
Abstract
Clinical trials and studies have implicated that E3 ubiquitin ligase BTBD3 (BTB Domain Containing 3) is a cancer-associated gene. However, the role and underlying mechanism of BTBD3 in colorectal cancer (CRC) is not fully understood yet. Herein, our study demonstrated that the mRNA and protein levels of BTBD3 were decreased in CRC tissues and associated with TYPO3 and Wnt/β-catenin pathway. Our results showed that circRAE1 knockdown and TYRO3 overexpression activated Wnt/β-catenin signaling pathway and the EMT process-associated markers, indicating that circRAE1/miR-388-3p/TYRO3 axis exacerbated tumorigenesis of CRC by activating Wnt/β-catenin signaling pathway. In addition, overexpression of BTBD3 reduced CRC cell migration and invasion in vitro and inhibited tumor growth in vivo. Our data demonstrated that BTBD3 suppressed CRC progression through negative regulation of the circRAE1/miR-388-3p/TYRO3 axis and the Wnt/β-catenin pathway. Our data further confirmed that BTBD3 bound and ubiquitinated β-catenin and led to β-catenin degradation, therefore blocked the Wnt/β-catenin pathway and suppressed the CRC tumorigenesis. This study explored the mechanism of BTBD3 involved in CRC tumorigenesis and provided a new theoretical basis for the prevention and treatment of CRC.
Similar content being viewed by others


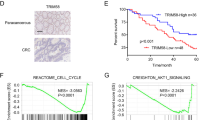
Introduction
Colorectal cancer (CRC) is one of the most common malignant cancers worldwide [1, 2]. Recurrence and metastasis of CRC lead to poor survival rate [3,4,5], and its pathogenesis remains to be fully elucidated. Recent studies implicate that circRNA is involved in the progression of cancer [6]. circRNA is a long non-coding RNA with a closed circular structure, formed by connecting the 3 ‘and 5’ ends. For example, circRNA_0000392 could act as a potential therapeutic target for the treatment of CRC by regulating the miR-193a-5p/PIK3R3/Akt axis [7]. In addition, circPACRGL promoted the proliferation, migration and invasion of CRC cells via miR-142-3p/miR-506-3p-TGF-β1 axis. A previous study has shown that the circular RNA has_circ_0060967 (circRAE1) promotes the development of CRC by modulating the miR-338-3p/TYRO3 axis [8]. Tyrosine kinase receptors TYRO3 is a member of the TYRO3-AXL-MER (TAM) family [9], which promotes cell growth and metastasis in CRC [10], breast cancer [11] and other malignancies [12]. In gastric cancer, TYRO3 promoted the development of cancer by activation of the Wnt/β-catenin signaling [13].
The canonical Wnt signaling pathway plays a notable role in the occurrence and development of CRC [14,15,16]. β-catenin, glycogen synthase kinase 3β (GSK3β), adenomatous polyposis coli (APC) and scaffolding protein 1 (AXIN1) are key proteins in Wnt/β-catenin signaling pathway [17,18,19], AXIN1 and APC promote the interaction between GSK3β and β-catenin [17]. Based on our bioinformatic search, it indicated that BTBD3 may be involved in Wnt signaling pathway. BTBD3 protein includes the BTB (BR-C, ttk and bab)/POZ (Pox virus and Zinc finger) and PHR (PAM, highwire, and RPM) domain. The BTB domain may function as substrate-specific adaptors of a Cullin E3 ubiquitin-protein ligase complex [20]. It also may belong to a family of putative transcription factors with essential roles in development, differentiation, and oncogenesis [21]. It has two splice variants coding for 482 and 385 amino acids, respectively. The role and underlying mechanism of BTBD3 on Wnt/β-catenin pathway and CRC is still limited at present [22]. Clinical trials and studies have implicated that BTBD3 is a cancer-associated gene [23, 24]. It also found that BTBD3 was upregulated in hepatocellular carcinoma tissues and cell lines, and promoted the invasion ad metastasis of cancer cells [23]. Zhang et al. predicted that BTBD3 was one of the targets of hsa-let-7i which played an important role in the occurrence and development of CRC and the expression of BTBD3 has clinical diagnostic value for cancer [23].
Based on the previous study, our study explored the role and underlying mechanism of BTBD3 in CRC in vitro and in vivo. Our study investigated the protein interaction between β-catenin, BTBD3 and TYRO3 and further elucidated that the TYRO3/Wnt/β-catenin pathway in CRC was regulated by BTBD3-mediated ubiquitination in vitro and in vivo. This study explored the mechanism of BTBD3 involved in CRC tumorigenesis and provided a new theoretical basis for the prevention and treatment of CRC.
Materials and methods
Materials
MCCOY’S 5 A, RPMI Medium 1640, BCA Protein Assay Kit, SDS-PAGE loading buffer, Crystal violet, Anti-Vimentin Polyclonal antibody were bought from Solaibao Biological Technology Co., Ltd. β-actin antibody, Anti-beta Catenin antibody, Anti-APC Antibody, Anti-TYRO3 Antibody, Anti-BTBD3 Antibody, Anti-Ubiquitin (linkage-specific K63) were purchased from Abcam. Anti-E-cadherin Polyclonal antibody and AXIN1 Polyclonal Antibody were obtained from Proteintech. HRP tagged goat anti-rabbit reagent, HRP tagged donkey anti-goat reagent, HRP tagged goat anti-mouse reagent and HRP tagged goat anti-rat reagent were purchased from Servicebio Biotechnology Co., Ltd.
Tissue collection and receiver operating characteristic (ROC) analysis
30 cancer and adjacent tissue samples from colorectal cancer (CRC) patients after surgery were collected from Jan 1, 2020 to Dec 31, 2022. The diagnosis of colorectal cancer accorded to National guidelines for diagnosis and treatment of colorectal cancer 2015 in China and the pathological staging of colorectal cancer accorded to the TNM staging system developed by AJCC/UICC (American Cancer Council/Union for International Cancer Control). All enrolled patients were informed and provided the informed consent. This research protocol (NO: FJMUSAH2021-69) was reviewed and approved by the Ethics Committee of Fujian Medical University. The area under the ROC curve is a widely used performance metric for the binary classifier system being evaluated. We used ROC analysis to evaluate whether BTBD3 has clinical diagnostic value for CRC.
Cell lines and cell culture
Colorectal cancer cell lines HCT116, SW620, HT29 and SW480 were obtained from CTCC. HCT116 and HT29 cells were cultured in MCCOY’S 5 A supplemented with 10% fetal calf serum and 1% P/S solution. SW620 and SW480 cells were cultured in 1640 containing 10% fetal calf serum and 1% P/S solution. NCM460 cells were cultured in DMEM medium containing 10% fetal calf serum and 1% P/S solution. Cell lines were cultured at 37 °C with 5% CO2 incubator.
Cell transfection
The knockdown for circRAE1 (sh-circRAE1), overexpression-TYRO3 (oe-TYRO3), overexpression-BTBD3 (oe-BTBD3), oe-negative control (oe-NC) and shRNA-negative control (sh-NC) were obtained from Zolgene Biotechnology Co., Ltd. The sequences of shRNAs were as follows: circRAE1-shRNA1-F: 5’-GATCCGGATCAAAGCTCCAAACTATTCAAGAGATAGTTTGGAGCTTTGATCCTTTTTG-3’; circRAE1-shRNA1-R:5’- AATTCAAAAAGGATCAAAGCTCCAAACTATCTCTTGAATAGTTTGGAGCTTTGATCCG-3’.
RNA isolation and RT-qPCR
Total RNAs were extracted using a Trizol reagent. Then, the RNA was reversely transcribed using the First-Strand cDNA Synthesis kit according to the manufacturer’s instructions. The relative expression level of β-actin, circRAE1, TYRO3, miR-338-3P, BTBD3, and β-catenin mRNA was quantified using real-time-PCR with SYBR Green RT-QPCR Kit. The expression of target genes was determined through the comparative 2−ΔΔCt method. Primer sequences of target genes were as used as follows in Table 1.
Western blot analysis
The cells were lysed by cold RIPA buffer containing phenylmethanesulfonyl fluoride (PMSF) for 20 min on ice. The proteins were collected and sample concentrations were detected by BCA Kit. Electrophoresis, blotting and blocking were performed as previously described, then, the membranes were incubated with primary antibodies (β-actin 1:5000, Vimentin 1:2000, E-cadherin 1:5000, GSK3β 1:2000, APC 1:1000, AXIN1 1:500, TYRO3 1:1000, β-catenin 1:1000) at 4 ℃ overnight, incubated for 2 h with a secondary antibody at room temperature, the signals were measured using Versa DocTM imaging system.
CCK-8 assay
The cells in the logarithmic growth stage were plated in 96-well plates at a density of 4 × 104 cells/well and cultured for 0, 24, 48 and 72 h. The viability of HT29 and SW480 cells was determined using a CCK-8 kit, and the cells were cultured in an incubator at 37 °C with 5% CO2. After 1 h, the absorbance at 450 nm was determined by a microplate reader.
Plate cloning experiment
Cells in the logarithmic growth phase were digested using trypsin to prepare a single cell suspension and cultured in a 6-well plate (800 cells per well). The cells were incubated at 37 °C, 5% CO2. The culturing was terminated when macroscopic clones were observed in the culture plates, and the cells were carefully washed 1 time with phosphate-buffered saline (PBS). The cells were fixed with polyoxymethylene (4%) for 60 min at 4 ℃, washed 1 time with PBS, and stained with crystal violet solution for 2 min. The colony formation was observed and photographed under a microscope.
Cell migration and invasion assays
To assess cell migration, HT29 or SW480 cells (1 × 105 cells/well) were added to the Transwell inserts in a 24-well plate and the cells were cultured in 5% CO2 at 37 °C for 24 h. The cells in the lower compartment were stained with crystal violet for 15 min. Finally, the migrated cells were examined using a microscope. For the Transwell invasion assay, cells (1 × 105 cells/well) were plated in the Transwell upper chamber, which precoated the Matrigel for 2 h at 37 °C. The following steps were the same as those used in the Transwell migration assay.
Co-immunoprecipitation (Co-IP)
The cell suspension was collected and centrifuged. The supernatant was used for Co-IP. GammaBind Protein A/G Sepharose (GE Healthcare, 20 µL) was added to the supernatant and incubated for 1 h at 4 °C, which added 0.2–2 µg of antibody. The supernatant was incubated at 4 °C for overnight and then added to GammaBind Protein A/G Sepharose beads (80 µL). After 2 h, the supernatant was centrifuged and aspirated, 1 mL of lysate was added, and the beads were washed three times. Then, 80 µL of loading buffer was added, and the samples were heated at 100 °C for 10 min, followed by WB analysis.
Fluorescence in situ hybridization (FISH) experiment
The cell sliver was fixed in 4% paraformaldehyde for 20 min and then digested with proteinase K (20 µg·mL− 1) for 8 min. FITC-labeled probe and Cy3-labeled probe were added into each well. Cell nuclei were stained by 4, 6-diamidino-2-phenylindole (DAPI), followed by being observed under fluorescence microscopy (ECLIPSE NI, Nikon, Japan). The probe TYRO3 had the sequence of CATGACACTGTCAAAGGCTTGCCCAAA, the probe BTBD3 had the sequence of CTCTAATGGTGGGATAAAGACCCTGCC and the probe β-catenin had the sequence of TGAAGGACTGAGAAAATCCCTGTTCCC.
In vivo ubiqutination assay
293T Cells were transfected with siNC or SiBTBD3 + flag-β-catenin + HA-ub for 42 h and then 10 μm MG132 were added to the samples and further incubated for 6 h. Cells were collected and lysed with RIPA buffer. 40uL lysate was saved from each sample and the left samples were mixed with 1 µg control IgG and anti-Flag M2 antibody and rotated at 4oC overnight. Protein A/G beads were added to the IP samples and the samples were rotated for an additional 3 h. Input and pulldown IP complexes are boiled with SDS loading dye for 10 min and processed to WB. IP samples were probed with anti-HA antibody to detect ubiqutination and input was probed with anti-FLAG, anti-BTBD3 and anti-actin antibody.
Protein half-life assay with cycloheximide
6 × 105 cells in 35-mm dishes were transfected with Si-NC or SiBTBD3 for 24 h and then added with 300 µg/ml* cycloheximide (dissolved in DMSO) into each dish and further incubated for 0, 6 12 h. Cells were collected, lysed and processed to WB with anti-β-catenin, anti-BTBD3 and anti-actin antibody.
In vivo tumorigenesis and metastasis assays
Sixty BALB/c nude mice (4–6 weeks) were divided equally into six groups: NC group, sh-circRAE1 group, oe-BTBD3 group, sh-circRAE1 + LV-TYRO3 group, sh-circRAE1 + miR-338-3P inhibitors group, oe-BTBD3 + LV-TYRO3 group and oe-BTBD3 + oe-β-catenin group. SW480 cells (1 × 107 cells, 0.2 mL) were injected subcutaneously into the right flank region of the nude mice. At 21 days, the tumors were removed completely and weighed. Tumor volume (V) was calculated by the formula (width2 × length)/2.
The Colon and rectum tissue were fixed with formalin, and after dehydration with ethanol and xylene, samples were embedded in paraffin and sectioned for hematoxylin-eosin (HE) staining.
Statistical analysis
All the experiments were performed in three independent assays. All graphical values were presented as the mean ± standard error of the mean (SEM). Statistical analyses were performed using GraphPad Prism 6 (GraphPad Software, Inc., La Jolla, CA). Correlation analysis between BTBD3, TYRO3 and β-catenin was performed by Pearson correlation. P < 0.05 was considered to indicate a statistically significant difference.
Results
BTBD3 is associated with the progression of patients with CRC
To clarify the significance of BTBD3 in CRC, the analysis of clinicopathological features of 30 patients showed that BTBD3 was significantly related to the TNM stage (P < 0.05) (Table 2). ROC analysis result showed that the area under the curve (AUC) is 0.6543 > 0.5 (95% CI: 0.5180–0.7906; P < 0.05) with a sensitivity and specificity of 62.5% and 59.4%, respectively (Fig. 1A), indicating that BTBD3 has clinical diagnostic value for in CRC progression and metastasis. The relationship between BTBD3, TYRO3 and β-catenin expression in CRC tissues was analyzed using the Pearson correlation coefficient, this result showed that BTBD3 was negatively correlated with TYRO3 and β-catenin (Fig. 1B, C), while TYRO3 was positively correlated with β-catenin (Fig. 1D). This finding indicates that BTBD3 may be targeted TYRO3 and β-catenin.

The ROC analysis (A) and the relationship of BTBD3 and TYRO3 (B), BTBD3 and β-catenin (C), TYRO3 and β-catenin (D) expression in CRC tissues was analyzed using Pearson correlation coefficient. RT-qPCR was applied to detect circRAE1 (E), BTBD3 (F), TYRO3 (G) and β-catenin (H) expression in 30 pairs of CRC tissues and the adjacent non-tumor tissues. Western blotting was also used to detect the protein expression levels of β-catenin and BTBD3 in CRC tissues and the adjacent non-tumor tissues. (I, J) quantification of β-catenin (I) and BTBD3 (J) protein expression in WB by ImageJ software. (K) representative image of WB for β-catenin, BTBD3 and action
To further verify the role of BTBD3 in CRC progression, the mRNA expression levels of circRAE1, TYRO3, β-catenin and BTBD3 were examined in 30 pairs of CRC tissues and the adjacent non-tumor tissues obtained from the Second Affiliated Hospital of Fujian Medical University by RT-qPCR. The results showed that the expressions of circRAE1 (Fig. 1E), TYRO3 (Fig. 1F) and β-catenin (Fig. 1G) were significantly up-regulated while the expression of BTBD3 (Fig. 1H) was significantly downregulated in tumor tissues. The results of WB show the similar results (Fig. 1I-K). Compared with the normal tissue, tumor tissues significantly increased the β-catenin expression, while the protein expression levels of BTBD3 were significantly reduced. This suggested that BTBD3 may have a protective effect on tumorigenesis through negatively regulating Wnt/β-catenin signaling pathway.
Roles of BTBD3 in CRC cells
BTBD3 belongs to is one E3 Ubiquitin ligase and we examine whether BTBD3, like other E3 Ubiquitin ligase, regulates the occurrence and development of CRC by regulating Wnt signaling pathway. To investigate the roles of BTBD3 in CRC cells, we first detect the mRNA expression of BDBT3 in HT29 and SW480 by RT-qPCR and it showed that BDBT3 mRNA in CRC cell lines was significantly lower than in control cell line NCM460 cells (Fig. 2A). In contrast, the β-catenin expression of HT29 and SW480 was significantly higher than that of other cell lines (Fig. 2B), indicating that the expression of BDBT3 was negatively correlated with the occurrence and development of tumors.

The mRNA expression of BDBT3 (A) and β-catenin (B) in HCT116, SW620, HT29, SW480 and NCM460 cells as detected by RT-qPCR. Overexpression of BDBT3 suppressed HT29 and SW480 cell proliferation ability as determined by CCK8 (C) and colony formation assays (D). Overexpression of BTBD3 reduced cell migration (E) and invasion (F) detected by Cell migration and invasion assay
Compared with the oe-NC group, the cell proliferation ability (Fig. 2C) and cloning ability (Fig. 2D) were significantly decreased in the oe-BTBD3 group. To verify the effect of BTBD3 on the migration and invasion of HT29 and SW480 cells, we performed Transwell assays. The oe-BTBD3 group decreased the number of migratory (Fig. 2E) and invasive (Fig. 2F) cells compared with the oe-NC group, indicating that BTBD3 inhibits the occurrence and development of tumors.
The circRAE1/miR-388-3p/TYRO3 axis affected the development of CRC by regulating the Wnt/β-catenin pathway
To verify the effect of sh-circRAE1 and oe-TYRO3 treatment on the circRAE1/miR-388-3p/TYRO3 axis, the mRNA expression levels of circRAE1, miR-338-3p, TYRO3 were detected by RT-qPCR. Our data indicated that sh-CircRAE1 treatment significantly decreased the expression levels of circRAE1 (Fig. 3A), increased the expression level of miR-338-3p (Fig. 3B) and decreased the mRNA expression levels of TYRO3 (Fig. 3C) in HT 29 and SW480 Cells (sh-CircNC + oe-TYPO3 vs. sh-CircRAE1 + oe-TYPO3). Overexpression of TYPO3 (sh-CircRAE1 + oe-NC vs. sh-CircRAE1 + oe-TYPO3) increased circRAE1 (Fig. 3A), decreased miR-338-3p (Fig. 3B) in HT 29 and SW620 Cells. This result demonstrated that circRAE1 and TYRO3 negatively regulated the mRNA expression levels of miR-338-3p.

RT-qPCR was used to detect the mRNA expression levels of circRAE1 (A), miR-338-3p (B) and TYRO3 (C) after circRAE1 knockdown and TYRO3 overexpression. Significant differences between groups are indicated as *P < 0.05, **P < 0.01, ***P < 0.001 (n = 3)
To observe a potential relationship between the expression of TYRO3, circRAE1 and the Wnt/β-catenin pathway, the protein expression levels of E-cadherin, Vimentin, TYRO3, GSK3β, AXIN1, APC and β-catenin were analyzed by WB in HT 29 and SW480 Cells (Fig. 4). The protein level of E-cadherin was significantly up-regulated by sh-CirRAE1(sh-CircNC + oe-TYOP3 vs. sh-CircRAE1 + oe-TYPO3) but down-regulated by overexpression of TYRO3 (sh-CircRAE1 + oe-NC vs. sh-CircRAE1 + oe-TYPO3), while the effect on vimentin was the opposite (Fig. 4), indicating that CircRAE1 and TYRO3 is involved in the EMT process. Studies have shown that the Wnt/β-catenin signaling pathway can mediate TYRO3-induced EMT [13], in this study, we further observed the effect of CircRAE1 and TYRO3 on the Wnt/β-catenin pathway (GSK3β, AXIN1, APC and β-catenin). Our data showed the protein expression levels of APC were significantly increased while the levels of GSK3β, AXIN1 and β-catenin were decreased significantly in the sh-circRAE1 + oe-TYRO3 group compared with the sh-circNC + oe-TYRO3 group. The protein level of APC was decreased, the levels of GSK3β, AXIN1 and β-catenin increased significantly in the sh-circRAE1 + oe-TYRO3 group compared with the sh-circRAE1 + oe-NC group. The result revealed that circRAE1 and TYRO3 oppositely affected the EMT process and the Wnt/β-catenin signaling pathway.

The relative expressions of E-cadherin, Vimentin, TYRO3, GSK3β, AXIN1, APC and β-catenin proteins in the HT29 cells (A) and SW480 cells (B) detected by western blotting. Right panel was quantification for WB by ImageJ
TYRO3 interacted with β-catenin in HT29 cells and SW620 cells
To verify how TYRO3 regulates the Wnt/β-catenin pathway, we detected the interaction between TYRO3 and β-catenin (a negative regulator of Wnt signaling [25]) by Co-IP studies and FISH (Fig. 5). The Co-IP results revealed that the TYRO3 and β-catenin coexist in the same complex (Fig. 5A) and oe-BTBD3 promoted the binding. The FISH experiment showed that TYRO3 and β-catenin were mainly colocalized in the cytoplasm (Fig. 5B), which further indicated that TYRO3 might regulate the Wnt signal through the interaction of β-catenin.

Co-IP assay for endogenous TYRO3 and β-catenin in HT29 cells (A) with oe-BTBD3. Representative FISH images of TYRO3 (green) and β-catenin (red) in HT29 cells (B) and SW480 cells (C). Scale bar: 20 µM
Effects of BTBD3 overexpression on the circRAE1/TYRO3 axis and Wnt/β-catenin pathway
To explore the effect of BTBD3 on the circRAE1/TYRO3 axis and Wnt/β-catenin pathway in CRC, the overexpression of BTBD3 was applied. Compared with the oe-NC group, the mRNA expression of circRAE1, TYRO3, β-catenin, GSK3β and AXIN1 were significantly decreased in the oe-BTBD3 group in HT29 (Fig. 6A) and SW480 (Fig. 6B). We further observed the protein expression levels of TYRO3, β-catenin, GSK3β and AXIN1 in the oe-BTBD3 group were significantly decreased compared to the oe-NC group in HT29 (Fig. 6C) and SW480 (Fig. 6D). Our data demonstrated that BTBD3 suppressed CRC progression through negative regulation of circRAE1/miR-388-3p/TYRO3 axis and the Wnt/β-catenin pathway.

The mRNA expression levels of circRAE1, TYRO3, GSK3β, AXIN1 and β-catenin in HT29 (A) and SW480 (B) detected by RT-qPCR. The protein expression levels of TYRO3, BTBD3, GSK3β, AXIN1 and β-catenin in HT29 (C) and SW480 (D) detected by Western blotting
BTBD3 interacted with β-catenin and TYRO3, and ubiqutinated β-catenin
To explore the relationship between BTBD3 and TYRO3, β-catenin, we focused on the protein interacting with BTBD3. The FISH experiment showed that TYRO3, β-catenin and BTBD3 were mainly colocalized in the cytoplasm (Fig. 7A). The Co-IP results revealed that BTBD3 interacted with β-catenin, and BTBD3 also interacted with TYRO3 (Fig. 7B), which confirmed that BTBD3, β-catenin and TYRO3 interacted with each other in the same complex. BTBD3 is an E3 ubiquitin ligase and we applied in vivo ubiqutination assay to detect whether BTBD3 can ubiqutinate theβ-catenin. The in vivo ubiqutination assay showed that BTBD3 ubiqutinated the β-catenin and BTB3 knockdown decreased the ubiqutination (Fig. 7C). Ubiqutination led to protein degradation and then protein half-life assay showed siBTBD3 knockdown increased the β-catenin half-life (Fig. 7D, E). These data supported that β-catenin was ubiquitinated by BTBD3 and accelerated the protein degradation.

Representative FISH images of TYRO3, β-catenin and BTBD3 in HT29 cells and SW480 cells (A), Scale bar: 20 µM. Co-IP assay of endogenous BTBD3 and β-catenin, BTBD3 and TYRO3 in HT29 cells (B). in vivo ubiquitination assay for β-catenin with BTBD3 (C). Protein half-life assay for β-catenin with BTBD3 (D, E). Effects of overexpression of TYRO3, β-catenin and BTBD3 on cell proliferation of HT29 (F) and SW480 (G), colony formation (H), migration (I) and invasion (J) of HT29 and SW480. (I, J) Magnification, ×200
Effects of BTBD3, TYRO3 and β-catenin on CRC cells invasion and metastasis
To further explore the function of BTBD3 and the relationship between BTBD3 and TYRO3, β-catenin, we observed invasion and metastasis of the cells in oe-BTBD3, oe-BTBD3 + oe-TYRO3 and oe-BTBD3 + oe-β-catenin groups. The results showed that overexpression of BTBD3 decreased the proliferation (CCK-8 assay: Fig. 7F, G; Cloning assay: Fig. 7H), migration (Fig. 7I) and invasion (Fig. 7J) of HT29 and SW480 cells, while overexpression of TYRO3 and β-catenin restored the capacity. As a tumor suppressor gene in CRC, BTBD3 had a significant negative correlation with TYRO3 and β-catenin, all of these results revealed that BTBD3 plays an inhibitory role in the occurrence and development of CRC.
The underlying mechanism of wnt pathway affecting the development of CRC in vivo
To confirm the underlying mechanism of Wnt pathway further in vivo, we established a xenograft model in nude mice. Tumor growth curves are shown in Fig. 8A. Tumor growth curves indicated that the sh-circRAE1 and oe-BTBD3-treated group had a smaller volume and sustained tumor inhibition, indicating that circRAE1/miR-388-3p/TYRO3 axis could promote but BTBD3 suppressed the tumorigenicity of CRC.

Growth curves of xenograft tumors (A). The mRNA expression levels of circRAE1, BTBD3, TYRO3 and β-catenin (B) were detected by RT-qPCR for the indicated treated xenograft tumors. The protein expression levels of BTBD3, TYRO3, GSK3β, APC and β-catenin (C) were detected by western blotting for the indicated treated xenograft tumors. HE staining of mouse colorectal and tumors for metastasis assays(D), magnification, ×200
Compared with the sh-circRAE1 group, the mRNA and protein expression of β-catenin in the sh-circRAE1 + oe-TYRO3 group and the sh-circRAE1 + miR-338-3P inhibitors group was significantly up-regulated. The mRNA (Fig. 8B) and protein (Fig. 8C) expression of β-catenin was significantly up-regulated in oe-BTBD3 + oe-TYRO3 group and oe-BTBD3 + oe-β-catenin group compared with oe-BTBD3 group. As shown in Fig. 8D, the HE results found that normal tumor cells retained their heteromorphism and ill-defined border, nuclei were enlarged in NC group, interstitium showed fibrous tissue proliferation and lymphocytic infiltration, and significant tumor cell necrosis was observed. However, the areas of tumor tissue necrosis in oe-BTBD3 group were significantly increased compared with the NC group, the oe-BTBD3 + oe-TYRO3 group and the oe-BTBD3 + oe-β-catenin group displayed strikingly opposite pathologic features. These results suggested that BTBD3 negatively correlated with TYRO3 and β-catenin, and inhibited the occurrence and development of CRC.
Discussion
Previous studies have demonstrated that circRAE1 acts as an oncogene in CRC progression by regulating the expression of epithelial-mesenchymal transition (EMT) related markers [8] including E-cadherin and vimentin [26, 27]. EMT is a critical process in tumor invasion and metastasis [28, 29], targeted reversal of EMT could be an effective strategy for treating cancer. In this study, our data supported that circRAE1/miR-388-3p/TYRO3 axis was involved in the CRC EMT process andBTBD3 suppressed circRAE1/miR-388-3p/TYRO3 axis and the Wnt/β-catenin pathway and therefore blocked the CRC progression. Our data further confirmed that BTBD3 bould and ubiquitinated β-catenin and led to β-catenin degradation, therefore inhibited Wnt/β-catenin pathway and CRC tumorigenesis.
Chien et al. confirmed that Tyro3 overexpression led to EMT in CRC [10], which was consistent with our experimental results. Studies indicated that TYRO3 (a member of the TAM receptor tyrosine kinase family) was a potential therapeutic target for a variety of cancers. For example, Dufour F et al. found that TYRO3 silencing induced apoptosis, and TYRO3 was a molecular target for growth inhibition in bladder cancer [30]. Al Kafri N et al. showed that the ProS1-TYRO3 interaction was primarily coupled to Erk signaling to support and protect cancer cell survival [31]. Shao H et al. demonstrated that TYRO3 mediated phosphorylation of ACTN4 to limit the migration and invasion of melanoma cells [32]. Recent studies have shown that the TYRO3 receptor plays an important role in CRC, when the TYRO3 receptor was knocked-down in vitro, it inhibited the proliferation of CRC cells [33]. Interestingly, TYRO3 can mediate cell growth, invasion and metastasis by activating the Wnt/β-catenin signaling-mediated EMT in human gastric cancer cells [13]. The disorder of the Wnt signaling pathway is considered as the pathological basis of CRC [34]. Our findings also demonstrated this view, the oe-TYRO3 could directly up-regulate the expression of β-catenin in HT29 and SW620 cells, which indicated circRAE1/miR-388-3p/TYRO3 axis promoting the Wnt/β-catenin signaling pathway and development of CRC.
In this study, we also found that BTBD3 expression inversely correlates with β-catenin in the four CRC cell lines and one normal colon epithelial cell line. Clinically, BTBD3 was also found downregulated in 30 pairs of CRC tissues and the adjacent non-tumor tissues, which indicated that BTBD3 may be a cancer-associated gene. We showed here that BTBD3 overexpression inhibited the colony formation, migration and invasion capacity of CRC cells in vivo and reduced tumor volume in vitro. Co-IP results further indicated that BTBD3 interacted with TYRO3 and β-catenin in vivo or in vitro. Interestingly, our data showed that BTBD3 ubiquitinated β-catenin which led to an increase in the degradation of β-catenin with a shorter protein half-life. Based on the aforementioned findings, it may be concluded that BTBD3 could directly interact with TYRO3 and β-catenin, ubiquitinated β-catenin, therefore blocked the Wnt/β-catenin pathway and inhibited the CRC tumorigenesis.
Conclusions
In summary, this study investigated the effect of circRAE1/miR-388-3p/TYRO3 axis and BTBD3 on Wnt/β-catenin pathway and CRC tumorigenesis in vitro and in vivo. circRAE1 knockdown and TYRO3 overexpression promoted Wnt/β-catenin signaling pathway and the EMT process associated markers, indicating that circRAE1/miR-388-3p/TYRO3 axis exacerbated tumorigenesis of CRC by activating Wnt/β-catenin signaling pathway. In addition, overexpression of BTBD3 reduced cell migration and invasion in vitro and inhibited tumor growth in vivo. BTBD3 ubiqutinated β-catenin which led to β-catenin degradation and blocked Wnt/β-catenin pathway, therefore suppressed the tumorigenesis of CRC. This study may provide novel therapeutic targets for CRC therapy.
Data availability
The data that support the findings of this study are available on request from the first author (K.Y.) or corresponding author (P.C.).
Abbreviations
- BTBD3:
-
BTB Domain Containing 3
- CRC:
-
Colorectal cancer
- TAM:
-
TYRO3-AXL-MER family
- GSK3β:
-
Alycogen synthase kinase 3β
- APC:
-
Adenomatous polyposis coli
- AXIN1:
-
Scaffolding protein 1
- AUC:
-
Area under the curve
- FISH:
-
Fluorescence in situ hybridization
- EMT:
-
Epithelial-mesenchymal transition
- ACTN4:
-
Alpha-actinin 4
References
Phipps O, Brookes MJ, Al-Hassi HO. Iron deficiency, immunology, and colorectal cancer. Nutr Rev. 2021;79(1):88–97.
Du F, Qiao C, Li X, Chen Z, Liu H, Wu S, Hu S, Qiu Z, Qian M, Tian D, et al. Forkhead box K2 promotes human colorectal cancer metastasis by upregulating ZEB1 and EGFR. Theranostics. 2019;9(13):3879–902.
Sun Y, Ji B, Feng Y, Zhang Y, Ji D, Zhu C, Wang S, Zhang C, Zhang D, Sun Y. TRIM59 facilitates the proliferation of colorectal cancer and promotes metastasis via the PI3K/AKT pathway. Oncol Rep. 2017;38(1):43–52.
Zong L, Yang Y, Zhang J, Dai L, Luo Y, Yang J, Luo X. Rhododendron molle G. Don Extract induces apoptosis and inhibits Migration in Human Colorectal Cancer cells and potential Anticancer Components Analysis. Molecules 2021, 26(10).
Wang HY, Li F, Liu N, Liu XY, Yang XH, Guo YM, Bei JX, Zeng YX, Shao JY. Prognostic implications of a molecular classifier derived from whole-exome sequencing in nasopharyngeal carcinoma. Cancer Med. 2019;8(6):2705–16.
Zhang C, Zhou X, Geng X, Zhang Y, Wang J, Wang Y, Jing J, Zhou X, Pan W. Circular RNA hsa_circ_0006401 promotes proliferation and metastasis in colorectal carcinoma. Cell Death Dis. 2021;12(5):443.
Shang A, Gu C, Wang W, Wang X, Sun J, Zeng B, Chen C, Chang W, Ping Y, Ji P, et al. Exosomal circPACRGL promotes progression of colorectal cancer via the miR-142-3p/miR-506-3p- TGF-β1 axis. Mol Cancer. 2020;19(1):117.
Du J, Xu J, Chen J, Liu W, Wang P, Ye K. circRAE1 promotes colorectal cancer cell migration and invasion by modulating miR-338-3p/TYRO3 axis. Cancer Cell Int. 2020;20:430.
Pagani S, Bellan M, Mauro D, Castello LM, Avanzi GC, Lewis MJ, Sainaghi PP, Pitzalis C, Nerviani A. New Insights into the Role of Tyro3, Axl, and Mer Receptors in Rheumatoid Arthritis. Disease markers 2020, 2020:1614627.
Chien CW, Hou PC, Wu HC, Chang YL, Lin SC, Lin SC, Lin BW, Lee JC, Chang YJ, Sun HS, Tsai SJ. Targeting TYRO3 inhibits epithelial-mesenchymal transition and increases drug sensitivity in colon cancer. Oncogene. 2016;35(45):5872–81.
Ekyalongo RC, Mukohara T, Funakoshi Y, Tomioka H, Kataoka Y, Shimono Y, Chayahara N, Toyoda M, Kiyota N, Minami H. TYRO3 as a potential therapeutic target in breast cancer. Anticancer Res. 2014;34(7):3337–45.
Dantas-Barbosa C, Lesluyes T, Loarer FL, Chibon F, Treilleux I, Coindre JM, Meeus P, Brahmi M, Bally O, Ray-Coquard I, et al. Expression and role of TYRO3 and AXL as potential therapeutical targets in leiomyosarcoma. Br J Cancer. 2017;117(12):1787–97.
Chen D, Liu Q, Cao G, Zhang W. TYRO3 facilitates cell growth and metastasis via activation of the Wnt/β-catenin signaling pathway in human gastric cancer cells. Aging. 2020;12(3):2261–74.
Nie X, Liu H, Liu L, Wang YD, Chen WD. Emerging roles of wnt ligands in human colorectal Cancer. Front Oncol. 2020;10:1341.
Yao H, Ren D, Wang Y, Wu L, Wu Y, Wang W, Li Q, Liu L. KCTD9 inhibits the Wnt/β-catenin pathway by decreasing the level of β-catenin in colorectal cancer. Cell Death Dis. 2022;13(9):761.
Eisa NH, Said E, Khodir AE, Sabry D, Ebrahim HA, Elsherbini DMA, Altemani R, Alnasser DM, Elsherbiny NM, El-Sherbiny M. Effect of Diacerein on HOTAIR/IL-6/STAT3, Wnt/β-Catenin and TLR-4/NF-κB/TNF-α axes in colon carcinogenesis. Environ Toxicol Pharmacol. 2022;95:103943.
Gajos-Michniewicz A, Czyz M. WNT signaling in Melanoma. Int J Mol Sci 2020, 21(14).
Zhang H, Qi Y, Geng D, Shi Y, Wang X, Yu R, Zhou X. Expression profile and clinical significance of wnt signaling in human gliomas. Oncol Lett. 2018;15(1):610–7.
Pergolizzi M, Bizzozero L, Maione F, Maldi E, Isella C, Macagno M, Mariella E, Bardelli A, Medico E, Marchiò C, et al. The neuronal protein neuroligin 1 promotes colorectal cancer progression by modulating the APC/β-catenin pathway. J Experimental Clin cancer Research: CR. 2022;41(1):266.
Xu L, Wei Y, Reboul J, Vaglio P, Shin TH, Vidal M, Elledge SJ, Harper JW. BTB proteins are substrate-specific adaptors in an SCF-like modular ubiquitin ligase containing CUL-3. Nature. 2003;425(6955):316–21.
Siggs OM, Beutler B. The BTB-ZF transcription factors. Cell Cycle. 2012;11(18):3358–69.
Schonrock N, Humphreys DT, Preiss T, Götz J. Target gene repression mediated by miRNAs miR-181c and miR-9 both of which are down-regulated by amyloid-β. J Mol Neuroscience: MN. 2012;46(2):324–35.
Zhang P, Ma Y, Wang F, Yang J, Liu Z, Peng J, Qin H. Comprehensive gene and microRNA expression profiling reveals the crucial role of hsa-let-7i and its target genes in colorectal cancer metastasis. Mol Biol Rep. 2012;39(2):1471–8.
Xiao W, Zhao W, Li L, Wu Q, Zhu L, Zhang Q, Dai W, Wang Y, Zhang B. Preliminary investigation of the role of BTB domain-containing 3 gene in the proliferation and metastasis of hepatocellular carcinoma. Oncol Lett. 2017;14(2):2505–10.
Kim MS, Kim SS, Ahn CH, Yoo NJ, Lee SH. Frameshift mutations of wnt pathway genes AXIN2 and TCF7L2 in gastric carcinomas with high microsatellite instability. Hum Pathol. 2009;40(1):58–64.
Zhu X, Li Q, Hu G, Wang J, Hu Q, Liu Z, Wu G, Zhong Y. BMS–345541 inhibits airway inflammation and epithelial–mesenchymal transition in airway remodeling of asthmatic mice. Int J Mol Med. 2018;42(4):1998–2008.
Che J, Yue D, Zhang B, Zhang H, Huo Y, Gao L, Zhen H, Yang Y, Cao B. Claudin-3 inhibits lung squamous cell Carcinoma Cell epithelial-mesenchymal transition and Invasion via suppression of the Wnt/β-catenin signaling pathway. Int J Med Sci. 2018;15(4):339–51.
Sui CJ, Xu M, Li WQ, Yang JM, Yan HZ, Liu HM, Xia CY, Yu HY. Co-culture of hepatoma cells with hepatocytic precursor (stem-like) cells inhibits tumor cell growth and invasion by downregulating Akt/NF-κB expression. Oncol Lett. 2016;12(5):4054–60.
Niu X, Huang B, Qiao X, Liu J, Chen L, Zhong M. MicroRNA-1-3p suppresses malignant phenotypes of Ameloblastoma through Down-regulating lysosomal Associated membrane protein 2-Mediated autophagy. Front Med. 2021;8:670188.
Dufour F, Silina L, Neyret-Kahn H, Moreno-Vega A, Krucker C, Karboul N, Dorland-Galliot M, Maillé P, Chapeaublanc E, Allory Y, et al. TYRO3 as a molecular target for growth inhibition and apoptosis induction in bladder cancer. Br J Cancer. 2019;120(5):555–64.
Al Kafri N, Hafizi S. Tumour-secreted protein S (ProS1) activates a Tyro3-Erk Signalling Axis and protects Cancer cells from apoptosis. Cancers 2019, 11(12).
Shao H, Wang A, Lauffenburger D, Wells A. Tyro3-mediated phosphorylation of ACTN4 at tyrosines is FAK-dependent and decreases susceptibility to cleavage by m-Calpain. Int J Biochem Cell Biol. 2018;95:73–84.
Schmitz R, Valls AF, Yerbes R, von Richter S, Kahlert C, Loges S, Weitz J, Schneider M, de Ruiz C, Ulrich A, Schmidt T. TAM receptors Tyro3 and Mer as novel targets in colorectal cancer. Oncotarget. 2016;7(35):56355–70.
Tang Q, Chen J, Di Z, Yuan W, Zhou Z, Liu Z, Han S, Liu Y, Ying G, Shu X, Di M. TM4SF1 promotes EMT and cancer stemness via the Wnt/β-catenin/SOX2 pathway in colorectal cancer. J Experimental Clin cancer Research: CR. 2020;39(1):232.
Acknowledgements
We sincerely thank all the medical staff of the Department of Gastrointestinal Surgery of the Second Affiliated Hospital of Fujian Medical University for their support for this work.
Funding
This work was financially supported by Science and Technology Project of Fujian Provincial Health Commission (2021CXA028). Key Clinical Specialty Discipline Construction Program of Fujian, P.R.C (Fujian Health Medicine and Politics [2022]884).
Ethics declarations
Ethical approval
The study was approved by the Ethical Review Committee of the Second Affiliated Hospital of Fujian Medical University. Ethical Review No. [2021] Fuyi Fueryi Ethical No. (69).
Consent for publication
This work is published under the standard license to publish agreement. After 12 months the work will become freely available and the license terms will switch to a Creative Commons Attribution 4.0 International (CCBY 4.0).
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Ye, K., Wang, PC., Chen, YX. et al. E3 ubiquitin ligase BTBD3 inhibits tumorigenesis of colorectal cancer by regulating the TYRO3/Wnt/β-catenin signaling axis. Cancer Cell Int 24, 306 (2024). https://doi.org/10.1186/s12935-024-03478-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12935-024-03478-z