
Abstract
Maonan nationality is a relatively conservative and isolated minority in China. Little is known about the association of the Slit-Robo Rho GTPase activating protein 2 gene (SRGAP2) single nucleotide polymorphisms (SNPs) and serum lipid levels in the Chinese populations. This study was performed to clarify the association of the SRGAP2 rs2483058 and rs2580520 SNPs and their haplotypes with serum lipid traits in the Maonan and Han populations. Genotyping of the 2 SNPs was performed in 2444 unrelated subjects (Han, 1210 and Maonan, 1234) by polymerase chain reaction and restriction fragment length polymorphism combined with gel electrophoresis, and then confirmed by direct sequencing. The allelic (rs2483058) and genotypic (rs2483058 and rs2580520) frequencies were different between the two ethnic groups. Four haplotypes were identified in our populations, and the rs2483058G-rs2580520C haplotype was the commonest one. The rs2483058C-rs2580520G haplotype was associated with an increased risk of dyslipidemia, and showed consistent association with serum total cholesterol (TC), high-density lipoprotein cholesterol (HDL-C), apolipoprotein (Apo) A1 levels, and the ApoA1/ApoB ratio. These results indicated that the SRGAP2 SNPs and their haplotypes were associated with serum lipid levels. Their haplotypes can explain much more serum lipid variation than any single SNP alone, especially for serum TC, HDL-C and ApoA1 levels.
Similar content being viewed by others

Introduction
During the past decades, cardiovascular disease (CVD) has become the leading cause which can give rise to the world’s largest mortality, morbidity, disability, functional decline, and healthcare costs1, 2. To evaluate risk severity, we general survey a standard lipid profile, just as total cholesterol (TC)3, triglyceride (TG)4, low-density lipoprotein cholesterol (LDL-C)5, apolipoprotein (Apo) B6, high-density lipoproteins cholesterol (HDL-C)7, 8, ApoA18 and the ratio ApoA1 to ApoB9, which is an efficient way for cardiovascular risk prediction and can be recommended from an integral component of approaches. Several researches in the past years about CVD risk factors have showed that the morbidity was usually different between men and women10, was also affected by age11 and ethnicity12, and was modified by behavioral choices13, poor diet14 and unhealthy lifestyle15, environmental factors16, and personal genetic profile17, 18. All these risk factors which have been taken to individual are important genetic components, however, there are lots of the true magnitude risk factor are uncertain to cluster, as well as on the role of genetic factors in risk factor clustering for individuals. In that case, the target of genome-wide association studies (GWASes) was to find out which part can identify common single nucleotide polymorphisms (SNPs) and calculate the numbers of the phenotypic variance is actually located by them19.
Some GWASes have demonstrated that several SNPs near the SLIT-ROBO Rho GTPase activating protein 2 gene (SRGAP2; also knows as: FNBP2; SRGAP3; SRGAP2A; ARHGAP34, Gene ID: 23380, HGNC ID: 19751, synonyms: FLJ33003, FLJ42565, KIAA0456, locus type: gene with protein product, chromosomal location: 1q32.1) may result in negatively regulate neuronal migration and induce neurite outgrowth20. In addition, it might also contribute to the higher susceptibility to neurodegenerative or psychiatric disorders of the human brain21. At the same time, a large number of surveys have showed that SRGAP2 expression was up regulated in multiple breast cancer cells. The mechanism was supposed to have a connection with lipid metabolism22. A previous GWAS on plasma lipid levels has identified the rs2483058 SNP near the SRGAP2 as hyperlipidemic locus in European23. In the meantime, several previous studies have showed that the association between the SRGAP2 rs2483058 SNP and serum lipid levels might have ethnic- and/or sex-specificity24,25,26. Besides these, another rs2580520 SNP has been clarified to contribute to the development of breast cancer27. As we have known that the hyperlipidemia may be an important part to result in breast cancer28, whether the SRGAP2 rs2483058 and rs2580520 SNPs are associated with serum lipid levels or whether they show ethnic- and/or sex-specific association as the previous reports remains dubious.
As we all know that China is a multi-ethnic country, including 56 nationalities. Han is the largest group and Maonan is one of the 55 minorities with a population of 107,166 (Rank 37) according to the sixth national census statistics of China in 2010. The Maonan people are mainly distributed in the Shangnan, Zhongnan, and Xianan townships of Huanjiang Maonan Autonomous County in the north of the Guangxi Zhuang Autonomous Region, which is situated in Southwestern China. Several previous studies have showed that the genetic relationship between Maonan nationality and other minorities in Guangxi29 was much closer than that between Maonan and Han nationalities30. In spite of a very small population, the Maonan ethnic group is well known in China for its long history and unique culture. The special customs and culture, including their clothing, intra-ethnic marriages, dietary habits and lifestyle factors are different from those of local Han Chinese31. They have their culture of consanguineous marriage to cousins of maternal side, suggesting that the genetic background of Maonan population may be less heterogeneous within the population. This study, therefore, was undertaken to detect the association of the SRGAP2 rs2483058 and rs2580520 SNPs and several environmental factors with serum lipid levels between males and females in the Maonan and Han populations.
Results
Demographic and biochemical characteristics
The demographic and biochemical characteristics of the participants according to ethnic group are presented in Table 1. The levels of body weight, body mass index (BMI), waist circumference, systolic blood pressure, diastolic blood pressure, pulse pressure and blood glucose and the percentages of subjects who consumed alcohol were higher in Maonan than in Han (P < 0.05- P < 0.001), whereas the levels of body height and the percentages of subjects who smoked cigarettes were lower in Maonan than in Han (P < 0.05). There was no significant difference in the levels of age, sex ratio, TC, TG, LDL-C, HDL-C, ApoA1, ApoB and the ratio of ApoA1 to ApoB (P > 0.05 for all).
Results of electrophoresis and genotyping
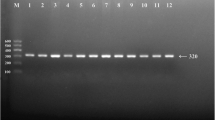
After the genomic DNA of the samples was amplified by polymerase chain reaction (PCR) and imaged by agarose gel electrophoresis for the SRGAP2 rs2483058 SNP, the PCR product of 382-bp nucleotide sequences could be seen in the samples (Figs 1 and 2). The GG (382 bp), GC (382-, 260- and 122-bp) and CC (260- and 122-bp) genotypes were shown, respectively. The PCR product of the rs2580520 SNP was 515-bp nucleotide sequences. The GG (515 bp), GC (515-, 389-, and 126-bp) and CC (389- and 126- bp) genotypes were shown, respectively.

Agarose gel electrophoresis (2%) of PCR products and genotyping of the SRGAP2 SNPs. (A) and (B) (rs2483058): Lane M, 100 bp marker ladder; lanes A1-A6, 382 bp band samples; lanes B1 and B4, CC genotype (260- and 122-bp); lanes B2, B3, B5 and B6, GC genotype (382-, 260- and 122-bp); Lanes B7 and B8, GG (382-bp). (C) and (D) (rs2580520): Lane M, 100 bp marker ladder; lanes C1-C6, 515 bp band samples; lanes D1 and D6, GG genotype (515-bp); lanes D3 and D5, GC genotype (515-, 389- and 126-bp); lanes D2 and D4, CC (389- and 126-bp).

The positions of the SRGAP2 rs2580520 and rs2483058 variants.
Genotypic and allelic frequencies
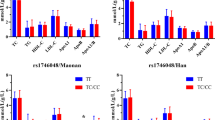
The genotypic distribution of the two loci was in Hardy-Weinberg equilibrium (P > 0.05 for all). The genotypic and allelic frequencies of the rs2483058 SNP were different between Maonan and Han, the frequencies of the rs2483058C allele (41.86% vs. 37.07%, P = 0.001) and rs2483058GC genotype (48.06% vs. 40.90%, P < 0.001) were higher in Maonan than in Han. Subgroup analysis showed that the rs2483058C allele frequency was higher in Han females than in Han males (39.06% vs. 33.87%, P = 0.01), but it was lower in Maonan females than in Maonan males (39.37% vs. 45.50%, P = 0.002). The genotypic frequencies of the rs2580520 SNP between Maonan and Han, and between Han males and Han females were also different (P < 0.05-P < 0.005; Table 2).
Genotypes and serum lipid levels
Tables 3 and 4 describe the association between genotypes and serum lipid levels. The levels of ApoA1 and the ratio of ApoA1 to ApoB in both Han and Maonan were different among the rs2483058 genotypes (P < 0.01 for all), the rs2483058C allele carriers had lower ApoA1 levels and the ApoA1/ApoB ratio than the rs2483058C allele non-carriers. The concentration of HDL-C in Maonan but not in Han was also different among the rs2483058 genotypes (P = 0.024), the rs2483058C allele carriers had lower HDL-C concentration than the rs2483058C allele non-carriers. The levels of TC, LDL-C and ApoB in Maonan but not in Han were different among the rs2580520 genotypes (P = 0.011-P = 0.001), the rs2580520C allele carriers had higher TC, LDL-C and ApoB levels than the rs2580520C allele non-carriers. Subgroup analysis showed that the levels of ApoA1 and the ratio of ApoA1 to ApoB in Han males and females and Maonan males were different among the rs2483058 genotypes (P < 0.01 for all), the rs2483058C allele carriers had lower ApoA1 levels and the ApoA1/ApoB ratio than the rs2483058C allele non-carriers. The levels of ApoA1 in Maonan females were also different among the rs2483058 genotypes (P < 0.001), the rs2483058C allele carriers had lower ApoA1 levels than the rs2483058C allele non-carriers. The levels of TC in Maonan males and females were different among the rs2580520 genotypes (P = 0.018-P = 0.001), the rs2580520C allele carriers had higher TC levels than the rs2580520C allele non-carriers. The ratio of ApoA1 to ApoB in Han males and Maonan males was also different among the rs2580520 genotypes (P = 0.013-P = 0.007), the rs2580520C allele carriers had lower the ApoA1/ApoB ratio than the rs2580520C allele non-carriers.
Haplotypes and serum lipid levels
A weak linkage disequilibrium (LD) was noted between the two SNPs (D′ = 0.58, r 2 = 0.33; Fig. 3). Thus, a haplotype analysis was conducted (Table 5). There were 4 haplotypes identified in our study populations. The haplotype of rs2483058G-rs2580520C was the commonest one (45%). The haplotype of rs2483058C-rs2580520G was associated with an increased risk of dyslipidemia (OR: 1.44, 95% CI: 1.17–1.78, P < 0.001), it showed consistent association with serum TC, HDL-C, ApoA1 and the ratio of ApoA1 to ApoB. Multivariate logistic analysis showed that the rs2483058C-rs2580520G haplotype was positively correlated with the incidence of hyperlipidemia in Han and Maonan according to stratified risk factors (gender, BMI, smoking, diabetes and blood pressure; Table 6). In addition, the rs2483058C-rs2580520G haplotype carriers had lower HDL-C and ApoA1 levels in Han and Maonan, lower ApoA1/ApoB ratio in Han, and higher TC in Maonan than the rs2483058C-rs2580520G haplotype non-carriers (Table 7).

The linkage disequilibrium (LD) of the SRGAP2 rs2483058 and rs2580520 SNPs in the combined population of Maonan and Han. The LD status is expounded by the (A) D′ = 0.58, (B) r 2 = 0.33.
Correlated factors for serum lipid parameters
Multivariable linear regression analyses showed that the levels of HDL-C and ApoA1 were associated with the rs2483058 genotypes and/or alleles in Han. The levels of TC were correlated with the rs2580520 genotypes and alleles, HDL-C and ApoA1 with the rs2483058 genotypes in Maonan (Table 8).
Serum lipid parameters were also correlated with several environmental factors such as sex, age, alcohol consumption, cigarette smoking, blood pressure, blood glucose, waist circumference, and BMI in both ethnic groups or in males and females (P < 0.05-P < 0.001; Tables 9 and 10).
Relative factors for serum lipid phenotypes
As shown in Fig. 4, Pearson correlation analysis demonstrated that the integrative variants and haplotype connected with the SRGAP2 rs2483058 and rs2580520 SNPs to lipid variables. Several environmental exposures such as age, gender, cigarette smoking, alcohol consumption and traditional cardiovascular risk factors such as BMI and blood pressure levels were also correlated with serum lipid phenotypes in the both ethnic groups.

Correlation between environmental exposures and serum lipid variables, as well as the candidate loci.
Discussion
The present study demonstrated that different SRGAP2 SNPs had different effect on serum lipid traits that (i) the levels of HDL-C, ApoA1 and the ratio of ApoA1 to ApoB in Maonan were different among the rs2483058 genotypes; (ii) the levels of TC in Maonan males and females and the ratio of ApoA1 to ApoB in Han and the level of LDL-C and ApoB in Maonan were different among the three rs2580520 genotypes; (iii) the levels of HDL-C and ApoA1 in males or females were different in Maonan and Han in rs2483058 genotypes; and (iv) the levels of TC and the ratio of ApoA1 to ApoB in males or females were different in Maonan and Han in rs2580520 genotypes.
In the current study, we found that the rs2483058C allele carriers had lower ApoA1 levels and the ApoA1/ApoB ratio than the rs2483058C allele non-carriers in both ethnic groups. The rs2483058C allele carriers also had lower HDL-C concentration than the rs2483058C allele non-carriers in Maonan but not in Han. The rs2580520C allele carriers had higher TC, LDL-C and ApoB levels than the rs2580520C allele non-carriers in Maonan but not in Han. The levels of TG, LDL-C and ApoB in the two ethnic groups were not significantly different among the rs2483058 and rs2580520 genotypes. It is well-known that dyslipidemia is a complex disease caused by environmental and genetic factors. Previous family and twin studies have suggested that in numerous populations, about 40-60% of the variation in serum lipid profiles is genetically determined32,33,34,35,36.
In some previous studies, there were associations with intra-pair differences in HDL-C, rs2483058 in an intron of SRGAP2, where twins carrying the C allele were more sensitive to environmental factors37. In the present study, we showed that the genotypic frequencies of our populations were similar to those obtained in other populations and to data available in the International HapMap Project’s database (http://www.hapmap.org) for the populations. The frequency of CC, GC and GG genotypes was 42.9%, 42.9% and 14.2% in Utah residents with ancestry from Northern and Western Europe (CEU); 7.2%, 56.4% and 36.4% in Yoruba in Ibadan, Nigeria (YRI); 9.3%, 37.2% and 53.4% in Japanese in Tokyo, Japan (JPT) and 9.5%, 40.5% and 50% in Han Chinese in Beijing, China (HCB). The frequency of C and G alleles was 64.3% and 35.7% in CEU; 27.9% and 72.1% in YRI; 35.5% and 64.5% in JPT and 29.8% and 70.2% in HCB. The present study identified significant differences in the genotypic frequency of the rs2483058 SNP between the two ethnic groups. The frequency of the CC genotype was higher in the Maonan than in the Han population and the frequency of the C allele was higher in the Maonan than in the Han population (41.86% vs. 37.03%; P = 0.001). These results suggest that the prevalence of the SRGAP2 rs2483058 SNP may exhibit a racial/ethnic difference.
In some previous studies, Jiang et al. had found that the rs2580520 genotypes in Chinese women may have different breast cancer susceptibility, which may contribute to the development of breast cancer in this population27. Coleman et al. also showed that obesity has been attributed as a risk factor for breast cancer development for over five decades and the contribution of obesity to postmenopausal breast cancer risk is thought to be related to elevated level of circulating estrogens in these women, resulting from aromatase-mediated conversion of androgens to estrogens in peripheral adipose tissue (and possible dysregulation of aromatase expression) and lower level of sex hormone-binding globin38. It is well-known that cholesterol is an important material for the synthesis of estrogen and higher cholesterol can give rise to more estrogen synthetic that may be enhanced a possibility to be suffered from breast cancer39, 40. The results of the present study clearly showed that the levels of TC were higher in who carried rs2580520C allele than without carried rs2580520C allele.
Important intra- and inter-genetic LD associations have been found in this study. These LD patterns in SRGAP2 were rather weak and low specific to the population under study and indicated the functional dependencies of the encoded proteins. In the present study, haplotype analysis with the two SNPs further supported the association between SRGAP2 polymorphisms and serum lipid levels in our study populations. The haplotype of rs2483058G-rs2580520C was the commonest one and represented about 45% of the samples. The haplotypes of rs2483058C-rs2580520C, rs2483058G-rs2580520G and rs2483058C-rs2580520G were associated with increased risk of dyslipidemia, but only rs2483058C-rs2580520G haplotype with a significant meaning. In addition, carriers of rs2483058C-rs2580520G haplotype had decreased serum concentration of HDL-C, ApoA1 and the ratio of ApoA1 to ApoB and increased serum concentration of TC. We also found that haplotypes could explain much more serum lipid variation than any single SNP alone.
The environmental factors just as dietary patterns, lifestyle, obesity, physical activity, and hypertension would also be play an important role in modifying serum lipid levels41,42,43. The dietary habits were different between the Han and Maonan populations. Rice is the Maonan people’s staple food supplemented with corn, sweet potato and other grains. Maonan people preferred to eat spicy and acid food with lots of oil and salt. This preference of high in carbohydrates may be related to the higher blood glucose levels, weight, BMI and waist circumference in Maonan than in Han people. In the meantime, rich oil and salt can give rise to higher blood pressure, serum TC, LDL-C and ApoB levels in Maonan than in Han people. There were lots of past studies had proved that diet alone could account for the variability on serum lipid levels44, 45. In the present study, we also found that serum lipid parameters were also correlated with several environmental factors such as age, alcohol consumption, cigarette smoking, blood pressure, blood glucose, waist circumference, and BMI. It is commonly accepted that the high-fat diet especially containing large quantities of saturated fatty acids, raise serum cholesterol concentrations and predispose subjects to coronary artery disease46. We also showed that the percentages of individuals who consumed alcohol were higher in Maonan than in Han. Although the effects of alcohol intake on LDL-C appear to vary by specific patient types or patterns of alcohol intake, and perhaps by population and sex hormone, this topic has been the focus of much recent research47. A recent study in older Italian individuals (65–84 years old) has found that alcohol intake reached higher serum LDL-C levels48. Another recent study of Turks also found increased LDL-C, ApoB and TG with alcohol in men, whereas decreased TG and did not change LDL-C or ApoB with alcohol in women49. Therefore, the results of exposure to different lifestyle and environmental factors probably further modify the association of genetic variations and serum lipid levels in our study populations.
The present study has some shortcomings. At first, the size of our study populations is a bit small, which might not have the power to detect the LD across the SRGAP2 locus. Next, the levels of body weight, waist circumference and the percentages of subjects who smoked cigarettes or consumed alcohol were higher in Maonan than in Han. Although age, BMI, blood pressure, cigarette smoking, and alcohol consumption have been adjusted for the statistical analysis, we cannot completely exclude the influence of these factors on serum lipid levels among different genotypes in both nationalities. In addition, because we selected the SNPs from literature and did not cover the extensive SRGAP2 locus, we might miss some information from other SNPs.
In summary, the genotypic and allelic frequencies of the SRGAP2 rs2483058 and rs2580520 SNPs were different between Maonan and Han. There were four haplotypes identified in our study populations. The SRGAP2 SNPs and rs2483058C-rs2580520G haplotype were closely sexually dimorphic associated with serum lipid traits. The haplotypes can explain much more serum lipid variation than any single SNP alone, especially for serum TC, HDL-C and ApoA1 levels.
Materials and Methods
Subjects
The study populations including 1210 unrelated subjects (465 males, 38.43% and 745 females, 61.57%) of Han and 1234 unrelated participants (500 males, 40.52% and 734 females, 59.48%) of Maonan were randomly selected from our previous stratified randomized samples. The participants were all agricultural workers from Huanjiang County, Guangxi Zhuang Autonomous region, People’s Republic of China. The participants’ age ranged from 25 to 80 years with a mean age of 55.77 ± 13.89 years in Han and 56.31 ± 13.80 years in Maonan; respectively. The age distribution and gender ratio were matched between the two groups. All participants were essentially healthy with no history of CVD such as coronary artery disease, stroke, diabetes, hyper- or hypo-thyroids, and chronic renal disease. They were free from medications known to affect serum lipid levels. The investigations were carried out following the rules of the Declaration of Helsinki of 1975 (http://www.wma.net/en/30publications/10policies/b3/), revised in 2008. The Ethics Committee of the First Affiliated Hospital, Guangxi Medical University approved the study protocol (No: Lunshen-2014-KY-Guoji-001; Mar. 7, 2014) prior to data collection, and all participants provided informed consent by signature or by fingerprint (if the participant was illiterate) after they had been informed of the objectives, benefits, medical items and confidentiality agreement of personal information. All procedures and methods were performed in accordance with the relevant ethical guidelines.
Epidemiological survey
The epidemiological survey was carried out using internationally standardized methods, following a common protocol50. Information on demographics, socioeconomic status, and lifestyle factors was collected with standardized questionnaires. Alcohol consumption was categorized into groups of grams of alcohol per day: 0 (non-drinker), <25 and ≥25. Smoking status was categorized into groups of cigarettes per day: 0 (non-smoker), <20 and ≥20. Several parameters such as blood pressure, height, weight, waist circumference, and BMI were measured. The methods of measuring above parameters were referred to previous studies51.
Biochemical analyses
A fasting venous blood sample of 5 ml was drawn from the participants. A part of the sample (2 mL) was collected into glass tubes and used to determine serum lipid levels. Another part of the sample (3 mL) was transferred to tubes with anticoagulants (4.80 g/L citric acid, 14.70 g/L glucose and 13.20 g/L tri-sodium citrate) and used to extract deoxyribonucleic acid (DNA). Measurements of serum TC, TG, HDL-C, and LDL-C levels in the samples were performed by enzymatic methods with commercially available kits (RANDOX Laboratories Ltd., Ardmore, Diamond Road, CrumlinCo. Antrim, United Kingdom, BT29 4QY; Daiichi Pure Chemicals Co., Ltd., Tokyo, Japan). Serum ApoA1 and ApoB levels were detected by the immunoturbidimetric immunoassay using a commercial kit (RANDOX Laboratories Ltd.). All determinations were performed with an auto-analyzer (Type 7170 A; Hitachi Ltd., Tokyo, Japan) in the Clinical Science Experiment Center of the First Affiliated Hospital, Guangxi Medical University52, 53.
DNA amplification and genotyping
Genomic DNA of the samples was isolated from peripheral blood leucocytes according to the phenol-chloroform method52, 53. The extracted DNA was stored at 4 °C until analysis. Genotyping of the SRGAP2 rs2483058 and rs2580520 SNPs were performed by polymerase chain reaction and restriction fragment length polymorphism (PCR-RFLP). PCR amplification was performed using rs2483058: 5′-CAGGGGGCACAGATAGTGGA-3′ as the forward and 5′-GCCAAGTTTGACTACGTGGG-3′ and rs2580520: 5′-AACAGGTTGGGGTGAGCATA-3′ as the forward and 5′-CCCCATCAGTACATCGTGGT-3′ as reversed primer pair (Sangon, Shanghai, People’s Republic of China), respectively. Each 25 μL PCR reaction mixture consisted of 2.0 μL genomic DNA, 1.0 μL each primer (10 μmol/L), 12.5 μL of 2 × Taq PCR Master mix (constituent: 0.1 U Taq polymerase/μL, 500 μM dNTP each and PCR buffer.), and 8.5 μL of ddH2O (DNase/RNase-free). PCR was performed with an initialization step of 95 °C for 5 min, followed by 30 s denaturing at 95 °C, 30 s of annealing at 60 °C and 35 s of elongation at 72 °C for 32 cycles. The amplification was completed by a final extension at 72 °C for 7 min. Following electrophoresis on a 2.0% agarose gel with 0.5 µg/mL ethidium bromide, the amplification products were visualized under ultraviolet light. Subsequently, each restriction enzyme reaction was performed with 5.0 μL amplified DNA, 8.8 μL nuclease-free water, 1.0 μL of 10 × buffer solution and 0.2 μL AvaI for rs2483058 and HPY188I for rs2580520 restriction enzyme in a total volume of 15 µL digested at 37 °C overnight. After restriction enzyme digestion of the amplified DNA, genotypes were identified by electrophoresis on 2% ethidium-bromide stained agarose gels and visualized with UV illumination. Genotypes were scored by an experienced reader blinded to the epidemiological and serum lipid results. Both of six samples (GG, GC and CC genotypes in two; respectively) detected by the PCR-RFLP were also confirmed by direct sequencing with an ABI Prism 3100 (Applied Biosystems) in Shanghai Sangon Biological Engineering Technology & Services Co., Ltd., People’s Republic of China.
Diagnostic criteria
The normal values of serum TC, TG, HDL-C, LDL-C, ApoA1, ApoB levels and the ApoA1/ApoB ratio in our Clinical Science Experiment Center were 3.10–5.17, 0.56–1.70, 1.16–1.42, 2.70–3.10 mmol/L, 1.20–1.60, 0.80–1.05 g/L and 1.00–2.50, respectively. The individuals with TC > 5.17 mmol/L and/or TG > 1.70 mmol/L were defined as dyslipidemia54. Hypertension was diagnosed according to the 1999 and 2003 criteria of the World Health Organization-International Society of Hypertension Guidelines for the management of hypertension55, 56. The diagnostic criteria of overweight and obesity were according to the Cooperative Meta-analysis Group of China Obesity Task Force. Normal weight, overweight and obesity were defined as a BMI < 24, 24–28 and > 28 kg/m2, respectively57.
Statistical analyses
The statistical analyses were performed with the statistical software package SPSS 22.0 (SPSS Inc., Chicago, Illinois). The quantitative variables were presented as mean ± standard deviation (serum TG levels were presented as medians and interquartile ranges for not a normal distribution). Allele frequency was determined via direct counting, and the Hardy-Weinberg equilibrium was verified with the standard goodness-of-fit test. The genotype distribution between the two groups was analyzed by the chi-square test. General characteristics between two ethnic groups were compared by the Student’s unpaired t-test. The association between genotypes and serum lipid parameters was tested by covariance analysis (ANCOVA). Any SNPs associated with serum lipid profiles at the value of P < 0.025 (corresponding to P < 0.05 after adjusting for 2 independent tests by the Bonferroni correction) were considered statistically significant. Gender, age, BMI, blood pressure, alcohol consumption and cigarette smoking were adjusted for the statistical analysis. Haploview (Broad Institute of MIT and Harvard, USA, version 4.2) analyzed the haplotype frequencies and pair-wise LD among the detected SNPs. Unconditional logistic regression was used to assess the correlation between the risk of hyperlipidemia and genotypes. Multivariable linear regression analyses with stepwise modeling were used to determine the correlation between the genotypes (GG = 1, GC = 2, CC = 3) and several environmental factors with serum lipid levels in males and females of Han and Maonan populations. Two sides P value < 0.05 was considered statistically significant. The heart-map of inter-locus models was measured by R software (version 3.3.0)58.
References
Yazdanyar, A. & Newman, A. B. The burden of cardiovascular disease in the elderly: morbidity, mortality, and costs. 25, 563–577 (2009).
Pandya, A., Gaziano, T. A., Weinstein, M. C. & Cutler, D. More americans living longer with cardiovascular disease will increase costs while lowering quality of life. Health affairs (Project Hope) 32, 1706–1714 (2013).
Matthan, N. R. et al. Sex-specific differences in the predictive value of cholesterol homeostasis markers and 10-year cardiovascular disease event rate in Framingham Offspring Study participants. Journal of the American Heart Association 2, e005066–e005066 (2013).
Lindman, A. S., Veierød, M. B., Tverdal, A., Pedersen, J. I. & Selmer, R. Nonfasting triglycerides and risk of cardiovascular death in men and women from the Norwegian Counties Study. European journal of epidemiology 25, 789–798 (2010).
Orekhov, A. N., Bobryshev, Y. V., Sobenin, I. A., Melnichenko, A. A. & Chistiakov, D. A. Modified low density lipoprotein and lipoprotein-containing circulating immune complexes as diagnostic and prognostic biomarkers of atherosclerosis and type 1 diabetes macrovascular disease. International journal of molecular sciences 15, 12807–12841 (2014).
Furtado, J. D. et al. Effect of protein, unsaturated fat, and carbohydrate intakes on plasma apolipoprotein B and VLDL and LDL containing apolipoprotein C-III: results from the OmniHeart Trial. The American journal of clinical nutrition 87, 1623–1630 (2008).
Chapman, M. J. et al. Triglyceride-rich lipoproteins and high-density lipoprotein cholesterol in patients at high risk of cardiovascular disease: evidence and guidance for management. European heart journal 32, 1345–1361 (2011).
Hamon, S. C. et al. Evidence for consistent intragenic and intergenic interactions between SNP effects in the APOA1/C3/A4/A5 gene cluster. Human heredity 61, 87–96 (2006).
Tamang, H. K. et al. Apo B/Apo A-I Ratio is Statistically A Better Predictor of Cardiovascular Disease (CVD) than Conventional Lipid Profile: A Study from Kathmandu Valley, Nepal. Journal of clinical and diagnostic research: JCDR 8, 34–36 (2014).
Vishram, J. K. K. et al. Correction: Impact of Age and Gender on the Prevalence and Prognostic Importance of the Metabolic Syndrome and Its Components in Europeans. The MORGAM Prospective Cohort Project. PloS one 10, e0128848 (2015).
Wald, N. J., Simmonds, M. & Morris, J. K. Screening for future cardiovascular disease using age alone compared with multiple risk factors and age. PloS one 6, e18742 (2011).
Goh, L. G. H., Dhaliwal, S. S., Welborn, T. A., Lee, A. H. & Della, P. R. Ethnicity and the association between anthropometric indices of obesity and cardiovascular risk in women: a cross-sectional study. BMJ open 4, e004702 (2014).
Chiuve, S. E. et al. Lifestyle-based prediction model for the prevention of CVD: the Healthy Heart Score. Journal of the American Heart Association 3, e000954–e000954 (2014).
Weihua Mmed, L., Yougang, W. & Jing, W. Reduced or modified dietary fat for preventing cardiovascular disease. The Journal of cardiovascular nursing 28, 204–205 (2013).
Booth, F. W., Roberts, C. K. & Laye, M. J. Lack of exercise is a major cause of chronic diseases. Comprehensive Physiology 2, 1143–1211 (2012).
Chum, A. O. & apos, C. P. Cross-sectional associations between residential environmental exposures and cardiovascular diseases. BMC public health 15, 438 (2015).
Thanassoulis, G. & Vasan, R. S. Genetic cardiovascular risk prediction: will we get there? Circulation 122, 2323–2334 (2010).
Thanassoulis, G. et al. A genetic risk score is associated with incident cardiovascular disease and coronary artery calcium: the Framingham Heart Study. Circulation. Cardiovascular genetics 5, 113–121 (2012).
Becker F. et al. Genetic testing and common disorders in a public health framework: how to assess relevance and possibilities. Background Document to the ESHG recommendations on genetic testing and common disorders. European journal of human genetics: EJHG 19 Suppl 1, S6–44 (2011).
Charrier, C. et al. Inhibition of SRGAP2 function by its human-specific paralogs induces neoteny during spine maturation. Cell 149, 923–935 (2012).
Ma, Y. et al. The inverse F-BAR domain protein srGAP2 acts through srGAP3 to modulate neuronal differentiation and neurite outgrowth of mouse neuroblastoma cells. PloS one 8, e57865 (2013).
Jeemon, P., Pettigrew, K., Sainsbury, C., Prabhakaran, D. & Padmanabhan, S. Implications of discoveries from genome-wide association studies in current cardiovascular practice. World journal of cardiology 3, 230–247 (2011).
Asselbergs, F. W. et al. Large-scale gene-centric meta-analysis across 32 studies identifies multiple lipid loci. American journal of human genetics 91, 823–838 (2012).
Teslovich, T. M. et al. Biological, clinical and population relevance of 95 loci for blood lipids. Nature 466, 707–713 (2010).
Legato, M. J. Gender and the heart: sex-specific differences in normal anatomy and physiology. The journal of gender-specific medicine: JGSM: the official journal of the Partnership for Women's Health at Columbia 3, 15–18 (2000).
Legato, M. J. Postpartum depression and gender-specific justice. The journal of gender-specific medicine: JGSM: the official journal of the Partnership for Women's Health at Columbia 5, 7–8 (2002).
Jiang, Y. et al. Genetic variants at 1p11.2 and breast cancer risk: a two-stage study in Chinese women. PLoS One 6, e21563 (2011).
Crespi, E., Bottai, G. & Santarpia, L. Role of inflammation in obesity-related breast cancer. Curr Opin Pharmacol 31, 114–122 (2016).
Deng, Q. et al. Genetic relationships among four minorities in Guangxi revealed by analysis of 15 STRs. Journal of genetics and genomics = Yi chuan xue bao 34, 1072–1079 (2007).
Yao, Y. et al. The association between HLA-A, -B alleles and major histocompatibility complex class I polymorphic Alu insertions in four populations in China. Tissue antigens 73, 575–581 (2009).
Li, X.-M. et al. Distribution of food-borne parasitic diseases and dietary habits in human population in Guangxi. Zhongguo ji sheng chong xue yu ji sheng chong bing za zhi = Chinese journal of parasitology & parasitic diseases 27, 151–155 (2009).
Ruixing, Y. et al. Effects of demographic, dietary and other lifestyle factors on the prevalence of hyperlipidemia in Guangxi Hei Yi Zhuang and Han populations. Eur J Cardiovasc Prev Rehabil 13, 977–984 (2006).
Ruixing, Y. et al. Comparison of demography, diet, lifestyle, and serum lipid levels between the Guangxi Bai Ku Yao and Han populations. J Lipid Res 48, 2673–2681 (2007).
Ruixing, Y. et al. Interactions of the apolipoprotein C-III 3238C > G polymorphism and alcohol consumption on serum triglyceride levels. Lipids Health Dis 9, 86 (2010).
Yin, R. X., Li, Y. Y., Liu, W. Y., Zhang, L. & Wu, J. Z. Interactions of the apolipoprotein A5 gene polymorphisms and alcohol consumption on serum lipid levels. PLoS One 6, e17954 (2011).
Pilia, G. et al. Heritability of cardiovascular and personality traits in 6,148 Sardinians. PLoS Genet 2, e132 (2006).
Surakka, I. et al. A genome-wide association study of monozygotic twin-pairs suggests a locus related to variability of serum high-density lipoprotein cholesterol. Twin Res Hum Genet 15, 691–699 (2012).
Coleman, W. B. Obesity and the breast cancer methylome. Curr Opin Pharmacol 31, 104–113 (2016).
Wei, L. J. et al. A case-control study on the association between serum lipid level and the risk of breast cancer. Zhonghua Yu Fang Yi Xue Za Zhi 50, 1091–1095 (2016).
Gomes-Rochette, N. F. et al. Association of PvuII and XbaI polymorphisms on estrogen receptor alpha (ESR1) gene to changes into serum lipid profile of post-menopausal women: Effects of aging, body mass index and breast cancer incidence. PLoS One 12, e0169266 (2017).
Bermudez, O. I., Velez-Carrasco, W., Schaefer, E. J. & Tucker, K. L. Dietary and plasma lipid, lipoprotein, and apolipoprotein profiles among elderly Hispanics and non-Hispanics and their association with diabetes. Am J Clin Nutr 76, 1214–1221 (2002).
Slentz, C. A. et al. Inactivity, exercise training and detraining, and plasma lipoproteins. STRRIDE: a randomized, controlled study of exercise intensity and amount. J Appl Physiol (1985) 103, 432–442 (2007).
Erem, C., Hacihasanoglu, A., Deger, O., Kocak, M. & Topbas, M. Prevalence of dyslipidemia and associated risk factors among Turkish adults: Trabzon lipid study. Endocrine 34, 36–51 (2008).
Joffe, Y. T., Collins, M. & Goedecke, J. H. The relationship between dietary fatty acids and inflammatory genes on the obese phenotype and serum lipids. Nutrients 5, 1672–1705 (2013).
Serrano-Martinez, M. et al. To what extent are the effects of diet on coronary heart disease lipid-mediated? Int J Cardiol 95, 35–38 (2004).
Yu-Poth, S. et al. Effects of the National Cholesterol Education Program’s Step I and Step II dietary intervention programs on cardiovascular disease risk factors: a meta-analysis. Am J Clin Nutr 69, 632–646 (1999).
Brinton, E. A. Effects of ethanol intake on lipoproteins and atherosclerosis. Curr Opin Lipidol 21, 346–351 (2010).
Perissinotto, E. et al. Alcohol consumption and cardiovascular risk factors in older lifelong wine drinkers: the Italian Longitudinal Study on Aging. Nutr Metab Cardiovasc Dis 20, 647–655 (2010).
Onat, A. et al. Associations of alcohol consumption with blood pressure, lipoproteins, and subclinical inflammation among Turks. Alcohol 42, 593–601 (2008).
An epidemiological study of cardiovascular and cardiopulmonary disease risk factors in four populations in the People's Republic of China. Baseline report from the P.R.C.-U.S.A. Collaborative Study. People's Republic of China–United States Cardiovascular and Cardiopulmonary Epidemiology Research Group. Circulation 85, 1083–1096 (1992).
Guo, T. et al. Association of the Trp316Ser variant (rs1801690) near the apolipoprotein H (β2-glycoprotein-I) gene and serum lipid levels. International journal of clinical and experimental pathology 8, 7291–7304 (2015).
Guo, T. et al. Suppressor of cytokine signaling 3 A + 930–>G (rs4969168) polymorphism is associated with apolipoprotein A1 and low-density lipoprotein cholesterol. International journal of clinical and experimental pathology 8, 7305–7317 (2015).
Zeng, X.-N. et al. Association of the MLXIPL/TBL2 rs17145738 SNP and serum lipid levels in the Guangxi Mulao and Han populations. Lipids in health and disease 12, 156 (2013).
Ramazauskiene, V., Petkeviciene, J., Klumbiene, J., Kriaucioniene, V. & Sakytė, E. Diet and serum lipids: changes over socio-economic transition period in Lithuanian rural population. BMC public health 11, 447 (2011).
Chalmers, J. et al. World Health Organization-International Society of Hypertension Guidelines for the management of hypertension. Guidelines sub-committee of the World Health Organization., vol. 21: Taylor & Francis, 1999: 1009–1060 (1999).
Whitworth, J. A. World Health Organization I. S. o. H. W. G. World Health Organization (WHO)/International Society of Hypertension (ISH) statement on management of hypertension., vol. 21, 1983–1992 (2003).
Zhou, B.-F. & China, C. M.-A. G. o. t. W. G. o. O. i. Predictive values of body mass index and waist circumference for risk factors of certain related diseases in Chinese adults–study on optimal cut-off points of body mass index and waist circumference in Chinese adults. Biomedical and environmental sciences: BES 15, 83–96 (2002).
Zhang, Z., Murtagh, F., Van Poucke, S., Lin, S. & Lan, P. Hierarchical cluster analysis in clinical research with heterogeneous study population: highlighting its visualization with R. Ann Transl Med 5, 75 (2017).
Acknowledgements
The authors acknowledge the essential role of the funding of the National Natural Science Foundation of China (No. 81460169) and the Innovation Project of Guangxi Graduate Education in this motif.
Author information
Authors and Affiliations
Contributions
Liu Miao conceived the study, participated in the design, undertook genotyping, performed the statistical analyses, and drafted the manuscript. Rui-Xing Yin conceived the study, participated in the design, carried out the epidemiological survey, collected the samples, and helped to draft the manuscript. Shuo Yang collaborated to the genotyping. Jin-Zhen Wu, Wei-Xiong Lin and Shang-Ling Pan carried out the epidemiological survey and collected the samples. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Miao, L., Yin, RX., Wu, JZ. et al. The SRGAP2 SNPs, their haplotypes and G × E interactions on serum lipid traits. Sci Rep 7, 11626 (2017). https://doi.org/10.1038/s41598-017-10950-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-10950-6
- Springer Nature Limited
This article is cited by
-
A novel lncRNA-miRNA-mRNA triple network identifies lncRNA TWF1 as an important regulator of miRNA and gene expression in coronary artery disease
Nutrition & Metabolism (2019)
-
LIPG SNPs, their haplotypes and gene-environment interactions on serum lipid levels
Lipids in Health and Disease (2019)
-
Circulating miR-3659 may be a potential biomarker of dyslipidemia in patients with obesity
Journal of Translational Medicine (2019)
-
A novel circRNA-miRNA-mRNA network identifies circ-YOD1 as a biomarker for coronary artery disease
Scientific Reports (2019)
-
TRIB1 and TRPS1 variants, G × G and G × E interactions on serum lipid levels, the risk of coronary heart disease and ischemic stroke
Scientific Reports (2019)