
Abstract
Background
Antimicrobial resistance is growing substantially, which necessitates the search for novel therapeutic options. Terbinafine, an allylamine antifungal agent that exhibits a broad spectrum of activity and is used in the treatment of dermatophytosis, could be a possible option to disarm S. aureus virulence.
Methods
Terbinafine inhibitory effect on staphyloxanthin was characterized by quantitative measurement of staphyloxanthin intermediates and molecular docking. The effect of terbinafine on S. aureus stress survival was characterized by viable counting. The anti-biofilm activity of terbinafine on S. aureus was assessed by the crystal violet assay and microscopy. Changes in S. aureus membrane following treatment with terbinafine were determined using Fourier transform infrared (FTIR) analysis. The synergistic action of terbinafine in combination with conventional antibiotics was characterized using the checkerboard assay. qRT-PCR was used to evaluate the impact of terbinafine on S. aureus gene expression. The influence of terbinafine on S. aureus pathogenesis was investigated in mice infection model.
Results
Terbinafine inhibits staphyloxanthin biosynthesis through targeting dehydrosqualene desaturase (CrtN). Docking analysis of terbinafine against the predicted active site of CrtN reveals a binding energy of − 9.579 kcal/mol exemplified by the formation of H-bonds, H-arene bonds, and hydrophobic/hydrophilic interactions with the conserved amino acids of the receptor pocket. Terbinafine treated S. aureus was more susceptible to both oxidative and acid stress as well as human blood killing as compared to untreated cells. Targeting staphyloxanthin by terbinafine rendered S. aureus more sensitive to membrane acting antibiotics. Terbinafine interfered with S. aureus biofilm formation through targeting cell autoaggregation, hydrophobicity, and exopolysaccharide production. Moreover, terbinafine demonstrated a synergistic interaction against S. aureus when combined with conventional antibiotics. Importantly, terbinafine attenuated S. aureus pathogenesis using mice infection model. qRT-PCR revealed that terbinafine repressed expression of the transcriptional regulators sigB, sarA, and msaB, as well as icaA in S. aureus.
Conclusions
Present findings strongly suggest that terbinafine could be used safely and efficiently as an anti-virulent agent to combat S. aureus infections.
Similar content being viewed by others

Background
Staphylococcus aureus is a commensal bacterium that asymptomatically colonizes humans. However, S. aureus could be an extremely versatile pathogen in humans, causing life-threatening infections. S. aureus possesses a wide range of virulence factors that enable it to thrive within the host [1]. Diseases caused by S. aureus are diverse, ranging from minor skin infections to highly severe infections including pneumonia, endocarditis, and sepsis [2]. S. aureus infections are problematic because of the frequent emergence of antibiotic resistant strains of S. aureus [3]. The emergence of methicillin-resistant S. aureus (MRSA) and vancomycin-resistant MRSA (VRSA) limits the usage of antibiotics and renders treatment more challenging [4]. In view of these facts, new antibacterial agents are urgently needed that target S. aureus with no pressure on the bacterium in order to minimize the development of antibiotic resistance [5].
Staphyloxanthin is considered the hallmark virulence factor of S. aureus [6]. Staphyloxanthin is a golden-yellow carotenoid pigment found in the plasma membrane and acts as an antioxidant against host immune response. Furthermore, staphyloxanthin maintains the structural integrity of the bacterial membrane and is associated with bacterial survival under stressful conditions [7, 8]. Thus, disrupting staphyloxanthin biosynthesis has become an innovative anti-infective approach to alter S. aureus virulence with the advantage of minimizing the emergence of antibiotic resistant strains [9]. Several enzymes are involved in staphyloxanthin biosynthesis. First, CrtM, a dehydrosqualene synthase, catalyzes the condensation of two molecules of farnesyl diphosphate to form 4,4′-diapophytoene, which is further dehydrogenated to 4, 4′-diaponeurosporene by CrtN, a dehydrosqualene desaturase. Finally, 4, 4′-diaponeurosporene undergoes oxidation, glycosylation, and esterification to give staphyloxanthin [10].
In addition to staphyloxanthin and its role in staphylococcal pathogenesis, S. aureus forms biofilm, which is an adherent microbial community on both biotic and abiotic surfaces [11]. Biofilm formation is mediated by self-secreted polymeric substances that provides a hydrated matrix structure that protects bacterial cells against both antimicrobials and host immune response [12]. Biofilm-related infections account for increased morbidity and mortality and could result in prolonged durations of hospitalization [13]. Therefore, the abolishment of bacterial biofilm formation has been reported to be an efficient strategy to counteract bacterial infections [14].
Naphthalene-containing compounds are known to have a variety of biological activities, including antimicrobial properties [15, 16]. The Food and Drug Administration has approved numerous naphthalene derivatives as therapeutics, such as naftifine and terbinafine. Terbinafine is an allylamine fungicidal, administered either topically or orally, and used in the treatment of superficial fungal infections of the skin and nails. Additionally, terbinafine has clinically relevant antibacterial properties that may be useful in mixed bacterial and fungal infections as athlete’s foot [17, 18]. Naftifine has been shown previously to have potent staphyloxanthin inhibiting activity [19]. Therefore, based on the structural similarity, the present study aims to characterize the anti-virulent potential of terbinafine against S. aureus. The influence of terbinafine on staphyloxanthin biosynthesis, biofilm formation, as well as S. aureus host pathogenesis will be fully uncovered herein.
Materials and methods
Bacterial strains and growth conditions
Staphylococcus aureus ATCC 6538 and five clinical isolates recovered from patients admitted to the Zagazig University Hospital in Egypt were included in this study. Clinical isolates were selected based on their pigment content; four isolates were pigmented, and one isolate was non-pigmented. Clinical S. aureus isolates were further characterized using 16S rRNA gene sequencing and results were deposited in GenBank (https://www.ncbi.nlm.nih.gov/) under accession numbers ON032523, ON032524, ON032525, ON032526 and ON032527. Bacteria were grown aerobically in a trypticase soy broth (TSB). For biofilm and virulence assays, bacterial cells were grown in TSB supplemented with 1% glucose (TSBG).
Determination of the minimum inhibitory concentration (MIC) of terbinafine
The MIC of terbinafine (Mash Premiere, Egypt) against S. aureus ATCC 6538 was determined as described before [20]. Results were recorded and MIC values were determined in triplicate. The effect of terbinafine on S. aureus viability was determined by growing bacteria in the presence of sub-MIC of terbinafine (132, 164, and 328 µg/mL) and bacterial growth was compared with untreated bacteria. Similarly, the impact of terbinafine on S. aureus metabolic activity was assessed by the alamar blue assay as previously described [21]. Resazurin solution (6.5 mg/mL) was added to both terbinafine treated and untreated bacteria and incubated in the dark for 4 h. The fluorescence signals of the supernatant were recorded at 590 nm emission and 560 nm excitation wavelengths [21].
Investigation of the inhibitory effect of terbinafine on staphyloxanthin biosynthesis.
Carotenoid pigment was extracted from both terbinafine treated and untreated S. aureus and estimated as described before [22]. S. aureus strains were grown in TSB with varying concentrations of terbinafine (3.28, 8.2, 16.4, 24.6, 32.8, and 49.2 µg/mL) at 37 °C for 24 h. Cells suspensions were adjusted to an optical density of 2 at 600 nm (4 × 109 colony forming units (CFUs) in a 10 mL volume). Bacterial cells were harvested by centrifugation and resuspended in 99% methanol and agitated for 30 min at 55 °C in the dark. Following centrifugation, the supernatant absorbance was measured at 465 nm using a Bio-Tek synergy HT microplate reader, USA. The half maximal inhibitory concentration (IC50) of terbinafine was estimated as previously described [23]. Additionally, staphyloxanthin biosynthesis intermediates were determined in response to terbinafine treatment. The absorbance of methanol-extracted carotenoids containing staphyloxanthin intermediates was measured using a plate reader (Synergy HT, BioTek) at wavelengths 286 nm, 435 nm, 455 nm, and 465 nm for 4,4′-diapophytoene 4,4′-diaponeurosporene 4,4′diaponeurosporenic acid, and staphyloxanthin, respectively [24].
Molecular docking analysis
The sequence of amino acids of CrtN (WP_000686169.1) was obtained as a primary structure in FASTA form from the NCBI database, and the predicted 3D structure of the receptor was obtained from the IntFOLD Server (Version 5.0) [25]. Terbinafine was drawn into the Marvin Sketch of Marvin site (http://www.chemaxon.com) to generate the lowest energy conformer. The Dock module of Molecular Operating Environment (MOE 2019.0102) was used in molecular docking analysis [26]. All hydrogen atoms with their standard geometry were inserted to the protein predicted 3D structure, and then their energy was minimized. Using the flexible ligand mode, terbinafine was docked into the rigid binding pocket of the protein. The placement phase generates poses from ligand conformations. The ligand free energy of binding from a predicted pose is calculated using the GBVI/WSA ΔG as a force field-based scoring function [27].
Characterization of terbinafine impact on S. aureus susceptibility to H2O2, acid stress, and whole blood killing
The sensitivity of S. aureus to H2O2 oxidative stress, acid stress, and whole blood killing following treatment with terbinafine was determined and compared with untreated cells [28]. Briefly, S. aureus ATCC 6538 and the non-pigmented S. aureus isolate were cultured in TSB with sub-MIC of terbinafine (328 μg/mL). Bacterial pellets were collected and adjusted to a concentration of 107 CFU/mL. For oxidative stress, H2O2 (1.5%) was added, and bacterial survival was assessed every 15 min over a 1-h period. For acid stress, S. aureus cells (106 CFU/mL) were exposed to acid stress at pH 4 (adjusted with acetic acid) and bacterial survival was assessed. Bacterial survival to whole blood killing was evaluated as follows: freshly drawn heparinized human blood and S. aureus cultures (6 × 108 CFU/mL) were mixed in 3:1 volume ratio and incubated for 2 h. For all experiments, bacterial viability was assessed by counting surviving bacteria after serial dilution in phosphate buffered saline (PBS) and plating on TSA and compared to control untreated cells.
Fourier transform infrared (FTIR) spectroscopy analysis
Changes in S. aureus membrane upon terbinafine exposure were determined using FTIR analysis [29]. S. aureus ATCC 6538 overnight culture in the presence of terbinafine (328 µg/mL) was adjusted to 9 × 108 CFU/mL. Bacterial suspension was centrifuged and collected cells were subjected to FTIR analysis (Bruker Alpha FTIR) and the spectrum was scanned in the range of 4000–500 cm−1. The FTIR spectra of treated cells were plotted as transmittance against wave number and compared to untreated S. aureus.
Evaluation of the effect of terbinafine on S. aureus survival to polymyxin B and biofilm formation
The effect of terbinafine on S. aureus survival to the membrane acting antibiotic polymyxin B was determined. Overnight S. aureus cultures (5 × 106 CFU/mL) in the presence of terbinafine were treated with 1 mM polymyxin B and incubated for 30 min. Bacterial viability was assessed by serial dilution and plating on TSA and compared with untreated bacteria [30]. The biofilm quantitative assay was performed as described before [31] using the crystal violet (CV) method. S. aureus ATCC 6538 (106 CFU/mL) was allowed to form biofilm for 48 h. Formed biofilm was assayed by staining with CV solution and glacial acetic acid (33%) for CV solubilization. The absorbance was measured at 570 nm and the percentage of biofilm inhibition was calculated as follows: % of inhibition = [(Control OD570 nm−Treated OD570 nm)/Control OD570 nm] × 100. In addition, the inhibitory effect of terbinafine on S. aureus biofilm was assessed using both light and scanning electron microscopes [32]. Biofilms were developed on polystyrene discs in the presence and absence of terbinafine (132, 164 and 328 μg/mL) as described above. Discs were washed twice with PBS, fixed by 2.5% glutaraldehyde for 2 h, and dehydrated using ethanol. Finally, the discs were air dried and gold coated before imaging using JEOL scanning microscope (JSM-T100, Japan).
Characterization of the influence of terbinafine on S. aureus auto-aggregation, surface hydrophobicity, exopolysacchraide (EPS) formation and cell autolysis
Staphylococcus aureus auto-aggregation assay was performed as reported earlier [33]. Overnight S. aureus ATCC 6538 culture in TSBG containing terbinafine was centrifuged and then bacteria were resuspended in PBS and allowed to stand at 37 °C for 20 h. Cell density of the upper portion of PBS containing cells was measured at OD600 in comparison with untreated cells. As described earlier [34], the bacterial surface hydrophobicity index was determined. S. aureus ATCC 6538 grown in TSBG containing terbinafine (132, 164, and 328 μg/mL) was collected by centrifugation. Bacterial suspensions were adjusted to initial absorbance (Ai = 1.0), toluene was added, vortexed, and the absorbance of aqueous phase (Af) was measured. The hydrophobicity index (HI) was expressed as: HI = (Ai−Af)/Ai × 100% and compared to the HI of control untreated bacteria. EPS production by S. aureus was quantified using the phenol–sulfuric acid method as previously mentioned [35]. Glass slides were immersed in TSBG containing S. aureus with and without terbinafine and incubated for 24 h. Glass slides were removed, washed, and formed cell suspensions were mixed with 5% phenol/H2SO4. The mixture was left for 1 h and the supernatant absorbance was measured at 490 nm. Finally, the impact of terbinafine on bacterial induced cell autolysis was characterized. S. aureus ATCC 6538 culture in TSB containing 1 M NaCl and terbinafine (328 µg/mL), adjusted at A580 of 0.7, was centrifuged and washed. Cell pellets were resuspended in autolysis buffer (50 mM Tris–HCl and 0.1% Triton X-100) and incubated. Bacterial autolysis was measured at A580 over 3 h, where the decrease in optical density indicates a higher cell death rate [36].
Characterization of terbinafine effect on S. aureus deoxyribonuclease (DNase), esterase, and lipase activity
DNase agar and tween substrate plates were prepared, and after solidification, cups were cut into the agar medium. Supernatant of overnight cultures of terbinafine treated and untreated S. aureus were added into wells and incubated. DNase activity was observed as a zone of clearance around the well after addition of 1 N HCL, while a white precipitation zone appeared around wells boundary, indicative of esterase and lipase activity [37, 38].
qRT-PCR analysis
The qRT-PCR was performed to investigate the impact of terbinafine on the expression of S. aureus genes; sarA, icaA, icaR, agrA, crtM, crtN, sodA, sodM, katA, sigB, msaB, and yjbH. Briefly, an overnight culture of S. aureus ATCC 6538 grown in the presence of terbinafine was subjected to RNA extraction using the TRIzol reagent. Extracted RNA was purified using the Qiagen RNeasy minikit and reverse-transcribed into single-stranded complementary DNA (cDNA) using the QuantiTect-Reverse Transcription Kit, and cDNA was amplified using Maxima SYBR Green/Fluorescein qPCR Master Mix. The expression level of tested genes was normalized to 16S rRNA, and the 2 −∆∆CT method was used to calculate relative gene expression [39]. Primers used in the current study are listed in Table 1 [40,41,42,43,44,45,46].
Checkerboard assay
The checkerboard assay was performed to measure the synergy between terbinafine and antibiotics targeting S. aureus; ampicillin, cefotaxime, azithromycin, ciprofloxacin, and gentamycin [47]. The effect of the combination between terbinafine and selected antibiotics was evaluated by calculating the fractional inhibitory concentration index (FICI) [48] according to the following formula:
FIC of terbinafine = MIC terbinafine in combination/MIC of terbinafine alone; FIC of antibiotic = MIC of antibiotic in combination/MIC of antibiotic alone; hence FIC index (FICI) = FIC of terbinafine + FIC of antibiotic. “Synergy” was defined when FICI was ≤ 0.5; while “additive” in which 0.5 ≤ FICI ≤ 1.0; moreover “indifferent” when the FICI is between 1 and 4.
In vivo characterization of the influence of terbinafine on S. aureus virulence using mice infection model
The impact of terbinafine on S. aureus pathogenesis was determined [49, 50]. Briefly, overnight cultures of S. aureus ATCC 6538 in the presence of terbinafine (treated cells) and non-pigmented S. aureus were adjusted to 2.5 × 107 CFU/mL in PBS. Five mice groups, each containing six mice, were included in the experiment. Untreated, terbinafine-treated, and non-pigmented S. aureus were injected into the abdominal cavity of 3-week-old albino mice (the first, second, and third groups, respectively). As negative controls, uninjected mice (the fourth group) and PBS-injected mice (the fifth group) were included. Mice were sacrificed at 24 h post-infection and the spleen, liver, and kidney were recovered aseptically and employed for bacterial load determination. Results were represented and expressed as the mean (CFU/g) ± standard errors. In addition, fragments of organs were fixed in buffered formalin (10%) for histopathological examination. The statistical analysis was examined by Mann–Whitney U analysis (P < 0.05 is considered significant).
Statistical analyses
All experiments were carried in triplicate and the findings were expressed as the mean ± standard error. Unless otherwise stated, statistical analyses were conducted with GraphPad Prism 5 software using Student t-tests or one-way ANOVA.
Results
Terbinafine inhibits the biosynthesis of staphyloxanthin at sub-MIC concentrations
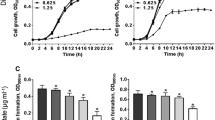
The MIC of terbinafine against S. aureus ATCC 6538 was determined as 2624 μg/mL. Results indicate that sub-MICs (132, 164, and 328 μg/mL) of terbinafine don’t interfere with S. aureus growth (Fig. 1A). Furthermore, the alamar blue assay shows that terbinafine-treated S. aureus was as metabolically active as untreated control cells (Fig. 1B). The inhibitory effect of terbinafine on staphyloxanthin biosynthesis in S. aureus ATCC 6538 was evaluated. Terbinafine inhibited staphyloxanthin biosynthesis in a dose dependent way with an IC50 of 36 µM (Fig. 2A–B). Similarly, terbinafine inhibited staphyloxanthin biosynthesis in S. aureus clinical isolates at micromolar concentrations (66–95 µM) (Fig. 2C–F).

Terbinafine had no effect on S. aureus growth or viability at sub-MICs (132, 164, and 328 µg/mL). A CFU/mL of terbinafine treated cells and untreated cells with no significant difference, indicating that sub-MIC had non bactericidal activity. B Terbinafine treated cells were metabolically active as control cells, confirmed by the alamar blue assay. Data shown represent the mean ± standard error from triplicate experiments

Terbinafine inhibited staphyloxanthin biosynthesis. A Terbinafine inhibition of S. aureus pigment in a dose dependent manner. B Dose inhibition curve with maximal inhibitory concentration (IC50) of terbinafine against S. aureus ATCC 6538. Dose inhibition curve with maximal inhibitory concentration (IC50) of terbinafine against S. aureus clinical isolates from wound infection (C), burn infection (D), respiratory tract infection (E) and urinary tract infection (F). Data shown represent the mean ± standard error from triplicate experiments
Terbinafine exhibits its activity via interference with CrtN
Staphylococcus aureus pigment methanolic extract was quantified spectrometrically upon treatment with sub-MIC of terbinafine. Importantly, treatment of S. aureus with terbinafine results in a significant accumulation of the CrtN substrate, 4,4′-diapophytoene. On the contrary, the amounts of subsequent staphyloxanthin biosynthesis intermediates (4,4′-diaponeurosporene and 4,4′-diaponeurosporenic acid) were significantly reduced in terbinafine treated S. aureus as compared to untreated bacteria (Fig. 3A). Furthermore, the molecular docking analysis validated terbinafine interaction with CrtN with a characteristic binding mode and affinity, exhibiting a binding energy of − 9.579 kcal/mol. Docking analysis results demonstrate that the tertiary amine group in the middle of terbinafine structure featured a conspicuous bifurcated H-bond with the conserved amino acids Val10 and Thr11 in the core of CrtN active site. Moreover, the flapping terminal of the ligand was fixed by an H-arene bond between ter.butyl group of the ligand and the conserved aromatic amino acid Tyr436 which enhanced the ligand/receptor complex stability. The overall recognition of the ligand inside the CrtN receptor hot spot was also augmented by the hydrophobic/hydrophilic interactions of the ligand and receptor (Fig. 3B–D).

A Terbinafine treatment led to the accumulation of 4, 4′-diapophytoene. B Ligand binding residues prediction by for CrtN; predicted ligand binding residues are shown as blue sticks. C Domain boundary prediction for CrtN; the model was colored according to the predicted domains. The putative binding mode of terbinafine into the predicted active site of CrtN receptor in 2D (D) and 3D structure (E). The blue and cyan shadows of the ligand and active site amino acids, respectively, indicated strong hydrophobic/hydrophilic interactions. Data shown represent the mean ± standard of error from triplicate experiments. Using Student’s t test, P value < 0.05 was considered statistically significant
Terbinafine treated S. aureus is more susceptible to environmental stresses
When compared to untreated S. aureus, terbinafine-treated S. aureus were more easily killed by hydrogen peroxide (bacterial survival of 28.1 ± 1.68% vs. 58.9% ± 2.01, respectively; Fig. 4A). Similarly, terbinafine treated S. aureus was more susceptible to acidic stress relative to untreated cells (bacterial survival of 52% ± 2.55 vs 75% ± 1.24, respectively; Fig. 4B). Importantly, terbinafine markedly sensitized S. aureus to human blood killing. The survival of the terbinafine treated S. aureus was almost three times lower than that of the untreated S. aureus (12.1% ± 2.194 vs. 34.8% ± 2.17, respectively; Fig. 4C).

Terbinafine treatment senstised S. aureus ATCC 6538 to; A hydrogen peroxide, B acetic acid (pH = 4 ± 0.5) and C human blood cells. (•) untreated cells, (▲) terbinafine treated and (■) non-pigmented isolate. Data shown represent the mean ± standard of error from triplicate experiments. Using ANOVA test, P value < 0.05 was considered statistically significant
Terbinafine alters S. aureus membrane rigidity and increases bacterial susceptibility to membrane targeting antibiotics
FTIR spectroscopic examination of terbinafine treated S. aureus revealed significant alterations in the spectrum profile as compared with untreated bacteria (Fig. 5A). The variation in bands corresponding to membrane phospholipids (1600–1200 cm−1) and polysaccharides (1100–1000 cm−1) confirmed the inhibitory effect of terbinafine on S. aureus staphyloxanthin. In support of these findings, terbinafine treated S. aureus showed a higher susceptibility to polymxin B than untreated cells, indicating alteration of bacterial membrane integrity upon bacterial treatment with terbinafine (Fig. 5B).

A) The FTIR spectra of S. aureus ATCC 6538 cells and terbinafine treated cells within wavenumber 4000–500 cm−1. Control untreated and terbinafine-treated cells are represented by red and blue lines, respectively. B Polymxin B survival assay indicating increased sensitivity of terbinafine treated cells to polymxin B than untreated cells. Data shown represent the mean ± standard of error from triplicate experiments. Using Student’s t test, P value < 0.05 was considered statistically significant
Terbinafine possesses a potent anti-biofilm activity against S. aureus
Light and scanning electron microscopy (SEM) assessed the antibiofilm potential of terbinafine against S. aureus. Both light microscopy and SEM images of untreated cells revealed a firm established biofilm with a multilayer of aggregates and adherent cells (Fig. 6A, B, respectively). However, terbinafine treated cells showed a loose biofilm with scattered cells. Additionally, terbinafine showed a dose dependent biofilm inhibition on both polystyrene and polypropylene surfaces (Fig. 6C, D, respectively).

Terbinafine inhibited S. aureus biofilm formation in a dose dependent manner as observed from; light microscopic (A) and SEM (B) images. Quantitative crystal violet biofilm inhibition assay on; C polystyrene plate and D polypropylene tube upon terbinafine treatment. Data shown represent the mean ± standard of error from triplicate experiments. using Student’s t test, P value < 0.05 was considered statistically significant
Terbinafine diminishes cell auto-aggregation, surface hydrophobicity, and EPS production in S. aureus and enhances bacterial autolysis
In comparison with untreated S. aureus, terbinafine treated bacteria were highly dispersed and did not show any obvious aggregates (Fig. 7A). Additionally, both the hydrophobicity index and EPS production were significantly disrupted in terbinafine treated S. aureus cells as compared to untreated cells (Fig. 7B, C, respectively). Moreover, terbinafine treated S. aureus was more susceptible to induced autolysis than untreated bacteria. The initial optical density was significantly reduced to 64% ± 2.8 in terbinafine treated cells vs. 81% ± 2.2 in untreated cells (Fig. 8).

A Reduced auto-aggregation in terbinafine treated S. aureus ATCC 6538 in a dose dependent manner. B Hydrophobicity index of terbinafine treated cells was reduced in a dose dependent manner. C Inhibition of EPS in terbinafine treated cells compared to untreated cells. Data shown represent the mean ± standard of error from triplicate experiments. Using Student’s t test, P value < 0.05 was considered statistically significant

Terbinafine treated cells were more prone to Triton X-100 induced autolysis. Data shown represent the mean ± standard of error from triplicate experiments. Using Student’s t test, P value < 0.05 was considered statistically significant
Terbinafine does not interfere with DNase, esterase, or lipase activity
The influence of terbinafine treatment on S. aureus virulence enzymes, DNase, esterase, or lipase has been characterized. Both terbinafine-treated and untreated S. aureus exhibited clearance zones with equal diameters, indicating no change in DNase activity. Similarly, both terbinafine-treated and untreated S. aureus showed white precipitation zones with no difference in diameters indicating no significant effect on either esterase or lipase activity upon treatment of S. aureus with terbinafine (Data is not shown).
Terbinafine alters the expression of S. aureus virulence and biofilm genes
Quantitative RT-PCR showed that terbinafine does not affect the expression of staphyloxanthin biosynthesis genes, crtM and crtN. However, the expression of biofilm related genes such as sarA and icaA was significantly down-regulated and agr A was upregulated in S. aureus upon terbinafine treatment. Similarly, the expression of transcription regulators msaB and sig B was significantly repressed in terbinafine-treated S. aureus cells compared to untreated cells. On the other hand, the oxidative stress responsive gene (sodM) was upregulated in S. aureus following exposure to terbinafine (Fig. 9).

Transcriptional profile of S. aureus virulence genes upon terbinafine treatment. Quantitative RT-PCR revealed decreased expression of biofilm related genes and virulence regulators in terbinafine treated S. aureus relative to untreated bacteria. The data shown are the means ± standard errors from triplicate experiments with three technical replicates each. Using Student’s t test, P value < 0.05 was considered statistically significant
Terbinafine synergizes antibiotics against S. aureus
The synergistic inhibitory potential of terbinafine with commonly used antibiotics as ampicillin, cefotaxime, azithromycin, ciprofloxacin, and gentamicin against S. aureus was determined using the checkerboard assay. Importantly, terbinafine showed a distinct synergistic effect in combination with all tested antibiotics, with FICIs ranging from 0.14 to 0.31 (Table 2).
Terbinafine weakens S. aureus pathogenesis in vivo
Mice were infected with S. aureus (pigmented and terbinafine treated bacteria), and signs of bacterial infection were compared to mice injected with non-pigmented and untreated bacteria. Spleen, liver, and kidney isolated from mice infected with pigmented S. aureus were congested, exhibiting a significant increase in weight as compared to those mice injected with either terbinafine-treated or non-pigmented bacteria (Fig. 10A). Furthermore, the numbers of colonizing bacteria were determined in organs isolated from infected mice. Importantly, pigmented S. aureus colonized more significantly mice organs; spleen, liver, and kidney (5255 ± 11, 222,492 ± 23, 10,297 ± 16 CFUs/g, respectively) as compared to both terbinafine treated bacteria (506 ± 13, 1245 ± 11, 2137 ± 363.3 CFUs/g, respectively) and non-pigmented S. aureus (507 ± 11, 1506 ± 22, 1938 ± 362.8 CFUs/g, respectively) (Fig. 10B). Additionally, histopathological examination of organ sections isolated from mice infected with pigmented S. aureus revealed tissue inflammatory responses with irreversible tissue necrosis and fibrosis. On contrast, organs isolated from mice inoculated with either terbinafine treated or non-pigmented S. aureus showed normal tissues with minimal pathological changes (Fig. 10C).

Terbinafine altered S. aureus mice pathogenesis. A Increase in organ weight of pigmented bacteria inoculated mice compared to terbinafine treated and non-pigmented inoculated mice. B Bacterial load of liver, spleen and kidney of each group. C Histopathological organ section from pigmented, terbinafine treated and non-pigmented isolate stained by hematoxylin and eosin stain. (I) Liver with diffuse areas of caseous necrosis. (II) Degeneration of some renal tubules represented in cloudy swelling (III) Focal necrosis of some spleen lymphocytes in the white pulp. (IV) Normal hepatic parenchyma with normal tissue architecture and cellular details. (V) Focal cystic dilation of some renal tubules (arrowhead). (VI) Spleen with depleted white pulp lymphocytes. (VII) Hepatic blood vessels with diffuse congestion (arrows) and dilated sinusoids. (VIII) Kidney with diffused cystic dilation of some renal tubules in the renal medulla. (IX) Spleen with focal vacuolar degeneration. Each symbol represents the value for an individual mouse and horizontal bars indicate the means. Using Mann–Whitney U analysis, P value < 0.05 was considered statistically significant
Discussion
Staphylococcus aureus is a leading pathogen causing infections with high morbidity and mortality rates that impose a heavy financial burden on hospitals worldwide [1]. Antibiotic-resistant S. aureus is on the rise, highlighting the need for novel therapeutic strategies. Thus, introducing of antimicrobials that hinder S. aureus virulence without impeding bacterial growth has been proposed [5]. The current study shows, for the first time, fully detailed evidence for terbinafine anti-virulent activity against S. aureus. Our findings clearly demonstrate that terbinafine alleviates S. aureus staphyloxanthin production, biofilm formation, as well as host pathogenesis.
Staphyloxanthin is a carotenoid pigment that functions as an antioxidant, mediating bacterial survival under oxidative stress and is involved in host pathogenesis and disease progression [28]. Present data demonstrates that terbinafine possesses a dose dependent inhibition of staphyloxanthin in S. aureus. To precisely unveil the molecular mechanism of terbinafine, staphyloxanthin biosynthesis intermediates were extracted and quantified from S. aureus following treatment with terbinafine. Interestingly, terbinafine treatment led to the accumulation of CrtN substrates, 4, 4′-diapophytoene, and a low abundance of the subsequent intermediates, suggesting interference of terbinafine with CrtN. In line with current results, Chen et al. [19] reported that naftifine, an allylamine antifungal, has pigment inhibitory activity through targeting CrtN and attenuates S. aureus virulence. Importantly, qRT-PCR analysis indicates that terbinafine has no effect on the expression of either crtM or crtN. On the other hand, the molecular docking study performed herein validated terbinafine interaction with CrtN. Similarly, Ye et al. [51] has reported that a novel cationic peptide MSI-1 significantly inhibited staphyloxanthin production through binding with CrtN without any change in the expression of crtM or crtN. The molecular docking results strongly indicate that terbinafine inhibitory activity on staphyloxanthin is likely to be due to direct interference with CrtN enzyme activity.
As a consequence of staphyloxanthin inhibition, staphyloxanthin induced protective activity was disrupted and terbinafine treated S. aureus was more susceptible to both oxidative and acidic stress relative to untreated bacteria [28]. This higher susceptibility to stressful conditions suggests that S. aureus would be more sensitive to reactive oxygen species and acidic pH acting inside the phagolysosome, limiting S. aureus intracellular survival [52, 53]. Furthermore, terbinafine sensitized S. aureus cells to human blood cells, which could result in increased immune clearance by the host. In addition to inhibition of staphyloxanthin biosynthesis, the increased sensitization of S. aureus to stressful conditions upon terbinafine treatment could be attributed to the down regulation of sigB and msaB, which have been reported to be involved in S. aureus survival under various stresses [43, 54]. Pandely et al. demonstrated that msaABCR operon regulates an oxidative stress defense mechanism facilitating the persistence and recurrence of staphylococcal infections [55]. Accordingly, terbinafine would be valuable to tackle recurrent and persistent staphylococcal infections.
The decrease in S. aureus membrane fluidity has been linked to membrane staphyloxanthin content, elucidating the role of staphyloxanthin in regulating membrane stiffness [56]. FTIR analysis has been recently utilized for studying S. aureus membrane biophysical characters [57]. In the current study, the FTIR spectrum of terbinafine treated S. aureus showed changes in membrane lipids and polysaccharides comparable to untreated cells [29]. Since S. aureus has intrinsic resistance to membrane-targeting antibiotics, the altered biophysical properties of the bacterial membrane rendered terbinafine treated S. aureus more susceptible to polymxin B killing. Accordingly, Valliammai reported that thymol efficiently inhibited staphyloxanthin biosynthesis and senstizied S. aureus cells to polymyxin B [30]. Thus, these findings could suggest the use of staphyloxanthin inhibitors such as terbinafine in concurrent with membrane targeting antibiotics to offset S. aureus resistance.
The propensity to form biofilm is considered to be a chief virulence factor of several pathogens, including S. aureus, resulting in persistent and recurrent infections within the host [12]. S. aureus commonly contaminates indwelling medical devices. S. aureus is capable of forming biofilms on polystyrene and polypropylene plastic polymers that are routinely used in the manufacture of medical devices [13]. In the present study, terbinafine significantly reduced S. aureus ability to form biofilm on both polystyrene and polypropylene surfaces in a dose dependent manner. The naphthalene moiety incorporation has recently reported by Ghameshlouei et al. [58] to improve biofilm inhibitory activity of oxadiazole derivatives. Terbinafine antibiofilm activity can be explained by its repressor effect on sarA and msaB, which have been strongly implicated in S. aureus biofilm formation. Interestingly, the mutation of sarA and absence of msaABCR in S. aureus were found to limit biofilm formation and lead to defect in biofilm maturation, respectively [59, 60].
Among bacterial biofilms, bacterial cell auto-aggregation and surface hydrophobicity play a major role in the initial steps of biofilm formation. Current results demonstrate a reduction in both S. aureus auto-aggregation and surface hydrophobicity upon terbinafine treatment. This reduction in bacterial auto-aggregation would be advantageous as S. aureus auto-aggregation has been linked to the increased pathogenicity, drug resistance, and host immune evasion [61]. Sarker et al. [62] also reported that reduction in cell surface hydrophobicity could hinder microbial colonization of hydrophobic surfaces and, consequently, biofilm development. Furthermore, EPS serves as a barrier to shield bacteria from antimicrobial agents and increases biofilm resistance to mechanical forces such as fluid flow and shear stress inside the catheter [63]. EPS is a polysaccharide intercellular adhesin (PIA), which is encoded by the icaADBC operon. Terbinafine treatment showed a dose dependent reduction in EPS content and, subsequently, in biofilm buildup and maturation. These findings were further validated by the repression of icaA, sigB, and sarA, which play a role in PIA biosynthesis. In addition to the induction of icaR, which binds to DNA region upstream icaA, causing suppression of icaADBC operon transcription [64]. Notably, Yazdani et al. [65] and Fowler et al. [66] have found an association between expression of ica genes and biofilm formation ability.
Combination therapy has been introduced to overcome bacterial resistance to conventional antibiotics [67]. Upon terbinafine treatment, S. aureus exhibited an increased rate of cell death. This increased bacterial autolytic activity could be beneficial in potentiating the bactericidal activity of cell wall-active antibiotics [68]. The increase in S. aureus induced autolysis was explained by repression of msaB, which has been recently reported to regulate the rate of cell death [69]. Accordingly, the current data show a synergistic effect between terbinafine and β-lactam antibiotics ampicillin and cefotaxime. This synergism could be attributed to the functional membrane microdomain disassembly as a result of staphyloxanthin inhibition by terbinafine, which would interfere with PBP2a oligmerization, rendering S. aureus more susceptible to penicillin [70]. Moreover, msaB has been reported to control peptidoglycan cross linking, affecting S. aureus susceptibility to cell wall acting antibiotics like β-lactams. Antibiotics such as ciprofloxacin and gentamicin alter bacterial cellular respiration, resulting in a lethal level of intracellular damaging reactive species [71]. Therefore, the combination of terbinafine with ciprofloxacin or gentamicin would sensitize S. aureus to oxidative stress generated by both immune cells and antibiotics. Interestingly, Márió Gajdács has recently reported that terbinafine could be used as adjuvant as it efficiently reduced MIC value of ciprofloxacin [72]. These results clearly suggest the use of terbinafine as an adjuvant in combination therapy to overcome S. aureus resistance to conventional antibiotics.
Staphyloxanthin has been found to have a substantial role in host pathogenesis. The effect of terbinafine on S. aureus virulence was evaluated in vivo using mice infection model. Interestingly, terbinafine treated S. aureus exhibited a weakened virulence potential, causing fewer serious and reversible lesions compared to untreated S. aureus. This is in line with Chen et al. [19], who reported that naftafine attenuated S. aureus virulence in mouse infection models. This attenuated virulence could attributed to staphyloxanthin inhibition that rendered S. aureus more susceptible to neutrophils killing and innate immune inactivation in mouse infection model [73]. Additionally, Blevins et al., revealed that mutations in the virulence regulatory genes sigB and/or sarA reduced S. aureus capacity to cause in vivo pathogenesis [74, 75]. It has been shown that SigB plays an important role in S. aureus acclimation during chronic infections [76].
Conclusion
The current study demonstrates the anti-virulence potential of terbinafine against S. aureus. Terbinafine revealed significant anti-staphyloxanthin activity, sensitizing S. aureus to stressful conditions and host killing. Staphyloxanthin inhibition by terbinafine disrupted cell membrane permeability, leading to increased sensitivity to membrane targeting antibiotics. Additionally, terbinafine exhibited a dose dependent inhibition of S. aureus biofilm formation through interfering with the several stages of biofilm formation regarding cell autoaggregation, cell surface hydrophobicity, and EPS production. Interestingly, terbinafine synergized the activity of conventional antibiotics and alleviated S. aureus pathogenesis in the host. Terbinafine anti-virulent activity against S. aureus is multifactorial. In addition to its anti-pigment potential, terbinafine represses global virulence regulators in S. aureus, such as MsaB, SarA, and SigB. The findings of the current study are valuable and highlight the importance of terbinafine in the management of S. aureus infections, providing evidence that terbinafine could be repurposed as an anti-virulent agent against S. aureus.
Availability of data and materials
The authors confirm that the data supporting the findings of this study are available within the article.
Ethics approval and consent to participate
Experiments involving human participants were performed in accordance to the Declaration of Helsinki and approved by Zagazig University Institutional Review Board (ZU-IRB). All procedures in the animal study section were carried out in accordance with the ethical standards of the Zagazig University-Institutional Animal and Use Committee (ZU-IACUC), which was granted approval number (ZU-IACUC/3/F/158/2019).
Abbreviations
- MRSA:
-
Methicillin-resistant S. aureus
- VRSA:
-
Vancomycin-resistant
- PIA:
-
Polysaccharide intracellular adhesion
- ica:
-
Intracellular adhesion
- TSB:
-
Trypticase soya broth
- MIC:
-
Minimum inhibitory concentration
- MH:
-
Muller Hinton
- PBS:
-
Phosphate buffered saline
- TSA:
-
Trypticase soya agar
- MOE:
-
Molecular operating environment
- CFUs:
-
Colony forming units
- FTIR:
-
Fourier infrared spectroscopy
- CV:
-
Crystal violet
- SEM:
-
Scanning electron microscopy
- HI:
-
Hydrophobicity index
- EPS:
-
Exopolysaccharide
- DNase:
-
Deoxyribonuclease
- FICI:
-
Fractional inhibitory concentration index
References
Foster TJ. The Staphylococcus aureus “superbug.” J Clin Investig. 2004;114(12):1693–6.
Lowy FD. Staphylococcus aureus infections. N Engl J Med. 1998;339(8):520–32.
Turner NA, et al. Methicillin-resistant Staphylococcus aureus: an overview of basic and clinical research. Nat Rev Microbiol. 2019;17(4):203–18.
Shorr AF. Epidemiology of staphylococcal resistance. Clin Infect Dis. 2007;45(Suppl 3):S171–6.
Dickey SW, Cheung GYC, Otto M. Different drugs for bad bugs: antivirulence strategies in the age of antibiotic resistance. Nat Rev Drug Discov. 2017;16(7):457–71.
Popov I, et al. Study of the membranes of pigment-free mutant of Staphylococcus aureus. Biokhimiia. 1976;41(6):1116–20.
Mishra NN, et al. Carotenoid-related alteration of cell membrane fluidity impacts Staphylococcus aureus susceptibility to host defense peptides. Antimicrob Agents Chemother. 2011;55(2):526–31.
Clauditz A, et al. Staphyloxanthin plays a role in the fitness of Staphylococcus aureus and its ability to cope with oxidative stress. Infect Immun. 2006;74(8):4950–3.
Xue L, et al. Staphyloxanthin: a potential target for antivirulence therapy. Infect Drug Resist. 2019;12:2151–60.
Pelz A, et al. Structure and biosynthesis of staphyloxanthin from Staphylococcus aureus. J Biol Chem. 2005;280(37):32493–8.
Heilmann C. Adhesion mechanisms of staphylococci. Adv Exp Med Biol. 2011;715:105–23.
Parsek MR, Fuqua C. Biofilms 2003: emerging themes and challenges in studies of surface-associated microbial life. J Bacteriol. 2004;186(14):4427–40.
Percival SL, et al. Healthcare-associated infections, medical devices and biofilms: risk, tolerance and control. J Med Microbiol. 2015;64(Pt 4):323–34.
van Tilburg Bernardes E, Lewenza S, Reckseidler-Zenteno S. Current research approaches to target biofilm infections. Postdoc J. 2015;3(6):36–49.
Laverty G, et al. Ultrashort cationic naphthalene-derived self-assembled peptides as antimicrobial nanomaterials. Biomacromol. 2014;15(9):3429–39.
Chen Y-Y, et al. Novel naphthalimide aminothiazoles as potential multitargeting antimicrobial agents. ACS Med Chem Lett. 2017;8(12):1331–5.
Nolting S, Bräutigam M. Clinical relevance of the antibacterial activity of terbinafine: a contralateral comparison between 1% terbinafine cream and 0.1% gentamicin sulphate cream in pyoderma. Br J Dermatol. 1992;126(Suppl 39):56–60.
Rokade YB, Sayyed R. Naphthalene derivatives: a new range of antimicrobials with high therapeutic value. Rasayan J Chem. 2009;2:972–80.
Chen F, et al. Small-molecule targeting of a diapophytoene desaturase inhibits S. aureus virulence. Nat Chem Biol. 2016;12(3):174–9.
Wikler MA. Methods for dilution antimicrobial susceptibility tests for bacteria that grow aerobically: approved standard. CLSI (NCCLS). 2006;26:M7-A7.
Gargotti M, et al. Comparative studies of cellular viability levels on 2D and 3D in vitro culture matrices. Cytotechnology. 2018;70(1):261–73.
Kossakowska-Zwierucho M, et al. Factors determining Staphylococcus aureus susceptibility to photoantimicrobial chemotherapy: RsbU activity, staphyloxanthin level, and membrane fluidity. Front Microbiol. 2016;7:1141–1141.
Wang Y, et al. Discovery of potent benzofuran-derived diapophytoene desaturase (CrtN) inhibitors with enhanced oral bioavailability for the treatment of methicillin-resistant Staphylococcus aureus (MRSA) infections. J Med Chem. 2016;59(7):3215–30.
Leejae S, Hasap L, Voravuthikunchai SP. Inhibition of staphyloxanthin biosynthesis in Staphylococcus aureus by rhodomyrtone, a novel antibiotic candidate. J Med Microbiol. 2013;62(3):421–8.
McGuffin LJ, et al. IntFOLD: an integrated web resource for high performance protein structure and function prediction. Nucleic Acids Res. 2019;47(W1):W408–13.
Inc, C.C.G., Molecular operating environment (MOE) version 2019.0102. 2019, Chemical Computing Group Inc. 1010 Sherbooke St. West, Suite# 910, Montreal
Labute P. The generalized Born/volume integral implicit solvent model: Estimation of the free energy of hydration using London dispersion instead of atomic surface area. J Comput Chem. 2008;29(10):1693–8.
Liu GY, et al. Staphylococcus aureus golden pigment impairs neutrophil killing and promotes virulence through its antioxidant activity. J Exp Med. 2005;202(2):209–15.
Wongthong S, et al. Attenuated total reflection: Fourier transform infrared spectroscopy for detection of heterogeneous vancomycin-intermediate Staphylococcus aureus. World J Microbiol Biotechnol. 2020;36(2):22.
Valliammai A, et al. Staphyloxanthin inhibitory potential of thymol impairs antioxidant fitness, enhances neutrophil mediated killing and alters membrane fluidity of methicillin resistant Staphylococcus aureus. Biomed Pharmacother. 2021;141: 111933.
Stepanovic S, C irkovic, I. and Ruzicka, F., et al. Quantification of biofilm in microtiter plates: overview of testing conditions and practical recommendations for assessment of biofilm production by staphylococci. APMIS. 2007;115(8):891–9.
Gowrishankar S, Duncun Mosioma N, Karutha Pandian S. Coral-associated bacteria as a promising antibiofilm agent against methicillin-resistant and -susceptible Staphylococcus aureus biofilms. Evid Based Complement Alternat Med. 2012;2012:862374.
Sorroche FG, et al. A positive correlation between bacterial autoaggregation and biofilm formation in native Sinorhizobium meliloti isolates from Argentina. Appl Environ Microbiol. 2012;78(12):4092–101.
Nostro A, et al. Modifications of hydrophobicity, in vitro adherence and cellular aggregation of Streptococcus mutans by Helichrysum italicum extract. Lett Appl Microbiol. 2004;38(5):423–7.
Favre-Bonté S, Köhler T, Van Delden C. Biofilm formation by Pseudomonas aeruginosa: role of the C4-HSL cell-to-cell signal and inhibition by azithromycin. J Antimicrob Chemother. 2003;52(4):598–604.
Manna C, et al. The protective effect of the olive oil polyphenol (3, 4-dihydroxyphenyl)-ethanol counteracts reactive oxygen metabolite-induced cytotoxicity in Caco-2 cells. J Nutr. 1997;127(2):286–92.
Jeffries CD, Holtman DF, Guse DG. Rapid method for determining the activity of microorganisms on nucleic acids. J Bacteriol. 1957;73(4):590–1.
Plou F, et al. Analysis of Tween 80 as an esterase/lipase substrate for lipolytic activity assay. Biotechnol Tech. 1998;12:183–6.
Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods. 2001;25(4):402–8.
Silva LN, et al. Myricetin protects Galleria mellonella against Staphylococcus aureus infection and inhibits multiple virulence factors. Sci Rep. 2017;7(1):1–16.
Sapp AM, et al. Contribution of the nos-pdt Operon to virulence phenotypes in methicillin-sensitive Staphylococcus aureus. PLoS ONE. 2014;9(10): e108868.
Bai J-R, et al. Antibiofilm activity of shikimic acid against Staphylococcus aureus. Food Control. 2018;95:327–33.
Sahukhal GS, Elasri MO. Identification and characterization of an operon, msaABCR, that controls virulence and biofilm development in Staphylococcus aureus. BMC Microbiol. 2014;14(1):154.
Engman J, et al. The YjbH adaptor protein enhances proteolysis of the transcriptional regulator Spx in Staphylococcus aureus. J Bacteriol. 2012;194(5):1186–94.
Oogai Y, Kawada-Matsuo M, Komatsuzawa H. Staphylococcus aureus SrrAB affects susceptibility to hydrogen peroxide and co-existence with Streptococcus sanguinis. PLoS ONE. 2016;11(7): e0159768.
Nakonieczna J, et al. Superoxide dismutase is upregulated in Staphylococcus aureus following protoporphyrin-mediated photodynamic inactivation and does not directly influence the response to photodynamic treatment. BMC Microbiol. 2010;10:323.
Ooi N, et al. Redox-active compounds with a history of human use: antistaphylococcal action and potential for repurposing as topical antibiofilm agents. J Antimicrob Chemother. 2014;70(2):479–88.
European Committee for Antimicrobial Susceptibility Testing (EUCAST) of the European Society of Clinical Microbiology and Infectious Dieases (ESCMID). Terminology relating to methods for the determination of susceptibility of bacteria to antimicrobial agents. Clin Microbiol Infect. 2000;6(9):503–8.
Deshmukh HS, et al. Critical role of NOD2 in regulating the immune response to Staphylococcus aureus. Infect Immun. 2009;77(4):1376–82.
Singh VK, et al. Significance of four methionine sulfoxide reductases in Staphylococcus aureus. PLoS ONE. 2015;10(2): e0117594.
Ye X, et al. Combats drug-resistant S. aureus by affecting bacterial viability and inhibiting carotenoid pigment production. Microbiol Res. 2022;255:126909.
Mandell GL. Bactericidal activity of aerobic and anaerobic polymorphonuclear neutrophils. Infect Immun. 1974;9(2):337–41.
Lang S, et al. Identification of a novel antigen from Staphylococcus epidermidis. FEMS Immunol Med Microbiol. 2000;29(3):213–20.
Chan PF, et al. The Staphylococcus aureus alternative sigma factor sigmaB controls the environmental stress response but not starvation survival or pathogenicity in a mouse abscess model. J Bacteriol. 1998;180(23):6082–9.
Pandey S, Sahukhal GS, Elasri MO. The msaABCR operon regulates the response to oxidative stress in Staphylococcus aureus. J Bacteriol. 2019;201(21):e00417-e419.
Seel W, et al. Carotenoids are used as regulators for membrane fluidity by Staphylococcus xylosus. Sci Rep. 2020;10(1):330.
López G-D, et al. Carotenogenesis of Staphylococcus aureus: new insights and impact on membrane biophysical properties. Biochimica et Biophysica Acta (BBA) Mol Cell Biol Lipids. 2021;1866(8):158941.
Ghameshlouei S, Zarrabi Ahrabi N, SarveAhrabi Y. In vitro and In Silico evaluation of biological activity of a new series of oxadiazole compounds against Esp gene expression in Enterococcus faecalis biofilm. Gene, Cell and Tissue. 2021. https://doi.org/10.5812/gct.112403.
Sahukhal GS, Batte JL, Elasri MO. msaABCR operon positively regulates biofilm development by repressing proteases and autolysis in Staphylococcus aureus. FEMS Microbiol Lett. 2015;362(4):1–10.
Beenken KE, Blevins JS, Smeltzer MS. Mutation of sarA in Staphylococcus aureus limits biofilm formation. Infect Immun. 2003;71(7):4206–11.
Trunk T, Khalil HS, Leo JC. Bacterial autoaggregation. AIMS microbiology. 2018;4(1):140.
Sarker RK, et al. Degradation of low-density poly ethylene (LDPE) by Enterobacter cloacae AKS7: a potential step towards sustainable environmental remediation. Arch Microbiol. 2020;202(8):2117–25.
McCarthy H, et al. Methicillin resistance and the biofilm phenotype in Staphylococcus aureus. Front Cell Infect Microbiol. 2015;5:1–1.
Joo H-S, Otto M. Molecular basis of in vivo biofilm formation by bacterial pathogens. Chem Biol. 2012;19(12):1503–13.
Yazdani R, et al. Detection of icaAD gene and biofilm formation in Staphylococcus aureus isolates from wound infections. Iranian J Public Health. 2006;35:25–8.
Fowler VG Jr, et al. The intercellular adhesin locus ica is present in clinical isolates of Staphylococcus aureus from bacteremic patients with infected and uninfected prosthetic joints. Med Microbiol Immunol. 2001;189(3):127–31.
Bulusu KC, et al. Modelling of compound combination effects and applications to efficacy and toxicity: state-of-the-art, challenges and perspectives. Drug Discov Today. 2016;21(2):225–38.
Trotonda M, et al. Role of mgrA and sarA in methicillin-resistant Staphylococcus aureus autolysis and resistance to cell wall-active antibiotics. J Infect Dis. 2009;199(2):209–18.
BibekSahukhal CGGS, Elasri MO. Role of the msaABCR operon in cell wall biosynthesis, autolysis, integrity, and antibiotic resistance in Staphylococcus aureus. Antimicrob Agents Chemother. 2019;63(10):e00680-e719.
García-Fernández E, et al. Membrane microdomain disassembly inhibits MRSA antibiotic resistance. Cell. 2017;171(6):1354-1367.e20.
Dwyer DJ, et al. Antibiotics induce redox-related physiological alterations as part of their lethality. Proc Natl Acad Sci. 2014;111(20):E2100–9.
Gajdács M. Non-antibiotic pharmaceutical agents as antibiotic adjuvants. Acta Biologica Szegediensis. 2020;64:17–23.
Liu C-I, et al. A cholesterol biosynthesis inhibitor blocks Staphylococcus aureus virulence. Science. 2008;319(5868):1391–4.
Blevins JS, et al. Role of sarA in the pathogenesis of Staphylococcus aureus musculoskeletal infection. Infect Immun. 2003;71(1):516–23.
Kullik I, Giachino P, Fuchs T. Deletion of the alternative sigma factor sigmaB in Staphylococcus aureus reveals its function as a global regulator of virulence genes. J Bacteriol. 1998;180(18):4814–20.
Tuchscherr L, et al. Sigma factor SigB is crucial to mediate Staphylococcus aureus adaptation during chronic infections. PLoS Pathog. 2015;11(4): e1004870.
Funding
Open access funding provided by The Science, Technology & Innovation Funding Authority (STDF) in cooperation with The Egyptian Knowledge Bank (EKB). Not applicable.
Author information
Authors and Affiliations
Contributions
Conceptualization, MA, and FY; methodology, MA, and FY; docking analysis, BM; validation, MA, and FY; investigation, MA, and FY; data curation, MA, and FY; writing-original draft preparation, MA, and FY; writing-review and editing, MA and FY; visualization, MA; FY and NY; supervision, MA and NY; All authors have read and approved the final manuscript.
Corresponding author
Ethics declarations
Consent for publication
Written informed consent for publication was obtained.
Competing interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Askoura, M., Yousef, N., Mansour, B. et al. Antibiofilm and staphyloxanthin inhibitory potential of terbinafine against Staphylococcus aureus: in vitro and in vivo studies. Ann Clin Microbiol Antimicrob 21, 21 (2022). https://doi.org/10.1186/s12941-022-00513-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12941-022-00513-7