consistently golden brown. Body size: 1733 µm. Head: length 384 µm, width 364 µm. Interocular distance 150 µm. Labrum is 183 µm long. Teeth on hypopharynx and maxilla were rudimentary. Palps: formula similar to that of female specimens, i.e. 5,3 = 2,1,4 (p5 > p3 = p2 > p1 > p4). Palpomere measurements: p1 98 µm, p2 108 µm, p3 108 µm, p4 68 µm and p5 161 µm. A group of about 15 club-like Newstead’s sensilla observed on middle of third palpal segment (p3). No such structure found on other palpal segments. One distal spiniform seta observed on p3, two setae on p4 and eight setae on p5. Antenna: f1 230 µm, f2 103 µm, f3 103 µm (f1 > f2 + f3). Ascoidal formula: each antenna (f1–f13) has a pair of ascoids (almost reaching the next antennal segment), one on each side. Ascoid length in f2: 51 µm. No SS present on f1 to f10. Four SS on f11, 10 on f12, 10 on f13 and 20 on f14 were observed. Single distal papilla observed on f1 to f3. Papilla absent from f4 to f11. One papilla on f11, three on f12, three on f13 and two on f14 were observed.
Cibarium: no visible horizontal teeth, denticles or fore-teeth, pigment patch was absent on the dorsal plate of the cibarium. Pharynx: nearly slender in shape with marginally broad base, comparatively lightly armed compared with female specimen, with a small group of minute spicules, length 158 µm, width 44 µm, pharyngeal armature depth 43 µm. Pharynx is about 3.6 times as long as wide.
Wings: length 1478 µm, width 477 µm. Length of principal vein sections: alpha 300 µm, beta 226 µm, gamma 110 µm, delta (R1 overlap) 50 µm, R5 length 927 µm. Wing index (alpha/beta): 1.33.
Fore leg: coxa 275 µm, trochanter 75 µm, femur 593 µm, tibia 835 µm, tarsomeres: T1 550 µm, T2 250 µm, T3 150 µm, T4 125 µm, T5 100 µm.
Genitalia: length of sperm pump is 204 µm, length of aedeagal duct is 175 µm, length of sperm pump + length of aedeagal duct: 378 µm, ratio of length of sperm pump/length of aedeagal duct is 1.17. Aedeagal duct is much shorter than the length of the spermathecal duct of female specimen. The aedeagal filament has no striation, i.e. it is smooth, straight, with rounded ends and overall slender throughout the duct. Ejaculatory apodeme of the male genitalia; 152 µm long. Gonocoxite of length 203 µm and with a median tuft of 48–50 internal setae. Gonostyle with a length of 150 µm and four thick spines: one apical, two subapical or subterminal and one basal with average length of 108 µm. Complex paramere with two lobes: long upper lobe with length of 148 µm and about 60 strong upward-facing setae, another shorter lobe (ventral process) without any such setae. Parameral sheath or aedeagus is thick distal end with tapering end and is finger-like in structure with a length of 102 µm. Length of epandrial lobes is 226 µm (Fig. 3).
Diagnosis
The characteristics of uniformly erect hairs on abdominal tergites (2–6) found in the specimen are particular to the genus Phlebotomus. Cibarium has spicules, i.e. fine tooth-like structures, but not in a consistent row. Pigment patch is absent. Pharynx is armed with a minute group of teeth in the middle and behind it a few concentric lines. Palps extending further than antenna III (f3). Spermatheca has regular segmentation. Apical segment of the spermatheca is not enlarged, i.e. it has a short neck. Chitinous arch is well developed along with distinct horizontal teeth in the cibarium region of the head. Terminalia is short or medium sized. Paramere with or without short ventral process. Gonostyle with four long spines and coxite without basal process. These taxonomic characters confirm the addition of the species into subgenus Anaphlebotomus of genus Phlebotomus. Cibarium had a few distinct horizontal teeth in female but was rudimentary in male specimens. Maxillary teeth range between 15–18 on the internal side and 9–11 on the external side in the mouth parts of females. Ascoid on second antennal segment (f2) almost crossed the joint between f2 and f3 in both sexes. Spermatheca was tubular in shape with 13–15 clear distinct segmentations. The individual spermathecal duct was 15–20 times (585–780 µm) the length of individual spermatheca and was highly coiled. In males, gonostyle had four thick spines, one apical, two subapical or subterminal and one basal. Paramere with one ventral process and aedeagus with thick distal area and tapered at the proximal end. The aedeagal duct was short in length and was smooth and slender in shape. These features are distinctive in holotype female and allotype male Ph. (Ana.) ajithii n. sp. specimens.
Variability
The morphometric characteristics indicated that the holotype and paratype of female and allotype and paratype of males were similar (Table 3). Holotype, allotype and paratype female and male were collected from the same type of habitats but different districts. All specimens of Ph. (Ana.) ajithii n. sp. showed similarities in the taxonomic characteristics. In addition, DNA barcode sequences of the collected specimens from different districts showed variation in four nucleotides and an overall negligible genetic distance (K2P) within the specimens, thus suggesting a single taxonomic group. However, the genetic distance from the other most comparable congeners was 11.7% [Ph. (Ana.) colabaensis: 16.7%; Ph. (Ana.) stantoni: 17.7%] (Fig. 4).

Phylogenetic tree of mitochondrial cytochrome c oxidase subunit I (COI) gene sequences for species of Phlebotomus (Anaphlebotomus) ajithii n. sp. along with Ph. (Ana.) colabaensis, Ph. stantoni, Ph. (Euphlebotomus) argentipes and Ph. (Eup.) mascomai; outgroup; Ph. (Phlebotomus) papatasi; M male, F female
Type materials
Phlebotomus (Ana.) ajithii constitutes about 4.9% (69 specimen) of the total species composition. The type locality of Ph. ajithii n. sp. is cattle sheds and adjacent rooms from Kadaar tribal community of Mullurkara (Thalapilly taluk) (GPS coordinates 10.69"N, 76.25"E, altitude: 103 m above sea level). This new species has also been caught from same habitat in different districts as described in Table 1. Paratype females were obtained from the Kadaar tribe of Mullurkara tribal settlements Thrissur, Kerala, India. Paratype males were collected from Ottamala settlement, taluk-Vellarikund, Panchayat-Panathadi, District-Kasaragod, Kerala, India: (12.48"N, 75.33"E, altitude: 158 m above sea level). Voucher specimens, comprising holotype female and allotype male, were mounted separately on microscopic glass slides and serially numbered with details about the place and date of collection, habitat type, etc., and were deposited at the museum, ICMR-VCRC), Puducherry-605 006, India. Additionally, paratype female and male were subsequently submitted to the National Museum, Zoological Survey of India, Alipur, Kolkata, India.
Phlebotomus (Ana.) ajithii samples processed for molecular analysis were submitted to National Center for Biotechnology Information (NCBI) GenBank with accession numbers PP501172-PP501177, PQ192578-PQ192583 and PQ192585-PQ192588.
Type specimen: female holotype (voucher KS-265[VCRC]) and male allotype (voucher HS-1310[VCRC]) are deposited in the museum of ICMR-VCRC, Puducherry, India.
Etymology
The new species Ph. (Ana.) ajithii is named after Mr. PM Ajithlal (Technical Officer ‘C’ [Retired]), ICMR-VCRC, Field Station, Kottayam, Kerala) in recognition of his unwavering commitment and lifelong dedication to the field of public health entomology.
ZooBank registration
Following section 8.5 of the ICZN, 2012 amended version [27], details of the new species have been submitted to ZooBank. The life science identifiers (LSID) associated with the record are urn:lsid:zoobank.org:pub:FFEFC2C3-B79C-4155-A3F3-56F0AB972E0D and urn:lsid:zoobank.org:act:90BDFC54-5BCD-4168-9AAC-2A25E7F75B1A.
Natural infection assessment of Leishmania parasite
None of the Ph. (Ana.) ajithii samples were found positive for Leishmania parasite.
Discussion and conclusion
The distribution of sand flies from the rain forests of Western Ghats in India was abridged and updated by Lewis [13, 25]. The present systematic entomological survey was carried out in various tribal villages situated in the Western Ghats region of different districts in Kerala. These areas were selected because of their epidemiological relevance, i.e. case reports of leishmaniasis. These areas provide favourable macro- and microhabitat (rainfall, organic-rich soil and variety of hosts as blood sources, etc.) for the proliferation and abundance of sand flies all year. Additionally, this investigation provides data on the record of a new sand fly species from these tribal settlements.
There are several genera of sand flies, of which genus Phlebotomus Rondani and Berté in Rondani 1840, comprises many species from the Old World [13, 24]. Subgenera Euphlebotomus and Anaphlebotomus are the only two subgenera in which some species share common female characteristics, exhibiting pharyngeal armature, being armed and having cibarial teeth or spicules. However, females of both subgenera can be distinguished based on the apical segment of the spermatheca [13]. Species under the subgenus Euphlebotomus exhibit the peculiar characteristics of apical segments of the spermatheca being differentiated or enlarged compared with the others by a deeper furrow [13, 24, 30].
The subgenus Anaphlebotomus was one of the smallest groups under the genus Phlebotomus, including a total of only three species from the Oriental region [13, 24]. Phlebotomus (Ana.) colabaensis, Ph. hoepplii and Ph. stantoni are the species of the subgenus Anaphlebotomus which were recorded from the country [13, 24]. In 2023, this was revised to five species by Shah et al. in the review article entitled “Faunal richness and checklist of sand flies (Diptera: Psychodidae) in India” with two species records from India, i.e. Phlebotomus (Ana.) chiyankiensis Singh, Phillips Singh and Ipe, 2009, and Ph. (Ana.) palamauensis Singh, Phillips Singh and Ipe, 2007 [23]. However, as discussed by Renaux et al. in 2023, the validation of these species cannot be confirmed based only on the drawings and morphological descriptions provided by the authors [31]. Furthermore, we also accept the revision of the systematics of the subgenera Anaphlebotomus and reinstated the validity of Phlebotomus maynei, based on examination of its holotype by Renaux et al. [31]. In this continuation, a new species record, Ph. (Ana.) ajithii n. sp., is described in this article.
Referring to the standard taxonomic keys, the female specimens of Ph. (Ana.) ajithii n. sp. were found to be similar to females and males of the congener, such as Ph. (Ana.) colabaensis, Ph. stantoni and Phlebotomus shadenae [13, 24, 31]. However, based on some peculiar taxonomic characteristics, those plausible new species specimens were separated. In Ph. (Ana.) colabaensis female, the labrum is about 300 µm long. Hypopharynx has about 19 teeth on each side and maxilla with 11 lateral and 23 ventral teeth. Ascoid in f2 is about 60 µm. Spermatheca is slightly carrot shaped with a small end segment. Spermathecal duct is long, i.e. about four to five times length of the individual spermatheca which joins to the common duct [13, 24]. In Ph. (Ana.) colabaensis male, the terminalia has four spines on the gonostyle with two being apical and subapical and one basal in position. Sperm pump is 150 µm long. Gonocoxite and gonostyle are about 150 and 230 µm long, respectively. Aedeagus or parameal sheath is pyramidal in shape and sharply pointed [31]. In Ph. (Ana.) stantoni female, the labrum is about 230–260 µm long. Hypopharynx has about 16 teeth on each side and maxilla with nine lateral and 18 ventral teeth. Ascoid in f2 is about 80 µm. Spermatheca is fusiform or spindle shaped with about 15–16 segments; the neck is thick and short with a more or less oblong head. The spermathecal duct is slightly longer than the length of the individual spermatheca and is striated. Common duct is very long, i.e. about 1.5 times the length of the spermatheca with thick walls [13, 24]. In Ph. (Ana.) stantoni male, labrum is about 169 µm long. Gonostyle is 77 µm long, and sperm pump is 143 µm long. Ejaculatory apodeme is about 107 µm long. Gonocoxite is 190 µm long with 43 internal gonocoxal tuft setae. Aedeagus or parameal sheath is 88 µm long [31]. For Ph. shadenae female, labrum is 187 µm long. Hypopharynx has about 15 teeth on each side and maxilla with 8 lateral and 15 ventral teeth. Individual spermathecal duct is 69 µm long and has spermatheca with > 15 rings along with the presence of a sessile head carried by a broad process [31]. In Ph. (Ana.) shadenae male, labrum is about 167 µm long. Gonostyle is 84 µm long with four thick spines, one of which is terminal and one subterminal, and two are basal in position. Sperm pump is 139 µm long. Ejaculatory apodeme is about 107 µm long. Gonocoxite is 207 µm long with 56 internal gonocoxal tuft setae. Aedeagus or parameal sheath is 88 µm long and is finger-like in shape. Aedeagal ducts are straight, smooth and overall tapering and slender with rounded ends [31]. However, Ph. (Ana.) ajithii n. sp. female has about 270-µm-long labrum. The hypopharynx has about 18 teeth on each side, and the maxilla has 10 lateral and 17 ventral teeth. Ascoid present on f2 is about 68 µm. The apical segment of the spermatheca is not enlarged (short neck), which is one of the key features that differs between the two subgenera, i.e. Euphlebotomus and Anaphlebotomus [13, 24, 25]. Spermatheca is tubular and slightly narrow towards the duct end with about 13–15 segmentations. Spermathecal ducts are narrow with striations. The individual spermathecal duct is long, about 15–20 times (585–780 µm) the length of the spermatheca, and is highly coiled. It has a short common spermathecal duct of 54 µm length (Fig. 2). In the Phlebotomus (Ana.) ajithii n. sp. male, labrum is about 183 µm long. Ascoid length on f2 is 51 µm. Gonostyle is 150 µm long with four thick spines, one apical, two subapical or subterminal and one basal in position, with average length of 108 µm. Sperm pump is 204 µm long. Ejaculatory apodeme is about 152 µm long. Gonocoxite is 203 µm long with 48–50 internal gonocoxal tuft setae. Aedeagus or parameal sheath is 102 µm long and has a thick distal end with tapering end. Aedeagal ducts have no striation, i.e. they are its smooth and straight with rounded ends and are overall slender throughout the duct.
In addition, molecular taxonomy by DNA barcoding followed by phylogenetic analysis also confirmed the association within the specimens of Ph. (Ana.) ajithii n. sp. with a very minimal genetic distance and four nucleotide variations. However, the overall genetic distance (GD) is 11.7% with the congener species. Phlebotomus (Ana.) colabaensis has 16.7% and Ph. (Ana.) stantoni has 17.7% GD with Ph. ajithii. The population genetic parameters analysed using MEGA 7.0 software also confirmed a very high genetic diversity (HST = 0.969) and trifling gene flow (Nm = 0.002). Hence, based on these taxonomic differences and molecular analysis, Ph. (Ana.) ajithii n. sp. is divergent from the other already reported and described species under the subgenus Anaphlebotomus. This species was mainly collected from indoor human dwellings since species of the genus Phlebotomus are mainly incriminated in the transmission of diseases [32,33,34,35,36], thus, infection assessment was carried out. However, none of the specimens were positive for the Leishmania parasite in qPCR targeting kDNA. In conclusion, the Western Ghats is an important biodiversity hotspot with a few database on entomological surveys of sand flies. The current study tried to fill this void and also report a new sand fly species.
Availability of data and materials
The sequences generated in the present study were deposited in NCBI GenBank under accession nos. PP501172-PP501177, PQ192578-PQ192583 and PQ192585-PQ192588.
Abbreviations
- COI:
-
Cytochrome c oxidase subunit I
- NTD:
-
Neglected tropical disease
- VL:
-
Visceral leishmaniasis
- CL:
-
Cutaneous leishmaniasis
- ICZN:
-
International Code of Zoological Nomenclature
- kDNA:
-
Kinetoplast-minicircle DNA
- NCBI:
-
National Center for Biotechnology Information
- ICMR:
-
Indian Council of Medical Research
References
WHO. Leishmaniasis. [cited 2024 May 14]. Available from: https://www.who.int/news-room/fact-sheets/detail/leishmaniasis.
WWF. About the Western Ghats. [cited 2024 May 14]. Available from: https://www.wwfindia.org/about_wwf/critical_regions/western_ghats2/about_the_western_ghats/.
Myers N, Mittermeier RA, Mittermeier CG, Da Fonseca GAB, Kent J. Biodiversity hotspots for conservation priorities. Nature. 2000;403:853–8.
UNESCO. Western Ghats. [cited 2024 April 08]. Available from: https://whc.unesco.org/en/list/1342/.
Chandran M. On the ecological history of the Western Ghats. Curr Sci. 1997;76:146–55.
Simi S, Anish T, Jyothi R, Vijayakumar K, Philip R, Paul N. Searching for cutaneous leishmaniasis in tribals from Kerala, India. J Global Infect Dis. 2010;2:95.
Murhekar MV, Kasabi GS, Mehendale SM, Mourya DT, Yadav PD, Tandale BV. On the transmission pattern of Kyasanur Forest disease (KFD) in India. Infect Dis Poverty. 2015;4:37.
Sukumaran A, Pradeepkumar AS. One Health approach: A platform for intervention in emerging public health challenges of Kerala state. Int J One Health. 2015;1:14–25.
Saini P, Kumar NP, Ajithlal PM, Joji A, Rajesh KR, Reena KJ, et al. Visceral leishmaniasis caused by Leishmania donovani zymodeme MON-37, Western Ghats, India. Emerg Infect Dis. 2020;26.
Kumar NP, Srinivasan R, Anish TS, Nandakumar G, Jambulingam P. Cutaneous leishmaniasis caused by Leishmania donovani in the tribal population of the Agasthyamala Biosphere Reserve forest, Western Ghats, Kerala, India. J Med Microbiol. 2015;64:157–63.
Saini P, Fathima PA, Aiswarya RS, Ajithlal PM, Rajesh KR, Simi SM, et al. Cutaneous and visceral leishmaniasis caused by the same zymodeme of Leishmania donovani in Kerala, India. Am J Trop Med Hyg. 2024;110:59–63.
Srinivasan R, Jambulingam P, Kumar NP, Selvakumar M, Edwin B, Kumar TD. Temporal distribution and behaviour of sand flies (Diptera: Psychodidae) in a cutaneous leishmaniasis focus of the Kani Tribe settlements in the Western Ghats, India. Acta Trop. 2015;148:147–55.
Lewis D. The phlebotomine sandflies (Diptera: Psychodidae) of the Oriental region. Bull Br Mus (Natural History); 1978.
Kaul SM. Phlebotomine sandflies (Diptera: Psychodidae) from Western Ghats in Kerala and Tamil Nadu States, India. Part I. Description of four new species. J Commun Dis. 1993;25:75–82.
Ilango K, Dhanda V, Srinivasan R, Sadanand AV, Lane RP. Phlebotomine sandflies (Diptera: Psychodidae) of Tamil Nadu and Pondicherry, southern India, in relation to visceral leishmaniasis. Ann Trop Med Parasitol. 1994;88:413–31.
Ilango K. A review of the Sergentomyia babu species complex (Diptera: Psychodidae: Phlebotominae) with description of a new species from India. Orient Insects. 2004;38:77–83.
Ilango K. Moth flies & sand flies (Diptera: Psychodidae) of India. Chennai: Rec Zool Surv India. 2009.
Renganathan S, Purushothaman J. Description of Sergentomyia (Parrotomyia) rectangulata n. sp. (Diptera: Psychodidae: Phlebotominae) from southern India. Mem Inst Oswaldo Cruz. 2010;105:792–5.
Srinivasan R, Jambulingam P. Description of a new species of sand fly Sergentomyia (Parrotomyia) vadhanurensis (Diptera: Psychodidae) from Puducherry Union Territory, India. J Am Mosq Control Assoc. 2011;27:15–20.
Srinivasan R, Jambulingam P, Vanamail P. Sand fly (Diptera: Psychodidae) abundance and species diversity in relation to environmental factors in parts of coastal plains of Southern India. J Med Entomol. 2013;50:758–63.
Srinivasan R, Jambulingam P, Kumar NP. Sergentomyia (Neophlebotomus) monticola, a new species of sand fly (Diptera: Psychodidae) from the Western Ghats, Thiruvananthapuram District, Kerala, India. Acta Trop. 2014;137:74–9.
Jambulingam P, Srinivasan R, Gopalakrishnan S. A report on occurrence of Phlebotomine sand flies (Diptera: Psychodidae) and two new country records from Andaman & Nicobar Islands, a Union territory of India. Zootaxa. 2022;5093:241–6.
Shah H, Fathima P, Kumar NP, Kumar A, Saini P. Faunal richness and checklist of sandflies (Diptera: Psychodidae) in India. Asian Pac J Trop Med. 2023;16:193.
Lewis DJ. Phlebotomine sandflies (Diptera: Psychodidae) from the Oriental Region. Syst Entomol. 1987;12:163–80.
Kalra N, Bang Y. Manual on entomology in visceral leishmaniasis. New Delhi, India: World Health Organization; 1988.
Galati EAB, Galvis-Ovallos F, Lawyer P, Léger N, Depaquit J. An illustrated guide for characters and terminology used in descriptions of Phlebotominae (Diptera, Psychodidae). Parasite. 2017;24:26.
ICZN International Commission on Zoological Nomenclature. Amendment of articles 8, 9, 10, 21 and 78 of the International Code of Zoological Nomenclature to expand and refine methods of publication. Bull Zool Nomencl. 2012;69:161–9.
Kumar NP, Srinivasan R, Jambulingam P. DNA barcoding for identification of sand flies (Diptera: Psychodidae) in India. Mol Ecol Resour. 2012;12:414–20.
Castelli G, Bruno F, Reale S, Catanzaro S, Valenza V, Vitale F. Molecular diagnosis of leishmaniasis: quantification of parasite load by a real-time PCR assay with high sensitivity. Pathogens. 2021;10:865.
Muller F, Depaquit J, Léger N. Phlebotomus (Euphlebotomus) mascomai n. sp. (Diptera–Psychodidae). Parasitol Res. 2007;101:1597–602.
Renaux Torres M-C, Pellot C, Somwang P, Khositharattanakool P, Vongphayloth K, Randrianambinintsoa FJ, et al. Phlebotomine sand flies (Diptera, Psychodidae) from Pha Tong cave, Northern Thailand with a description of two new species and taxonomical thoughts about Phlebotomus stantoni. PLoS Negl Trop Dis. 2023;17:e0011565.
Davami MH, Motazedian MH, Kalantari M, Asgari Q, Badzohre A, Mohammadpour I. First microscopical and molecular-based characterization of Leishmania major within naturally infected Phlebotomus salehi (Diptera; Psychodidae) in Fars province, southern Iran. Ann Trop Med Parasitol. 2011;105:485–91.
Satoskar AR, Salotra P, Bumb RA, Khandelwal K, Ghiya BC, Dodd C, et al. Clinco-epidemiologic study of cutaneous Leishmaniasis in Bikaner, Rajasthan, India. Am J Trop Med Hyg. 2013;89:111–5.
Srinivasan R, Kumar NP, Jambulingam P. Detection of natural infection of Leishmania donovani (Kinetoplastida: Trypanosomatidae) in Phlebotomus argentipes (Diptera: Psychodidae) from a forest ecosystem in the Western Ghats, India, endemic for cutaneous leishmaniasis. Acta Trop. 2016;156:95–9.
Jambulingam P, Pradeep Kumar N, Nandakumar S, Paily KP, Srinivasan R. Domestic dogs as reservoir hosts for Leishmania donovani in the southernmost Western Ghats in India. Acta Trop. 2017;171:64–7.
Lata S, Kumar G, Ojha VP, Dhiman RC. Detection of Leishmania donovani in Wild-Caught Phlebotomine Sand Flies in Endemic Focus of Leishmaniasis in Himachal Pradesh. India J Med Entomol. 2022;59:719–24.
Acknowledgements
We are extremely grateful to the Director of ICMR-VCRC, for her constant support, guidance and encouragement in conducting the surveys. We also thank the Director, Directorate of Scheduled Tribe Development Department, and the Chief Conservator of Forest and Chief Wild Life Warden, Government of Kerala, for their kind permission to carry out the survey in the tribal hamlets of Western Ghats. We extend our sincere gratitude to all of the staff at the ICMR-VCRC Field Station in Kerala for their assistance during this study. FPA acknowledges a fellowship provided by the Council of Scientific and Industrial Research, New Delhi (File no. 09/1319(0001)/2020-EMR-I).
Funding
This study is supported by the Indian Council of Medical Research, New Delhi, grant no. 6/9-7(331)/2020/ECD-II and 6/9-7(271)/KA/2021-ECD-II), and Indian Council of Medical Research-Vector Control Research Centre (project grant no. IM-1905).
Author information
Authors and Affiliations
Contributions
Conceptualization: HKS, PAF, PS; data curation and investigation: HKS, PAF; formal analysis: HKS, PAF, PS; methodology: HKS, PAF, JJ, PS; project administration and resources: PS; Writing original draft: HKS, PAF; writing, review and editing: HKS, PAF, PS.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
All authors have approved the final version of the manuscript and provide consent to publish.
Competing interests
The authors declare that there is no conflict of interest involved in this study.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Shah, H.K., Fathima, P.A., Jicksy, J. et al. Report of a new species of sand fly, Phlebotomus (Anaphlebotomus) ajithii n. sp. (Diptera: Psychodidae), from Western Ghats, India. Parasites Vectors 17, 388 (2024). https://doi.org/10.1186/s13071-024-06468-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13071-024-06468-2