
Abstract—
The results of the palynological study of the Paleogene and the underlying Cretaceous sediments of the Cis-Donets Monocline, drilled by borehole 1238, are presented. Analysis of dinoflagellate cysts enabled the recognition of the Apectodinium hyperacanthum, Axiodinium augustum, and Ochetodinium romanum/Samlandia chlamydophora zone intervals and the Rhombodinium draco–Deflandrea spinulosa layers in the Paleogene part of the section. The age of regional formations and beds was updated: the Buzinovka Formation is dated by the early Thanetian; the Veshenskaya Formation is late Thanetian–earliest Ypresian age; the Surovikino and Osinovaya Beds are early–middle Ypresian; and the Ventsy and “Poltava” Beds are of the late Rupelian–Chattian age. Two major stratigraphic hiatuses, corresponding to the Maastrichtian–Selandian and the upper Ypresian–lower Rupelian, were recognized in the borehole 1238 section. The analysis of the quantitative fluctuations of different palynomorph groups through the section permitted to reconstruct changes in depositional environments of the Peri-Tethys Basin margin during the early and late Paleogene: the Buzinovka and Veshenskaya formations and the Surovikino and Osinovaya Beds were accumulated in an open-marine environments during the long late Paleocene–early Ypresian transgressive stage, whereas the Ventsy and “Poltava” beds were deposited in the half-landlocked basin during the Chattian.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
INTRODUCTION
The principal task of the present-day geological studies of the Paleogene is the reconstruction of the evolution of marine basins during various time intervals and related possible migrations of various fauna and flora groups. It is obvious that the deepest parts of paleobasins are naturally characterized by the most complete stratigraphic successions and paleontological characteristics, whereas the marginal parts, as a rule, demonstrate wide facies variations, often the absence or low saturation of fossils. This often leads to disagreement in understanding of the age and volume of local and regional lithostratigraphic units.
The subregion of the eastern surrounding of Donbass is one of these marginal marine basins in the south of the Russian Platform (Akhmetiev and Beniamovsky, 2003). Initially this territory belonged to the Donets Syneclise of the Dnieper–Donets Depression (Leonov, 1961) and later, to the subregion of the Voronezh Anteclise (Semenov, 1965, 1975). Generally, during the Paleogene, the Dnieper–Donets Basin, Voronezh Anteclise, and the eastern part of the Greater Donbass represented a single marine area inside the Peri-Tethys, through which, during several Paleogene intervals, sublatitudinal communications with the seas on the territory of the present-day Belarus, Poland, the Baltics, as well as the former North Sea Basin were carried out.
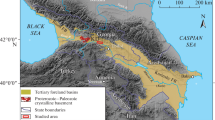
Three structural-facies zones (SFZs), differing in lithological structure and thicknesses (and sometimes in the enlistment) of their lithostratigraphic units and taking into account their facies and structural features, have been established in the Regional Paleogene Stratigraphic Scheme for the Subregion of the Eastern Greater Donbass, approved in 2000 by the Interdepartmental Stratigraphic Committee of Russia (hereafter the Scheme of 2000): (1) the Cis-Donets Monocline; (2) directly Eastern Donbass; and (3) the southern and eastern surroundings of the Donbass (Fig. 1) (Postanovleniya…, 2001; Akhmetiev and Beniamovsky, 2003). The established SFZs reflect facies transitions from the shallow-water sediments in the north to the deeper ones in the south, sometimes taking the effect of local consedimentary structures (depressions and uplifts) into consideration.

Location of studied sections (a) and the structural-facies zonation of the eastern Greater Donbass Subregion (b) (modified after Akhmetiev and Beniamovsky, 2003).
The lithostratigraphic units for the Cis-Donets Monocline, which occupies the northernmost position in the subregion and is characterized by the terrigenous and terrigenous–siliceous facies, were established first by Leonov (1936, 1939, 1961) and later by Semenov (1965) (Fig. 2); it should be noted that the age of most of them was based on the malacofauna data.

Paleogene stratigraphical schemes of the subregion of the eastern surrounding of Donbass.
According to Akhmetiev and Beniamovsky (2003), the early Paleogene horizons on the territory of the Cis-Donets Monocline correspond to eponymous formations. Thus, in the Scheme of 2000, for the whole subregion of eastern Greater Donbass, the Sumy suprahorizon is used as a regional Paleocene subdivision and includes the Tatsinskaya, Buzinovka, and Veshenskaya horizons; the Sal horizon (with three subhorizons), the Kuberla, Keresta, Kuma, Kiev, and Obukhov horizons are used for the Eocene; and, finally, the Khadum suprahorizon and the Tsimlyanskaya, Solenaya, Kalmykia, and Bereka horizons are used for the Oligocene (Fig. 2) (Postanovleniya…, 2001). Thus, the stratigraphic scheme applies the horizons of Northern Ukraine (Sumy Superhorizon; Kiev, Obukhov, and Bereka horizons); the lithostratigraphic units of Semenov (1975) for the Voronezh Anteclise; and from the schemes of the Lower Volga region, Scythian Platform, and the North Caucasus. On that point, it was noted (Akhmetiev and Beniamovsky, 2003, p. 48) that the Paleogene horizons of Northern Ukraine were used for the SFZs of the Cis-Donets Monocline. At the present time, different groups of specialists use local lithostratigraphic units for the subdivision of the Paleogene deposits in this region, proposed for Northern Ukraine and the Voronezh Anteclise as well as for the Volga region and Scythian Platform. This complicates the understanding of the spatial and temporal relations between the sequences and their exact stratigraphic ages.
It is important to note that during the last time Ukrainian and Russian specialists introduced a number of modifications and precisions to the stratigraphic scheme of the northern regions of Ukraine (for instance, the Sumy Superhorizon was abolished) (Zosimovich et al., 2005, 2013, 2015, 2016, 2017; Zosimovich, 2013; Zosimovich and Shevchenko, 2014, 2015; Oreshkina and Iakovleva, 2007; Iakovleva, 2015; Musatov and Ryabokon’, 2017) as well as to the schemes of the Voronezh Anteclise (Gorbatkina and Iosifova, 2004; Shpul’, 2005, 2006, 2007, 2008, 2009, 2010; Bugrova et al., 2016; Musatov, 2020) and the Pre-Caspian and Volga regions (Vasil’eva, 2013, 2019; Musatov et al., 2014; Musatov and Bogachkin, 2019a, 2019b; Unifitsirovannaya…, 2015; Zastrozhnov et al., 2019).
Recently, Beniamovsky (2016, 2017a, 2017b, 2017c), on the basis of new or recently published data, undertook the revision of the Scheme of 2000 for the subregions of the eastern part of the Greater Donbass and the Voronezh Anteclise, which he united into a single Voronezh–Cis-Donets subregion. Beniamovsky proposed his own structural-facies zonation and used all formations, horizons, and superhorizons from Semenov’s scheme in the draft version of the new regional scheme. It should be noted that almost all local lithostratigraphic units in this version of the regional stratigraphic scheme for the subregion contain hiatuses and that none of them has conformable contacts (Beniamovsky, 2017b); nevertheless, no explanations for these facts were given in publications.
The insufficient paleontological knowledge of the Paleogene of southern Russian Platform, the application of different local lithostratigraphic units (formations, beds) by different groups of specialists, the absence of a robust definition of the stratotypes of regional stages, and, finally, the necessity to update the unified regional Paleogene chronostratigraphic scheme determine the importance of new integrated studies in the region of the Cis-Donets Monocline and Voronezh Anteclise.
The object of our studies in 2018 was the territory of the Cis-Donets Monocline, where we tried to revise the stratotypes of the lithostratigraphic units of the Paleogene, introduced by Leonov as beds (1936, 1939, 1961) and upgraded to formations by Semenov (1965). However, only some of these formations had paleontological age based on the malacofauna. It should be also noted that Leonov and Semenov considered the Danian Stage as a part of the Cretaceous System. For this reason, the characteristics of the Lower Paleocene rocks were not included in the publications of these two authors.
During our fieldwork, we found that most of the type sections of the Kumovka, Buzinovka, Veshenskaya, and Surovikino formations introduced by Leonov are presently inaccessible for direct observation because of thick grass and forest coverage. Unfortunately, the stratotypes of the Sheptukhovka and Osinovaya formations established by Semenov and located in the Russian–Ukrainian border zone, are presently not available for examination. It was found that the Paleogene deposits in the studied region are exposed in the isolated small outcrops, as “windows” on gully or clough slopes, or in shallow opencast mines; however, their contacts with the underlying and overlying strata have not been observed anywhere. Field observations and analysis of published data suggest that the Paleogene deposits in the Cis-Donets Monocline are represented mostly by monotonous sequences of sands, sandstones, siltstones, and opokas with rare clay interbeds and lenses, and pebble or phosphoritic horizons. Our integrated lithological–petrographic and micropaleontological research in more than ten natural Paleogene outcrops of the Paleogene revealed the absence of foraminifers, diatoms algae, and silicoflagellates, as well as dinoflagellate cysts and spores and pollen in all collected samples. The paleontological characteristics (mostly by dinoflagellate cysts and diatoms) in the studied region were obtained only from a few boreholes.
The aims of this study were the detailed integrated palynostratigraphic analysis of the Paleogene sediments in the section of borehole no. 1238, age update for local and regional stratigraphic units of the Cis-Donets Monocline, correlation of these deposits with the successions in adjacent regions, and reconstruction of depositional environments.
MATERIALS AND METHODS
In the scope of the present study, we analyzed 63 samples from borehole no. 1238, drilled in the vicinity of Bazkovskaya Stanitsa of the Rostov oblast (54°89′10′′ N, 77°27′47′′ E) by the North Caucasus Geological Production Association and the Karpinsky Russian Geological Research Institute (VSEGEI) in the scope of geological survey work on the Geological Sheet M-37-XXIV with the objective to subdivide the Paleogene sequence (Fig. 1).
A field lithological description (from base to top) of the section drilled by borehole no. 1238 according to the scheme of Leonov (1961) is given below (Fig. 3). The lithological description was made by A.S. Zastrozhnov and S.V. Popov; the subdivision, by G.N. Aleksandrova.

Lithology and stratigraphic subdivision of the geological section of borehole no. 1238. The colors inside the lithological column correspond to rock colors.
Boguchar Formation:
Bed 1, 100–91.7 m. Gray and light-gray compact, massive, micaceous marl with occasional mud eater burrows filled by the light marly material.
Buzinovka Formation:
Bed 2, 91.7–90.2 m. Resting upon it with a well-defined boundary are sands dark gray with a greenish tint, glauconite–quartz, micaceous, inequigranular and coarse with small (10–15 mm diameter) pebbles and coarse phosphorite gravels at the base and grading upward into medium and clayey sands.
Bed 3, 90.2–86.5 m. Resting upon them with a well-defined boundary are dark gray to black compact silty clays, which grade upward into clayey siltstones with a greenish tint (by glauconite), slightly siliceous, slightly bioturbated, with interbeds (up to 0.05 m) of sandstones (siltstones) of the same composition.
Bed 4, 86.5–82.0 m. Dark gray medium to fine clayey sand with glauconite at the base of the bed and medium to coarse sand with glauconite grains in the upper meter.
Bed 5, 82.0–81.7 m. Hard silicified fine (up to siltstone) glauconitic sandstone with a distinctive emerald green color.
Veshenskaya Formation:
Bed 6, 81.7–5.5 m. Light-gray opoka-like clay, very light, up to the tripolith (5 cm); above is slightly siliceous micaceous clay with glauconitic inclusions. The contact with the underlying rocks is distinct, with a basal horizon, composed by gravel-size quartz and phosphorite grains (1–4 mm in diameter).
Bed 7, 75.5–71.5 m. Resting upon it with indistinct contact are dark gray sands with a greenish tint, clayey, slightly siliceous, fine-grained, micaceous, with gray and light-gray zones, which imparts a mottled appearance to the rock.
Bed 8, 71.5–67.5 m. Gray micaceous, slightly siliceous clayey siltstones.
Surovikino Beds:
Bed 9, 67.5–60.5 m. Well-sorted fine and medium quartz sand, white at the base (0.3 m) and with a yellowish tint higher in the sequence. The contact with the underlying rocks is not recovered.
Osinovaya Beds:
Bed 10, 60.5–59.6 m. Clayey siltstone with bunches of fine clayey sand, with ferruginization patches, interbeds, and bunches. The color of rock is brown–crimson, ocherous.
Bed 11, 59.6–58.8 m. Brown “coffee” non-calcareous clays with fine gray glauconite–quartz sand bunches at the top of the bed.
Bed 12, 58.8–58.1 m. Light-gray fine opoka-like light glauconite–quartz sandstone.
Bed 13, 58.1–57.4 m. Brown, slightly greenish clays with indistinct platy parting and numerous mud eater burrows filled with pale fine glauconite–quartz sand.
Bed 14, 57.4–56.5 m. Light-gray, slightly yellowish-brown fine siltstone, slightly micaceous, with ferrugination stains.
Ventsy Beds:
Bed 15, 56.5–53.5 m. Greenish medium- and fine-grain sandstone at the base (0.2 m). Above is gray micaceous siltstone with dark gray clay lenses and thin lenticular interbeds, which imparts a stratified appearance to the rock, with numerous sorted small fragments of plant tissues. The contact with the underlying rocks is not recovered.
“Poltava” Beds:
Bed 16, 53.5–52.0 m. Compact variegated sandy clay: greenish gray with interbeds of red and yellow ferrugination (ferrugination intensity increases toward the base of the bed) and light-gray siltstones, which imparts a banded structure to the rock. The contact with the underlying rocks is not recovered.
Bed 17, 52.0–45.3 m. Light-gray and greenish gray siltstone with yellowish ferrugination interbeds; micaceous. The ferruginous interbeds contain occasional sandstone fragments. Greenish gray clay occurs in the interval of 46.3–46.6 m.
Bed 18, 45.3–38.8 m. Dusty greenish-gray sand with interbeds of red and yellow ferrugination. Light-gray siltstone interbeds up to 5 cm thick at 44.1–44.2, 42.8, and 42.3 m depth. The contact with underlying rocks is erosional.
Bed 19, 38.8–36.5 m. Fine greenish light-gray sand with thin interbeds of light-gray siltstone and ferrugination. The latter impart a banded structure to the rock (9–10 interbeds per 1 m of core). The contact with the underlying rocks is marked by an increase in the number of ferruginated interbeds.
Bed 20, 36.5–30.7 m. Resting upon it with a well-defined boundary is fine yellow-green ferruginated sand with white cemented sand inclusions and thin interbeds of light-gray clayey material.
Bed 21, 30.7–28.5 m. Resting upon it with a well-defined boundary is light-brown massive fine sand.
Bed 22, 28.5–23.1 m. Resting upon it with a well-defined boundary is yellow fine micaceous slightly cemented sandstone with indistinctly layered texture, with ferrugination in the form of red sinters. A white sand interbed in the interval of 26.2–26.1 m depth.
Bed 23, 23.1–20.6 m. Resting upon it with a well-defined boundary is a unit of alternating beds (25 cm) of compact crimson, gray, and yellow clays; ocherous siltstones; and soft fine red sandstones—the weathering crust on the underlying sediments.
The section is overlain by solid brown Quaternary loam 20 m thick.
Palynological samples were prepared according to the standard techniques using at the Laboratory of Paleofloristics, Geological Institute, Russian Academy of Sciences (GIN RAS): (1) the treatment of the samples with 10% hydrochloric acid (HCl) in order to remove carbonates; (2) the treatment with hot tetrasodium pyrophosphate (Na4P2O7∙10H2O) solution in order to disaggregate the clayey material and the subsequent washing of samples every two hours to remove argillaceous compounds; (3) the centrifugation of samples with a heavy liquid (K2CdI4) with a density of 2.25 in order to separate the organic fraction from the heavier mineral particles; (4) the treatment with 70% hydrofluoric acid (HF) in order to dissolve siliceous matter; (5) the treatment with 10% hydrochloric acid to remove fluorosilicate gels; and (6) washing of samples in distilled water and covering them with glycerine. The macerate was not sieved.
The dinocyst taxonomy follows the DINOFLAJ3 Index (Williams et al., 2017). The quantitative palynological study of the material was performed in two steps: (1) a minimum of 200–250 marine and terrestrial palynomorphs (dinoflagellate cysts, acritarchs, prasinophytes, angiosperm and gymnosperm pollen, and foraminiferal linings) were counted in each productive sample; (2) a minimum of 200 dinocysts were counted; subsequently, remaining material was scanned for rare taxa. Paleoecological interpretation of quantitative fluctuations in the Paleogene palynological assemblages was based on publications (Brinkhuis, 1994; see Table 1 in Iakovleva and Heilmann-Clausen, 2020). For interpreting possible paleoenvironments, dinoflagellate cysts were combined in 17 groups of morphologically close taxa: (1) Deflandrea-group (Deflandrea, Lejeunecysta, Lentinia, Senegalinium); (2) wetzelielloids; (3) Phthanoperidinium; (4) Fromea fragilis; (5) Lingulodinium machaerophorum; (6) Areoligera-group (Areoligera, Glaphyrocysta, Emmetrocysta, Adnatosphaeridium, Membranilarnacia); (7) Thalassiphora; (8) Ochetodinium romanum; (9) Elytrocysta brevis; (10) Cribroperidinium; (11) Carpatella rossica; (12) Cordosphaeridium-group (Cordosphaeridium, Fibrocysta); (13) Operculodinium; (14) Diphyes/Dapsilidinium; (15) Spiniferites-group (Achomosphaera, Hystrichosphaeropsis, Hystrichostrogylon, Spiniferella, Spiniferites, Rottnestia); (16) Impagidinium; (17) other gonyaulacoids.
The chronostratigraphic age is given according to Ogg et al. (2016).
Palynological material is stored at the Laboratory of Paleofloristics, GIN RAS (Moscow).
RESULTS
The main objective of this study was the palynological analysis of the Paleogene sediments, drilled by the borehole no. 1238. Stratigraphic and quantitative data are shown in Figs. 4–6. Palynological data from the underlying Upper Cretaceous deposits recovered at the bottom of this core are given in brief. The most typical dinoflagellate cyst taxa from the Cretaceous–Paleogene deposits are shown in Plates I–XI.

Stratigraphic distribution of dinocyst species in the Paleogene part of the section of borehole no. 1238.

(Contd.)

(Contd.)

(Contd.)

Quantitative distribution of principal palynomorph groups in the Paleogene section of the borehole no. 1238.

Quantitative distribution of dinocyst ecogroups in the Paleogene section of the borehole no. 1238.

Plate I . Dinocysts from the Santonian–Campanian sediments from the borehole no. 1238. Magnification is given in microns (µm). (1) Odontochitina costata (Alberti) Clark et Verdier, 92 m depth; (2) Manumiella sp., 92 m depth; (3, 6, 9, 12) Cribroperidinium wilsonii subsp. trabeculosum Slimani et Louwye, 92 m depth; (4, 5) Laciniadinium firmum (Harland) Morgan, 92 m depth; (7, 10) Xenascus ceratioides (Deflandre) Lentin et Williams, 92 m depth; (8, 11) Isabelidinium microarmum (McIntyre) Lentin et Willams: (8) 96 m depth, (11) 92 m depth; (13) Chatangiella vnigrii (Vozzhennikova) Lentin et Willams, 93 m depth; (14, 15) Spinidinium echinoideum (Cookson et Eiselanck) Lentin et Willams, 92 m depth.

Dinocysts from the Santonian–Campanian sediments from the borehole no. 1238. Magnification is given in microns (µm). (1, 2) Hystrichosphaeropsis quasicribrata (Wetzel) Gocht, 92 m depth; (3, 4) Chatangiella vnigrii (Vozzhennikova) Lentin et Willams, 96 m depth; (5) ?Alterbidinium sp., 92 m depth; (6) Subtilisphaera perlucida (Alberti) Jain et Millepied, 96 m depth; (7, 12) Isabelidinium sp., 92 m depth; (8) Isabelidinium pellucidum (Deflandre et Cookson) Lentin et Willams, 92 m depth; (9, 10) Apteodinium deflandrei (Clarke et Verdier) Lucas-Clarck, 92 m depth; (11, 14) Isabelidinium acuminatum (Cookson et Eisenack) Stover et Evitt: (11) 92 m depth, (14) 93 m depth; (13) Isabelidinium microarmum (McIntyre) Lentin et Willams, 93 m depth; (15, 16) Alisocysta sp. 2 sensu Heilmann-Clausen, 1985, 90 m depth; (17, 18) Conneximura fimbriata (Morgenroth) May, 86.7 m depth.

Dinocysts from the Paleogene sediments from the borehole no. 1238. Magnification is given in microns (µm). (1, 5) Axiodinium augustum (Harland) Williams et al., 70.1 m depth; (2, 6) Dracodinium samlandicum (Eisenack) Costa et Downie, 59.3 m depth; (3, 4) Samlandia chlamydophora Eisenack, 59.3 m depth; (7, 8) Cerebrocysta bartonensis Bujak, 59.3 m depth; (9, 10, 13) Impagidinium wardii Iakovleva et Heilmann-Clausen, 58.0 m depth; (11) Dracodinium brevicornutum (Heilmann-Clausen) Williams et al., 54.0 m depth; (12) Dracodinium sp., latiepeliform archeopyle, 58.0 m depth; (14, 15) Phthanoperidinium sp., 58 m depth; (16, 17) Membranilarnacia compressa Bujak, 58 m depth; (18, 19) Spiniferites ramosus (Ehrenberg) Mantell, 59.3 m depth; (20) Ochetodinium romanum Damassa, 59.3 m depth.

Dinocysts from the Paleogene sediments from the borehole no. 1238. Magnification is given in microns (µm). (1, 4, 7) Dracodinium samlandicum (Eisenack) Costa et Downie, 58 m depth; (2, 3) Fibrocysta bipolaris (Cookson et Eisenack) Stover et Evitt, 59.3 m depth; (5) Fibrocysta essentialis (De Coninck) Brinkhuis et Zachariasse, 59.3 m depth; (6) Charlesdowniea coleothrypta (Williams et Downie) Lentin et Vozzhennikova, 59.3 m depth; (8, 9, 11, 12) Hystrichokolpoma spinosum Wilson, 59.3 m depth; (10, 13, 14) Impagidinium wardii Iakovleva et Heilmann-Clausen, 57.8 m depth; (15) Heteraulacacysta leptalea Eaton, 59.3 m depth; (16, 17) Deflandrea denticulata Alberti, 70.1 m depth; (18) Deflandrea cf. denticulata Alberti, 59.3 m depth.

Dinocysts from the Paleogene sediments from the borehole no. 1238. Magnification is given in microns (µm). (1, 2, 6) Axiodinium prearticulatum Williams et al., 58.0 m depth; (3–5) Impagidinium wardii Iakovleva et Heilmann-Clausen, 58.0 m depth; (7, 8) Dracodinium sp. A, latiepeliform archeopyle, 58.0 m depth; (9, 10) Membranilarnacia compressa Bujak, 59.3 m depth; (11, 12) Phthanoperidinium sp., 58.0 m depth; (13, 14) Eatonicysta ursulae (Morgenroth) Stover et Evitt, 58.0 m depth; (15) Diphyes pseudoficusoides Bujak, 58.0 m depth.

Dinocysts from the Paleogene sediments from the borehole no. 1238. Magnification is given in microns (µm). (1, 4, 9) Axiodinium prearticulatum Williams et al., 59.3 m depth; (2, 3) Dracodinium sp. A, 57.8 m depth; (5) Axiodinium prearticulatum Williams et al., 58.0 m depth; (6, 7) Carpatella rossica Iakovleva, 81.4 m depth; (8) Dracodinium sp. A, 58.0 m depth; (9) Charlesdowniea coleothrypta (Williams et Downie) Lentin et Vozzhennikova, 59.3 m depth; (10, 11) Diphyes ficusoides Islam, 58.0 m depth; (12, 13) Kallosphaeridium yorubaense Jan du Chene et Adediran, 58.0 m depth; (14) Fibrocysta vectensis (Eaton) Stover et Evitt, 58.0 m depth; (15) Membranilarnacia compressa Bujak, 58.0 m depth.

Dinocysts from the Paleogene sediments from the borehole no. 1238. Magnification is given in microns (µm). (1‒3) Cordosphaeridium gracile (Eisenack) Davey et Williams, 57.8 m depth; (4–7) Ochetodinium romanum Damassa, 59.3 m depth; (8) Axiodinium prearticulatum Williams et al., 58.0 m; (9, 12) Samlandia chriskingii Iakovleva et Heilmann-Clausen, 59.3 m depth; (10, 11) Diphyes pseudoficusoides Bujak, 58.0 m depth; (13, 14) Hystrichosphaeropsis sp., 58.0 m depth.

Dinocysts from the Paleogene sediments from the borehole no. 1238. Magnification is given in microns (µm). (1, 4) Thalassiphora dominiquei Iakovleva et Heilmann-Clausen, 59.3 m depth; (2) Cribroperidinium tenuitabulatum (Gerlach) Helenes, 59.3 m depth; (3, 7) Rhombodinium draco Gocht, 54.0 m depth; (5, 6) Eisenackia circumtabulata Drugg, 59.3 m depth; (8, 11) Piladinium columna (Michoux) Williams et al., 59.3 m depth; (9) Dracodinium eocaenicum (Agelopoulos) Williams et al., 54.0 m depth; (10) Dracodinium sp. A, 57.8 m depth; (12, 13) Ochetodinium romanum Damassa, 59.3 m depth; (14) Rhombodinium draco Gocht, free operculum, 54.0 m depth.

Dinocysts from the Paleogene sediments from the borehole no. 1238. Magnification is given in microns (µm). (1, 2, 4, 5) Cordosphaeridium inodes (Klumpp) Eisenack, 57.8 m depth; (3, 6, 11, 15) Achilleodinium biformoides (Eisenack) Eaton, 59.3 m depth; (7–10) Hystrichokolpoma cinctum Klumpp, 59.3 m depth; (12–14) Hystrichokolpoma cf. cinctum Klumpp, 57.8 m depth.

Dinocysts from the Paleogene sediments from the borehole no. 1238. Magnification is given in microns (µm). (1, 5, 9, 13) Hystrichosphaeropsis sp., 57.8 m depth; (2, 6, 10) Piladinium columna (Michoux) Williams et al., 59.3 m depth; (3, 7, 11) Hystrichosphaeropsis sp., 57.0 m depth; (4, 8, 12) Areoligera sentosa-group, 58 m depth; (14) Tectatodinium pellitum Wall, 59.3 m depth; (15) Phthanoperidinium sp., 58.0 m depth; (16) Piladinium columna (Michoux) Williams et al., 59.3 m depth; (17, (18) Deflandrea oebisfeldensis Alberti, 70.1 m depth.

Dinocysts from the Paleogene sediments from the borehole no. 1238. Magnification is given in microns (µm). (1, 2, 4, 5) Cribroperidinium tenuitabulatum (Gerlach) Helenes, 58.0 m depth; (3, 6) Charlesdowniea coleothrypta (Williams et Downie) Lentin et Vozzhennikova, 59.3 m depth; (7, 8) Pentadinium laticinctum Gerlach, 59.3 m depth; (9, 10) Dracodinium samlandicum (Eisenack) Costa et Downie, 58.0 m depth; (11–13) Piladinium columna (Michoux) Williams et al., 58.0 m depth.
All samples from Bed 1, attributed to the Upper Cretaceous Boguchar Formation, contain quantitatively rich palynomorph assemblages, which, however, contain mostly poorly preserved dinocysts, single fern spores and angiosperm pollen, and single prasinophytes (Palambages morulosa), acritarchs (Paralecaniella indentata, Michystridium sp.), and foraminiferal linings.
Unfortunately, only 22 of 47 palynological samples from the Paleogene part of the sequence revealed quantitatively rich palynological assemblages; the rest of them either contained a small number or were completely barren of palynomorphs. The Paleocene–lower Eocene palynological assemblages are characterized by the dominance of dinoflagellate cysts (30–90%), common proportion (up to ~30%) of angiosperm pollen and spores, and a single peak in the abundance of foraminiferal linings. Only at the base of the middle part of the section, attributed to the Oligocene Ventsy Beds, the assemblage is marked by the dominance (~40%) of the bisaccate pollen of conifers.
Dinocyst assemblage from the Boguchar Formation is characterized by the predominance of Trythirodinium suspectum and Trythirodinium cf. rhomboideum as well as by the taxonomic diversity and common occurrence of the genera Chatangiella (Ch. vnigrii, Ch. bondarenkoi, Ch. cassidea, Ch. victoriense), Isabelidinium (I. acuminatum, I. belfastense, I. microarmum, I. magnum), and Spiniferites. The assemblage also contains the species Acantaulax wilsonii, Apteodinium deflandrei, Connexmura fimbriata, Circulodinium distinctum, Alisogymnium euclaense, Dinogymnium acuminatum, D. westralinum, Raetiadinium truncigerum, Exochosphaeridium bifidum, Manumiella cretacea, Odontochitina costata, Laciniadinium rhombiforme, L. arcticum, Lanternosphaeridium lanosum, Spinidinium echinoideum, S. uncinatum, Fromea chytra, Xenascus gochtii, and Form 2 sensu Lebedeva. In general, this dinocyst assemblage is taxonomically close to the Campanian assemblages of the Polar Cis-Urals, Ust-Yenisey region, and the Omsk Trough in Western Siberia (Lebedeva, 2005, 2006, 2007; Lebedeva et al., 2013). Taking into account the presence of species Odontochitina costata, Chatangiella vnigrii, Manumiella cretacea, Lanternosphaeridium lanosum, and Form 2 sensu Lebedeva as well as the absence of stratigraphically important species Surculosphaeridium? longifurcatum, this assemblage can be preliminarily compared with one of the Layers with Odontochitina costata established in the Lower Volga region, in the upper part of the Mezino–Lapshinovka Formation and in the lower part of the Pudovkino Formation of Santonian to Campanian transitional age (Aleksandrova et al., 2012). It should be noted, however, that the common occurrence of the genus Trythirodinium in the Lower Volga region was established stratigraphically below, in the upper Santonian part of the Layers with Surculosphaeridium? longifurcatum–Heterosphaeridium difficile. Consequently, sediments in the interval of 100–91.7 m depth in borehole no. 1238 are preliminarily dated as late Santonian–early Campanian.
In terms of abundance, the dinocyst assemblage from the Boguchar Formation is characterized by the prevalence of the peridinioid genera Trythirodinium, Chatangiella, and Isabelidinium, which may indicate accumulation in littoral environments with a significant nutrient supply.
A dinocyst assemblage including the stratigraphically important species Deflandrea oebisfeldensis, Deflandrea denticulata, and Alisocysta sp. 2 sensu Heilmann-Clausen, as well as Areoligera coronata, Areoligera senonensis, Palaeotetradinium minusculum, Trigonopyxidia ginella, Tanyosphaeridium xanthiopyxides, Hystrichosphaeridium tubuferum, Cerodinium cf. medcalfii, Cerodinium speciosum, etc., was recognized in the lower part of the Paleogene section, which was attributed to the Buzinovka Formation (91.7–85.5 m). The presence of Deflandrea denticulata allows to attribute the sediments of the Buzinovka Formation to the interval of the Alisocysta margarita dinocyst Zone from zonations of Western Siberia (Iakovleva and Aleksandrova, 2013) and Western Europe (Powell, 1992) and to the interval of Layers with Alisocysta sp. 2 of the Volga region (Aleksandrova, 2013), and, consequently, to date them by early Thanetian (~58.5–56.5 Ma).
Palynological assemblage from the Buzinovka Formation is characterized by the dominance of dinoflagellate cysts (30–70%); the abundance of acritarchs and green algae does not exceed 5%; angiosperm pollen varies from 10 to 30%; and spores do not exceed 10%. The dinoflagellate cyst assemblage, in its turn, is marked by the absolute dominance of the gonyaulacoid groups, especially Areoligera (55–5%), Membranosphaera (45–0%), Spiniferites (10–35%), Fromea (up to 20%), and Cordosphaeridium (up to 15%); Impagidinium was encountered as single finds; the peridinioid dinocysts, represented by the group of deflandroids, account for not more than 15% of the assemblage.
The second dinocyst assemblage was recognized in the lower part of the Veshenskaya Formation (81.7–76.4 m depth) by the occurrence of species Apectodinium homomorphum and Apectodinium quinquelatum and is attributed to the interval of the late Thanetian Apectodinium hyperacanthum dinocyst Zone from zonations of the Volga region (Aleksandrova, 2013), Western Siberia (Iakovleva and Aleksandrova, 2013), and Western Europe (Powell, 1992). Species Phthanoperidinium crenulatum, Tectatodinium pellitum, Areoligera medusettiformis, Carpatella rossica, Hystrichokolpoma rigaudiae, Melitasphaeridium pseudorecurvatum, etc., appear within the assemblage. The stratigraphic age of the lower Veshenskaya Formation is late Thanetian (~56.5–55.8 Ma).
Palynological assemblage of the lower part of the Veshenskaya Formation is marked by substantial dominance of dinoflagellate cysts (35–80%) with up to 10% acritarchs, up to 30% of land plant spores, 7–20% angiosperm pollen, and up to 12% bisaccate pollen of gymnosperms. Dinocyst assemblage in this interval is characterized by the dominance of the peridinioid group of deflandroids (up to 55%) and peaks in the abundance of the gonyaulacoid species Carpatella rossica (up to 40%); the Areoligera- and Spiniferites-groups do not exceed 15% each; Impagidinium represents few percent. The dominance of the deflandroid cysts and the noticeable abundance of continental palynomorphs may suggest that the lower part of the Veshenskaya Formation accumulated in marine shallow water environments with reduced salinity and nutrient supply from the flooded land. The proportions of dinocyst groups by the end of the interval (relative decrease in deflandroids and a slight increase in Areoligera-, Cordosphaeridium-, and Spiniferites-groups) indicate a gradual normalization of water salinity.
The third dinocyst assemblage, corresponding to the Axiodinium augustum Zone of the eastern Peri-Tethys (Iakovleva, 2017a), was recognized by the appearance of Axiodinium augustum in the upper part of the Veshenskaya Formation (73.6–70.1 m depth). The dinocyst assemblage in this interval is quite diversified: it contains, for the first time in this section, species Apectodinium summissum, Adnatosphaeridium robustum, Caligodinium amiculum, Dapsilidinium pseudocolligerum, Hystrichokolpoma granulatum, Impagidinium dispertitum, etc. The interval between the first and last occurrences of Axiodinium augustum characterizes the Apectodinium (=Axiodinium) augustum Zone in Western Siberia (Iakovleva and Aleksandrova, 2013) and Southern England (Powell et al., 1996), Subzone D5A in Northwestern Europe (Costa and Manum, 1988), Viborg-6 Zone in Denmark (Heilmann-Clausen, 1988), and Subzone P6b in the North Sea (Mudge and Bujak, 1994).
It is widely known that the base of the Apectodinium (=Axiodinium) augustum Zone in many regions of the world coincides with the beginning of a carbon isotope excursion (CIE) event, associated with a short-term (~200 000 years) geological event, the Paleocene–Eocene Thermal Maximum (PETM), whereas the upper boundary of this zone is just slightly younger than the end of CIE and, consequently, slightly younger than the PETM event (Frieling et al., 2014; Shcherbinina et al., 2016). Considering that the greater part of the interval of Ax. augustum Zone corresponds to the PETM event (~55.8–55.6 Ma) and the associated global CIE event, we attribute the upper part of the Veshenskaya Formation to earliest Ypresian (~55.8– 55.6 Ma).
Palynological assemblage from this interval is characterized by the dominance of dinoflagellate cysts (50–70%) and a relative increase in acritarchs (up to 25%). Angiosperms are 25–10% of the assemblage, whereas the spores of land plants and the bisaccate pollen of conifers do not exceed 5–10% each. Dinoflagellate cyst assemblage is dominated by the gonyaulacoid groups Spiniferites (~35–45%), Areoligera (10–25%), and Operculodinium (up to 10%). Peridinioid deflandroids do not exceed 15%, and Apectodinium spp. are represented by few percent. Such ratios of marine and continental palynomorphs as well as the dominance of Spiniferites and Areoligera in dinocyst associations indicate open-marine environments with normal water salinity and the transgressive stage of basin evolution.
The borehole section interval of 67.4–59.7 m depth, attributed to the Surovikino Beds, is almost barren of palynomorphs. Only single specimens of Areoligera coronata, Corodosphaeridium gracile, Spiniferites ramosus and Hystrichosphaeridium tubiferum with a wide stratigraphic distribution were noted here.
The fourth dinocyst assemblage was recognized in the Osinovaya Beds in the interval of 59.3–57.0 m depth. Based on the presence of species Ochetodinium romanum, Piladinium columna, and Samlandia chlamydophora, this taxonomically diverse (more than 85 taxa) and quantitatively rich assemblage corresponds to the Ochetodinium romanum/Samlandia chlamydophora dinocyst zone from the eastern Peri-Tethys Zonation (Iakovleva, 2017). Species Samlandia chriskingii, Areoligera sentosa-group, Heteralaucacysta everriculata, Homotryblium tasmaniense, Wetzeliella articulata, Hystrichosphaeropsis obscura, Membranilarnacia glabra, Thalassiphora gracilis, Turbiosphaera galatea, etc., were also noted in the interval. The stratigraphic age of this part of the Osinovaya Beds is mid Ypresian (~51.3–50.8 Ma).
Palynological assemblage in the interval of 59.3–57.0 m depth is almost entirely represented by dinoflagellate cysts (more than 95%), whereas the foraminiferal linings and the angiosperms account for one or two percent. Noticeable changes in acritarch abundance (~24%) were encountered only at the top of the Osinovaya Beds (57.0 m depth). Dinocyst assemblage is primarily characterized by significantly varying percentages of the Cordosphaeridium-group, from dominant (20–71%) to insignificant (4–6%); the content of peridinioid dinocysts (deflandroids and wetzelielloids) here is up to 19% and 20%, respectively; the content of the Spiniferites-group varies from 6 to 19%, and it is almost absent at the top of the succession (57.0 m depth). The genus Impagidinium (Impagidinium wardii) attains up to 5% of the assemblage. Such characteristics of palynological assemblage may indicate once again a mid Ypresian transgressive cycle in basin evolution history and the accumulation of the Osinovaya Beds in shallow-water marine environments with normal water salinity, but considering the notable abundance of the peridinioids in the dinocyst assemblage, we may suggest nutrient supply from a flooded land.
The fifth dinocyst assemblage was recognized in the Ventsy Beds in the interval of 56.5–54.0 m, and we ascribed it to the Layers with Rhombodinium draco and Deflandrea spinulosa. It is characterized by the presence of Rhombodinium draco, Wetzeliella ovalis, Glaphyrocysta semitecta, Areosphaeridium diktyoplokum, Charlesdowniea? rotundata, and Phthanoperidinium geminatum, the first co-occurrence of which in the eastern Peri-Tethys is observed at the end of Lutetian and in Bartonian in the interval of the Rhombodinium draco dinocyst Zone (Iakovleva, 2017). However, the comparison of this assemblage with the middle and upper Eocene dinocyst assemblages established in the sections in Northern Ukraine, Voronezh Anteclise, and the Scythian Plate indicates substantial differences between them (Zaporozhets, 1998, 2001; Oreshkina and Iakovleva, 2007; Andreeva-Grigorovich and Shevchenko, 2007; Andreeva-Grigorovich et al., 2011; Aleksandrova et al., 2011; Vasil’eva, 2013, 2019; Iakovleva, 2015). Thus, the predominance of Rhombodinium draco and Deflandrea spinulosa and common Thalassiphora pelagica and Wetzeliella articulata allow us to correlate the dinocyst assemblage from the Ventsy Beds in borehole no. 1238 with the Rhombodinium draco Subzone of the Chiropteridium partispinatum Zone, established earlier in the interval of transition from the Solenaya to the Kalmykia Formation in borehole 45A in the Yergeni (130–94 m; Zaporozhets, 1998) and in the upper Morozkina Balka Fm–lower Batalpashinsk Fm in the Belaya River section in the North Caucasus (Zaporozhets and Akhmet’ev, 2017), where sediments are dated by late Rupelian–early Chattian (Zaporozhets, 1998; Zaporozhets and Akhmet’ev, 2017; Zastrozhnov et al., 2019). It should be noted that Zaporozhets indicates the abundance of reworked Paleogene and even Mesozoic dinocysts within the dinocyst assemblage corresponding to the Rhombodinium draco Subzone in different parts of the Peri-Tethys. It is quite possible that a part of taxa in the dinocyst assemblage from the Ventsy Beds can also be reworked. The recognized dinocyst assemblage from the Ventsy Beds is also close to one from the transitional Rupelian–Chattian interval in the Danish Basin (Śliwinska et al., 2012).
The most part of the “Poltava” Beds, represented by sands with secondary clay layers, contains rare palynomorphs; only the palynological assemblage from the depth of 54.0 m (siltstone) was quantitatively rich. A significant change in the dominant palynomorph groups (in comparison to the underlying sediments) was noted here: dominance of the bisaccate pollen of conifers (46%); approximately equal quantitative content of angiosperms (26%) and dinoflagellate cysts (23%); and a very small content (4%) of acritarchs. Dinocyst assemblage, in its turn, is characterized by the absolute predominance of peridinioids—deflandroids (38%), wetzelielloids (33%), and Phthanoperidinium (5%); the gonyaulacoids occur in small numbers. Such ratios of palynomorph groups suggest the accumulation of the lower part of the “Poltava” Beds in littoral environments with reduced water salinity and substantial nutrient supply.
DISCUSSION
Originally Leonov (1936, 1939) subdivided the Paleogene deposits of the Middle Don River Basin into five formations with numerical designation (I–V, Fig. 2). Later, he replaced these formations by secondary local lithostratigraphic units (beds) or distinguished the beds within his formations (Leonov, 1961). Thus, Formation I became the Buzinovka Beds, corresponding to the Proleika Formation in the Lower Volga region; Formation II was subdivided into the Veshenskaya and Surovikino Beds, corresponding to the subformations of the Tsaritsyn Formation of the Lower Volga region; Formation III was subdivided into the Osinovaya and Chir Beds, compared with the Stalingrad Formation (Mechetka and Yelshanka Beds, respectively) in the Lower Volga region. Finally, Formation IV was subdivided into the Kiev and Kharkov Beds from the unified Paleogene chronostratigraphic scheme of Northern Ukraine, which Leonov correlated with the Aksai Beds and the lower part of the Balykley Beds in the Lower Volga region. Higher in the succession, Leonov recognized the Luchin Beds, which he correlated yet again with the upper part of the Balykley Beds in the Lower Volga region. The uppermost part of the Paleogene sequence, Formation V, was assigned to the Ventsy Beds, compared with the Maykop Formation in the Lower Volga region, and the so-called “Poltava” Beds, corresponding, with a bit of convention, to the Poltava Group of Ukraine.
For the subdivision of the Paleocene and lower Eocene, Semenov (1965) adopted most of the secondary local lithostratigraphic units introduced earlier by Leonov (1961), such as the Buzinovka, Veshenskaya, and Surovikino Beds; but, in addition, this author upgraded them into formations (Fig. 2). Semenov also introduced two new formations, Kumovka (the lowermost Paleogene) and Sheptukhovka. Semenov unified the Kumovka, Buzinovka, Veshenskaya, and Surovikino formations into the Don Group (lower Paleocene–lowermost Eocene). In turn, the Sheptukhovka Formation (upper part of the lower Eocene), introduced by Semenov, and the middle Eocene Khripun and Osinovaya formations were unified into the Boguchar Group. As regards the upper Eocene in the Middle Don region, Semenov subdivided it into the Sergeyevka, Tishki, and Kasyanovka formations, while he subdivided the Oligocene deposits into the Kantemirovka and Zhuravka formations. The age of all these formations was based mostly on comparison of the malacofauna assemblages with those from the Dnieper region (Ukraine). Later, these formations were given in rank of regional horizons in the chronostratigraphic scheme of 1975 of the Voronezh Anteclise (Semenov, 1975). It should be noted that the formations established by Semenov are largely identical in lithology and faunistic composition and have the most typical features on the territory to the west from the Cis-Donets Monocline, which, beginning from the approval of Unified Scheme of 2000 is already attributed to the SFZ of the Voronezh Anteclise, while the Buzinovka and Veshenskaya horizons from Semenov’ scheme of 1975 are reserved for the eastern Greater Donbass subregion.
The upper Santonian–lower Campanian deposits from the studied borehole no. 1238 section are overlain by the upper Paleocene (lower Thanetian) marine deposits of the Buzinovka Formation. Based on this, we suggest an important (~18 Ma) hiatus, corresponding to the upper Campanian–Selandian. The absence of the Danian–Selandian deposits in the borehole 1238 area more likely indicates the uplifted position of this part of the Russian Platform during the early Paleocene.
The Buzinovka Formation within the Cis-Donets Monocline is represented by a succession (8–16 m) of fine greenish-gray glauconite–quartz sands with discontinuous siliceous sandstone interbeds; a horizon of non-rounded phosphorite nodules is present in the middle of the sequence, and a second relatively thin phosphoritic horizon is sometimes noted below (Leonov, 1961). This formation with its typical features is restricted to the Cis-Donets Monocline and is not recognized west of the Tikhaya River mouth or in the Voronezh Anteclise.
The exploration of the surroundings of the Verkhnyaya Buzinovka village, which is a stratotype region for the Buzinovka Beds (Leonov, 1961), showed that there is only one natural Paleogene outcrop, where only the upper part of the Buzinovka Formation with an apparent thickness of 6.2 m and its contact with the overlying Veshenskaya Formation are observed (Figs. 7a, 7b). The Buzinovka Formation is represented by a member of fine-grained quartz–glauconite sandstones with a phosphorite horizon (0.5 m) in the lower part. A bioturbation in the form of silicified mud eater burrows (up to 1.5 cm in diameter) is noted at the top of formation. The Veshenskaya Formation overlies with the irregular transgressive boundary. Its basal part is composed of dark gray massive compact opoka, which makes up a distinct edge in the wall.

Photographs of the outcrops of the Buzinovka Formation from the Cis-Donets Monocline: (a) outcrop of the upper Buzinovka Formation in the environs of the Verkhnyaya Buzinovka village; (b) the upper Buzinovka Formation exposed by stripping in the environs of the Verkhnyaya Buzinovka village; (c) outcrop of the lower Buzinovka Formation in the environs of the Biryukovsky village; (d) phosphoritic horizon and the contact of the Buzinovka and Veshenskaya formations in the vicinity of the Biryukovsky village.
One of the few most complete outcrops of the lower Buzinovka Formation is situated farther west in the vicinity of the Biryukovsky village (Figs. 7c, 7d). This section demonstrates distinctly visible rhythmically cyclic structure of the lower part of the Buzinovka Fm: it is represented by quartz–glauconite greenish-gray inequigranular, predominantly medium- and fine-grained sands with occasional quartz gravel inclusions, transgressively overlying the chalk. The base of the formation is abrupt; the basal level (8–10 cm) is composed of dark clay with small phosphorites, rounded flatted pebbles, and reworked belemnites. The sands contain gray slightly micaceous hard sandstone interbeds. The apparent top of the formation is represented by a member of nodular phosphorites with numerous quartz and black siltstone inclusions (up to 10 mm in diameter), which grades upward into a stratiform phosphorite slab, containing mollusk shell fragments. The thickness of the Buzinovka Formation is 5 m. Based on the assumption of Leonov that phosphoritic levels occur in the middle part of the Buzinovka Fm, we suggest the erosion of the upper part of formation in the section near the Biryukovsky village. The Buzinovka Fm is transgressively, with traces of erosion such as abundant quartz gravel and sand pockets, overlapped by the Veshenskaya Formation. The Buzinovka Fm has the similar features in the vicinity of the Kumovka village, where, however, it is poorly exposed.
The Buzinovka Horizon in the Scheme of 2000 contains a mollusk assemblage with Lyostrea reussi, benthic foraminifers of the BF3 Zone, and the SP3 spore and pollen assemblage and corresponds to the lower Thanetian (Akhmetiev and Beniamovsky, 2003), which is now confirmed by our dinocyst data. Based on the new palynological data indicating that the Buzinovka Formation corresponds to the interval of the early Thanetian Alisocysta margarita dinocyst Zone, this formation can be correlated with the Saratov Formation of the Ulyanovsk–Saratov Trough and the Precaspian Syneclise and with the lower Thanetian Novouzenskaya Formation in the Precaspian region (Unifitsirovannaya…, 2015). According to the palynological results from the borehole no. 1238, the Buzinovka Formation was accumulated during a transgressive period, in the inner neritic zone with normal salinity.
The Veshenskaya Formation with a thickness of up to 15 m, overlying the Buzinovka Fm, is composed of compact opoka-like or tripolith-like sandy clays, often facially replaced by loose clayey, sometimes opoka-like sandstones or compact clayey sands (Leonov, 1961). At the south ending of the Don–Medveditsa Arch, the clays turn into variegated quartz–glauconite sands interbedded with analogous sandstones. This formation transgressively overlaps different levels of the underlying deposits, and its base is characterized by the appearance of gravel and pebble. The Veshenskaya Fm is more widespread than the Buzinovka Formation: to Boguchar town in the west and to Novokhopersk in the north. In the Scheme of 2000, the Veshenskaya Horizon contains a mollusk assemblage with Arctica (=Cyprina) morrisi, benthic foraminifers of the BF3 Zone, and the SP3 spore and pollen assemblage and corresponds to the upper Thanetian (Akhmetiev and Beniamovsky, 2003).
Outcrops of the Veshenskaya Formation are widespread in the Cis-Donets Monocline; we identified them in the environs of the Kumovka, Verkhnyaya Buzinovka, and Surovikino villages and the Biryukovsky and Kalininsky villages, as well as in the stratotype region near Bazkovskaya Stanitsa. All these outcrops represent only separate parts of this formation; unfortunately, we did not find any complete outcrops (Figs. 8a, 8b). In general, our observations confirm Leonov’s data: the Veshenskaya Formation was accumulated during the continued marine transgression in open-marine environments.

Photographs of the outcrops of the Veshenskaya Formation and the Surovikino and “Poltava” Beds from the Cis-Donets Monocline: (a) outcrops of the upper Veshenskaya Formation in the environs of the Kumovka village; (b) outcrops of the lower Veshenskaya Formation in the environs of Bazkovskaya village; (c) contact of the Veshenskaya Formation and the Surovikino Beds in the environs of the Surovikino village; (d) horizon with decapod burrows and holes in the Surovikino Beds in the environs of the Surovikino village; (e) bioturbation in the “Poltava” Beds in the environs of Bazkovskaya village; (f) low-angle cross-bedding of the “Poltava” Beds in the environs of Bazkovskaya village.
Based on the new palynological data, a dinocyst assemblage corresponding to the interval of the late Thanetian Apectodinium hyperacanthum Zone was recognized in the lower Veshenskaya Formation in the borehole no. 1238 (81.7–76.4 m). This does confirm the late Thanetian age of sediments, but only for the lower part of the formation. Based on the dinoflagellate cysts, the lower part of the Veshenskaya Formation is correlated with the upper Thanetian Kamyshin Formation of the Ulyanovsk–Saratov Trough (Aleksandrova, 2013; Unifitsirovannaya…, 2015) and the Nalchik Formation of Northern Caucasus (Shcherbinina et al., 2016). As regards the upper part of the formation, drilled by the borehole no. 1238, it corresponds to the interval of the initial Eocene Axiodinium (=Apectodinium) augustum dinocyst Zone. This part of the Veshenskaya Formation can be correlated in Peri-Tethys with the coeval sediments of the Nalchik Formation in the North Caucasus (Shcherbinina et al., 2016), the Bostandyk Formation in the Northern and Central Precaspian region (Vasilyeva and Musatov, 2010а; Vasilyeva and Musatov, 2012), the Proleika Formation in Volgograd region (Aleksandrova et al., 2011), the sandy-clayey formation in Eastern Crimea (Aleksandrova and Shcherbinina, 2011), and the Kalinin Formation from the Lower Volga region (Dyupa Gulch; Aleksandrova and Radionova, 2006). Moreover, according to Oreshkina et al. (unpublished data), a borehole drilled in the extreme west of the Cis-Donets Monocline (close to Boguchar town, Voronezh region) (Fig. 1) uncovered a stratigraphically younger part of the Veshenskaya Formation corresponding to the interval of the early Ypresian Stenodinium meckelfeldense dinocyst Zone (~54.8–53.4 Ma). Consequently, based on dinocyst data, the general age of the Veshenskaya Formation can be estimated as late Thanetian–part of the lower Ypresian (~56.5–53.4 Ma).
The Surovikino Beds, with a thickness of 8–10 m, overlying the Veshenskaya Fm in the studied region, are represented by light-gray and yellowish medium to fine powdery quartz sands, slightly glauconitic in the lower horizons, with discontinuous compact siliceous sandstone interbeds and lenses (Leonov, 1961). Starting from the 1950s, the Surovikino Beds were attributed to the lower Eocene and correlated with the lower parts of the Bakhchisaray Formation in Crimea, the Kanev Formation in Ukraine, and the Upper Tsaritsyn Beds in the Volgograd sector of the Lower Volga region (Leonov, 1939, 1961; Semenov, 1965, 1975; Geologiya…, 1969; Stratigraficheskaya…, 1987).
During the recent fieldwork the complete outcrops of the Surovikino Beds were not found; only their contact with the Veshenskaya Formation (Fig. 8c) was observed in the outcrop near the Surovikino village, while an isolated outcrop of, most probably, the middle part of the Surovikino Beds was discovered near the village of Kumovka. It should be noted that the Surovikino outcrop section revealed a level with numerous decapod burrows with a well pronounced tubercular texture, very close to the Ophiomorpha nodosa ichnospecies from the lower Ypresian Kalinin Formation in the Lower Volga region (Yanin and Beniamovsky, 2018) (Fig. 8d). Lithology of the Surovikino Beds suggests that they were deposited in a shallow-water marine paleoenvironment.
Unfortunately, the Surovikino Beds in borehole 1238 revealed only few dinocysts, which do not permit to precise the age of sediments. In the borehole section, they are situated between the upper Thanetian–lower Ypresian Veshenskaya Formation and the Osinovaya Beds with the mid Ypresian dinocyst assemblage of the Ochetodinium romanum/ Samlandia chlamydophora Zone, which confirms the early Ypresian age of the Surovikino Beds.
The overlying Osinovaya Beds rest transgressively in the Cis-Donets Monocline, which is confirmed by our palynological data. The base of these beds in the studied region is characterized by the presence of coarse sediments and one or two siliceous sandstone interbeds. These deposits change upward by brownish clays or fine sands (4–5 m), which, in turn, are overlain by variegated greenish-gray compact fine quartz–glauconite sands up to 8–10 m in thickness, replacing next by medium quartz sands (Leonov, 1961). The Osinovaya Beds are not characterized paleontologically.
It should be noted that during the fieldwork the natural outcrops of the Osinovaya Beds in the Cis-Donets SFZ were not found. Based on the lithological characteristics of these beds, only their lower part in the interval of 60.5-57.0 m depth was recognized in the borehole no. 1238, whereas their upper horizons are probably eroded. Our palynological data from borehole 1238 (60.5–57.4 m; Ochetodinium romanum/ Samlandia chlamydophora Zone) imply the mid Ypresian age of the Osinovaya Beds.
It should be recalled that, from the middle of last century (Semenov, 1965; Geologiya…, 1969; Stratigrafiya…, 1975) to the present day (Gosudarstvennaya…, 2009), it is generally accepted that the Osinovaya Beds (=Osinovaya Formation) rest erosively (as indicated by a pebble or conglomerate layer) on the Surovikino Beds, correspond to the lower part of the Buchak Horizon, and are therefore attributed to the middle Eocene (Lutetian). It must be emphasized that Semenov took as the stratotype of the Osinovaya Formation the outcrops near the village of Osinovaya on the Aidar River in Lugansk region of Ukraine (Fig. 1), where, according to Leonov (1961), the Osinovaya Beds condense and become coarser, whereas the overlying Chir Beds are either absent because of erosion or poorly exposed. As a result, the recognition of these lithostratigraphic units in the Aidar River basin was conventional and required confirmation by additional data. In the type section in the Lipovskii gully (the right bank of the Don River, in the vicinity of the Chir River mouth), Leonov correlated the Osinovaya Beds to the lower part of the Stalingrad Formation, the Mechetka Beds of the Lower Volga region, and with the Cherkessk Formation of the Rostov Arch, the Cis-Caucasian part of the Scythian Platform, and, to some degree, with the lowermost Buchak Stage of Ukraine (Leonov, 1961), which was recorded in the Regional Scheme of the Paleogene Deposits (Geologiya…, 1969).
According to recent data, the Cherkessk Formation in the North Caucasian Kheu River reference section is Ypresian (early Eocene); within this stratigraphic interval, among others, the mid-Ypresian Ochetodinium romanum/Samlandia chlamydophora dinocyst zone was recognized (Iakovleva, 2017; Shcherbinina et al., 2020). Consequently, the Osinovaya Beds correspond indeed to a part of the Cherkessk Formation of North Caucasus and a part of the Bostandyk Formation in the Precaspian region (Vasilyeva and Musatov, 2010a, 2010b, 2012; Vasilyeva, 2013; Musatov et al., 2014), the Simferopol Formation in Crimea (King et al., 2018), of the Kulsary Formation in Northern Ustyurt (Iakovleva and Heilmann-Clausen, 2020), to a part of the Lower Sal subhorizon of the Scheme of 2000 (Akhmetiev and Beniamovsky, 2003), and to the upper part of the Proleika Horizon of the Lower Volga region (Unifitsirovannaya…, 2015) (Fig. 2).
Long-term discussions are related to the age and genesis of the sandstone member in the upper part of the Paleogene in southern Russia. Most researchers tend to correlate it with the so-called Poltava Stage of the scheme proposed by Sokolov (1893) and to date it by Oligocene–Early Miocene. Due to the absence of fossils in this member, the Oligocene–early Miocene age was determined by comparison with the Paleogene lithostratigraphic units of the Dnieper River Basin, Scythian Platform, or the Lower Volga region (Leonov, 1936, 1939, 1961; Semenov, 1965, 1975; Semenov and Stolyarov, 1988; Stratigraficheskaya…, 1987; Voronina et al., 1988; Popov et al., 1993; Gorbatkina and Iosifova, 2004; etc.).
These sediments with a thickness of up to 30 m in the Cis-Donets Monocline rest transgressively and erosively on different Cretaceous and Paleogene horizons. The reliably marine Oligocene sediments, attributed to the Maykop Group, spread over the boundary between the structural-facies zones of the Cis-Donets Monocline and the eastern surroundings of the Donbass, in the environs of the Tsimlyansk hydroelectric complex. Based on the geographic location and lithologic characteristics, the facies transitional from the Maykop to the Poltava sediments, established in the lower reaches of the Chir River, were assigned to the Ventsy Beds (Leonov, 1961). They are composed of dark (gray to black) humus-rich clays with numerous thin lenticular fine-grained quartz sand interbeds with a total thickness of up to 4 m.
Farther north are continental sediments classified as the “Poltava Group” (Leonov, 1961; Semenov, 1965), which is represented by inequigranular quartz sands, varicolored (white, yellow, red, crimson, gray, greenish), loose, sometimes slightly clayey, compacted. Thin varicolored clay interbeds and lenses (1–4 m) are also reported here, predominantly in the lower part, as well as ferruginous sandstone nodules. It was assumed that the Donbass and the Don–Medveditsa Arch were the line of facial boundary between the Maykop and Poltava sediments.
Quantitatively rich palynological data from the Ventsy Beds in borehole no. 1238 (57.0–54.0 m depth), where the Layers with Rhombodinium draco–Deflandea spinulosa of the late Rupelian–early Chattian are now recognized, finally bring some certainty on the stratigraphic position of the lower boundary of the upper Paleogene horizons in the Cis-Donets Monocline and on the problems of paleogeographic reconstructions of the depositional environments. We found that the Ventsy Beds in borehole no. 1238 rest erosively on the Lower Eocene Osinovaya Beds, which implies an important hiatus corresponding to the upper Ypresian and most of Rupelian (~18 Ma). Unfortunately, the overlying “Poltava” Beds, including those that constitute the uplifted parts of the Cis-Donets Monocline, do not contain fossil palynomorphs in borehole no. 1238, and no other fossils in the outcrops were found during the fieldwork. As a result, the age of the upper boundary of the “Poltava” Beds still cannot be determined exactly.
Leonov (1961) interpreted the Ventsy Beds as nearshore marine or even lagoonal deposits. The diversified dinocyst assemblage confirms the marine genesis of the Ventsy Beds, while their regular underlying of typical Poltava facies suggests that the Ventsy Beds are the base of the upper sedimentary succession.
During our fieldwork we also examined the “Poltava” Beds in natural outcrops and opencast mines, where they are represented by thick members of varicolored quartz sandstones with low-angle cross bedding, clay levels, and extensive bioturbation (Figs. 8e, 8f). On the basis of which a conclusion was made about their shallow-marine genesis.
Taking into account the data of Goretsky (1982), the Cis-Donets Monocline area was also the discharge zone of a large paleoriver, which he named the “Solenaya River”; its flow was directed from north to south, and the alluvium was composed mostly by sands, which perhaps explains one of the sources of terrigenous material supply (Fig. 9).

Paleogeographic map of paleobasins during the Chattian (modified after Popov et al., 2009).
According to the lithological and paleontological data, the marginal marine depositional environments of the Poltava Group are also presumed for the western regions of the southern Russian Platform up to southern Belarus and northern Ukraine (Savko et al., 1995, 2012; Zinova, 2001; Sklyarenko, 2010; Zosimovich and Shevchenko, 2014, 2015; Boiko and Sklyarenko, 2014; etc.). It is assumed that during this time there was a latitudinal marine strait, which connected the basin of the Cis-Donets Monocline (as a part of the Paratethys) to the West European basins (Fig. 9). All that said and based on our evidences of development of marine facies up to the westernmost regions of the Cis-Donets Monocline, it is obvious that the Oligocene marine transgression extended much farther north than it was anticipated earlier (Leonov, 1961; Voronina et al., 1988; Popov et al., 1993, 2009; Semenov and Stolyarov, 1988; Stolyarov, 1991; Stolyarov and Ivleva, 2004a, 2004b).
Palynological data on the dynamics of changes in dinocyst, prasinophyte, and spore and pollen assemblages during the Solenaya and Kalmykia time in the adjacent regions of the Lower Don, Lower Volga, Yergeni, and North Caucasus (Zaklinskaya, 1953; Ananova, 1974; Prakticheskaya…, 1990; Zaporozhets, 1998; Akhmetiev and Zaporozhets, 2014; Zaporozhets and Akhmetiev, 2015, 2017; Zastrozhnov et al., 2019) permit to reconstruct the stages of accumulation of the Ventsy and “Poltava” Beds. Thus, the first stage is characterized by an increase in abundance of the pollen of Taxodiaceae (Taxodium, Glyptostrobus, Sequoia, Metasequoia) and the thermophilic and hydrophilic plants (Juglans, Carpinus, Fagus, Tilia, Ulmus, Quercus, Rhus, Myricaceae, Acer, Fraxinus, Hammamelis, Ilex, Liquidambar, Nyssa, Nelumbo) during the Rupelian, which was due to the development of a major and led to significant formation of marshes on the adjacent lands and climate warming, while the peak of warming corresponds to the early Morozkina Balka time in the North Caucasus and the early Solenaya time in the Yergeni. This time interval corresponds to the formation of thin, predominantly argillaceous facies of the uppermost Khadum Superhorizon in the North Caucasus. Marine phytoplankton assemblages reflect the unstable probably brackish depositional environments with restricted water exchange.
The second stage, corresponding to the late Morozkina Balka (or late Solenaya) time, is marked by free communication between the Paratethys and the North European basins in a still warm climate, as evidenced by the first taxonomically diversified dinocyst associations, mollusk finds, and the composition of spore and pollen assemblages (Zastrozhnov et al., 2019).
The third stage corresponds to the end of the late Solenaya and the beginning of the early Kalmykia time. According to spore–pollen data, this stage is characterized by the cooling of the climate, still remaining highly humid, while the peridinioid genera Rhombodinium and Deflandrea started to dominate in dinocyst assemblage. The beginning of the third stage is also marked by the maximum abundance of reworked palynomorphs, whereas the palynological associations are marked by abundant Pinaceae (Pinus, Picea, Tsuga, Abies) and the predominance of Betulaceae (Alnus, Betula, Corylus) and Salicaceae among the deciduous angiosperms, which indicates climate humidification, increase in denudation, and the uplift of the surrounding land. The combination of these factors led to the increase in river discharges into the marine basin and to the beginning of the accumulation of thick sandstones of the Kalmykia Formation along the northern periphery of the Paratethys (Stolyarov, 1991; Stolyarov and Ivleva, 2004a, 2004b). The late Kalmykia time is characterized by a dinocyst assemblage dominated by the genus Deflandrea and the depletion of the benthic molluscan fauna and foraminifers, which in general again reflects the environments of a restricted marine basin.
We believe that the pollen assemblage dominated by Pinaceae in conjunction with common Rhombodinium and Deflandrea of the Ventsy Beds can be correlated with those of the late Solenaya time, while the overlying “Poltava” Beds could accumulate during the early Kalmykia time. This allows correlating the Ventsy Beds with the upper Solenaya Subformation and, in turn, the “Poltava” Beds with the Kalmykia Formation of the Yergeni and the Lower Volga region.
Based on lithological characteristics and stratigraphic position of the Ventsy and Poltava Beds in borehole no. 1238, these units can be correlated with the Bereka Formation of the Voronezh Anteclise (Gorbatkina and Iosifova, 2004) and with the Zmiev and Sivash formations from Northern Ukraine (Stratigraficheskaya…, 1987; Zosimovich and Shevchenko, 2014, 2015).
CONCLUSIONS
Based on the dinocyst data, the sediments at the bottom of the borehole no. 1238, drilled in the Cis-Donets Monocline, are dated as late Santonian–early Campanian. The overlying marine Paleogene deposits revealed the intervals of dinocyst zones from the chronostratigraphic schemes of the Volga region and the Eastern Peri-Tethys: Alisocysta margarita, Apectodinium hyperacanthum, Axiodinium augustum, and Ochetodinium romanum/Samlandia chlamydophora, as well as Layers with Rhombodinium draco–Deflandea spinulosa, which enabled us to precise the age of the sediments. Thus, our data confirm the early Thanetian age of the Buzinovka Horizon; the age of the Veshenskaya Horizon has been revised and corresponds to the late Paleocene–initial Eocene. The overlying Surovikino and Osinovaya beds correspond to a part of the lower–middle Ypresian. The upper part of the Osinovaya Beds and the Chir Beds are missing in this borehole, which requires the search of more complete and continuous sections. Lithological composition and stratigraphic age of these beds permit to correlate them with the Proleika Formation in the Lower Volga region and a part of the Kanev Formation in Northern Ukraine. The middle and upper Eocene and the most part of the lower Oligocene sediments are not established on this territory. The Ventsy and Poltava Beds are correlated with the upper Solenaya Subformation and the lower part of the Kalmykia Formation of the Yergeni Highland. Two important hiatuses have been recognized in the borehole no. 1238 section: first one corresponds to the upper Campanian–Selandian and the other one, to the upper Ypresian–lower Rupelian.
We demonstrated that the local lithostratigraphic units proposed by Leonov in 1961 are quite applicable for the subdivision of the Paleogene of the Cis-Donets Monocline. We have established that the sediment deposition within the Cis-Donets Monocline is timed to the period of a major late Paleocene–early Eocene marine transgression and to the time of the end of the early Oligocene–late Oligocene regression in conditions of restricted marine basin. The first phase is represented by the Buzinovka and Veshenskaya formations and the Surovikino and Osinovaya Beds, while the second one is represented by the Ventsy and “Poltava” beds.
REFERENCES
Akhmetiev, M.A. and Beniamovsky, V.N Stratigraphic chart of marine Paleogene on the South of European Russia, Byull. Mosk. O–va Ispyt. Prir., Otd. Geol., 2003, vol. 78, no. 5, pp. 40–51.
Akhmetiev, M.A. and Zaporozhets, N.I., Paleogene events in Central Eurasia: their role in the flora and vegetation cover evolution, migration of phytochore boundaries, and climate changes, Stratigr. Geol. Correl., 2014, vol. 22, no. 3, pp. 312–335.
Aleksandrova, G.N., Paleocene dinocysts from the Central and Lower Volga Region: Sratigraphy and paleoenvironments, Cand. (Geol.-Mineral.) Dissertation, Moscow: Geol. Inst. Ross. Akad. Nauk, 2013.
Aleksandrova, G.N. and Radionova, E.P., On the late Paleocene stratigraphy of the Saratov Volga Region: Micropaleontological characteristics of the Kamyshin Formation, Dyupa Gully section, Paleontol. J., 2006, vol. 40, no. 5, pp. 543–557.
Aleksandrova, G.N. and Shcherbinina, E.A., Stratigraphy and paleoenvironmental interpretation of the Paleocene–Eocene transition in the eastern Crimea, Stratigr. Geol. Correl., 2011, vol. 19, no. 4, pp. 424–449.
Aleksandrova, G.N., Beniamovsky, V.N., Zaporozhets, N.I., Zastrozhnov, A.S., Zastrozhnov, S.I., Tabachnikova, I.P., Oreshkina, T.V., and Zakrevskaya, E.Yu., Paleogene of the southwestern Volgograd Region (borehole 13, Gremyach’e area). 1. Biostratigraphy, Stratigr. Geol. Correl., 2011, vol. 19, no. 3, pp. 310–336.
Aleksandrova, G.N., Beniamovsky, V.N., Vishnevskaya, V.S., and Zastrozhnov, A.S., New data on the Upper Cretaceous biostratigraphy of the Lower Volga Region, Stratigr. Geol. Correl., 2012, vol. 20, no. 5, pp. 426–465.
Ananova, E.N., Pyl’tsa v neogenovykh otlozheniyakh yuga Russkoi ravniny (Pollen in the Neogene Sediments of the South of Russian Plain), Leningrad: Leningrad. Gos. Univ., 1974 [in Russian].
Andreeva-Grigorovich, A.S. and Shevchenko, T.V., Dinocyst zonation of the Paleogene deposits of Ukraine, in Paleontologichni doslidzhennya v Ukraini: Zb. nauk. prats' (Coll. Sci. Works “Paleontological Research in Ukraine”), Kyiv: Inst. Geol. Nauk Nat. Akad. Nauk Ukraini, 2007, pp. 211–214.
Andreeva-Grigorovich, A.S., Zaporozhets, N.I., Shevchenko, T.V., Aleksandrova, G.N., Vasil’eva, O.V., Iakovleva, A.I., Stotland, A.B., and Savitskaya, N.A., Atlas dinotsist paleogena Ukrainy, Rossii i sopredel’nykh stran (Atlas of Paleogene Dinocysts of Ukraine, Russia and Adjacent Countries), Kiev: Nauk. Dumka, 2011 [in Russian].
Beniamovsky, V.N., Proposals for the refinement of the Paleogene stratigraphic scheme of the Voronezh–Donets Region, in Materiali Soveshch. “Sostoyanie stratigraficheskoi bazy tsentra i yugo-vostoka Vostochno-Evropeiskoi platformy” (Moskva, VNIGNI, 23–25 noyabrya 2015 g.) (Proc. Conf. “State of the Stratigraphic Base of the Central and Southeastern Parts of the East European Platform” (Moscow, All-Russ. Res. Geol. Oil Inst., November 23–25, 2015)), Moscow: Vseross. Nauchno-Issled. Geol. Neft. Inst., 2016, pp. 122–128, 134–139.
Beniamovsky, V.N., Proposals for structural–facies zoning and subregional horizons of the Paleogene of the Voronezh–Donets Subregion, in PALEOSTRAT-2017. Godichnoe sobranie (nauchn. konf.) sekt. paleontol. MOIP i Moskovskogo otd. Paleontol. obshch. pri RAN. Moskva, 26-28 yanvarya 2017 g. Progr. tez. dokl. (Proc. Ann. Meet. (Sci. Conf.) Paleontol. Sect. Moscow Soc. Nat. Sci. and Moscow Branch Paleontol. Soc. “PALEOSTRAT-2017,” Moscow, January 26–28, 2017), Alekseev, A.S., Ed., Moscow: Paleontol. Inst. Ross. Akad. Nauk, 2017a, pp. 15–16.
Beniamovsky, V.N., Paleontological and paleogeographic justification of Paleocene–Eocene regional horizons in the Voronezh–Donets Region of the South European Russia, in Mater. LXIII sess. Paleontol. obshch. pri RAN (3–7 aprelya 2017 g., Sankt-Peterburg) (Proc. LXIII Sess. Paleontol. Soc. Russ. Acad. Sci. (April 3–7, 2017, St. Petersburg)), St. Petersburg: Vseross. Nauchno-Issled. Geol. Inst., 2017b, pp. 31–33.
Beniamovsky, V.N., An updated stratigraphic scheme (a new generation) of Paleogene deposits of the southern part of the Russian Plate and the northern part of the Scythian Plate – a key basin in the sublatitudinal communication system of the Paleogene epicontinental seas of Eurasian Peri-Tethys, in Mater. XXII Mezhd. nauchn. konf. (shkoly) po morskoi geologii “Geologiya morei i okeanov.” T. I (Proc. XXII Int. Sci. Conf. (School) on Marine Geology “Geology of Seas and Oceans.” Vol. I), Moscow: IO Ross. Acad. Nauk, 2017c, pp. 45–49.
Boiko, N.I. and Sklyarenko, G.Yu., Titanium–zirconium placers of the Poltava Formation on the southern slope of the Voronezh Anteclise, in Problemy mineralogii, petrografii i metallogenii. Nauchn. Ch. pamyati P.N. Chirvinskogo (P.N. Chirvinsky’s Lection “Problems of Mineralogy, Petrography, and Metallogeny”), 2014, vol. 17, pp. 181–187.
Brinkhuis, H., Late Eocene to Early Oligocene dinoflagellate cysts from the Priabonian type-area (Northeast Italy): biostratigraphy and paleoenvironmental interpretation, Palaeogeogr., Palaeoclimatol., Palaeoecol., 1994, vol. 107, pp. 121–163.
Bugrova, E.M., Beniamovsky, V.N., Tabachnikova, I.P., Ryabokon’, T.S., and Radionova, E.P., The Kantemirovka Paleogene reference section of the Voronezh Region, Stratigr. Geol. Correl., 2016, vol. 24, no. 6, pp. 602–624.
Costa, L.I. and Manum, S.B., The distribution of the interregional zonation of the Paleogene (D1–D15) and the Miocene (D16–D20), in The Northwest European Tertiary Basin. Results of the International Geological Correlation Program, Project No. 124, Geol. Jb., Hannover, 1988, A100, pp. 321–330.
Frieling, J., Iakovleva, A.I., Reichart, G.-J., Aleksandrova, G.N., Gnibidenko, Z.N., Schouten, S., and Sluijs, A., Paleocene–Eocene warming and biotic response in the epicontinental West Siberian Sea, Geology, 2014, vol. 42, no. 9, pp. 767–770.
Geologiya SSSR. Tom 46. Rostovskaya, Volgogradskaya, Astrakhanskaya oblasti i Kalmytskaya ASSR. Chast’ 1. Geologicheskoe opisanie (Geology of the USSR. Vol. 46. Rostov, Volgograd, and Astrakhan Regions and Kalmyk ASSR. Part 1. Geological Description), Belov, F.A., Egorov, A.I., and Pogrebnov, N.I., Eds., Moscow: Nedra, 1969 [in Russian].
Gorbatkina, T.E. and Iosifova, Yu.I., New Paleogene stratigraphic units of the Voronezh Antecline – Vorobyevskaya and Pasekovskaya formations, Vestn. Voronezh. Gos. Univ. Geol., 2004, no. 7, pp. 28–44.
Goretsky, G.I., Paleopotamologicheskie eskizy Paleo-Dona i Pra-Dona (Paleopotamological Sketches of the Paleo-Don and Pra-Don Rivers), Minsk: Nauka Tekhn., 1982 [in Russian].
Gosudarstvennaya geologicheskaya karta Rossiiskoi Federatsii. Masshtab 1 : 1000000 (tret’e pokolenie). Seriya Tsentral’no-Evropeiskaya. List M-38 - Volgograd. Ob"yasnitel’naya zapiska (The 1 : 1 000 000 State Geological Map of the Russian Federation (3rd ed). Ser. Central European. Sheet M-38 (Volgograd). Explanatory Note), Zastrozhnov, S.I., Ed., St. Petersburg: Kartfabr. Vseross. Nauchno-Issled. Geol. Inst., 2009 [in Russian].
Heilmann-Clausen, C., The Danish Subbasin, Paleogene dinoflagellates, in The Northwest European Tertiary Basin. Results of the International Geological Correlation Program, Project No. 124, Geol. Jb., Hannover, 1988, A100, pp. 339–343.
Iakovleva, A.I., Middle–late Eocene dinoflagellate cysts from NE Ukraine (Borehole no. 230, Dnepr-Donets Depression): stratigraphic and paleoenvironmental approach, Acta Palaeobot., 2015, vol. 55, no. 1, pp. 19–51.
Iakovleva, A.I., Detalization of the Eocene dinocyst zonation for Eastern Peri-Tethys, Bull. Moscow Soc. Nat. Geol. Ser., 2017, vol. 92, no. 2, pp. 32–48.
Iakovleva, A.I. and Aleksandrova, G.N., To the question of dinocyst zonation of the Paleocene–Eocene deposits of Western Siberia, Bull. Moscow Soc. Nat. Geol. Ser., 2013, vol. 88, no. 1, pp. 59–81.
Iakovleva, A.I. and Heilmann-Clausen, C., Early and middle Eocene dinoflagellate cysts from the Aktulagay section, Kazakhstan, Palynology, 2020. https://doi.org/10.1080/01916122.2019.1705933
King, Ch., Iakovleva, A.I., Heilmann-Clausen, C., and Steurbaut, E., Ypresian (early Eocene) stratigraphy of the Suvlu-Kaya reference section in the Bakhchisaray area (Crimea), Newsl. Stratigr., 2018, vol. 51, no. 2, pp. 167–208.
Lebedeva, N.K., Dinocyst biostratigraphy of the Upper Cretaceous deposits in the Usa River Basin (Polar Cis-Urals), Stratigr. Geol. Correl., 2005, vol. 13, no. 3, pp. 310–326.
Lebedeva, N.K., Dinocyst biostratigraphy of the Upper Cretaceous of Northern Siberia, Paleontol. J., 2006, vol. 40, no. 5, pp. S604–S621.
Lebedeva, N.K., Dinocyst-based biostratigraphy of the Upper Cretaceous deposits of North Siberia, in Sb. nauchn. tr. “Melovaya sistema Rossii i blizhnego zarubezh’ya: problemy stratigrafii i paleogeografii” (Proceedings “Cretaceous System of Russia and CIS Countries: Problems of Stratigraphy and Paleogeography”), Pervushov, E.M., Ed., Saratov: Saratov. Gos. Univ., 2007, pp. 108–122.
Lebedeva, N.K., Aleksandrova, G.N., Shurygin, B.N., Ovechkina, M.N., and Gnibidenko, Z.N., Paleontological and magnetostratigraphic data on the Upper Cretaceous deposits from borehole no. 8 (Russkaya Polyana District, Southwestern Siberia), Stratigr. Geol. Correl., 2013, vol. 21, no. 1, pp. 48–78.
Leonov, G.P., Paleogene deposits of the Stalingrad Volga Region and their relationships with related formations of the Don and Dnieper river basins, Byull. Mosk. O–va Ispyt. Prir., Otd. Geol., 1936, vol. XIV, no. 4, pp. 287–321.
Leonov, G.P., Stratigraphy of the Paleogene deposits of the Middle Don River basin, Uch. Zap. MGU. Geol., 1939, vol. 26, pp. 3–51.
Leonov, G.P., Osnovnye voprosy regional’noi stratigrafii paleogena Russkoi plity (The Main Questions of the Paleogene Regional Stratigraphy of the Russian Plate), Moscow: Mosk. Gos. Univ., 1961 [in Russian].
Mudge, D.C. and Bujak, J.P., Eocene stratigraphy of the North Sea Basin, Mar. Petrol. Geol., 1994, vol. 11, pp. 166–181.
Musatov, V.A., Lutetian or Bartonian? Nannoplankton-based age of the Sergeevskaya Formation in the Kantemirovka section of the Voronezh anteclise and correlation with the adjacent regions, Nedra Povolzh’ya Prikaspiya, 2020, vol. 101, pp. 4–26.
Musatov, V.A. and Bogachkin, A.B., The Bartonian Stage of the Middle Eocene of the European Russia. Stratigraphic interval and boundary determination criteria. Article 1. The accuracy limits of the zonal stratigraphy of the Lutetian–Bartonian interval based on nannoplankton and the age of Bartonian strata in England, Nedra Povolzh’ya Prikaspiya, 2019a, vol. 98, pp. 3–24.
Musatov, V.A. and Bogachkin, A.B., Bartonian Stage of the Middle Eocene within the European Russia. Stratigraphic interval and boundary determination criteria. Article 2. Detailed zonal stratigraphy of the Lutetian–Bartonian interval based on nannoplankton and paleomagnetic characteristics of Caucasian sections, Nedra Povolzh’ya Prikaspiya, 2019b, vol. 99, pp. 3–34.
Musatov, V.A. and Ryabokon’, T.S., The distribution of nannofossils and foraminifers in the Kiev Formation reference section (the village of Khalep’e (Kiev Dnieper Region, Ukraine), in Mater. XXXVIII sess. Paleontol. Obshch. NAN Ukrainy “40 let Paleontologicheskomu obshchestvu Ukrainy“ (Kanev, 23-26 maya 2017 g.) (Proc. XXXVIII Sess. Paleontol. Soc. Ukraine Nat. Acad. Sci. “The 40th Anni. Paleontol. Soc. Ukraine” (Kanev, May 23–26, 2017)), Kyiv: Inst. Geol. Nauk Nat. Akad. Nauk. Ukraine, 2017, pp. 117–119.
Musatov, V.A., Vasilyeva, O.N., and Muzylev, N.G., Results of the study of nannoplankton and dinocysts from the neostratotype of the Keresty Formation in the Southern Ergeni, in PALEOSTRAT-2014. Godichnoe sobranie (nauchn. konf.) sektsii paleontol. MOIP i Mosk. otd. Paleontol. Obshch. pri RAN (Moskva, 27–29 yanvarya 2014 g.) (Proc. Ann. Meet. (Sci. Conf.) Paleontol. Sect. Moscow Soc. Nat. Sci. and Moscow Branch Paleontol. Soc. “PALEOSTRAT-2014,” (Moscow, January 27–29, 2014)), Alekseev, A.S., Ed., Moscow: Paleotol. Inst., Ross. Akad. Nauk, 2014, pp. 53–54.
Ogg, J.G., Ogg, F.M., and Gradstein, F.M., A Concise Geologic Time Scale 2016, Elsevier, 2016.
Oreshkina, T.V. and Iakovleva, A.I., New paleontological data on the Buchak, Kiev, and Obukhov formations of the northern side of the Dnieper–Donets depression (borehole 230, settl. Strelech’e), in Paleontol. doslidzhennya v Ukraïni: istoriya, suchasnii stanta perspektivi. Zbor. nauk. pr. IGN NAN Ukraïni (Proceedings “Paleontological Researches in Ukraine: History, Modern State, and Perspectives”), Kyiv, 2007, pp. 233–237.
Popov, S.V., Akhmetiev, M.A., Zaporozhets, N.I., Voronina, A.A., and Stolyarov, A.S., Evolution of the Eastern Paratethys in the Late Eocene–Early Miocene, Stratigr. Geol. Correl., 1993, vol. 13, no. 6, pp. 10–39.
Popov, S.V., Akhmetiev, M.A., Lopatin, A.V., Bugrova, E.M., Sychevskaya, E.K., Shcherba, I.G., Andreeva-Grigorovich, A.I., Zaporozhets, N.I., Nikolaeva, I.A., and Kopp, M.L., Paleogeografiya i biogeografiya basseinov Paratetisa. Chast’ 1. Pozdnii eotsen-rannii miotsen (Paleogeography and Biogeography of the Paratethys Basins. Part 1: Late Eocene–Early Miocene), Moscow: Nauchn. Mir, 2009 [in Russian].
Postanovleniya Mezhvedomstvennogo stratigraficheskogo komiteta i ego postoyannykh komissii. Vyp. 32 (Resolutions of the Interdepartmental Stratigraphic Committee and its Permanent Commissions. Vol. 32), St. Petersburg: Vseross. Nauchno-Issled. Geol. Inst., 2001 [in Russian].
Powell, A.J., A Stratigraphic Index of Dinoflagellate Cysts, British Micropaleontol. Soc. Publ. Ser., 1992.
Powell, A.J., Brinkhuis, H., and Bujak, J.P., Upper Paleocene–lower Eocene dinoflagellate cyst sequence biostratigraphy of southeast England, in Correlation of the Early Paleogene in Northwest Europe. Geol. Soc. Spec. Publ., 1996, vol. 101, pp. 145–183.
Prakticheskaya palinostratigrafiya (Practical Palynostratigraphy), Panova, L.A., Oshurkova, M.V., Romanovskaya, G.M., Eds., Leningrad: Nedra, 1990 [in Russian].
Savko, A.D., Belyaev, V.I., and Ikonnikov, N.N., Titan-tsirkonievye rossypi Tsentral’no-Chernozemnogo raiona (Titanium-Zirconium Placers of the Central Chernozem Region), Voronezh: Voronezh. Gos. Univ., 1995 [in Russian].
Savko, A.D., Zvonarev, A.E., and Ivanov, D.A., Zirconium-titanium placers of the Voronezh anteclise: Types, epochs and factors of formation, and forecast, Geol. Ore Deposits, 2012, vol. 54, no. 1, pp. 60–80.
Semenov, V.P., Paleogen Voronezhskoi anteklizy (Paleogene of the Voronezh Anteclise), Voronezh: Voronezh. Gos. Univ., 1965 [in Russian].
Semenov, V.P., Voronezh anteclise, in Stratigrafiya SSSR. Paleogenovaya sistema (Stratigraphy of the USSR. Paleogene System), Grossgeim, V.A. and Korobkov, I.A., Eds., Moscow: Nedra, 1975, pp. 183–191.
Semenov, G.I. and Stolyarov, A.S., Problems of stratigraphy of the Maikop deposits of the Volga–Don region, Byull. Mosk. O–va Ispyt. Prir., Otd. Geol., 1988, vol. 63, no. 2, pp. 70–83.
Shcherbinina, E.A., Gavrilov, Yu.O., Iakovleva, A.I., Golovanova, O.V., Pokrovsky, B.G., and Aleksandrova, G.N., Environmental dynamics during the Paleocene–Eocene thermal maximum (PETM) in the northeastern Peri-Tethys revealed by high-resolution micropaleontological and geochemical studies of the Caucasian key-section, Palaeogeogr., Palaeoecol., Palaeoclimatol., 2016, vol. 456, pp. 60–81.
Shcherbinina, E., Iakovleva, A., Gavrilov, Yu., Golovanova, O., and Myzylöv, N., Lower Eocene sedimentary succession and microfossil biostratigraphy in the central northern Caucasus basin, Geol. Acta, 2020, vol. 18, no. 1, pp. 1–15.
Shpul, V. G., New data on phytostratigraphy of the southeastern slope of the Voronezh anteclise in the Eocene–Oligocene time, Vestn. Voronezh. Gos. Univ. Geol., 2005, no. 1, pp. 55–69.
Shpul, V. G., Eocene biostratigraphy of the northeastern slope of the Voronezh anteclise, in Problemi biostratigrafii nizhnego proterozoyu i fanerozoyu Ukraini: zb. nauk. prats'. Inst. Geol. Nauk (Proceedings Inst. Geol. Nauk “Problems of the Lower Proterozoic and Phanerozoic Biostratigraphy”), Kiev, 2006, pp. 208–214.
Shpul, V. G., Eocene-Oligocene flora of theVoronezh anteclise according to palynological data, in Paleontologichni doslidzhennya v ukraini: istoriya, suchasnii stan ta perspektivi: sbornik statei (Proceedings “Paleontological Researches in Ukraine: History, Modern State, and Perspectives”), Kiev, 2007, pp. 255–260.
Shpul, V. G., The Paleocene–Lower Eocene palynostratigraphy of the northeastern slope of the Voronezh anteclise, in Biosfernye osnovy postroeniya stratigraficheskikh skhem fanerozoya Ukrainy. Mater. XXXI Sess. Paleontol. Obshch. NAN Ukrainy, 19-22 maya 2008 g. (Proc. XXXI Sess. Paleontol. Soc. Ukraine Nat. Acad. Sci. “Biosphere Grounds for Developing the Phanerozoic Stratigraphic Schemes of Ukraine,” May 19–22, 2008), Simferopol, Kiev, 2008, pp. 144–149.
Shpul, V. G., Paleocene–Eocene floras of the Voronezh anteclise based on the palynological data, in Vikopna fauna i flora Ukraini: paleoekologichnii ta stratigrafichnii aspekti: zb. Nauk. prats' Inst. Geol. nauk NAN Ukraini (Coll. Sci. Works Inst Geol. Sci. Ukraine Nat. Acad. Sci. “Fossil Fauna and Flora of Ukraine: Paleoecological and Stratigraphic Aspects”), Kiev, 2009, pp. 220–225.
Shpul, V. G., The significance of palynology for substantiating regional stratigraphic subdivisions of the Paleogene of the Voronezh anteclise, in Mater. 4-go Mezhd. Simp. “Evolyutsiya zhizni na Zemle”, 10–12 noyabrya 2010 g. (Proc. 4th Int. Symp. “Evolution of the Life in the Earth,” November 10–12, 2010), Tomsk, 2010, pp. 426–429.
Sklyarenko, G.Yu., Titanium-zirconium placers in the Miocene deposits of the Poltava Group on the southeastern slope of the Voronezh anteclise, Cand. (Geol.-Mineral.) Dissertation, Rostov-on-Don, Yuzhn. Federal. Univ., 2010.
Śliwińska, K.K., Abrahamsen, N., Beyer, C., Brunings-Hansen, T., Thomsen, E., Ulleberg, K., and Heilmann-Clausen, C., Bio- and magnetostratigraphy of Rupelian–mid Chattian deposits from the Danishland area, Rev. Palaeobot. Palynol., 2012, vol. 172, pp. 48–69.
Sokolov, N.A., Lower Tertian Deposits of the Southern Russia, in Tr. Geol. Kom. (Trans. Geol. Com.), 1893, vol. IX, no. 2.
Stolyarov, A.S., Paleogeography of the Ciscaucasus, Volga–Don, and Southern Mangyshlak Region in the Late Eocene–Early Oligocene, Byull. Mosk. O-va Ispyt. Prir., Otd. Geol., 1991, vol. 66,no. 4, pp. 64–79.
Stolyarov, A.S. and Ivleva, E.I., Upper Oligocene rocks in the Ciscaucasus, Volga–Don and Mangyshlak regions (central part of the East Paratethys): Communication 1. Main compositional and structural features, Lithol. Miner. Resour., 2004a, vol. 39, no. 3, pp. 23–35.
Stolyarov, A.S. and Ivleva, E.I., Upper Oligocene sediments in the Ciscaucasus, Volga–Don, and Mangyshlak regions (central part of the Eastern Paratethys): Communication 2. Facies–paleogeographic sedimentation settings, Lithol. Miner. Resour., 2004b, vol. 39, no. 4, pp. 308–316.
Stratigraficheskaya skhema paleogenovykh otlozhenii Ukrainy (unifitsirovannaya) (Unified Stratigraphic Scheme of Paleogene Deposits of Ukraine), Makarenko, D.E., Ed., Kyiv: Nauk. Dumka, 1987 [in Russian].
Stratigrafiya SSSR. Paleogenovaya sistema (Stratigraphy of the USSR. Paleogene System). Grossgeim, V.A. and Korobkov, I.A., Eds., Moscow: Nedra, 1975 [in Russian].
Unifitsirovannaya stratigraficheskaya skhema paleogenovykh otlozhenii Povolzhsko-Prikaspiiskogo subregiona. Ob"yasnitel’naya zapiska (Unified Stratigraphic Scheme of the Paleogene Deposits of the Volga–Pre-Caspian Subregion. Explanatory Note), Akhmetiev, M.A., Ed., Moscow: Vseross. Nauchno-Issled. Geol. Neft. Inst., 2015 [in Russian].
Vasilyeva, O.N., The Middle Eocene organic-walled phytoplankton in the Keresta-1 parastratotype section (Ergeni, Rostov Region), in Ezhegodnik-2012. Tr. IGG UrO RAN. Vyp. 160 (Yearbook-2012. Trans. Inst. Geol. Geophys. Ur. Br. Russ. Acad. Sci. Vol. 160), Ekaterinburg: Izd. Inst. Geol. Geokhim. Ural. Otd. Ross. Akad. Nauk, 2013, pp. 13–21.
Vasilyeva, O.N., Dinocyst zonation of the Middle Eocene deposits of the Volgograd Volga Region, in Ezhegodnik-2018 (Yearbook-2018), Ekaterinburg: Izd. Inst. Geol. Geokhim. Ural. Otd. Ross. Akad. Nauk, 2019, pp. 3–11.
Vasilyeva, O.N. and Musatov, V.A., Paleogene biostratigraphy of the North Caspian Region based on dinocysts and nannofossils (Novouzensk Reference Borehole). Article 1. Substantiation of age of sediments and correlation, Stratigr. Geol. Correl., 2010a, vol. 18, no. 1, pp. 83–104.
Vasilyeva, O.N. and Musatov, V.A., Paleogene biostratigraphy of the North Caspian region based on dinocysts and nannofossils from the Novouzensk borehole. Article 2: Biotic events and paleoecological settings, Stratigr. Geol. Correl., 2010b, vol. 18, no. 2, pp. 179–199.
Vasilyeva, O.N. and Musatov, V.A., The Paleogene Dinoflagellate Cyst and Nannoplankton Biostratigraphy of the Caspian Depression, Intech: Stratigraphic Analysis of Layered Deposits, 2012, vol. 7, pp. 161–194.
Voronina, A.A., Kurgalimova, G.G., Popov, S.V., Semenov, G.I., and Stolyarov, A.S., Biostratigraphy and facies features of the Maikop deposits of the Volga–Don Region, Izv. AN SSSR. Ser. Geol., 1988, no. 9, pp. 39–50.
Williams, G.L., Fensome, R.A., and MacRae, R.A., The Lentin and Williams Index of fossil dinoflagellates 2017 edition, Am. Assoc. Stratigr. Palynol. Contrib. Ser., 2017, no. 48.
Yanin, B.T. and Beniamovsky, V.N., The burrows of decapod crustaceans from Paleogene deposits of the Middle Volga Region: Paleogeographic implications, Moscow Univ. Geol. Bull., 2018, vol. 73, no. 3, pp. 141–149.
Zaklinskaya, E.D., Description of some spore–pollen species extracted from the Tertiary deposits of the Pasekovsky quarry, Voronezh Region, in Tr. GIN AN SSSR. Ser. Geol. Vyp. 142 (Trans. Geol. Inst. USSR Acad. Sci. Ser. Geol. Vol. 142), 1953, no. 59, pp. 60–102.
Zaporozhets, N.I., New data on the Eocene and Oligocene phytostratigraphy of Severnye Ergeni (the south of the Russian Platform), Stratigr. Geol. Correl., 1998, vol. 6, no. 3, pp. 262–279.
Zaporozhets, N.I., Palynological assemblages from the Middle Eocene Kuma horizon and its age analogues in adjacent regions, Stratigr. Geol. Correl., 2001, vol. 9, no. 6, pp. 603–622.
Zaporozhets, N.I. and Akhmetiev, M.A., Assemblages of organic-walled phytoplankton, pollen, and spores from the Solenaya Horizon (lower Oligocene) of Western Eurasia, Stratigr. Geol. Correl., 2015, vol. 23, no. 3, pp. 326–350.
Zaporozhets, N.I. and Akhmetiev, M.A., Paleobotanical study of a section of the Oligocene–Lower Miocene Maikop Group along the Belaya River above the city of Maikop, Ciscaucasia, Stratigr. Geol. Correl., 2017, vol. 25, no. 6, pp. 638–658.
Zastrozhnov, A.S., Popov, S.V., Beniamovsky, V.N., Musatov, V.A., Akhmetiev, M.A., Zaporozhets, N.I., Bogachkin, A.B., and Stolyarov, A.S., The Oligocene Reference Section in the South of the East European Platform (Northern Ergeni), Stratigr. Geol. Correl., 2019, vol. 27, no. 4, pp. 442–474.
Zinova, R.A., The Solenovian horizon described by G.I. Goretsky and its age analogues, Litosfera, 2001, no. 14, pp. 32–38.
Zosimovich, V.Yu., The Paleogene regional stages of the North Ukrainian paleosedimentation province, in Mater. Mezhd. nauchn. konf. “Stratigrafiya osadochnykh obrazovanii verkhnego proterozoya i fanerozoya” (Proc. Int. Conf. “Stratigraphy of the Upper Paleozoic and Phanerozoic Sedimentary Rocks”), Kiev: Lat & K, 2013, pp. 66–67.
Zosimovich, V.Yu. and Shevchenko, T.V., The development stages of sedimentary basins of the Northern Ukraine during the Paleogene, in Zb. nauk. pr. IGN NAN Ukraïni (Proceedings Inst. Geol. Sci. Nat. Acad. Sci. Ukraine), 2014, vol. 7, pp. 83–100.
Zosimovich, V.Yu. and Shevchenko, T.V., Paleogene deposits of the Northern Ukraine sedimentation paleoprovince, in Zb. nauk. pr. IGN NAN Ukraïni (Proceedings Inst. Geol. Sci. Nat. Acad. Sci. Ukraine), 2015, vol. 8, pp. 68–121.
Zosimovich, V.Yu., Zernets’kii, B.F., Andreeva-Grigorovich, A.S., Lyul’eva, S.A., Maslun, N.V., Ryabokon’, T.S., and Shevchenko, T.V., The Paleogene regional stages of the platform Ukraine, in Biostratigrafichni kriteriï rozchlenuvannya ta korelyatsiï vidkladiv fanerozoyu Ukraïni. Zb. nauk. pr. IGN NAN Ukraïni (Proceedings Inst. Geol. Sci. Nat. Acad. Sci. Ukraine “Biostratigraphic Criteria of Subdivision and Correlation of the Phanerozoic Deposits of Ukraine”), Kiev, 2005, pp. 118–132.
Zosimovich, V.Yu., Shevchenko, T.V., and Tsyba, N.N., The “Kanev Stage” neostratotype, in Zb. nauk. pr. IGN NAN Ukraïni (Proceedings Inst. Geol. Sci. Nat. Acad. Sci. Ukraine), 2013, vol. 6, no. 1, pp. 98–110.
Zosimovich, V.Yu., Zernetsky, B.F., Olshtynskaya, A.P., Ryabokon’, T.S., and Shevchenko, T.V., Correlation of Paleogene regional stages of Northern and Southern Ukraine, in Stratotipovi ta oporni rozrizi fanerozois’kikh vidkladiv Ukraïni: suchasnii stan paleontologichnoï vivchenosti ta perspektivi podal’shikh doslidzhen'. Mater. XXXVI ses. Paleontol. tov-va NAN Ukraïni (L’viv, 24-26 veresnya 2015 r.) (Proc. XXXVI Sess. Paleontol. Soc. Nat. Acad. Sci. Ukraine “Stratotypes and Reference Sections of the Phanerozoic Deposits of Ukraine: Modern State of Paleontological Knowledge and Perspectives of Further Research,” Lvov, September 24–26, 2015), Kiev: Inst. Geol. Nauk Nat. Akad. Nauk Ukrainy, 2015, pp. 53–54.
Zosimovich, V.Yu., Shevchenko, T.V., Ryabokon’, T.S., and Ol’shtynskaya, A.P., Regional stratigraphic subdivisions of the Paleogene of the Northern Ukraine, in Sostoyanie stratigraficheskoi bazy tsentra i yugo-vostoka Vostochno-Evropeiskoi platformy. Mater. soveshch. (Moskva, VNIGNI, 23–25 noyabrya 2015 g.) (Proc. Conf. “State of the Stratigraphic Base of the Central and Southeastern Parts of the East European Platform” (Moscow, All-Russ. Res. Geol. Oil Inst., November 23–25, 2015)), Moscow: Vseross. Nauchno-Issled Geol. Neft. Inst., 2016, pp. 140–144.
Zosimovich, V.Yu., Zernetsky, B.F., Olshtynskaya, A.P., Ryabokon’, T.S., and Shevchenko, T.V., Correlation of the Paleogene regional stages of the Northern and Southern Ukraine, in Aktual’ni problemi ta perspektivi rozvitku geologiï: nauka i virobnitstvo. Mater. Mizhnarod. Geol. forumu (19-24 chervnya 2017 r., m. Odesa, Ukraïna) (Proc. Int. Geol. Forum “Actual Problems and Perspectives of the Development of Geology: Science and Practice (June 19–24, 2017. Odessa, Ukraine)), Kiev: UkrDGRI, 2017, pp. 141–144.
ACKNOWLEDGMENTS
We thank the North Caucasus Geological Production Association for the core material and our colleagues A.S. Zastrozhnov (Karpinsky Russian Geological Research Institute), S.V. Popov (Paleontological Institute, Russian Academy of Sciences), S.V. Grishin (Geological Institute, Russian Academy of Sciences), and N.I. Panchenko (Volgograd) for assistance in fieldwork. We are grateful to N.K. Lebedeva (Trofimuk Institute of Petroleum Geology and Geophysics, Siberian Branch, Russian Academy of Sciences), S.V. Popov (Paleontological Institute, Russian Academy of Sciences), E.Yu. Baraboshkin, Yu.B. Gladenkov, and M.A. Rogov for valuable remarks and comments.
Funding
The research was performed within the framework of the State program no. 0135-2019-0044 (Geological Institute, Russian Acad. Sci.). Fieldwork and analytical studies were supported by the Russian Foundation for Basic Research (project no. 18-05-00505a).
Author information
Authors and Affiliations
Corresponding author
Additional information
Translated by E. Murashova
Reviewers E.Yu. Baraboshkin, N.K. Lebedeva, and S.V. Popov
Rights and permissions
About this article

Cite this article
Iakovleva, A.I., Aleksandrova, G.N. The Paleogene of the Cis-Donets Monocline and Its Palynological Characteristics. Stratigr. Geol. Correl. 29, 65–103 (2021). https://doi.org/10.1134/S0869593821010093
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1134/S0869593821010093