
Abstract
Background
Despite the success of antiretroviral therapy (ART), people living with HIV (PLWH) suffer from a high burden of pulmonary diseases, even after accounting for their smoking status. Cytotoxic CD8 T-cells are likely implicated in this phenomenon and may act as a double-edged sword. While being essential in viral infection control, their hyperactivation can also contribute to lung mucosal tissue damage. The effects of HIV and smoking on pulmonary mucosal CD8 T-cell dynamics has been a neglected area of research, which we address herein.
Methods
Bronchoalveolar lavage (BAL) fluid were obtained from ART-treated PLWH (median duration of supressed viral load: 9 years; smokers: n = 14; non-smokers: n = 21) and HIV-uninfected controls (smokers: n = 11; non-smokers: n = 20) without any respiratory symptoms or active infection. Lymphocytes were isolated and CD8 T-cell subsets and homing markers were characterized by multiparametric flow cytometry.
Results
Both smoking and HIV infection were independently associated with a significant increase in frequencies of total pulmonary mucosal CD8 T-cell. BAL CD8 T-cells were primarily CD69 + expressing CD103 and/or CD49a, at least one of the two granzymes (GzmA/GzmB), and little Perforin. Higher expression levels of CD103, CD69, and GzmB were observed in smokers versus non-smokers. The ex vivo phenotype of GzmA + and GzmB + cells revealed increased expression of CD103 and CXCR6 in smokers, while PLWH displayed elevated levels of CX3CR1 compared to controls.
Conclusion
Smoking and HIV could promote cytotoxic CD8 T-cell retention in small airways through different mechanisms. Smoking likely increases recruitment and retention of GzmB + CD8 Trm via CXCR6 and CD103. Heightened CX3CR1 expression could be associated with CD8 non-Trm recruitment from the periphery in PLWH.
Similar content being viewed by others
Introduction
Despite the success of antiretroviral therapy (ART), people living with HIV (PLWH) suffer from a disproportionate burden of infectious and non-infectious pulmonary diseases, suggesting that their lung immunity is not fully restored [1,2,3]. Moreover, smoking prevalence in PLWH is almost twice as high compared to the general population, contributing to low grade chronic systemic inflammation and decreased ART efficacy in these individuals [1, 2, 4,5,6]. Helleberg and others report that both all-cause and non-AIDS-related mortalities are higher among PLWH who smoke than among those who do not and that this population loses more life-years to smoking than to HIV infection itself (12.3 years versus 5.1 years respectively) [7]. Importantly, both smoking and HIV are independent risk factors for pulmonary complications such as lung cancer, emphysema, and Chronic Obstructive Pulmonary Disease (COPD) [1, 2, 8,9,10].
Within this context, lung mucosal CD8 T-cells play a critical role in controlling chronic viral infection, while potentially harming surrounding tissues and contributing to pulmonary pathologies frequently seen in PLWH [1, 2]. HIV is rapidly seeded within the lung during primary infection, where proximity of millions of alveoli provide a large surface area for cell-to-cell viral spread [1, 11, 12]. Our team have also previously shown that HIV-DNA persists within pulmonary CD4 and double negative T-cells despite a decade of ART [13, 14]. Importantly, CD8 T-cells are critical for HIV control. Animal studies have shown that CD8 T-cell depletion in SIV-infected Rhesus Macaques treated with short-term ART, leads to increased plasma viremia, which is reversible with CD8 T-cell repopulation [15]. CD8 T-cells may accumulate in the lung upon HIV seeding within that tissue. In fact, asymptomatic PLWH frequently develop “CD8 T-cell alveolitis” characterized by accumulation of HIV-specific pulmonary T-cells, which is associated with respiratory symptoms and worse clinical outcomes [1, 16, 17]. While CD8 T-cells are crucial for the clearance of HIV-infected cells and opportunistic infections, chronic inflammation and antigen stimulation result in impairment of their antiviral functions in PLWH [1, 2, 17]. Although some of these functions do recover in the peripheral blood after ART initiation, this is not the case for pulmonary CD8 T-cells [18, 19]. In the context of other chronic pulmonary viral infections and lung cancers, CD8 T-cells also lose their cytotoxic activity during late-stage differentiation due to immune exhaustion [20,21,22,23], increasing the risk of opportunistic infections and malignancies [24, 25]. Moreover, these cells might induce excessive expansion of other CD8 T-cells in the vicinity via T-cell receptor independent mechanisms, known as “bystander activation” [26, 27]. Collectively, chronic immune activation during HIV infection is one of the likely mechanisms driving accumulation of functionally impaired CD8 T-cells displaying reduced proliferation, poor effector functions, and high expression of inhibitory receptors in the lung as reported by our team and others [19, 26, 28,29,30].
Cigarette smoke can have both pro- and anti-inflammatory effects by inducing the production of TNF-α, IL-1β, IL-6, IL-8, and monocyte chemoattractant protein-1 [31, 32]. However, other studies indicate that nicotine, the most addictive component of tobacco, does the opposite, where it can decrease levels of IL-6, IL-8, and IL-10 through the engagement of α7 nicotinic acetylcholine receptor [33,34,35]. Increased production of pro-inflammatory cytokines in smokers has a significant effect on their BAL fluid immune cell concentration and composition: BAL fluid from smokers has increased number of neutrophils, macrophages and CD8 T-cells compared to non-smokers [36, 37]. Subsequently, BAL CD4/CD8 T-cell ratio in smokers is reduced. Studies also show that these CD8 and CD4 T-cells are skewed towards a type 1 phenotype (Tc1 and Th1 cells respectively), which are IFN-γ producing cells that help fight intracellular infections but are also capable of causing tissue destruction in COPD patients [38,39,40]. We recently demonstrated that pulmonary CD8 T-cells show lower ex vivo perforin expression, lower ability to upregulate degranulation upon in vitro stimulation, and limited HIV-specific killing capacity compared with blood CD8 T-cells regardless of HIV or smoking status [30]. On the one hand, this could potentially contribute to a suboptimal anti-HIV immune response and the establishment of viral reservoirs within the lungs. On the other hand, this low cytotoxic profile might be part of a normal physiological cell state that acts as a safeguard against undue tissue damage to a vital organ. Compared to the blood, pulmonary mucosa is exposed to high levels of oxygen, airborne particles, and microbes. Furthermore, nutrients are scarce in that tissue, which in turn, can affect CD8 T-cell metabolism and function [41, 42]. Transcriptomic analysis studies in murine models also report that, in contrast to cells from the pulmonary interstitium and the circulation, CD8 T-cells from the airways display lower expression of genes involved in cytotoxic function [41]. This property of tissue-resident memory CD8 T-cells (CD8 Trm) during homeostatic conditions might be reversed in context of chronic inflammation, resulting in tissue destruction. In chronic inflammatory lung conditions, like smoking and COPD, increased CD8 T-cell cytotoxicity and reduced apoptosis in the small airways can compromise the integrity of the mucosal barrier by killing alveolar epithelial cells via perforin, granzyme-B, and TNF-α [43,44,45,46,47,48,49,50]. Notably, we have previously reported significant reductions in proportions of senescent pulmonary CD28-CD57 + CD8 T-cells in smoking PLWH, which was not observed in seronegative controls, suggesting differential dynamics of CD8 T-cells in these study participants [30].
Overall, CD8 T-cells’ cytotoxic functions are needed for pathogen clearance and infection control but can wreak havoc if left unchecked. In the context of pulmonary mucosa, we do not yet know at what differentiation or tissue migration stage CD8 T-cells begin to lose their cytotoxic functions. In this study, we aimed to investigate pulmonary CD8 T-cell dynamics in PLWH and uninfected individuals, while accounting for the impact of tobacco smoking. Across four different study groups, we performed a comprehensive flow-cytometric characterization of CD8 T-cell phenotypes based on their expression of effector proteins, tissue-residency markers, and chemokine receptors involved in T-cell migration.
Methods
Study population
ART treated (suppressed VL ≥ 1 year and CD4 count ≥ 350 cells/mm3) smokers and non-smokers in addition to HIV-uninfected smokers and non-smokers were recruited at the McGill University Health Centre (MUHC). All participants underwent bronchoscopies for research purposes only. Participants underwent spirometric testing several weeks prior to bronchoscopy to ensure the absence of any undiagnosed obstructive airflow disease. The participants were also screened for the absence of active pulmonary infection and any other acute pulmonic process. Participants were labelled as smokers if they smoked at least 1 tobacco cigarette daily. 50–100 ml of BAL fluid were obtained via bronchoscopies performed by a respirologist and matched 40 ml blood samples were collected by venipuncture the same day.
Lymphocyte isolation
As per a recent protocol optimized and published by our team, BAL fluid was centrifuged within the first hour following the bronchoscopy to isolate the cells and remove any BAL liquid. Lymphocytes were isolated from both blood and BAL as previously described [13, 30, 51]. All samples underwent cryopreservation prior to subsequent flow cytometric analysis in batch.
Flow cytometry
Live pulmonary CD8 T-cells were characterized ex vivo using CD49a/CD69/CD103 (Trm markers), CD45RA/CCR7 (T-cell memory subsets), CXCR3/CXCR6 (CD8 T-cell migration from the interstitium), CX3CR1/KLRG1 (CD8 T-cell migration from peripheral circulation) [52,53,54,55,56,57], and granzyme-A/granzyme-B/perforin (cytotoxicity). The cells were stratified based on expression of granzyme-A/granzyme-B/perforin and CD103 (E-cadherin receptor)/CD69 (S1P1 signaling inhibitor)/CD49a (collagen IV receptor). The antibodies used are listed in Supplementary Table 1. Samples were acquired on a 5-laser BD Fortessa-X20 and the results were analyzed by FlowJo software V10.9 (BD Bioscience). Cell frequencies obtained from gates whose parent population was less than 100 cells were excluded from analyses. Due to the constrained quantity of BAL cells and interindividual discrepancies in their CD8 T-cell proportions, we were unable to evaluate all markers for every participant. The exact sample size for each data set is specified accordingly in the Results section. T-distributed stochastic neighbor embedding plots were generated using BAL CD3 + lymphocytes pooled from four study groups (n = 5 participant per group; 3905 CD3 + live BAL lymphocytes per group; 15,620 cells in total). Distinct CD8 T-cell populations were identified using FlowSOM, a visualization tool that clusters cells based on chosen flow cytometry markers. FlowSOM arranges cell clusters via a Self-Organizing Map in a Minimum Spanning Tree and groups similar clusters into metaclusters – what we refer to as cell populations [58].
Statistical analyses
Data were analyzed using GraphPad Prism V10 (San Diego, CA). For all categorical variables, Fisher’s exact test was used. For all quantitative variables, the Kolmogorov-Smirnov test was first used to determine the distribution of variables. The Kruskal-Wallis test was then used to determine significant differences between more than two study groups. Wilcoxon matched-pairs signed rank test and Mann-Whitney rank-sum test were used to compare paired and unpaired study variables respectively. The figures depict only statistically significant p-values (* p < 0.05, ** p < 0.01, *** p < 0.001, **** p < 0.0001).
Results
Participant characteristics
Most of the study participants were male Caucasians (Table 1). PLWH were slightly older compared to uninfected control donors (median age 56 vs. 40 years old). All participating PLWH were under ART with suppressed viral loads and stable CD4 T-cell counts (median: 531 cells/mm3). The final sample size per experiment varied based on the BAL cell viability and cell count after thawing. Details for each experiment performed per sample can be found in Supplementary Table 2.
HIV and smoking are independently associated with increased BAL CD8 T-cell frequencies
To evaluate the differences in CD8 T-cell proportions between study groups, BAL CD3 + live singlets expressing the CD8 co-receptor, but not CD4, were gated and their frequencies evaluated within the total live CD3 + lymphocyte pool (Supplementary Fig. 1a). Both smoking and positive HIV status were independently associated with significantly higher BAL CD8 T-cell frequencies (Fig. 1a).

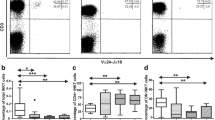
Impact of chronic HIV infection and smoking on BAL CD8 T-cell frequencies and the expression of Trm markers. (a) Frequencies of CD8 + CD4- cells in total BAL CD3 + live lymphocytes are shown. (b-d) Expression levels of CD103, CD69, and CD49a were assessed within the total CD8 T-cells pool in BAL (HIV-NS: n = 8; HIV-SM: n = 6; HIV + NS: n = 6; HIV + SM: n = 6). Data points were stratified by smoking status (right), HIV status (middle), or both (left). Comparisons were made using Mann–Whitney rank-sum test (* p < 0.05, ** p < 0.01, *** p < 0.001, **** p < 0.0001)
The majority of BAL CD8 T-cells are tissue-resident
To have a broader view of BAL CD8 T-cell dynamics and effector function, we quantified expression levels of tissue-residency markers (CD103, CD69, CD49a) and effector proteins (GzmA, GzmB, Perforin) in this cell population (Supplementary Fig. 1b). Across all study groups, nearly all BAL CD8 T-cells were positive for at least one Trm marker (Supplementary Fig. 1c, Figs. 1b-d and 2a). Among these populations, the most prominent were CD69 + subsets. Median cell frequencies within total CD8 T-cells are as follows: CD103 + CD69 + CD49a+ (median: 54%), CD103-CD69 + CD49a+ (median: 15%), CD103 + CD69 + CD49a- (median: 6%), CD103-CD69 + CD49a- (median: 11%) and the CD8 non-Trm subset CD103-CD69-CD49a- (median: 5.50%). Frequencies of the remaining CD69- CD8 T-cell subsets were very low (median ≤ 1% per subset). The majority of BAL CD8 T-cells were also positive for GzmA and/or GzmB, with low frequencies of Perforin + cells (Fig. 2b).

Distribution of Trm subsets and cytotoxic effector subsets among total BAL CD8 T-cells. (a) Trm subsets stratified by CD103/CD69/CD49a expression in BAL based on median cell population frequencies in each study group. (b) Cytotoxic effector CD8 T-cell subsets stratified by GzmA/GzmB/Perforin expression in BAL and blood based on median cell population frequencies in each study group. (HIV-NS: n = 8; HIV-SM: n = 6; HIV + NS: n = 6; HIV + SM: n = 6)
It’s worth mentioning that many of the experiments were conducted during the COVID-19 pandemic, a time when bronchoscopies were not allowed, which restricted us to only using frozen samples. During that time, we examined the effect of cryopreservation using matched fresh and frozen peripheral blood mononuclear cells (HIV-NS: n = 2) on CD8 T-cell subset markers measured in our flow cytometric panels (Trm: CD103/CD69/CD49a; cytotoxicity: GzmA/GzmB/Perforin, memory: CD45RA/CCR7). We observed no considerable differences between fresh and frozen samples for the exception of one marker – CD69 – that showed a mean increase of 18.83% on CD8 T-cells after cryopreservation (Supplementary Table 3). Although we observed a decrease in live CD3 + BAL lymphocytes, no substantial change in BAL CD8 T-cell viability was observed in frozen versus fresh samples (Supplementary Table 4). To verify whether T-cell viability could confound our results, we performed Spearman’s rank correlation analysis and observed no significant relationship between levels of CD69 and T-cell viability (Supplementary Fig. 3).
Smokers display higher frequencies of CD103+, CD69 + and GzmB + CD8 T-cells in BAL along with increased CD8 T-cell recruitment within pulmonary mucosa
Only smoking, but not HIV status, was associated with higher frequencies of CD103+, CD69+ (Fig. 1b, c) and GzmB + CD8 T-cells in BAL, while GzmA and perforin expression remained unchanged based on smoking and HIV status (Fig. 3a-c). Since only GzmB + CD8 T-cells are increased in smokers and because GzmB + CD8 T-cells have been associated with COPD severity [59], we further gated on GzmB + cells in our analyses to measure their expression of markers of tissue-residency (CD103, CD69, CD49a), transepithelial migration (CXCR6, CXCR3) [55, 56], proliferative capacity (Ki67), and peripheral infiltration (CX3CR1, KLRG1) [57]. Among smoking participants, GzmB + BAL CD8 T-cells displayed significantly higher levels of CD103 and CXCR6 along with significantly reduced frequencies of CD49a + cells compared to non-smokers (Fig. 4a-c), suggesting increased transepithelial migration and retention of interstitial CD8Trm in the airway lumen. Similarly, significantly higher levels of CXCR6 were observed in GzmA + cells in smokers (Fig. 4e). Notably, in PLWH, we observed significantly higher levels of CX3CR1 on both GzmB + and GzmA + BAL CD8 T-cells (Fig. 4d, f) with a few participants exhibiting particularly elevated levels. Because CX3CR1 is exclusively expressed by non-tissue resident cells coming from the periphery [60], these data suggest increased recruitment of circulating effector CD8 T-cells into the airways in some PLWH. Lastly, we found no significant differences in levels of CXCR3, CX3CR1, KLRG1, and Ki-67 expression among study groups (Supplementary Table 5).

Expression of CD8 T-cell cytotoxic effector molecules GzmA/B and perforin among study groups. Expression levels of GzmA (a), GzmB (b), and Perforin (c) were assessed within the total CD8 T-cell pool in BAL (HIV-NS: n = 8; HIV-SM: n = 6; HIV + NS: n = 6; HIV + SM: n = 6). Data points were stratified by smoking status (right), HIV status (middle), or both (left). Comparisons were made using Mann–Whitney rank-sum test (* p < 0.05, ** p < 0.01, *** p < 0.001, **** p < 0.0001)

Increased migration and retention of GzmB + CD8 Trm from the interstitium to alveolar space in smokers. (a-d) Expression levels of CD103, CD49a, CXCR6, and CX3CR1 in GzmB + BAL CD8 T-cells (HIV-NS: n = 8; HIV-SM: n = 6; HIV + NS: n = 6; HIV + SM: n = 5), (e-f) as well as CXCR6 and CX3CR1 in GzmA + BAL CD8 T-cells (HIV-NS: n = 8; HIV-SM: n = 5; HIV + NS: n = 6; HIV + SM: n = 5) are shown. Data points were stratified by smoking status (right), HIV status (middle), or both (left). Comparisons were made using Mann–Whitney rank-sum test (* p < 0.05, ** p < 0.01, *** p < 0.001, **** p < 0.0001)
FlowSOM analysis reveals five distinct BAL CD8 T-cell populations that vary by smoking status
To better identify and describe BAL CD8 T-cell subsets distributions, we generated a t-distributed stochastic neighbor embedding (tSNE) plot on BAL CD3 + lymphocytes pooled from four study groups and identified 5 distinct populations using FlowSOM clustering and visualization technique (Fig. 5). In line with our afore mentioned observations, an apparent divide in expression of markers associated with tissue-resident (CD103, CD69, CD49a in Populations 1, 2, 4 and 5) and non-tissue resident (CX3CR1, KLRG1 in Population 3) cells was observed. Among the populations expressing tissue-residency markers, distinct differences in levels of CD103 and GzmA/B are also present. Lastly, we observed significantly higher frequencies of cells in Population 4 in smokers versus non-smokers (NS median = 10.95; SM median = 34.75; p-value = 0.009), which expressed intermediate levels of GzmB and high levels of CD103, both of which were found to be significantly increased in smoking participants in our previous analyses. Interestingly, this population displayed low levels of GzmA compared to other populations. To confirm this, we performed additional analyses of CD8 T-cell effector subsets based on their combined expression of GzmA/GzmB/Perforin and compared their frequencies between different study groups (Supplementary Fig. 2). Among the GzmB + subsets, only GzmA-GzmB + Perforin- cell frequencies were significantly higher in smokers versus non-smokers – a trend that was consistent regardless of HIV status.

FlowSOM analysis reveals five distinct BAL CD8 T-cell populations that vary by smoking status. (a) t-distributed stochastic neighbor embedding (tSNE) plot of sub-population in BAL CD3 + lymphocytes pooled from four study groups (n = 5 per group; 3905 CD3 + live BAL lymphocytes per group; 15,620 cells in total). Five different CD8 T-cell populations identified using FlowSOM clustering and visualization technique are shown. (b) tSNE plots of BAL CD8 T-cell populations stratified by either HIV, smoking status, or both with (c) their corresponding cell frequencies shown as bar plots. (d) The heat map displays relative expression of markers used for the FlowSOM analysis in each population identified
Reduction in naïve and terminally differentiated BAL CD8 T-cells subsets in smokers
We have previously reported significantly higher levels of CD45RA-CD28 + BAL CD8 T-cells in smokers versus non-smokers [30]. Given that we observed a significant increase in GzmB + BAL CD8 T-cells in smokers, we aimed to elucidate whether this increase is linked to cell differentiation stage, since more differentiated cells are known to have more pronounced effector functions [61]. We found that effector memory (EM; CCR7-CD45RA-) and terminally differentiated (TD; CCR7-CD45RA+) cells harbored the highest frequencies of GzmA and GzmB positive cells, with highest levels of perforin found in the CCR7-CD45RA + cell subset (Fig. 6a). Among cross-group comparisons, we have observed a reduction in naïve (N; CCR7 + CD45RA+) and TD BAL CD8 T-cell subsets in smoking versus non-smoking participants (Fig. 6b) with a similar skew in proportions toward central memory (CM; CCR7 + CD45RA-). Furthermore, we found that both CM and TD cells have significantly higher levels of GzmB in smokers versus non-smokers (Fig. 6c).

Significantly higher levels of GzmB in effector memory (EM) and terminally differentiated (TD) BAL CD8 T-cells cells in smokers. (a) Expression levels of GzmA, GzmB, and Peforin in BAL CD8 T-cell memory subsets (Naïve/ CCR7 + CD45RA+: HIV-NS n = 6, HIV-SM n = 2, HIV + NS n = 5, HIV + SM n = 1; central memory (CM) / CCR7 + CD45RA-: HIV-NS n = 6, HIV-SM n = 4, HIV + NS n = 4, HIV + SM n = 3; effector memory (EM) CCR7-CD45RA-: HIV-NS n = 8, HIV-SM n = 5, HIV + NS n = 6, HIV + SM n = 4), terminally differentiated (TD) CCR7-CD45RA+: HIV-NS n = 8, HIV-SM n = 3, HIV + NS n = 6, HIV + SM n = 2; (b) Distributions of BAL CD8 T-cell memory subsets between different study groups (HIV-NS: n = 8; HIV-SM: n = 6; HIV + NS: n = 6; HIV + SM: n = 6); (c) Frequencies of GzmB + cells in CM (left) and TD (right) BAL CD8 T-cell subsets stratified by participant smoking status. Wilcoxon matched-pairs signed rank test and Mann-Whitney rank-sum test were used to compare paired and unpaired study variables respectively (* p < 0.05, ** p < 0.01, *** p < 0.001, **** p < 0.0001)
Discussion
Herein, we provide a comprehensive in-depth flow cytometric description of airway CD8 T-cells encompassing their tissue-residency characteristics, effector proteins, markers of migration, and differentiation. We report that HIV and smoking status have distinct and independent effects on lung mucosal CD8 T-cell dynamics. We have found that, while both HIV and smoking are independent factors that can promote higher CD8 T-cell frequencies in BAL, they do so through different mechanisms. Our results suggest that smoking promotes increased migration of GzmA + and GzmB + CD8 T-cells via CXCR6 and augmented retention in the pulmonary mucosa through CD103. Conversely, HIV promotes their infiltration from the periphery via the fractalkine receptor CX3CR1. A significant increase in GzmB + cells in smokers versus non-smokers was evident in CM (CCR7 + CD45RA-) and TD (CCR7-CD45RA+) cell subsets. Importantly, among all GzmB + CD8 T-cells, smoking was associated with enrichment of the GzmA-GzmB + Perforin- CD8 T-cell population only.
While CXCR6 is expressed under normal homeostatic conditions in lung CD8 T-cells, it is upregulated substantially during inflammatory responses in context of acute lung infections, chronic lung disease, and aging [62,63,64,65]. In context of smoking, increased migration and retention of CD8 T-cells armed with effector proteins like GzmB could be one of the contributors towards increased risk of COPD development in smokers and its subsequent tissue-destructive clinical presentation. Previously, we have reported that, unlike in the blood, BAL CD8 T-cells display high levels of degranulation marker CD107a [30]. Combined with our current observation of enriched GzmB + cells in smokers, collectively this could lead to accumulation GzmB in the extracellular space. Notably, GzmB is a serine protease capable of cleaving extracellular matrix proteins, which could contribute to its degradation and, subsequently, emphysema observed in COPD patients [59]. Our finding that smoking leads to increased levels of GzmB + CD8 T-cells in the lung is in line with previous studies documenting increased CD8 T-cell cytotoxicity in the blood and airways of smokers [66,67,68]. Furthermore, our observation of increased retention of these GzmB + cells via CD103 is consistent with a recent study by Corleis et al., which reported an increase in CD8 T-cell frequencies in endobronchial brushings of HIV-1 infected smokers [69]. We further show that this is likely caused by increased recruitment of lung interstitial CD8 Trm to the alveolar space via CXCR6, which has also been implicated in COPD and lymphocytic alveolitis [70, 71]. Notably, Freeman et al., have reported that increased CXCR6 expression by CD8 T-cells is positively correlated with COPD severity, meaning that CXCL16/CXCR6 axis blockade might constitute a new therapeutic approach for mitigating some of the damage seen in smoking COPD patients [64].
Significantly higher GzmB levels in CM CD8 T-cells in smokers along with a skewed trend in CD8 memory phenotype from TD toward CM, and the lack of a significant increase in Ki-67 could be indicative of increased cellular trafficking (rather than homeostatic proliferation) of younger memory cells into the alveolar space, and simultaneous clearing of older TD cells. While, by definition, CD8 Trm cells do not express CCR7 and we did not observe increased levels of CX3CR1 in smokers, it is does not rule out a potential increase in CD8 T-cell infiltration not only from the interstitial CD8 Trm cell pool but also the CCR7 + CM cell pool from the peripheral circulation. While we used CX3CR1 as a marker for circulating CD8 T-cells, since it is not expressed by tissue-resident cells, some studies have pointed out a distinction between CX3CR1 + and CX3CR1- circulating CD8 T-cell memory subsets [60], leaving open the possibility that lack of CX3CR1 on BAL CD8 T-cells in smokers might indicate that the infiltrating CD8 T-cells are at an earlier differentiation stage.
Considerably elevated levels of CX3CR1 observed in some of our study participants who are PLWH might be linked to the residual lung HIV reservoir that persists despite long-term suppressive ART, which our group and others have reported on previously [13, 72, 73]. Importantly, residual HIV proteins found in BAL fluid of PLWH on ART are capable of mediating T-cell chemotaxis [74, 75]. Existing literature also postulates that CX3CR1 + CD8 T-cells can act as a double-edged sword, contributing to viral control [76] but also increasing the risk of inflammatory diseases in PLWH [77]. Enrichment in CX3CR1 + circulating memory CD8 T-cells in PLWH on ART has been previously reported by Mudd et al. [77]. In their work, the authors argued that the expanded CD8 T-cells in PLWH on ART could preferentially localize to vascular endothelium and contribute to cardiovascular disease onset. Importantly, the CX3CL1-CX3CR1 axis has been implicated in various chronic inflammatory lung diseases, such as COPD and pulmonary fibrosis [78], whose rates are also increased in PLWH on ART [1, 78]. One of the pathogenic mechanisms observed in these chronic lung conditions is CD8 T-cell accumulation within the lung vessel wall and parenchyma [17, 59, 78]. In summary, increased frequencies of BAL CD8 T-cells, mediated through chemokine receptors like CX3CR1, could be one of the culprits behind increased lung disease rates in PLWH on ART.
One of this study’s challenges and subsequent limitations was gathering of sufficient biological material. Because a large portion of the experiments was performed during the COVID-19 pandemic, all bronchoscopies were prohibited, limiting us to frozen samples only. This has significantly impacted cell viability and subsequent cell count for ongoing experiments, which is reflected in the varying sample sizes between data sets. To verify whether the drop in cell viability and recovery had an effect on other markers being measured, we have performed additional tests and analyses comparing fresh and frozen BAL and PBMC samples using our flow cytometric panels after the ban on bronchoscopies was lifted. We observed no prominent changes except for one marker – CD69 – whose frequencies were higher in frozen versus fresh CD8 T-cells coming from the same donors. CD69 is known to be increased upon T-cell activation and cell stress, both of which are likely during the heat shock the cells are put under upon thawing [79, 80]. A similar increase in CD69 after cryopreservation has also been previously reported in circulating human NK cells [81]. Collectively, because all BAL samples in this study were cryopreserved and then processed in the same manner, we do not anticipate that this would affect our cross-group comparisons.
Conclusions
Overall, results of this study provide an additional puzzle piece to the bigger picture of the pulmonary immune system in PLWH on ART and in smokers. We have provided an extensive description of GzmB + CD8 T-cell populations that are enriched in context of smoking and how they compare between PLWH and seronegative controls. We report that the profound effect on CD8 T-cell frequencies in BAL observed in context of smoking and HIV is likely mediated through different migration/homing mechanisms, and significant differences in CD8 T-cell phenotypes between the two groups. Collectively, these findings can help inform further studies exploring new targets to design therapeutics that could help alleviate chronic emphysema and bronchiolitis in PLWH on ART. Importantly, such therapeutic approaches might need to be tailored to smoking status.
Data availability
No datasets were generated or analysed during the current study.
References
Alexandrova Y, Costiniuk CT, Jenabian MA. Pulmonary Immune Dysregulation and viral persistence during HIV infection. Front Immunol. 2021;12:808722.
Costiniuk CT, Jenabian MA. The lungs as anatomical reservoirs of HIV infection. Rev Med Virol. 2014;24(1):35–54.
Cillóniz C, García-Vidal C, Moreno A, Miro JM, Torres A. Community-acquired bacterial pneumonia in adult HIV-infected patients. Expert Rev Anti-infective Therapy. 2018;16(7):579–88.
Mdodo R, Frazier EL, Dube SR, Mattson CL, Sutton MY, Brooks JT, et al. Cigarette smoking prevalence among adults with HIV compared with the general adult population in the United States: cross-sectional surveys. Ann Intern Med. 2015;162(5):335–44.
Feldman JG, Minkoff H, Schneider MF, Gange SJ, Cohen M, Watts DH, et al. Association of cigarette smoking with HIV prognosis among women in the HAART era: a report from the women’s interagency HIV study. Am J Public Health. 2006;96(6):1060–5.
Van Eeden S, Leipsic J, Paul Man SF, Sin DD. The relationship between lung inflammation and Cardiovascular Disease. Am J Respir Crit Care Med. 2012;186(1):11–6.
Helleberg M, Afzal S, Kronborg G, Larsen CS, Pedersen G, Pedersen C, et al. Mortality attributable to Smoking among HIV-1–Infected individuals: a Nationwide, Population-based Cohort Study. Clin Infect Dis. 2013;56(5):727–34.
Sigel K, Wisnivesky J, Gordon K, Dubrow R, Justice A, Brown ST, et al. HIV as an independent risk factor for incident lung cancer. AIDS. 2012;26(8):1017–25.
Koshiol J, Rotunno M, Consonni D, Pesatori AC, De Matteis S, Goldstein AM, et al. Chronic obstructive pulmonary disease and altered risk of lung cancer in a population-based case-control study. PLoS ONE. 2009;4(10):e7380.
Agustí A, Edwards LD, Rennard SI, MacNee W, Tal-Singer R, Miller BE, et al. Persistent systemic inflammation is associated with poor clinical outcomes in COPD: a novel phenotype. PLoS ONE. 2012;7(5):e37483.
Clarke JR, Mitchell DM, Coker RJ, Israel-Biet D. Is the lung important as a privileged site for the human immunodeficiency virus? Thorax. 1998;53(2):81–2.
Costiniuk CT, Jenabian MA. Cell-to-cell transfer of HIV infection: implications for HIV viral persistence. J Gen Virol. 2014.
Costiniuk CT, Salahuddin S, Farnos O, Olivenstein R, Pagliuzza A, Orlova M, et al. HIV persistence in mucosal CD4 + T cells within the lungs of adults receiving long-term suppressive antiretroviral therapy. AIDS. 2018;32(16):2279–89.
Meziane O, Salahuddin S, Pham TNQ, Farnos O, Pagliuzza A, Olivenstein R et al. HIV infection and persistence in Pulmonary Mucosal double negative T cells in vivo. J Virol. 2020;94(24).
Cartwright EK, Spicer L, Smith SA, Lee D, Fast R, Paganini S, et al. CD8(+) lymphocytes are required for maintaining viral suppression in SIV-Infected macaques treated with short-term antiretroviral therapy. Immunity. 2016;45(3):656–68.
Guillon J-M, Autran B, Denis M, Fouret P, Plata F, Mayaud CM, et al. Human immunodeficiency virus-related Lymphocytic Alveolitis. Chest. 1988;94(6):1264–70.
Neff CP, Chain JL, MaWhinney S, Martin AK, Linderman DJ, Flores SC, et al. Lymphocytic alveolitis is associated with the accumulation of functionally impaired HIV-specific T cells in the lung of antiretroviral therapy-naive subjects. Am J Respir Crit Care Med. 2015;191(4):464–73.
Warren JA, Clutton G, Goonetilleke N. Harnessing CD8(+) T cells under HIV antiretroviral therapy. Front Immunol. 2019;10:291.
Brenchley JM, Knox KS, Asher AI, Price DA, Kohli LM, Gostick E, et al. High frequencies of polyfunctional HIV-specific T cells are associated with preservation of mucosal CD4 T cells in bronchoalveolar lavage. Mucosal Immunol. 2008;1(1):49–58.
Trautmann L, Janbazian L, Chomont N, Said EA, Gimmig S, Bessette B, et al. Upregulation of PD-1 expression on HIV-specific CD8 + T cells leads to reversible immune dysfunction. Nat Med. 2006;12(10):1198–202.
Braun MW, Iwakuma T. Regulation of cytotoxic T-cell responses by p53 in cancer. Transl Cancer Res. 2016;5(6):692–7.
Durgeau A, Virk Y, Corgnac S, Mami-Chouaib F. Recent advances in targeting CD8 T-Cell immunity for more effective Cancer immunotherapy. Front Immunol. 2018;9:14.
Wang Y, Kim TH, Fouladdel S, Zhang Z, Soni P, Qin A, et al. PD-L1 expression in circulating Tumor cells increases during radio(chemo)therapy and indicates poor prognosis in non-small cell Lung Cancer. Sci Rep. 2019;9(1):566.
Buggert M, Japp AS, Betts MR. Everything in its right place: resident memory CD8 + T cell immunosurveillance of HIV infection. Curr Opin HIV AIDS; 2018.
Gulzar N, Copeland KF. CD8 + T-cells: function and response to HIV infection. Curr HIV Res. 2004;2(1):23–37.
Cao W, Mehraj V, Kaufmann DE, Li T, Routy J-P. Elevation and persistence of CD8 T-cells in HIV infection: the Achilles heel in the ART era. J Int AIDS Soc. 2016;19(1):20697.
Doisne JM, Urrutia A, Lacabaratz-Porret C, Goujard C, Meyer L, Chaix ML, et al. CD8 + T cells specific for EBV, cytomegalovirus, and influenza virus are activated during primary HIV infection. J Immunol. 2004;173(4):2410–8.
Seddiki N, Kaufmann DE. Editorial overview: cell dysfunction and exhaustion in HIV infection. Curr Opin HIV AIDS. 2014;9(5):437–8.
Fenwick C, Joo V, Jacquier P, Noto A, Banga R, Perreau M, et al. T-cell exhaustion in HIV infection. Immunol Rev. 2019;292(1):149–63.
Meziane O, Alexandrova Y, Olivenstein R, Dupuy FP, Salahuddin S, Thomson E, et al. Peculiar phenotypic and cytotoxic features of pulmonary mucosal CD8 T cells in people living with HIV receiving long-term antiretroviral therapy. J Immunol. 2021;206(3):641–51.
de Boer WI, Sont JK, van Schadewijk A, Stolk J, van Krieken JH, Hiemstra PS. Monocyte chemoattractant protein 1, interleukin 8, and chronic airways inflammation in COPD. J Pathol. 2000;190(5):619–26.
Doz E, Noulin N, Boichot E, Guénon I, Fick L, Le Bert M, et al. Cigarette smoke-induced pulmonary inflammation is TLR4/MyD88 and IL-1R1/MyD88 signaling dependent. J Immunol. 2008;180(2):1169–78.
Arnson Y, Shoenfeld Y, Amital H. Effects of tobacco smoke on immunity, inflammation and autoimmunity. J Autoimmun. 2010;34(3):J258–65.
Nizri E, Irony-Tur-Sinai M, Lory O, Orr-Urtreger A, Lavi E, Brenner T. Activation of the cholinergic anti-inflammatory system by nicotine attenuates neuroinflammation via suppression of Th1 and Th17 responses. J Immunol. 2009;183(10):6681–8.
Lindblad SS, Mydel P, Jonsson I-M, Senior RM, Tarkowski A, Bokarewa M. Smoking and nicotine exposure delay development of collagen-induced arthritis in mice. Arthritis Res Therapy. 2009;11(3):1–8.
Karimi R, Tornling G, Grunewald J, Eklund A, Sköld CM. Cell recovery in Bronchoalveolar Lavage Fluid in smokers is dependent on cumulative smoking history. PLoS ONE. 2012;7(3):e34232.
Forsslund H, Mikko M, Karimi R, Grunewald J, Wheelock ÅM, Wahlström J, et al. Distribution of T-Cell subsets in BAL fluid of patients with mild to moderate COPD depends on current smoking status and not airway obstruction. Chest. 2014;145(4):711–22.
Yu MQ, Liu XS, Wang JM, Xu YJ. CD8(+) Tc-lymphocytes immunodeviation in peripheral blood and airway from patients of chronic obstructive pulmonary disease and changes after short-term smoking cessation. Chin Med J (Engl). 2013;126(19):3608–15.
Harrison OJ, Foley J, Bolognese BJ, Long E 3rd, Podolin PL, Walsh PT. Airway infiltration of CD4 + CCR6 + Th17 type cells associated with chronic cigarette smoke induced airspace enlargement. Immunol Lett. 2008;121(1):13–21.
Barnes PJ, Cosio MG. Characterization of T lymphocytes in chronic obstructive pulmonary disease. PLoS Med. 2004;1(1):e20–e.
Hayward SL, Scharer CD, Cartwright EK, Takamura S, Li Z-RT, Boss JM, et al. Environmental cues regulate epigenetic reprogramming of airway-resident memory CD8 + T cells. Nat Immunol. 2020;21(3):309–20.
Roberts LM, Evans TJ, Bosio CM. T cell metabolism is dependent on anatomical location within the lung. ImmunoHorizons. 2019;3(9):433.
Freeman CM, Han MK, Martinez FJ, Murray S, Liu LX, Chensue SW, et al. Cytotoxic potential of lung CD8(+) T cells increases with chronic obstructive pulmonary disease severity and with in vitro stimulation by IL-18 or IL-15. J Immunol. 2010;184(11):6504–13.
Chrysofakis G, Tzanakis N, Kyriakoy D, Tsoumakidou M, Tsiligianni I, Klimathianaki M, et al. Perforin expression and cytotoxic activity of Sputum CD8 + lymphocytes in patients with COPD. Chest. 2004;125(1):71–6.
Angelis N, Porpodis K, Zarogoulidis P, Spyratos D, Kioumis I, Papaiwannou A, et al. Airway inflammation in chronic obstructive pulmonary disease. J Thorac Disease. 2014;6(Suppl 1):S167–72.
Velotti F, Barchetta I, Cimini FA, Cavallo MG. Granzyme B in Inflammatory diseases: apoptosis, inflammation, Extracellular Matrix Remodeling, epithelial-to-mesenchymal transition and fibrosis. Front Immunol. 2020;11:587581.
Shiratsuchi N, Asai K, Kanazawa H, Kyoh S, Tochino Y, Kodama T, et al. Measurement of soluble perforin, a marker of CD8 + T lymphocyte activation in epithelial lining fluid. Respir Med. 2011;105(12):1885–90.
Olloquequi J, Ferrer J, Montes JF, Rodriguez E, Montero MA, Garcia-Valero J. Differential lymphocyte infiltration in small airways and lung parenchyma in COPD patients. Respir Med. 2010;104(9):1310–8.
Siena L, Gjomarkaj M, Elliot J, Pace E, Bruno A, Baraldo S, et al. Reduced apoptosis of CD8 + T-lymphocytes in the airways of smokers with mild/moderate COPD. Respir Med. 2011;105(10):1491–500.
Williams M, Todd I, Fairclough LC. The role of CD8 + T lymphocytes in chronic obstructive pulmonary disease: a systematic review. Inflamm Research: Official J Eur Histamine Res Soc [et al]. 2021;70(1):11–8.
Salahuddin S, Thomson E, Méziane O, Farnos O, Pagliuzza A, Chomont N et al. Processing of Bronchoalveolar Lavage Fluid and matched blood for alveolar macrophage and CD4 + T-cell immunophenotyping and HIV Reservoir Assessment. J Visualized Experiments: JoVE. 2019(148).
Morris SE, Farber DL, Yates AJ. Tissue-Resident Memory T cells in mice and humans: towards a quantitative Ecology. J Immunol. 2019;203(10):2561.
Herndler-Brandstetter D, Ishigame H, Shinnakasu R, Plajer V, Stecher C, Zhao J, et al. KLRG1(+) Effector CD8(+) T cells lose KLRG1, differentiate into all memory T cell lineages, and convey enhanced protective immunity. Immunity. 2018;48(4):716–e298.
Bottois H, Ngollo M, Hammoudi N, Courau T, Bonnereau J, Chardiny V et al. KLRG1 and CD103 Expressions Define Distinct Intestinal Tissue-Resident Memory CD8 T Cell Subsets Modulated in Crohn’s Disease. Front Immunol. 2020;11.
Takamura S, Kato S, Motozono C, Shimaoka T, Ueha S, Matsuo K, et al. Interstitial-resident memory CD8 + T cells sustain frontline epithelial memory in the lung. J Exp Med. 2019;216(12):2736–47.
Wein AN, McMaster SR, Takamura S, Dunbar PR, Cartwright EK, Hayward SL, et al. CXCR6 regulates localization of tissue-resident memory CD8 T cells to the airways. J Exp Med. 2019;216(12):2748–62.
Buggert M, Vella LA, Nguyen S, Wu VH, Chen Z, Sekine T, et al. The identity of human tissue-emigrant CD8(+) T cells. Cell. 2020;183(7):1946–e6115.
Van Gassen S, Callebaut B, Van Helden MJ, Lambrecht BN, Demeester P, Dhaene T, et al. FlowSOM: using self-organizing maps for visualization and interpretation of cytometry data. Cytometry Part A. 2015;87(7):636–45.
Ngan DA, Vickerman SV, Granville DJ, Man SFP, Sin DD. The possible role of granzyme B in the pathogenesis of chronic obstructive pulmonary disease. Ther Adv Respir Dis. 2009;3(3):113–29.
Gerlach C, Moseman EA, Loughhead SM, Alvarez D, Zwijnenburg AJ, Waanders L, et al. The chemokine receptor CX3CR1 defines three Antigen-experienced CD8 T cell subsets with distinct roles in Immune Surveillance and Homeostasis. Immunity. 2016;45(6):1270–84.
Caserta S, Borger J, Zamoyska R. Central and Effector Memory CD4 and CD8 T-Cell responses to Tumor-Associated antigens. Crit Rev Immunol. 2012;32:97–126.
McMaster S, Wilson J, Wein A, Dunbar P, Matloubian M, Kohlmeier J. Local antigen and inflammation in the lung induce the preferential establishment and maintenance of CXCR6 and CD49a-expressing antigen-specific CD8 TRM cells in the lung parenchyma and airways (MUC1P.900). J Immunol. 2015;194(1Supplement):641–1.
Ashhurst AS, Flórido M, Lin LCW, Quan D, Armitage E, Stifter SA et al. CXCR6-Deficiency improves the control of pulmonary Mycobacterium tuberculosis and Influenza Infection Independent of T-Lymphocyte Recruitment to the lungs. Front Immunol. 2019;10.
Freeman CM, Curtis JL, Chensue SW. CC chemokine receptor 5 and CXC chemokine receptor 6 expression by lung CD8 + cells correlates with chronic obstructive pulmonary disease severity. Am J Pathol. 2007;171(3):767–76.
Daniel JP, Surita D, Richard L, Richard P, Stephen G, Clive SM et al. The CXCR6/CXCL16 axis links inflamm-aging to disease severity in COVID-19 patients. bioRxiv. 2021:2021.01.25.428125.
Wasén C, Turkkila M, Bossios A, Erlandsson M, Andersson KM, Ekerljung L, et al. Smoking activates cytotoxic CD8(+) T cells and causes survivin release in rheumatoid arthritis. J Autoimmun. 2017;78:101–10.
Hodge S, Hodge G, Nairn J, Holmes M, Reynolds PN. Increased airway granzyme b and perforin in current and ex-smoking COPD subjects. Copd. 2006;3(4):179–87.
Martos SN, Campbell MR, Lozoya OA, Wang X, Bennett BD, Thompson IJB, et al. Single-cell analyses identify dysfunctional CD16 + CD8 T cells in smokers. Cell Rep Med. 2020;1(4):100054.
Corleis B, Cho JL, Gates SJ, Linder AH, Dickey A, Lisanti-Park AC, et al. Smoking and human immunodeficiency virus 1 infection promote Retention of CD8(+) T cells in the Airway Mucosa. Am J Respir Cell Mol Biol. 2021;65(5):513–20.
Agostini C, Cabrelle A, Calabrese F, Bortoli M, Scquizzato E, Carraro S, et al. Role for CXCR6 and its ligand CXCL16 in the pathogenesis of T-Cell Alveolitis in Sarcoidosis. Am J Respir Crit Care Med. 2005;172(10):1290–8.
Marques P, Collado A, Escudero P, Rius C, González C, Servera E, et al. Cigarette smoke increases endothelial CXCL16-Leukocyte CXCR6 adhesion in Vitro and in vivo. Potential consequences in Chronic Obstructive Pulmonary Disease. Front Immunol. 2017;8:1766.
Collini PJ, Bewley MA, Mohasin M, Marriott HM, Miller RF, Geretti AM, et al. HIV gp120 in the lungs of antiretroviral therapy-treated individuals impairs alveolar macrophage responses to Pneumococci. Am J Respir Crit Care Med. 2018;197(12):1604–15.
Gundavarapu S, Mishra NC, Singh SP, Langley RJ, Saeed AI, Feghali-Bostwick CA, et al. HIV gp120 induces mucus formation in human bronchial epithelial cells through CXCR4/α7-nicotinic acetylcholine receptors. PLoS ONE. 2013;8(10):e77160.
Iyengar S, Schwartz DH, Hildreth JE. T cell-tropic HIV gp120 mediates CD4 and CD8 cell chemotaxis through CXCR4 independent of CD4: implications for HIV pathogenesis. J Immunol. 1999;162(10):6263–7.
Kitchen SG, LaForge S, Patel VP, Kitchen CM, Miceli MC, Zack JA. Activation of CD8 T cells induces expression of CD4, which functions as a chemotactic receptor. Blood. 2002;99(1):207–12.
Hudson WH, Gensheimer J, Hashimoto M, Wieland A, Valanparambil RM, Li P, et al. Proliferating transitory T cells with an effector-like transcriptional signature emerge from PD-1 + stem-like CD8 + T cells during chronic infection. Immunity. 2019;51(6):1043–e584.
Mudd JC, Panigrahi S, Kyi B, Moon SH, Manion MM, Younes SA, et al. Inflammatory function of CX3CR1 + CD8 + T cells in treated HIV infection is modulated by platelet interactions. J Infect Dis. 2016;214(12):1808–16.
Zhang J, Patel JM. Role of the CX3CL1-CX3CR1 axis in chronic inflammatory lung diseases. Int J Clin Exp Med. 2010;3(3):233–44.
Yassouf MY, Zhang X, Huang Z, Zhai D, Sekiya R, Kawabata T, et al. Biphasic effect of mechanical stress on lymphocyte activation. J Cell Physiol. 2022;237(2):1521–31.
Koliński T, Marek-Trzonkowska N, Trzonkowski P, Siebert J. Heat shock proteins (HSPs) in the homeostasis of regulatory T cells (Tregs). Cent Eur J Immunol. 2016;41(3):317–23.
Gómez-Mora E, Carrillo J, Urrea V, Rigau J, Alegre J, Cabrera C et al. Impact of Long-Term Cryopreservation on Blood Immune cell markers in myalgic Encephalomyelitis/Chronic fatigue syndrome: implications for Biomarker Discovery. Front Immunol. 2020;11.
Acknowledgements
We thank all the participants who underwent research procedures in this study as well as research nurse Josée Girouard for nursing care and the respiratory therapists for assistance with bronchoscopies throughout this study.
Funding
This study was funded by the Canadian Institutes of Health Research (CIHR, operating grant 153082 to CTC, ES, MAJ; and the Canadian HIV Cure Enterprise [CanCURE] Team Grant HB2 164064) to CTC, JE, MAJ; and in part, by the AIDS and Infectious Diseases Network of the Réseau SIDA et maladies infectieuses du Fonds de recherche du Québec-Santé (FRQ-S) to CTC, MAJ. YA was supported by an FRQ-S MSc scholarship. AY was supported by an FRQ-S doctoral scholarship. JE holds the CIHR Canada Research Chair Tier 1 in Pathophysiology of Cell Death in Host-Pathogen Interactions. CTC holds an FRQ-S Junior 2 Clinician-researcher career award. MAJ holds the tier 2 CIHR Canada Research Chair in Immuno-Virology. The funding institutions played no role in the design, collection, analysis, and interpretation of data.
Author information
Authors and Affiliations
Contributions
Study design: CTC, MAJ. Study conceptualization: CTC, JE, MAJ. Supervision: MAJ. Funding acquisition: CTC, MAJ. Performance of the experiments and data generation: YA, AY. Discussion and data analysis and interpretation: YA, AY, JE, CTC, MAJ. Access to BAL specimens and clinical data: RO, MO, ES, CTC. Manuscript writing and formatting: YA, MAJ. All authors contributed to the refinement of the study and reviewed, revised, and approved the final version of the manuscript.
Corresponding author
Ethics declarations
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Ethics approval
This study has been ethically approved by the Research Institute of the McGill University Health Centre (#15–031) and Université du Québec à Montréal (UQAM) (#602) and procedures performed in accordance with the Declaration of Helsinki All participants provided written informed consent beforehand.
Additional information
Abstracts of data included in the paper have previously been presented at the AIDS 2022 Conference, Montreal, QC, Canada; and, at the IAS 2023 conference, Brisbane, Australia.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Alexandrova, Y., Yero, A., Olivenstein, R. et al. Dynamics of pulmonary mucosal cytotoxic CD8 T-cells in people living with HIV under suppressive antiretroviral therapy. Respir Res 25, 240 (2024). https://doi.org/10.1186/s12931-024-02859-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12931-024-02859-2